Human Immunodeficiency Virus Type I and Its Coat Protein gpUO Apoptosis and Activate JNK a Mitogen-Activated Protein Kr Human Neurons Induce .nd ERK . nases in Annie Lannuzel,* Jean Vianney Barnier,t Christiane Hery,* Huynh Van Tan,$ Bernard Guibert,? FranGoise Gray,$ Jean Didier Vincent,? and Marc Tardieu* Detection of apoptotic neurons and microglial cells in the brains of human immunodeficiency virus type 1 (HIV-1)- infected patients bas suggested that programmed cell death may be implicated in the physiopathology of HIV-1 enceph- alopathy. To analyze in vitro the intracellular signals induced by HIV-1 in human neurons and the associated neuronal death, we tested cultured human central nervous system (CNS) cells for apoptosis induced by HIV-1 and gp120 and for signaling pathways activated by gp120. HIV-1 and gp120 induced apoptosis of neurons and microglial cells but not of astrocytes or transformed microglial cells. Gp120 activated c-Jun N-terminal kinase UNK) and p42 extracellular- regulated kinase (ERK) in primary CNS cells, with an early peak of activation at 2 to 5 minutes that was not present when pure microglial or astrocyte cultures were tested, followed by a late and sustained activation (10 and 60 minutes) in primary and enriched glial cell cultures as well as in transformed microglial cells. This demonstrates that gp120 could be an effector of HIV-1-induced apoptosis in the CNS and act directly on neuronal and glial cells. Lannuzel A, Barnier JV, Hery C, Huynh VT, Guibert B, Gray F, Vincent JD, Tardieu M. Human immunodeficiency virus type 1 and its coat protein gp120 induce apoptosis and activate JNK and ERK mitogen-activated protein kinases in human neurons. Ann Neurol 1997;42:847-856 Human immunodeficiency virus type 1 (HIV-1) infec- tion can induce neuronal alterations and brain dysfunc- tion, as indicated by the emergence of neurological symptoms in patients at the stage of severe immuno- deficiency and by studies using nuclear magnetic reso- nance spectroscopy [l-41. O n pathological examina- tion of HIV-1-infected brains, a 40% neuronal loss has been demonstrated, accompanied by extensive dis- ruption of the dendriric arbor [5-7]. More recently, apoptotic neurons and apoptotic microglial cells have also been detected, suggesting that programmed cell death is implicated in the physiopathology of the HIV- 1 encephalopathy [8-lo]. Several viral and cellu- lar factors could be responsible for the induced neuro- nal death, among which the envelope glycoprotein gp120 of HIV-1 was considered very early, because in vitro it is toxic for rodent neurons (for review, see Ref- erences 11-16) and induces apoptosis [17, 181. Its mechanisms of action appeared to depend on inhibi- tion of neurotrophic factors [13, 131 or on the activa- tion of voltage-gated calcium channels and/or gluta- mate receptors of the N-methyl-D-aspartate (NMDA) subtype, induced either by gp12O itself or by soluble factors released by gpI20-activated microglial cells [I 5, 17, 201. In a previous study, we demonstrated that en- velope proteins gp120 and gpl60 induce neurotoxicity in human embryonic neurons through a potentiation of NMDA-induced increase of intracellular calcium concentration ([Ca”’],) and an alteration of [Ca2’], homeostasis 1211. Apoptosis is a programmed cell death occurring in both physiological and pathological conditions in re- sponse to various stimuli originating from either within or outside the cell [22]. Apoptosis is usually in balance with cellular differentiation and multiplication. The initial stimulus induces activation of intracellular trans- From the *Laboratoire “Virus, Neurone, Irnmunitb,” Facult6 de Mkdecine Paris-Sud, Universitk Paris-Sud; tInstitut Alfred Fessard, CNRS, Gif/Yvette; $Laboratoire “Signalisation Cellulaire et Para- sites,” UFR Cochin, Universitt RenC Descartes, Paris; and SDC- partement de Neuropathologie, HBpital Raymond PoincarC, Garches, France. Received Jan 16, 1997, and in revised form Jun 2. Accepted for publication Aug 1, 1997. Address correspondence to Dr Tardieu, Laboratoire Virus, Neurone, ImmunitC, Facultk de MCdecine, 63 rue Gabriel PCri, 94276 Le Kremlin-Bidtre Cedex, France. Copyright 0 1997 by the American Neurological Association 847

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Human Immunodeficiency Virus Type I and Its Coat Protein gpUO
Apoptosis and Activate JNK a Mitogen-Activated Protein Kr
Human Neurons
Induce .nd ERK . nases in
Annie Lannuzel,* Jean Vianney Barnier,t Christiane Hery,* Huynh Van Tan,$ Bernard Guibert,? FranGoise Gray,$ Jean Didier Vincent,? and Marc Tardieu*
Detection of apoptotic neurons and microglial cells in the brains of human immunodeficiency virus type 1 (HIV-1)- infected patients bas suggested that programmed cell death may be implicated in the physiopathology of HIV-1 enceph- alopathy. To analyze in vitro the intracellular signals induced by HIV-1 in human neurons and the associated neuronal death, we tested cultured human central nervous system (CNS) cells for apoptosis induced by HIV-1 and gp120 and for signaling pathways activated by gp120. HIV-1 and gp120 induced apoptosis of neurons and microglial cells but not of astrocytes or transformed microglial cells. Gp120 activated c-Jun N-terminal kinase UNK) and p42 extracellular- regulated kinase (ERK) in primary CNS cells, with an early peak of activation at 2 to 5 minutes that was not present when pure microglial or astrocyte cultures were tested, followed by a late and sustained activation (10 and 60 minutes) in primary and enriched glial cell cultures as well as in transformed microglial cells. This demonstrates that gp120 could be an effector of HIV-1-induced apoptosis in the CNS and act directly on neuronal and glial cells.
Lannuzel A, Barnier JV, Hery C, Huynh VT, Guibert B, Gray F, Vincent JD, Tardieu M. Human immunodeficiency virus type 1 and its coat protein gp120 induce apoptosis and activate JNK and ERK mitogen-activated protein
kinases in human neurons. Ann Neurol 1997;42:847-856
Human immunodeficiency virus type 1 (HIV-1) infec- tion can induce neuronal alterations and brain dysfunc- tion, as indicated by the emergence of neurological symptoms in patients at the stage of severe immuno- deficiency and by studies using nuclear magnetic reso- nance spectroscopy [l-41. O n pathological examina- tion of HIV-1-infected brains, a 40% neuronal loss has been demonstrated, accompanied by extensive dis- ruption of the dendriric arbor [5-7]. More recently, apoptotic neurons and apoptotic microglial cells have also been detected, suggesting that programmed cell death is implicated in the physiopathology of the HIV- 1 encephalopathy [8-lo]. Several viral and cellu- lar factors could be responsible for the induced neuro- nal death, among which the envelope glycoprotein gp120 of HIV-1 was considered very early, because in vitro it is toxic for rodent neurons (for review, see Ref- erences 11-16) and induces apoptosis [17, 181. Its
mechanisms of action appeared to depend on inhibi- tion of neurotrophic factors [13, 131 or on the activa- tion of voltage-gated calcium channels and/or gluta- mate receptors of the N-methyl-D-aspartate (NMDA) subtype, induced either by gp12O itself or by soluble factors released by gpI20-activated microglial cells [I 5 , 17, 201. In a previous study, we demonstrated that en- velope proteins gp120 and gpl60 induce neurotoxicity in human embryonic neurons through a potentiation of NMDA-induced increase of intracellular calcium concentration ([Ca”’],) and an alteration of [Ca2’], homeostasis 1211.
Apoptosis is a programmed cell death occurring in both physiological and pathological conditions in re- sponse to various stimuli originating from either within or outside the cell [22]. Apoptosis is usually in balance with cellular differentiation and multiplication. The initial stimulus induces activation of intracellular trans-
From the *Laboratoire “Virus, Neurone, Irnmunitb,” Facult6 de Mkdecine Paris-Sud, Universitk Paris-Sud; tInstitut Alfred Fessard, CNRS, Gif/Yvette; $Laboratoire “Signalisation Cellulaire et Para- sites,” UFR Cochin, Universitt RenC Descartes, Paris; and SDC- partement de Neuropathologie, HBpital Raymond PoincarC, Garches, France.
Received Jan 16, 1997, and in revised form Jun 2. Accepted for publication Aug 1, 1997.
Address correspondence to Dr Tardieu, Laboratoire Virus, Neurone, ImmunitC, Facultk de MCdecine, 63 rue Gabriel PCri, 94276 Le Kremlin-Bidtre Cedex, France.
Copyright 0 1997 by the American Neurological Association 847

duction pathways including mitogen-activated protein (MAP) kinases (MAPKs), which converge toward en- donuclease and protease activations [23]. In mamma- lian cells, three different MAPKs have been described, ie, extracellular signal-regulated kinases (ERKs), c-Jun N-terminal kinases (JNKs, also called SAPKs for stress- activated protein kinases), and p38 [24]. ’They belong to three different protein kinase cascades, Raf-MEK- ERK, SEK-JNWSAPK, and MKK3/MKK6-p38. Clas- sic hypotheses consider apoptosis to be associated with activation of the SEK-JNK/SAPK and MKK3/MKK6- p38 pathways, and regulation of growth and differen- tiation with activation of the Raf-MEK-ERK pathway [24]. This view should probably be amended, because a balance between activation of JNK/p38 and inhibition of the ERK pathway has been demonstrated to lead to apoptosis in PC12 cells [25].
The present study was performed to analyze the neu- ronal death induced by HIV-1 in human central ner- vous system (CNS) cells and the intracellular signals produced after HIV- 1 envelope glycoprotein contact with CNS cells. We observed that HIV-1 and soluble gpI2O induced apoptosis of neurons and microglial cells but not of astrocytes. Gp120 also induced an ac- tivation of MAPKs in human neurons, astrocytes, and microglial cells, although the kinetics of activation dif- fered between the different cell types.
Materials and Methods Cells and Cultures Human CNS cell cultures were prepared from 8- to 10- week-old embryos obtained after elective abortion in com- pliance with the recommendations of the French National Ethics Committee and after approval by the local ethics committee.
PRIMARY CULTURES. Primary cultures of human CNS were prepared as described [21, 261. Spinal cord and prosen- cephalon were dissected, trypsinized, centrifuged, resus- pended, and placed in MCDB 153 (Polylabo, France) com- pleted with 2 mM glutamine, 7.5% fetal calf serum, lo5 IU/L penicillin, and 0.1 g/L streptomycin. The cell suspen- sion was distributed at a density of 7 X 105/22 mm2 culture well coated with collagen. Cultures were placed in a 10% C0,-humidified incubator at 37°C. The medium was re- moved 48 hours after plating and changed by one-half every 3 to 4 days. Cultures consisted of nerve cell adhesion mole- cule- and neurofilament-positive neuronal clusters lying on a monocellular layer containing 50 to 70% glial fibrillary acidic protein (GFAP)-positive astrocytes and 30 to 50% CD68/KiM-7-positive microglial cells associated with fibro- blasts, as described [27].
CULTURES ENRICHED IN MICROGLIA OR ASTROCYTES. Ten to 15 days after plating, the microglial cells were re- leased from primary cultures by circular shaking and adher- ent cells were selected as described j26, 281. They were sub-
sequently grown in minimal essential medium containing 6 g/L glucose, 2 mM glutamine, 10% fetal calf serum, lo5 IU/L penicillin, and 0.1 g/L streptomycin for 3 weeks to 2 months. The culture contained between 80 and 90% CD681 KiM-7-positive microglial cells. The adherent cells remain- ing after release of microglial cells were trypsinized, plated, and passaged several times to obtain purified cultures of as- trocytes (>90% GFAP-positive cells).
Microglial cell lines were established by transfection of primary human embryonic microglial cells with a plasmid encoding for the large T antigen of SV40 by lipofection as described [28].
HIV-1 Isolate, Recombinant gp 120, and Induction o f Apoptosis In experiments testing the induction of apoptosis, we used the monotropic strain 9533 of HIV-1 or two different re- combinant gp120 proteins. The first recombinant gp120 was obtained from the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH, Rockville, MD. It was produced by transfection of the Chinese hamster ovary (CHO) cell line with a plasmid containing the SF2 envelope coding sequence G l ~ ” - A r g ~ ~ ’ [29-3 11. This enve- lope glycoprotein was purified by gel scanning to more than 90%. Impurities are human transferrin (-30 mg/ml), bovine serum albumin (55 nglml), bovine fetuin (7 ng/ml), and bo- vine IgG (<I ng/ml). The second recombinant gp120 ob- tained from Neosystem (Strasbourg, France) was expressed on baculovirus from the HIVl-IIIB strain and was 95% pure. To induce apoptosis, cells were incubated with 5 ng of HIV-1 (incubation for 18 hours, two washes, and test for apoptosis after 2 to 6 hours; in such conditions no produc- tive viral replication occurred [32]) or with 25 to 250 pM gp120 for different lengths of time (time range tested, 1-24 hours), and apoptosis was revealed by the terminal dUTP nick-end labeling (TUNEL) technique (Apoptag, Appligene, Oncor, Gaithersburg, MD).
Biochemical Studies of Transduction Pathways PREPARATION OF CELLS. Twenty- to 40-day-old primary cultures were incubated in the presence of gp120 (dose range tested, 25 pM to 1 nM) in a 10% C0,-humidified incuba- tor at 37°C. Cells were then washed with cold phosphate- buffered saline containing 0.1 mM Na,VO, and lysed in a specific lysing buffer for each type of experiment. Insoluble material was removed by centrifugation at 15,000 g for 15 minutes at 4°C. The protein concentration of cell lysates was determinated in duplicate by the microassay method of Brad- ford (Bio-Rad protein assay). For pharmacological studies, we used the NMDA receptor antagonist (DL-2-amino-5- phosphonopentanoic acid or AP5, 50 pM, Tocris Cookson, Bristol, UK), an inhibitor of Ca2’ release from sarcoplasmic reticulum (dantrolene, 30 pM, Tocris Cookson) and the di- hydropyridine L-type Ca2+ channel antagonist nifedipine (50 p M , Sigma Chemical, St Louis, MO). These pharmaco- logical substances were applied 15 minutes before stimula- tion by gp120. To chelate extracellular Ca2’, 1 mM EGTA was added in the medium for 2 and 30 minutes.
848 Annals of Neurology Vol 42 No 6 December 1997

WESTERN BLOT ANALYSIS. After a 30-minute equilibration period in Krebs buffer (126 mM NaCI, 3.5 mM KCI, 1.2 mM NaH,P04, 1.3 mM MgCI,, 2.0 mM CaCI,, 10 mM glucose, 25 mM NaHCO,, pH 7.3) cells were treated and lysed in a 0.2% Triton X-100 lysing buffer (0.2% Triton X- 100 [wt/vol], 80 mM P-glycerophosphate, 200 mM EGTA, 15 mM MgCI,, 1 mM Na3V04, 1 mM phenyl- methylsulfonyl fluoride, 10 mg/ml aprotinin, 10 mglml leu- peptin). Equal amounts of protein from cell lysates were sep- arated in sodium dodecyl sulfate (SDS)/10% polyacrylamide gels and electrophoretically transferred to nitrocellulose membranes in Tris-glycine buffer with 20% methanol (wtivol). Membranes were blocked in Tris-buffered saline (10 mM Tris-HC1, pH 7.5, 150 mM NaCI) containing 0.05% Tween 20 (wdvol) and 0.25% gelatin (wdvol). The blots were then incubated with primary antibody (anti- phosphotyrosine monoclonal antibody [mAb]; PY20, sc-508, Santa Cruz Biotechnologies, Santa-Cruz, CA), anti-ERK mAb (sc-94, Santa Cruz, Biotechnologies), and a secondary horseradish peroxidase-linked antirabbit IgG. The blots were visualized by chemiluminescence (Amersham ECL system).
JNK A N D ERK KINASE ASSAYS. Plasmids encoding the fu- sion proteins glutathione-Stransferase (GST)-Elk(307-408) and GST-cJun (1-79) were transfected in BL21(Lys E) bac- teria. After induction with 1 mM isopropyl P-D- thiogalactopyranoside for 3 hours at 37°C the fusion pro- teins were purified by glutathione-Sepharose affinity chromatography (glutathione-Sepharose 4B, Pharmacia) as described [33]. The amount of protein was estimated by gel electrophoresis and Bradford quantification.
JNK kinase activity was measured by a solid-phase kinase assay [34]. In brief, cells were prepared and lysed as de- scribed above in a lysing buffer containing 0.2% Triton X- 100, 20 mM HEPES, pH 7.4, 300 mM NaCl, 2.5 mM MgCI,, 0.4 mM EDTA, 1 mM Na,V04, 1 mM dithiothre- itol (DTT), 20 mM pnitrophenyl phosphate (PNPP), 20 mM P-glycerophosphate, 1 mM 4-(2-amino- ethy1)benzenesulfonyl fluoride (AEBSF), and 2 pglml leu- peptin. Protein samples (100 pg) from cell lysates were di- luted fourfold with a buffer containing 20 mM HEPES, pH 7.4, 2.5 mM MgCl,, 20 mM P-glycerophosphate, 20 mM PNPP, 1 mM AEBSF, and 2 pglml leupeptin. The lysates were incubated for 2 hours at 4°C with 10 pl of GSH aga- rose suspension to which 10 mg of GST-c-Jun protein was bound. After incubation, protein samples were washed four times with buffer containing 20 mM HEPES, pH 7.4, 50 mM NaCI, 2.5 mM MgCI,, 0.1 mM EDTA, and 0.05% Triton X-100, and one last time with 1 ml of kinase buffer (20 mM HEPES, pH 7.4, 2.5 mM MgCI,, 20 mM 6- glycerophosphate, 20 mM PNPP, 0.1 mM Na,VO,, and 2 mM DTT). The pellet was resuspended in 20 pI of kinase buffer and the reaction was initiated by adding the substrate GST-c-Jun (1 pg/40 ml) and 3.7 pl [y-32P]ATP (2.5 pCi/40 1.1) (Amersham). After incubation at 30°C for 20 minutes, assays were terminated by the addition of 20 pl of 2X Laemmli sample buffer. The samples were boiled for 5 minutes at 95°C and analyzed by SDS/10% polyacrylamide gel electrophoresis. The gels were dried and subjected to au-
toradiography. The level of 32P incorporation into the sub- strates was then evaluated by a PhosphoImager.
ERK kinase activity was measured as described [35]. In brief, the cells were prepared and lysed as described above in a lysing buffer containing 1% Triton X-100, 20 mM Tris- acetate, pH 7.0, 1 mM EDTA, 1 mM EGTA (4Ht), 0.27 M Saccharose, 1 mM Na,V04, 10 mM P-glycerophosphate, 5 mM NaF, 5 mM sodium pyrophosphate, 1 mM benzami- dine, 4 pg/ml leupeptin, and 0.1% P-mercaptoethanol (wt/ vol). Protein samples containing 100 pg of protein were di- luted 10-fold with a buffer containing 50 mM Tris-HCI, pH 7.3, 2 mM Na,-EDTA, 2 mM EGTA, 100 p M Na,VO,, 100 FM benzamidine, 1 pg/ml leupeptin, and 1 mM DTT. ERK kinase activity was purified by anion-exchange chroma- tography on micro-Sepharose Q columns (Q-Sepharose Fast Flow, Pharmacia; minicolumn “Bio-Spin” 0.64 X 5 cm, Bio-Rad). The protein elution was performed in 500 pl of an elution buffer (0.4 M NaCI, 50 mM Tris-HCI, pH 7.3, 2 mM Na,-EDTA, 2 mM Na,-EGTA, 100 p M Na,VO,, 100 p M benzamidine, 1 pgiml leupeptin, and 1 mM DTT). A 20-pI fraction of purified sample was then mixed with 20 p1 of a buffer containing 40 mM HEPES, 30 mM MgCI,, 2 mM DTT, 40 mM PNPP, and 40 mM P-glycerophosphate. The reaction was initiated by adding an enzyme substrate (myelin basic protein [MBP] 10 pg/40 pl [Sigma] or GST- Elk protein 0.5 pg/40 pl) and [Y-~~P]ATP 2.5 pCi/40 p1 (Amersham). After incubation at 30°C for 20 minutes, assays were terminated by the addition of 10 pl of 5X Laemmli sample buffer. The samples were analyzed by SDS/10% polyacrylamide gel electrophoresis. Gels were then dried and subjected to autoradiography. Radioactivity was quantified with a PhosphoImager.
Results HIV-1 and Recombinant gpl20 Induce In Vitro Apoptosis of Human Neurons and Microglial Cells but Not of Astroytes To understand the mechanism of the previously ob- served gp 120-induced death in human embryonic neu- rons, we used the T U N E L technique to test for the presence of apoptotic cells in primary CNS cultures in- cubated with HIV-1 particles. Apoptotic cells were nu- merous in cultures labeled with TUNEL reagents 20 hours after the onset of incubation with HIV-1 parti- cles (15- to 38-day-old cultures, n = 3) , whereas the number of labeled cells in control cultures remained lower than one cell per field (Fig 1). The apoptotic cells were found mostly in the neuronal clusters but also in scattered foci in the monolayer containing as- trocytes and microglial cells, preventing evaluation of the percentage of positive cells. The labeled nuclei had either a normal appearance or were shrunken and pyk- notic. In some instances, the labeled cells showed frag- mentation of the nuclei into multiple apoptotic bodies. To test the involvement of gp120, primary CNS cul- tures were incubated with 250 pM gp120 and tested after 6 hours (n = 5) . Apoptotic cells were abundant in neuronal clusters and less frequent in the monolayer
Lannuzel et al: HIV-1-Induced Apoptosis in Human Neurons 849
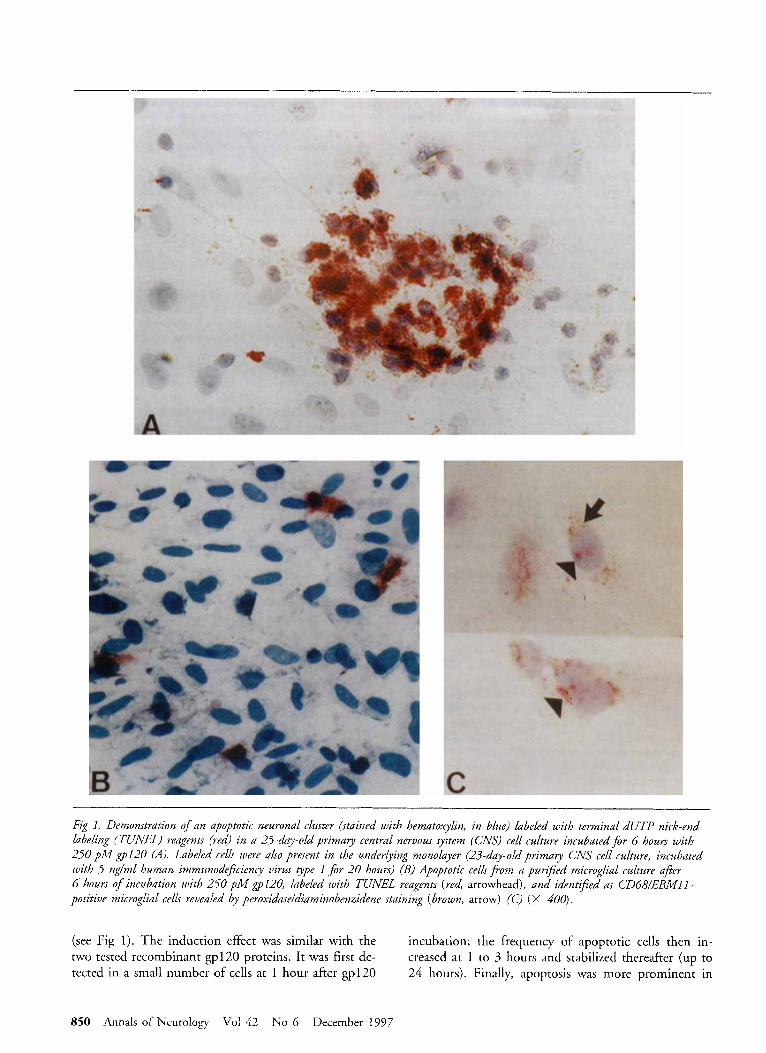
Fig I. Demonstration of an apoptotic neuronal cluster (stained with hernatoxylin, in blue) labeled with terminal dUTP nick-end labeling (TUNEL) reagents (red) in a 25-day-old primary central nervous system (CNS) cell culture incubatedfor 6 hours with 250 pM gp120 (A). Labeled cells were also present in the underlying monolayer (23-day-old primay CNS cell culture, incubated with 5 ng/ml human immunodeficiency virus qpe 1 for 20 hours) (B) Apoptotic cellsfiom a purzj5ed microglial culture after 6 hours o f incubation with 250 pM ~ 1 2 0 , labeled with TUNEL reagents (red arrowhead), and identified as CD68/EBMlI- positive microglial cells revealed by peroxidase/diaminobenzidene staining (brown, arrow) (C) (X 400).
(see Fig 1). The induction effect was similar with the two tested recombinant gp12O proteins. It was first de- tected in a small number of cells at 1 hour after gp120
incubation; the frequency of apoptotic cells then in- creased at I to 3 hours and stabilized thereafter (up to 24 hours). Finally, apoptosis was more prominent in
850 Annals of Neurology Vol 42 No 6 December 1997
cultures more than 15 days old than in cultures less than 7 days old (n = 3), suggesting that an in vitro maturation of embryonic cells was required for suscep- tibility to apoptosis.
The cell specificity of HIV-1- or gp120-induced ap- optosis was subsequently analyzed on purified cultures of astrocytes and microglial cells. Apoptosis was in- duced by HIV-1 in purified microglial cells (10-30% of positive cells after 6 hours; n = 4) or by gp120 (250 pM, 60-80% of positive cells after 6 hours; n = 4), whereas astrocytes appeared resistant (1% of apoptotic cells at 6 and 24 hours after HIV-1 incubation, n = 3, or gp120 incubation, n = 3). The resistance of astro- cytes was also supported by the detection, in double- staining experiments on HIV- 1-incubated primary cul- tures, of only 2% of the GFAP-positive cells, which were also labeled by TUNEL reagents. Apoptosis in microglial cells was detected with gpl2O concentrations as low as 25 pM (40-80% increase in the number of positive cells over the dose range tested, 25-250 pM; see Fig 1C for double staining). In sharp contrast with the induction of apoptosis in purified microglial cells, SV40-transformed microglial cells were resistant to gpl2O-induced apoptosis (50-250 pM, 6-hour delay; n = 3).
We also tested the effects of an NMDA antagonist, an L-type Ca2+ channel blocker, and an inhibitor of Ca”’ release from sarcoplasmic reticulum on the HIV-1- and gpl20-induced apoptosis. Apoptosis re- mained unchanged when primary cultures were treated
with AP5 (50 pM for 1 hour), nifedipine (50 and 100 FM for 1 hour), and dantrolene (30 pM, for 1 hour) (n = 3).
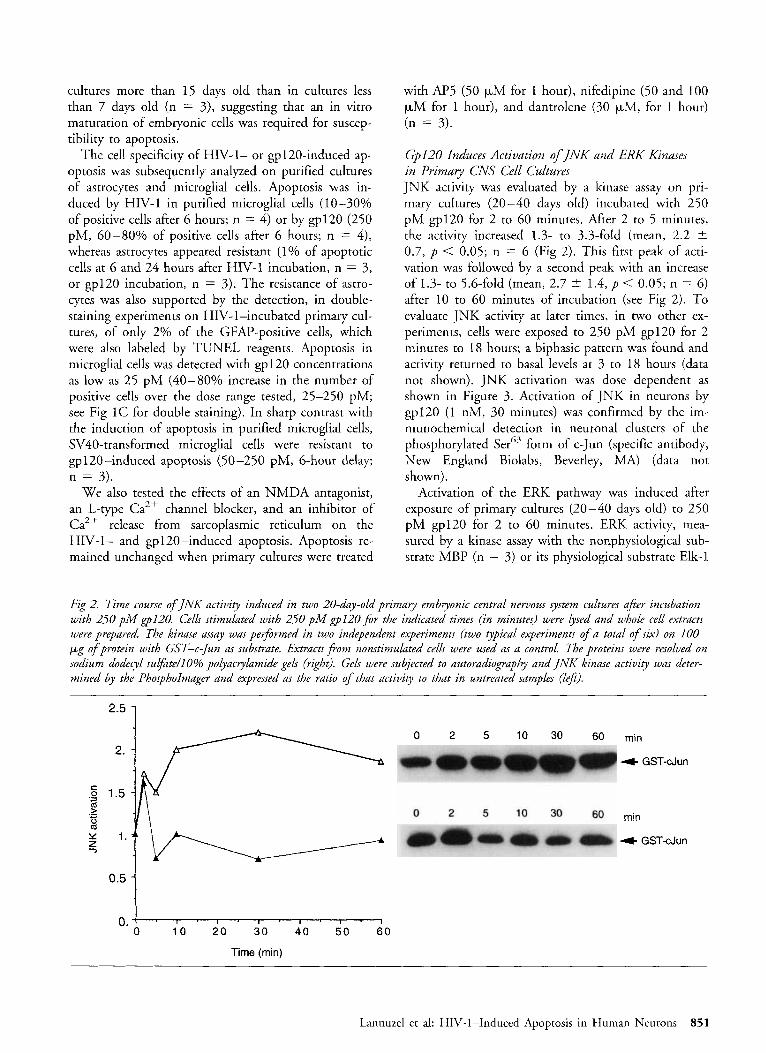
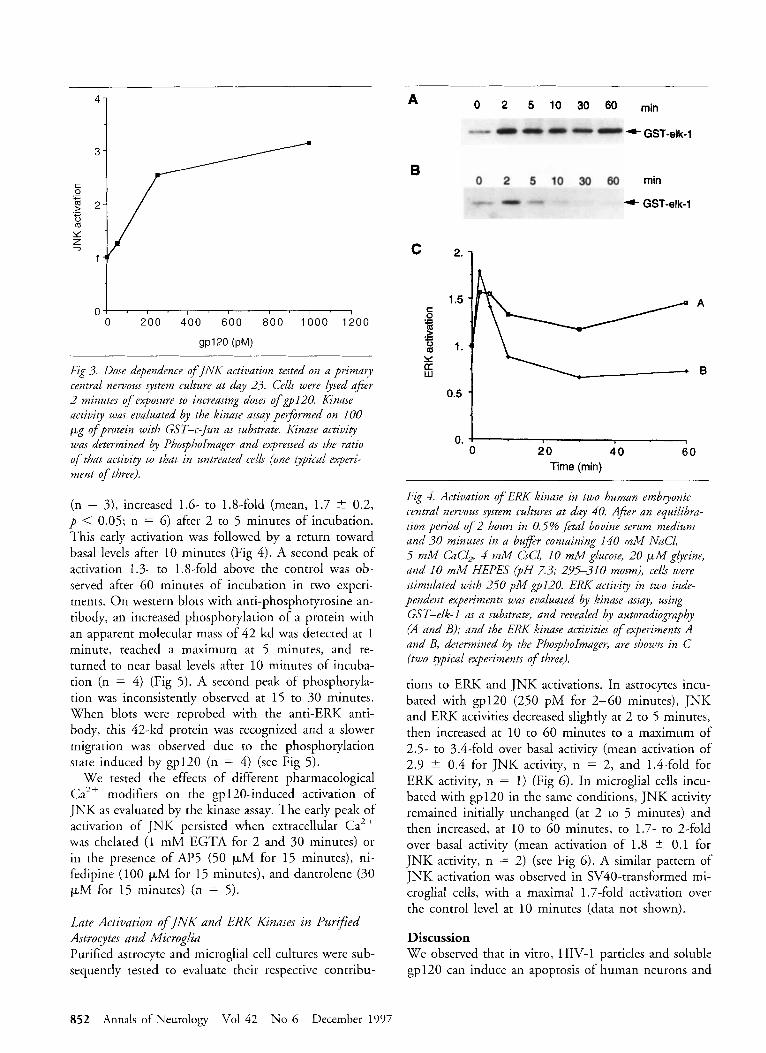
Gp120 Induces Activation of INK and ERK K znases in Primary CNS Cell Cultures JNK activity was evaluated by a kinase assay on pri- mary cultures (20-40 days old) incubated with 250 pM gp120 for 2 to 60 minutes. After 2 to 5 minutes, the activity increased 1.3- to 3.3-fold (mean, 2.2 2 0.7, p < 0.05; n = 6 (Fig 2). This fitst peak of acti- vation was followed by a second peak with an increase of 1.3- to 5.6-fold (mean, 2.7 t 1.4, p < 0.05; n = 6) after 10 to 60 minutes of incubation (see Fig 2). To evaluate JNK activity at later times, in two other ex- periments, cells were exposed to 250 pM gp120 for 2 minutes to 18 hours; a biphasic pattern was found and activity returned to basal levels at 3 to 18 hours (data not shown). JNK activation was dose dependent as shown in Figure 3. Activation of JNK in neurons by gp12O (1 nM, 30 minutes) was confirmed by the im- munochemical detection in neuronal clusters of the phosphorylated Serb3 form of c-Jun (specific antibody, New England Biolabs, Beverley, MA) (data not shown).
Activation of the ERK pathway was induced after exposure of primary cultures (20-40 days old) to 250 pM gp120 for 2 to 60 minutes. ERK activity, mea- sured by a kinase assay with the nonphysiological sub- strate MBP (n = 3) or its physiological substrate Elk-1
Fig 2. Time course of JNK activity induced in two 20-day-old primary embryonic central nervous system cultures afer incubation with 250 pM ~ 1 2 0 . Cells stimulated with 250 pM ~ 1 2 0 f i r the indicated times (in minutes) were lysed and whole cell extracts were prepared. The kinase assay wac performed in two independent experiments (two typical experiments of a total of six) on 100 pg o f protein with GST-c-jun a substrate. Extracts j?om nonstimukzted cells were used as a control. The proteins were resolved on sodium dodecyl sulfdte/lO% polyacrylamide gels (right). Gels were subjected to autoradiography and INK kinase activity was deter- mined by the Phospholmager and expressed as the ratio o f that activity to that in untreated samples (lef2).
0 2 5 10 30 60 min
4 GST-dun
min
4- GST-cJun
0 10 20 30 40 50 60
Time (min)
Lannuzel et al: HIV-1-Induced Apoptosis in Human Neurons 851
A 0 2 5 10 30 60 min
4- GST-elk-1
B rnin
* GST-elk-1
0 1 0 200 400 600 800 1000 1200
Fig 3. Dose dependence o f J N K activation tested on a primary central nervous system culture at day 23. Cells were lysed after 2 minutes of exposure to increasing doses o f ~ 1 2 0 . Kinase activity was evaluated by the kinase assay performed on 100 Fg of protein with GST-c-Jun as substrate. Kinase activity was determined by PhosphoImager and expressed as the ratio of that activity to that in untrPated cells (one gpical experi- ment of three).
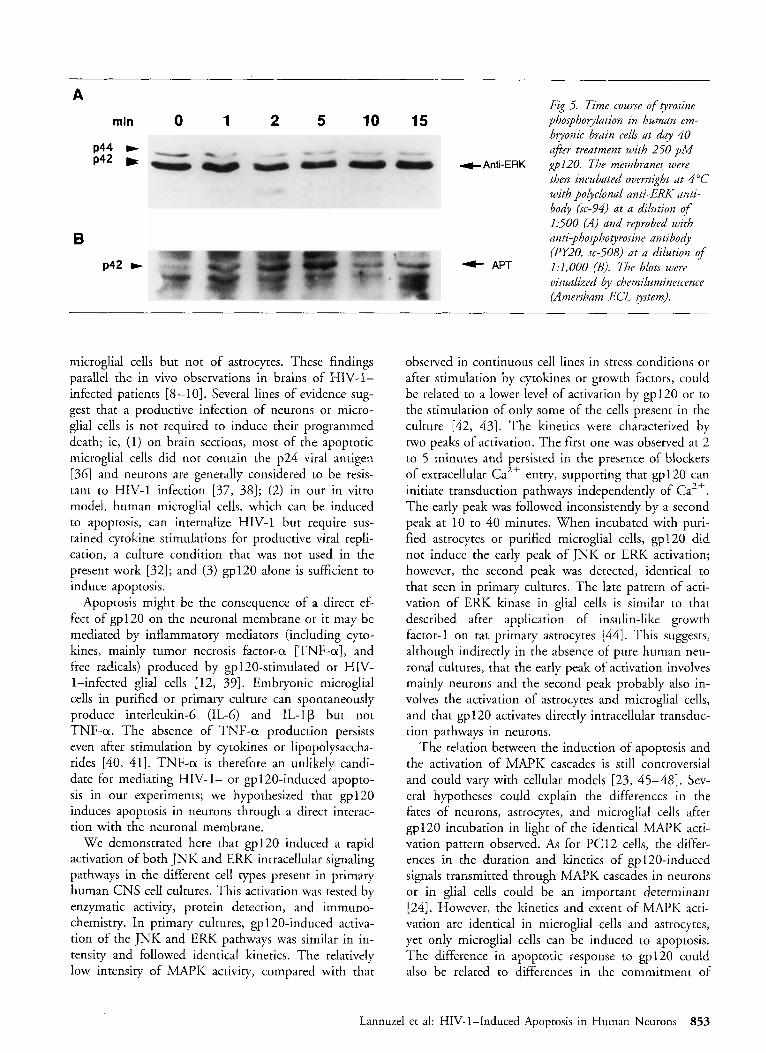
(n = 3), increased 1.6- to 1.8-fold (mean, 1.7 -+ 0.2, p < 0.05; n = 6) after 2 to 5 minutes of incubation. This early activation was followed by a return toward basal levels after 10 minutes (Fig 4). A second peak of activation 1.3- to 1.8-fold above the control was ob- served after 60 minutes of incubation in two experi- ments. O n western blots with anti-phosphotyrosine an- tibody, an increased phosphorylation of a protein with an apparent molecular mass of 42 kd was detected at 1 minute, reached a maximum at 5 minutes, and re- turned to near basal levels after 10 minutes of incuba- tion (n = 4) (Fig 5) . A second peak of phosphoryla- tion was inconsistently observed at 15 to 30 minutes. When blots were reprobed with the anti-ERK anti- body, this 42-kd protein was recognized and a slower migration was observed due to the phosphorylation state induced by gp120 (n = 4) (see Fig 5 ) .
We tested the effects of different pharmacological Ca2+ modifiers on the gp 120-induced activation of JNK as evaluated by the kinase assay. The early peak of activation of JNK persisted when extracellular Ca2+ was chelated (1 mM EGTA for 2 and 30 minutes) or in the presence of AP5 (50 pM for 15 minutes), ni- fedipine (100 pM for 15 minutes), and dantrolene (30 pM for 15 minutes) (n = 5) .
Late Activation of JNK and ERK Kinases in PurzJied Asrrocytes and Microglia Purified astrocyte and microglial cell cultures were sub- sequently tested to evaluate their respective contribu-
0.5 0. 0 20 4 0 60
Time (min)
Fig 4. Activation o f E M kinase in two human embryonic central nervous y t e m cultures at day 40. A$er an equilibra- tion period o f 2 hours in 0.5% fetal bovine serum medium and 30 minutes in a buffer containing 140 mM NaC1, 5 mM CaC12, 4 mM CsCl, 10 mM glucose, 20 p M glycine, and 10 mM HEPES (PH 7.3; 295-31 0 moswl), cells were stimulated with 250 p M ~ 1 2 0 . ERK activity in two inde- pendent experiments was evaluated by kinase assay, using GST-elk-1 as a substrate, and revealed by autoradiography (A and B); and the ERK kinase activities o f experiments A and B, determined by the PhosphoImager, are shown in C (two typical experiments of three).
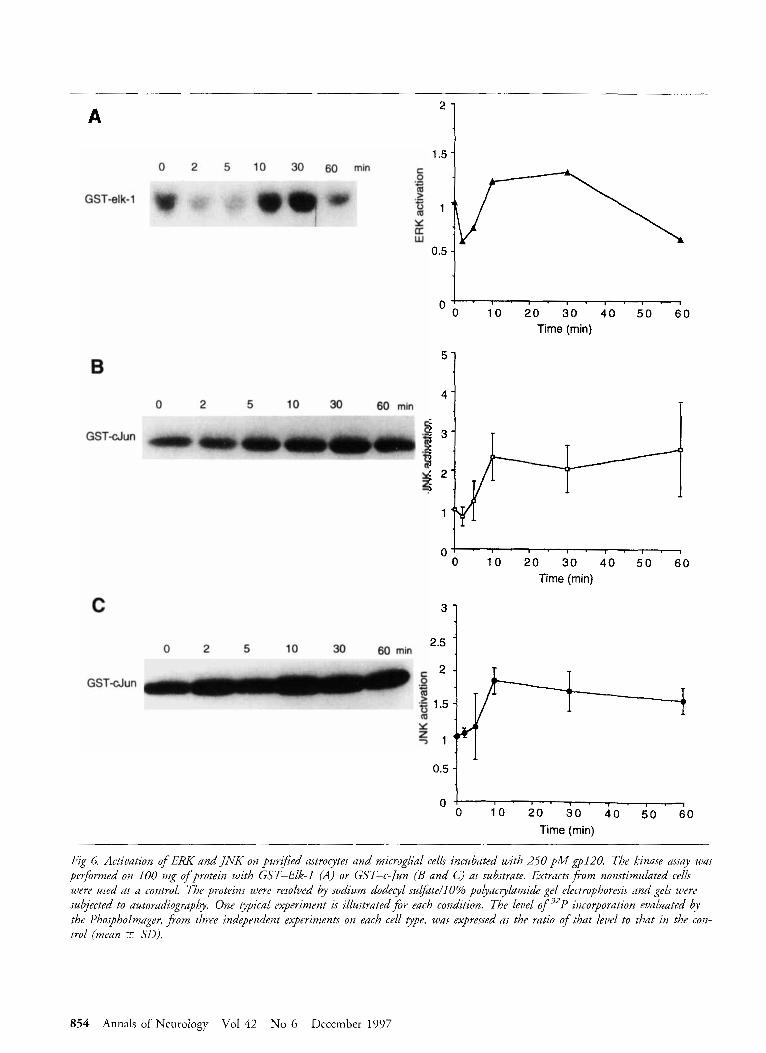
tions to ERK and JNK activations. In astrocytes incu- bated with gp120 (250 pM for 2-60 minutes), JNK and ERK activities decreased slightly at 2 to 5 minutes, then increased at 10 to 60 minutes to a maximum of 2.5- to 3.4-fold over basal activity (mean activation of 2.9 +- 0.4 for JNK activity, n = 2, and 1.4-fold for ERK activity, n = 1) (Fig 6). In microglial cells incu- bated with gp120 in the same conditions, JNK activity remained initially unchanged (at 2 to 5 minutes) and then increased, at 10 to 60 minutes, to 1.7- to 2-fold over basal activity (mean activation of 1.8 5 0.1 for JNK activity, n = 2) (see Fig 6 ) . A similar pattern of JNK activation was observed in SV40-transformed mi- croglial cells, with a maximal l .7-fold activation over the control level at 10 minutes (data not shown).
Discussion We observed that in vitro, HIV-1 particles and soluble gp120 can induce an apoptosis of human neurons and
852 Annals of Neurology Vol 42 No 6 December 1997
A
min 0 1 2 5 10 15
p44 * P42
B p42 +
Fig 5. Time course of tyrosine phosphoyllation in human em- bryonic brain cells at day 40 ajer treatment with 250 piw
+Anti-ERK ~ 1 2 0 . The membranes were then incubated overnight at 4°C with polyclonal anti-ERK anti- body (sc-94) at a dilution of 1500 (A) and reprobed with anri-phosphotyrosine antibody (PY20, sc-508) at a dilution of 1:1,000 (B). The blots were visualized by chemiluminescence (Amersham ECL system).
+ APT
microglial cells but not of astrocytes. These findings parallel the in vivo observations in brains of HIV-1- infected patients [&lo]. Several lines of evidence sug- gest that a productive infection of neurons or micro- glial cells is nor required to induce their programmed death; ie, (1) on brain sections, most of the apoptotic microglial cells did not contain the p24 viral antigen [36] and neurons are generally considered to be resis- tant to HIV-1 infection [37, 381; (2) in our in vitro model, human microglial cells, which can be induced to apoptosis, can internalize HIV-1 but require sus- tained cytokine stimulations for productive viral repli- cation, a culture condition that was not used in the present work [32]; and (3) gp120 alone is sufficient to induce apoptosis.
Apoptosis might be the consequence of a direct ef- fect of gp12O on the neuronal membrane or it may be mediated by inflammatory mediators (including cyto- kines, mainly tumor necrosis factor-a [TNF-a], and free radicals) produced by gpl20-stimulated or HIV- 1-infected glial cells [ 12, 391. Embryonic microglial cells in purified or primary culture can spontaneously produce interleukin-6 (IL-6) and IL-1p but not TNF-a. The absence of TNF-a production persists even after stimulation by cytokines or lipopolysaccha- rides [40, 411. TNF-a is therefore an unlikely candi- date for mediating HIV-1- or gpl20-induced apopto- sis in our experiments; we hypothesized that gp 120 induces apoptosis in neurons through a direct interac- tion with the neuronal membrane.
We demonstrated here that gp120 induced a rapid activation of both JNK and ERK intracellular signaling pathways in the different cell rypes present in primary human CNS cell cultures. This activation was tested by enzymatic activity, protein detection, and immuno- chemistry. In primary cultures, gp 120-induced activa- tion of the JNK and ERK pathways was similar in in- tensity and followed identical kinetics. The relatively low intensity of MAPK activity, compared with that
observed in continuous cell lines in stress conditions or after stimulation by cyrokines or growth factors, could be related to a lower level of activation by gp12O or to the stimulation of only some of the cells present in the culture [42, 431. The kinetics were characterized by two peaks of activation. The first one was observed at 2 to 5 minutes and persisted in the presence of blockers of extracellular Ca2+ entry, supporting that gp 120 can initiate transduction pathways independently of Ca2+. The early peak was followed inconsistently by a second peak at 10 to 40 minutes. When incubated with puri- fied astrocytes or purified microglial cells, gp12O did not induce the early peak of JNK or ERK activation; however, the second peak was detected, identical to that seen in primary cultures. The late pattern of acti- vation of ERK kinase in glial cells is similar to that described after application of insulin-like growth factor-1 on rat primary astrocytes [44]. This suggests, although indirectly in the absence of pure human neu- ronal cultures, that the early peak of activation involves mainly neurons and the second peak probably also in- volves the activation of astrocytes and microglial cells, and that gp 120 activates directly intracellular transduc- tion pathways in neurons.
The relation between the induction of apoptosis and the activation of MAPK cascades is still controversial and could vary with cellular models [23, 45-48]. Sev- eral hypotheses could explain the differences in the fates of neurons, astrocytes, and microglial cells after gp120 incubation in light of the identical MAPK acti- vation pattern observed. As for PC12 cells, the differ- ences in the duration and kinetics of gp 120-induced signals transmitted through MAPK cascades in neurons or in glial cells could be an important dererminant [24]. However, the kinetics and extent of MAPK acti- vation are identical in microglial cells and astrocytes, yet only microglial cells can be induced to apoptosis. The difference in apoptotic response to gp12O could also be related to differences in the commitment of
Lannuzel et al: HIV-1-Induced Apoptosis in Human Neurons 853
A
5'
4 -
0 2 5 10 30 60 min
'1 1.5 i
GST-elk-1
6
O 0 , 1 ' 0 ' i o ' 40. 4 0 ' 5 0 ' $ 0 Time (rnin)
C
2.5 0 2 5 10 30 60 min
GST-dun
0 0 10 20 30 40 50 60
Time (min)
Fig 6. Activation of ERK and JNK on purified astroeytes and microglial cells incubated with 250 p M $pl20. The kinase assay was performed on 100 mg ofprotein with GST-Elk-I (A) or GST-c-/un (B and C) as substrate. Extractsfvom nonstimulated cells were used as a control. The proteins were resolved by sodium dodecyl su@te/lO% polyacrylamide gel electrophoresis and gels were subjected to autoradiopapby. One typical experiment is illustrated for each condition. The level o f 32P incorporation evaluated by the Phospholmuger, fiom three independpnt experiments on each cell type, was expressed as the ratio of that level to that in the con- trol (mean -+ SO).
854 Annals of Neurology Vol 42 No 6 December 1997

cells [49]; a cell in a postmitotic state could respond to a signal by apoptosis, whereas an actively dividing cell could not [50]. In our culture conditions, astrocytes, which are resistant to apoptosis, have the highest rate of division, whereas microglial cells divide very slowly and neurons are postmitotic. The resistance of SV40- transformed microglial cells to gpl2O-induced apopto- sis also supports the relationship between induction of apoptosis and rate of division.
Neuronal apoptosis after HIV-1 contact may result from the combination of several intracellular signaling pathways. Biochemical studies in cultured primary hu- man CNS cells will allow evaluation of the differences in apoptosis induction between neuronal and glial cells.
This study was supported by grants from the Agence Nationale de Recherche sur le Sida, Institut National de la Santt et de la Recher- che Medicale, and Universiti: Paris XI.
We thank Dr C. Maignier for his support of our studies through a long collaboration in tissue collection. We thank, for the gifts of reagents, Dr N. Haigwood of Chiron Corporation for gp120 from SF2 and Dr S. Zolla-Pazner for monoclonal antibody specific to HIV-1 V3, which was obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH, B. Derijard and R. Davis for the gift of the pGEX-c-Jun (1-79) plas- mid, and R. Hipskind for the gift of the pGEX-Elk(307-428) plas- mid. We also thank Dr R. Fagard and Dr P. Vernier for helpful discussions, L. Wingertsmann for expert technical assistance, M. Grelier for secretarial assistance, and J. P. Bouillot for help with the photography.
References 1.
2.
3.
4.
5 .
6.
7.
8.
9.
Dal Pan GJ, McArthur JH, Aylward E, et al. Patterns of cere- bral atrophy in HIV-1-infected individuals: results of a quanti- tative MRI analysis. Neurology 1992;42:2125-2130 McArthur JC, Hoover DR, Bacellar H, et al. Dementia in AIDS patients: incidence of risk factors. Neurology 1993;43: 2245-2252 Meyerhoff DJ, MacKay S, Bachman L, et al. Reduced brain N-acetylaspartate suggests neuronal loss in cognitively impaired human immunodeficiency virus-seropositive individuals: in vitro ' H magnetic resonance spectroscopy imaging. Neurology
Paley M, Cozzone PJ, Alonso J, et al. A multicenter proton magnetic resonance spectroscopy study of neurological compli- cations of AIDS. AIDS Res Hum Retroviruses 1996;12:213- 22 1 Everall IP, Luthert PJ, Lantos PL. Neuronal loss in the frontal cortex in HIV infection. Lancet 1991;337:1119-1121 Weis S, Haug H, Budka H . Neuronal damage in the cerebral cortex of AIDS brains: a morphometric study. Acta Neuro- pathol 1993;85:185-189 Masliah E, Ge N, Achim CL, et al. Selective neuronal vulner- ability in HIV encephalitis. J Neuropathol Exp Neurol 1992;
Adle-Biassette H , Levy Y, Colombel M, et al. Neuronal apop- tosis in HIV infection in adults. Neuropathol Appl Neurobiol 1995;21:218-227 Gelbard HA, James H, Sharer L, et al. Apoptotic neurons in brains of pediatric patients with HIV-1 encephalitis and pro- gressive encephalopathy. Neuropathol Appl Neurobiol 1995;2 1 : 208 -2 17
1993;43:509-5 15
5 1~585-593
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22. 23.
24.
25.
26.
27.
28.
29.
30
31
Lannuzel et
Petito CK, Roberts B. Evidence of apoptotic cell death in HIV encephalitis. Am J Pathol 1995;146:1121-1130 Epstein LG, Gendelman HE. Human immunodeficiency virus type-1 infection of the nervous system: pathogenic mechanisms. Ann Neurol 1993;33:429-436 Gendelman HE, Lipton SA, Tardieu M, et al. The neuropatho- genesis of HIV-I infection. J Leukoc Biol 1994;56:589-598 Brenneman DE, Westbrook GL, Fitzgerald SP, et al. Neuronal cell killing by the envelope protein of HIV and its prevention by vasoactive intestinal peptide. Nature 1988;335:639-642 Dreyer EB, Kaiser PK, Offermann JT, Lipton SA. HIV-1 coat protein neurotoxicity prevented by calcium channel antagonists. Science 1990;248:364-367 Lipton SA. Calcium channel antagonists and human immuno- deficiency virus coat protein-mediated neuronal injury. Ann Neurol 199 1;30: 1 10-1 14 Lipton SA. Synergistic effects of HIV coat protein and NMDA receptor-mediated neurotoxicity. Neuron I991;7:111-118 Muller WEG, Schroder HC, Ushijima H , et al. Gp120 of HIV-1 induces apoptosis in rat cortical cell cultures; prevention by memantine. Eur J Pharmacol 1992;226:209-214 Dawson VL, Dawson TM, Uhl GR, Snyder SH. Human im- munodeficiency virus type 1 protein neurotoxicity mediated by nitric oxide in primary cortical cultures. Proc Natl Acad Sci USA 1993;90:3256-3259 Lee MR, H o DD, Gurney ME. Functional interaction and par- tial homology between human immunodeficiency virus and neuroleukin. Science 1987;237: 1047-105 1 Perovic S, Schleger C, Pergande G, et al. The triaminopyridine flupirtine prevents cell death in rat cortical cells induced by N-methyl-D-aspartate and gp120 of HIV-1. Eur J Pharmacol 1994;288:27-33 Lannuzel A, Lledo PM, Lamghitnia HO, et al. HIV-1 envelope proteins gp120 and gpl60 potentiate NMDA-induced [Ca2+], increase, alter [Ca*+], homeostasis and induce neurotoxicity in human embryonic neurons. Eur J Neurosci 1995;7:2285-2296 Bredesen DA. Neural apoptosis. Ann Neurol 1995;38:839-851 Xia Z, Dickens M, Raingeaud J, et al. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995;l: 1326-133 1 Seger R, Krebs EG. Rgz MAPK signalling cascade. FASEB J 1995;9:726-735 Minden A, Lin A, McMahon M, et al. Differential activation of ERK and JNK mitogen-activated protein kinases by Raf-l and MEKK. Science 1994;266:1719-1722 Peudenier S, Hi:ry C, Monragnier L, et al. Human microglial cells: characterization in cerebral tissue and in primary culture, and study of their susceptibility to HIV-1 infection. Ann Neu- rol 1991;29:152-161 Janabi N, Chabrier S, Tardieu M. Endogenous nitric oxide ac- tivates prostaglandin F2u production in human microglial cells but not in astrocytes. J Immunol 1996;157:2129-2135 Janabi N, Peudenier S, Heron B, et al. Establishment of human microglial cell lines after transfection of primary cultures micro- glial cells with the SV40 large T antigen. Neurosci Lett 1995;
Levy JA, Hoffman AD, Kramer SM, er al. Isolation of lympho- cytopathic retrovirus from San Francisco patients with AIDS. Science 1984;225:840-842 Haigwood NL, Barker CB, Higgins KW, et al. Evidence for neutralizing antibodies directed against conformational epitopes of HIV-1 gp120. In vaccines 90, Cle-step purification of polypeptides expressed in Escbericbia coli as fusions with glutathione-S-transferase. Gene 1988;67:31-40 Sanchez-Pescador R, Power MD, Barr PJ, et al. Nucleotide se- quence and expression of an AIDS-associated retrovirus (ARV- 2). Science 1985;227:484-492
195: 105-108
al: HIV-1-Induced Apoptosis in H u m a n Neurons 855

32. Janabi N, di Stefano M, Chiodi F, et al. The susceptibility of human microglial cells to HIVl depends on the state of differentiation: its modulation by IFNy and TNFa. J Neuro- virol 1996;2:16 (Abstract)
33. Smith SB, Johnson KG. Single-step purification of polypeptides expressed in Escherichia coli as fusions with glutathione-S- transferase. Gene 1988;67:3 1-40
34. Hibi M, Lin A, Minden A, et al. Identification of an oncoprotein- and UV-responsive protein kinase that binds and potentiates the c-Jun activation domain. Genes Dev 1993;7: 2135-2148
35. Chabre 0, Cornillon F, Bottari SP, et a]. Hormonal regulation of mitogen-activated protein kinase activity in bovine adenocor- tical cells: cross-talk between phosphoinosides, adenosine 3'5'- monophosphate, and tyrosine kinase receptor pathways. Endo- crinology 1995; 136:956 -964
36. Krajewski S, James HJ, Ross J, et al. Expression of pro- and anti-apoptosis gene products in brains from pediatric patients with HIV-1 encephalitis. Neuropathol Appl Neurobiol 1997; 23:242-253.
37. Gabuzda DH, Ho DD, de la Monte SM, et al. Imniunohisto- chemical identification of HTLV-Ill antigen in brains of pa- tients with AIDS. Ann Neurol 1986;20:289-295
38. Takahashi K, Wesselingh SL, Griffin DE, et al. Localization of HIV-1 in human brain using polymerase chain reactiodin situ hybridization and immunocytochemistry. Ann Neurol 1996;39:
39. Yeung MC, Pulliam L, Lau AS. The HIV envelope protein gp120 is toxic to human brain-cell cultures through the induc- tion of interleukin-6 and tumor necrosis factor-a. AIDS 1995; 9: 137-143
40. SCbire G, Emdie D, Wallon C, et al. In vitro production of IL-6, and tumor necrosis factor-a by human embryonic inicroglial and neural cells. J Immunol 1993;150:1517-1523
705-71 1
41. Janabi N, Mirshahi A, Wolfrom C. et al. Effect of interferon y and TNFa on the differentiation/activation of human glid cells: implication for the TNFa receptor 1. RKS Virol 1996;147: 147-153
42. Canipos-Gonzales R, Kindy MS. Tyrosine phosphorylation of the microtubule-associated protein lilnase after transient ischemia in the gerbil brain. J Neurochem 1992;59:1955-1958
43. Murphy TH, Blatter LA, Bhat RV, et al. Differential regulation of calcium/calmodulin-dependent protein kinase 11 and p42 MAP kinase activity by synaptic transmission. J Neurosci 1994; 14:1320-1331
44. Tournier C, Pomerance M, Gavaret JM, et al. MAP kinase cas- cade in astrocytes. Glia 1994;10:81-88
45. Liu Z, Hsu H, Goeddel DV, et al. Dissection of TNFa recep- tor 1 effector functions: JNK activation is not linked to apop- tosis while NF-KB activation prevents cell death. Cell 1996;87:
46. Lenczowscki JM, Dominguez L, Eder AM, et al. Lack of a role for Jun kinase and AP-1 in fas-induced apoptosis. Mol Cell Biol 1997;17: 170-181
47. Verheij M, Base R, Lin XH, et a[. Requirement for ceraniide- initiated SAPK/JNK signaling in stress-induced apoptosis. Na- ture 1996;380:75-79
48. Heady LE, Storey B, Fanger GR, et al. GTPase deficient Gal , and Ga, induce PC12 differentiation and persistent activation of c-Jun NH,-terminal kinases. Mol Cell Biol 1996;16:648- 658
49. Kranenburg 0, van der Eb AJ, Zantema A. Cyclin D1 is an essential mediator of apoptotic neuronal cell death. EMBO J 1996; 1 5:46-54
50. Ross ME. Cell division and the nervous system: regulating the cycle from neural differentiation to death. Trends Neurosci 1996;19:62-68
565-576
856 Annals of Neurology Vol 42 No 6 December 1997
Related Documents











![Human Mitogen-activated Protein Kinase Kinase 4 as a ......(CANCERRESEARCH57. 4177—4182,October 1, 1997] Advances in Brief Human Mitogen-activated Protein Kinase Kinase 4 as](https://static.cupdf.com/doc/110x72/6082557b7810d746a5071f39/human-mitogen-activated-protein-kinase-kinase-4-as-a-cancerresearch57.jpg)