JOURNAL OF CLINICAL MICROBIOLOGY, 0095-1137/01/$04.0010 DOI: 10.1128/JCM.39.6.2178–2183.2001 June 2001, p. 2178–2183 Vol. 39, No. 6 Copyright © 2001, American Society for Microbiology. All Rights Reserved. Human Babesiosis in Japan: Isolation of Babesia microti-Like Parasites from an Asymptomatic Transfusion Donor and from a Rodent from an Area Where Babesiosis Is Endemic QIANG WEI, 1 MASAYOSHI TSUJI, 1 * AYA ZAMOTO, 1 MASATOSHI KOHSAKI, 2 TOSHIMITSU MATSUI, 3 TSUNEZO SHIOTA, 4 SAM R. TELFORD III, 5 AND CHIAKI ISHIHARA 1 School of Veterinary Medicine, Rakuno-Gakuen University, Ebetsu 069-8501, 1 Hoygo Red Cross Blood Center, Kobe 651-0062, 2 Third Division of the Department of Medicine, Kobe University School of Medicine, Kobe 650-0017, 3 and Kyoto Prefectural University of Medicine, Kyoto 602-8566, 4 Japan, and Harvard School of Public Health, Boston, Massachusetts 02115 5 Received 16 January 2001/Returned for modification 20 March 2001/Accepted 2 April 2001 To determine the source of infection for the Japanese index case of human babesiosis, we analyzed blood samples from an asymptomatic individual whose blood had been transfused into the patient. In addition, we surveyed rodents collected from near the donor’s residence. Examination by microscopy and PCR failed to detect the parasite in the donor’s blood obtained 8 months after the donation of the blood that was transfused. However, we were able to isolate Babesia parasites by inoculating the blood sample into SCID mice whose circulating red blood cells (RBCs) had been replaced with human RBCs. A Babesia parasite capable of propagating in human RBCs was also isolated from a field mouse (Apodemus speciosus) captured near the donor’s residential area. Follow-up surveys over a 1-year period revealed that the donor continued to be asymptomatic but had consistently high immunoglobulin G (IgG) titers in serum and low levels of parasitemia which were microscopically undetectable yet which were repeatedly demonstrable by inoculation into animals. The index case patient’s sera contained high titers of IgM and, subsequently, rising titers of IgG antibodies, both of which gradually diminished with the disappearance of the parasitemia. Analysis of the parasite’s rRNA gene (rDNA) sequence and immunodominant antigens revealed the similarity between donor and patient isolates. The rodent isolate also had an rDNA sequence that was identical to that of the human isolates but that differed slightly from that of the human isolates by Western blot analysis. We conclude that the index case patient acquired infection by transfusion from a donor who became infected in Japan, that parasitemia in an asymptomatic carrier can persist for more than a year, and that A. speciosus serves as a reservoir of an agent of human babesiosis in Japan. Human babesiosis due to Babesia microti (25, 26), a hemo- sporidian of small rodents, has increasingly been reported wherever Lyme borreliosis (11) and human granulocytic ehr- lichiosis (16) are endemic. The three zoonotic agents share a life cycle that depends on rodents and a tick of the Ixodes persulcatus species complex, and this association of microbes seems to be widely distributed through the temperate zone (26). Although B. microti has long been known to parasitize rodents in northern North America (4, 6, 24, 30), Europe (7, 29), and Eurasia (22, 28), cases of parasitization of humans have been reported almost exclusively from the United States (5, 25). Cases have recently been documented in Taiwan (21) and Japan (13). Increased awareness will facilitate the detec- tion of B. microti babesiosis in areas where the protozoon is enzootic. We have previously reported on the Japanese index case of human babesiosis, which occurred in 1999 at Kobe City in Hyogo Prefecture, Japan (13). The patient had received a blood transfusion before the onset of babesiosis. Of the eight blood donors involved in the transfusion, only a single individ- ual was found to be positive for specific antibody and parasite DNA, suggesting that the patient was infected via a transfusion of blood from this asymptomatic donor (20). Since neither the patient nor the donor had a history of travel abroad, the in- fection had presumably originated within Japan. Although no human case of babesiosis was reported in the country until 1999, human infections might simply have been undetected for many years. Indeed, more than 15 years ago, Shiota et al. (22) documented the presence of B. microti-like parasites in Japa- nese field mice (Apodemus speciosus), demonstrating that babesiosis has long been enzootic among Japanese wild ro- dents. However, because the original parasite described in their report is no longer available, the identity between that strain and the Kobe strain, which was isolated from the index case patient in our previous study (20), is not known. The objective of the present study was to determine the source of infection for the Japanese index case of human babesiosis. Using SCID mice whose circulating red blood cells (RBCs) had been replaced with those of humans (designated hu-RBC-SCID mice) (27), we were able to isolate Babesia parasites not only from the asymptomatic blood donor but also from a field mouse captured near the donor’s residential area. The antigenic and genotypic characteristics of these isolates were compared to those from the patient, confirming that B. microti is enzootic and zoonotic in Japan. * Corresponding author. Mailing address: School of Veterinary Medicine, Rakuno-Gakuen University, 582-1 Bunkyodai-Midorima- chi, Ebetsu 069-8501, Japan. Phone and fax: 81-11-386-3144. E-mail: [email protected]. 2178 on August 22, 2018 by guest http://jcm.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JOURNAL OF CLINICAL MICROBIOLOGY,0095-1137/01/$04.0010 DOI: 10.1128/JCM.39.6.2178–2183.2001
June 2001, p. 2178–2183 Vol. 39, No. 6
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Human Babesiosis in Japan: Isolation of Babesia microti-LikeParasites from an Asymptomatic Transfusion Donor and from a
Rodent from an Area Where Babesiosis Is EndemicQIANG WEI,1 MASAYOSHI TSUJI,1* AYA ZAMOTO,1 MASATOSHI KOHSAKI,2
TOSHIMITSU MATSUI,3 TSUNEZO SHIOTA,4 SAM R. TELFORD III,5 AND CHIAKI ISHIHARA1
School of Veterinary Medicine, Rakuno-Gakuen University, Ebetsu 069-8501,1 Hoygo Red Cross Blood Center,Kobe 651-0062,2 Third Division of the Department of Medicine, Kobe University School of Medicine,
Kobe 650-0017,3 and Kyoto Prefectural University of Medicine, Kyoto 602-8566,4 Japan,and Harvard School of Public Health, Boston, Massachusetts 021155
Received 16 January 2001/Returned for modification 20 March 2001/Accepted 2 April 2001
To determine the source of infection for the Japanese index case of human babesiosis, we analyzed bloodsamples from an asymptomatic individual whose blood had been transfused into the patient. In addition, wesurveyed rodents collected from near the donor’s residence. Examination by microscopy and PCR failed todetect the parasite in the donor’s blood obtained 8 months after the donation of the blood that was transfused.However, we were able to isolate Babesia parasites by inoculating the blood sample into SCID mice whosecirculating red blood cells (RBCs) had been replaced with human RBCs. A Babesia parasite capable ofpropagating in human RBCs was also isolated from a field mouse (Apodemus speciosus) captured near thedonor’s residential area. Follow-up surveys over a 1-year period revealed that the donor continued to beasymptomatic but had consistently high immunoglobulin G (IgG) titers in serum and low levels of parasitemiawhich were microscopically undetectable yet which were repeatedly demonstrable by inoculation into animals.The index case patient’s sera contained high titers of IgM and, subsequently, rising titers of IgG antibodies,both of which gradually diminished with the disappearance of the parasitemia. Analysis of the parasite’s rRNAgene (rDNA) sequence and immunodominant antigens revealed the similarity between donor and patientisolates. The rodent isolate also had an rDNA sequence that was identical to that of the human isolates but thatdiffered slightly from that of the human isolates by Western blot analysis. We conclude that the index casepatient acquired infection by transfusion from a donor who became infected in Japan, that parasitemia in anasymptomatic carrier can persist for more than a year, and that A. speciosus serves as a reservoir of an agentof human babesiosis in Japan.
Human babesiosis due to Babesia microti (25, 26), a hemo-sporidian of small rodents, has increasingly been reportedwherever Lyme borreliosis (11) and human granulocytic ehr-lichiosis (16) are endemic. The three zoonotic agents share alife cycle that depends on rodents and a tick of the Ixodespersulcatus species complex, and this association of microbesseems to be widely distributed through the temperate zone(26). Although B. microti has long been known to parasitizerodents in northern North America (4, 6, 24, 30), Europe (7,29), and Eurasia (22, 28), cases of parasitization of humanshave been reported almost exclusively from the United States(5, 25). Cases have recently been documented in Taiwan (21)and Japan (13). Increased awareness will facilitate the detec-tion of B. microti babesiosis in areas where the protozoon isenzootic.
We have previously reported on the Japanese index case ofhuman babesiosis, which occurred in 1999 at Kobe City inHyogo Prefecture, Japan (13). The patient had received ablood transfusion before the onset of babesiosis. Of the eightblood donors involved in the transfusion, only a single individ-
ual was found to be positive for specific antibody and parasiteDNA, suggesting that the patient was infected via a transfusionof blood from this asymptomatic donor (20). Since neither thepatient nor the donor had a history of travel abroad, the in-fection had presumably originated within Japan. Although nohuman case of babesiosis was reported in the country until1999, human infections might simply have been undetected formany years. Indeed, more than 15 years ago, Shiota et al. (22)documented the presence of B. microti-like parasites in Japa-nese field mice (Apodemus speciosus), demonstrating thatbabesiosis has long been enzootic among Japanese wild ro-dents. However, because the original parasite described intheir report is no longer available, the identity between thatstrain and the Kobe strain, which was isolated from the indexcase patient in our previous study (20), is not known.
The objective of the present study was to determine thesource of infection for the Japanese index case of humanbabesiosis. Using SCID mice whose circulating red blood cells(RBCs) had been replaced with those of humans (designatedhu-RBC-SCID mice) (27), we were able to isolate Babesiaparasites not only from the asymptomatic blood donor but alsofrom a field mouse captured near the donor’s residential area.The antigenic and genotypic characteristics of these isolateswere compared to those from the patient, confirming thatB. microti is enzootic and zoonotic in Japan.
* Corresponding author. Mailing address: School of VeterinaryMedicine, Rakuno-Gakuen University, 582-1 Bunkyodai-Midorima-chi, Ebetsu 069-8501, Japan. Phone and fax: 81-11-386-3144. E-mail:[email protected].
2178
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

MATERIALS AND METHODS
Human blood specimens. The detailed clinical course of the patient has beenreported elsewhere (13). Heparinized blood samples were obtained from thepatient, who provided informed consent, at the hospital of the Kobe UniversitySchool of Medicine. Some samples were obtained on clinically important days: 28December 1998 (admission and blood transfusion), 24 May 1999 (diagnosis andstart of treatment with quinine and clindamycin), 27 July 1999 (5-day follow-upafter discharge from hospital), 31 August 1999 (5 days after readmission due torecrudescence), and 11 November 1999 (4-week follow-up after the final dis-charge from the hospital). Blood samples from the asymptomatic donor and hisor her family members (father, brother, and sister) were obtained with informedconsent at the Hyogo Red Cross Blood Center of the Japanese Red CrossSociety.
Experimental animals. NOD/shi-scid mice (8), which were originally obtainedfrom the Central Institute of Experimental Animals, Kawasaki, Japan, weremaintained in the laboratory animal facility in Rakuno-Gakuen University.Golden Syrian hamsters and C.B-17 scid mice were purchased from SLC, Inc.(Hamamatsu, Japan) and Japan CLEA (Tokyo, Japan), respectively. All animalswere housed in isolators at temperatures between 22 and 25°C and were providedwith a g-ray-irradiated pellet diet and autoclaved tap water. All mice and ham-sters were splenectomized and used for experiments after the surgical woundshad healed completely. Animal experimentation was carried out according to theLaboratory Animal Control Guidelines of Rakuno-Gakuen University.
Parasite isolation. The blood samples from the patient, the asymptomaticdonor, and the field mice were washed with phosphate-buffered saline (PBS; pH7.2) by repeated centrifugation, and aliquots of RBCs were inoculated intohu-RBC-SCID mice or splenectomized hamsters for parasite isolation. To pre-pare hu-RBC-SCID mice, the peripheral RBCs in splenectomized NOD/shi-scidmice were replaced with human type O RBCs by the method described for ourprevious studies (17, 20). The rate of replacement with human RBCs in hu-RBC-SCID mice was monitored by flow cytometry (Cyto ACE150; JASCO Co., Tokyo,Japan) with biotin-labeled anti-human RBC mouse immunoglobulin G (IgG)Fab fragment (27) and phycoerythrin-labeled streptavidin (Life Technologies,Rockville, Md.) and was maintained at over 90% by repeatedly transfusing 0.5 mlof a packed cell volume of human RBCs (approximately 6 3 109) into the miceat 2- to 4-day intervals, together with administration of anti-mouse RBC mono-clonal rat antibody clone 2E11 (17) and anti-erythropoietin rabbit serum (17).Blood samples were collected from the tail veins of the inoculated animals, andGiemsa-stained thin-smeared blood films were prepared to microscopically de-termine the percent parasitemia. The isolates were further propagated by sub-passage into new animals. For the preparation of antigens or DNA, blood(parasitemia levels, 20 to 80%) was harvested by cardiocentesis from anesthe-tized animals, washed in PBS, and stored without cryopreservatives at 280°C.
Amplification and sequencing of parasite rDNA. Parasite DNAs were pre-pared from the frozen blood stocks described above with a whole-blood DNAextraction kit (GenTLE; TaKaRa Biochemical, Otsu, Japan). Sequences encod-ing the eukaryotic nuclear small-subunit rRNA gene (rDNA) were amplifiedfrom the DNA samples by PCR with the primer set described by Medlin et al.(15). The specific PCR products, approximately 1.8 kb, were cloned and se-quenced as described in our previous report (20). Nested PCR was used to detectparasites in human or rodent blood samples by a previously published protocol(19) except for minor modifications of the primers used. DNA samples wereprepared from 500 ml of the heparinized whole-blood specimens with a DNAExtractor WB kit (Wako Pure Chemical Industries, Osaka, Japan); 1/10 of eachspecimen was used for the first round of PCR with the primer set of Bab1A(59-GTCTTAGTATAAGCTTTTATACAGCG-39) and Bab4A (59-GATAGGTCAGAAACTTGAATGATACATCG-39), followed by the second round of PCRwith 1 ml of the first-round PCR product and the primer set of Bab2A (59-CAGTTATAGTTTATTTGATGTTCGTTTTAC-39) and Bab3A (59-CGGCAAAGCCATGCGATTCGCTAAT-39). The results of this nested PCR were con-firmed by another nested PCR done independently in a different laboratory withnewly designed primer sets: Bab5 (59-AATTACCCAATCCTGACACAGG-39)and Bab8 (59-TTTCGCAGTAGTTCGTCTTTAACA-39) for the first-round am-plification, followed by Bab6 (59-GACACAGGGAGGTAGTGACAAGA-39)and Bab7 (59-CCCAACTGCTCCTATTAACCATTAC-39) for the second-round amplification.
Antigenic analyses. The indirect immunofluorescent-antibody (IFA) test wascarried out by the method described previously (20), except that secondaryantibody was replaced with fluorescein isothiocyanate-conjugated affinity puri-fied goat IgG F(ab9)2 antibodies (anti-human IgM [heavy and light chains; IBL,Fujioka, Japan], anti-human IgG (Fc) [IBL], anti-human IgG, IgA, and IgM[Protos Immunoresearch, San Francisco, Calif.]). A mixture of fluorescein iso-
thiocyanate-conjugated anti-mouse IgG plus IgM and anti-rat IgG plus IgM(Jackson ImmunoResearch Laboratories, Inc., West Grove, Pa.) was used fordetection of antibodies in A. speciosus mouse serum. For the titration of specificIgM in human sera, samples were treated with protein G-conjugated Sepharose(Amersham Pharmacia Biotech, Uppsala, Sweden) to remove IgG antibodies.Western blot analysis was performed as described previously (2). Frozen stocksof Babesia-infected RBCs were thawed and washed five times at 4°C in 10 mMTris-HCl–10 mM EDTA (pH 7.5) by centrifugation at 10,000 3 g for 10 min. Theresulting pellets were resuspended in 125 mM Tris-HCl (pH 6.5) containing 5%b-mercaptoethanol, 2% sodium dodecyl sulfate, 10% glycerol, and 0.1% bromo-phenol blue, heated at 98°C for 5 min, and vigorously vortexed. The sampleswere diluted such that each contained material from equivalent numbers ofinfected RBCs and were subjected to sodium dodecyl sulfate-polyacrylamide gelelectrophoresis with 7.5% acrylamide gels. Proteins were electrophoreticallytransferred to Fluorotrans membranes (Pall BioSupport, Port Washington, N.Y.)for 1 h at 100 V. After being blocked with PBS containing 0.5% casein, themembranes were reacted with appropriately diluted immune sera, followed byreaction with secondary antibodies (alkaline phosphatase-conjugated AffiniPuregoat anti-mouse IgG [heavy and light chains] or anti-Syrian hamster IgG [heavyand light chains]; Jackson ImmunoResearch Laboratories, Inc.). Immunoreac-tive antigens were detected with a BCIP/NBT Alkaline Phosphatase SubstratesKit IV (Vector Laboratories, Inc., Burlingame, Calif.).
RESULTS
Isolation of parasites from an asymptomatic carrier. Afterestablishing the diagnosis of B. microti babesiosis in the indexcase patient, we initiated a search for the blood donors whocontributed to the units that had presumably infected the pa-tient. As reported previously (20), we identified one individualwhose serum contained antibody specific for B. microti. On 27July 1999, we were able to obtain a blood sample from theimplicated donor. The sample proved to be negative both bymicroscopy of a Giemsa-stained thin blood smear and bynested PCR specific for B. microti rDNA. However, we wereable to isolate Babesia parasites by inoculation of the bloodinto hu-RBC-SCID mice (Fig. 1) and into splenectomizedhamsters as well. It required approximately a month beforeparasitemia became detectable in the mice and the hamsters,indicating that the sensitivities of these experimental animalsfor parasite isolation were comparable and that the donor hada very low level of parasitemia. We also examined the bloodsamples from the donor’s family members (father, brother, andsister) for detection of parasite-specific antibody and parasiteDNA and for isolation of parasites by inoculation into ham-sters, but all examinations gave rise to negative results.
Parasite isolation from a field mouse. We set a total of 200Sherman live traps at various sites within approximately 7 kmof the donor’s residence and captured two field mice, both ofwhich were identified as A. speciosus. Microscopy revealed thatone of the mice was parasitemic, and its serum containedantibody specific for the Kobe strain. The other mouse wasnegative by examination of a blood smear and by an IFA test.The blood sample from the positive mouse was divided intothree portions and was inoculated into a hamster, an SCIDmouse, and an hu-RBC-SCID mouse. The level of parasitemiaincreased the fastest in the hu-RBC-SCID mouse, followed bythose in the hamster and the SCID mouse, in that order (Fig.2). Flow cytometric analysis with the parasitized RBCs ob-tained from the hu-RBC-SCID mouse revealed the presenceof parasites within human RBCs (data not shown).
Analysis of rDNA sequence. Approximately 1.8 kb of therDNA target sequence was obtained from the parasites iso-lated from the asymptomatic donor and from the Apodemus
VOL. 39, 2001 HUMAN BABESIOSIS IN JAPAN 2179
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from
mouse. Both sequences were exactly identical to that previous-ly reported for the Kobe strain (DDBJ accession no. AB032434).
Antigenic analysis. To determine whether the patient, do-nor, or mouse isolates might differ antigenically, we performedpairwise comparisons of their IFA titers. Whereas the serafrom both the patient and the donor demonstrated comparabletiters of antibodies against all the three parasite isolates (IFAtiters, 1:12,800 or 1:25,600 with both serum samples), theApodemus mouse serum showed a higher titer against homol-ogous parasite antigens than heterologous parasite antigens(IFA titers, 1:3,200 and 1:800, respectively). When the profilesof the parasite antigens were analyzed by Western blotting, theisolates from the patient and the donor were indistinguishable
but differed slightly from the Apodemus mouse-derived para-site (Fig. 3). The patient’s convalescent-phase sera mainlyrecognized three immunodominant proteins with estimatedmolecular masses of approximately 28, 97, and 140 kDa,respectively, whereas the donor’s sera recognized a much widervariety of additional antigens. Parasite antigens recognized bythe Apodemus mouse serum sample greatly differed from thoserecognized by the human serum sample.
Follow-up survey. Follow-up of the patient and the asymp-tomatic donor demonstrated that both remained chronicallyparasitemic and seroreactive (Table 1). Because we found thatthe presence of IgG antibody significantly interfered with de-
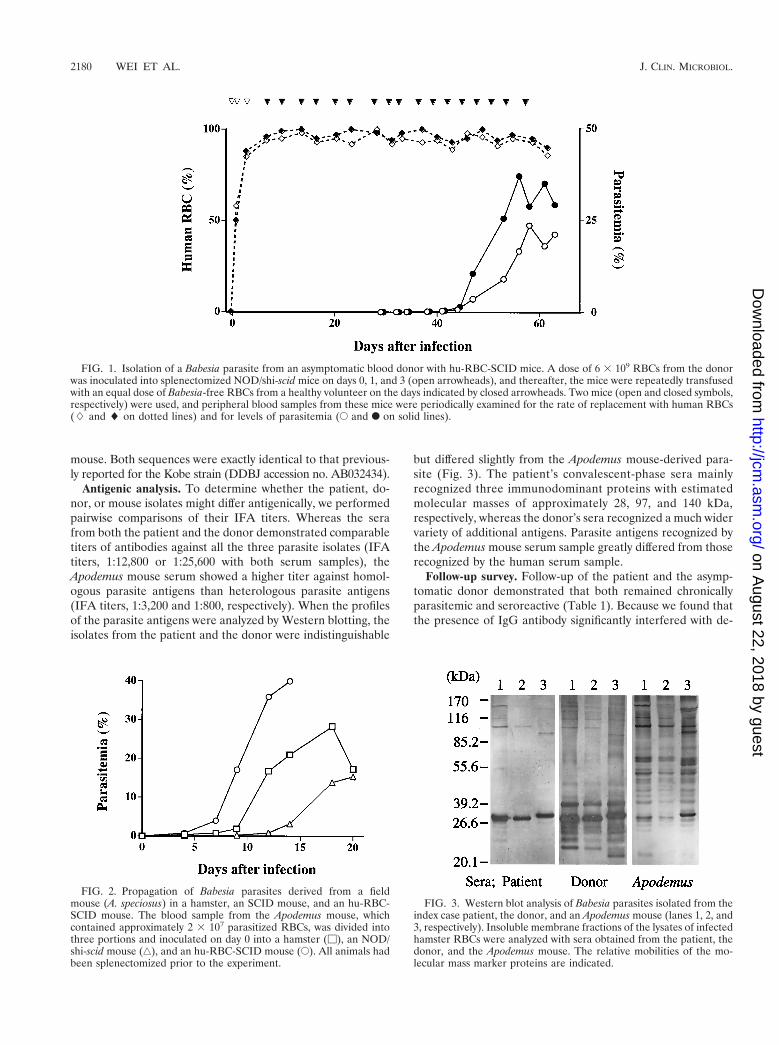
FIG. 1. Isolation of a Babesia parasite from an asymptomatic blood donor with hu-RBC-SCID mice. A dose of 6 3 109 RBCs from the donorwas inoculated into splenectomized NOD/shi-scid mice on days 0, 1, and 3 (open arrowheads), and thereafter, the mice were repeatedly transfusedwith an equal dose of Babesia-free RBCs from a healthy volunteer on the days indicated by closed arrowheads. Two mice (open and closed symbols,respectively) were used, and peripheral blood samples from these mice were periodically examined for the rate of replacement with human RBCs(L and l on dotted lines) and for levels of parasitemia (E and F on solid lines).
FIG. 2. Propagation of Babesia parasites derived from a fieldmouse (A. speciosus) in a hamster, an SCID mouse, and an hu-RBC-SCID mouse. The blood sample from the Apodemus mouse, whichcontained approximately 2 3 107 parasitized RBCs, was divided intothree portions and inoculated on day 0 into a hamster (M), an NOD/shi-scid mouse (‚), and an hu-RBC-SCID mouse (E). All animals hadbeen splenectomized prior to the experiment.
FIG. 3. Western blot analysis of Babesia parasites isolated from theindex case patient, the donor, and an Apodemus mouse (lanes 1, 2, and3, respectively). Insoluble membrane fractions of the lysates of infectedhamster RBCs were analyzed with sera obtained from the patient, thedonor, and the Apodemus mouse. The relative mobilities of the mo-lecular mass marker proteins are indicated.
2180 WEI ET AL. J. CLIN. MICROBIOL.
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

termination of the IgM titer by the IFA test, the serum sampleswere absorbed with protein G-Sepharose prior to the IgM titerdetermination. Throughout the survey period, the donor hadapparently been healthy and did not show any clinical symp-toms of babesiosis. This individual’s serum consistently con-tained high specific IgG titers. Babesia parasites were repeat-edly isolated from the donor’s blood by inoculation intosplenectomized hamsters, whereas detection of parasite DNAby rDNA-based PCR gave variable results. The patient hadhigh IgM titers at both the acute and the recrudescent phases.A rapid rise in IgG titer was observed after the recrudescentphase, which was followed by gradual decreases in both IgGand IgM titers, in accordance with the disappearance of par-asitemia. All the isolates obtained at various time points fromthe patient and the asymptomatic carrier exhibited virtuallyidentical banding profiles when the parasite antigens were an-alyzed by Western blotting (Fig. 4).
DISCUSSION
In the present study we have conclusively demonstrated thatthe Japanese index case of human babesiosis was acquired bya blood transfusion from an asymptomatic donor who acquiredthe infection in Japan. The parasite was isolated from theasymptomatic blood donor and was indistinguishable fromthose isolated from the index case patient by rDNA sequencingand immunodominant antigen profiles. A mouse trapped near
the residence of the donor was also infected with a geneticallyindistinguishable Babesia isolate. As we reported previously,neither the index case patient nor the donor had traveled fromJapan. Thus, there is no doubt that the index case was autoch-thonous, although the degree of endemicity appears to be low,as the donor’s family members were uninfected.
Because the donor denied prior exposure to ticks and toengaging in any relevant outdoor activities, exactly how he orshe became infected is unknown. However, the donor wasprobably infected around his or her residential area, inasmuchas a field mouse infected with B. microti-like parasites wasfound there and the parasites from the mouse and the donorhad identical rRNA sequences and similar antigenic proper-ties. This field mouse was identified as A. speciosus, whichShiota et al. (22) previously identified as being commonly in-fected with B. microti-like parasites in Shiga Prefecture. Thismouse has previously been identified as a main Japanese res-ervoir for Borrelia burgdorferi sensu lato (18), and it seemsprobable that it concurrently serves as a main reservoir of B.microti in Japan. The Babesia sp. in A. speciosus mice describedby Shiota et al. (22), however, may be different from thatobtained in the present study, because we recently found thata B. microti-like parasite isolated from a field mouse (A. spe-ciosus) captured in the town of Yamanaka in Shiga Prefecture,Japan, the same place where the prototype parasite was foundtwo decades ago, had a rDNA sequence (DDBJ accession no.
TABLE 1. Results of follow-up surveys of the patient and the donor
Sample source and date(yr-mo-day)
Detection of parasites Reciprocal IFA titers
Microscopya PCRb Inoculationc IgM IgG
Patient1998-12-28d NDe 2 ND ,100 ,1001999-05-24 1 1 1 12,800 1,6001999-06-25 2 1 ND 12,800 8001999-07-27 2 1 1 3,200 1,6001999-08-31 1 1 1 12,800 102,4001999-10-02 2 1 ND 12,800 51,2001999-11-01 ND 2 ND 1,600 25,6001999-11-29 ND 2 ND 800 25,6001999-12-27 ND 2 ND 400 6,4002000-01-29 ND ND ND 200 3,2002000-03-08 ND 2 ND 200 3,2002000-05-22 2 2 2 200 1,6002000-07-19 2 2 2 100 800
Donor1998-12-22 ND 1 ND 200 25,6001999-06-26 ND 2 ND 100 25,6001999-07-27 2 2 1 50 25,6001999-08-31 2 1 1 25 12,8001999-10-02 2 1 1 50 25,6001999-11-11 2 2 1 200 51,2001999-12-20 2 2 2 100 51,2002000-01-31 2 2 1 50 25,6002000-03-06 2 2 1 100 25,6002000-04-24 ND 1 ND ND ND
a Giemsa-stained thin-smeared blood films were microscopically observed; 1, parasitemia detectable; 2, not detectable.b Amplification by nested PCR. Results were confirmed in different laboratories by independent tests with different primers sets; 1, amplification positive; 2,
negative.c Inoculation into splenectomized hamster. Blood samples from the animals were observed for 2 months; 1, parasitemia detected; 2, not detected.d Because a blood sample from the patient was not available on this day, the results were estimated with extracts from the cross-match test papers made prior to blood
transfusion.e ND, not done.
VOL. 39, 2001 HUMAN BABESIOSIS IN JAPAN 2181
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

AB050732) which differed from those of the Kobe strain(DDBJ accession no. AB032434) and B. microti parasites in theUnited States (accession no. U09833) (M. Tsuji and T. Shiota,unpublished data). There therefore appears to be at least twotypes of indigenous B. microti-like parasites in Japan, and ep-idemiological studies of these parasites are ongoing. We areactively attempting to identify the tick vectors, regions of en-demicity, and prevalence of infection in wildlife and humans.
We were able to demonstrate that the hu-RBC-SCID mousemodel, which was developed in our previous study (20, 27),serves as an excellent experimental tool for isolation of Babesiaparasites from a human carrier with a very low level of para-sitemia. The sensitivity of this animal model was comparable tothat of splenectomized hamsters, which have previously beenreported to have the greatest susceptibility to B. microti (3).Moreover, the use of hu-RBC-SCID mice enabled us, for thefirst time, to demonstrate clearly that rodent-derived parasitesare readily infective for human RBCs. Because an in vitrocultivation system with human RBCs has not yet been madeavailable for B. microti, inoculation into the hu-RBC-SCIDmice is the sole method for assessment of the infectivity ofanimal-derived parasites for humans.
The dynamics of the specific antibody responses in the pa-tient and the asymptomatic donor provide evidence for pre-munition (concomitant immunity), first described during theseminal investigations of the mode of perpetuation of Texascattle fever due to Babesia bigemina (23). The index case pa-tient was discharged from the hospital in late July 1999, aftertreatment with quinine, clindamycin, and atovaquone. Amonth later, the patient experienced a recrudescent para-sitemia (13), which resulted in boosting of not only IgG titersbut also IgM titers. If recrudescence and reappearance of aspecific IgM response are common during human babesiosis,detection of a high serum IgM antibody titer may not neces-sarily be taken as evidence for a recently acquired primary
infection. Whereas the index case patient’s IgG titer dimin-ished with parasite clearance, the donor consistently had veryhigh IgG titers. This observation is consistent with premuni-tion, in which parasites persist despite a significant antibodyresponse; parasitemias are controlled to a very low level by theimmune response but generate enough antigenic stimuli tomaintain immunity. However, antigenic variation, which hasbeen suggested for cattle babesiosis (1), was not observed:virtually identical immunoblot profiles were seen with parasitesisolated at various times from both the patient and the asymp-tomatic donor.
Consistent with previous reports (10), our monitoring of theindex case patient and donor also demonstrated that para-sitemia may persist for an extended period, implying an asymp-tomatic carrier state. The donor described in the present studydid not show any clinical signs or symptoms of babesiosis, andthe level of parasitemia was so low that a highly sensitive test,such as nested PCR, often failed to detect it. Blood donatedfrom such individuals may therefore produce additional trans-fusion-associated infections. On the other hand, our ongoingepidemiological survey indicates that the prevalence of asymp-tomatic carriers in Japan is quite low (M. Tsuji, unpublisheddata). While effective chemotherapeutic protocols against hu-man babesiosis (9, 31, 32) have already been established, aneffective blood screening method for prevention of the trans-fusion-acquired infections has not yet been available even inthe regions in the United States where babesiosis is endemic(14), where over 20 transfusion-associated cases have beenreported (12). Practical measures that might be taken in Japanand at other locations where B. microti is endemic would be toinform physicians and patients about the risk of transfusion-acquired babesiosis and to develop a posttransfusion test forthe rapid, sensitive, and specific detection of Babesia parasites.
ACKNOWLEDGMENTS
We thank M. Ohtake, Y. Saito, and H. Murayama, Rakuno-GakuenUniversity, for excellent technical assistance.
This work was supported in part by grants-in-aid from the Minis-try of Education, Science and Culture of Japan (grants 11660316and 12450139) and by Gakujutsu-Frontier Cooperative Research atRakuno-Gakuen University.
REFERENCES
1. Allred, D. A., R. M. Cinque, T. J. Lane, and K. P. Ahrens. 1994. Antigenicvariation of parasite-derived antigens on the surface of Babesia bovis-in-fected erythrocytes. Infect. Immun. 62:91–98.
2. Arai, S., M. Tsuji, S.-J. Kim, K. Nakada, R. Kirisawa, M. Ohta, and C.Ishihara. 1998. Antigenic and genetic diversities of Babesia ovata in persis-tently infected cattle. J. Vet. Med. Sci. 60:1321–1327.
3. Brandt, F., G. R. Healy, and M. Welch. 1977. Human babesiosis: the isolationof Babesia microti in golden hamsters. J. Parasitol. 63:934–937.
4. Fay, F. H., and R. L. Rausch. 1969. Parasitic organisms in the blood ofarvicoline rodents in Alaska. J. Parasitol. 55:1258–1265.
5. Gorenflot, A., K. Moubri, E. Precigout, B. Carcy, and T. P. M. Schetters.1998. Human babesiosis. Ann. Trop. Med. Parasitol. 92:489–501.
6. Healy, G. R., A. Spielman, and N. Gleason. 1976. Human babesiosis: reser-voir of infection on Nantucket Island. Science 192:479–480.
7. Hussein, H. S. 1980. Ixodes trianguliceps: seasonal abundance and role in theepidemiology of Babesia microti infection in north-western England. Ann.Trop. Med. Parasitol. 74:531–539.
8. Koyanagi, Y., Y. Tanaka, J. Kira, M. Ito, K. Hioki, N. Misawa, Y. Kawano,K. Yamasaki, R. Tanaka, Y. Suzuki, Y. Ueyama, E. Terada, T. Tanaka, M.Miyasaka, T. Kobayashi, Y. Kumazawa, and N. Yamanoto. 1997. Primaryhuman immunodeficiency virus type 1 viremia and central nervous systeminvasion in a novel hu-PBL-immunodeficient mouse strain. J. Virol. 71:2417–2424.
9. Krause, P. J., T. Lepore, V. K. Sikand, J. J. Gadbaw, Jr., G. Burke, S. R.
FIG. 4. Western blot analysis of Babesia parasites isolated from thepatient and the donor at various sampling times. Isolates were ob-tained from the patient’s blood samples obtained on 24 May 1999, 27July 1999, and 31 August 1999 (lanes 1, 2, and 3, respectively) and fromthe donor’s blood samples obtained on 27 July 1999, 31 August 1999,2 October 1999, 11 November 1999, 31 January 2000, and 6 March2000 (lanes 4 through 9, respectively). Parasite antigens were immu-nostained with pooled sera obtained from the donor.
2182 WEI ET AL. J. CLIN. MICROBIOL.
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

Telford III, P. Brassard, D. Pearl, J. Azlanzadeh, D. Christianson, D.McGrath, and A. Spielman. 2000. Atovaquone and azithromycin for thetreatment of babesiosis. N. Engl. J. Med. 343:1454–1458.
10. Krause, P. J., A. Spielman, S. R. Telford III, V. K. Sikand, K. McKay, D.Christianson, R. J. Pollack, P. Brassard, J. Magera, R. Ryan, and D. H.Persing. 1998. Persistent parasitemia after acute babesiosis. N. Engl. J. Med.339:160–165.
11. Krause, P. J., S. R. Telford III, A. Spielman, V. Sikand, R. Ryan, D. Chris-tianson, G. Burke, P. Brassard, R. Pollack, J. Peck, and D. H. Persing. 1996.Concurrent Lyme disease and babesiosis: evidence for increased severity andduration of illness. JAMA 275:1657–1660.
12. Linden, J. V., S. J. Wong, F. K. Chu, G. B. Schmidt, and C. Bianco. 2000.Transfusion-associated transmission of babesiosis in New York State. Trans-fusion 40:285–289.
13. Matsui, T., R. Inoue, K. Kajimoto, A. Tamekane, Y. Katayama, M. Shi-moyama, K. Chihara, A. Saito-Ito, and M. Tsuji. 2000. First documentationof human babesiosis in Japan. Jpn. J. Clin. Hematol. 41:628–634 (In Japa-nese with English summary.)
14. McQuiston, J. H., J. E. Childs, M. E. Chamberland, and E. Tabor. 2000.Transmission of tick-borne agents of disease by blood transfusion: a reviewof known and potential risks in the United States. Transfusion 40:274–284.
15. Medlin, L., H. J. Elwood, S. Stickel, and M. L. Sogin. 1988. The character-ization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions.Gene 71:491–499.
16. Mitchell, P. D., K. D. Reed, and J. M. Hofkes. 1996. Immunoserologicevidence of coinfection with Borrelia burgdorferi, Babesia microti, and humangranulocytic Ehrlichia species in residents of Wisconsin and Minnesota.J. Clin. Microbiol. 34:724–727.
17. Nakamura, Y., M. Tsuji, S. Arai, and C. Ishihara. 1995. A method for rapidand complete substitution of the circulating erythrocytes in SCID mice withbovine erythrocytes and use of the substituted mice for bovine hemoprotozoainfections. J. Immunol. Methods 188:247–254.
18. Nakao, M., and K. Miyamoto. 1995. Mixed infection of different Borreliaspecies among Apodemus speciosus mice in Hokkaido, Japan. J. Clin. Mi-crobiol. 33:490–492.
19. Persing, D. H., D. Mathiesen, W. F. Marshall, S. R. Telford, A. Spielman,J. W. Thomford, and P. A. Conrad. 1992. Detection of Babesia microti bypolymerase chain reaction. J. Clin. Microbiol. 30:2097–2103.
20. Saito-Ito, A., M. Tsuji, Q. Wei, S. He, T. Matsui, M. Kohsaki, S. Arai, T.Kamiyama, K. Hioki, and C. Ishihara. 2000. Transfusion-acquired, autoch-thonous human babesiosis in Japan: isolation of Babesia microti-like para-
sites with hu-RBC-SCID mice. J. Clin. Microbiol. 38:4511–4516.21. Shih, C.-M., L.-P. Liu, W.-C. Chung, S. J. Ong, and C.-C. Wan. 1997. Human
babesiosis in Taiwan: asymptomatic infection with a Babesia microti-likeorganism in a Taiwanese woman. J. Clin. Microbiol. 35:450–454.
22. Shiota, T., H. Kurimoto, N. Haguma, and Y. Yoshida. 1984. Studies onBabesia first found in murine in Japan: epidemiology, morphology and ex-perimental infection. Zentbl. Bakteriol. Mikrobiol. Hyg. Reihe A 256:347–355.
23. Smith, T., and F. L. Kilbourne. 1893. Investigation into the nature, causa-tion, and prevention of Texas or southern cattle fever. Bull. Bur. Anim. Ind.U.S. Dept. Agric. 1:177–304.
24. Steketee, R. W., M. R. Eckman, E. C. Burgess, J. N. Kuritsky, J. Dickerson,W. L. Schell, M. S. Godsey, Jr., and J. P. Davis. 1985. Babesiosis in Wis-consin: a new focus of disease transmission. JAMA 53:2675–2678.
25. Telford, S. R., III, and J. H. Maguire. 1999. Babesiosis, p. 767–773. In R. L.Guerrant, D. H. Walker, and P. F. Weller (ed.), Tropical infectious diseases:principles, pathogens, and practice, vol. 1. Churchill Livingstone, London,England.
26. Telford, S. R., III, and A. Spielman. 1998. Babesiosis of humans, p. 349–359.In L. Collier, A. Balows, and M. Sussman (ed.), Topley and Wilson’s micro-biology and microbial infections, 9th ed., vol. 5. Arnold, London, England.
27. Tsuji, M., C. Ishihara, S. Arai, R. Hiratai, and I. Azuma. 1995. Establish-ment of a SCID mouse model having circulating human red blood cells anda possible growth of Plasmodium falciparum in the mouse. Vaccine 13:1389–1392.
28. van Peen, P. F. D., S. J. Chang, A. R. Banknieder, and F. J. Santana. 1977.Piroplasms from Taiwanese rodents. J. Protozool. 24:310–312.
29. Walter, G. 1984. Transmission and course of parasitemia of Babesia microti(Hannover I strain) in the bank vole (Clethrionomys glareolus) and field vole(Microtus agrestis). Acta Trop. 41:259–264. (In German.)
30. Watkins, R. A., S. E. Moshier, W. D. O’Dell, and A. J. Pinter. 1991. Spleno-megaly and reticulocytosis caused by Babesia microti infections in naturalpopulations of the montane vole, Microtus montanus. J. Protozool. 38:573–576.
31. Wittner, M., J. Lederman, H. B. Tanowitz, G. S. Rosenbaum, and L. M.Weiss. 1996. Atovaquone in the treatment of Babesia microti infections inhamsters. Am. J. Trop. Med. Hyg. 55:219–222.
32. Wittner, M., K. S. Rowin, H. B. Tanowitz, J. F. Hobbs, S. Saltzman, B. Wenz,R. Hirsch, E. Chisholm, and G. R. Healy. 1982. Successful chemotherapy oftransfusion babesiosis. Ann. Intern. Med. 96:601–604.
VOL. 39, 2001 HUMAN BABESIOSIS IN JAPAN 2183
on August 22, 2018 by guest
http://jcm.asm
.org/D
ownloaded from
Related Documents











