Human Amniotic Fluid Stem Cells Can Integrate and Differentiate into Epithelial Lung Lineages GIANNI CARRARO, a LAURA PERIN, a SARGIS SEDRAKYAN, a STEFANO GIULIANI, a CATERINA TIOZZO, b JOOEUN LEE, a GIANLUCA TURCATEL, a STIJN P. DE LANGHE, a BARBARA DRISCOLL, a SAVERIO BELLUSCI, a PARVIZ MINOO, b ANTHONY ATALA, c ROGER EDWARD DE FILIPPO, a DAVID WARBURTON a a Developmental Biology, Regenerative Medicine and Surgery Program, Saban Research Institute, Children’s Hospital Los Angeles, Keck School of Medicine and School of Dentistry, Los Angeles, California, USA; b Women’s & Children’s Hospital, Keck School of Medicine, University of Southern California, Los Angeles, California; c Wake Forest Institute for Regenerative Medicine, Wake Forest University School of Medicine, Winston-Salem, North Carolina Key Words. Amniotic fluid stem cells • Lung injury • Tissue repair • Regenerative medicine ABSTRACT A new source of stem cells has recently been isolated from amniotic fluid; these amniotic fluid stem cells have significant potential for regenerative medicine. These cells are multipo- tent, showing the ability to differentiate into cell types from each embryonic germ layer. We investigated the ability of human amniotic fluid stem cells (hAFSC) to integrate into murine lung and to differentiate into pulmonary lineages after injury. Using microinjection into cultured mouse embryonic lungs, hAFSC can integrate into the epithelium and express the early human differentiation marker thyroid transcription fac- tor 1 (TTF1). In adult nude mice, following hyperoxia injury, tail vein-injected hAFSC localized in the distal lung and ex- pressed both TTF1 and the type II pneumocyte marker sur- factant protein C. Specific damage of Clara cells through naph- thalene injury produced integration and differentiation of hAFSC at the bronchioalveolar and bronchial positions with expression of the specific Clara cell 10-kDa protein. These results illustrate the plasticity of hAFSC to respond in different ways to different types of lung damage by expressing specific alveolar versus bronchiolar epithelial cell lineage markers, de- pending on the type of injury to recipient lung. STEM CELLS 2008;26:2902–2911 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION Stem cells have the potential to contribute to the self-renewal and repair of the lung [1]. The existence of endogenous adult stem/progenitor cells has been reported in various key regions of the lung. Proximal airways contain basal cells capable of main- taining cellular turnover [2]. In contrast, the distal bronchioal- veolar compartment does not contain basal cells, but at least two different kinds of progenitor cells have been identified: a variant of Clara cells [3] that lie adjacent to neuroendocrine bodies, and bronchioalveolar stem cells [4] located at the bronchioalveolar duct junction [5]. However, the potential ability of these endog- enous stem/progenitor cells to repair injured lung is clearly limited when the injury is severe enough to cause irreversible respiratory failure. Exogenous stem cells have a feasible potential to be used as cellular therapy to contribute to or supplement endogenous lung repair mechanisms. Bone marrow-derived stem cells expressing green fluorescence protein can populate both proximal and distal lung airway in an irradiated mouse [6]. Embryonic stem cells have also been used to produce epithelial lung lineages in vivo and in culture [7, 8]. Recently, a new source of human amniotic fluid stem cells (hAFSC) has been isolated [9]. These cells represent 1% of the population of cells obtained from amniocentesis and are char- acterized by the expression of the receptor for stem cell factor c-Kit. hAFSC are multipotent, showing the ability to differen- tiate into lineages belonging to all three germ layers, and can be propagated easily in vitro without the need of a feeder layer. hAFSC express the markers OCT4 and SSEA-4, both of which are typical of the undifferentiated state of embryonic stem cells (ESC). However, hAFSC do not express some of the other typical markers of ESC, such as SSEA-3, and instead express mesenchymal and neuronal stem cell markers (CD29, CD44, CD73, CD90, and CD105) that are normally not expressed in Author contributions: G.C.: conception and design, data analysis and interpretation, manuscript writing, assembly of data; L.P.: conception and design, hAFSC culture; S.S.: tail vein injection of hAFSC; S.G.: bioluminescence experiments; C.T.: real-time PCR experiments; J.L.: mouse preparation, intratracheal administration of hAFSC; G.T.: cytospin experiments; S.P.D.L.: naphthalene injury experiments; B.D.: conception and design, data analysis and interpretation; S.B.: manuscript writing, collection and assembly of data; P.M.: data analysis and interpretation; A.A.: data interpretation; R.E.D.F.: collection and assembly of data, financial support; D.W.: manuscript writing, final approval of manuscript, financial support. Correspondence: David Warburton, DSc, M.D., FRCP, FRCS, Developmental Biology, Regenerative Medicine and Surgery Program, Saban Research Institute, Children’s Hospital Los Angeles, Keck School of Medicine and School of Dentistry, 4661 Sunset Boulevard MS 35, Los Angeles, California 90027, USA. Telephone: 323-669-5422; Fax: 323-671-3613; e-mail: [email protected] Received January 29, 2008; accepted for publication August 5, 2008; first published online in STEM CELLS EXPRESS August 21, 2008. ©AlphaMed Press 1066-5099/2008/$30.00/0 doi: 10.1634/stemcells.2008-0090 TISSUE-SPECIFIC STEM CELLS S TEM CELLS 2008;26:2902–2911 www.StemCells.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Human Amniotic Fluid Stem Cells Can Integrate and Differentiateinto Epithelial Lung Lineages
GIANNI CARRARO,a LAURA PERIN,a SARGIS SEDRAKYAN,a STEFANO GIULIANI,a CATERINA TIOZZO,b
JOOEUN LEE,a GIANLUCA TURCATEL,a STIJN P. DE LANGHE,a BARBARA DRISCOLL,a SAVERIO BELLUSCI,a
PARVIZ MINOO,b ANTHONY ATALA,c ROGER EDWARD DE FILIPPO,a DAVID WARBURTONa
aDevelopmental Biology, Regenerative Medicine and Surgery Program, Saban Research Institute, Children’sHospital Los Angeles, Keck School of Medicine and School of Dentistry, Los Angeles, California, USA; bWomen’s& Children’s Hospital, Keck School of Medicine, University of Southern California, Los Angeles, California;cWake Forest Institute for Regenerative Medicine, Wake Forest University School of Medicine, Winston-Salem,North Carolina
Key Words. Amniotic fluid stem cells • Lung injury • Tissue repair • Regenerative medicine
ABSTRACT
A new source of stem cells has recently been isolated fromamniotic fluid; these amniotic fluid stem cells have significantpotential for regenerative medicine. These cells are multipo-tent, showing the ability to differentiate into cell types fromeach embryonic germ layer. We investigated the ability ofhuman amniotic fluid stem cells (hAFSC) to integrate intomurine lung and to differentiate into pulmonary lineages afterinjury. Using microinjection into cultured mouse embryoniclungs, hAFSC can integrate into the epithelium and express theearly human differentiation marker thyroid transcription fac-tor 1 (TTF1). In adult nude mice, following hyperoxia injury,
tail vein-injected hAFSC localized in the distal lung and ex-pressed both TTF1 and the type II pneumocyte marker sur-factant protein C. Specific damage of Clara cells through naph-thalene injury produced integration and differentiation ofhAFSC at the bronchioalveolar and bronchial positions withexpression of the specific Clara cell 10-kDa protein. Theseresults illustrate the plasticity of hAFSC to respond in differentways to different types of lung damage by expressing specificalveolar versus bronchiolar epithelial cell lineage markers, de-pending on the type of injury to recipient lung. STEM CELLS2008;26:2902–2911
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Stem cells have the potential to contribute to the self-renewaland repair of the lung [1]. The existence of endogenous adultstem/progenitor cells has been reported in various key regions ofthe lung. Proximal airways contain basal cells capable of main-taining cellular turnover [2]. In contrast, the distal bronchioal-veolar compartment does not contain basal cells, but at least twodifferent kinds of progenitor cells have been identified: a variantof Clara cells [3] that lie adjacent to neuroendocrine bodies, andbronchioalveolar stem cells [4] located at the bronchioalveolarduct junction [5]. However, the potential ability of these endog-enous stem/progenitor cells to repair injured lung is clearlylimited when the injury is severe enough to cause irreversiblerespiratory failure.
Exogenous stem cells have a feasible potential to be used ascellular therapy to contribute to or supplement endogenous lung
repair mechanisms. Bone marrow-derived stem cells expressinggreen fluorescence protein can populate both proximal anddistal lung airway in an irradiated mouse [6]. Embryonic stemcells have also been used to produce epithelial lung lineages invivo and in culture [7, 8].
Recently, a new source of human amniotic fluid stem cells(hAFSC) has been isolated [9]. These cells represent 1% of thepopulation of cells obtained from amniocentesis and are char-acterized by the expression of the receptor for stem cell factorc-Kit. hAFSC are multipotent, showing the ability to differen-tiate into lineages belonging to all three germ layers, and can bepropagated easily in vitro without the need of a feeder layer.hAFSC express the markers OCT4 and SSEA-4, both of whichare typical of the undifferentiated state of embryonic stem cells(ESC). However, hAFSC do not express some of the othertypical markers of ESC, such as SSEA-3, and instead expressmesenchymal and neuronal stem cell markers (CD29, CD44,CD73, CD90, and CD105) that are normally not expressed in
Author contributions: G.C.: conception and design, data analysis and interpretation, manuscript writing, assembly of data; L.P.: conceptionand design, hAFSC culture; S.S.: tail vein injection of hAFSC; S.G.: bioluminescence experiments; C.T.: real-time PCR experiments; J.L.:mouse preparation, intratracheal administration of hAFSC; G.T.: cytospin experiments; S.P.D.L.: naphthalene injury experiments; B.D.:conception and design, data analysis and interpretation; S.B.: manuscript writing, collection and assembly of data; P.M.: data analysis andinterpretation; A.A.: data interpretation; R.E.D.F.: collection and assembly of data, financial support; D.W.: manuscript writing, finalapproval of manuscript, financial support.
Correspondence: David Warburton, DSc, M.D., FRCP, FRCS, Developmental Biology, Regenerative Medicine and Surgery Program, SabanResearch Institute, Children’s Hospital Los Angeles, Keck School of Medicine and School of Dentistry, 4661 Sunset Boulevard MS 35, LosAngeles, California 90027, USA. Telephone: 323-669-5422; Fax: 323-671-3613; e-mail: [email protected] Received January 29,2008; accepted for publication August 5, 2008; first published online in STEM CELLS EXPRESS August 21, 2008. ©AlphaMed Press1066-5099/2008/$30.00/0 doi: 10.1634/stemcells.2008-0090
TISSUE-SPECIFIC STEM CELLS
STEM CELLS 2008;26:2902–2911 www.StemCells.com

ESC. Amniotic fluid stem cells (AFSC) are also negative forhematopoietic stem cell markers (CD45, CD34, and CD133).Therefore, hAFSC can be considered as an intermediate type ofstem or progenitor cell between ESC and adult stem cellsresident in differentiated organs. De Coppi et al. [19] reportedthat hAFSC can be encouraged to differentiate into specializedfunctional populations, such as neural, hepatic, and osteogeniclineages. However, it is not known whether hAFSC have theability to participate in lung development or repair after injury.Herein we show that hAFSC can integrate into developing aswell as injured lung tissues and differentiate therein into lungepithelial lineages.
MATERIALS AND METHODS
Isolation, Culture, and Labeling of hAFSCSamples of human amniotic fluid from male fetuses (12–18-weekgestation) were provided by Genzyme Genetics Corporation (Mon-rovia, CA, http://www.genzymegenetics.com). The stem cell popu-lation was isolated using magnetic-activated cell sorting (MiltenyiBiotec, Auburn, CA, http://www.miltenyibiotec.com) against thecell surface marker c-kit. Pluripotential characteristics of the clonaland subclonal groups were tested according to published protocols[9]. Clones were cultured in Petri dishes in medium containing�-minimal essential medium supplemented with 20% Chang B and2% Chang C solutions, 20% fetal bovine serum (FBS), 1%L-glutamine, and 1% antibiotics (Gibco-BRL, Rockville, MD,http://www.gibcobrl.com). Clonal hAFSC populations were la-beled with the fluorescent cell surface marker chloromethylbenz-amide-1,1�-diactaolecyl-3,3,3�,3�-tetramethylindocarbocyanineperchlorate (CM-Dil) (Invitrogen, Carlsbad, CA, http://probes.invitrogen.com) to track the cells during and after injection.Briefly, the cells were incubated with a working solution ofCM-Dil at 1 mg/ml for 5 minutes at 37°C, followed by incuba-tion for 15 minutes at 4°C, followed by three washes withphosphate-buffered saline (PBS).
Organ Culture and MicroinjectionhAFSC were trypsinized, and 104 cells were loaded into a 15-�mdiameter transfer tip (Eppendorf AG, Hamburg, Germany, http://www.eppendorf.com), guided by a micromanipulator (TransferManNK2; Eppendorf) and a CellTram Oil injector (Eppendorf). Embry-onic mouse lungs were explanted at day 11.5 of gestation (E11.5).Immediately before injection, lungs were placed on a polyethylene-terephthalate track-etched membrane (Sterlitech Corp., Kent, WA,http://www.sterlitech.com) and microinjected through the trachea.Directly after microinjection, the lungs were placed on 8-�m porefilters (VWR, Marietta, GA, http://www.vwrsp.com); laid on thesurface of 800 �l of Dulbecco’s modified Eagle’s medium(DMEM)/Ham’s F-12 medium (F12) containing 10% FBS, 50units/ml penicillin, and 50 �g/ml streptomycin; placed in a 37°Cincubator; humidified; and kept at 7% CO2 for 2–10 days. For eachtime point, four embryonic lungs were injected and placed on thesame filter. Each time point was repeated al least in triplicate.
Injuries and Cell AdministrationA sex-mismatched model was used for all of our experiments inwhich hAFSC derived from male donors were administered tofemale nude mice. For the hyperoxia treatment, mice were exposedto short-term hyperoxia, as described previously. Briefly, mice wereplaced in a 90 � 42 � 38-cm Plexiglas chamber and exposed tohumidified �99% oxygen for 72 hours (Terra Universal, Fullerton,CA, http://www.terrauniversal.com). Control mice were kept inroom air during the treatment period. For the tail vein injection,�106 hAFSC were prepared in sterile PBS, and the injection wasperformed with a sterile 28-G1⁄2 insulin syringe. Six mice were usedto generate data for each time point and each experimental condi-tion. Mice exposed to hyperoxia were sacrificed after 1 week, 15
days, or 40 days, respectively. Control mice were also sacrificedafter 1 week, 15 days, or 40 days.
Naphthalene (Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) was dissolved to 30 mg/ml in corn oil (Sigma-Aldrich) andadministered at 275 mg/kg. Nude mice, 3 months old (JacksonLaboratories), were given naphthalene or corn oil alone byintraperitoneal injection. Mice were housed in plastic cages on a12-hour light/12-hour dark cycle with food and water ad libitum.Four mice were used to generate data for each time point andeach experimental condition. Mice exposed to naphthalene weresacrificed after 1 week, 15 days, or 40 days. Control mice werealso sacrificed after 1 week, 15 days, or 40 days.
For intratracheal instillation of hAFSC, mice were anesthetizedwith 5% isoflurane. After incision with a scalpel, trachea wasvisualized by depressing muscle tissue, and 2.5 � 105 hAFSC wereinjected. Mice were left in a prewarmed cage for recovery. Motrin(McNeil CHI, Fort Washington, PA, http://www.motrin.com) wasadded to drinking water. Three mice were used to generate data foreach time point and each experimental condition. Mice exposed tonaphthalene were sacrificed after 1 week, 15 days, or 40 days.Control mice were also sacrificed after 1 week, 15 days, or 40 days.For both naphthalene and oxygen injury experiments, mice weresacrificed, and lung tissue was either processed for the isolation ofgenomic DNA or total RNA or fixed and embedded in paraffin.
Live ImaginghAFSC with stable expression of firefly luciferase were used for tailvein injection. Prior to imaging, mice were injected with 125 mg/kgluciferin (Promega, Madison, WI, http://www.promega.com), asdescribed [10]. The animals were anesthetized, and luciferase bio-luminescence was monitored under a detector (Saban ResearchInstitute Small Animal Imaging Core facility) 4 hours after injectionof the substrate. The animals were also monitored at the followingtime points after stem cell injection: 1 day, 2 days, 3 days, 1 week,2 weeks, and 3 weeks. Images with all animals were taken at thesame intensity of signal, with the exception of the first 4 hours afterinjection, when the intensity was reduced to compensate for thestronger signal. Fifteen animals were analyzed at each time point.
Histology and ImmunohistochemistryLung tissues were fixed in 4% paraformaldehyde at 20 cm waterinflation pressure, embedded in paraffin, and cut into 3–5-�m-thicksections. Primary antibodies used for immunohistochemistry in-cluded thyroid transcription factor 1 (TTF1), pulmonary pro-surfac-tant protein C (pro-SPC) (Seven Hills, Cincinnati, http://www.sevenhillsbioreagents.com), Clara cell 10-kDa protein (CC10)(Santa Cruz Biotechnology Inc., Santa Cruz, CA, http://www.scbt.com), pan-cytokeratin (Sigma-Aldrich), podoplanin (PDPN) (De-velopmental Studies Hybridoma Bank, Iowa City, IA, http://www.uiowa.edu/�dshbwww), and F4/80 (Abcam, Cambridge, MA,http://www.abcam.com). Sections were counterstained with 4�,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlin-game, CA, http://www.vectorlabs.com) and photographed with aLeica microscope (Leica, Heerbrugg, Switzerland, http://www.leica.com).
In Situ Hybridization and ImmunofluorescenceChromogenic in situ hybridization (CISH) for Y chromosomewas performed on formalin-fixed, paraffin-embedded tissue sec-tions according to the manufacturer’s instructions (ZymedSPOT-Light CISH; Invitrogen). CISH signals were detected un-der light microscopy using a �40 objective lens. Fluorescence insitu hybridization (FISH) for X and Y chromosome was per-formed using commercially available probes (Vysis, DownersGrove, IL, http://www.vysis.com). For paraffin-embedded lungs,the sections were deparaffinized in Histochoice clearing agent(Sigma-Aldrich) and graded ethanol, incubated in boiling anti-gen-unmasking solution (Vector Laboratories) for 25 minutes,and equilibrated at room temperature. Slides were then immersedin 0.2 N HCl for 10 minutes, rinsed two times in PBS, andincubated at 78°C in NaSCN for 20 minutes. Slides were thenwashed in PBS and dehydrated with graded ethanol. Probe was
2903Carraro, Perin, Sedrakyan et al.
www.StemCells.com

prewarmed at 75°C for 5�, applied to the target area, sealed withcoverslips (Grace Bio-Labs, Bend, OR, http://www.gracebio.com), and hybridized overnight at 42°C. The next day, thecoverslips were removed, and the slides were incubated in 0.4�standard saline citrate (SSC; Bio-Rad, Hercules, CA, http://www.bio-rad.com)/0.3% Nonidet P40 (Sigma-Aldrich) at 75°Cfor 10 minutes; then, slides were immersed in 2� SSC/0.1%Nonidet P40 for 2 minutes and rinsed in PBS. Once dried, slideswere counterstained with 4�-6-diamidino-2-phenylindole (VectorLaboratories) and visualized with a fluorescence microscope. ForFISH on single cells, slides retrieved from �80°C storage wereallowed to dry for 5 minutes, and then probe was added asdescribed above and slides were incubated overnight at 42°C.The next day, slides were immersed in 2� SSC at 42°C for 15minutes, and then coverslips were removed and slides were leftin 0.4� SSC/0.3% Nonidet P40 for 2 minutes at 70°C. After awash in PBS for 5 minutes, slides were counterstained andanalyzed as described above. When combined immunostainingwas performed, primary antibody was incubated overnight at4°C, and secondary antibody was incubated for 30 minutes atroom temperature.
Tissue AnalysisFISH and immunohistochemistry were analyzed using a Leicafluorescence microscope. The percentage of integration was de-termined by counting CM-Dil-positive cells on the total numberof cells from three different experiments using five randomlychosen areas for each experiment. The percentage of differenti-ation was determined by counting the cells that were positive forboth Y chromosome and the marker for epithelial cell pancyto-keratin.
DNA and RNA Isolation and Reverse Transcription-Polymerase Chain ReactionGenomic DNA was extracted from lung using the DNeasy Blood &Tissue Kit (Qiagen, Hilden, Germany, http://www1.qiagen.com)following the protocol provided. Total RNA was extracted fromlung using TRIzol (Invitrogen, Carlsbad, CA, http://www.invitrogen.com). The purity of the RNA was analyzed by measuringthe absorbance ratio at 260 and 280 nm using a spectrophotometer,and the integrity of the RNA was confirmed by analyzing for thepresence of intact 28S and 18S RNA following agarose gel elec-trophoresis. Reverse transcription (RT) was performed starting from1 �g of RNA using Superscript-III (Invitrogen) according to themanufacturer’s protocols. Polymerase chain reaction (PCR) wasperformed with 50 ng of cDNA, 500 nM each primer, and a totalvolume of 25 �l using PCR supermix (Roche Diagnostics, Basel,Switzerland, http://www.roche-applied-science.com) and the fol-lowing primer pairs: TTF1 forward, CCAAGCGCATCCAATCT-CAA; TTF1 reverse, GGCAGAGTGTGCCCAGAGTG; CC10forward, CACCCTGGTCACACTGGCTC; CC10 reverse,GGAGGGTGTCCACCAGCTTC; SPC forward, TTGGTCCT-TCACCTCTGTCC; SPC reverse, CTCCAGAACCATCTC-CGTGT; PDPN forward, CCAGCGAAGACCGCTATAAG;PDPN reverse, GGTCACCGTGGATTCTGAGT. Primers were de-signed to discriminate between human and murine mRNA. PCRconditions were as follows: 28 cycles of 95°C for 30 seconds, 60°Cfor 30 seconds, and 72°C for 30 seconds; and 72°C extension for 2minutes. Loading conditions were checked using a mouse �-actin(mACTB), and the presence of human DNA in each sample wasanalyzed with human �-actin (hACTB).
Real-Time PCRReal-time PCR was performed using LightCycler 480 Probe MasterMix in a LightCycler 480 instrument (Roche Diagnostics). HumanCXCR4 (hCXCR4) and stromal cell-derived factor 1 (SDF1) ex-pression was calculated following normalization to ACTB levels byrelative quantification [11]. hCXCR4 was first amplified withexternal primers hCXCR4big-forward (F) (ATGCTTTCCTTG-GAGCCAAA) and hCXCR4big-reverse (R) (ATGCTTTCCTT-GGAGCCAAA). Real-time PCR was performed with hCXCR4-Forward (CTGTGAGCAGAGGGTCCAG) and hCXCR4-
Reverse (ATGAATGTCCACCTCGCTTT) and with probe 55from the Universal ProbeLibrary (Roche Diagnostics). Real-timePCR for SDF1 was performed with the following primers andprobe: SDF1-Forward (CTGTGCCCTTCAGATTGTTG), SDF1-Reverse (TAATTTCGGGTCAATGCACA), and probe 41 fromthe Universal ProbeLibrary (Roche Diagnostics). Real-time PCRfor the human Y chromosome was performed using the followingprimers and probe for Sry genes: SRY-Forward (TCGCATTTT-TCAGGACAGC), SRY-Reverse (CGTTGACTACTTGCCCT-GCT), and probe 16 from the Universal ProbeLibrary (RocheDiagnostics). A standard curve was obtained with a syntheticoligonucleotide (Operon, Alameda, CA, http://www.operon.com). Absolute quantification was performed with the secondderivative maximum method. Real-time PCR conditions were asfollows: 45 cycles of 95°C for 10 seconds, 60°C for 30 seconds.
Single-Cell PreparationHanks’ balanced saline solution (HBSS)-perfused lung sampleswere finely minced with razor blades and enzymatically digested for1 hour at 37°C in a solution of 1 mg/ml collagenase IV, 0.1 mg/mlhyaluronidase (Sigma-Aldrich), and 30 U/ml DNaseI (Roche Diag-nostics) in DMEM/F12 medium. The resulting digest was thenfiltered through a 40 �m Falcon cell strainer (BD Biosciences, SanDiego, http://www.bdbiosciences.com) to remove debris andwashed with HBSS. A sample chamber positioned on the top of amicroscope slide was loaded with �105 cells and centrifuged in aCytospin2 (Thermo Shandon Inc., Pittsburgh, http://www.thermo.com) at 350 rpm for 5 minutes. The slide was dried and stored at�80°C until it was used.
Statistical AnalysisComparisons between two groups were made using an unpaired t test. Avalue of p � .05 was considered statistically significant. Analyses weredone using GraphPad Prism software (GraphPad Software, Inc., San Di-ego, http://www.graphpad.com). Data are shown as mean � SD.
Figure 1. hAFSC integrate into embryonic lung epithelium and mesen-chyme and express the early marker of differentiation TTF1. Embryoniclung at embryonic stage 11.5 was microinjected with 104 hAFSC andcultured for 1 week. Immunohistochemistry for TTF1 (green) showedepithelial cells. hAFSC labeled with CM-Dil (red) were found in theepithelium (A), as well as the mesenchyme (B). Nuclei were counterstainedwith 4�,6-diamidino-2-phenylindole (blue) (�63). (C): Reverse transcrip-tion-polymerase chain reaction from embryonic lungs a week after themicroinjection showed that hAFSC start to express TTF1. Primers weredesigned to discriminate human from mouse TTF1. Abbreviations: ACTB,�-actin; hAFSC, human amniotic fluid stem cells; hLung, human lung;m-eLung, mouse embryonic lung; TTF1, thyroid transcription factor 1.
2904 Pulmonary Differentiation of AFSC
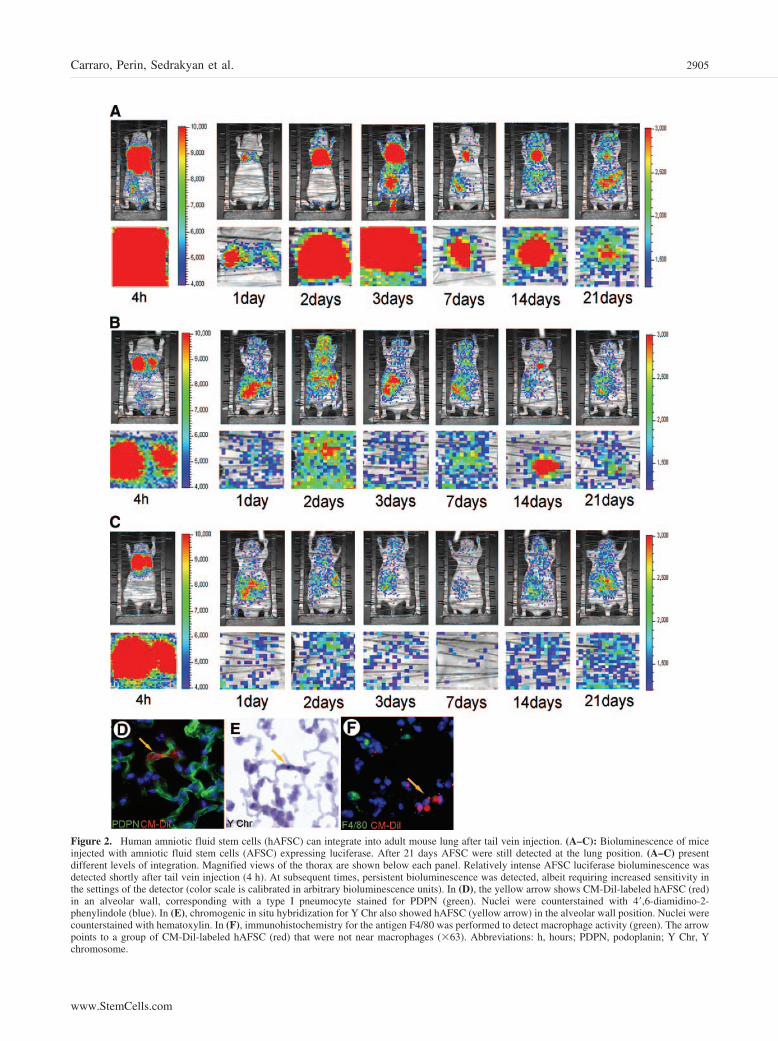
Figure 2. Human amniotic fluid stem cells (hAFSC) can integrate into adult mouse lung after tail vein injection. (A–C): Bioluminescence of miceinjected with amniotic fluid stem cells (AFSC) expressing luciferase. After 21 days AFSC were still detected at the lung position. (A–C) presentdifferent levels of integration. Magnified views of the thorax are shown below each panel. Relatively intense AFSC luciferase bioluminescence wasdetected shortly after tail vein injection (4 h). At subsequent times, persistent bioluminescence was detected, albeit requiring increased sensitivity inthe settings of the detector (color scale is calibrated in arbitrary bioluminescence units). In (D), the yellow arrow shows CM-Dil-labeled hAFSC (red)in an alveolar wall, corresponding with a type I pneumocyte stained for PDPN (green). Nuclei were counterstained with 4�,6-diamidino-2-phenylindole (blue). In (E), chromogenic in situ hybridization for Y Chr also showed hAFSC (yellow arrow) in the alveolar wall position. Nuclei werecounterstained with hematoxylin. In (F), immunohistochemistry for the antigen F4/80 was performed to detect macrophage activity (green). The arrowpoints to a group of CM-Dil-labeled hAFSC (red) that were not near macrophages (�63). Abbreviations: h, hours; PDPN, podoplanin; Y Chr, Ychromosome.
2905Carraro, Perin, Sedrakyan et al.
www.StemCells.com
RESULTS
hAFSC Engraft In Vitro into Epithelium andMesenchyme of Embryonic Lung Explants and Startto Express the Early Differentiation Marker TTF1To investigate the ability of hAFSC to integrate in murine lungtissues, we cultured lungs between E11.5 and E12.5 for short-term experiments. Approximately 104 CM-Dil-labeled hAFSCwere microinjected intratracheally into the embryonic lungs thatwere then analyzed at different time points. After 1 week,hAFSC had integrated into both epithelium and mesenchyme ofembryonic lungs (Fig. 1A, 1B). To assess whether hAFSCdifferentiate into lung epithelial-specific lineages after microin-jection, we used TTF1, a transcription factor that is expressed inearly embryonic lung epithelium, as a marker. Reverse tran-scription (RT)-PCR performed with TTF1 human-specific prim-ers showed that hAFSC do express TTF1 at 7 days after themicroinjection (Fig. 1C).
hAFSC Home In Vivo to Adult Mouse LungWe also examined whether hAFSC were able to uptake andintegrate into adult mouse lung. For these experiments we didnot damage the lung of the recipient mice, and approximately106 luciferase-labeled hAFSC were tail vein-injected per nudemouse. hAFSC were monitored as the cells passed through thecirculation. Figure 2B is representative of eight different exper-iments (of 15 total experiments) and shows that AFSC localizein the lung right after the injection. During the next few days,AFSC distribute around the body and sublocalize mainly in thearea of liver and lung, although a relatively weak signal issometimes seen in the head. A range of efficiency of uptake ofhAFSC into the lung was observed in luciferase experiments.Figure 2A and 2C shows two examples of the maximum and theminimum level of engraftment. The graph in Figure 3B showsthat hAFSC have an engraftment of 1.6% � 0.65% 1 week afterthe injection in control mice and that the engraftment goes downto 0.47% � 0.18% after 40 days. Engrafted CM-Dil-labeledcells were detectable 2 weeks after injection by fluorescence(Fig. 2D) and up to 7 months from the injection by CISH for Ychromosome (Fig. 2E). All these in vivo experiments were doneusing uninjured nude mice. Mice monitored up to 7 months fromthe injection did not show any neoplasia arising from thehAFSC. Also, intramuscular injection of hAFSC in nude micedid not produce any tumor formation (data not shown).
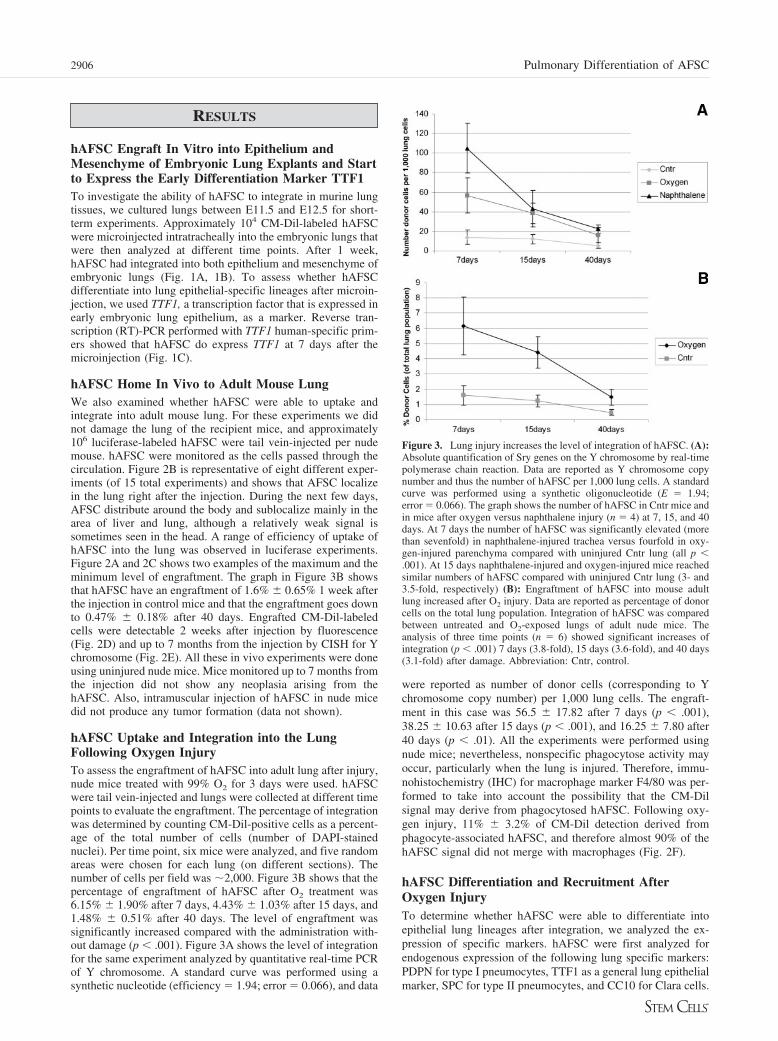
hAFSC Uptake and Integration into the LungFollowing Oxygen InjuryTo assess the engraftment of hAFSC into adult lung after injury,nude mice treated with 99% O2 for 3 days were used. hAFSCwere tail vein-injected and lungs were collected at different timepoints to evaluate the engraftment. The percentage of integrationwas determined by counting CM-Dil-positive cells as a percent-age of the total number of cells (number of DAPI-stainednuclei). Per time point, six mice were analyzed, and five randomareas were chosen for each lung (on different sections). Thenumber of cells per field was �2,000. Figure 3B shows that thepercentage of engraftment of hAFSC after O2 treatment was6.15% � 1.90% after 7 days, 4.43% � 1.03% after 15 days, and1.48% � 0.51% after 40 days. The level of engraftment wassignificantly increased compared with the administration with-out damage (p � .001). Figure 3A shows the level of integrationfor the same experiment analyzed by quantitative real-time PCRof Y chromosome. A standard curve was performed using asynthetic nucleotide (efficiency � 1.94; error � 0.066), and data
were reported as number of donor cells (corresponding to Ychromosome copy number) per 1,000 lung cells. The engraft-ment in this case was 56.5 � 17.82 after 7 days (p � .001),38.25 � 10.63 after 15 days (p � .001), and 16.25 � 7.80 after40 days (p � .01). All the experiments were performed usingnude mice; nevertheless, nonspecific phagocytose activity mayoccur, particularly when the lung is injured. Therefore, immu-nohistochemistry (IHC) for macrophage marker F4/80 was per-formed to take into account the possibility that the CM-Dilsignal may derive from phagocytosed hAFSC. Following oxy-gen injury, 11% � 3.2% of CM-Dil detection derived fromphagocyte-associated hAFSC, and therefore almost 90% of thehAFSC signal did not merge with macrophages (Fig. 2F).
hAFSC Differentiation and Recruitment AfterOxygen InjuryTo determine whether hAFSC were able to differentiate intoepithelial lung lineages after integration, we analyzed the ex-pression of specific markers. hAFSC were first analyzed forendogenous expression of the following lung specific markers:PDPN for type I pneumocytes, TTF1 as a general lung epithelialmarker, SPC for type II pneumocytes, and CC10 for Clara cells.
Figure 3. Lung injury increases the level of integration of hAFSC. (A):Absolute quantification of Sry genes on the Y chromosome by real-timepolymerase chain reaction. Data are reported as Y chromosome copynumber and thus the number of hAFSC per 1,000 lung cells. A standardcurve was performed using a synthetic oligonucleotide (E � 1.94;error � 0.066). The graph shows the number of hAFSC in Cntr mice andin mice after oxygen versus naphthalene injury (n � 4) at 7, 15, and 40days. At 7 days the number of hAFSC was significantly elevated (morethan sevenfold) in naphthalene-injured trachea versus fourfold in oxy-gen-injured parenchyma compared with uninjured Cntr lung (all p �.001). At 15 days naphthalene-injured and oxygen-injured mice reachedsimilar numbers of hAFSC compared with uninjured Cntr lung (3- and3.5-fold, respectively) (B): Engraftment of hAFSC into mouse adultlung increased after O2 injury. Data are reported as percentage of donorcells on the total lung population. Integration of hAFSC was comparedbetween untreated and O2-exposed lungs of adult nude mice. Theanalysis of three time points (n � 6) showed significant increases ofintegration (p � .001) 7 days (3.8-fold), 15 days (3.6-fold), and 40 days(3.1-fold) after damage. Abbreviation: Cntr, control.
2906 Pulmonary Differentiation of AFSC
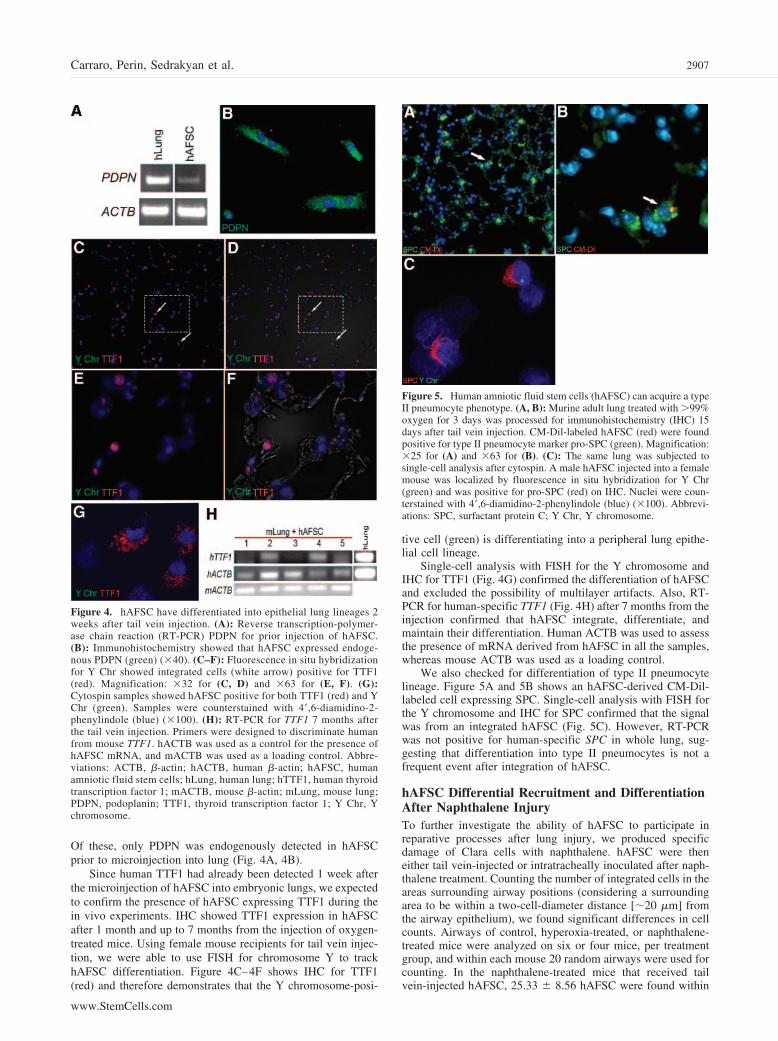
Of these, only PDPN was endogenously detected in hAFSCprior to microinjection into lung (Fig. 4A, 4B).
Since human TTF1 had already been detected 1 week afterthe microinjection of hAFSC into embryonic lungs, we expectedto confirm the presence of hAFSC expressing TTF1 during thein vivo experiments. IHC showed TTF1 expression in hAFSCafter 1 month and up to 7 months from the injection of oxygen-treated mice. Using female mouse recipients for tail vein injec-tion, we were able to use FISH for chromosome Y to trackhAFSC differentiation. Figure 4C–4F shows IHC for TTF1(red) and therefore demonstrates that the Y chromosome-posi-
tive cell (green) is differentiating into a peripheral lung epithe-lial cell lineage.
Single-cell analysis with FISH for the Y chromosome andIHC for TTF1 (Fig. 4G) confirmed the differentiation of hAFSCand excluded the possibility of multilayer artifacts. Also, RT-PCR for human-specific TTF1 (Fig. 4H) after 7 months from theinjection confirmed that hAFSC integrate, differentiate, andmaintain their differentiation. Human ACTB was used to assessthe presence of mRNA derived from hAFSC in all the samples,whereas mouse ACTB was used as a loading control.
We also checked for differentiation of type II pneumocytelineage. Figure 5A and 5B shows an hAFSC-derived CM-Dil-labeled cell expressing SPC. Single-cell analysis with FISH forthe Y chromosome and IHC for SPC confirmed that the signalwas from an integrated hAFSC (Fig. 5C). However, RT-PCRwas not positive for human-specific SPC in whole lung, sug-gesting that differentiation into type II pneumocytes is not afrequent event after integration of hAFSC.
hAFSC Differential Recruitment and DifferentiationAfter Naphthalene InjuryTo further investigate the ability of hAFSC to participate inreparative processes after lung injury, we produced specificdamage of Clara cells with naphthalene. hAFSC were theneither tail vein-injected or intratracheally inoculated after naph-thalene treatment. Counting the number of integrated cells in theareas surrounding airway positions (considering a surroundingarea to be within a two-cell-diameter distance [�20 �m] fromthe airway epithelium), we found significant differences in cellcounts. Airways of control, hyperoxia-treated, or naphthalene-treated mice were analyzed on six or four mice, per treatmentgroup, and within each mouse 20 random airways were used forcounting. In the naphthalene-treated mice that received tailvein-injected hAFSC, 25.33 � 8.56 hAFSC were found within
Figure 4. hAFSC have differentiated into epithelial lung lineages 2weeks after tail vein injection. (A): Reverse transcription-polymer-ase chain reaction (RT-PCR) PDPN for prior injection of hAFSC.(B): Immunohistochemistry showed that hAFSC expressed endoge-nous PDPN (green) (�40). (C–F): Fluorescence in situ hybridizationfor Y Chr showed integrated cells (white arrow) positive for TTF1(red). Magnification: �32 for (C, D) and �63 for (E, F). (G):Cytospin samples showed hAFSC positive for both TTF1 (red) and YChr (green). Samples were counterstained with 4�,6-diamidino-2-phenylindole (blue) (�100). (H): RT-PCR for TTF1 7 months afterthe tail vein injection. Primers were designed to discriminate humanfrom mouse TTF1. hACTB was used as a control for the presence ofhAFSC mRNA, and mACTB was used as a loading control. Abbre-viations: ACTB, �-actin; hACTB, human �-actin; hAFSC, humanamniotic fluid stem cells; hLung, human lung; hTTF1, human thyroidtranscription factor 1; mACTB, mouse �-actin; mLung, mouse lung;PDPN, podoplanin; TTF1, thyroid transcription factor 1; Y Chr, Ychromosome.
Figure 5. Human amniotic fluid stem cells (hAFSC) can acquire a typeII pneumocyte phenotype. (A, B): Murine adult lung treated with �99%oxygen for 3 days was processed for immunohistochemistry (IHC) 15days after tail vein injection. CM-Dil-labeled hAFSC (red) were foundpositive for type II pneumocyte marker pro-SPC (green). Magnification:�25 for (A) and �63 for (B). (C): The same lung was subjected tosingle-cell analysis after cytospin. A male hAFSC injected into a femalemouse was localized by fluorescence in situ hybridization for Y Chr(green) and was positive for pro-SPC (red) on IHC. Nuclei were coun-terstained with 4�,6-diamidino-2-phenylindole (blue) (�100). Abbrevi-ations: SPC, surfactant protein C; Y Chr, Y chromosome.
2907Carraro, Perin, Sedrakyan et al.
www.StemCells.com
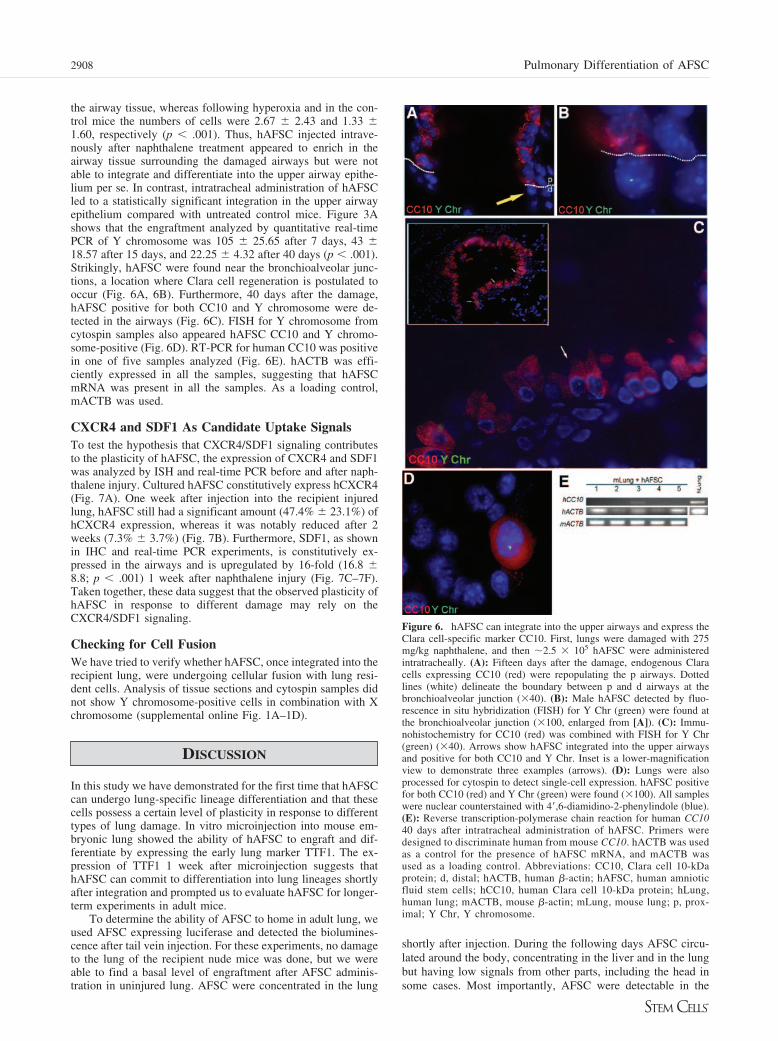
the airway tissue, whereas following hyperoxia and in the con-trol mice the numbers of cells were 2.67 � 2.43 and 1.33 �1.60, respectively (p � .001). Thus, hAFSC injected intrave-nously after naphthalene treatment appeared to enrich in theairway tissue surrounding the damaged airways but were notable to integrate and differentiate into the upper airway epithe-lium per se. In contrast, intratracheal administration of hAFSCled to a statistically significant integration in the upper airwayepithelium compared with untreated control mice. Figure 3Ashows that the engraftment analyzed by quantitative real-timePCR of Y chromosome was 105 � 25.65 after 7 days, 43 �18.57 after 15 days, and 22.25 � 4.32 after 40 days (p � .001).Strikingly, hAFSC were found near the bronchioalveolar junc-tions, a location where Clara cell regeneration is postulated tooccur (Fig. 6A, 6B). Furthermore, 40 days after the damage,hAFSC positive for both CC10 and Y chromosome were de-tected in the airways (Fig. 6C). FISH for Y chromosome fromcytospin samples also appeared hAFSC CC10 and Y chromo-some-positive (Fig. 6D). RT-PCR for human CC10 was positivein one of five samples analyzed (Fig. 6E). hACTB was effi-ciently expressed in all the samples, suggesting that hAFSCmRNA was present in all the samples. As a loading control,mACTB was used.
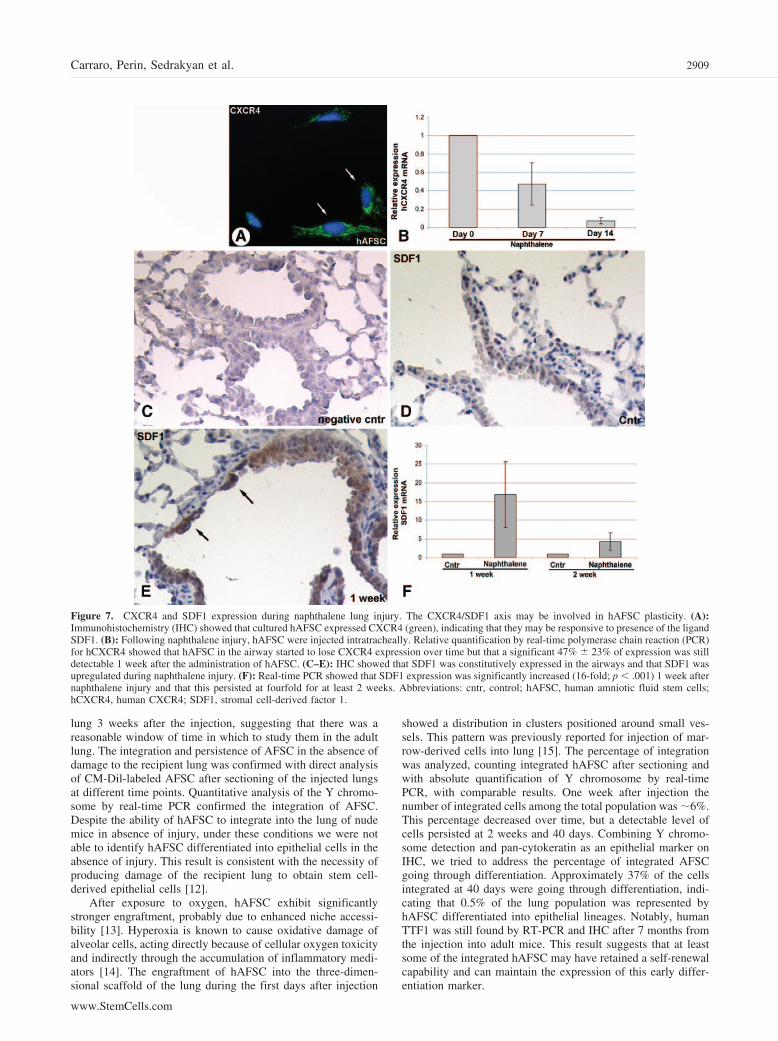
CXCR4 and SDF1 As Candidate Uptake SignalsTo test the hypothesis that CXCR4/SDF1 signaling contributesto the plasticity of hAFSC, the expression of CXCR4 and SDF1was analyzed by ISH and real-time PCR before and after naph-thalene injury. Cultured hAFSC constitutively express hCXCR4(Fig. 7A). One week after injection into the recipient injuredlung, hAFSC still had a significant amount (47.4% � 23.1%) ofhCXCR4 expression, whereas it was notably reduced after 2weeks (7.3% � 3.7%) (Fig. 7B). Furthermore, SDF1, as shownin IHC and real-time PCR experiments, is constitutively ex-pressed in the airways and is upregulated by 16-fold (16.8 �8.8; p � .001) 1 week after naphthalene injury (Fig. 7C–7F).Taken together, these data suggest that the observed plasticity ofhAFSC in response to different damage may rely on theCXCR4/SDF1 signaling.
Checking for Cell FusionWe have tried to verify whether hAFSC, once integrated into therecipient lung, were undergoing cellular fusion with lung resi-dent cells. Analysis of tissue sections and cytospin samples didnot show Y chromosome-positive cells in combination with Xchromosome (supplemental online Fig. 1A–1D).
DISCUSSION
In this study we have demonstrated for the first time that hAFSCcan undergo lung-specific lineage differentiation and that thesecells possess a certain level of plasticity in response to differenttypes of lung damage. In vitro microinjection into mouse em-bryonic lung showed the ability of hAFSC to engraft and dif-ferentiate by expressing the early lung marker TTF1. The ex-pression of TTF1 1 week after microinjection suggests thathAFSC can commit to differentiation into lung lineages shortlyafter integration and prompted us to evaluate hAFSC for longer-term experiments in adult mice.
To determine the ability of AFSC to home in adult lung, weused AFSC expressing luciferase and detected the biolumines-cence after tail vein injection. For these experiments, no damageto the lung of the recipient nude mice was done, but we wereable to find a basal level of engraftment after AFSC adminis-tration in uninjured lung. AFSC were concentrated in the lung
shortly after injection. During the following days AFSC circu-lated around the body, concentrating in the liver and in the lungbut having low signals from other parts, including the head insome cases. Most importantly, AFSC were detectable in the
Figure 6. hAFSC can integrate into the upper airways and express theClara cell-specific marker CC10. First, lungs were damaged with 275mg/kg naphthalene, and then �2.5 � 105 hAFSC were administeredintratracheally. (A): Fifteen days after the damage, endogenous Claracells expressing CC10 (red) were repopulating the p airways. Dottedlines (white) delineate the boundary between p and d airways at thebronchioalveolar junction (�40). (B): Male hAFSC detected by fluo-rescence in situ hybridization (FISH) for Y Chr (green) were found atthe bronchioalveolar junction (�100, enlarged from [A]). (C): Immu-nohistochemistry for CC10 (red) was combined with FISH for Y Chr(green) (�40). Arrows show hAFSC integrated into the upper airwaysand positive for both CC10 and Y Chr. Inset is a lower-magnificationview to demonstrate three examples (arrows). (D): Lungs were alsoprocessed for cytospin to detect single-cell expression. hAFSC positivefor both CC10 (red) and Y Chr (green) were found (�100). All sampleswere nuclear counterstained with 4�,6-diamidino-2-phenylindole (blue).(E): Reverse transcription-polymerase chain reaction for human CC1040 days after intratracheal administration of hAFSC. Primers weredesigned to discriminate human from mouse CC10. hACTB was usedas a control for the presence of hAFSC mRNA, and mACTB wasused as a loading control. Abbreviations: CC10, Clara cell 10-kDaprotein; d, distal; hACTB, human �-actin; hAFSC, human amnioticfluid stem cells; hCC10, human Clara cell 10-kDa protein; hLung,human lung; mACTB, mouse �-actin; mLung, mouse lung; p, prox-imal; Y Chr, Y chromosome.
2908 Pulmonary Differentiation of AFSC
lung 3 weeks after the injection, suggesting that there was areasonable window of time in which to study them in the adultlung. The integration and persistence of AFSC in the absence ofdamage to the recipient lung was confirmed with direct analysisof CM-Dil-labeled AFSC after sectioning of the injected lungsat different time points. Quantitative analysis of the Y chromo-some by real-time PCR confirmed the integration of AFSC.Despite the ability of hAFSC to integrate into the lung of nudemice in absence of injury, under these conditions we were notable to identify hAFSC differentiated into epithelial cells in theabsence of injury. This result is consistent with the necessity ofproducing damage of the recipient lung to obtain stem cell-derived epithelial cells [12].
After exposure to oxygen, hAFSC exhibit significantlystronger engraftment, probably due to enhanced niche accessi-bility [13]. Hyperoxia is known to cause oxidative damage ofalveolar cells, acting directly because of cellular oxygen toxicityand indirectly through the accumulation of inflammatory medi-ators [14]. The engraftment of hAFSC into the three-dimen-sional scaffold of the lung during the first days after injection
showed a distribution in clusters positioned around small ves-sels. This pattern was previously reported for injection of mar-row-derived cells into lung [15]. The percentage of integrationwas analyzed, counting integrated hAFSC after sectioning andwith absolute quantification of Y chromosome by real-timePCR, with comparable results. One week after injection thenumber of integrated cells among the total population was �6%.This percentage decreased over time, but a detectable level ofcells persisted at 2 weeks and 40 days. Combining Y chromo-some detection and pan-cytokeratin as an epithelial marker onIHC, we tried to address the percentage of integrated AFSCgoing through differentiation. Approximately 37% of the cellsintegrated at 40 days were going through differentiation, indi-cating that 0.5% of the lung population was represented byhAFSC differentiated into epithelial lineages. Notably, humanTTF1 was still found by RT-PCR and IHC after 7 months fromthe injection into adult mice. This result suggests that at leastsome of the integrated hAFSC may have retained a self-renewalcapability and can maintain the expression of this early differ-entiation marker.
Figure 7. CXCR4 and SDF1 expression during naphthalene lung injury. The CXCR4/SDF1 axis may be involved in hAFSC plasticity. (A):Immunohistochemistry (IHC) showed that cultured hAFSC expressed CXCR4 (green), indicating that they may be responsive to presence of the ligandSDF1. (B): Following naphthalene injury, hAFSC were injected intratracheally. Relative quantification by real-time polymerase chain reaction (PCR)for hCXCR4 showed that hAFSC in the airway started to lose CXCR4 expression over time but that a significant 47% � 23% of expression was stilldetectable 1 week after the administration of hAFSC. (C–E): IHC showed that SDF1 was constitutively expressed in the airways and that SDF1 wasupregulated during naphthalene injury. (F): Real-time PCR showed that SDF1 expression was significantly increased (16-fold; p � .001) 1 week afternaphthalene injury and that this persisted at fourfold for at least 2 weeks. Abbreviations: cntr, control; hAFSC, human amniotic fluid stem cells;hCXCR4, human CXCR4; SDF1, stromal cell-derived factor 1.
2909Carraro, Perin, Sedrakyan et al.
www.StemCells.com

Pro-SPC expression by hAFSC has been observed duringour analysis by IHC, and this was confirmed with Y chromo-some on single cells, suggesting that engraftment as type IIpneumocytes can occur. However, the major caveat on thisconclusion is that we could not detect SPC expression withRT-PCR with human-specific primers.
After naphthalene injury, hAFSC showed localizationwithin the bronchial tissue versus at bronchioalveolar junctions,where the damage to Clara cells had occurred, depending on therespective route of administration: i.v. versus intratracheal.Naphthalene is an aromatic hydrocarbon used to injure the lungby specifically destroying Clara cells that express the cyto-chrome P450 (Cyp2f2) [16]. hAFSC given by tail vein injectionafter naphthalene injury accumulated in the upper airway, in-creased numbers compared with after oxygen treatment. Thisbehavior suggests a certain level of plasticity of AFSC inresponding to different types of lung damage.
The chemokine SDF1 is involved in progenitor cell traffick-ing [17], and the CXCR4/SDF1 signaling axis is involved in thehoming of circulating stem cells into bleomycin-injured lung[18]. We found that cultured hAFSC express hCXCR4 andretain a significant amount of hCXCR4 expression 7 days afterthe administration of the cells to a naphthalene-injured lung.SDF1, which is the ligand of CXCR4, is constitutively ex-pressed in the airways, and its level of expression has a peakafter 7 days from naphthalene injury. Taken together, theseobservations are consistent with a possible involvement of theCXCR4/SDF1 signaling axis on the plasticity of hAFSC ob-served in this study.
hAFSC added intratracheally after naphthalene injury ini-tially had an higher level of epithelial integration, possibly dueto the direct administration (Fig. 3A). During the followingdays, there was a progressive reduction of integrated cells from15 to 40 days. After naphthalene injury, intratracheally admin-istered hAFSC were found at bronchioalveolar positions whereCC10-positive cells had been damaged. Also, FISH for Y chro-mosome and IHC for CC10 on tissue sections, as well assingle-cell cytospin, showed that hAFSC in these conditions canstart to express CC10, indicating that they may differentiate intoClara cells (Fig. 6D).
We tried to determine whether hAFSC, once integrated intothe recipient lung, were undergoing cellular fusion with endog-enous resident cells (supplemental online Fig. 1). During theanalysis of tissue and cytospin samples, we never found Ychromosome-positive cells in combination with the X chromo-
some. However, considering the small absolute number of Ychromosome-positive cells identified, we are unable to posi-tively exclude the possibility that hAFSC may occasionally fusewith resident cells of recipient mouse lung.
CONCLUSION
Additional experiments and different approaches are needed tooptimize hAFSC integration and differentiation. The possibilityof deriving specific lung lineages from stem cells in vitro hasalready been shown [19]. Thus, the induction of the hAFSC intospecific lung epithelial lineages before administration may beone approach to improving their integration, survival, and dif-ferentiation. Our data with embryonic lung organ culture and theexpression of TTF1 1 week after the microinjection suggest thatculture of hAFSC before injection in vivo into the adult animalmay perhaps increase the percentage of integration and hope-fully of differentiation. We conclude that the abilities of hAFSC,demonstrated herein, to express specific epithelial lung markersand to respond in a selective way to different types of lungdamage support further evaluation of hAFSC as a resource forcell-based therapy for the lung.
ACKNOWLEDGMENTS
We thank E. Herzog for the suggestions for FISH and IHCdouble staining. This work was supported by NIH NationalHeart, Lung, and Blood Institute Grants HL60231, 44060,44977, and 75773 (to D.W.), NIH KO8 (to R.E.D.F.), a Cali-fornia Institute for Regenerative Medicine Fellowship (to L.P.and C.T.), and Childrens Hospital Los Angeles Career Devel-opment Fellowship (to S.P.D.L.). The hybridoma antibody forpodoplanin developed by Andrew Farr was obtained from theDevelopmental Studies Hybridoma Bank developed under theauspices of the National Institute of Child Health and HumanDevelopment and maintained by the Department of BiologySciences, University of Iowa, Iowa City, IA.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Warburton D, Berberich MA, Driscoll B. Stem/progenitor cells inlung morphogenesis, repair, and regeneration. Curr Top Dev Biol2004;64:1–16.
2 Hong KU, Reynolds SD, Watkins S et al. Basal cells are a multipotentprogenitor capable of renewing the bronchial epithelium. Am J Pathol2004;164:577–588.
3 Hong KU, Reynolds SD, Giangreco A et al. Clara cell secretoryprotein-expressing cells of the airway neuroepithelial body microen-vironment include a label-retaining subset and are critical for epithe-lial renewal after progenitor cell depletion. Am J Respir Cell Mol Biol2001;24:671– 681.
4 Kim CF, Jackson EL, Woolfenden AE et al. Identification of bron-chioalveolar stem cells in normal lung and lung cancer. Cell 2005;121:823– 835.
5 Giangreco A, Reynolds SD, Stripp BR. Terminal bronchioles harbor aunique airway stem cell population that localizes to the bronchoalveolarduct junction. Am J Pathol 2002;161:173–182.
6 Krause DS, Theise ND, Collector MI et al. Multi-organ, multi-lineageengraftment by a single bone marrow-derived stem cell. Cell 2001;105:369–377.
7 Coraux C, Nawrocki-Raby B, Hinnrasky J et al. Embryonic stem cellsgenerate airway epithelial tissue. Am J Respir Cell Mol Biol 2005;32:87–92.
8 Rippon HJ, Polak JM, Qin M et al. Derivation of distal lung epithelialprogenitors from murine embryonic stem cells using a novel three-stepdifferentiation protocol. STEM CELLS 2006;24:1389–1398.
9 De Coppi P, Bartsch G Jr, Siddiqui MM et al. Isolation of amniotic stemcell lines with potential for therapy. Nat Biotechnol 2007;25:100–106.
10 Wang X, Rosol M, Ge S et al. Dynamic tracking of human hematopoieticstem cell engraftment using in vivo bioluminescence imaging. Blood2003;102:3478–3482.
11 Herzog EL, Van Arnam J, Hu B et al. Threshold of lung injury requiredfor the appearance of marrow-derived lung epithelia. STEM CELLS2006;24:1986–1992.
12 Pfaffl MW. A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res 2001;29:e45.
13 Pagano A, Barazzone-Argiroffo C. Alveolar cell death in hyperoxia-induced lung injury. Ann N Y Acad Sci 2003;1010:405–416.
14 Adamson IY, Bowden DH. Bleomycin-induced injury and metaplasia ofalveolar type 2 cells. Relationship of cellular responses to drug presencein the lung. Am J Pathol 1979;96:531–544.
15 Ceradini DJ, Kulkarni AR, Callaghan MJ et al. Progenitor cell traffickingis regulated by hypoxic gradients through HIF-1 induction of SDF-1. NatMed 2004;10:858–864.
2910 Pulmonary Differentiation of AFSC

16 Kotton DN, Ma BY, Cardoso WV et al. Bone marrow-derived cells asprogenitors of lung alveolar epithelium. Development 2001;128:5181–5188.
17 Stripp BR, Maxson K, Mera R et al. Plasticity of airway cell proliferationand gene expression after acute naphthalene injury. Am J Physiol 1995;269:L791–L799.
18 Gomperts BN, Belperio JA, Rao PN et al. Circulating progenitor epithe-lial cells traffic via CXCR4/CXCL12 in response to airway injury.J Immunol 2006;176:1916–1927.
19 Ali NN, Edgar AJ, Samadikuchaksaraei A et al. Derivation of type IIalveolar epithelial cells from murine embryonic stem cells. Tissue Eng2002;8:541–550.
See www.StemCells.com for supplemental material available online.
2911Carraro, Perin, Sedrakyan et al.
Related Documents











