Human muscle stem cells Racquel N Cooper, Gillian S Butler-Browne and Vincent Mouly Stem cells are unspecialized cells that have been defined in many different ways but they have two important characteristics that distinguish them from other cells in the body. First, they can replenish their numbers for long periods through cell division. Second, after receiving certain chemical signals, they can produce, through asymmetric cell division, a progeny that can differentiate or transform into specialized cells with specific functions, such as heart, nerve or muscle. In recent years, stem cells have received much attention owing to their potential use in cell-based therapies for human neurodegenerative diseases such as Parkinson’s disease, stroke and muscular dystrophies. However, many questions need to be resolved before stem cells with myogenic potential are used in clinical standard protocols. Addresses INSERM U787, Institut de Myologie, 105 bd de l’Ho ¨ pital, 75013 Paris, France Corresponding author: Mouly, Vincent ([email protected]) Current Opinion in Pharmacology 2006, 6:1–6 This review comes from a themed issue on Musculoskeletal Edited by Geoff Goldspink and Brendon Noble 1471-4892/$ – see front matter # 2006 Elsevier Ltd. All rights reserved. DOI 10.1016/j.coph.2006.01.007 Introduction: skeletal muscle stem cells Skeletal muscle needs to employ stem cells for its main- tenance and repair. It contains a potent myogenic stem cell population, called satellite cells, with the ability to self-renew as well as to generate myogenic precursor cells that can regenerate muscle in vivo. However, owing to the gradual decrease in number of satellite cells during the evolution of muscular dystrophies, leading to their final exhaustion [1,2], many attempts have been undertaken to isolate cells able to efficiently repair adult skeletal mus- cle, following the paradigm of haematopoietic stem cells which reconstitute blood cells. Satellite cells Since its identification over 40 years ago, the satellite cell has been a popular candidate for the adult skeletal muscle stem cell. Located beneath the basal lamina of mature skeletal muscle fibres, they are ideally positioned for repair of degenerating muscle fibres. These dormant cells are activated to proliferate upon muscle injury or when heavily used during activities such as weight lifting or running. This proliferation step is necessary to generate sufficient numbers of myoblasts for muscle differentia- tion and myotube formation. In humans and mice, these mononuclear cells are most plentiful at birth (estimated at 32% of sublaminar nuclei) [3]. Their frequency declines post-natally, stabilizing to between 1% and 5% of skeletal muscle nuclei in adult mice [4]. In humans, the propor- tion of satellite cells in skeletal muscles also decreases with age, which could explain the decreased efficiency of muscle regeneration in older subjects [5,6]. Satellite cells from aged muscle also display reduced proliferative and fusion capacity, as well as a tendency to accumulate fat, all of which probably contribute to deteriorating regenera- tion capacity [7]. Recent studies carried out in mice and involving single myofibre transplantations have allowed the direct inves- tigation of the function and behaviour of satellite cell populations [8 ]. It was found that fibres containing as little as seven satellite cells on a transplanted myofibre could generate over 100 new myofibres containing thou- sands of myonuclei. These results indicate that at least some cells in the position of satellite cells (i.e. on the edge of the fibres) present an extended proliferative potential and can repopulate extensively the host’s muscle with an efficiency unknown in any other experimental situation. Whether this population of progenitor, which also pro- vides cells in the satellite compartment, corresponds to bona fide stem cells or simply progenitors with extended proliferative potential is still unknown (see review by Collins, this issue). The identification of multiple stem cell populations resident in skeletal muscle has added further complexity to our understanding of the process of muscle regenera- tion. In adult human tissue, skeletal muscle-derived stem cells (MDSCs) appear to be a distinct population of immature progenitors of satellite cells, but their func- tional properties remain unclear. Muscle-derived stem cells The idea that the regeneration of adult muscle is wholly accomplished by satellite cells has been challenged by the demonstration that muscle also contains a population of adult stem cells termed MDSCs. These cells constitute a unique population that appears to be distinct from muscle satellite cells [9]. MDSCs exhibit the capacity to reconstitute the entire haematopoietic repertoire after intravenous injection into lethally irradiated mice [10,11], although the cells that showed this potential were COPHAR 355 www.sciencedirect.com Current Opinion in Pharmacology 2006, 6:1–6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Human muscle stem cellsRacquel N Cooper, Gillian S Butler-Browne and Vincent Mouly
Stem cells are unspecialized cells that have been defined in
many different ways but they have two important
characteristics that distinguish them from other cells in the
body. First, they can replenish their numbers for long periods
through cell division. Second, after receiving certain chemical
signals, they can produce, through asymmetric cell division, a
progeny that can differentiate or transform into specialized cells
with specific functions, such as heart, nerve or muscle. In
recent years, stem cells have received much attention owing to
their potential use in cell-based therapies for human
neurodegenerative diseases such as Parkinson’s disease,
stroke and muscular dystrophies. However, many questions
need to be resolved before stem cells with myogenic potential
are used in clinical standard protocols.
Addresses
INSERM U787, Institut de Myologie, 105 bd de l’Hopital, 75013 Paris,
France
Corresponding author: Mouly, Vincent ([email protected])
Current Opinion in Pharmacology 2006, 6:1–6
This review comes from a themed issue on
Musculoskeletal
Edited by Geoff Goldspink and Brendon Noble
1471-4892/$ – see front matter
# 2006 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.coph.2006.01.007
Introduction: skeletal muscle stem cellsSkeletal muscle needs to employ stem cells for its main-
tenance and repair. It contains a potent myogenic stem
cell population, called satellite cells, with the ability to
self-renew as well as to generate myogenic precursor cells
that can regenerate muscle in vivo. However, owing to the
gradual decrease in number of satellite cells during the
evolution of muscular dystrophies, leading to their final
exhaustion [1,2], many attempts have been undertaken to
isolate cells able to efficiently repair adult skeletal mus-
cle, following the paradigm of haematopoietic stem cells
which reconstitute blood cells.
Satellite cells
Since its identification over 40 years ago, the satellite cell
has been a popular candidate for the adult skeletal muscle
stem cell. Located beneath the basal lamina of mature
skeletal muscle fibres, they are ideally positioned for
repair of degenerating muscle fibres. These dormant cells
COPHAR 355
www.sciencedirect.com
are activated to proliferate upon muscle injury or when
heavily used during activities such as weight lifting or
running. This proliferation step is necessary to generate
sufficient numbers of myoblasts for muscle differentia-
tion and myotube formation. In humans and mice, these
mononuclear cells are most plentiful at birth (estimated at
32% of sublaminar nuclei) [3]. Their frequency declines
post-natally, stabilizing to between 1% and 5% of skeletal
muscle nuclei in adult mice [4]. In humans, the propor-
tion of satellite cells in skeletal muscles also decreases
with age, which could explain the decreased efficiency of
muscle regeneration in older subjects [5,6]. Satellite cells
from aged muscle also display reduced proliferative and
fusion capacity, as well as a tendency to accumulate fat, all
of which probably contribute to deteriorating regenera-
tion capacity [7].
Recent studies carried out in mice and involving single
myofibre transplantations have allowed the direct inves-
tigation of the function and behaviour of satellite cell
populations [8�]. It was found that fibres containing as
little as seven satellite cells on a transplanted myofibre
could generate over 100 new myofibres containing thou-
sands of myonuclei. These results indicate that at least
some cells in the position of satellite cells (i.e. on the edge
of the fibres) present an extended proliferative potential
and can repopulate extensively the host’s muscle with an
efficiency unknown in any other experimental situation.
Whether this population of progenitor, which also pro-
vides cells in the satellite compartment, corresponds to
bona fide stem cells or simply progenitors with extended
proliferative potential is still unknown (see review by
Collins, this issue).
The identification of multiple stem cell populations
resident in skeletal muscle has added further complexity
to our understanding of the process of muscle regenera-
tion. In adult human tissue, skeletal muscle-derived stem
cells (MDSCs) appear to be a distinct population of
immature progenitors of satellite cells, but their func-
tional properties remain unclear.
Muscle-derived stem cells
The idea that the regeneration of adult muscle is wholly
accomplished by satellite cells has been challenged by
the demonstration that muscle also contains a population
of adult stem cells termed MDSCs. These cells constitute
a unique population that appears to be distinct from
muscle satellite cells [9]. MDSCs exhibit the capacity
to reconstitute the entire haematopoietic repertoire after
intravenous injection into lethally irradiated mice [10,11],
although the cells that showed this potential were
Current Opinion in Pharmacology 2006, 6:1–6

2 Musculoskeletal
identified as having a haematopoietic origin [12]. Further-
more, muscle side population (SP) cells, which can be
isolated by their specific exclusion of the vital dye Hoechst
33342 [10,13], are a fraction of MDSCs that expresses stem
cell antigen-1, a haematopoietic stem cell marker, but no
satellite cell markers such as M-cadherin or Pax7 [14]. In
addition, transplanted SP cells isolated from bone marrow
or muscle actively participate in the formation of skeletal
myotubes during regeneration [13]. Significantly, Pax7-
deficient muscle entirely lacks myogenic satellite cells, yet
contains muscle SP cells that exhibit a high level of
haematopoietic progenitor activity [15].
This review focuses on recent advances in skeletal mus-
cle stem cell research and in particular human muscle
stem cells.
Muscle stem cell plasticitySatellite cells have long been considered monopotential
stem cells with the ability to give rise only to cells of the
myogenic lineage. However, recent experiments have
identified the existence of adult stem cells present in
most tissues that appear to exhibit the ability to differ-
entiate into many different cell types after reintroduction
in vivo [10,14,16–18]. Importantly, these cells have been
recognized as a unique population that appears to be
distinct from muscle satellite cells.
Treatment of C2C12 myoblasts, an immortalized cell line
derived from mouse limb muscle, with bone morphoge-
netic protein activates osteogenic markers, whereas treat-
ment with adipogenic inducers such as fatty acids and
thiazolidinediones activates adipocyte markers [19].
Interestingly, satellite cells derived from intact single
fibre cultures (and thought to be more representative
of true myogenic stem cells) spontaneously form adipo-
cytes and osteoblasts when cultured on Matrigel, a solu-
ble basement membrane matrix lacking strong osteogenic
or adipogenic signals [20]. The adipogenic potential of
myoblasts has also been observed in ageing mouse mus-
cles [7]. The finding that undifferentiated cells in adult
myoblast cultures co-express MyoD, Runx2 and PPARg,
key regulators for myogenesis, osteogenesis and adipo-
genesis, respectively, supports the hypothesis that satel-
lite cells have a multipotential predisposition [21] that is
revealed depending upon the environment. Interestingly,
recent data point to a possible role for wnt signalling in
maintaining the physiological balance between adipo-
genic and myogenic potential of satellite cells. Mice
deficient for Wnt 10b display an increased fat infiltration
within muscle, a phenomenon that has been described
during physiological ageing where wnt10b is downregu-
lated [22].
The plasticity of muscle stem cells has also been demon-
strated using ex vivo approaches. Mouse muscle stem cells
isolated via serial preplating contribute to regenerating
Current Opinion in Pharmacology 2006, 6:1–6
myofibres when injected directly into dystrophic muscle,
and are also detected among differentiated vascular and
nerve populations [18,23]. Furthermore, these cells,
which express the myoblast markers desmin and MyoD,
are sufficient to completely heal skull defects in vivowhen engineered to express bone morphogenetic protein
[24]. These MDSCs are also capable of reconstituting
bone marrow in lethally irradiated mice [25].
Nevertheless, these studies do have a limitation in that
they used animal models, and few studies have focused
on human stem cells. Alessandri et al. [26] have recently
shown that adult human skeletal muscle includes a
population of progenitor stem cells that can generate
stem cells of the same lineage and cells with neurogenic
properties. Consequently, human skeletal muscle could
represent an important alternative source to isolate plur-
ipotent stem cells for the development of cell-based
therapies for human myogenic and neurogenic diseases.
Likewise, neural stem cells that have been isolated from
adult human skeletal muscle offer a possible alternative
source of neural stem cells for tissue engineering of the
central nervous system [27].
Other stem cells with myogenic potentialIn recent years, many data have been published indicat-
ing the ability of multipotent adult stem cell populations
from a given tissue to differentiate into myogenic cells.
The majority of these studies were performed in rodent
models, whereas a few used human cell sources. Pioneer-
ing studies were performed by Ferrari and colleagues in
1998 [28], in which unfractionated bone marrow-derived
cells were capable of myogenic conversion. However,
repair by bone marrow-derived cells never attained
>1% of total muscle fibres during the lifespan of trans-
planted mdx mouse muscles and did not improve murine
muscular dystrophy [29]. Gussoni et al. [13] transplanted
several thousand haematopoietic stem cells from male
mice into female mdx mice and were able to show that
donor cells efficiently replenished bone marrow of reci-
pients; cells from males expressing dystrophin were found
at low levels in host muscles, indicating differentiation of
transplanted cells into muscle. Blau and colleagues have
shown more recently that bone marrow cells can partici-
pate in muscle regeneration by first being converted to
satellite cells [30].
Intra-arterial injections of wild-type mesoangioblasts —
stem cells isolated from blood vessels — into mice
suffering from limb girdle muscle dystrophy results in
complete functional recovery of all affected muscles [31].
This presents a promising solution to difficulties encoun-
tered with myoblast transplantation therapy, and makes
all muscles accessible for treatment. This is especially
important for the treatment of muscles such as the dia-
phragm, impairment of which results in severe respiratory
problems.
www.sciencedirect.com
Human muscle stem cells Cooper, Butler-Browne and Mouly 3
However, one would have to scale up the procedure to
repair the large muscles found in humans. This would
require large numbers of cells, and human mesoangio-
blast stem cell cultures have not yet been described.
Human-derived AC133, a newly identified glycosylated
polypeptide expressed on a population of circulating
human hematopoeitic/endothelial progenitor cells [32],
can repopulate bone marrow and differentiate into mature
endothelial cells. Torrente et al. [32] have shown that,
after injection of human AC133+ cells into skeletal muscle
of scid/mdx mice, an immunodeficient strain of mice that
lacks dystrophin, up to 10% of human dystrophin-positive
fibres could be detected, suggesting that blood-derived
human AC133+ cells participated in muscle regeneration
of dystrophic muscles. It should be noted that, although
some of these AC133+ cells were of circulating origin, no
data have been published concerning a systemic delivery
of these cells, even in animal models, which would be a
pre-requisite to successful cell treatment of major dys-
trophies such as Duchenne muscular dystrophy (DMD).
Rodriguez et al. [33] have more recently demonstrated the
isolation of adipogenic, myogenic and osteogenic lineages
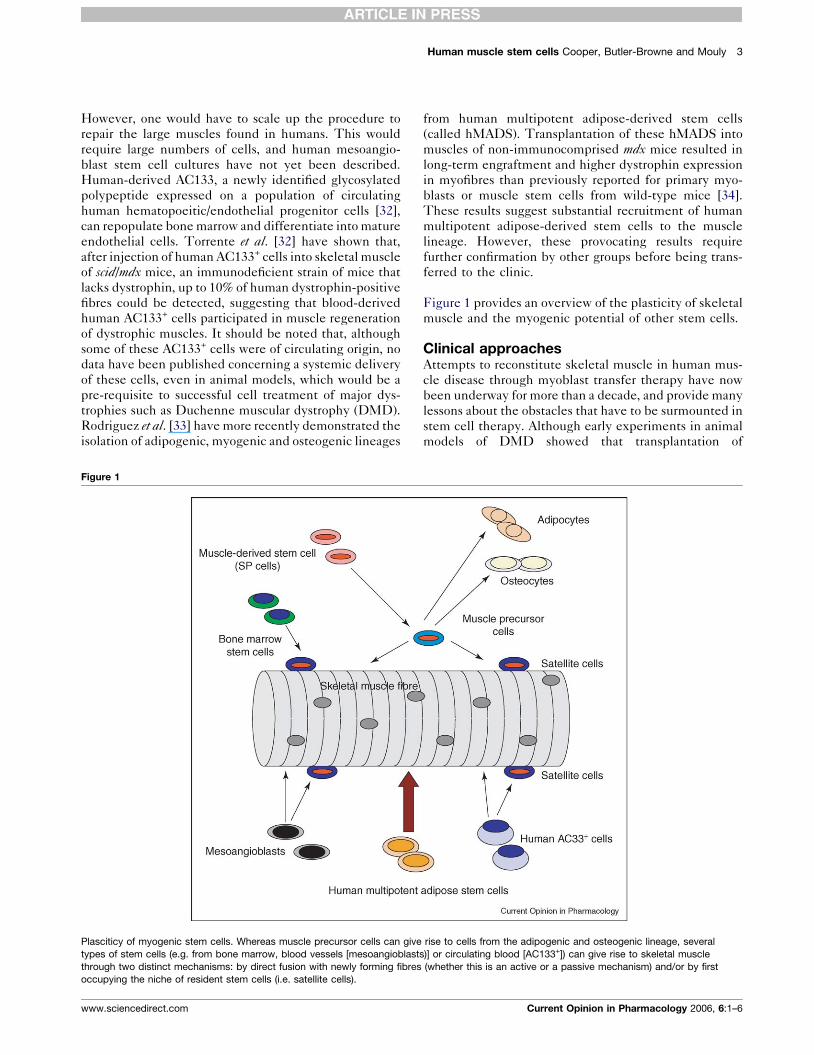
Figure 1
Plasciticy of myogenic stem cells. Whereas muscle precursor cells can give
types of stem cells (e.g. from bone marrow, blood vessels [mesoangioblast
through two distinct mechanisms: by direct fusion with newly forming fibres
occupying the niche of resident stem cells (i.e. satellite cells).
www.sciencedirect.com
from human multipotent adipose-derived stem cells
(called hMADS). Transplantation of these hMADS into
muscles of non-immunocomprised mdx mice resulted in
long-term engraftment and higher dystrophin expression
in myofibres than previously reported for primary myo-
blasts or muscle stem cells from wild-type mice [34].
These results suggest substantial recruitment of human
multipotent adipose-derived stem cells to the muscle
lineage. However, these provocating results require
further confirmation by other groups before being trans-
ferred to the clinic.
Figure 1 provides an overview of the plasticity of skeletal
muscle and the myogenic potential of other stem cells.
Clinical approachesAttempts to reconstitute skeletal muscle in human mus-
cle disease through myoblast transfer therapy have now
been underway for more than a decade, and provide many
lessons about the obstacles that have to be surmounted in
stem cell therapy. Although early experiments in animal
models of DMD showed that transplantation of
rise to cells from the adipogenic and osteogenic lineage, several
s)] or circulating blood [AC133+]) can give rise to skeletal muscle
(whether this is an active or a passive mechanism) and/or by first
Current Opinion in Pharmacology 2006, 6:1–6

4 Musculoskeletal
committed muscle stem cells from normal muscle
resulted in partial restoration of the deficient dystrophin
protein, a high percentage of introduced cells died within
a short time after transplant [35]. Although such a high
level of cell death still needs to be estimated when human
cells are injected, clinical trials in DMD have showed no
significant clinical improvement. Roadblocks to the use-
fulness of this therapy include limited migration and
proliferative capacity of donor cells in dystrophic muscle,
problems with poor donor cell survival, immunorejection
and inefficient myogenic contribution [36]. Difficulties in
delivering myoblasts to a wide range of muscle tissue
remain to be overcome. No systemic delivery of comm-
ited myogenic cells has ever been successful, although
only systemic delivery could reply to the extended degen-
eration observed in many muscle degenerative diseases,
such as DMD, where the majority of the musculature is
targeted by the disease. Despite enormous hope and
enthusiasm for this strategy by investigators and the
muscular dystrophy community when it was introduced,
and although some progress has been made, we are still a
long way from efficient therapy.
Adult stem cells have been shown in animal models to
repair heart damage, provide therapeutic benefit for
stroke, and reverse diabetes. In human patients, these
cells have been employed successfully to relieve lupus,
multiple sclerosis and arthritis, whereas the regenerative
potential of embryonic stem cells has not yet been tested,
as efficacy and safety tests are still underway. However,
although embryonic stem cells might not be immuno-
genic per se, they may turn to be so when they engage to a
more differentiated pathway.
Stem cell research for the treatment of degenerative
diseases is in its infancy and many issues require resolu-
tion before clinical application. The problem of using
autologous or allogenic cells is crucial to the development
of therapeutic trials. Until tolerance to allogenic cells can
be safely induced, the injection of allogenic cells will
require heavy immuno-depressive treatment, which can-
not be sustained for prolonged periods of time owing to
the secondary effects of these treatments. On the other
hand, autologous cells cannot be used if they are already
exhausted by the degenerative process. There have,
however, been a few clinical trials employing autologous
adult human muscle stem cells to treat heart disease and
some muscular dystrophies.
Myocardial regeneration
The first demonstration of successful treatment for heart
disease using the patient’s own adult muscle stem cells
was performed in 2000 by Menasche et al. [37]. Auto-
logous human myoblasts were isolated from the quad-
riceps of the patient, propagated and expanded ex vivountil enough cells are obtained to be injected into the
ventricular wall. After eight months of follow-up, the
Current Opinion in Pharmacology 2006, 6:1–6
grafted cells were functionally integrated and augmented
the function of the patients’ heart. These encouraging
results underline the potential of this new approach and
further clinical trials are now underway in Europe and the
US to investigate the effectiveness of muscle stem cell
therapy in improving function of failing hearts. Autolo-
gous myoblasts are considered possible donor cells
because they possess an inherent capacity for contracti-
lity. They are resistant to ischemia and do not raise
immunologic and ethical concerns.
Muscular dystyrophies
From the results obtained with heart infarct, it was
recently proposed that autologous myoblasts could ame-
liorate the conditions of patients suffering from muscular
dystrophies in cases when the disease leaves enough
muscles spared by the pathological evolution. One such
example is fascio-scapulo-humeral muscular dystrophy,
where myoblasts isolated from muscles spared by the
disease showed normal regenerative capacity [38]. In
oculo-pharyngeal muscular dystrophy, only pharyngeal
and eyelid muscles are targeted by the disease in most
patients. A Phase I clinical trial involving autologous
transfer of myoblasts into pharyngeal muscles has
recently been launched (Perie, Lacau St Guily, Marolleau,
Butler-Browne and Mouly, unpublished).
Perspectives and conclusionsFollowing the successful use of blood stem cells to
reconstitute cells damaged in diseases such as leukaemia,
researchers have been interested in the use of other types
of stem cells to repair adult tissue damaged through injury
or degenerative disease. However, it has until now been
difficult to isolate pure populations of adult stem cells in
large numbers. Exciting results recently obtained by
Montarras et al. [39�] reveal the development of a new
purification procedure that gives direct access to muscle
stem cells that can both repair and contribute to the
progenitor cell population of damaged muscles. The
isolated cells are more efficient contributors to muscle
repair than are the cultured muscle precursor cells pre-
viously used, as 20 000 purified muscle stem cells were
sufficient to promote significant muscle repair when
grafted in dystrophic mouse muscles as compared with
one million cultured muscle precursor cells. It is sug-
gested that the higher regenerative capacity of freshly
isolated stem cells reflects the ability of these cells to
more effectively colonise grafted muscle compared with
cultured cells, which undergo modifications that make
them less efficient, partly because they tend to differ-
entiate too quickly and thereby lose their ability to
regenerate damaged cells. Collins et al. [8�] have recently
demonstrated in the mouse that the implantation of single
fibres with their resident satellite cells into regenerating
muscles of host mice results in an extremely efficient
participation of the cells originally resident to the
implanted fibre to the host’s regeneration (see review
www.sciencedirect.com

Human muscle stem cells Cooper, Butler-Browne and Mouly 5
by Collins, this issue). Although not yet applicable to
human therapy, the aforementioned results provide new
insight for therapeutic use of muscle precursors cells in
humans.
Stem cell therapy holds huge promise for many illnesses
that are presently incurable. This field is currently in its
early stages of development, and a huge amount of work is
a prerequisite to fulfilling these promises. Information
about scientific achievements needs to be communicated
to the public properly, in a rigorous and cautious way, so
that it does not raise excessive expectations. Despite the
numerous obstacles, the potential for stem cell-based
therapy in the treatment of muscular dystrophies and
degenerative diseases of all organ systems provides a
stimulating field for continued research.
AcknowledgementsThe laboratory has received support from Association Francaise contre lesMyopathies (AFM), the Parent Project, Universite Pierre et Marie Curie,CNRS, INSERM and EC (Program Ageing Muscle in the 5th FP andMYORES Network of Excellence, contract 511978, from the EuropeanCommission 6th Framework Programme). The authors wish to thank allthe other members of the laboratory, as well as Martin Catala and WoodyWright for fruitful discussions and Terry Partridge for sharing observationsand concepts.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest�� of outstanding interest
1. Decary S, Hamida CB, Mouly V, Barbet JP, Hentati F,Butler-Browne GS: Shorter telomeres in dystrophic muscleconsistent with extensive regeneration in young children.Neuromuscul Disord 2000, 10:113-120.
2. Heslop L, Morgan JE, Partridge TA: Evidence for a myogenicstem cell that is exhausted in dystrophic muscle. J Cell Sci2000, 113:2299-2308.
3. Cardasis CA, Cooper GW: An analysis of nuclear numbers inindividual muscle fibers during differentiation and growth: asatellite cell-muscle fiber growth unit. J Exp Zool 1975,191:347-358.
4. Bischoff R: The satellite cell and muscle regeneration. InMyology Volume 1, edn 2. Edited by Engel AG, Franzini-ArmstrongC.New York: McGraw-Hill, Inc; 1994:97-118.
5. Decary S, Mouly V, Hamida CB, Sautet A, Barbet JP,Butler-Browne GS: Replicative potential and telomere lengthin human skeletal muscle: implications for satellite cell-mediated gene therapy. Hum Gene Ther 1997, 8:1429-1438.
6. Renault V, Thornell LE, Eriksson PO, Butler-Browne G, Mouly V:Regenerative potential of human skeletal muscle duringaging. Aging Cell 2002, 1:132-139.
7. Taylor-Jones JM, McGehee RE, Rando TA, Lecka-Czernik B,Lipschitz DA, Peterson CA: Activation of an adipogenic programin adult myoblasts with age. Mech Ageing Dev 2002,123:649-661.
8.�
Collins CA, Olsen I, Zammit PS, Heslop L, Petrie A, Partridge TA,Morgan JE: Stem cell function, self-renewal, and behavioralheterogeneity of cells from the adult muscle satellite cellniche. Cell 2005, 122:289-301.
By transplanting freshly isolated single mouse fibres with their residentsatellite cells into irradiated host muscles, these authors show that as fewas seven satellite cells can generate over 100 new myofibers containingthousands of myonuclei, as well as new satellite cells that can be
www.sciencedirect.com
recruited through subsequent cycles of regeneration. This efficiency ismuch higher than that observed in any experimental study involvingtransplantation of cells into skeletal muscle. Whether the isolated fibrecontains a more potent progenitor that cannot be isolated in culture, but ismaintained within the freshly isolated fibre, is not yet known. Intrerest-ingly, these results can only be obtained if the mouse recipient muscle hasbeen irradiated prior to the transplantation of the fibre. This set of resultsopens a new field of research to understand how such an efficiency canbe reached, suggesting that cells located at the periphery of the fibreshave real stem potential.
9. Deasy BM, Li Y, Huard J: Tissue engineering with muscle-derived stem cells. Curr Opin Biotechnol 2004, 15:419-423.
10. Jackson KA, Mi T, Goodell MA: Hematopoietic potential of stemcells isolated from murine skeletal muscle. Proc Natl Acad SciUSA 1999, 96:14482-14486.
11. Kawada H, Ogawa M: Hematopoietic progenitors and stemcells in murine muscle. Blood Cells Mol Dis 2001, 27:605-609.
12. McKinney-Freeman SL, Jackson KA, Camargo FD, Ferrari G,Mavilio F, Goodell MA: Muscle-derived hematopoietic stemcells are hematopoietic in origin. Proc Natl Acad Sci USA 2002,99:1341-1346.
13. Gussoni E, Soneoka Y, Strickland CD, Buzney EA, Khan MK,Flint AF, Kunkel LM, Mulligan RC: Dystrophin expression inthe mdx mouse restored by stem cell transplantation.Nature 1999, 401:390-394.
14. Asakura A, Seale P, Girgis-Gabardo A, Rudnicki MA:Myogenic specification of side population cells in skeletalmuscle. J Cell Biol 2002, 159:123-134.
15. Seale P, Sabourin LA, Girgis-Gabardo A, Mansouri A, Gruss P,Rudnicki MA: Pax7 is required for the specification of myogenicsatellite cells. Cell 2000, 102:777-786.
16. Gussoni E, Blau HM, Kunkel LM: The fate of individual myoblastsafter transplantation into muscles of DMD patients. Nat Med1997, 3:970-977.
17. Torrente Y, Tremblay JP, Pisati F, Belicchi M, Rossi B, Sironi M,Fortunato F, El Fahime M, D’Angelo MG, Caron NJ et al.:Intraarterial injection of muscle-derived CD34(+)Sca-1(+)stem cells restores dystrophin in mdx mice. J Cell Biol 2001,152:335-348.
18. Qu-Petersen Z, Deasy B, Jankowski R, Ikezawa M, Cummins J,Pruchnic R, Mytinger J, Cao B, Gates C, Wernig A et al.:Identification of a novel population of muscle stem cells inmice: potential for muscle regeneration. J Cell Biol 2002,157:851-864.
19. Teboul L, Gaillard D, Staccini L, Inadera H, Amri EZ, Grimaldi PA:Thiazolidinediones and fatty acids convert myogenic cells intoadipose-like cells. J Biol Chem 1995, 270:28183-28187.
20. Asakura A, Komaki M, Rudnicki M: Muscle satellite cells aremultipotential stem cells that exhibit myogenic, osteogenic,and adipogenic differentiation. Differentiation 2001, 68:245-253.
21. Wada MR, Inagawa-Ogashiwa M, Shimizu S, Yasumoto S,Hashimoto N: Generation of different fates from multipotentmuscle stem cells. Development 2002, 129:2987-2995.
22. Vertino AM, Taylor-Jones JM, Longo KA, Bearden ED,Lane TF, McGehee RE Jr, MacDougald OA, Peterson CA:Wnt10b deficiency promotes coexpression of myogenicand adipogenic programs in myoblasts. Mol Biol Cell 2005,16:2039-2048.
23. Qu Z, Balkir L, van Deutekom JC, Robbins PD, Pruchnic R,Huard J: Development of approaches to improve cell survivalin myoblast transfer therapy. J Cell Biol 1998, 142:1257-1267.
24. Lee JY, Qu-Petersen Z, Cao B, Kimura S, Jankowski R,Cummins J, Usas A, Gates C, Robbins P, Wernig A et al.: Clonalisolation of muscle-derived cells capable of enhancing muscleregeneration and bone healing. J Cell Biol 2000, 150:1085-1100.
25. Cao B, Zheng B, Jankowski RJ, Kimura S, Ikezawa M, Deasy B,Cummins J, Epperly M, Qu-Petersen Z, Huard J: Muscle stemcells differentiate into haematopoietic lineages but retainmyogenic potential. Nat Cell Biol 2003, 5:640-646.
Current Opinion in Pharmacology 2006, 6:1–6

6 Musculoskeletal
26. Alessandri G, Pagano S, Bez A, Benetti A, Pozzi S, Iannolo G,Baronio M, Invernici G, Caruso A, Muneretto C et al.: Isolation andculture of human muscle-derived stem cells able todifferentiate into myogenic and neurogenic cell lineages.Lancet 2004, 364:1872-1883.
27. Schultz SS, Lucas PA: Human stem cells isolated from adultskeletal muscle differentiate into neural phenotypes. JNeurosci Methods 2005, 152:144-155.
28. Ferrari G, Cusella-De Angelis G, Coletta M, Paolucci E,Stornaiuolo A, Cossu G, Mavilio F: Muscle regeneration bybone marrow derived myogenic progenitors. Science 1998,279:1528-1530.
29. Ferrari G, Stornaiuolo A, Mavilio F: Failure to correct murinemuscular dystrophy. Nature 2001, 411:1014-1015.
30. Corbel SY, Lee A, Yi L, Duenas J, Brazelton TR, Blau HM,Rossi FM: Contribution of hematopoietic stem cells to skeletalmuscle. Nat Med 2003, 9:1528-1532.
31. Sampaolesi M, Torrente Y, Innocenzi A, Tonlorenzi R, D’Antona G,Pellegrino MA, Barresi R, Bresolin N, De Angelis MG, Campbell KPet al.: Cell therapy of alpha-sarcoglycan null dystrophic micethrough intra-arterial delivery of mesoangioblasts. Science2003, 301:487-492.
32. Torrente Y, Belicchi M, Sampaolesi M, Pisati F, Meregalli M,D’Antona G, Tonlorenzi R, Porretti L, Gavina M, Mamchaoui Ket al.: Human circulating AC133(+) stem cells restoredystrophin expression and ameliorate function in dystrophicskeletal muscle. J Clin Invest 2004, 114:182-195.
33. Rodriguez AM, Pisani D, Dechesne CA, Turc-Carel C,Kurzenne JY, Wdziekonski B, Villageois A, Bagnis C,Breittmayer JP, Groux H et al.: Transplantation of a multipotentcell population from human adipose tissue induces dystrophin
Current Opinion in Pharmacology 2006, 6:1–6
expression in the immunocompetent mdx mouse. J Exp Med2005, 201:1397-1405.
34. Mueller GM, O’Day T, Watchko JF, Ontell M: Effect of injectingprimary myoblasts versus putative muscle-derived stem cellson mass and force generation in mdx mice. Hum Gene Ther2002, 13:1081-1090.
35. Beauchamp JR, Morgan JE, Pagel CN, Partridge TA: Dynamics ofmyoblast transplantation reveal a discrete minority ofprecursors with stem cell-like properties as the myogenicsource. J Cell Biol 1999, 144:1113-1122.
36. Negroni E, Butler-Browne GS, Mouly V: Myogenic stem cells:regeneration and cell therapy in human skeletal muscle.Pathol Biol 2005.
37. Menasche P, Hagege AA, Scorsin M, Pouzet B, Desnos M,Duboc D, Schwartz K, Vilquin JT, Marolleau JP: Myoblasttransplantation for heart failure. Lancet 2001, 357:279-280.
38. Vilquin JT, Marolleau JP, Sacconi S, Garcin I, Lacassagne MN,Robert I, Ternaux B, Bouazza B, Larghero J, Desnuelle C: Normalgrowth and regenerating ability of myoblasts from unaffectedmuscles of facioscapulohumeral muscular dystrophypatients. Gene Ther. 2005, 12:1651-1662.
39.�
Montarras D, Morgan J, Collins C, Relaix F, Zaffran S, Cumano A,Partridge T, Buckingham M: Direct isolation of satellite cells forskeletal muscle regeneration. Science 2005, 309:2064-2067.
The characteristics of Pax3+ myogenic cells were determined by FACSusing Pax3-GFP mice as a source of cells. Grafting of as few as 104 freshlyisolated cells, sorted by FACS using the same characteristics, showedthat these freshly isolated cells were 10-50 times more efficient thanmyogenic cells expanded in culture. These data can be explained eitherby the presence of a population of cells that is lost during expansion inculture, or by modifications of the cells during the expansion in vitro.
www.sciencedirect.com
Related Documents











