©2006 FASEB The FASEB Journal express article 10.1096/fj.05-5343fje. Published online February 7, 2006. HSP induction mediates selective clearance of tau phosphorylated at proline-directed Ser/Thr sites but not KXGS (MARK) sites Chad A. Dickey,* Judith Dunmore,* Bingwei Lu, † Ji-Wu Wang, † Wing C. Lee,* Adeela Kamal, ‡ Francis Burrows, ‡ Christopher Eckman,* Michael Hutton,* and Leonard Petrucelli* *Mayo Clinic Jacksonville, College of Medicine, Jacksonville Florida; † Stanford University, Stanford, California, USA; and ‡ Conforma Therapeutics Corporation, San Diego, California Corresponding author: Michael Hutton, Mayo Clinic Jacksonville, 4500 San Pablo Rd., Jacksonville, FL 32224. E-mail: [email protected] ABSTRACT Neurofibrillary tangles (NFTs) are a characteristic neuropathological feature of Alzheimer’s disease (AD), and molecular chaperones appear to be involved in the removal of disease- associated hyperphosphorylated tau, a primary component of NFTs. Here, novel HSP90 inhibitors were used to examine the impact of chaperone elevation on clearance of different tau species in transfected cells using a unique quantitative assay. The HSP90 inhibitors reduced levels of tau phosphorylated at proline-directed Ser/Thr sites (pS202/T205, pS396/S404) and conformationally altered (MC-1) tau species, an epitope that is immeasurable by standard Western blot techniques. The selective clearance of these phospho-tau species and MC-1 tau was mediated via the proteasome, while lysosomal-mediated tau degradation seems to lack specificity for certain tau species, suggesting a more general role in total tau removal. Interestingly, tau phosphorylated at S262/S356 within the tau microtubule binding domain was minimally affected by chaperone induction. Overall, our data show that chaperone induction results in the selective clearance of specific phospho-tau and conformationally altered tau species mediated by the proteasome; however, the apparent stability of pS262/S356 tau may also explain why MARK is able to regulate normal tau function yet still be linked to the initiation of pathogenic tau hyperphosphorylation in AD. Key words: neurofibrillary tangles • Alzheimer’s disease • proteasome long with senile plaques composed mainly of Aß, accumulation of hyperphosphorylated species of the microtubule associated protein tau into intraneuronal filamentous inclusions (tangles and neurophil threads) is a characteristic pathological feature of Alzheimer’s disease (AD). Indeed, tau inclusions in neurons, and sometimes glia, are observed in multiple neurodegenerative conditions that are known collectively as tauopathies. The discovery of mutations in the tau gene that cause familial frontotemporal dementia (1–3) demonstrated that tau dysfunction is sufficient to cause neurodegeneration. This, in turn, refocused attention on therapeutic approaches that might prevent or reverse the pathological accumulation of tau in disease. A Page 1 of 18 (page number not for citation purposes)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

©2006 FASEB
The FASEB Journal express article 10.1096/fj.05-5343fje. Published online February 7, 2006.
HSP induction mediates selective clearance of tau phosphorylated at proline-directed Ser/Thr sites but not KXGS (MARK) sites Chad A. Dickey,* Judith Dunmore,* Bingwei Lu,† Ji-Wu Wang,† Wing C. Lee,* Adeela Kamal,‡ Francis Burrows,‡ Christopher Eckman,* Michael Hutton,* and Leonard Petrucelli*
*Mayo Clinic Jacksonville, College of Medicine, Jacksonville Florida; †Stanford University, Stanford, California, USA; and ‡Conforma Therapeutics Corporation, San Diego, California
Corresponding author: Michael Hutton, Mayo Clinic Jacksonville, 4500 San Pablo Rd., Jacksonville, FL 32224. E-mail: [email protected]
ABSTRACT
Neurofibrillary tangles (NFTs) are a characteristic neuropathological feature of Alzheimer’s disease (AD), and molecular chaperones appear to be involved in the removal of disease-associated hyperphosphorylated tau, a primary component of NFTs. Here, novel HSP90 inhibitors were used to examine the impact of chaperone elevation on clearance of different tau species in transfected cells using a unique quantitative assay. The HSP90 inhibitors reduced levels of tau phosphorylated at proline-directed Ser/Thr sites (pS202/T205, pS396/S404) and conformationally altered (MC-1) tau species, an epitope that is immeasurable by standard Western blot techniques. The selective clearance of these phospho-tau species and MC-1 tau was mediated via the proteasome, while lysosomal-mediated tau degradation seems to lack specificity for certain tau species, suggesting a more general role in total tau removal. Interestingly, tau phosphorylated at S262/S356 within the tau microtubule binding domain was minimally affected by chaperone induction. Overall, our data show that chaperone induction results in the selective clearance of specific phospho-tau and conformationally altered tau species mediated by the proteasome; however, the apparent stability of pS262/S356 tau may also explain why MARK is able to regulate normal tau function yet still be linked to the initiation of pathogenic tau hyperphosphorylation in AD.
Key words: neurofibrillary tangles • Alzheimer’s disease • proteasome
long with senile plaques composed mainly of Aß, accumulation of hyperphosphorylated species of the microtubule associated protein tau into intraneuronal filamentous inclusions (tangles and neurophil threads) is a characteristic pathological feature of
Alzheimer’s disease (AD). Indeed, tau inclusions in neurons, and sometimes glia, are observed in multiple neurodegenerative conditions that are known collectively as tauopathies. The discovery of mutations in the tau gene that cause familial frontotemporal dementia (1–3) demonstrated that tau dysfunction is sufficient to cause neurodegeneration. This, in turn, refocused attention on therapeutic approaches that might prevent or reverse the pathological accumulation of tau in disease.
A
Page 1 of 18(page number not for citation purposes)

Hyperphosphorylation of tau at multiple sites throughout the protein and abnormal folding of this typically linear protein are inevitably associated with the formation of the neurofibrillary lesions in AD. Two distinct phosphorylation motifs are present within the tau protein; proline-directed serine/threonine sites (4) and KXGS motifs within the four microtubule binding repeat domains of tau (serine residues at 262, 293, 324 and 356). These KXGS sites have been identified as substrates for the microtubule affinity regulating kinase (MARK) (5), which plays a normal role in the regulation of tau function, causing tau to be released from microtubules and thus axonal dynamics; however, recent evidence suggests that prior phosphorylation by MARK (the PAR-1 Drosophila ortholog) is required to initiate the pathogenic cascade of hyperphosphorylation by other kinases that are associated with the formation of neurofibrillary tau lesions in tauopathy (6). Along with hyperphosphorylation, an early event in the process of tau aggregation is a conformational change that occurs, allowing the N terminus of tau to interact with its microtubule binding region (7). Tau in this folded state can be detected with the MC-1 antibody that recognizes amino acids 7 through 9 and 312 through 342 only when these residues are adjacent to each other (7).
Understanding the mechanism by which these hyperphosphorylated and conformationally altered tau species are removed by neurons is, therefore, of considerable interest for therapeutic development. Recent studies have suggested the involvement of multiple degradation pathways in this process that require the action of molecular chaperones. In particular, HSP27 appears to direct ubiquitin-independent proteasome-mediated clearance of phospho-tau species (8, 9), and the HSP70 cochaperone CHIP (carboxyl terminus of the Hsc70-interacting protein) appears to be a critical ligase required for ubiquitin-dependent tau degradation (10, 11).
One group of compounds that can activate this chaperone system is the HSP90 inhibitors, and recent evidence demonstrates the impact that drugs such as these may have on neurodegenerative diseases of abnormal protein aggregation. HSP90 inhibition elicits a two-pronged effect, resulting in the proteasome-mediated degradation of HSP90 “client” proteins and activation of heat shock factor 1 (HSF1; Ref. 12). Geldanamycin (GA), the original HSP90 inhibitor, proved to be hepatotoxic; however, several less toxic GA derivatives have been generated, including 17-allylamino 17-demethoxy-geldanamycin (17-AAG) and 17-dimethyl-amino-ethylamino-17-demethoxygeldanamycin (17-DMAG), both of which are presently in clinical trials as putative anticancer agents (13). These compounds are quite large and therefore lack the appropriate characteristics for a CNS therapeutic. Therefore, we have developed several low molecular weight HSP90 inhibitors that may offer sufficient BBB penetrance to make systemic administration for neurological disorders feasible (A. Kamal et al., unpublished observations).
In this study, we used a novel fluorescent cell-based assay to screen a small series of these low molecular weight HSP90 inhibitors for their effects on phospho-tau protein levels in a transfected cell model. Robust induction of HSP70, HSP40, and HSP27 through HSP90 inhibition was shown to result in selective reductions in proline-directed Ser/Thr phospho-tau species and in conformationally altered (MC-1 positive) tau species. Importantly, we also found that tau phosphorylated at S262/S356 by MARK remains largely unaffected by these compounds. We discuss the implications of these findings for the regulation of tau function by phosphorylation and for the potential use of HSP90 inhibitors as therapies for human tauopathy.
Page 2 of 18(page number not for citation purposes)

MATERIALS AND METHODS
Cell culture and expression vectors
Chinese hamster ovary (CHO) cells and human embryonic kidney (HEK293) cells stably transfected with the shortest isoform of tau containing 4 microtubule binding domains and harboring the P301L point mutation in exon 10 (4R0N P301L) were used in Western blot analysis, immunoprecipitations and in-cell Western blot assays for phospho-tau species and conformationally altered (MC-1) tau. These transfected cell lines were used because overexpression of the mutant construct resulted in enhanced levels of these tau species in cell culture. These lines were grown in Opti-MEM media +10% FBS + pen-strep in the presence of the G418 selection marker to maintain high numbers of transfected cells. New vials of the lines were also started once every month. The human neuroglioma-derived cell line (H4) was used for analysis of HSP90 inhibitor effects on endogenous total tau levels using in-cell Western blot assays due to preliminary work showing moderate levels of both 4 and 3 repeat tau expression in this line. Cells were passaged every 3−4 days in Opti-MEM medium + 10% FBS + pen-strep and a new line was started once per month.
Primary cultures were generated as described previously (14). Briefly, cortical neurons from 2 day-old postnatal mice were dissociated with papain, and cells were plated on top of cortical glia cell monolayers at a density of 80,000 (colocalization) cells per cover slip in preconditioned media, by adding to glial feeder layers 24 h before plating neurons.
Antibodies
Rabbit polyclonal total tau and ubiquitin antibodies were obtained from DAKO (1:500, 1:1000); Mouse monoclonal total tau antibody (Tau5; Standard Western only 1:1000); E1, human specific tau antibody (Mayo Clinic; Standard Western only 1:5000); CP13 (phospho-tau S202/T205; 1:100, 1:1000), PHF-1 (phospho-tau S396/404; 1:100, 1:1000) and MC-1 (tau conformation-specific; in-cell Western blot analysis only 1:50) monoclonal antibodies were obtained from Dr. Peter Davies (Albert Einstein College of Medicine); 12E8 (phospho-tau S262/356; 1:500, 1:1000) monoclonal antibody was obtained from Dr. Seubert (Table 1; Elan Pharmaceuticals; (15, 16). Glyceraldehyde-3-phosphate dehydrogenase monoclonal antibody was obtained from Biodesign International (1:1500, 1:5000); TUB2.1 tubulin antibody was obtained from Sigma (ICC 1:500); HSP70, HSP40 and HSP27 (All standard Western blot analysis only 1:1000) antibodies were obtained from Stressgen; HRP-conjugated secondary antibodies were obtained from Jackson Immunoresearch (Standard Western blot analysis only 1:2000); IR-Dye conjugated antibodies were obtained from Molecular Probes (680) and Rockland Labs (800) (Both in-cell Western blot analysis only 1:500). Anti-rabbit IgG conjugated to Alexa Fluor 568 and Anti-mouse IgG conjugated to Alexa Fluor 568 were used for fluorescent ICC diluted 1:1000.
HSP90 inhibitors and cell treatments
Conforma Therapeutics Corporation generated the panel of HSP90 inhibitors used in these studies. Conforma Therapeutics screened this series of HSP90 inhibitors for potency using a flow cytometric assay detecting degradation of HER-2 in MCF-7 breast cancer cells (manuscript submitted). The compounds with their chemical formulas, molecular weights, and IC50s in the HER-2 degradation assay were EC82 (C14H15BrN6O, Mr=363, IC50=45 nM), EC86
Page 3 of 18(page number not for citation purposes)

(C14H17ClN6O2, Mr=337, IC50=60 nM), EC102 (C19H25IN6OS, Mr=512, IC50=80 nM), EC104 (C18H20ClN6O3PS2, Mr=499, IC50=33 nM), EC105 (C13H12BrClN6, Mr=368, IC50= 15 nM), EC107 (C14H17ClN6O2, Mr=427, IC50=32 nM), EC108 (C14H15BrClN5O, Mr=385, IC50=15 nM), and 17-AAG (C31H43N3O8, Mr=586, IC50=5 nM). HSP90 inhibitors were suspended in DMSO to a stock concentration of 1 mM. The proteasomal inhibitor MG-132 was obtained from A.G. Scientific; the lysosomal inhibitor chloroquine was obtained from Calbiochem.
For in-cell Western blot assays, cells were passaged into 96-well clear bottom plates (BD Falcon), incubated for 24 or 48 h before treatment with HSP90 inhibitors (48 h was used only in assays of the conformationally altered, MC-1-positive, tau species to allow sufficient initial accumulation of this species). Treatment with the inhibitors (1-μM final concentration) or vehicle alone was for 24 h. The plates were then prepared for fluorescent analysis as described below. For studies using the proteasomal inhibitors, the final concentration of MG132 was 5 µM and was coadministered directly with the HSP90 inhibitors. For preparation of homogenates for normal Western blot analyses, cells were passaged into 6-well plates, incubated overnight, and treated the next day with an HSP90 inhibitor (1 μM) or vehicle. Cells were then incubated for 24 h and harvested in lysis buffer containing 50 mM Tris-HCl pH 7.4, 1 M NaCl, 0.1% Triton-X, 5 mM EDTA plus 1% SDS, PMSF, and both a protease and phosphatase inhibitor cocktail.
In-cell Western blot assays
Following 24 or 48 h drug treatment (24 h was largely used because of cell overgrowth at the 48-h time point), 50 µl of media from the 96-well plates was removed for LDH analysis to detect toxicity (Cytotox 96 Assay Kit, Promega), and the remaining media was aspirated. Cells were fixed with 3.7% formaldehyde in PBS for 20 min and then permeabilized with three 10-min washes in PBS + 0.1% Triton-X 100 using an ELX405 auto plate washer (BioTek Instruments). A proprietary lipid-based blocking buffer (Li-Cor) was added for 2 h, followed by overnight incubation with primary antibodies in a 1:1 buffer of LBB and PBS + 0.2% Tween20. After three 10-min washes in PBS + 0.1% Tween-20, secondary detection was carried out using two species-specific infrared fluorescent dye conjugated antibodies (see above) in 1:1 buffer of LBB and PBS + 0.4% Tween20. After an hour incubation and washing, targets were simultaneously visualized using the Odyssey Infrared Imaging Scanner (Li-Cor, Lincoln, NB) with the 680-nm fluorophore emitting a red color and the 800-nm fluorophore emitting a green color. Relative fluorescent units for HSP90 inhibitor-treated samples were divided by vehicle controls to determine percent change in expression. A student t test was used to determine significant toxicity effects due to drug treatment based on reduced levels of GAPDH. A separate Student’s t test was used to determine significant differences between tau species levels in treated vs. untreated cells.
Western blot analysis
Cell lysates, prepared as described above, were sonicated, and protein concentrations were measured by a standard BCA assay (Pierce). The samples were then heated in Laemmli’s buffer, and equal amounts of protein were loaded into 12-well 10% Tris-HCl gels (Bio-Rad). Following transfer, blots were blocked with Blotto (5% NFDM in TBS+0.1% Triton-X 100) for 2 h, then antibodies were applied at appropriate dilutions in Blotto overnight at 4°C. Membranes were washed 3 × 10 min in TBST and incubated with secondary antibody for 1 h. Membranes were then washed 3 × 10 min, and protein expression was visualized by ECL treatment and exposure
Page 4 of 18(page number not for citation purposes)

to film. Bands were quantified using Scion Image by analyzing pixel density and correcting protein levels for GAPDH (data not shown).
Fluorescent immunocytochemistry in primary cortical neurons
Primary cortical neurons were grown on coverslips for 7 days following harvest from 2-day postnatal mouse pups. After 24-h treatment with a 1-µM concentration of the HSP90 inhibitor or vehicle, these were then fixed and permeabilized with 4% formaldehyde in DPBS + 0.1% saponin for 30 min and blocked for 1 h with 5% goat serum + 5% FBS in DPBS. Polyclonal anti-total tau antibody (DAKO) and the TYR 2.1 anti-tubulin antibody were diluted in DPBS + Tween-20 (DPBST) + 0.1% goat serum and incubated overnight at 4°C. Coverslips were washed the following day 3 times for 10 min in DPBST, and Alexa Fluor secondary antibodies were added for 1 h. Vectamount was used to mount the coverslip to slides, and cells were imaged on a standard fluorescent microscope (Olympus) without changing light intensity.
RESULTS
Recent evidence has suggested that pharmacologic manipulation of the heat shock response by inhibiting HSP90 may have therapeutic potential for the treatment of neurodegenerative protein aggregation disorders (8, 11, 17, 18). Here, we demonstrated that treatment of primary cortical neurons with the classical HSP90 inhibitor, GA, reduced tau protein levels in a dose-dependent manner that inversely correlated with HSP70 induction (Fig. 1). However, the use of these typical HSP90 inhibitor compounds (i.e., GA and its derivatives, 17-AAG and 17-DMAG) lacks practical clinical relevance for the treatment of CNS disorders due to poor BBB permeability. Therefore, a small panel of novel, low molecular weight HSP90 inhibitors was generated, and we have developed novel 96-well microplate-based in-cell Western assays that used near-infrared imaging to analyze the effects of these compounds on tau biology in cell culture. Here, we have demonstrated that several of these novel compounds were able to reduce total tau levels with minimal toxicity (Fig. 2A). Although chaperones such as HSP27 and CHIP have been identified as having an essential role in the degradation of tau (10, 11), it is interesting to note that those compounds which elicited a greater than 10-fold induction in HSP70 levels also produced the greatest reduction in total tau levels following drug exposure (Fig. 2B; (19)).
To address whether chaperone activation through HSP90 inhibitor treatment was capable of selectively reducing abnormal phospho-tau species, we refined our in-cell Western blot technique by using antibodies specific for 1) phospho-S202/T205 tau, a reported Ser/Thr-Pro phosphorylation site for GSK3β and MAPK, 2) phospho-S396/S404 tau, a reported Ser/Thr-Pro phosphorylation site for CdK5 and 3) phospho-S262/S356, a reported KXGS phosphorylation site for MARK. In addition, because of the nondenaturing fixation conditions used in this in-cell assay, we were also able to quantitatively measure levels of the MC-1 conformational epitope, an aberrant early structural motif that is formed by interaction between the N-terminal domain of tau and the microtubule-binding domains. Because these phospho- and conformationally altered tau species represent a small fraction of total tau, we increased the dynamic range of the assay by using a stably transfected CHO cell line overexpressing 4R0N P301L tau (CHO tau (P301L)).
In-cell Western analysis of the CHO tau(P301L) cell line demonstrated that it generated 3–4 times the amount of the phospho-tau species pS202/T205, pS396/S404, and pS262/S356 when compared with normal H4 neuroglioma cells (not shown). In addition, MC-1 tau (conformational
Page 5 of 18(page number not for citation purposes)

dependent) levels were slightly increased (0.5-fold higher) in the 4R0N P301L tau cell line as compared with the H4 cell line.
The CHO tau (P301L) cell line was treated with 1 μM of each HSP90 inhibitor and the levels of total tau (Fig. 3A), the three phospho-tau species (S202/T205, S96/S404, and S262/S356; Fig. 3B), and MC-1 positive tau (Fig. 3C) were determined by in-cell Western blot analysis. The HSP90 inhibitors EC82, 86, 102, 104, 105, and 17-AAG, which generate the largest increase in chaperone levels (Fig. 2), significantly reduced total tau levels by 25–55%, in the 4R0N P301L tau cell line, relative to vehicle-treated cells (Fig. 3A). In contrast, EC107 and EC108, which generate the smallest increase in chaperone levels (Fig. 2), had no effect on total tau levels (Fig. 3A). Five of the compounds (EC82, 86, 102, 104, and 105) selectively reduced the proportion of total tau phosphorylated at sites S202/T205 and S396/S404 by 50% or greater (P<0.001, Student’s t test), while leaving levels of tau phosphorylated at the MARK sites S262/S356 largely unchanged (maximum reduction=18%) (Fig. 3B). 17-AAG significantly reduced all three phospho-species; however, the reductions in S202/T205 (74%) and S396/S404 (77%) were much greater than that of S262/S356 (30%). One of the compounds (EC107) failed to significantly (P>0.05) reduce any of the phospho-tau species, while EC108 produced only a small (18%) reduction in tau phosphorylated at S262/S356 that was significant.
The same compounds that produced >50% reductions in the levels of S202/T205 and S396/S404 phospho-tau species also significantly reduced the levels of conformationally altered tau with the MC-1 epitope (Fig. 3B). Both EC104 and 17-AAG eliminated >90% MC-1 positive-tau, while EC82, EC86, EC102, and EC105 reduced this species by 50–75%. Neither EC107 nor EC108 had any effect on MC-1 tau levels.
The MC-1 positive tau species and Tau phosphorylated at the proline directed Ser/Thr sites, S202/T205 and S396/S404, was reduced to a greater extent by EC82, 86, 102, 104, 105 and 17-AAG than total tau. These differences in the extent of the reduction were significant (t test pvalue of <0.05) and demonstrate that the HSP90 inhibitors selectively clear these tau protein species that accumulate in human tauopathy.
GAPDH levels remained unchanged after treatment with each of the compounds except for EC104, which produced a small (15%) but significant reduction. This finding along with LDH analysis (not shown) suggests that the observed reductions in total tau and in specific tau species produced by HSP90 inhibitor treatment are unlikely to have been caused by toxicity in the 4R0N P301L tau cell line.
To verify the in-cell Western blot findings, we repeated our analyses of different tau species after treatment of the 4R0N P301L tau cell line with the HSP90 inhibitors 17-AAG, EC-102, and EC107 using standard Western blots (Fig. 4). Although Western blots lack the quantitative acuity of the in-cell Western blot analysis, it is clear that treatment with 17-AAG and EC102 again produced an induction of the chaperones HSP70, HSP40, and HSP27, concomitant with reductions in total tau, pS202/T205 tau, and pS396/S404 tau. In contrast, tau phosphorylated by MARK at sites S262 and S356 was not reduced to the same extent, although 17-AAG again caused a significant reduction (60%) in this species, as measured by pixel density (also detected by the in-cell Western blot analysis results in Fig. 2B). As expected, EC107 caused a marginal induction of HSPs and, as in the in-cell Western blot results, had little effect on the levels of each
Page 6 of 18(page number not for citation purposes)

of the tau species. Overall, the standard Western blot analysis verified the original in-cell Western assays performed in the CHO tau (P301L).
One possible result of HSP90 inhibitor treatment might have been neuronal microtubule instability either because of tau depletion or because of non-specific toxic effects on the cells due to HSP90 client-protein degradation. To determine the effects of HSP90 inhibitor treatment on the microtubule network, we cultured primary cortical neurons from 2 day-old Swiss Webster pups and treated mature neurons (day 7–10) with 17-AAG, EC102 and EC107. After 24 h treatment, the colocalization of tau with the microtubule network was visualized by fluorescent immunostaining for total tau (red) and tubulin (green). Tau within postmitotic neurons, treated with these compounds remained colocalized with intact tubulin (Fig. 5). Neuritic processes were also maintained, consistent with previous observations by Dai and colleagues (20) (Fig. 5). Indeed, we observed no qualitative difference in the microtubule network between control (vehicle treated) and HSP90 inhibitor treated neurons (Fig. 5; EC102 not shown).
To determine the mechanism of tau degradation elicited by HSP90 inhibition, we again treated the CHO tau (P301L) cell line with the panel of HSP90 inhibitors, this time in the presence of the proteasomal inhibitor MG-132. Tau protein species were again quantified using the In-Cell Western blot assay. Treatment with MG-132 completely blocked all HSP90 inhibitor-mediated reduction of total tau and tau phosphorylated at proline-directed Ser/Thr sites (S202/T205 and S396/S404; Fig. 6A). Proteasomal inhibition also blocked the clearance of conformationally altered tau species (MC-1) produced by HSP90 inhibition (not shown). Exposure to MG-132 alone however did not significantly alter total tau levels (5%±6% reduction in total tau levels compared with vehicle treated alone and 6%±6% compared with GAPDH levels). In addition, no evidence of toxicity was observed based on LDH assays and unchanged GAPDH levels (not shown). Overall, these data indicate that chaperone-mediated clearance of tau species is proteasome-dependent in CHO tau (P301L) cells. In contrast, cells cotreated with HSP90 inhibitors and the lysosomal inhibitor, chloroquine, for 24 h maintained the selective reductions in pS202/T205 and pS396/S404 phospho-tau species (Fig. 6B); however, the levels of tau species in cells cotreated with the HSP90 inhibitors (EC82, 86, 102, 104, 105, and 17-AAG) and with chloroquine were significantly higher (P<0.05) than in cells treated with the HSP90 inhibitors alone. Chloroquine treatment alone did not significantly reduce tau levels compared with vehicle or GAPDH levels (1%±8% and 3%±5% reductions, respectively). This result suggests that although HSP90 inhibitor mediated clearance of tau is proteasome-dependent, lysosomal function is likely also necessary for this process to work at maximal efficiency.
DISCUSSION
High-throughput fluorescent assays were developed to measure the levels of different tau species within cells as a means of identifying compounds that might have therapeutic benefit in tauopathy (19). These in-cell Western assays were used to examine the effects of a panel of seven novel small-molecule HSP90 inhibitors on the clearance of different tau species in the CHO tau(P301L) cell line (Fig. 3). The size of these novel compounds and their predicted pharmacokinetic properties suggests that they will cross the blood-brain barrier at levels sufficient to impact chaperone levels within the brain.
These studies showed that HSP90 inhibitors producing the greatest induction of molecular chaperones were able to selectively lower levels of tau phosphorylated at the proline-directed
Page 7 of 18(page number not for citation purposes)

Ser/Thr sites pS202/T205 and pS396/S404. Tau phosphorylated at these epitopes accumulates during the development of AD and other tauopathies, whereas these species are absent or present at low levels in the normal adult human brain. In addition, because of the nondisruptive conditions of the assay, we were also able to show selective reductions in conformationally altered tau that is detected by the MC-1 antibody. The MC-1 epitope is formed when tau takes on a folded conformation in which the N terminus interacts with the microtubule binding region (21). Formation of this abnormal folded conformation is thought to be one of the earliest pathological changes observed in patients with tauopathy and is associated with the initiation of tau filament formation. (21). This is the first time that this disease-associated tau conformational epitope has been quantitatively analyzed in vitro as part of a high-throughput assay.
The mechanism by which HSP90 inhibition led to enhanced tau clearance in H4 and CHO tau(P301L) cells is an obvious question raised by our experiments. The reduction in total tau and in the specific modified tau species broadly follows the degree of elevation in HSPs (27 and 70) generated by each inhibitor (Figs. 1, 2); however, the complexity of the heat shock response would predict that other factors are equally if not more important for the degradation of tau (i.e., CHIP, HSP40, HSP90, etc.), particularly as HSP70 levels are not linearly dose responsive in this system. Still, the results presented here consistently indicate that induction of de novo chaperone (HSP) synthesis is the primary mechanism of HSP90 inhibitor-mediated tau clearance. Additional studies are ongoing to determine whether induction of one specific HSP is sufficient or whether the totality of the heat shock response is crucial for tau removal. In this respect, it is important to note that both HSP70 (10) and HSP27 (8) have previously been shown to play crucial roles in two independent pathways involved in the selective degradation of hyperphosphorylated tau species via the proteasome. The relationship between the increase in HSPs and tau clearance further implies that either 1) HSP90-inhibiting compounds that significantly lower tau are able to elevate HSPs beyond some threshold or 2) a defined time period of HSP induction is required to significantly enhance tau clearance.
Our findings further suggest that HSP-mediated tau clearance is likely proteasome-dependent, as evidenced by the failure of the HSP90 inhibitors to cause reductions in any of the assayed tau species in the presence of a proteasomal inhibitor (Fig. 5A). The fact that proteasomal inhibition blocks the clearance of phospho-tau species (pS202/T205 and pS396/S404) also makes it unlikely that the reduction in the levels of these species, produced by the HSP90 inhibitors, was mediated by alteration in the activity of specific kinases/phosphatases rather than by enhanced proteolytic degradation. It should be pointed out that proteasomal inhibition alone is sufficient to induce HSP expression independent of other stimuli. Despite this potential complication, tau degradation was indeed prevented by proteasomal inhibition, regardless of whether or not HSP expression was enhanced by the presence of MG132. It is also interesting to note, that although treatment with the lysosomal inhibitor chloroquine (Fig. 5B) did not block the reduction in tau species mediated by HSP90 inhibitor treatment it did significantly ablate the extent to which tau levels were reduced. It is therefore possible that although proteasome-dependent, the lysosome is a secondary site of HSP-mediated tau degradation.
One of the most striking results of these studies was the minimal effect that HSP90 inhibitor treatment had on levels of tau phosphorylated by MARK, (pS262/S356 tau species). This is in direct contrast to the selective degradation of tau phosphorylated at the proline-directed Ser/Thr sites, S202/T205 and S396/S404, in the CHO tau(P301L) cell line. Therefore, it appears that tau phosphorylated within the microtubule binding domains by MARK is likely resistant to
Page 8 of 18(page number not for citation purposes)

degradation by HSP90 inhibition. A potential reason why tau phosphorylated at S262/S356 is not reduced by HSP90 inhibition, while tau species phosphorylated at the proline-directed S202/T205 and S396/S404 sites are reduced, could be the differences in the respective protein recognition motifs. Specifically, the KXGS motifs (S262/S356) that are phosphorylated by MARK/PAR-1 may not be recognized by the relevant chaperones induced by HSP90 inhibition because they lack the proline residue that is a consistent feature of the other Ser/Thr phosphorylation sites within tau (22).
The functional significance of the apparent resistance of tau phosphorylated at S262/S356 to degradation is an important question. Transient phosphorylation of tau by MARK, within the microtubule binding domains, causes tau release from the microtubules and consequently microtubule destabilization. MARK is thus able to regulate microtubule dynamics, which is important for axonal growth and other processes that require neuronal plasticity. It is possible that tau phosphorylated by MARK needs to be resistant to HSP-mediated degradation so that it can provide a pool of unbound tau that is then available to rapidly bind microtubules after dephosphorylation. In this way the microtubule network could then be restablized as required after axonal growth or synaptic remodeling (5).
The resistance of pS262/S356 tau to degradation by HSP induction may also partially explain the reported role of tau phosphorylated by MARK in the initiation of pathogenesis in tauopathy. Nishimura et al. (6) recently demonstrated in a Drosophila model of tauopathy, that other deleterious tau phosphorylation events on proline-directed Ser/Thr sites require prior phosphorylation by PAR-1 (the Drosophila homologue of MARK) that allows tau to dissociate from the microtubules. This result implicated MARK/PAR-1 as the “master” kinase for subsequent pathological phosphorylation and accumulation of tau in the disease state (6). As a result, the resistance of pS262/S356 tau to molecular chaperone-mediated degradation, could have the secondary consequence of allowing the initiation of pathogenic tau phosphorylation on proline-directed Ser/Thr sites during the development of tauopathy.
Taken together, our results suggest that HSP90 inhibitors that penetrate the blood-brain barrier may have the potential for modifying the pathogenic accumulation of abnormal tau species during pathogenesis in tauopathy. Importantly, however, MARK phosphorylated tau (pS262/S356) was resistant to HSP90 inhibitor-mediated degradation. The resistance of pS262/S356 tau to HSP90 inhibitor induced degradation may reflect the role of MARK in regulating microtubule stability, and in addition, it may also explain how accumulation of these phospho-tau species initiates the pathogenic cascade that leads to the development of neurodegenerative tauopathy.
ACKNOWLEDGMENTS
The work presented here received generous support from the NIA (PO1 AG17216 to MH), The Florida Alzheimer's Center Research Initiative, the Byrd Institute and the Institute for the study of Aging. We would like to thank Dr. Peter Seubert for graciously providing the 12E8 antibody, Dr. Peter Davies for the PHF-1, CP13, and MC1 antibodies, and Dr. Lester Binder for the Tau5 antibody.
Page 9 of 18(page number not for citation purposes)

REFERENCES
1. Hutton, M., Lendon, C. L., Rizzu, P., Baker, M., Froelich, S., Houlden, H., Pickering-Brown, S., Chakraverty, S., Isaacs, A., Grover, A., et al. (1998) Association of missense and 5′-splice-site mutations in tau with the inherited dementia FTDP-17. Nature 393, 702–705
2. Dumanchin, C., Camuzat, A., Campion, D., Verpillat, P., Hannequin, D., Dubois, B., Saugier-Veber, P., Martin, C., Penet, C., Charbonnier, F., et al. (1998) Segregation of a missense mutation in the microtubule-associated protein tau gene with familial frontotemporal dementia and parkinsonism. Hum. Mol. Genet. 7, 1825–1829
3. Spillantini, M. G., Murrell, J. R., Goedert, M., Farlow, M. R., Klug, A., and Ghetti, B. (1998) Mutation in the tau gene in familial multiple system tauopathy with presenile dementia. Proc. Natl. Acad. Sci. USA 95, 7737–7741
4. Biernat, J., Wu, Y. Z., Timm, T., Zheng-Fischhofer, Q., Mandelkow, E., Meijer, L., and Mandelkow, E. M. (2002) Protein kinase MARK/PAR-1 is required for neurite outgrowth and establishment of neuronal polarity. Mol. Biol. Cell 13, 4013–4028
5. Drewes, G., Ebneth, A., Preuss, U., Mandelkow, E. M., and Mandelkow, E. (1997) MARK, a novel family of protein kinases that phosphorylate microtubule-associated proteins and trigger microtubule disruption. Cell 89, 297–308
6. Nishimura, I., Yang, Y., and Lu, B. (2004) PAR-1 kinase plays an initiator role in a temporally ordered phosphorylation process that confers tau toxicity in Drosophila. Cell 116, 671–682
7. Weaver, C. L., Espinoza, M., Kress, Y., and Davies, P. (2000) Conformational change as one of the earliest alterations of tau in Alzheimer's disease. Neurobiol. Aging 21, 719–727
8. Shimura, H., Miura-Shimura, Y., and Kosik, K. S. (2004) Binding of tau to heat shock protein 27 leads to decreased concentration of hyperphosphorylated tau and enhanced cell survival. J. Biol. Chem. 279, 17957–17962
9. Bagatell, R., Whitesell, L., Shimura, H., Miura-Shimura, Y., Kosik, K. S., Zhou, Y., Gu, G., Goodlett, D. R., Zhang, T., Pan, C., et al. (2004) Altered Hsp90 function in cancer: a unique therapeutic opportunity. Mol. Cancer Ther. 3, 1021–1030
10. Shimura, H., Schwartz, D., Gygi, S. P., and Kosik, K. S. (2004) CHIP-Hsc70 complex ubiquitinates phosphorylated tau and enhances cell survival. J. Biol. Chem. 279, 4869–4876
11. Petrucelli, L., Dickson, D., Kehoe, K., Taylor, J., Snyder, H., Grover, A., De Lucia, M., McGowan, E., Lewis, J., Prihar, G., et al. (2004) CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum. Mol. Genet. 13, 703–714
12. McDonough, H., and Patterson, C. (2003) CHIP: a link between the chaperone and proteasome systems. Cell Stress Chaperones 8, 303–308
Page 10 of 18(page number not for citation purposes)

13. Kamal, A., Boehm, M. F., and Burrows, F. J. (2004) Therapeutic and diagnostic implications of Hsp90 activation. Trends Mol. Med. 10, 283–290
14. Petrucelli, L., O'Farrell, C., Lockhart, P. J., Baptista, M., Kehoe, K., Vink, L., Choi, P., Wolozin, B., Farrer, M., Hardy, J., et al. (2002) Parkin protects against the toxicity associated with mutant alpha-synuclein: proteasome dysfunction selectively affects catecholaminergic neurons. Neuron 36, 1007–1019
15. Seubert, P., Mawal-Dewan, M., Barbour, R., Jakes, R., Goedert, M., Johnson, G. V., Litersky, J. M., Schenk, D., Lieberburg, I., Trojanowski, J. Q., et al. (1995) Detection of phosphorylated Ser262 in fetal tau, adult tau, and paired helical filament tau. J. Biol. Chem. 270, 18917–18922
16. Sun, L., Liu, S. Y., Zhou, X. W., Wang, X. C., Liu, R., Wang, Q., and Wang, J. Z. (2003) Inhibition of protein phosphatase 2A- and protein phosphatase 1-induced tau hyperphosphorylation and impairment of spatial memory retention in rats. Neuroscience 118, 1175–1182
17. Shen, H.Y., He, J.C., Wang, Y., Huang, Q.Y. and Chen, J.F. (2005) Geldanamycin induces heat shock protein 70 and protects against MPTP-induced dopaminergic neurotoxicity in mice. J Biol Chem.
18. Waza, M., Adachi, H., Katsuno, M., Minamiyama, M., Sang, C., Tanaka, F., Inukai, A., Doyu, M., and Sobue, G. (2005) 17-AAG, an Hsp90 inhibitor, ameliorates polyglutamine-mediated motor neuron degeneration. Nat. Med. 11, 1088–1095
19. Dickey, C. A., Eriksen, J., Kamal, A., Burrows, F., Kasibhatla, S., Eckman, C. B., Hutton, M., and Petrucelli, L. (2005) Development of a high throughput drug screening assay for the detection of changes in tau levels—proof of concept with HSP90 inhibitors. Curr. Alzheimer Res. 2, 231–238
20. Dai, Q., Zhang, C., Wu, Y., McDonough, H., Whaley, R. A., Godfrey, V., Li, H. H., Madamanchi, N., Xu, W., Neckers, L., et al. (2003) CHIP activates HSF1 and confers protection against apoptosis and cellular stress. EMBO J. 22, 5446–5458
21. Hyman, B. T., Van Hoesen, G. W., Wolozin, B. L., Davies, P., Kromer, L. J., and Damasio, A. R. (1988) Alz-50 antibody recognizes Alzheimer-related neuronal changes. Ann. Neurol. 23, 371–379
22. Trinczek, B., Biernat, J., Baumann, K., Mandelkow, E. M., and Mandelkow, E. (1995) Domains of tau protein, differential phosphorylation, and dynamic instability of microtubules. Mol. Biol. Cell 6, 1887–1902
Received October 25, 2005; accepted December 15, 2005
Page 11 of 18(page number not for citation purposes)

Table 1 Tau antibody nomenclature
Antibody Host Species Tau Epitope Dako Tau Rabbit All isoforms, non-species specific E1 Rabbit All isoforms, human specific Tau5 Mouse All isoforms, non-species specific CP13 Mouse Tau phosphorylated by GSK3β/CdK5 at pS202/T205 PHF-1 Mouse Tau phosphorylated by GSK3β/CdK5 at pS396/S404 12E8 Mouse Tau phosphorylated by MARK(PAR-1) at pS262/S356 MC1 Mouse Pre-paired helical filament conformational epitope
Page 12 of 18(page number not for citation purposes)
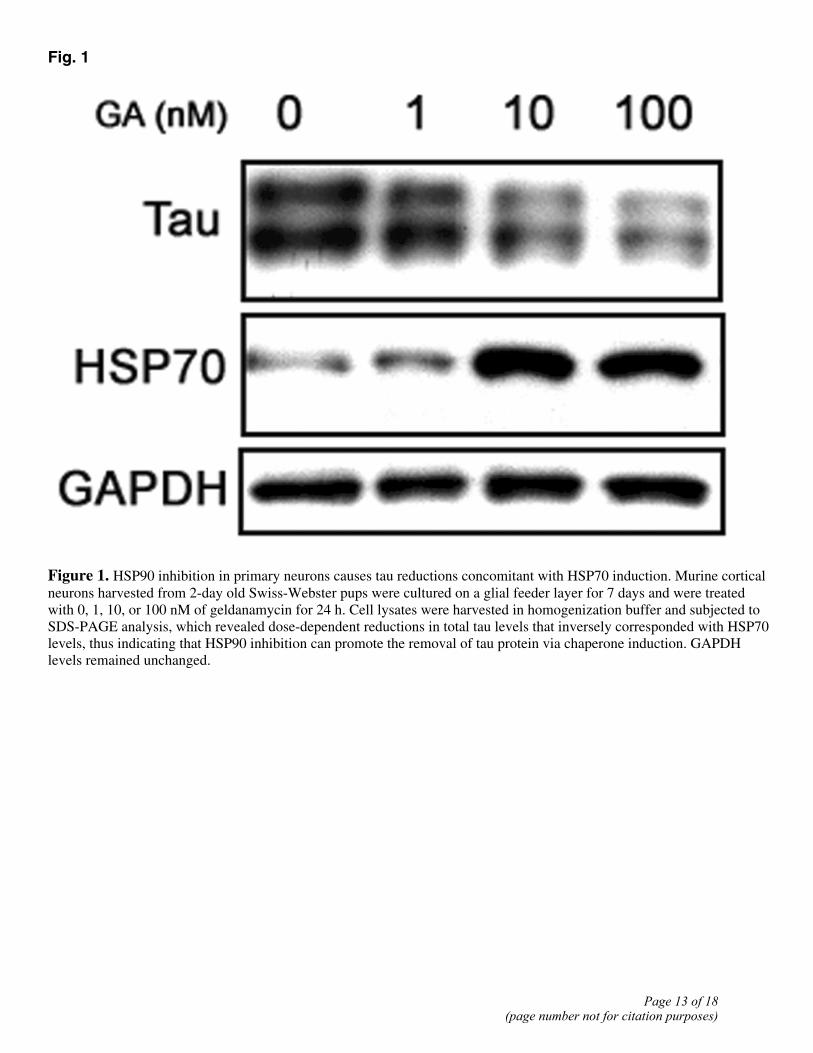
Fig. 1
Figure 1. HSP90 inhibition in primary neurons causes tau reductions concomitant with HSP70 induction. Murine cortical neurons harvested from 2-day old Swiss-Webster pups were cultured on a glial feeder layer for 7 days and were treated with 0, 1, 10, or 100 nM of geldanamycin for 24 h. Cell lysates were harvested in homogenization buffer and subjected to SDS-PAGE analysis, which revealed dose-dependent reductions in total tau levels that inversely corresponded with HSP70 levels, thus indicating that HSP90 inhibition can promote the removal of tau protein via chaperone induction. GAPDH levels remained unchanged.
Page 13 of 18(page number not for citation purposes)
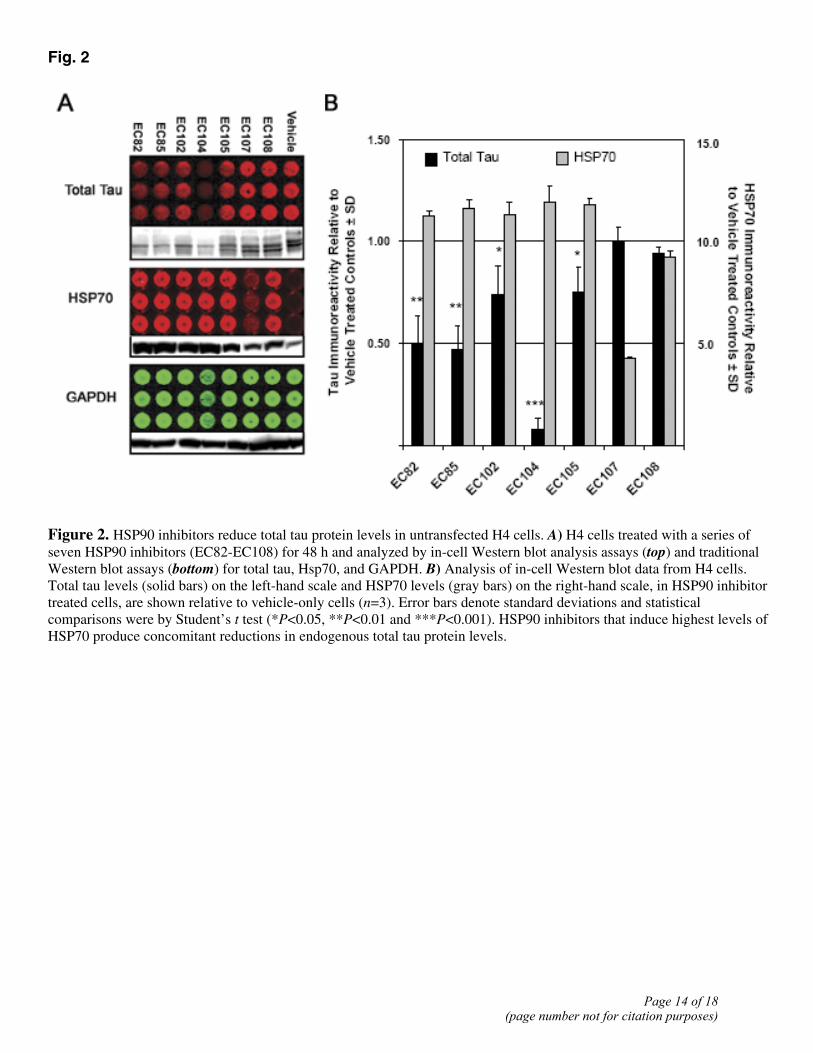
Fig. 2
Figure 2. HSP90 inhibitors reduce total tau protein levels in untransfected H4 cells. A) H4 cells treated with a series of seven HSP90 inhibitors (EC82-EC108) for 48 h and analyzed by in-cell Western blot analysis assays (top) and traditional Western blot assays (bottom) for total tau, Hsp70, and GAPDH. B) Analysis of in-cell Western blot data from H4 cells. Total tau levels (solid bars) on the left-hand scale and HSP70 levels (gray bars) on the right-hand scale, in HSP90 inhibitor treated cells, are shown relative to vehicle-only cells (n=3). Error bars denote standard deviations and statistical comparisons were by Student’s t test (*P<0.05, **P<0.01 and ***P<0.001). HSP90 inhibitors that induce highest levels of HSP70 produce concomitant reductions in endogenous total tau protein levels.
Page 14 of 18(page number not for citation purposes)
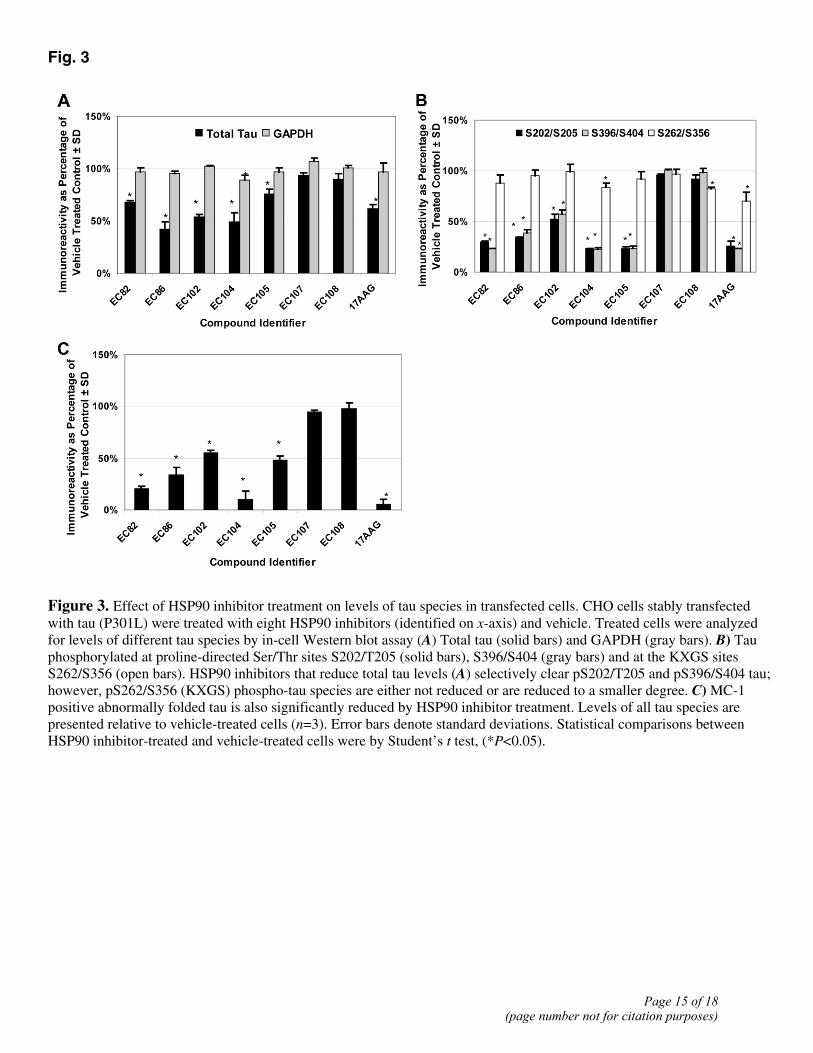
Fig. 3
Figure 3. Effect of HSP90 inhibitor treatment on levels of tau species in transfected cells. CHO cells stably transfected with tau (P301L) were treated with eight HSP90 inhibitors (identified on x-axis) and vehicle. Treated cells were analyzed for levels of different tau species by in-cell Western blot assay (A) Total tau (solid bars) and GAPDH (gray bars). B) Tau phosphorylated at proline-directed Ser/Thr sites S202/T205 (solid bars), S396/S404 (gray bars) and at the KXGS sites S262/S356 (open bars). HSP90 inhibitors that reduce total tau levels (A) selectively clear pS202/T205 and pS396/S404 tau; however, pS262/S356 (KXGS) phospho-tau species are either not reduced or are reduced to a smaller degree. C) MC-1 positive abnormally folded tau is also significantly reduced by HSP90 inhibitor treatment. Levels of all tau species are presented relative to vehicle-treated cells (n=3). Error bars denote standard deviations. Statistical comparisons between HSP90 inhibitor-treated and vehicle-treated cells were by Student’s t test, (*P<0.05).
Page 15 of 18(page number not for citation purposes)
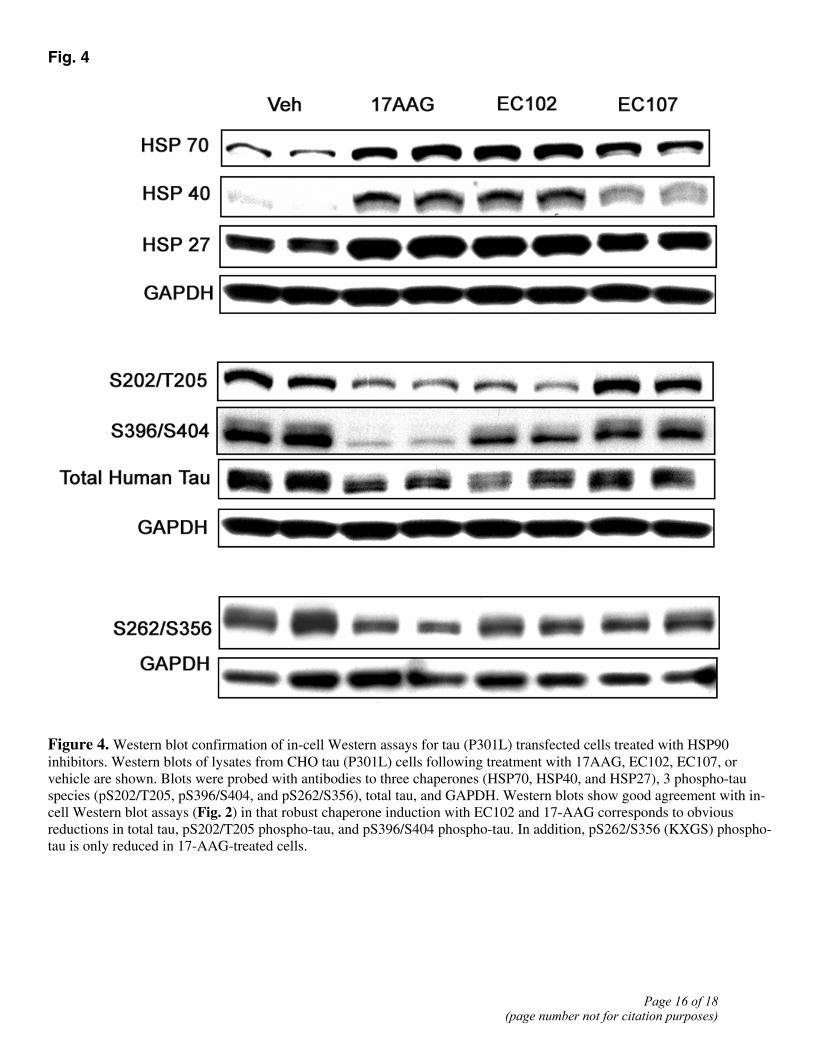
Fig. 4
Figure 4. Western blot confirmation of in-cell Western assays for tau (P301L) transfected cells treated with HSP90 inhibitors. Western blots of lysates from CHO tau (P301L) cells following treatment with 17AAG, EC102, EC107, or vehicle are shown. Blots were probed with antibodies to three chaperones (HSP70, HSP40, and HSP27), 3 phospho-tau species (pS202/T205, pS396/S404, and pS262/S356), total tau, and GAPDH. Western blots show good agreement with in-cell Western blot assays (Fig. 2) in that robust chaperone induction with EC102 and 17-AAG corresponds to obvious reductions in total tau, pS202/T205 phospho-tau, and pS396/S404 phospho-tau. In addition, pS262/S356 (KXGS) phospho-tau is only reduced in 17-AAG-treated cells.
Page 16 of 18(page number not for citation purposes)

Fig. 5
Figure 5. Treatment of primary cortical neurons with HSP90 inhibitor 17-AAG does not result in altered tau distribution. Primary neurons were treated with a 1-µM concentration of 17AAG, EC107 (negative control), or vehicle. The distribution of human tau was then determined by immunohistochemistry with the E1 anti-tau antibody (middle). The microtubule network was visualized with an anti-tubulin monoclonal antibody (top). Colocalization between tau and the microtubule network is visualized in the merged image (bottom). Treatment with 17-AAG, which produces the greatest reduction in tau species (Figs. 2, 3), has no obvious effect on the colocalization of tau with microtubules in the primary neurons.
Page 17 of 18(page number not for citation purposes)
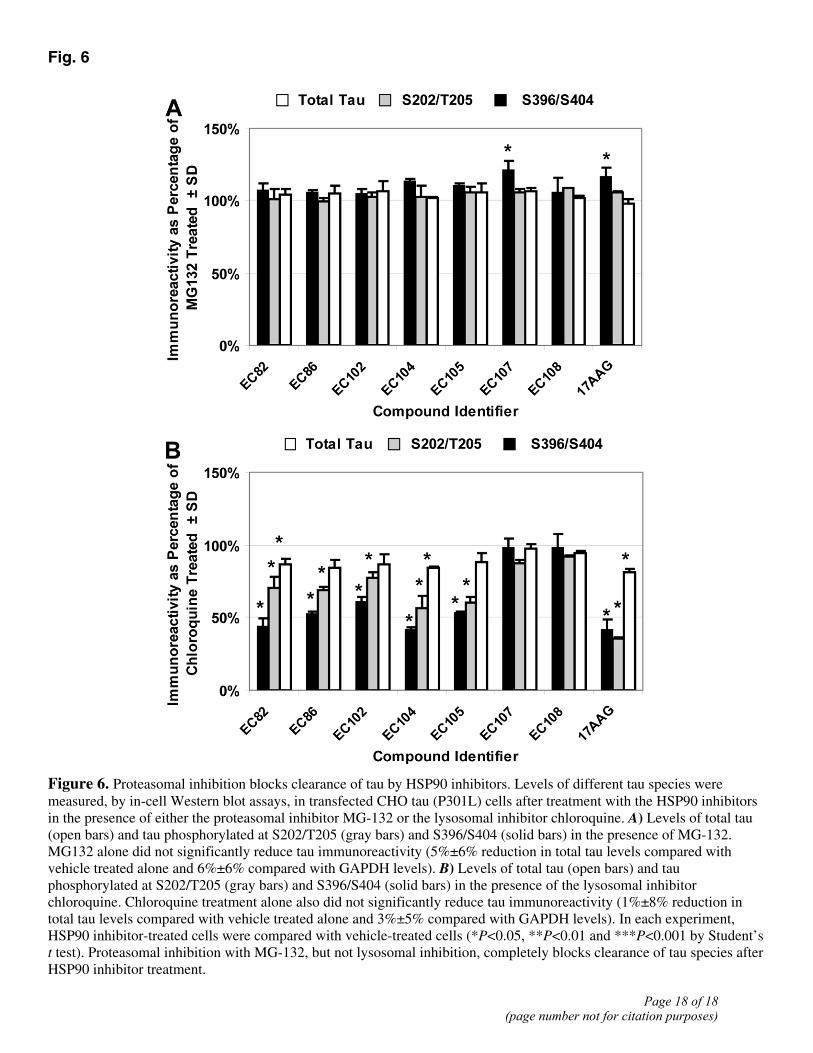
Fig. 6
Figure 6. Proteasomal inhibition blocks clearance of tau by HSP90 inhibitors. Levels of different tau species were measured, by in-cell Western blot assays, in transfected CHO tau (P301L) cells after treatment with the HSP90 inhibitors in the presence of either the proteasomal inhibitor MG-132 or the lysosomal inhibitor chloroquine. A) Levels of total tau (open bars) and tau phosphorylated at S202/T205 (gray bars) and S396/S404 (solid bars) in the presence of MG-132. MG132 alone did not significantly reduce tau immunoreactivity (5%±6% reduction in total tau levels compared with vehicle treated alone and 6%±6% compared with GAPDH levels). B) Levels of total tau (open bars) and tau phosphorylated at S202/T205 (gray bars) and S396/S404 (solid bars) in the presence of the lysosomal inhibitor chloroquine. Chloroquine treatment alone also did not significantly reduce tau immunoreactivity (1%±8% reduction in total tau levels compared with vehicle treated alone and 3%±5% compared with GAPDH levels). In each experiment, HSP90 inhibitor-treated cells were compared with vehicle-treated cells (*P<0.05, **P<0.01 and ***P<0.001 by Student’s t test). Proteasomal inhibition with MG-132, but not lysosomal inhibition, completely blocks clearance of tau species after HSP90 inhibitor treatment.
Page 18 of 18(page number not for citation purposes)
Related Documents











