1 HUMAN ALTERATION OF GROUNDWATER-SURFACE WATER INTERACTIONS 1 (SAGITTARIO RIVER, CENTRAL ITALY): IMPLICATION FOR FLOW REGIME, CONTAMINANT 2 FATE AND INVERTEBRATE RESPONSE 3 4 5 Mariachiara Caschetto 1 , Maurizio Barbieri 1 , Diana M. P. Galassi 2 , Lucia Mastrorillo 3 , Sergio Rusi 4 , 6 Fabio Stoch 2 , Alessia Di Cioccio 2 , Marco Petitta 1 7 8 9 1 Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”, P.le Aldo Moro 5, 00185 Roma, Italy 10 11 2 Dipartimento della Vita, della Salute e delle Scienze Ambientali, Università de L’Aquila, Via Vetoio, Coppito 12 67100 L'Aquila, Italy 13 14 3 Dipartimento di Scienze Geologiche, Università “Roma Tre”, Largo San Leonardo Murialdo, 00146 Roma, 15 Italy 16 17 4 Dipartimento di Ingegneria e Geologia (INGEO) Sezione di Geologia, Università G. d'Annunzio, Via dei 18 Vestini 30, 66013 Chieti, Italy 19 20 21 22 23 24 25 26 27 28 Corresponding author: Marco Petitta 29 E-mail: [email protected] 30 Phone: 0039-06-49914834 31 Fax: 0039-06-4454729 32 33 34 35 36 37 38 39 40 41

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
HUMAN ALTERATION OF GROUNDWATER-SURFACE WATER INTERACTIONS 1 (SAGITTARIO RIVER, CENTRAL ITALY): IMPLICATION FOR FLOW REGIME, CONTAMINANT 2 FATE AND INVERTEBRATE RESPONSE 3 4 5 Mariachiara Caschetto1, Maurizio Barbieri1, Diana M. P. Galassi2, Lucia Mastrorillo3, Sergio Rusi4, 6 Fabio Stoch2, Alessia Di Cioccio2, Marco Petitta1 7 8 9 1Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”, P.le Aldo Moro 5, 00185 Roma, Italy 10 11 2Dipartimento della Vita, della Salute e delle Scienze Ambientali, Università de L’Aquila, Via Vetoio, Coppito 12 67100 L'Aquila, Italy 13 14 3Dipartimento di Scienze Geologiche, Università “Roma Tre”, Largo San Leonardo Murialdo, 00146 Roma, 15 Italy 16 17 4Dipartimento di Ingegneria e Geologia (INGEO) Sezione di Geologia, Università G. d'Annunzio, Via dei 18 Vestini 30, 66013 Chieti, Italy 19 20 21 22 23 24 25 26 27 28 Corresponding author: Marco Petitta 29 E-mail: [email protected] 30 Phone: 0039-06-49914834 31 Fax: 0039-06-4454729 32 33 34 35 36 37 38 39 40
41

2
ABSTRACT 42
Many rivers world-wide are undergoing severe man-induced alterations which are reflected also in 43
changes of the degree of connectivity between surface waters and groundwater. Pollution, 44
irrigation withdrawal, alteration of freshwater flows, road construction, surface water diversion, soil 45
erosion in agriculture, deforestation and dam building have led to some irreversible species losses 46
and severe changes in community composition of freshwater ecosystems. 47
Taking into account the impact of damming and flow diversion on natural river discharge, the 48
present study is aimed at (i) evaluating the effects of anthropogenic changes on 49
groundwater/surface water interactions; (ii) analysing the fate of nitrogenous pollutants at the 50
floodplain scale; and (iii) describing the overall response of invertebrate assemblages to such 51
changes. 52
Hydrogeological, geochemical, and isotopic data revealed short- and long-term changes in 53
hydrology, allowing the assessment of the hydrogeological setting and the evaluation of potential 54
contamination by nitrogen compounds. Water isotopes allowed distinguishing a shallow aquifer 55
locally fed by zenithal recharge and river losses, and a deeper aquifer/aquitard system fed by 56
surrounding carbonate aquifers. This system was found to retain ammonium and, through the 57
shallow aquifer, release it in surface running waters via the hyporheic zone of the river bed. All 58
these factors influenced river ecosystem health. As many environmental drivers entered in action 59
offering a multiple - component artificial environment, a clear relationship between river flow 60
alteration and benthic and hyporheic invertebrate diversity was not found, being species response 61
driven by the combination of three main stressors: ammonium pollution, man-induced changes in 62
river morphology and altered discharge regime. 63
64
Key words: river, nitrogen cycle, stable isotope, surface/groundwater interaction, GDE, Italy 65
66
67

3
INTRODUCTION 68
The key role of hydrological processes on ecological functioning in rivers is largely emphasized 69
and the cause-effect relationship between them already described in the Water Framework 70
Directive (WFD) of the European Community (EC, 2000). Nevertheless, there is still great 71
uncertainty in assessing a clear link between flow alteration and ecological response, in part 72
because along with man-induced flow alteration, several other environmental parameters may 73
change and not always as “dependent variables.” Additionally, if alteration of river flow may lead to 74
bank and riverbed sediment erosions with severe changes in river bed sediment composition, there 75
are other factors that contribute to the alteration of the abiotic facies of rivers. For instance, 76
agriculture, urbanization and industrialization determine the pollution of soil and water in both 77
surface-flowing water and groundwater. 78
Dams lead to severe changes in river discharge regimes, altering the natural distribution and timing 79
of stream flow by high-magnitude peaks during dam release, with a cascade effect on river 80
morphology, alteration of riffle-pools sequences, and an increase in sediment and bank erosion 81
downstream (Boulton et al., 1998; Salem et al., 2012). These man-induced alterations co-occur in 82
the Sagittario River floodplain in central Italy with indiscriminate land use for urbanization, 83
agriculture and industrialization. Moreover, water diversion for irrigation coexists with damming, 84
further altering the natural flow regime, and urbanization and agriculture in the floodplain increases 85
the nitrogenous contamination. 86
Hydropower dams release high-discharge water downstream, leading to floods in the alluvial plain, 87
where urban and agricultural settlements are located (May et al., 2011). In these areas, local 88
authorities introduce additional interference by means of river course rectification and 89
channelization by erasing riparian vegetation. Moreover, where discharge is very high and the river 90
flows close to roads, railways and urbanized areas, the river flow is moved out of its original 91
topology, making parts of the river artificially perched. Changes in river hydrology and dynamics 92
can directly or indirectly influence the chemical characteristics of surface water and groundwater 93
(EC, 2012; Rao et al., 2012) by altering the connectivity between the river and the underlying 94
aquifer that has become disconnected in the artificial perched stretch, as in the Sagittario River 95

4
(Banks et al., 2011). This condition alters the topology of the groundwater-dependent ecosystems 96
(GDEs) (Eamus and Froend, 2006), consequently affecting freshwater invertebrate biodiversity 97
(Poff et al., 2010) at the interface between surface water and groundwater, in the so-called 98
hyporheic zone of the riverbed sediments, and on the surface benthic macroinvertebrate 99
communities. Moreover, understanding how hydrological and biological processes in the floodplain 100
and in the hyporheic zone may control the concentration and fate of nitrogen compounds in 101
watersheds is crucial (Landers et al., 2008; Di Lorenzo et al., 2012). In particular, nitrate 102
attenuation depends on the capacity of riparian vegetation and microbial communities to intercept 103
pollutants moving in surface runoff and groundwater flow (Hill, 1996; Cey et al., 1999; Petitta et al., 104
2009; Keskin, 2010; Wexler et al., 2011).In a groundwater multi-layer system (Stanford and Ward, 105
1992), as in the case under study, the interactions between groundwater and surface water that 106
are influenced by the alteration of river discharge and morphology can generate impacts on GDEs 107
(e.g., in terms of nitrogen contamination and invertebrate community compositions). 108
An integrated multidisciplinary monitoring approach was applied to man-altered stretches of the 109
Sagittario River (central Italy). This study was focused on assessing (i) surface water-groundwater 110
interactions, (ii) the influence of surface water and groundwater interactions on the nitrogen cycle 111
at the floodplain scale, and (iii) freshwater benthic and hyporheic invertebrate responses to multiple 112
disturbances. 113
114
STUDY AREA 115
The Sulmona Basin, a Pleistocene intramontane plain surrounded by carbonate ridges in central 116
Italy, is characterized by a complex hydrogeological setting where several high-discharge springs 117
feed the Aterno-Pescara hydrological basin, including the Sagittario River (Celico, 1978; Boni et 118
al., 1986; Conese et al., 2001; Barbieri et al., 2005; Desiderio et al., 2012). 119
The Sagittario River valley is the southern part of the Sulmona Basin, which extends over 120
approximately 143 km2 in the SSE-NNW direction (Fig. 1). It is a large depression filled with thick 121
(up to 500 m) fluvio-lacustrine deposits of Pleistocene-Holocene age and completely surrounded 122
by carbonate ridges (Miccadei et al., 1998). Previous studies (Desiderio et al., 2003, 2005b, 2012) 123

5
have identified the hydrogeological units (Fig. 1) of the alluvial fans, river terrace deposits, 124
lacustrine-marshy deposits and recent alluvial deposits. 125
The alluvial fans, including detrital deposits, have a thickness ranging between 10 and 100 m and 126
are located at the base of the surrounding carbonate ridges. This unit is extremely permeable and 127
hosts a single level aquifer, frequently connected with the surrounding hydrogeological units. Multi-128
level aquifers and perched aquifers are locally present, due to silt levels and to wedging by 129
lacustrine deposits. 130
The fluvial terrace forms a hydrogeological unit outcropping along the valley borders that is 131
composed of gravels and conglomerate deposits in a sandy-silty matrix with silty-sand and silt-clay 132
lenses with peat and that has a thickness varying from 10 to 60 m. This unit, similar to the alluvial 133
one, is extremely permeable, and it hosts a single-level aquifer that can occasionally be multi-level. 134
The lacustrine-marshy hydrogeological unit underlies the alluvial deposits and outcrops at the 135
edges of the narrow valleys of the main water courses, with a thickness up to some hundred 136
meters. The unit consists mainly of silts in a sandy and clayey matrix and has levels and lenses of 137
sands, gravels, clays, peats and volcanic clasts. It acts as a low permeability barrier and limits 138
groundwater transfers from the limestone aquifers and to the recent alluvial aquifer. Locally, the 139
lacustrine-marshy aquifer unit hosts confined aquifers within the more permeable deposits, 140
occasionally having artesian conditions. 141
The recent alluvial hydrogeological unit is located close to the main rivers and it is composed by 142
sandy and clayed silts with gravel-sand lenses of varying extension, having a thickness varying 143
between 10 and 20 m. The unit hosts a shallow aquifer interconnected with river surface waters, 144
with the alluvial fan unit and in some cases, with the underlying lacustrine-marshy unit and the 145
adjacent limestone aquifers. 146
The carbonate hydrogeological unit is located along the ridges surrounding the basin, composed of 147
limestone, marly limestone and dolomite, with a thickness ranging from a few hundred to a few 148
thousand meters. High hydraulic conductivity is mostly due to fracturing/fissuring and conduit flow, 149
as well as hosting aquifers with huge water supply potential. 150

6
Unconfined aquifers recognized in the alluvial units and within the multi-layer aquifer in the 151
lacustrine-marshy deposits are recharged by rain, runoff, and irrigation as well (Desiderio et al. 152
2003, 2012). Locally, the deeper levels are fed by regional carbonate aquifers. Available δ18O and 153
δD data (Desiderio et al., 2005a) indicate that sand-gravel aquifers are recharged at average 154
isotopic altitudes of 800 m a.s.l. The flow path of the alluvial aquifer moves from the alluvial fans 155
toward the terrace edges. The main aquifer drainage pathways are along some paleo-riverbeds 156
into recent alluvial deposits. The Sagittario River acts as the final destination for groundwater (Fig. 157
1). 158
A hydrological gauging station (Capocanale site) has been recording the discharge of the 159
Sagittario River since 1927. The monthly discharge regime shows two peaks in April and in 160
December (with a mean highest discharge of 8 m3s-1), with the lowest values from June to August 161
(from 2.5 to 4 m3s-1), demonstrating the base flow contribution from groundwater. In the last two 162
decades, the lowest mean discharge values were measured along the hydrologic year. The natural 163
regime of the river has been highly affected since the first half of the 20th century by a hydropower 164
dam located upstream, as shown by daily hydrographs (Fig. 2) related to years 2006, 2008, 2010 165
and 2011. In the first half of the year and in the last months of the year, weekly changes 166
correspond to the use of hydropower, which is active during weekdays, causing peaks in 167
discharge. Conversely, during the weekend, there is no overflow, and water is stocked in the dam, 168
as reflected in a shortage of approximately 50% of the river discharge. This artificial regime has 169
been employed for several years, but during 2010, the hydropower plant was stopped. As a result, 170
the 2010 hydrograph represents a more "natural" discharge, with peaks corresponding to runoff 171
contribution and low-discharge phases during dry summer periods. In addition, from June to 172
September, river diversions for irrigation purposes are active along the river, are concentrated 173
downstream from the gauging station, and are limitedly affecting the discharge hydrographs at the 174
Capocanale station. The selected years can be considered representative of rainy (2006), average 175
(2010-2011) and dry years (2008). 176
177
178

7
METHODS 179
A multidisciplinary approach has been adopted for evaluating the hydrological, hydrochemical and 180
biological status of the Sagittario River according to EU requirements (EC, 2012) by analyzing the 181
degree of connectivity between surface flowing waters and the underlying aquifers, its influence on 182
the nitrogen cycle and the response of freshwater invertebrate fauna. Three sampling surveys 183
were performed in different hydrological and environmental conditions along the hydrological year: 184
(1) between November and December 2010, when runoff and groundwater discharges are 185
progressively increasing the river discharge (initial recharge phase); (2) in June 2011, when the 186
highest river discharge is recorded (peak phase) and influenced by snow melting and abundant 187
runoff (Fig. 2); and (3) between August and October 2011, corresponding to the minimum 188
discharge value (exhaustion phase), which persists until fall, when the new rainfall season starts. 189
To assess the relationship between the Sagittario River and the water table, for each sampling 190
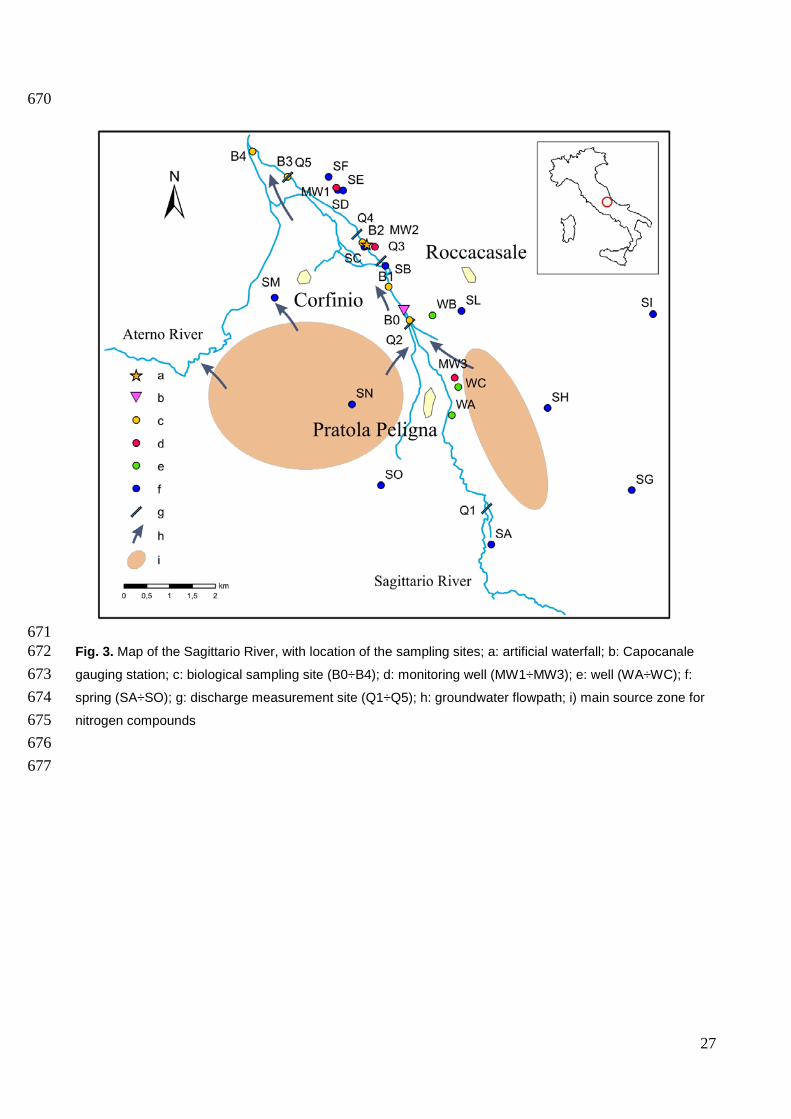
time, river discharge was measured with a portable flow meter at five gauging stations (Fig. 3). 191
Water samples were collected along the riverbed in 19 sites. Groundwater from 13 springs (S) and 192
3 wells (W) located in the Sagittario River alluvial plain was sampled (Fig. 3, Table 1). Due to the 193
limited availability of wells in the study area, 3 new monitoring wells (MW) were drilled close to the 194
Sagittario River (up to 300 m away, Fig. 3), to a depth of 26, 30 and 24 m. MW1 showed an 195
artesian condition, flushing groundwater up to 2 m above ground level. 196
Groundwater from monitoring wells MW2 and MW3 was sampled using Solinst straddle inflatable 197
packers for depth profiles. Three depths were monitored for each well: MW2 was investigated at 7, 198
12 and 18 m below the groundwater table (b.g.t.) and MW3 at 6, 14, and 20 m b.g.t. Temperature, 199
oxidation reduction potential (ORP), pH and electrical conductivity (EC) were measured using the 200
Oxi 349i/SET multiparametric probe. In the monitoring wells MW2 and MW3, vertical logs of the 201
same parameters were also obtained using a Hydrolab Reporter multiprobe 5.0. 202
Laboratory analyses including major ions δ18O and δD were performed. For chemical analysis, 50 203
mL of water were collected in PVS bottles, filtered (0.45 µm Millipore Filters) and then acidified with 204
0.1 N HNO3 to pH≤ 5. 205

8
Major ions analyses were performed by chromatographic technique (Metrohm 761 Compact IC) 206
with an analytical precision of ±0.5%. Groundwater and surface water samples (100 mL) for δ18O 207
and δD were analyzed at the Isotope Lab at the University of Parma. The international standard 208
used was the Vienna Standard Mean Ocean Water (VSMOW) for oxygen and hydrogen isotopes. 209
The analytical error was ±0.1‰ and ±1‰, respectively. The normalization procedure followed the 210
two-points or multi-point method based on a linear regression. The raw measured delta values for 211
at least two different analyzed standards were plotted on the x-axis, and the “true” commonly 212
accepted delta values, expressed in VSMOW scale, were plotted on the y-axis, a regression line 213
that covers a different range of delta values depending on the delta values of the standards 214
analyzed. During the normalization process, the raw delta value of the sample (measured versus 215
the working gas standard) was multiplied by the slope, and the value of the intercept was then 216
added (Skrzypek, 2013). 217
Samples for water chemistry from both surface flowing waters and hyporheic sites were collected 218
and stored in polyethylene bottles and kept refrigerated until analyzed by A.R.T.A. Abruzzo 219
(Department of L’Aquila). Dissolved oxygen, temperature, pH, oxygen concentration, and electrical 220
conductivity were measured in the field with a multiparametric probe (YSI 556 MPS). 221
The invertebrate sampling was conducted along with the hydrological and hydrochemical 222
monitoring surveys. Sampling sites were located as close as possible to the corresponding 223
discharge measurement sites. Faunal samples were collected at five sites along the Sagittario 224
River downstream of the dam (Fig. 3) by selecting an additional “reference site” (site B0) in a river 225
stretch that did not undergo rectification, river bank concretion, or riparian devegetation. Site B1 226
was placed at the terminal fan of the river-gaining sector where several alterations co-occurred, 227
such as rectification, river bank concretion, and riparian devegetation; site B2 was located in the 228
most altered river stretch, where the river path underwent diversion, was artificially perched, and 229
riparian vegetation was completely erased; B3 was located 500 m downstream of B2, where the 230
river comes back to its natural path after an artificial waterfall (Fig. 3); and B4 was located 1 km 231
upstream from the confluence to the Aterno River, river banks had consistent riparian vegetation 232
but rectification was still present. Sites B3 and B4 were the ones less affected by changes in river 233

9
morphology. To assess spatial and temporal biodiversity patterns in the hyporheic zone, each site 234
was sampled at 50 cm below the river bed and samples were taken using a Bou-Rouch pump (Bou 235
and Rouch, 1967) and mobile pipes were hammered at each sampling point-depth. For each site, 236
three spatial replicates were taken for three times along the hydrological year to maximize the 237
measured species richness along habitat heterogeneity for each transect. The piezometric head 238
was measured at each sampling date and site using the T-bar (Malard et al., 2002). 239
Faunal samples were extracted by filtering 10 L samples of water and fine sediments collected 240
through a hand net (mesh size of 60 μm) and then preserved in 60% ethyl alcohol in the field. All 241
fauna samples were accompanied by physicochemical analyses. The Crustacea Copepoda 242
resulted in the most abundant group in the hyporheic sites by far, and for this reason, they were 243
selected as the target group. Copepods were sorted, counted, identified to species level, and 244
assigned to two ecological categories (obligate groundwater dwellers, i.e., stygobionts, and non-245
obligate groundwater dwellers, i.e., non-stygobionts) according to the definition given by Galassi 246
(2001) and Galassi et al. (2009). Supplementary samples were also taken at the benthic surface of 247
riverbed sediments by sampling 25 x 25 cm2 on three different microhabitats recognizable along 248
the transect, to cover the highest habitat heterogeneity at each site. Benthic samples were filtered 249
through a drift net and preserved in 60% ethyl alcohol on the field. Individuals, predominantly 250
belonging to insect orders and macrocrustaceans, were sorted and identified at the species level 251
when achievable. The Margalef’s index (Margalef, 1958) was calculated from the total number of 252
species counted and their abundance from both hyporheic and benthic samples by cumulating 253
abundances and species richness of spatial replicates and sampling dates per sampling site. 254
255
RESULTS 256
River discharge 257
In the first survey, the highest value of 7.7 m3s-1 was measured downstream (Q4), upstream from 258
the confluence with the Aterno River, and the lowest value of 4.9 m3s-1 was measured at Q3 (Fig. 259
4). During the second survey, the highest discharge value of approximately 10.7 m3s-1 was 260
measured at Q2, at the confluence with the Velletta stream, and the lowest value at Q3. Q4 261

10
showed the highest discharge value, even in the third survey (3.4 m3s-1), whereas in Q1, close to 262
the Acqua Chiara spring (SA, Fig. 3), the lowest value of 1.8 m3s-1 was observed. According to the 263
hydrological cycle in central Italy, the seasonal variability among collected data showed a 264
significant increase in discharge between the first two surveys, followed by a decrease in the third 265
survey. In addition, data related to Q3 were collected 500 m downstream from the Capocanale 266
gauging station. A comparison between the measured values and the calculated values were 267
similar, with an error for each survey below 10% (Fig. 2). 268
In the first two surveys, discharge differences along monitoring sites followed the same scheme, 269
with alternation of increase and decrease of river discharge having comparable magnitude and 270
highlighting a similar response in terms of surface water-groundwater exchanges. The third survey 271
values are affected both by natural lower discharge and by additional human diversions for 272
agricultural purposes; consequently, discharge changes along the river showed a lower magnitude, 273
becoming negligible. Furthermore, it was possible to identify small-scale differences in river 274
discharge, where local river discharge increased or decreased (Fig. 4). In upstream stretches close 275
to the SA spring, up to Q2, the Sagittario River discharge increased at an average of approximately 276
0.7 m3s-1; from Q2 to Q3 the discharge decreases of approximately 2.7 m3s-1 (33% of the total river 277
discharge). Downstream from Q3, where the artificial waterfall is located, the discharge increased 278
to 2.6 m3s-1, with lower values during the third survey. Between Q4 and Q5, upstream from the 279
confluence with the Aterno River, local mean discharge lowered to approximately 1.1 m3s-1. 280
281
Water chemistry 282
All sampled waters in wells, springs and monitoring wells showed pH values close to neutrality or 283
slightly alkaline (from 6.84 to 8.02) (Table 1), except for the well WC during the second survey in 284
June 2011. Seasonal changes were observed in surface water temperature, with an average of 285
6.9°C in the first survey, and 14.7°C in the second and third surveys. Groundwater showed a less 286
pronounced variation, between 7.9°C in the first survey and 12.5°C in the second and third 287
surveys. The electrical conductivity ranged between 255 and 790 µScm-1 in springs, and between 288
478 and 693 µScm-1 in groundwater, including vertical logs. ORP ranged from +37 to +231 mV in 289

11
spring, as expected for oxygenated waters, except for SC in the second survey. Wells (W) and 290
monitoring wells (MW) were locally characterized by low ORP values until -188 mV, indicating a 291
reducing environment. Vertical data logs of MW2 and MW3 (Fig. 5) showed sharp changes in pH, 292
ORP, and electrical conductivity between 17 and 22 m b.g.l. 293
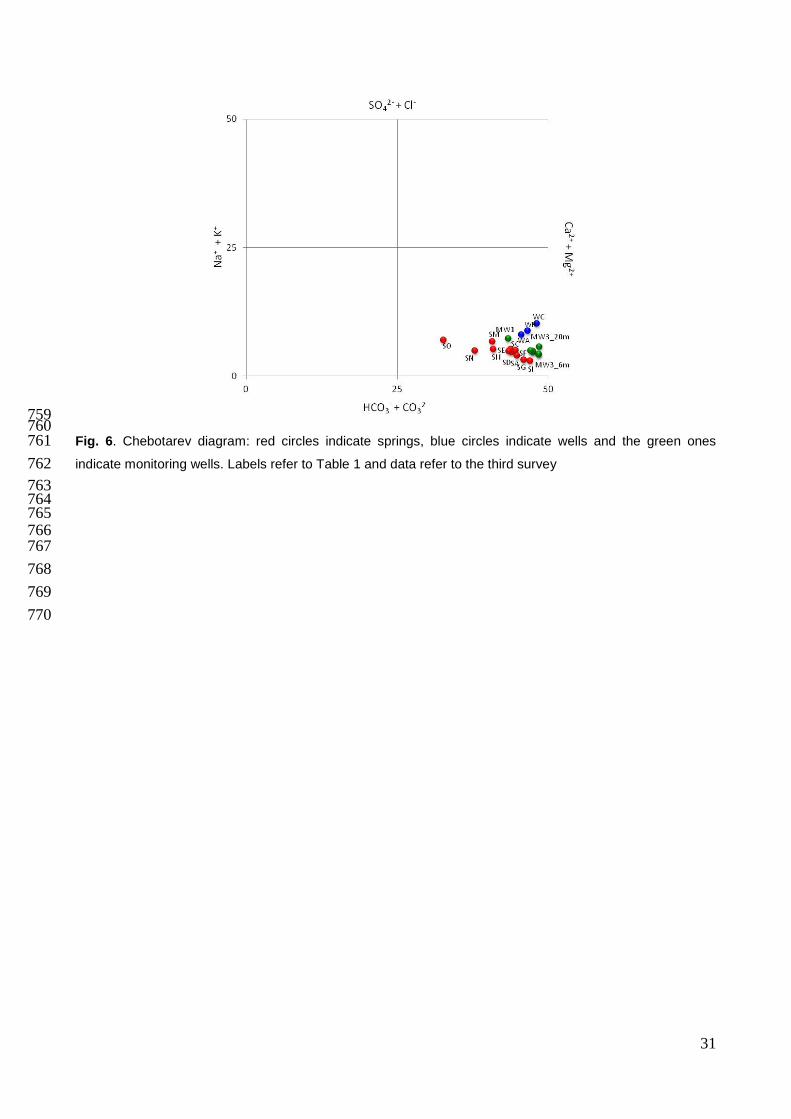
The geochemical facies (Fig. 6) of groundwater and surface water is mainly a Ca-HCO3 type. 294
Slight variations of measured values of SO42-, Ca2+, Mg2+, Na+ and K+ were also detected both 295
among the three seasonal surveys and across different sampling sites. Wells WB and WC showed 296
a steady chemical facies during the whole sampling period. Conversely, marked differences of 297
Ca2+ (65.9 mgL-1 in WB and 47.8 mgL-1 in WC during the third survey) and Cl- concentrations (11.9 298
mgL-1 in WB and 5.8 mgL-1 in WC during the third survey) were recorded in the same samples. WA 299
showed an intermediate chemical composition and a sharp seasonal variability. Differences in 300
spring waters were observed between the group SA-SC-SF close to the river bed and those at 301
higher elevation (SM, SN, SO) that showed a higher SO42-, Cl-, Na+, and K+ content. Monitoring 302
wells (MW) did not show chemical variation through vertical sampling, except MW3, which at the 303
deepest sampling point had a significant increase in HCO3-, K+ and NH4
+ content. MW3 showed 304
higher values of NH4+ and PO4
3- (up to 30.6 and 1.2 mgL-1, respectively), whereas the same ions 305
have not been found in MW1 and MW2. MW1 had a unique chemical composition, showing the 306
highest values of Ca2+ (68.9 mgL-1), SO42- (10.1 mgL-1) and Cl- (18.5 mgL-1). 307
During the three seasonal surveys, no clear trends were observed in nitrate concentration, ranging 308
from 0.1 to 100.9 mgL-1. Therefore, wide variability among springs, wells and monitoring wells was 309
proved by standard deviation (Table 2). The lowest average value was detected during the second 310
survey. The monitoring wells did not show differences along the water column at the temporal 311
scale. Conversely, slight differences among springs at different elevation were observed (Fig. 7). 312
All springs located on the left side of the Sagittario River (between 340 and 1200 m a.s.l.) showed 313
remarkable concentrations of nitrate. A similar trend, even if less pronounced, was observed in 314
wells located on the right border. 315
Ammonium concentrations showed the similar standard deviation values to the averages (Table 2). 316
Values ranged from under the instrumental detection limits to 30.9 mgL-1 of NH4+ observed in few 317

12
springs. Detectable ammonium concentrations were found only in SC (1.3 mgL-1, first survey) and 318
SE (2.5 mgL-1, second survey). Some wells and monitored wells revealed significant ammonium 319
concentration, reaching 21 and 14 mgL-1 in WB and WC, respectively. Low ammonium 320
concentrations were measured in MW1 and MW2 during the second survey, but they were under 321
the instrumental detection limits in October. MW3 showed high ammonium concentrations along 322
the entire sampling survey, with the maximum value measured at the deepest screen sampled (up 323
to 30 mgL-1). At the spatial scale (Fig. 8), the highest ammonium concentrations were measured 324
close to the river bed, mostly along the right bank, especially in wells and monitoring wells. 325
326 Isotope approach 327
The isotopic compositions of the sampled waters collected during all monitoring surveys showed a 328
variability range between -59.5/-74.8‰ for δD, and -8.6/-11.3‰ for δ18O (Table 3). Differences in 329
isotopic composition among sampling points were also observed over time, in particular from 330
springs SA, SC, SE and SF. 331
The isotopic data are consistent with the literature relationships for the global average (GML, 332
Rozanski et al., 1993), central Italy (cIML, Longinelli and Selmo, 2003) and the Abruzzi region 333
(RML, Barbieri et al., 2003). Moreover, such a straight line (Fig. 9) is also consistent with the 334
equation (δD = 5.05 δ18O – 16.04) concerning samples of the upper valley of the Sagittario River 335
(Petitta et al., 2010). Three different isotope signals representative of the study area were found in 336
groundwater samples (Fig. 9). A group of waters including spring SI and well WC showed the less 337
enriched δ18O and δD values: -10.9‰ and -71.4 in SI, -11.3‰ and -74.8‰ in WC (third survey). 338
More enriched values referred to a different group of springs (SN, SO and SM) (-8.6‰ δ18O, -339
59.5‰ δD in SN, third survey) and well WB (-9.1‰ δ18O, -63.5‰ δD, third survey). The third group 340
included springs and monitoring wells characterized by intermediate signals (-10‰ δ18O, -66.9‰ 341
δD in SC). No significant variations were detected among the vertical logs in MW2 and MW3. 342
343 344
345

13
Species diversity and abundance of freshwater invertebrates 346
The distribution pattern of benthic and hyporheic invertebrate fauna along the Sagittario River from 347
the third survey (Fig. 10) showed significant differences. Both benthic macroinvertebrates and 348
hyporheic copepods displayed higher species diversity (expressed as Margalef’s index) in the 349
uppermost stretch (B0), characterized by the presence of a consistent riparian buffer strip. 350
Although located downstream from the dam, the stretch showed microhabitat diversification, with 351
riffle-pool sequences, macrophytes, and mosses close to river banks, where current velocity was 352
lower. Macroinvertebrate diversity dramatically decreased in site B1, where river channel 353
rectification, bank concretion, very high current velocity, and the total absence of riparian 354
vegetation contributed to an impoverishment of microhabitats. Moreover, the erosion of bed 355
sediments created a benthic habitat inhospitable for almost all the macroinvertebrate taxa. 356
Conversely, the highest stygobiotic copepod species richness was found at this site. In 357
downstream river stretches (from B2 to B4), there was a significant decrease of both surface and 358
hyporheic diversity, which reached the lowest values in the terminal stretch (site B4), upstream 359
from the confluence with the Aterno River. 360
361 DISCUSSION 362
Surface water and groundwater interactions 363
According to the data of this study along the Sagittario River, in the upstream stretch between Q1 364
and Q2, an average increase in discharge of approximately 0.7 m3s-1 was recorded, whereas in the 365
second stretch, between Q2 and Q3, a water loss of approximately 33% of the total river discharge 366
was estimated (up to 2.7 m3s-1). Downstream, in the third stretch (Q3-Q4), immediately after the 367
artificial perched sector and downstream from the artificial waterfall occurrence, the river regained 368
nearly all the previous water loss. In the fourth stretch, between Q4 and Q5, a new downwelling 369
zone showed a water loss of approximately 1.1 m3s-1, with an average discharge of 5.4 m3s-1 (Fig. 370
4). 371
Along the river, two discharge peaks were observed, in correspondence with Q2 and Q4, with 372
particular reference to the first and second surveys (Fig. 4). The second survey showed higher 373

14
values for Q1 and Q2 in comparison to the first survey; nevertheless, the downstream sites Q3 and 374
Q4 showed identical values and trends. 375
The correspondence of exchanges values from Q3 to Q5, starting from different upstream values, 376
demonstrates that discharge in these stretches is largely affected by river modifications (e.g., the 377
artificial waterfall), which forces the hydraulic gradient between surface waters and water table. 378
Consequently, along this river stretch, the exchange amount of up to 2.5 m3s-1km-1 was magnified 379
by artificial riverbed elevation and morphology, which enhanced river-aquifer interactions, at 380
seasonal and annual scales. In the summer season (third survey), the lower discharge of the river 381
did not allow high-rate exchanges between Q2 and Q3, but the relationships among the river and 382
the groundwater table remained unchanged in downstream stretches, indicating a gaining sector 383
between Q3-Q4 and a losing sector in the last stretch Q4-Q5. 384
385
Groundwater conceptual model 386
Groundwater flow paths were consistent with a complex multilayer system; in fact, the study area is 387
characterized by (I) a shallow alluvial aquifer, fed by zenithal recharge and river losses, (II) a local 388
perched terraced aquifer and (III) a deep carbonate aquifer. They are separated by a lacustrine-389
marshy aquitard with sand and gravel lenses with high permeability (Fig. 11). Nevertheless, along 390
the river, the shallow aquifer interacts with both surface water and the hyporheic zone, immediately 391
below the river bed, as supported by multi-level sampling results. A high hydrological connectivity 392
has been identified between the Sagittario River and this complex groundwater system, 393
determined by exchanges between the river and the shallow alluvial aquifer. The deep aquifer-394
aquitard system, fed by high-altitude surrounding carbonate aquifers, contributed to river discharge 395
in upwelling zones where shallow and deep groundwater contributions were mixed due to a limited 396
thickness and/or higher permeability deposits of the aquitard (Fig. 11). Similarly, hydraulic 397
interactions also allowed the river to feed water back to the water table artificially disconnected 398
from the river, just as upstream the waterfall in the perched stretch between Q2 and Q3 (Figs. 3, 399
4). 400

15
Environmental isotopes, δ18O and δD, provided evidence of different recharge areas. The more 401
depleted isotope values (WC) indicated the existence of a deep flow path related to the carbonate 402
aquifer, having a similar recharge altitude of the spring SI, which was located in the upper terrace 403
on the right river bank (1275 m a.s.l.). A second group showing more enriched isotope signals 404
included surface water and groundwater coming from local and perched springs (SN, SO, SM) and 405
a WB well denoted a local recharge effect related to the terraced aquifer. The group showing 406
intermediate isotope signals, such as the monitoring wells and WA, suggests potential mixing 407
processes are occurring through the lacustrine-marshy deposits between the deep and shallow 408
aquifers. 409
410
The nitrogen cycle at floodplain scale 411
In this hydrogeological framework, it was possible to determine the source and fate of nitrogen 412
compounds in groundwater and surface water (Fig. 11). High nitrate concentrations were detected 413
in springs located on the terraced aquifer (up to 100 mgL-1 NO3-) at an elevation higher than the 414
Sagittario River alluvial plain. The nitrogen load is due to anthropogenic activities, mainly related to 415
agricultural activities occurring at the floodplain scale, using synthetic fertilizers and/or livestock 416
wastes. Nitrogenous compounds can transfer toward the plain through infiltration and/or leaching 417
processes following the shallow groundwater flow paths (Fig. 11), reaching wells and springs 418
closer to the riverbed (maximum concentrations: 100 mgL-1 NO3- and 31 mgL-1 NH4
+). The pollution 419
of surface river waters was prevented upstream, where the riparian buffer strips were still present. 420
Conversely, in the downstream stretches, river waters in upwelling zones (Fig. 11U) received 421
nitrogen from the shallow aquifer and from the deep flow path, reaching a maximum value of 422
approximately 5.4 and 1 mgL-1 of nitrate and ammonium (1.2 mgL-1 as N-NO3-, 0.6 mgL-1 as N-423
NH4+), respectively (Fig. 12). Nitrogen compounds detected in river water are likely due to 424
anthropogenic practices that, to manage the plain against flood risks, caused the eradication of 425
riparian vegetation (which is useful in lowering nitrogen contamination). In deeper flow paths, 426
however, the nitrogen content affecting groundwater can reach deep wells and monitoring wells as 427
ammonium, according to the lithology of the deep aquifer-aquitard, where peat and clay deposits 428

16
determine anoxic conditions, reduce infiltration processes and most likely increase denitrification. 429
Moreover, ammonium from the deep aquifer can locally transfer to the shallow aquifer too, where 430
the aquitard thickness lowers or disappears. As a result, a mixing between nitrates and ammonium 431
having different origins was observed (e.g., in WB). Ammonium can also reach river waters, along 432
the upwelling zones (due to the high content of organic matter, such as peat, in lacustrine deposits: 433
Desiderio et al., 2010). Following groundwater flowpaths, in downwelling zones located 434
downstream (Fig. 11L), nitrogen-enriched surface waters can transfer nitrates to the shallow 435
aquifer, increasing nitrogen content of local springs fed mainly by losses of surface water. 436
Consequently, the complex hydrogeological setting may explain nitrate or ammonium pollution at 437
the subsurface, as ammonium is uncommon in oxidized environments. Thus, the ecological 438
conditions of the Sagittario River in some upwelling zones can be affected by the presence of a 439
non-negligible concentration of nitrogen compounds. 440
441
Preliminary evaluation of freshwater invertebrate response 442
The distribution pattern of benthic and hyporheic invertebrate fauna along the Sagittario River (Fig. 443
10) predominantly followed the broad-scale pattern of water pollution as derived from the increase 444
in nitrate and ammonium concentrations. Both benthic macroinvertebrates and hyporheic 445
copepods displayed high species diversity, as measured by Margalef’s index, in the gaining sector 446
of the river (B0), characterized by higher microhabitat diversification, presence of riparian 447
vegetation and low concentrations of nitrate and ammonium. Macroinvertebrate diversity 448
decreased in site B1, and the Margalef’s index became half of that measured in site B0, due to 449
microhabitat impoverishment as a reflection of river channel rectification and concretion and very 450
high current velocity. Conversely, hyporheic copepod species richness was increased slightly in 451
site B1 by groundwater upwelling from the shallow aquifer, which enriched the site in stygobionts 452
(species which complete the entire life cycle in true groundwater habitats), such as the cyclopoids 453
Diacyclops italianus, Diacyclops maggii, and Eucyclops intermedius and the harpacticoids 454
Parastenocaris sp., Nitocrella psammophila, and Parapseudoleptomesochra italica. In downstream 455
river stretches, there was a significant decrease of macroinvertebrate diversity, with the lowest 456

17
species richness and abundance measured at site B4 (Fig. 10 a, b). The worsening of water quality 457
due to the input of nitrogenous compounds (especially ammonium) from the hypoxic aquifer 458
highlighted a continuous increase of the overall environmental alteration along an upstream-459
downstream gradient. In downstream stretches, the input of ammonium in the hyporheic zone 460
caused a higher response to pollution by hyporheic copepods, which almost disappeared at site B4 461
(Fig. 10 c, d). 462
Epibenthic macroinvertebrates and copepod assemblages were sensitive to both water pollution 463
and hydraulic alteration of the analyzed stretches, i.e., they were able to integrate the 464
environmental information derived from all stressors together. Hydraulic change among river 465
stretches was the major explanatory parameter for variation in total abundances; macrobenthic 466
abundances markedly increased in the perched stretch of B3 and therefore decoupled from 467
species richness, and the dominance of a single eurytopic species, the amphipod 468
Echinogammarus tibaldii, was observed. Hyporheic copepod abundances showed a marked 469
decrease in the same stretch of B3, where only a few stygoxene species were present. The low 470
abundances of the hyporheos were related to the reduced availability of subsurface habitats that 471
resulted in unsuitable colonization by surface copepods due to the aggressive erosion of 472
sediments. In addition, the losing facies of this artificial perched site determined an interruption in 473
connectivity between surface and groundwater, as reflected in the total absence of stygobiotic 474
species. The deterioration of the environmental conditions did not allow a recovery in assemblage 475
structure complexity, which remained compromised along the whole downstream river sector. 476
In the present study, nitrogen content also represented dependent variables following groundwater 477
and surface water exchanges. This study demonstrated that in river stretches with high ammonium 478
concentration (B3-Q5, Figs. 3, 4, 10, 12) and persistence (B4), the effects on abundance of 479
hyporheic meiofauna and epibenthic macroinvertebrates were evident. Hyporheic assemblages 480
were also sensitive in terms of species richness more than surface benthic invertebrates because 481
lower oxygen concentration in the hyporheic zone maintained over longer time reduced the 482
nitrogenous compounds, which were notoriously more toxic than the oxidized forms (Camargo and 483
Alonso, 2006; Dehedin et al., 2013). Conversely, nitrate contamination in the shallow aquifer was 484

18
negligible due to the peculiar conditions of the groundwater flow path, and its detrimental effect on 485
meiofauna was observed only at very high concentrations (Di Lorenzo and Galassi, in press). 486
487
CONCLUSIONS 488
This study analyzed the interaction among river flow alteration, the degree of connectivity between 489
surface flowing waters and the underlying aquifers, and the amount of water pollution by 490
nitrogenous compounds, and it included a preliminary evaluation of the large-scale response of 491
freshwater invertebrate fauna among stretches downstream from a hydropower plant. 492
The hydrological and hydrochemical information gathered in the present study provided information 493
to advance a conceptual model explaining the relationships between groundwater coming from a 494
multilayer aquifer-aquitard system, and surface water affected by morphological and discharge 495
regime disturbances. This hydrogeological setting has significant effects on the distribution of 496
nitrogen pollution along the stretches analyzed and on the invertebrate riverine biota. This altered 497
situation was heavily reflected in the worsening of the ecological status of the Sagittario River, as 498
measured by benthic and hyporheic invertebrate diversity and abundance, especially in gaining 499
stretches where groundwater is polluted by nitrogenous compounds. The GDEs are known to be 500
highly sensitive to such alterations (Dole-Olivier, 2011). 501
The alternation of gaining and losing stretches along the Sagittario River, even if allowing 502
significant water exchanges with the shallow alluvial aquifer, was obscured by anthropogenic 503
changes having both short- and long-time frequencies. Processes that lead to river-aquifer 504
interactions affecting river discharge are forced by anthropogenic practices concerning 505
geomorphologic disturbance and hydrologic regime alteration, mainly due to rectification of the 506
river path, artificial perched riverbed stretch (e.g., the artificial waterfall), summer withdrawals due 507
to channel diversions for agricultural use, and hydropower dam activity. At the floodplain scale, the 508
nitrogen cycle was affected by the complex groundwater flow system and also by anthropogenic 509
changes. The nitrogen content in groundwater and surface water did not reach high contamination 510
levels; conversely, significant concentrations of ammonium reached surface waters in downstream 511
stretches, fed by the hypoxic deep aquifer enriched of reduced nitrogenous compounds via the 512

19
hyporheic zone. This hydrogeological scenario, superimposed to nitrogen loads entering the 513
aquifer, was inexorably reflected on the biological integrity of local GDEs (Stanford and Ward, 514
1993). The integrity of the GDEs was thus threatened by multi-faceted pressures, as suggested by 515
the degree of occurrence of copepods in the hyporheic zone among stretches, and a similar trend, 516
although less marked, was also observed in the macroinvertebrate benthic fauna. Artificial perched 517
stretches are detrimental in dammed rivers for several reasons and determine a worsening of the 518
ecological status of river waters, as indicated by the severe decline of invertebrate biodiversity both 519
above and below the riverbed surface. Only tolerant species may survive in such conditions, 520
becoming dominant, if not the only “pioneer” species able to colonize the river in such extreme 521
conditions (such as, in this case, Echinogammarus tibaldii). 522
The interaction between groundwater and surface water in rivers by means of upwelling, outwelling 523
and downwelling exchanges represents a crucial “dimension” in urgent need to be evaluated for a 524
correct integrated approach to river management issues (Boulton et al., 1998; Tomlinson and 525
Boulton, 2010; Dole-Olivier, 2011; Di Lorenzo et al., 2013). 526
527
528
ACKNOWLEDGEMENTS 529
This study was supported by a project granted by Provincia dell’Aquila. We are much indebted to 530
Valerio Saladini, Lorenzo Pasqualini, Nicola Tirozzi and Matteo Mammone for field work 531
assistance. The Regional Agency for Environmental Protection (A.R.T.A.), Department of L’Aquila, 532
is greatly acknowledged for having supported the chemical monitoring. 533
534
REFERENCES 535
Banks EW, Simmons CT, Love AJ, Shand P (2011) Assessing spatial and temporal connectivity 536
between surface water and groundwater in a regional catchment: Implications for regional scale 537
water quantity and quality. J Hydrol 404:30–49 538
Barbieri M, D’ Amelio L, Desiderio G, Rusi S, Marchetti A, Nanni T, Petitta M, Rusi S, Tallini M 539
(2003) Gli isotopi ambientali (18O, 2H and 87/86Sr) nelle acque sorgive dell’Appennino Abruzzese: 540

20
considerazioni sui circuiti sotterranei negli acquiferi carbonatici. Atti I° Congresso Nazionale AIGA, 541
Chieti, 19-20 February 2003, pp 69–81 542
Barbieri M, Boschetti T, Tallini M (2005) Stable isotopes (2H, 18O and 87/86Sr) and hydrochemistry 543
monitoring for groundwater hydrodynamics analysis in a karst aquifer (Gran Sasso, Central Italy). 544
Appl Geochem 20(11):2063–2081 545
Boni C, Bono P, Capelli G (1986) Schema Idrogeologico dell’Italia Centrale. Mem Soc Geol It 546
36:991–1012 547
Bou C, Rouch R (1967) Un nouveau champ de recherches sur la faune aquatique souterraine. C R 548
Acad Sci 265:369–370 549
Boulton AJ, Findlay S, Marmonier P, Stanley EH, Valett HM (1998) The functional significance of 550
the hyporheic zone in streams and rivers. Ann Rev Ecol Syst 29:59–81 551
Camargo JA, Alonso Á (2006) Ecological and toxicological effects of inorganic nitrogen pollution in 552
aquatic ecosystems: A global assessment. Environ Int 32:831–849 553
Celico P (1978) Schema idrogeologico dell’Appennino carbonatico centro-meridionale. Memorie e 554
Note dell’Istituto di Geologia Applicata, Università di Napoli 14:5–97 555
Cey EE, Rudolph DL, Aravena R, Parking G (1999) Role of the riparian zone in controlling the 556
distribution and fate of agricultural nitrogen near a small stream in southern Ontario. J Contam 557
Hydrol 37:45–67 558
Conese M, Nanni T, Peila C, Rusi S, Salvati R (2001) Idrogeologia della Montagna del Morrone 559
(Appennino abruzzese): dati preliminari. Mem Soc Geol It 56:181–196 560
Dehedin A, Maazouzi C, Puijalon S, Marmonier P, Piscart C (2013) The combined effects of water 561
level reduction and an increase in ammonia concentration on organic matter processing by key 562
freshwater shredders in alluvial wetlands. Global Change Biol 19:763–774 563
Desiderio G, Rusi S, Nanni T (2003) Idrogeologia e qualità delle acque degli acquiferi della conca 564
intramontana di Sulmona (Abruzzo) Atti del 1° Convegno Nazionale dell’AIGA, 19 – 20 February 565
3003, Chieti, pp 315–342 566

21
Desiderio G, Ferracuti L, Rusi S, Tatangelo F (2005a) Il contributo degli isotopi naturali 18O e 2H 567
nello studio delle idrostrutture carbonatiche abruzzesi e delle acque mineralizzate nell’area 568
abruzzese e molisana. Giornale di Geologia Applicata 2:453–458 569
Desiderio G, Nanni T, Rusi S (2005b) Hydrogeology, aquifer vulnerability and potenzial hazard of 570
the Sulmona plain (Central Apennine, Italy) Fourth Congress on the Protection and Management of 571
Groundwater, Colorno, 21-23/9/2005, GEA Torino, pp 1–7 572
Desiderio G, Rusi S, Tatangelo F (2010) Caratterizzazione idrogeochimica delle acque sotterranee 573
abruzzesi e relative anomalie. Ital J Geosci 129(2):207–222 574
Desiderio G, Folchi Vici D'Arcevia C, Nanni T, Rusi S (2012) Hydrogeological mapping of the 575
highly anthropogenically influenced Peligna Valley intramontane basin (Central Italy). J Maps 576
8(2):165–168 577
Di Lorenzo T, Brilli M, Del Tosto D, Galassi DMP, Petitta M (2012) Nitrate source and fate at the 578
catchment scale of the Vibrata River and aquifer (Central Italy): an analysis by integrating 579
component approaches and nitrogen isotopes. Environ Earth Sci 67:2383–2398 580
Di Lorenzo T, Galassi DMP (in press) Agricultural impact in Mediterranean alluvial aquifers: do 581
groundwater communities respond? Fundam Appl Limnol. 582
Di Lorenzo T, Stoch F, Galassi DMP (2013) Incorporating the hyporheic zone within the river 583
discontinuum: Longitudinal patterns of subsurface copepod assemblages in an Alpine stream. 584
Limnologica. doi: 10.1016/j.limno.2012.12.003 585
Di Marzio WD, Castaldo D, Pantani C, Di Cioccio A, Di Lorenzo T, Sàenz ME, Galassi DMP (2009) 586
Relative Sensitivity of Hyporheic Copepods to Chemicals. Bull Environ Contam Toxicol 82:488–587
491 588
Dole–Olivier M-J (2011) The hyporheic refuge hypothesis reconsidered: a review of hydrological 589
aspects. Mar Freshw Res 62(11):1281–1302 590
Eamus D, Froend R (2006) Groundwater-dependent ecosystems: the where, what and why of 591
GDEs. Aust J Bot 54:91–96 592

22
EC - European Commission (2000) Directive 2000/60/EC of the European Parliament and of the 593
Council of 23 October 2000 establishing a framework for Community action in the field of water 594
policy. Off J Eur Commun L 327:1–72 595
EC- European Commission (2012) Communication from the commission to the European 596
Parliament, the Council, the European economic and social committee and the committee of the 597
regions. A blueprint to Safeguard Europe's Water Resources. Brussels 598
Galassi DMP (2001) Groundwater copepods: diversity patterns over ecological and evolutionary 599
scales. Hydrobiologia 453/454:227–253 600
Galassi DMP, Huys R, Reid JW (2009) Diversity, ecology and evolution of groundwater copepods. 601
Freshwater Biol 54:691–708 602
Hill AR (1996) Nitrate removal in stream riparian zones. J Environ Qual 25:743–755 603
Keskin TE (2010) Nitrate and heavy metal pollution resulting from agricultural activity: a case study 604
from Eskipazar (Karabuk, Turkey). Environ Earth Sci 61(4): 703–721 605
Landers DH, Simonich SL, Jaffe DA, Geiser LH, Campbell DH, Schwindt AR, Schreck CB, Kent 606
ML, Hafner WD, Taylor HE, Hageman KJ, Usenko S, Ackerman LK, Schrlau JE, Rose NL, Blett 607
TF, Erway MM (2008) The Fate, Transport, and Ecological Impacts of Airborne Contaminants in 608
Western National Parks (USA). EPA/600/R-07/138. US Environmental Protection Agency, Office of 609
Research and Development, NHEERL, Western Ecology Division, Corvallis, Oregon 610
Longinelli A, Selmo E (2003) Isotopic composition of precipitation in Italy; a first overall map. J 611
Hydrol 270:75–88 612
Malard F, Dole–Olivier M–J, Mathieu J, Stoch F (2002) Sampling Manual for the Assessment of 613
Regional Groundwater Biodiversity. Available at: http://www.pascalis-project.com (last accessed on 614
17 October 2008) 615
Margalef R (1958) Temporal succession and spatial heterogeneity in phytoplankton. In: 616
Perspectives in Marine biology, Buzzati-Traverso (ed.), Univ. Calif. Press, Berkeley, pp 323–347 617
May R, Jinno K, Tsutsumi A (2011) Influence of flooding on groundwater flow in central Cambodia. 618
Environ Earth Sci 63(1):151–161 619

23
Miccadei E, Barberi R, Cavinato G (1998) La geologia della Conca di Sulmona (Abruzzo, Italia 620
Centrale). Geologica Romana 34:59–86 621
Petitta M, Fracchiolla D, Aravena R, Barbieri M (2009) Application of isotopic and geochemical 622
tools for the evaluation of nitrogen cycling in an agricultural basin, the Fucino Plain, central Italy. J 623
Hydrol 372:124–135 624
Petitta M, Scarascia Mugnozza G, Barbieri M, Bianchi Fasani G, Esposito C (2010) Hydrodynamic 625
and isotopic investigations for evaluating the mechanisms and amount of groundwater seepage 626
through a rockslide dam. Hydrol Process 24(24):3510–3520 627
Poff NL, Richter BD, Arthington AH, Bunn SE, Naiman RJ, Kendy E, Acreman M, Apse C, Bledsoe 628
BP, Freeman MC, Henriksen J, Jacobson RB, Kennen JG, Merritt DM, O’keeffe JH, Olden JD, 629
Rogers K, Tharme RE, Warner A (2010) The ecological limits of hydrologic alteration (ELOHA): a 630
new framework for developing regional environmental flow standards. Freshwater Biol 55(1):147–631
170 632
Rao NS, Subrahmanyam A, Kumar SR, Srinivasulu N, Rao GB, Rao PS, Reddy GV (2012) 633
Geochemistry and quality of groundwater of Gummanampadu sub-basin, Guntur District, Andhra 634
Pradesh, India. Environ Earth Sci 67(5):1451–1471 635
Rozanski K, Araguás-Araguás L, Gonfiantini R (1993) Isotopic patterns in modern global 636
precipitation. Geophys Monogr Ser 78:1–36 637
Salem SBH, Chkir N, Zouari K, Cognard-Plancq AL, Valles V, Marc V (2012) Natural and artificial 638
recharge investigation in the Zeroud Basin, Central Tunisia: impact of Sidi Saad Dam storage. 639
Environ Earth Sci 66(4):1099–1110 640
Skrzypek G (2013) Normalization procedures and reference material selection in stable HCNOS 641
isotope analyses: an overview. Anal Bioanal Chem 405:2815–2823 642
Stanford JA, Ward JV (1992) Management of aquatic resources in large catchments: Recognizing 643
interactions between ecosystem connectivity and environmental disturbance In: Naiman, RJ (ed.) 644
Watershed Management. Springer-Verlag, New York, pp 91–124 645
Stanford JA, Ward JV (1993) An ecosystem perspective of alluvial rivers: connectivity and the 646
hyporheic corridor. J N Am Benthol Soc 12:48–60 647

24
Tomlinson M, Boulton AJ (2010) Ecology and management of subsurface groundwater dependent 648
ecosystems in Australia: a review. Mar Freshw Res 61:936–949 649
Wexler SK, Hiscock KM, Dennis PF (2011) Catchment-scale quantification of hyporheic 650
denitrification using an isotopic and solute flux approach. Environ Sci Technol 45:3963–3973 651
652

25
653
654 655
Fig. 1. Geolithological and hydrogeological scheme (modified from Desiderio et al., 2012). 656 a: Regional geolithological map. A- Carbonatic sequence (Meso-Cenozoic); B- “Argille varicolori” formation 657 (Cretaceous-Miocene); C– Terrigenous and evaporitic deposits (Miocene); D- Pelagic clayey deposits (Plio-658 Pleistocene); E- Alluvial deposits (Holocene); F- Thrusts; G- Faults. The rectangle indicates the area detailed 659 in b. b: Hydrogeological scheme. 1- Alluvial fan, 2- Lacustrine and marshy deposits, 3- Ancient fluvial 660 deposits, 4- Recent fluvial deposits, 5- Travertine deposits, 6- Carbonatic deposits, 7- piezometric contour 661 lines, 8- main groundwater flow lines of the alluvial plain, 9- groundwater flow divide 662
663

26
664 665 Fig. 2. Comparison of daily discharge (m3s-1) for 2006, 2008, 2010 and 2011 at the Capocanale gauging 666 station. Circles refer to direct discharge measurements: first survey in red, second survey in yellow and third 667 survey in black 668
669
27
670
671 Fig. 3. Map of the Sagittario River, with location of the sampling sites; a: artificial waterfall; b: Capocanale 672 gauging station; c: biological sampling site (B0÷B4); d: monitoring well (MW1÷MW3); e: well (WA÷WC); f: 673 spring (SA÷SO); g: discharge measurement site (Q1÷Q5); h: groundwater flowpath; i) main source zone for 674 nitrogen compounds 675 676
677

28
Table 1. Physico-chemical parameters. Data refer to the third survey. (n.a.: not available) 678 (T: Temperature; EC: Electrical conductivity; ORP: Oxidation Reduction Potential; m a.s.l.: meter above sea 679 level; m b.g.l.: meter below ground level) 680
681 682 683 684 685 686 687 688 689
690 691 692 693 694 695 696 697 698 699 700 701
702 703 704
705 706
707 708 709 710 711 712 713 714 715 716 717 718 719 720 721 722 723 724 725 726 727 728
Id Elevation Well depth
Water table depth
T pH EC 25°C ORP
m a.s.l. m b.g.l. m b.g.l. °C μS cm-1 mV
SA 314 16.7 7.4 385 130 SC 261 16.3 7.7 378 152 SD 256 15.0 7.5 398 139 SE 257 15.7 7.6 405 137 SF 253 21.7 7.1 578 121 SG 366 22.4 7.6 304 189 SH 359 16.2 7.0 613 223 SI 1275 19.6 7.1 255 231 SL 333 14.2 7.2 436 241 SM 350 16.7 7.7 633 156 SN 351 15.4 7.0 788 176 SO 343 15.6 7.7 749 177 WA 286 4 -1 19.0 7.2 502 -91 WB 274 45 n.a. 25.0 6.9 640 -91 WC 285 58 0 14.5 7.6 515 -188 MW1 255 26 +2 14.2 7.0 603 -60 MW2_7m 265 29 -2 14.0 6.9 508 -125 MW2_12m 265 29 -2 13.8 6.9 509 -134 MW2_18m 265 29 -2 13.6 6.9 510 -141 MW3_6m 287 24 -2 14.2 6.8 551 -93 MW3_14m 287 24 -2 13.8 6.9 551 -100 MW3_20m 287 24 -2 13.7 6.8 681 -126

29
729 730 731 732
733 734 735 Fig. 4. River discharge measured along three seasonal surveys. Horizontal axis approximately corresponds 736 to distance (km), vertical axis to river discharge (m3s-1). Q1÷ Q5 refer to Fig. 3 737 738 739 740 741 742 743 744 745 746 747 748 749 750 751 752
753

30
754 755
Fig. 5. Vertical Logs. Blue line refers to MW2, red line refers to MW3. a: circles refer to CE (µScm-1), squares 756 to temperature (°C). b: rhombus refers to ORP (mV), triangles refer to pH. Location of MW2 and MW3 is 757 shown in Fig. 3 758
31
759 760
Fig. 6. Chebotarev diagram: red circles indicate springs, blue circles indicate wells and the green ones 761 indicate monitoring wells. Labels refer to Table 1 and data refer to the third survey 762
763 764
765 766 767 768 769
770

32
771 Table 2. Statistical parameters of nitrate and ammonium concentrations (mgL-1) during the three sampling 772 surveys (u.d.l: under detection limits; #: number of samples; SD: standard deviation) 773
774 775 776 . 777 778 779 780 781 782 783 784 785 786 787 788 789 790 791 792 793 794 795 796 797 798 799 800 801 802 803 804 805 806 807 808
Survey # Average SD Min 1° Quartile Median 3° Quartile Max
NO3-
I 9 10 20 u.d.l. 2 3 4 63
II 16 2 1 1 1 2 3 4
III 22 10 22 u.d.l. 0 2 6 101
NH4+
I 9 3 6 u.d.l. 0 u.d.l. 1 16
II 16 3 5 u.d.l. 0 u.d.l. 4 19
III 22 4 9 u.d.l. 0 u.d.l. 0 31

33
809
810 811 812 Fig. 7. Map of distribution of nitrate concentrations for each sampling site (third survey). For monitoring wells, 813 the values showed in the map refer to the shallowest screen monitored 814 815 816 817
818

34
819 820 821 Fig. 8. Map of distribution of ammonium concentrations for each sampling site (third survey). For monitoring 822 wells, the values showed in the map refer to the shallowest screen monitored 823 824 825
826 827
828
829
830
831
832
833
834

35
Table 3. δ18O and δD data. S: springs, W: wells, MW: monitoring wells. Data refer to the international 835 standard, Vienna Standard Mean Ocean Water (VSMOW). Location is shown in Fig. 3. (n.a.: data not 836 available) 837 838
839 840 841 842
843

36
844 845 Fig. 9. δ18O and δD plot; S: springs, W: wells, MW: monitoring wells. Blue line from Petitta et al. (2010) is 846 related to the upper valley of the Sagittario River. Red line from Rosanski et al. (1993) is related to Global 847 Meteoric Line, green line from Barbieri et al. (2003) is related to the Abruzzi region and the Orange line from 848 Longinelli and Selmo (2003) is related to Central Italy Meteoric Line 849 850
851
852
853
854
855
856
857
858
859
860
861 862

37
863 864
865 866 867 Fig.10. Longitudinal changes of benthic macroinvertebrates (a, b) and hyporheic copepods (c, d) along the 868 Sagittario River. a, c: abundance; b, d: species diversity measured as Margalef’s index (Margalef, 1998), 869 both calculated on cumulative abundance and species richness of spatial and temporal replicates. B0÷B4 870 location is shown in Fig.3 871 872 873 874 875 876 877 878 879 880 881 882 883 884 885 886 887 888 889 890 891

38
892 893
894 895 896 Fig. 11. Conceptual model of groundwater flow system and nitrogen cycle. U: upstream section 897 representative of upwelling zones; L: downstream section representative of downwelling zones. a: alluvial 898 fan aquifer, b: alluvial shallow aquifer, c: local terraced aquifer, d: lacustrine-marshy aquitard with high 899 permeability lenses, e: deep carbonate aquifer, f: groundwater flow direction, g: recharge area, h: water 900 table, i: monitoring well, l: nitrogen transport in groundwater 901 902 903 904 905

39
906 Fig. 12. Nitrogen and ammonium concentrations in river waters. Data refer to the third survey. Blue line 907 represents nitrate (as N-NO3
- mgL-1), red line ammonium (as N-NH4+ mgL-1). B0÷B4 refer to Fig. 3 908
909
Related Documents











