Harvard-MIT Division of Health Sciences and Technology HST.722J: Brain Mechanisms for Hearing and Speech Course Instructor: Kenneth E. Hancock HST.722 Brain Mechanisms of Speech and Hearing Fall 2005 Dorsal Cochlear Nucleus September 14, 2005 Ken Hancock

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Harvard-MIT Division of Health Sciences and TechnologyHST.722J: Brain Mechanisms for Hearing and SpeechCourse Instructor: Kenneth E. Hancock
HST.722 Brain Mechanisms of Speech and HearingFall 2005
Dorsal Cochlear NucleusSeptember 14, 2005
Ken Hancock

Dorsal Cochlear Nucleus (DCN)
• Overview of the cochlear nucleus and its subdivisions• Anatomy of the DCN• Physiology of the DCN• Functional considerations

Dorsal Cochlear Nucleus (DCN)
• Overview of the cochlear nucleus and its subdivisions• Anatomy of the DCN• Physiology of the DCN• Functional considerations

The cochlear nucleus
DCN
VCN AN

AN fibers terminate in a “tonotopic” or “cochleotopic” pattern
I
IIIII
0.17 kHz
2.7 kHz
10.2 kHz
36.0 kHz
ANR
sgb
1 mm
DCN
AVCN
PVCN
36.0 kHz
10.2 kHz
2.7 kHz
0.17 kHz
Figure by MIT OCW.

Major subdivisions of the cochlear nucleus
PVCN
AVCNDCN
OAPD
AP
PV
MA
AA
VEST NERVE
AUDITORYNERVE
Spherical bushy
Globular bushy Multipolar
Octopus
Spherical bushy
Globular bushy Multipolar
Octopus
Figure by MIT OCW.
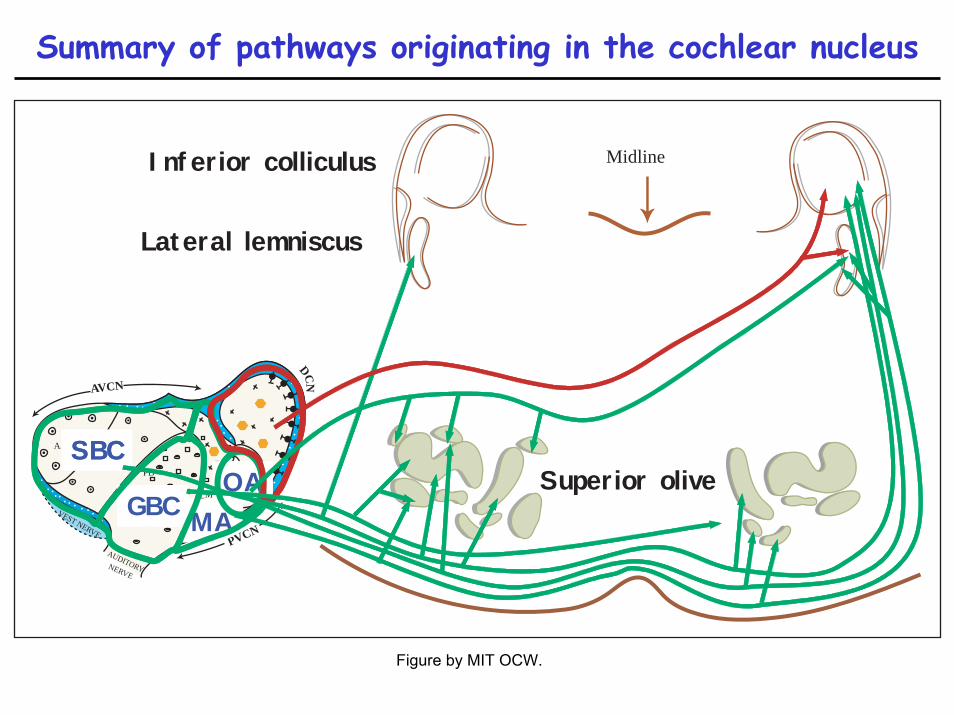
Summary of pathways originating in the cochlear nucleus
PVCN
AVCN
DCN
OAPD
AP
PV
MA
AA
VEST NERVE
AUDITORYNERVE
Midline
SBC
GBC MAOA
SBC
GBC MAOA
Inferior colliculus
Lateral lemniscus
Superior olive
Figure by MIT OCW.

Projections suggest DCN is a different animal than VCN
• (All roads lead to the inferior colliculus)• VCN projects directly to structures dealing with binaural
hearing and olivocochlear feedback• DCN ???
PVCN
AVCN
DCN
OAPD
AP
PV
MA
AA
VEST NERVE
AUDITORYNERVE
Midline
SBC
GBC MAOA
SBC
GBC MAOA
Inferior colliculus
Lateral lemniscus
Superior olive
Figure by MIT OCW.

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN• Physiology of the DCN• Functional considerations

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN• Physiology of the DCN• Functional considerations

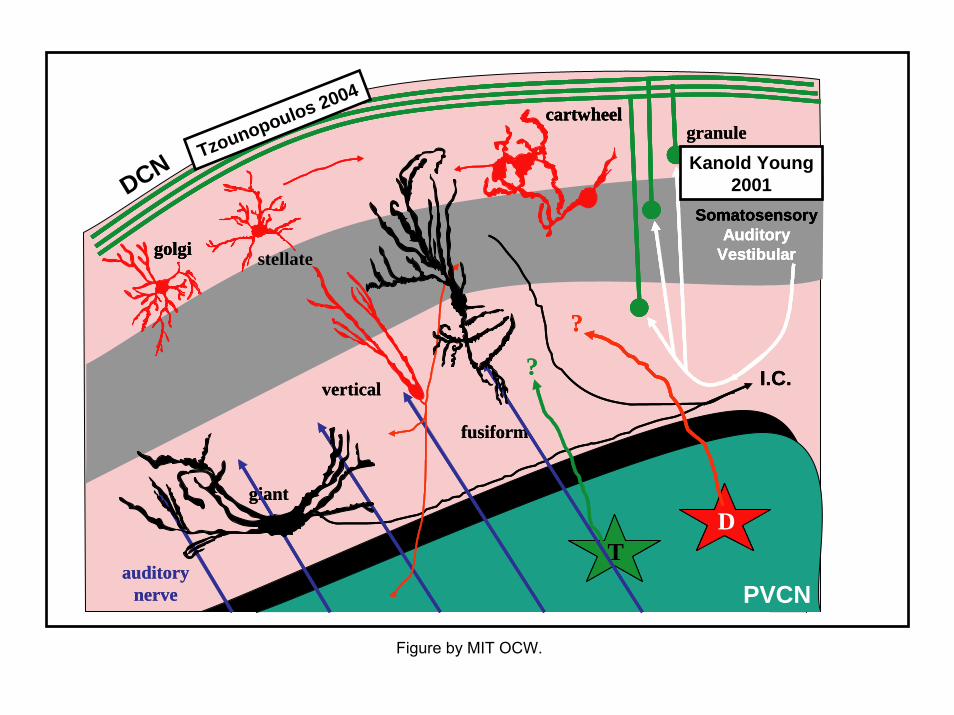
DCN
PVCN
I.C.I.C.
SomatosensoryAuditory
Vestibular
SomatosensoryAuditory
Vestibular
SomatosensoryAuditory
Vestibular
TD
?
?
TD
?
D
?
?
granulegranule
giant
fusiform
giant
fusiform
auditory nerve
auditory nerve
golgi
cartwheel
golgi
cartwheel
stellategolgi
cartwheel
verticalvertical
Kanold Young2001
Tzounopoulos 2004
Figure by MIT OCW.

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN• Functional considerations

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN• Functional considerations
Photograph of Eric Young removed due to copyright reasons. Please see: http://www.bme.jhu.edu/labs/chb/people/index.php?page=ABOUT&user=eyoung
Eric Young

Response Map classification scheme
~ Auditory Nerve increasing inhibition
200
10050
100
150
100
200
100
200
300
200
400
0 0
600
0 0 0
FREQUENCY FREQUENCY
Type I Type II Type III Type IV Type V
Sound Level Sound Level 10 dB10 dB
Soun
d Le
vel
Dis
char
ge R
ate
Excitatory Responses
Inhibitory Responses
BF Tone
Noise
Spont.Act
Figures by MIT OCW.

DCN: Vertical cells are type II and type III units
•Narrow V-shaped region of excitation•No spontaneous activity•Tone response >> noise response
•V-shaped region of excitation•Inhibitory sidebands
Evidence: Antidromic stimulation (Young 1980)
BF tonenoise
type IItype IItype II
BF tonenoise
type III
400
00.5 1 2 5 10 20
-95
-90
-80
-70
-60
-50
-40
-30
-20
Frequency, kHz
Rat
e, S
pike
s/s
Rat
e, S
pike
s/s
dB atten
Sound Level0
Rat
e
1.25 2
300
05 10 20
2030405060708090
100
Frequency, kHz
100
200
0Sound Level
Rat
e
300
200
100
BF
Figures by MIT OCW.
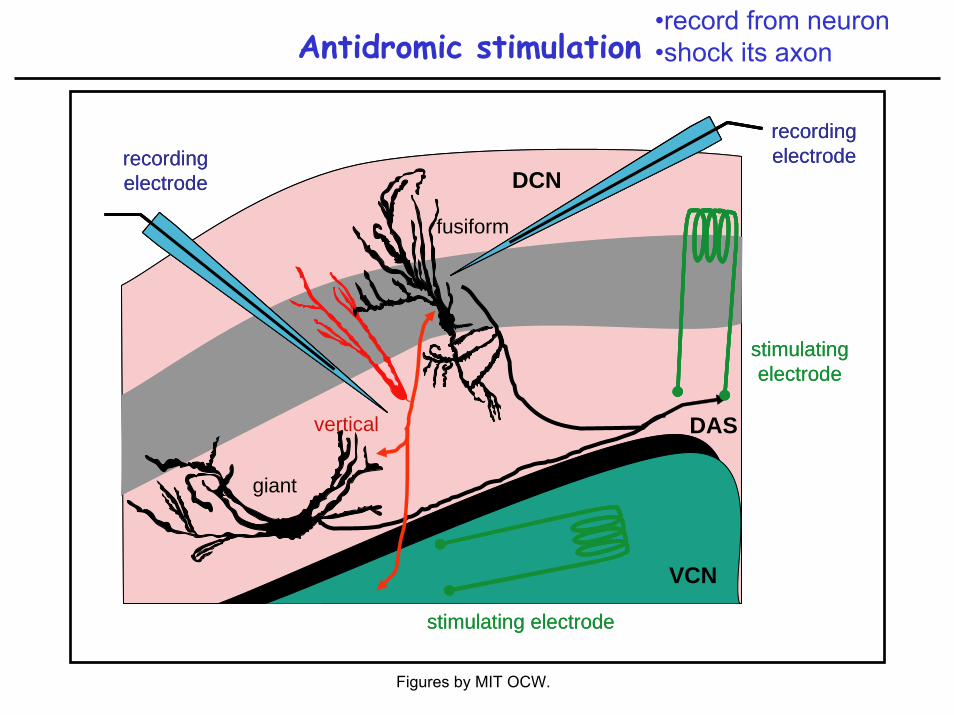
Antidromic stimulation•record from neuron•shock its axon
DAS
VCN
DCN
fusiform
giant
vertical
recordingelectrode
stimulatingelectrode
recordingelectrode
stimulatingelectrode
recordingelectrode
stimulating electrode
recordingelectrode
stimulating electrode
Figures by MIT OCW.

DCN: “Principal” cells are type III and type IV units
•“Island of excitation” & “Sea of inhibition”•BF rate-level curve inhibited at high levels•Noise rate-level curve ~ monotonic
Rat
e, sp
ikes
/sR
ate,
spik
es/s
1.25 2
300
05 10 20
2030405060708090100
Frequency, kHz
Frequency, kHz
50
100
150
100
200
0
0Sound Level
Sound Level
Rat
eR
ate
400
05 10 20 50
25
35
45
55
65
75
8595
dB attn
TYPE IIITYPE III
TYPE IVTYPE IV
Evidence:Antidromic stimulation (Young 1980)Intracellular recording and labeling (Rhode et al. 1983, Ding et al. 1996)
SpontSpont
Figures by MIT OCW.

Neural circuitry underlying DCN physiology:type II units inhibit type IV units
Rat
e, sp
ikes
/s
Frequency, kHz
50
100
150
0Sound level
Rat
eR
ate
400
05 10 20 50
25
35
45
55
65
75
8595
dB attn.TYPE IVTYPE IV
TYPE IITYPE II
SpontSpont
400
00.5 1 2 5 10 20
-95
-90
-80
-70
-60
-50
-40
-30
-20
Frequency, kHz
Rat
e, sp
ikes
/sdB attn.BF
Sound level0
300
200
100
BF tonenoise
"Reciprocal" responses
Giant
Fusiform
Vertical
Figures by MIT OCW.

Classic experiment: type II units inhibit type IV units• multiunit recording Voigt & Young (1980, 1990)
Spik
es/S
econ
d
Cross-correlogram
Milliseconds-10 -5 0 5 10
10
20
30
-20
-10
400
300
200
100
0110 60 10
BF Tone
Level, dB Attn
NoiseRat
e, S
pike
s
Rat
e, S
pike
s
200
100
0-25 25 75
BBN
BF
Level,dB
type IV firing rate given type II fires at t=0
inhibitory trough
Cross-correlogramtype IV firing rate given type II fires at t=0
inhibitory trough
-
type II unit type IV unittype II unittype II unit type IV unit
Figures by MIT OCW.

DCN physiology so far…
IV ~ principal cells
II
auditory nerve
~ vertical cells
+
+
• type II units inhibit type IV units
• BUT this analysis based on pure-tone responses
⇒ what happens with more general stimuli???
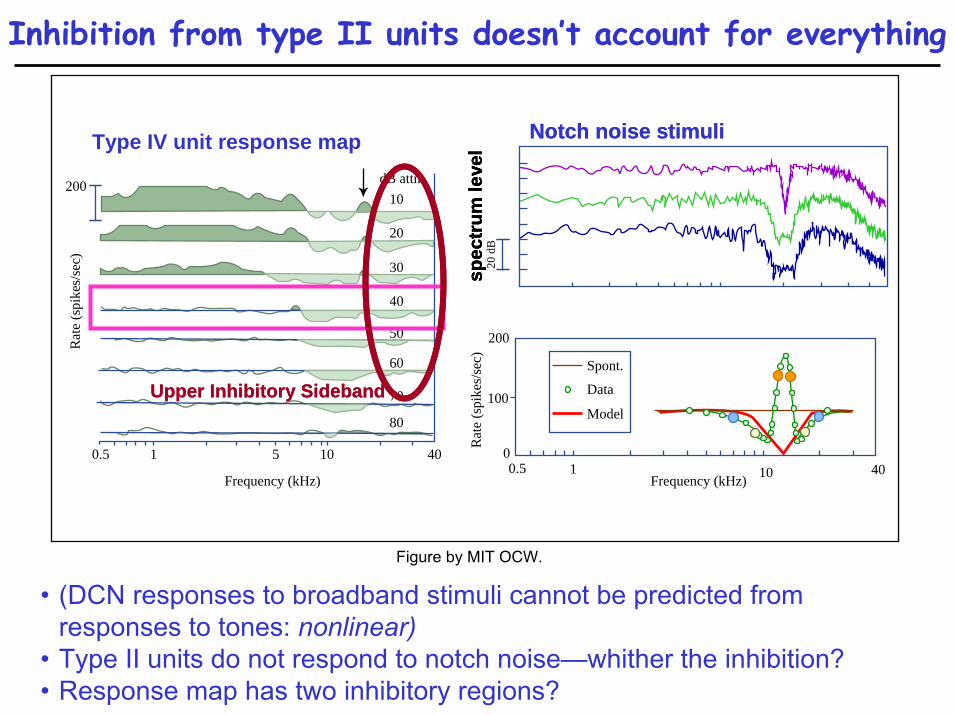
Inhibition from type II units doesn’t account for everything
• (DCN responses to broadband stimuli cannot be predicted from responses to tones: nonlinear)
• Type II units do not respond to notch noise—whither the inhibition?• Response map has two inhibitory regions?
0.5 1 5 10 40
Frequency (kHz) Frequency (kHz)
10
20
30
40
50
60
70
80
Rat
e (s
pike
s/se
c)
dB attn
0.5 1 10 40
Data
Model
Spont.
Rat
e (s
pike
s/se
c)
200
0
100
20 d
B
200
Type IV unit response map
spec
trum
leve
l
Notch noise stimuli
spec
trum
leve
lsp
ectr
um le
vel
Notch noise stimuli
Upper Inhibitory SidebandUpper Inhibitory Sideband
Figure by MIT OCW.

DCN notch noise sensitivity due to wideband inhibition
Broadband noise
AN input to type IV unit
frequency
leve
lle
vel
AN input to WBI
Notch noise
frequency
• strong AN input dominates
• type IV response is excitatory
• type IV loses greater portion of its excitatory input
• WBI input dominates
• type IV response is inhibitory
Nelken & Young 1994
WBIExcitatory
Inhibitory
Frequency
IV
Figure by MIT OCW.

PVCN: is the D-stellate cell the wideband inhibitor?
• such responses arise from radiate or stellate neurons (Smith & Rhode 1989)
• stellate cells send axons dorsally into the DCN, thus called “D-stellate cells” (Oertel et al. 1990)
• D-stellate cells are inhibitory (Doucet & Ryugo 1997)
• broadly-tuned, onset-chopper units are found in the PVCN (Winter & Palmer 1995)
• typically respond better to broadband noise than to tones
5-5
-15-25-35-45-55-65-75-85-95
-1051000050001000
Frequency (Hz)
Rel
ativ
e to
ne le
vel (
dB) 500
400
300
200
100
0100 50 0
1000
00 10 20
Noise
BF tone
Level, dB attn
Rat
e, sp
ikes
/s
Figure by MIT OCW. DCN
PVCN
DCN
PVCNa
Figure by MIT OCW.

Summary: Circuitry of DCN deep layer
Excitatory inputAN fibers?
Type II inhibitory inputs (strong)
UIS from ? source (weak)
Soun
d le
vel
Frequency
Excitatory
Inhibitory
400
300
200
100
0110 60 10
BF Tone
Level, dB attn
NoiseRat
e, sp
ike/
s
IV
Best frequency
500
400
300
200
100
0100 50 0
1000
00 10 20
Noise
BF tone
Level, dB attnR
ate,
spik
es/s
Inhibitory
Excitatory
Spirou and Young 1991
type IItype II
W.B.I.W.B.I.
“reciprocal” response properties
Figures by MIT OCW.
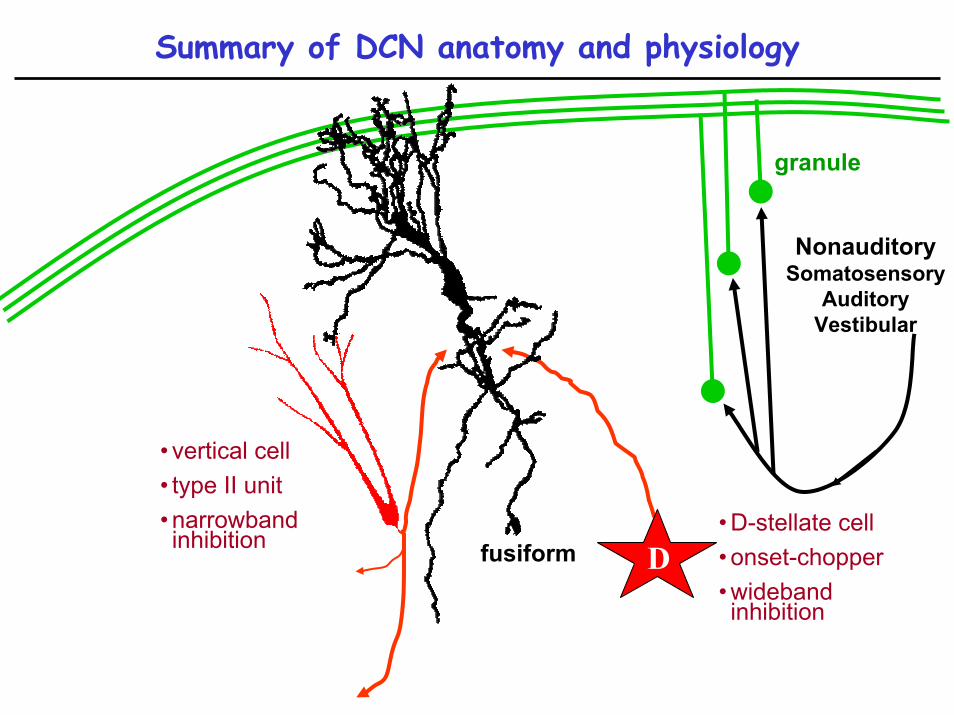
Summary of DCN anatomy and physiology
NonauditorySomatosensory
AuditoryVestibular
granule
fusiform
• vertical cell• type II unit• narrowband inhibition
D• D-stellate cell• onset-chopper• wideband inhibition

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN– diverse response properties– complex interconnections– highly nonlinear
• Functional considerations

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN– diverse response properties– complex interconnections– highly nonlinear
• Functional considerations
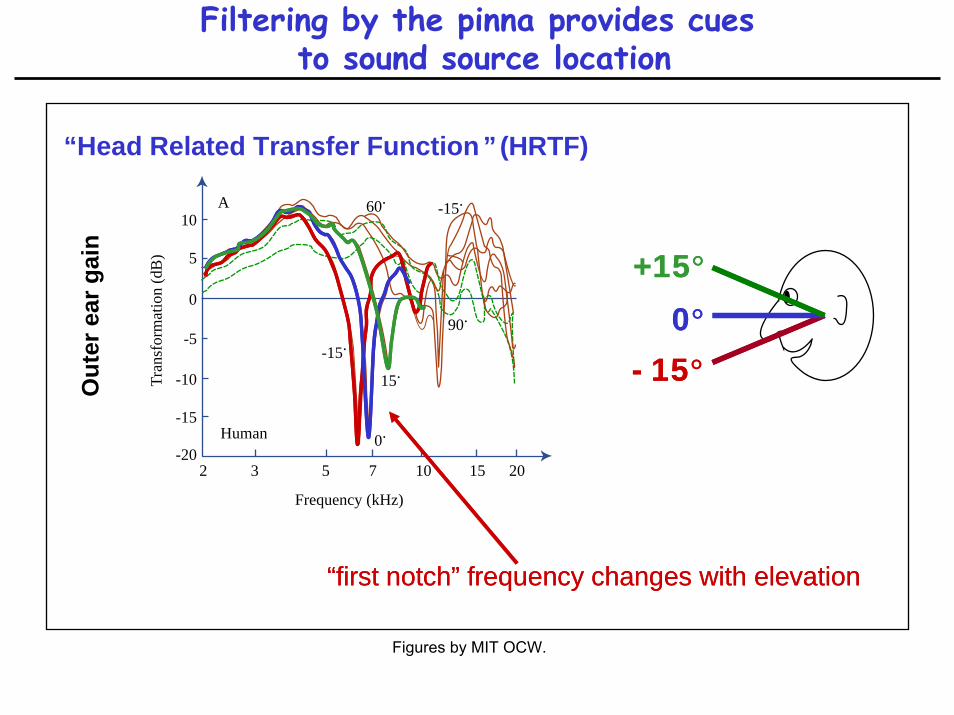
Filtering by the pinna provides cuesto sound source location
Human
A
-15.
-15.
15.
60.
90.
0.
2 3 5 7 10 15 20
10
0
5
-5
-10
-15
-20
Frequency (kHz)
Tran
sfor
mat
ion
(dB
)“Head Related Transfer Function ” (HRTF)
Out
er e
ar g
ain
-15°-15°0°0°
+15°+15°
“first notch” frequency changes with elevation“first notch” frequency changes with elevation
Figures by MIT OCW.

Type IV units are sensitive to HRTF first notch
• type IV units are inhibited by notches centered on BF
• null in DCN population response may code for sound source location
Reiss & Young2005
May 2000
Physiology ⇒
Behavior ⇒
Figures by MIT OCW.
30
-100
1020
30
-100
1020
-15dB
-35 dB
-55 dB
-75 dB
-95 dB300
3 5 10 30
3 5 10 300
100
200
Frequency / kHz
Stimulus notch frequency
Rat
e / s
pike
s per
seco
ndR
ate
/ spi
kes p
er se
cond
Gai
n / d
BEl = 30o, Az = 15o
El = 30o, Az = -15o
El = 30o, Az = -15o
El = 30o, Az = 15o
spont.

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– receives nonauditory inputs– has similar organization to cerebellar cortex
• Physiology of the DCN– diverse response properties– complex interconnections– highly nonlinear
• Functional considerations– coding sound source location based on pinna cues
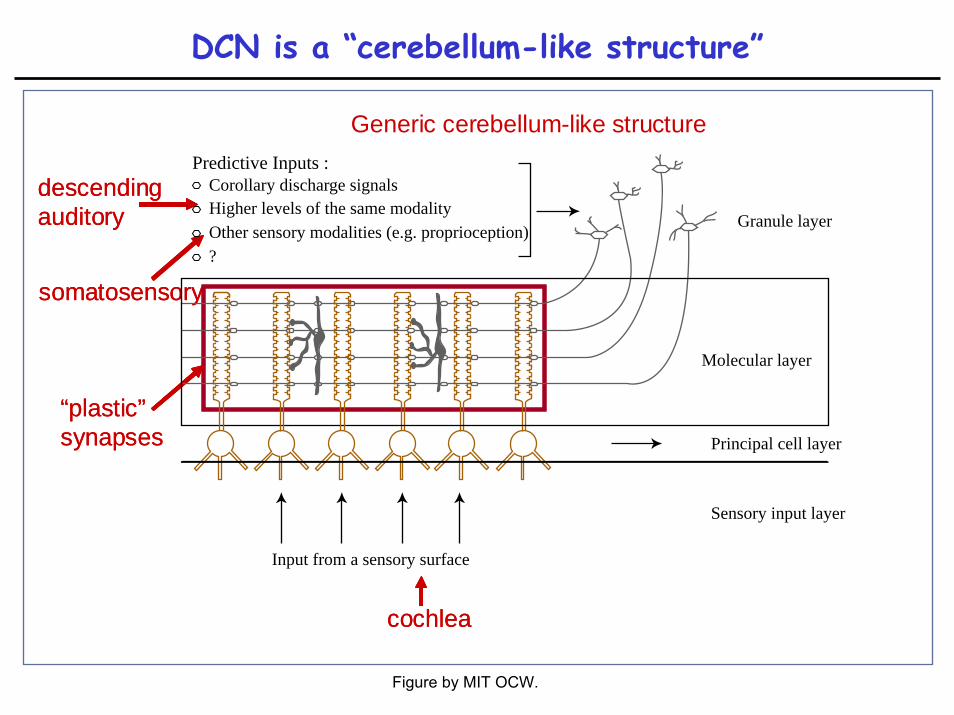
DCN is a “cerebellum-like structure”
Generic cerebellum-like structure
descending auditorydescending auditory
somatosensorysomatosensory
cochleacochlea
“plastic” synapses“plastic” synapses
Predictive Inputs :Corollary discharge signalsHigher levels of the same modalityOther sensory modalities (e.g. proprioception)?
Granule layer
Molecular layer
Principal cell layer
Sensory input layer
Input from a sensory surface
Figure by MIT OCW.

Synaptic plasticity: Long-Term Potentiation (LTP)
• “Classical” LTP demonstration at the hippocampal CA3-CA1 synapse• LTP evoked by tetanic stimulation (mechanism involves NMDA receptors)
Tzou
nopo
ulos
2004

Electric fish provide clues to cerebellum-like function
black ghost knifefish(Apteronotus albifrons)
Photograph removed due to copyright reasons. Please see the Nelson Lab home page: http://nelson.beckman.uiuc.edu
electric organ discharge
(EOD)
• electrical activity detected by electric lateral line• afferent activity transmitted to electric lateral line
lobe (ELL), analogous to DCN
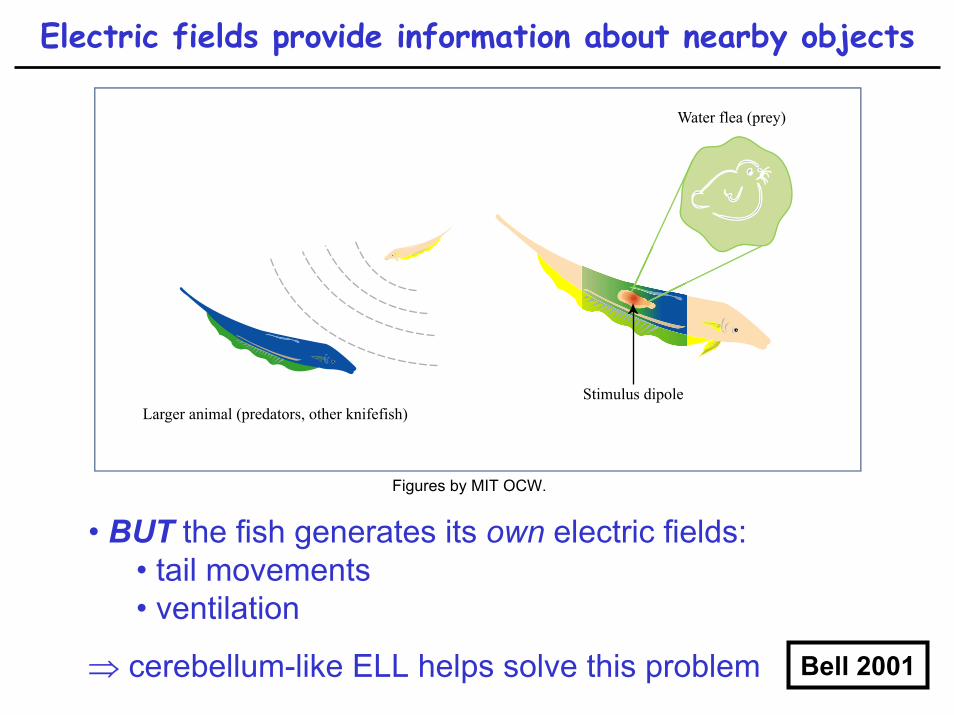
Electric fields provide information about nearby objects
• BUT the fish generates its own electric fields:• tail movements• ventilation
⇒ cerebellum-like ELL helps solve this problem Bell 2001
Figures by MIT OCW.

What do cerebellum-like structures do???
• Subtract the expected input pattern from the actual input pattern to reveal unexpected or novel features of a stimulus.
– DCN: pinna movement is expected to shift the first notch, independent of what the sound source is doing

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN– diverse response properties– complex interconnections– highly nonlinear
• Functional considerations: the DCN may…– code sound source location based on pinna cues– extract novel components of response

DCN may play a role in tinnitus
• percept of noise, ringing, buzzing, etc.
• affects up to 80% of the population
• 1 in 200 are debilitated
• (not voices in the head)
This makes no sense!
So why DCN? Because tinnitus…
• involves plasticity
• may involve somatosensory effects
Levine 1999

Dorsal Cochlear Nucleus
• Overview of the cochlear nucleus and its subdivisions– DCN projections do not reveal its function
• Anatomy of the DCN– more complex than other CN subdivisions– nonauditory inputs– similar organization to cerebellar cortex
• Physiology of the DCN– diverse response properties– complex interconnections– highly nonlinear
• Functional considerations: the DCN may…– code sound source location based on pinna cues– extract novel components of response– contribute to tinnitus

Selected References
Slide 5: Ryugo DK, May SK (1993) The projections of intracellularly labeled auditory nerve fibers to the dorsal cochlear nucleus of cats. J Comp Neurol 329:20-35.
Slide 6: Ehret G, Romand R, eds (1997) The Central Auditory System. New York: Oxford UniversityPress.
Slide 7: Ehret G, Romand R, eds (1997) The Central Auditory System. New York: Oxford UniversityPress.
Slide 12: Lorente de Nó R (1981) The Primary Acoustic Nuclei. New York: Raven.
Kane ES, Puglisi SG, Gordon BS (1981) Neuronal types in the deep dorsal cochlear nucleus of the cat. I. Giant neurons. J Comp Neurol 198:483-513.
Slide 15: Young ED (1984) Response characteristics of neurons of the cochlear nuclei. In: Hearing Science (Berlin CI, ed), pp 423-460. San Diego: College-Hill. ISBN: 0316091693.
Slides 16, 18, 19: Young ED (1984) Response characteristics of neurons of the cochlear nuclei. In: Hearing Science (Berlin CI, ed), pp 423-460. San Diego: College-Hill.
Young ED, Davis KA (2001) Circuitry and Function of the Dorsal Cochlear Nucleus. In: Integrative Functions in the Mammalian Auditory Pathway (Oertel D, Popper AN, Fay RR, eds). New York: Springer-Verlag.

Selected References, continued
Slide 20:VOIGT, H. F. AND YOUNG, E. D. Cross-correlation analysis of inhibitory interactions in dorsal cochlear nucleus. J. Neurophysiol. 64: 1590- 16 10, 1990.
Young ED, Davis KA (2001) Circuitry and Function of the Dorsal Cochlear Nucleus. In: Integrative Functions in the Mammalian Auditory Pathway (Oertel D, Popper AN, Fay RR, eds). New York: Springer-Verlag.
Slide 22:Spirou GA, Young ED (1991) Organization of dorsal cochlear nucleus type IV unit response maps and their relationship to activation by band-limited noise. J Neurophysiol 66:1750-1768.
Slide 23:Nelken I, Young ED (1994) Two separate inhibitory mechanisms shape the responses of dorsal cochlear nucleus type IV units to narrowband and wideband stimuli. J Neurophysiol 71:2446-2462.
Slide 24:Winter IM, Palmer AR (1995) Level dependence of cochlear nucleus onset unit responses and facilitation by second tones or broadband noise. J Neurophysiol 73:141-159.
Young ED, Davis KA (2001) Circuitry and Function of the Dorsal Cochlear Nucleus. In: Integrative Functions in the Mammalian Auditory Pathway (Oertel D, Popper AN, Fay RR, eds). New York: Springer-Verlag.
Oertel D, Wu SH, Garb MW, Dizack C (1990) Morphology and physiology of cells in slice preparations of the posteroventral cochlear nucleus of mice. J Comp Neurology 295:136-154.

Selected References, continued
Slide 25:Spirou GA, Young ED (1991) Organization of dorsal cochlear nucleus type IV unit response maps and their relationship to activation by band-limited noise. J Neurophysiol 66:1750-1768.
Nelken I, Young ED (1994) Two separate inhibitory mechanisms shape the responses of dorsal cochlear nucleus type IV units to narrowband and wideband stimuli. J Neurophysiol 71:2446-2462.
Young ED, Davis KA (2001) Circuitry and Function of the Dorsal Cochlear Nucleus. In: Integrative Functions in the Mammalian Auditory Pathway (Oertel D, Popper AN, Fay RR, eds). New York: Springer-Verlag.
Slide 26:Oertel D, Wu SH, Garb MW, Dizack C (1990) Morphology and physiology of cells in slice preparations of the posteroventral cochlear nucleus of mice. J Comp Neurology 295:136-154.
Slide 29: Geisler CD (1998) From Sound to Synapse: Physiology of the Mammalian Ear.: Oxford University Press.
Slide 30:Young ED, Spirou GA, Rice JJ, Voigt HF (1992) Neural organization and responses to complex stimuli in the dorsal cochlear nucleus. Philos Trans R Soc Lond B Biol Sci 336:407-413.
Slide 32: Bell CC (2001) Memory-based expectations in electrosensory systems. Curr Opin Neurobiol 11:481-487.
Slide 35:Zakon HH (2003) Insight into the mechanisms of neuronal processing from electric fish. Curr Opin Neurobiol13:744-750.
Related Documents











