How and why did the Lingulidae (Brachiopoda) not only survive the end-Permian mass extinction but also thrive in its aftermath? Yuanqiao Peng a,b, ⁎ , G.R. Shi a,b , Yongqun Gao a,b , Weihong He b , Shuzhong Shen c a School of Life and Environmental Sciences, Deakin University, Melbourne Campus, 221 Burwood Highway, Burwood, VIC 3125, Australia b Faculty of Earth Sciences, China University of Geosciences, Wuhan, Hubei Province 430074, PR China c Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, Jiangsu Province 210008, PR China Accepted 30 November 2006 Abstract The end-Permian mass extinction devastated most of the organisms in the sea and on land. However, a few Late Permian taxa survived the mass extinction and also the subsequent Early Triassic post-extinction catastrophic environments. Among them, the Lingulidae brachiopods were perhaps one of the most noted conquerors. Not only had they survived the mass extinction, but also they thrived in the Early Triassic marine realm, forming a nearly globally distributed Lingulidae fauna characterized by low taxonomic diversity but high abundance of individuals. The reasons that this group of brachiopods was able to achieve this remain unclear, and could possibly include a number of interacting factors. Among them are their wide ecological adaptations and their shell composition and appearance. The long-term evolutionary trends in the shell microstructure and exterior of the Lingulidae since the Late Devonian are thought to have advanced considerably by the end-Permian to the extent that the shells of the brachiopods were able to sustain stressful (e.g. anoxic) environments. In addition, a variety of short-term adaptive changes in the shell size (reducing shell size), shape (shells became more pointed and flatter anteriorly), and thickness (reducing shell thickness) are believed to have also aided the survival of the brachiopods through the highly toxic marine environments from the latest Permian to the Early Triassic. The long-term unchanged organophosphatic shell composition of Lingulidae brachiopods is considered to be another key feature that must have aided the survival of this group in the end-Permian to the Early Triassic Period, for organophosphatic shell composition would have prevented the brachiopods from dissolution in the highly acidic marine environment prevailing globally during the Permian–Triassic transition. The ability that the lingulids had to live in a wide range of environmental conditions, from high-latitude to low-latitude and from the shoreface to the relatively deep sea environments, is also considered vital for the group to have managed to survive the catastrophic event in the latest Permian and then thrive in the vacated ecological space in the ensuing Early Triassic disaster period. © 2007 Elsevier B.V. All rights reserved. Keywords: Permian–Triassic Boundary (PTB); Mass extinction; Early Triassic; Lingulidae; Adaptive changes 1. Introduction As the greatest of all mass extinctions in deep time, the Permian–Triassic Boundary (PTB) mass extinction witnessed more than 90% loss of all species in the marine realm, and about 70% loss of continental species in the non-marine realm (Erwin, 1993, 1994). No reef Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118 – 131 www.elsevier.com/locate/palaeo ⁎ Corresponding author. Present address: School of Life and Environmental Sciences, Deakin Universtiy, Melbourne Campus, 221 Burwood Highway, Burwood, VIC 3125, Australia. E-mail address: [email protected] (Y. Peng). 0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2006.11.039

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

alaeoecology 252 (2007) 118–131www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
How and why did the Lingulidae (Brachiopoda) not only survive theend-Permian mass extinction but also thrive in its aftermath?
Yuanqiao Peng a,b,⁎, G.R. Shi a,b, Yongqun Gao a,b, Weihong He b, Shuzhong Shen c
a School of Life and Environmental Sciences, Deakin University, Melbourne Campus, 221 Burwood Highway, Burwood, VIC 3125, Australiab Faculty of Earth Sciences, China University of Geosciences, Wuhan, Hubei Province 430074, PR China
c Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, Jiangsu Province 210008, PR China
Accepted 30 November 2006
Abstract
The end-Permian mass extinction devastated most of the organisms in the sea and on land. However, a few Late Permian taxasurvived the mass extinction and also the subsequent Early Triassic post-extinction catastrophic environments. Among them, theLingulidae brachiopods were perhaps one of the most noted conquerors. Not only had they survived the mass extinction, but alsothey thrived in the Early Triassic marine realm, forming a nearly globally distributed Lingulidae fauna characterized by lowtaxonomic diversity but high abundance of individuals. The reasons that this group of brachiopods was able to achieve this remainunclear, and could possibly include a number of interacting factors. Among them are their wide ecological adaptations and theirshell composition and appearance. The long-term evolutionary trends in the shell microstructure and exterior of the Lingulidaesince the Late Devonian are thought to have advanced considerably by the end-Permian to the extent that the shells of thebrachiopods were able to sustain stressful (e.g. anoxic) environments. In addition, a variety of short-term adaptive changes in theshell size (reducing shell size), shape (shells became more pointed and flatter anteriorly), and thickness (reducing shell thickness)are believed to have also aided the survival of the brachiopods through the highly toxic marine environments from the latestPermian to the Early Triassic. The long-term unchanged organophosphatic shell composition of Lingulidae brachiopods isconsidered to be another key feature that must have aided the survival of this group in the end-Permian to the Early Triassic Period,for organophosphatic shell composition would have prevented the brachiopods from dissolution in the highly acidic marineenvironment prevailing globally during the Permian–Triassic transition. The ability that the lingulids had to live in a wide range ofenvironmental conditions, from high-latitude to low-latitude and from the shoreface to the relatively deep sea environments, is alsoconsidered vital for the group to have managed to survive the catastrophic event in the latest Permian and then thrive in the vacatedecological space in the ensuing Early Triassic disaster period.© 2007 Elsevier B.V. All rights reserved.
Keywords: Permian–Triassic Boundary (PTB); Mass extinction; Early Triassic; Lingulidae; Adaptive changes
⁎ Corresponding author. Present address: School of Life andEnvironmental Sciences, Deakin Universtiy, Melbourne Campus,221 Burwood Highway, Burwood, VIC 3125, Australia.
E-mail address: [email protected] (Y. Peng).
0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2006.11.039
1. Introduction
As the greatest of all mass extinctions in deep time,the Permian–Triassic Boundary (PTB) mass extinctionwitnessed more than 90% loss of all species in themarine realm, and about 70% loss of continental speciesin the non-marine realm (Erwin, 1993, 1994). No reef

119Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
and no coal beds/seams have ever been found anywherearound the world in the Lower Triassic stratigraphicrecords, leading to suggestions of the so-called “reefgap” (Flugel, 1994) and “coal gap” (Retallack et al.,1996) respectively in geological history. In the mean-time, however, the Permian–Triassic transition is alsomarked by another feature of parallel significance,characterized by the survival of the so-called “Lilliput”fauna (Urbanek, 1993) that thrived in the Early Triassicpost-extinction period (Twitchett, 2001). Globally, thissurvival fauna was composed of small-sized and/or thin-shelled, low-diversity, high-abundance marine commu-nities, among which the Lingulidae brachiopods aremost notable (Xu and Grant, 1994; Rodland and Bottjer,2001).
Brachiopod faunas were very abundant and diversi-fied in the marine realm during the Late Paleozoic, butwere drastically reduced in species richness in the EarlyTriassic after nearly 87–90% of genera and 94–96% ofspecies became extinct at the end of the Permian (Shiand Shen, 2000; Shen and Shi, 2002). Compared tohundreds of species found in the Late Permian deposits,only a few species of brachiopods occur in the earliestTriassic strata. Among these low-diversity but high-density survivors, the Lingulidae proliferated most inthe earliest Triassic marine realm (Xu and Grant, 1994and references therein), forming a cosmopolitan Lingu-lidae assemblage, as the bivalve Claraia and theammonoid Ophiceras did in the aftermath of the end-Permian mass extinction (Hallam and Wignall, 1997).Despite this recognition, the taxonomic identities of theEarly Triassic Lingulidae remain problematic. Untilrecently, the Early Triassic Lingulidae had beendescribed or reported all as Lingula (see reviews in Xuand Grant, 1994), but Biernat and Emig (1993) haverecognized two probable distinct genera among theEarly Triassic lingulids: Lingularia and/or Barroisella.Likewise, after detailed examination of hundreds ofEarly Triassic lingulid specimens from South China, wehave also recognized several additional new genera andspecies (Peng and Shi, submitted for publication).
In this paper, the long-term evolutionary character-istics of lingulids have been discussed based on aglobal extensive collection of taxa on Lingulidaebrachiopods through time. In addition, we attempt toelucidate why and how the Lingulidae not onlysurvived the end-Permian mass extinction but alsothrived in the Early Triassic aftermath, based on adetailed analysis of lingulid's burrowing habits,capacity for adaptation, as well as shell microstructureand adaptive changes in shell geometry (e.g. size,shape, and thickness).
2. Method
The senior author constructed a comprehensivedatabase (Appendix 1) containing all reported Lingu-lidae genera and species and their size measurements(mainly width and length and, subordinately, height ifpossible) for 160 species occurrences and 1892measurements of the Lingulidae through geologicalhistory on the period level. The focus was especiallyon the Permian and the Triassic on the epoch level, theearliest Triassic Griesbachian is particularly separatedfrom Early Triassic as a single unit for data collecting.Most data were gathered from monographs of theLingulidae and other brachiopods from single sites orregions, although data were also gathered from generalpapers in which the Lingulidae were identified to thegenus level and/or figured to scale. Genus and specieslevel taxonomy follows the original authors except forcases where a clear consensus for an alternativeclassification has been suggested by subsequentworkers. Because the genus Lingula is only confirmedto appear from the Paleogene to the Present (Emig,2003), other specimens referred to this genus beforethe Paleogene are considered to be unconfirmedlingulids if their taxonomic positions have not beenreclassified.
All adult specimens described or figured inpublished monographs are included in the database.The sizes of individuals within a single species vary asa function of environment and age, among otherfactors, and no single specimen can account for all ofthese influences (Payne, 2005). In addition, within anyhigher-level classification, the number of individualswill vary among taxa (Payne, 2005). Therefore,determination of the average size of lingulids fromany interval depends upon how many taxa are used andhow taxa are weighted. For example, the mean size willchange depending on whether one specimen is selectedper species or all measured specimens are used.Commonly, the more data used, the closer mean sizecan be achieved.
Specimens were selected only when maximum shellwidth and/or maximum shell length could be deter-mined. The shell height of lingulids commonly cannotbe measured through figured specimens if it is notmentioned in the text, because most specimens arepreserved on bed planes and/or their figured imageslack lateral profiles to enable the measurement of shellheight. Measurements were taken from descriptionswhenever available, or measured from illustrationswith digital calipers to the nearest 0.1 mm from scaledillustrations.

120 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
3. Evolution of Lingulidae through time
Since appearing in the Late Devonian (Holmer andPopov, 2000; see Emig and Herrera, 2006 for newachievements), the Lingulidae have undergone a verylong period of evolution over the past 380 million years.The legacy of this group is today represented by twogenera: Lingula Bruguière 1797 and Glottidia Dall1870. Lingula is often considered to be one of the mostancient living fossils based on its morphologicalconsistency tracing back to their Cambrian relatives.As a result, most of the Phanerozoic lingulids have beenand are still referred to this genus by many researchers(Xu and Grant, 1994). However, as recently noted byEmig (2003), the lingulids as a group do demonstratesome traits of significant long-term evolutionarychanges (such as body size and shape) to enable themto better adapt to their burrowing lifestyles niche. It istrue that the shell ornamentation, especially theconcentric growth lines, appear to have remained verystable since the Late Devonian. Fossil and livingLingulidae species do, however, demonstrate consider-able differences in shell shape and, most importantly,inner structures. These differences have led Emig (2003)to argue that Lingula is not a living fossil. Likewise,Yegorov and Popov (1990), Biernat and Emig (1993)and Smirnova and Ushatinskaya (2001) have placedfossil lingulids in genera other than Lingula.
3.1. Evolution of the internal structures in Lingulidae
Based on a comparative study of internal structuresbetween the fossil Lingulidae and extant lingulids, Emig(2003) found that there are some morphological evolu-tionary changes among the members of the family,including the reduction of the pseudointerarea with onlyvestigial propareas, an asymmetrical disposition of thebody muscles, convergent main mantle canals (vasculalateralia) to become subparallel anteriorly, and a singleposterior adductor muscle of different shapes on bothventral and dorsal sides (Fig. 1). Some of theseevolutionary differences in morphological charactershad been recognized by previous workers, who employedthese differences to create several new genera such asLangella Oliveira (1930), Barroisella Cooper (1942),Apsilingula Williams (1977), Semilingula Popov (inYegorov and Popov, 1990), Lingularia Biernat andEmig (1993), and Credolingula Smirnova and Ushatins-kaya (2001). It is thus clear from the creation of these newgenera that the determination of the taxonomic identity offossil Lingulidae depends as much on their internalstructures as on their external linguliform shapes.
3.2. Shape evolution of the Lingulidae
The Lingulidae also show some trends of evolutionin their shell shapes through time. Generally, theLingulidae have increased their body size since theirfirst appearance in the Late Devonian, with theexception of two small reductions occurring in theTriassic and the Cretaceous (Fig. 2A, C). The long-termminimum body size of lingulids seems to have changedlittle, although the accuracy of this observation isdifficult to assess because of sampling discrepancyamong researchers for small-sized fossils (Jablonski,1996). Thus, the long-term increase of lingulid bodysize relies much on the increase of their maximum bodysize. It is also apparent that the body length of theLingulidae increased much faster than the body widthand height. This discrepancy in the rate of increase ofthe three shape parameters thus results in a systematicdecrease in the shell width/length, height/length andheight/width ratios of lingulids over time. Also of note isthe fact that although the whole shell height hasincreased over time, the height of the anterior shell ofthe lingulids actually decreased, resulting in a tendencytoward an increasingly sharper and more pointed shellshape in younger species (Emig, 2003). This would alsomean that the earliest Triassic (Griesbachian in thispaper) Lingulidae fossils are more bladed and morepointed anteriorly than those that lived either before orafter the Permian–Triassic mass extinction.
3.3. Diversity change of the Lingulidae through time
We have tabulated a detailed list of all Lingulidaegenera and species through geological time from allpublished literature. From this database, we havegenerated two simple diversity curves: one for Lingulidaegenera and the other for Lingulidae species (Figs. 3 and 4).Generally, the long-term diversity trend for the Lingulidaegenera does not seem to have changedmuch except for theinterval from the Permian to Jurassic when a significantfluctuation apparently occurred in the Middle Permian.From the Late Devonian to Early Permian (Cisuralian),the generic diversity of the Lingulidae was stable.However, this trend was rapidly reversed in the Cisuralianwith a rapid decrease in generic diversity, after which thegeneric diversity of the Lingulidae remained low until theJurassic when the diversity had clearly rebounded andformed another stable period from the Cretaceous toRecent (Fig. 3A). It should be evident from Fig. 3 that themost significant turning point for the Lingulidae in termsof their generic diversity did not coincide with the end-Permian mass extinction. Rather, it fell within the period
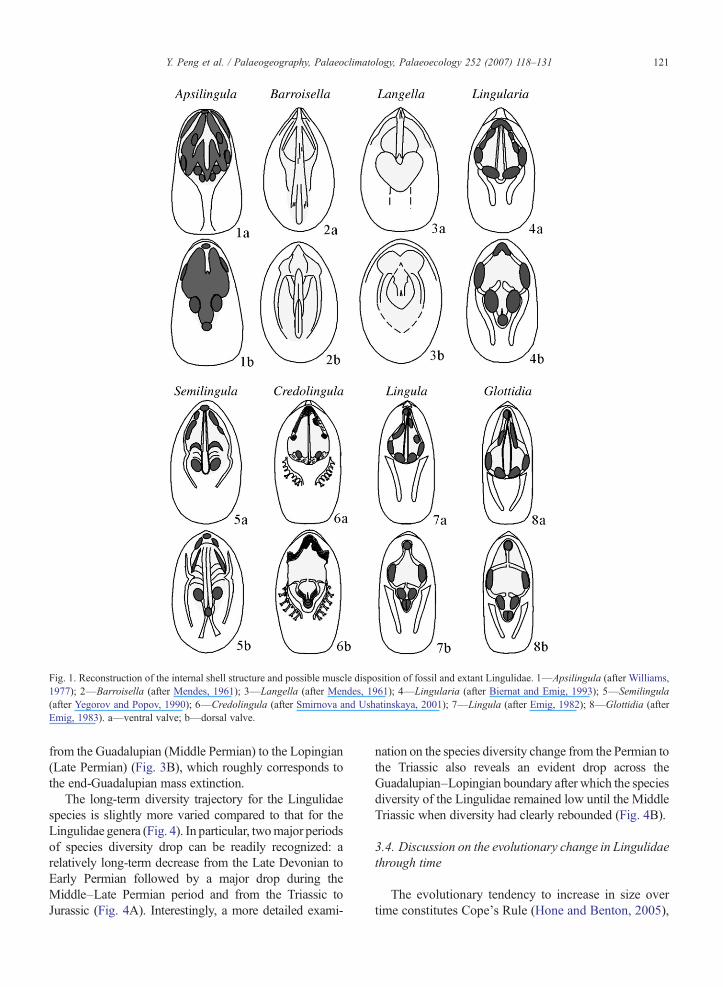
Fig. 1. Reconstruction of the internal shell structure and possible muscle disposition of fossil and extant Lingulidae. 1—Apsilingula (after Williams,1977); 2—Barroisella (after Mendes, 1961); 3—Langella (after Mendes, 1961); 4—Lingularia (after Biernat and Emig, 1993); 5—Semilingula(after Yegorov and Popov, 1990); 6—Credolingula (after Smirnova and Ushatinskaya, 2001); 7—Lingula (after Emig, 1982); 8—Glottidia (afterEmig, 1983). a—ventral valve; b—dorsal valve.
121Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
from the Guadalupian (Middle Permian) to the Lopingian(Late Permian) (Fig. 3B), which roughly corresponds tothe end-Guadalupian mass extinction.
The long-term diversity trajectory for the Lingulidaespecies is slightly more varied compared to that for theLingulidae genera (Fig. 4). In particular, twomajor periodsof species diversity drop can be readily recognized: arelatively long-term decrease from the Late Devonian toEarly Permian followed by a major drop during theMiddle–Late Permian period and from the Triassic toJurassic (Fig. 4A). Interestingly, a more detailed exami-
nation on the species diversity change from the Permian tothe Triassic also reveals an evident drop across theGuadalupian–Lopingian boundary after which the speciesdiversity of the Lingulidae remained low until the MiddleTriassic when diversity had clearly rebounded (Fig. 4B).
3.4. Discussion on the evolutionary change in Lingulidaethrough time
The evolutionary tendency to increase in size overtime constitutes Cope's Rule (Hone and Benton, 2005),
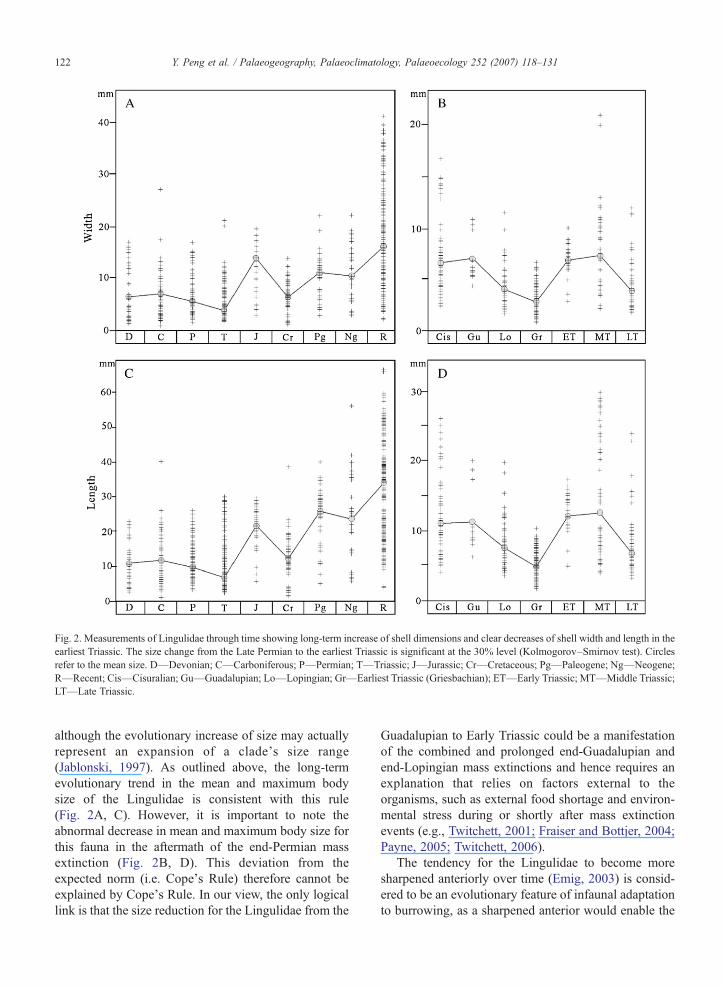
Fig. 2. Measurements of Lingulidae through time showing long-term increase of shell dimensions and clear decreases of shell width and length in theearliest Triassic. The size change from the Late Permian to the earliest Triassic is significant at the 30% level (Kolmogorov–Smirnov test). Circlesrefer to the mean size. D—Devonian; C—Carboniferous; P—Permian; T—Triassic; J—Jurassic; Cr—Cretaceous; Pg—Paleogene; Ng—Neogene;R—Recent; Cis—Cisuralian; Gu—Guadalupian; Lo—Lopingian; Gr—Earliest Triassic (Griesbachian); ET—Early Triassic; MT—Middle Triassic;LT—Late Triassic.
122 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
although the evolutionary increase of size may actuallyrepresent an expansion of a clade's size range(Jablonski, 1997). As outlined above, the long-termevolutionary trend in the mean and maximum bodysize of the Lingulidae is consistent with this rule(Fig. 2A, C). However, it is important to note theabnormal decrease in mean and maximum body size forthis fauna in the aftermath of the end-Permian massextinction (Fig. 2B, D). This deviation from theexpected norm (i.e. Cope's Rule) therefore cannot beexplained by Cope's Rule. In our view, the only logicallink is that the size reduction for the Lingulidae from the
Guadalupian to Early Triassic could be a manifestationof the combined and prolonged end-Guadalupian andend-Lopingian mass extinctions and hence requires anexplanation that relies on factors external to theorganisms, such as external food shortage and environ-mental stress during or shortly after mass extinctionevents (e.g., Twitchett, 2001; Fraiser and Bottjer, 2004;Payne, 2005; Twitchett, 2006).
The tendency for the Lingulidae to become moresharpened anteriorly over time (Emig, 2003) is consid-ered to be an evolutionary feature of infaunal adaptationto burrowing, as a sharpened anterior would enable the
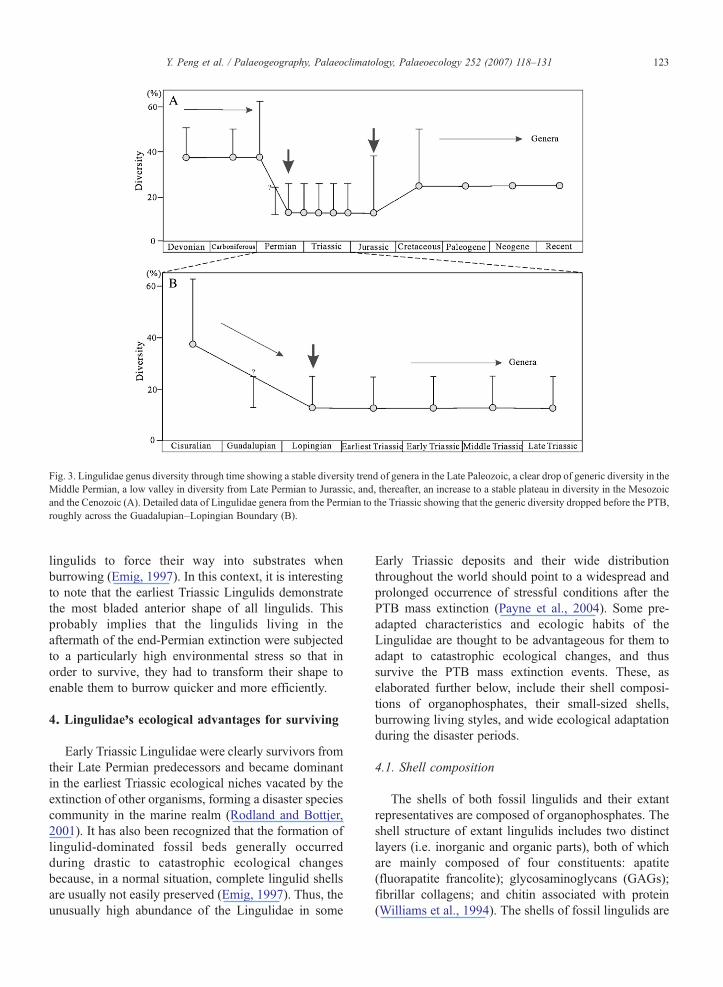
Fig. 3. Lingulidae genus diversity through time showing a stable diversity trend of genera in the Late Paleozoic, a clear drop of generic diversity in theMiddle Permian, a low valley in diversity from Late Permian to Jurassic, and, thereafter, an increase to a stable plateau in diversity in the Mesozoicand the Cenozoic (A). Detailed data of Lingulidae genera from the Permian to the Triassic showing that the generic diversity dropped before the PTB,roughly across the Guadalupian–Lopingian Boundary (B).
123Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
lingulids to force their way into substrates whenburrowing (Emig, 1997). In this context, it is interestingto note that the earliest Triassic Lingulids demonstratethe most bladed anterior shape of all lingulids. Thisprobably implies that the lingulids living in theaftermath of the end-Permian extinction were subjectedto a particularly high environmental stress so that inorder to survive, they had to transform their shape toenable them to burrow quicker and more efficiently.
4. Lingulidae's ecological advantages for surviving
Early Triassic Lingulidae were clearly survivors fromtheir Late Permian predecessors and became dominantin the earliest Triassic ecological niches vacated by theextinction of other organisms, forming a disaster speciescommunity in the marine realm (Rodland and Bottjer,2001). It has also been recognized that the formation oflingulid-dominated fossil beds generally occurredduring drastic to catastrophic ecological changesbecause, in a normal situation, complete lingulid shellsare usually not easily preserved (Emig, 1997). Thus, theunusually high abundance of the Lingulidae in some
Early Triassic deposits and their wide distributionthroughout the world should point to a widespread andprolonged occurrence of stressful conditions after thePTB mass extinction (Payne et al., 2004). Some pre-adapted characteristics and ecologic habits of theLingulidae are thought to be advantageous for them toadapt to catastrophic ecological changes, and thussurvive the PTB mass extinction events. These, aselaborated further below, include their shell composi-tions of organophosphates, their small-sized shells,burrowing living styles, and wide ecological adaptationduring the disaster periods.
4.1. Shell composition
The shells of both fossil lingulids and their extantrepresentatives are composed of organophosphates. Theshell structure of extant lingulids includes two distinctlayers (i.e. inorganic and organic parts), both of whichare mainly composed of four constituents: apatite(fluorapatite francolite); glycosaminoglycans (GAGs);fibrillar collagens; and chitin associated with protein(Williams et al., 1994). The shells of fossil lingulids are
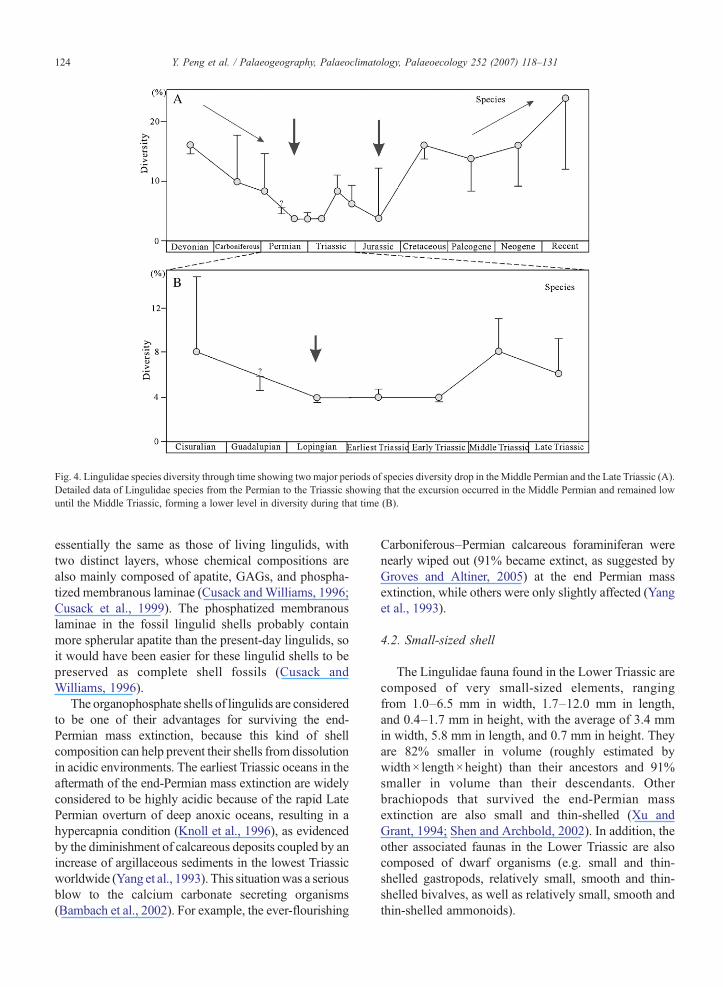
Fig. 4. Lingulidae species diversity through time showing two major periods of species diversity drop in the Middle Permian and the Late Triassic (A).Detailed data of Lingulidae species from the Permian to the Triassic showing that the excursion occurred in the Middle Permian and remained lowuntil the Middle Triassic, forming a lower level in diversity during that time (B).
124 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
essentially the same as those of living lingulids, withtwo distinct layers, whose chemical compositions arealso mainly composed of apatite, GAGs, and phospha-tized membranous laminae (Cusack andWilliams, 1996;Cusack et al., 1999). The phosphatized membranouslaminae in the fossil lingulid shells probably containmore spherular apatite than the present-day lingulids, soit would have been easier for these lingulid shells to bepreserved as complete shell fossils (Cusack andWilliams, 1996).
The organophosphate shells of lingulids are consideredto be one of their advantages for surviving the end-Permian mass extinction, because this kind of shellcomposition can help prevent their shells from dissolutionin acidic environments. The earliest Triassic oceans in theaftermath of the end-Permian mass extinction are widelyconsidered to be highly acidic because of the rapid LatePermian overturn of deep anoxic oceans, resulting in ahypercapnia condition (Knoll et al., 1996), as evidencedby the diminishment of calcareous deposits coupled by anincrease of argillaceous sediments in the lowest Triassicworldwide (Yang et al., 1993). This situationwas a seriousblow to the calcium carbonate secreting organisms(Bambach et al., 2002). For example, the ever-flourishing
Carboniferous–Permian calcareous foraminiferan werenearly wiped out (91% became extinct, as suggested byGroves and Altiner, 2005) at the end Permian massextinction, while others were only slightly affected (Yanget al., 1993).
4.2. Small-sized shell
The Lingulidae fauna found in the Lower Triassic arecomposed of very small-sized elements, rangingfrom 1.0–6.5 mm in width, 1.7–12.0 mm in length,and 0.4–1.7 mm in height, with the average of 3.4 mmin width, 5.8 mm in length, and 0.7 mm in height. Theyare 82% smaller in volume (roughly estimated bywidth× length×height) than their ancestors and 91%smaller in volume than their descendants. Otherbrachiopods that survived the end-Permian massextinction are also small and thin-shelled (Xu andGrant, 1994; Shen and Archbold, 2002). In addition, theother associated faunas in the Lower Triassic are alsocomposed of dwarf organisms (e.g. small and thin-shelled gastropods, relatively small, smooth and thin-shelled bivalves, as well as relatively small, smooth andthin-shelled ammonoids).

125Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
Reducing body size could either reflect selectiveextinction of larger species or evolution of large speciestoward smaller size (Payne, 2005). At period level, thesmallest lingulid specimens do not seem to havechanged much through time (Fig. 2A, C), thoughchanges in minimum size of fossils are difficult to assessbecause smallest species are often the most poorlysampled despite constituting a significant fraction of thediversity (Jablonski, 1996; Bouchet et al., 2002).However, at epoch level, both the smallest and largestlingulid specimens and the mean of all in the earliestTriassic Griesbachian are smaller than their ancestorsand descendants (Fig. 2B, D), probably reflecting tosome extent the evolution of species toward smaller sizeduring that time.
The small body size is considered an advantageousfeature for surviving mass extinctions, as, for example,demonstrated by graptolites during the Late Silurianbiotic crises that were characterized by a massoccurrence of very small, short, and slim graptolitesimmediately after the extinction event (Urbanek, 1993).Also, the foraminifer faunas across the Cretaceous–Paleogene boundary were characterized by a markedloss of large and complex-shelled species and thesurvival of relatively small, robust, and dwarfed ones(Keller and Perch-Nielsen, 1995). The end-Permianmass extinction is also no exception. As noted byHallam and Wignall (1997), larger, complex, and thick-shelled marine organisms suffered the most loss acrossthe PTB while many of the relatively small, smooth, andthin-shelled marine organisms survived (also seeTwitchett, 1999; Price-Lloyd and Twitchett, 2001).
So, why is being small better in terms of survival instressful environments? The small-sized faunas areconsidered more advantageous for the toleration ofanoxic conditions (Levin, 2003) and having morechances to survive when the nutrient supplies areinadequate, as would be so in the Early Triassic(Twitchett, 2001, 2006). In our view, any small marineorganisms living in the aftermath of the end-Permianmass extinction would have the following advantages:(1) small bodies would consume less oxygen and foodcompared to larger species, and (2) being small wouldenable the organisms to move more effectively thanbigger species. These interpretations can be readilysubstantiated with the modern Lingulidae fauna. Forexample, the modern lingulids show an infaunal habitand possess hemerythrin within the coelomocytes as ameans to store oxygen. Both adaptation can be regardedas manifestations of their tolerance to low oxygenconditions (Cloud, 1948). The end-Permian and EarlyTriassic sediments are considered by many to have
formed under anoxic environments (Wignall andHallam, 1992; Grice et al., 2005), as evidenced by theappearance of finely laminated black lithologies, largeamounts of framboidal pyrites preserved in the rocks,and a shortage of benthos coupled with a highabundance of lower organisms (i.e. fungi and/oralgae). This anoxic environment meant that those faunasunsuitable to dysaeroxic conditions would eventuallysuccumb and give way to those that had alreadydeveloped some pre-adapted features enabling them tosurvive and, in some cases, flourish in these adverseenvironments. Among these organisms that had pre-adapted features to anoxic environments, the Lingulidaeare probably most notable. The hemerythrin in extantLingula has an oxygen equilibrium that is reversiblyaltered by pH changes (Manwell, 1960), which meansthat Lingula can store more oxygen when environmentstend to be acidic, as would have been the case in the seaduring the PTB period (Knoll et al., 1996). As such,lingulids had advantages to obtain and store moreoxygen than other organisms in the acidic and anoxicenvironments during the PTB mass extinction period.
It is considered that small-bodied organisms have atrue advantage in dysoxic conditions, because smallbodies would have a larger surface area/volume ratio,and reduced diffusion distances. These are also thefeatures considered by marine biologists to explain thesuccess of small, thin, elongate taxa thriving within theoxygen minimum zones of modern oceans (Levin,2003). Small-bodied organisms may also have greatermetabolic flexibility that confers ability to use foodresources in the absence of oxygen (Zehnder, 1988).McClain and Rex (2001) have observed a positiverelationship between maximum body size and oxygenconcentration for nine species of turid gastropods fromthe western North Atlantic, and within a single species,Benthomangelia antonia Dall 1881. If we use this studyas an analogue, it is plausible that small-sized lingulidsand other Early Triassic dwarf taxa would have used ormaintained the same advantage in the aftermath of theend-Paleozoic mass extinction as the modern turidgastropods do in coping with low oxygen environments.
4.3. Burrowing lifestyle
Lingulids have retained their burrowing (infaunal)lifestyle in sediments since they first appeared ingeological history (Craig, 1952; Emig, 1997). Bothfossil and extant lingulids have a range of morpholog-ical, physiological and behavioral features for them toadapt to a burrowing mode of life that has remainedremarkably stable at least since the early Paleozoic

126 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
(Emig, 1997). These characteristics include an oblongoval or rectangular shell shape with straight subparallelto parallel lateral margins and a straight to slightlyconcave anterior margin for burrowing, and a pedicle toanchor to the bottom of burrows and to shift the positionof shells (Emig, 2003). Fossil burrows with lingulidshells in situ found in the Triassic of Vosges Mountainsshow the same burrowing structures as those of extantlingulids (Emig et al., 1978; Emig, 1997). Many fossillingulids are also found to be preserved in situ, lyingperpendicular to bedding, although the structures oftheir burrows are usually not preserved (Craig, 1952;Cherns, 1979).
The infaunal living style is likely to make lingulidsregularly experience low oxygen tension in thesurrounding water (Emig, 1997). Besides havingrelatively lower rates of oxygen consumption (seeabove), lingulids also have strong abilities to removeoxygen from water at low oxygen tension (Peck et al.,1997). In addition, lingulids might have some specialrespiratory pigments to transport or to store oxygenwithin their body tissues (Peck et al., 1997). Thus, allthese advantages supported Lingulidae to struggle in theanoxic environments during the Permian–Triassictransitional period.
In addition, taxa of protected or buffered habitatsusually have higher survival potential, because they arenot as greatly perturbed by the forcing mechanisms ofmass extinctions (Harries et al., 1996). Another goodexample for cave dwellers to survive the PTB massextinction events is demonstrated by the infauna(burrowers) of two important Permian bivalve families,
Fig. 5. Global distributions of the Early Triassic Lingulidae fauna. Base map oMcKerrow (1990). 1—Northwestern USA (Newell and Kummel, 1942; RoSouthern Alps of Italy (Assereto et al., 1973; Broglio Loriga et al., 1980); 4—WWest Pakistan (Rowell, 1970); 7—Western Australia (Dickins andMcTavish,(Bando, 1964); 10—Siberia, Russia (Dagys, 1965).
Nuculidae and Myophoriidae, represented respectivelyby Palaeonucula and Neoschizodus. Both range up intothe Triassic without clear changes in diversity (Yin inYang et al., 1993). In both cases, Yin (in Yang et al.,1993) suggested that the infaunas usually evolved muchslower than the epifaunas, and had higher stability tosurvive because of their infield ecotopes. However,other data, such as that from the PTB trace fossils(Twitchett, 1999; Twitchett and Barras, 2004), indicatethat clearly infaunal burrowers were also affected by theend-Permian mass extinction event. Thus, infaunallifestyle alone is not necessarily a pre-requisite forsurvival.
4.4. Wide ecological adaptation
The distribution pattern of extant lingulids (Lingulaand Glottidia) shows that they both have clear depth andlatitude limits (Craig, 1952; Emig, 1997). They occurlatitudinally within the 400 belt from temperate toequatorial areas (Emig, 1997), and their bathymetricdistributions are restricted to the continental shelf watersexcept for Glottidia albida, which extends into theupper part of the bathyal slope (477 m in deepestrecords) (Emig, 1997). This phenomenon is completelydifferent from their fossil counterparts.
Unlike their modern counterparts, the Early Triassiclingulids (and indeed fossil lingulids from othergeological periods) have been found almost globally,from high-latitude, polar, cool-temperate environmentsto low-latitude tropical and subtropical warm-watersettings (Fig. 5, and references herein). This would
f the Early Triassic reconstruction of the continents is after Scotese anddland and Bottjer, 2001); 2—Eastern Greenland (Spath, 1930); 3—estern Hungry (Hips, 1996); 5—Iran (Broglio Loriga et al., 1980); 6—
1963; Gorter, 1978; Tasch and Jones, 1979); 8—South China; 9—Japan

127Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
suggest that temperature probably did not play a majorrole in controlling the geographical and biogeographicaldistribution of the Early Triassic lingulids. Likewise,variations of sedimentary facies also did not seem tohave any significant control on the geographicaldistribution of Early Triassic lingulids, because EarlyTriassic lingulid species have been found in a widevariety of sediment types such as coarse sand, silt, mud,and lime. This is evidenced by data from southwesternMontana and western Wyoming, USA (Rodland andBottjer, 2001) and from South China. Additionally, ithas been suggested that lingulids are not just theindicators of marginal shallow marine environments ingeological history; they were also found in differentsediment types such as mud, silt, ironstone, limestone,and coarse sand (Graham, 1970), as well as in Paleozoicrocks believed to have formed in deep shelf and evenbasinal environments (Cherns, 1979; Pickerill et al.,1984, and references cited therein). Thus, it appears thatlingulids could regulate their osmotic equilibrium toadapt to different environments in geological history,qualifying them as one of the most geographicallywidespread generalist genera adapted to a wide varietyof environments. Thus, although the end-Permian globalregression brought about a drastic decrease of neriticareas of the oceans – a situation only to be reversed bythe earliest Triassic transgression – the lingulids werenot significantly affected by either the regression or thetransgression through the transition period between theLate Palaeozoic and Early Mesozoic.
There is also a remarkable similarity among theimmediate post-mass extinction faunas worldwidethroughout geological history, in that these faunas areinvariably characterized by low-diversity, high abun-dance, and geographically widespread eurytopic species(e.g. Erwin, 1998). In the case of the end-Permian massextinction, this remarkable phenomenon is evidentlydemonstrated by the lingulids which thrived in the EarlyTriassic. Taphonomic study of known, extant lingulidssuggests that dead lingulid shells usually tend to bequickly separated, degraded, and finally completely lostfrom the sediment within 2–3 weeks after their naturaldeath in normal sedimentary environments (Emig,1997). Therefore, for large amounts of Early Triassiclingulid shells to be preserved in good to excellentconditions in the fossil record around the world,catastrophic events must be invoked as the most likelycauses. This means that the wide and abundantappearance of lingulids in the Early Triassic sedimentsshould suggest a widespread and prolonged occurrenceof stressful environments following the end-Permianmass extinction events.
5. Lingulidae's life strategies for thriving
Small evolutionary changes usually provide organ-isms with an opportunity to obtain more advantages tosurvive and thrive when facing disasters and naturalselection (Darwin, 1859). Surviving might not be sodifficult for some organisms if they have already hadsome pre-adapted characteristics for surviving, such isdiscussed above. However, the survivors would stillhave to change to adapt to stressful environments in theaftermath, or they would become extinct soon after. Forexample, though some small-sized non-lingulid bra-chiopods also survived the end-Permian mass extinc-tion, they remained rare in the earliest Triassic andbecame extinct shortly thereafter (Chen et al., 2005). Incontract, the Early Triassic Lingulidae continued theirevolutionary innovations by, for example, reducingbody size, shell thickness, and shell width/length,thickness/length, and thickness/width ratios to enablethem to continually adapt to the rapidly changing anddeteriorating environment during the end-Permian massextinction process and its aftermath in the Early Triassicperiod.
5.1. Reducing body size
As discussed above, the ability to maintain a smallbody size or even further reduce body size is one of thekey strategies for organisms to survive catastrophes,especially when food supplies are sharply reduced, aswould be the case during the end-Permian massextinction. Although lingulids were already very smallin maximum and mean size in the Lopingian – meanwidth 4.08 mm; mean length 7.64 mm; and mean height1.7 mm – they further reduced their maximum body sizeand mean size in the earliest Triassic – mean width2.87 mm; mean length 4.92 mm; and mean height0.7 mm – for better adaptation to the catastrophe. Theydid not resume their normal size range until situationswere completely recovered in the Middle Triassic(Fig. 2). The same phenomenon can be observed inother associated disaster organisms struggling in theEarly Triassic marine realm. For example, the EarlyTriassic paper-pecten Claraia species are considered tohave evolved from their Late Permian ancestors living inrelatively deep water environments (Yang et al., 2001;Gao et al., 2004). Late Permian Claraia species areusually of thick shells with strong radial and/orconcentrated ornaments (Yin, 1983, 1985; Yang et al.,2001; Gao et al., 2004; also see Claraioides in Kotlyaret al., 2004). Their descendents in the earliest Triassic,however, have thinner shells, usually have very weak or

128 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
no ornamentation on the shells, and have an averagereduction in shell size. The advantages of being thin-and relatively smooth-shelled for Claraia and otherorganisms were probably to enable them to obtain moreoxygen through shell exchange of dissolved gas underanoxic conditions and to move more effectively in thesea. Also, it might reflect a decrease in predation pres-sure in the Early Triassic aftermath when most faunasbecame extinct in the end-Permian. Similarly, ammo-noids also exhibit clear adaptive changes in theirmorphology, with Late Permian types being generallybigger with strong ornaments of ribs and nodules,in contrast to their Early Triassic counterparts (e.g.Ophiceras) that are typically small with smooth shells orweak ornaments (Yang in Yang et al., 1993). The EarlyTriassic foraminiferan assemblage is also composed ofdwarf types compared to their Late Permian ancestors(KaJenny-Deshusses, 1991; Tong in Yang et al., 1993).The size decrease of organisms (Lilliput effect) in theEarly Triassic also include the Hindeodus conodontlineage (e.g. Kozur, 1996), the fossil fish (Twitchett,2001), and the burrowing infaunas (Twitchett, 1999).
Furthermore, a reduced body size for the Lingulidaewould enable them to burrow faster and more efficiently.This is evident from the study of Emig (1997) whodemonstrated that extant Lingula with shell lengthexceeding 17 to 20 mm often fail to re-burrow. Theabnormally reduced body size in the PTB strata thusmay be considered as one of the effective, short-termadaptive changes the Lingulidae needed in order toallow them to survive in the PTB catastrophic seas.
5.2. Reducing body width/length, height/length andheight/width ratios
Animal body size and shape are usually controlledand mediated by their habitat architectures. In general, ifviewed from the mechanical principles, the slimmer (i.e.smaller width/length ratio) and flatter (i.e. smallerheight/length and height/width ratios) an object is, theeasier it is inserted into and taken out of another object,because the surface area to withstand a force is smallerwhile the intensity of pressure is greater. Thesemechanical problems are also what burrowing faunaswould face in order to better adapt to their livingenvironments. If we apply these mechanical principlesto the Early Triassic lingulids, it would help us betterunderstand why they not only had to reduce their bodysize, but they also became more sharpened and flattenedanteriorly. Clearly, possessing a sharper and flatteranterior would have allowed the Lingulidae to better fitinto their living caves, because that type of physical
form would make it much easier for them to burrow andto withdraw into, as well as to protrude out from, theircaves. In particular, as we have stated, living in cavesshould be advantageous for lingulids to survive in theanoxic environments during the Permian–Triassicperiod. Thus, making themselves more suitable toburrowing should be another short-term, temporaryliving strategy for lingulids to survive and thrive in theend-Permian and earliest Triassic catastrophes.
5.3. Reducing shell thickness
In an anoxic environment, reducing shell thicknessshould be another adaptive strategy for the organisms toget more oxygen directly and efficiently through shellgas exchange of dissolved gas (Levin, 2003). A goodexample of this is from Southern Alps where earliestTriassic Lingulidae fossils are cosmopolitan and verythin-shelled in comparison to those occurring in theoverlying strata of the Early Triassic Siusi Memberwhere lingulids were occasionally found (BroglioLoriga et al., 1980). As discussed above, the bivalveClaraia and the ammonoid Ophiceras are other goodexamples showing reduction of shell thickness as astrategy to survive and thrive after the PTB massextinction.
6. Conclusions
Viewed from their long-term evolutionary history,the Lingulidae fauna clearly possesses several pre-adapted morphological advantages which appear tohave enhanced their survival rate through the PTB massextinction. These pre-adapted advantages include anorganophosphate shell composition, small body size,burrowing habitat, and an ability to adapt to a widerange of ecological and environmental conditions.During their Early Triassic survival interval, thelingulids thrived because they further amended someof their pre-adapted morphological features by under-taking adaptive changes in response to the drasticallydeteriorated and increasingly anoxic marine environ-ment following the end-Permian mass extinction. As aresult, the lingulids in the Early Triassic aftermathfurther reduced body size to enable them to cope withfood shortage, they further modified their body shape(by reducing body width/length, height/length, andheight/width ratios) to allow them to better fit theirburrowing habitat, and they further reduced their shellthickness in order to increase the efficiency of oxygenexchange. We believe that these adaptive changes andfurther enhancements not only enabled the lingulids to

129Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
survive the end-Permian mass extinction, but also gavethem the opportunity to thrive in prolonged EarlyTriassic stressful environments in the marine realm.
Acknowledgement
Profs. Fengqing Yang and Zhiping Wang (ChinaUniversity of Geosciences) and Dr. Wenzhong Li(Nanjing Institute of Geology and Palaeontology) areacknowledged for their help to collect the Lingulidaesamples. The authors are grateful to Dr. Mike Orchard(Geological Survey of Canada), Dr. Richard Twitchett(University of Plymouth, UK) and two anonymousreviewers for their critical and constructive reviews,comments, and assistance with English. This work is apart of the research programs supported by the NationalScience Foundation of China (grant nos. 40225005,40232025, 40321202, 40325004 and 40328003). Thispaper has also been supported in part by a research grantfrom the Australian Research Council (LX 0348047 toG. R. Shi). Yuanqiao Peng and Yongqun Gao acknowl-edge support of the Australian Commonwealth Govern-ment, and Deakin University for the award of anInternational Postgraduate Research Scholarship (IPRS).
Appendix A. Supplementary data
Supplementary data associated with this article canbe found, in the online version, at doi:10.1016/j.palaeo.2006.11.039.
References
Assereto, R., Bosellini, A., Fantini, S.N., Sweet, W.C., 1973. ThePermian–Triassic boundary in the Southern Alps (Italy). In:Logan, A., Hills, L.V. (Eds.), The Permian and Triassic Systemsand their Mutual Boundary. Memoirs, vol. 2. Canadian Society ofPetroleum Geologists, Calgary, pp. 176–199.
Bambach, R.K., Knoll, A.H., Sepkoski, J.J., 2002. Anatomical andecological constraints on Phanerozoic animal diversity in themarine realm. Proceedings of the National Academy of Sciences ofthe United States of America 99, 6854–6859.
Bando, Y., 1964. On some Lower and Middle Triassic ammonoidsfrom Japan. Transactions and Proceedings of the PalaeontologicalSociety of Japan 56, 332–345.
Biernat, G., Emig, C.C., 1993. Anatomical distinctions of theMesozoiclingulide brachiopods. Acta Palaeontologica Polonica 38, 1–20.
Bouchet, P., Lozouet, P., Maestrati, P., Heros, V., 2002. Assessing themagnitude of species richness in tropical marine environments:exceptionally high numbers of molluscs at a New Caledonia site.Biological Journal of the Linnean Society 75, 421–436.
Broglio Loriga, C., Neri, C., Posenato, R., 1980. The “Lingula zone”of Scythian (Lower Triassic) stratigraphy and palaeoecology.Annali dell'Università di Ferrara (new series). Scienze Geologichee Paleontologiche 6, 91–130.
Chen, Z.Q., Kaiho, K., George, A.D., 2005. Survival strategies ofbrachiopod faunas from the end-Permian mass extinction.Palaeogeography, Palaeoclimatology, Palaeoecology 224,232–269.
Cherns, L., 1979. The environmental significance of Lingula in theLudlow Series of the Welsh Borderland and Wales. Lethaia 12,35–46.
Cloud, P.E., 1948. Notes on recent brachiopods. American Journal ofScience 246, 241–250.
Cooper, S.A., 1942. New genera of North American brachiopods.Washington Academy of Sciences Journal 32, 228–235.
Craig, Y., 1952. A comparative study of the ecology and palaeoecol-ogy of Lingula. Transactions of Edinburgh Geological Society 15,110–120.
Cusack, M., Williams, A., 1996. Chemico-structural degradation ofCarboniferous lingulid shells. Philosophical Transactions of theRoyal Society of London. Series B 351, 33–49.
Cusack, M., Williams, A., Buckman, J.O., 1999. Chemico-structuralevolution of linguloid brachiopod shells. Palaeontology 42,799–840.
Dagys, A.S., 1965. Triasovye Brakhiopodi Sibiri. Akademiia NaukUSSR, Sibirskoe Otdelenie, Institut Geologii I Geofiziki, Moscow.186 pp. (in Russian).
Darwin, C., 1859. On the Origin of Species by Means of NaturalSelection. John Murray, London. 490 pp.
Dickins, J.M., McTavish, R.A., 1963. Lower Triassic marine fossilsfrom the Beagle Ridge (BMR 10) Bore, Perth basin, westernAustralia. Journal of the Geological Society of Australia 10,123–140.
Emig, C.C., 1982. Taxonomie du genre Lingula (BrachiopodesInarticulés). Bulletin du Muséum National d'Histoire Naturellede Paris 4, 337–367 (in French).
Emig, C.C., 1983. Taxonomie du genre Glottidia (BrachiopodesInarticulés). Bulletin du Muséum National d'Histoire Naturelle deParis 5, 469–489 (in French).
Emig, C.C., 1997. Ecology of inarticulated brachiopods. In: Kaesler,R.L. (Ed.), Treatise on Invertebrate Paleontology, Part H,Brachiopoda. The Geological Society of America, pp. 473–495(revised).
Emig, C.C., 2003. Proof that Lingula (Brachiopoda) is not a living-fossil, and emended diagnoses of the family Lingulids. Notebookson Geology-Letter 2003/01. (CG2003_L01_CCE).
Emig, C.C., Herrera, Z., 2006. Dignomia munsterii (Brachiopoda,Lingulata) from the Ordovician of Bolivia, with redescription ofthe genus. Geodiversitas 28, 227–237.
Emig, C.C., Gall, J.C., Pajaud, D., Plaziat, J.C., 1978. Réflexionscritiques sur l'écologie et la systématique des lingules actuelles etfossiles. Géobios 11, 573–609 (in French).
Erwin, D.H., 1993. The Great Paleozoic Crisis: Life and Death in thePermian. Columbia University Press, New York. 327 pp.
Erwin, D.H., 1994. The Permo-Triassic extinction. Nature 367,231–236.
Erwin, D.H., 1998. The end and the beginning: recoveries from massextinction. Trends in Ecology and Evolution 13, 344–349.
Flugel, E., 1994. Pangean shelf carbonates: controls and paleoclimatesignificance of the Permian and Triassic reefs. In: Klein, G.V.(Ed.), Pangea: Paleoclimate, Tectonics and Sedimentation duringAccretion, Zenith and Breakup of a Supercontinent. GeologicalSociety of America Special Paper, vol. 288, pp. 247–266.
Fraiser, M.L., Bottjer, D.J., 2004. The non-actualistic Early Triassicgastropod fauna: a case study of the Lower Triassic SinbadLimestone Member. Palaios 19, 259–275.

130 Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
Gao, Y.Q., Yang, F.Q., Peng, Y.Q., 2004. Significance of Claraia fromthe Late Permian of South Guizhou, China. Alcheringa 28,469–476.
Gorter, J.D., 1978. Triassic environments in the Canning Basin,western Australia. Journal of Australian Geology and Geophysics3, 25–33.
Graham, D.K., 1970. Scottish Carboniferous Lingulacea. Bulletin ofGeological Survey of Great Britian 31, 139–184.
Grice, K., Cao, C.Q., Love, G.D., Böttcher, M.E., Twitchett, R.J.,Grosjean, E., Summons, R.E., Turgeon, S.C., Dunning,W., Jin, Y.G.,2005. Photic zone euxinia during the Permian–Triassic superanoxicevent. Science 307, 706–709.
Groves, J.R., Altiner, D., 2005. Survival and recovery of calcareousforaminifera pursuant to the end-Permian mass extinction.Comptes Rendus Palevol 4, 419–432.
Hallam, A., Wignall, P.B., 1997. Mass Extinction and Their Aftermath.Oxford University Press, Oxford. 320 pp.
Harries, P.J., Kauffman, E.G., Hansen, T.A., 1996. Models for bioticsurvival following mass extinction. In: Hart, M.B. (Ed.), BioticRecovery from Mass Extinction Events. Geological SocietySpecial Publication, vol. 102, pp. 41–60.
Hips, K., 1996. Stratigraphic and facies evaluation of the LowerTriassic formations in the Aggtelek-Rudabánya Mountains, NEHungary. Acta Geologica Hungarica 39, 369–411.
Holmer, L.E., Popov, L.E., 2000. Lingulidae. In: Kaesler, R.L (Ed.),Treatise on Invertebrate Paleontology, Part H, Brachiopoda(revised). The Geological Society of America and The Universityof Kansas, pp. 35–39.
Hone, D.W.E., Benton, M.J., 2005. The evolution of large size: howdoes Cope's Rule work? Trends in Ecology and Evolution 20, 4–6.
Jablonski, D., 1996. Body size and macroevolution. In: Erwin, D.H.,Jablonski, D., Lipps, J.H. (Eds.), Evolutionary Paleobiology.University of Chicago Press, Chicago, pp. 256–289.
Jablonski, D., 1997. Body-size evolution in Cretaceous mollusks andthe status of Cope's rule. Nature 385, 250–252.
KaJenny-Deshusses, C., 1991. The Permian–Triassic of the Gartner-kofel-1 Core (Carnic Alps, Austria): foraminifera and algae of theCore and the outcrop section. Abhandlungen der GeologischenBundesanstalt 45, 99–108.
Keller, G., Perch-Nielsen, K., 1995. Cretaceous–Tertiary (K/T) massextinction: effect of global change on calcareous microplankton.In: Board on Earth Sciences and Resources, Commission onGeosciences, Environment, and Resources, National ResearchCouncil (Eds.), Effects of Past Global Change on Life. NationalAcademy Press, Washington D. C., 72–93.
Knoll, A.H., Bambach, R.K., Canfield, D.E., Grotzinger, J.P., 1996.Comparative earth history and Late Permian mass extinction.Science 273, 452–457.
Kotlyar, G.V., Zakharov, Y.D., Polubotko, I.V., 2004. Late Changh-singian fauna of the northwestern Caucasus Mountains, Russia.Journal of Paleontology 78, 513–527.
Kozur, H., 1996. The conodonts Hindeodus, Isarcicella and Sweeto-hindeodus in the uppermost Permian and lowermost Triassic.Geologica Croatica 49, 81–115.
Levin, L.A., 2003. Oxygen minimum zone benthos: adaptation andcommunity response to hypoxia. Oceanography and MarineBiology: an Annual Review 41, 1–45.
Manwell, C.P., 1960. Comparative physiology: blood pigments.Annual Review of Physiology 22, 191–244.
McClain, C.R., Rex, M.A., 2001. The relationship between dissolvedoxygen concentration andmaximumsize in deep-sea turrid gastropods:an application of quantile regression. Marine Biology 13, 681–685.
Mendes, J.C., 1961. Langella, novo gênero de lingulídeo da SérieTubarão. Geologia 5, 1–8 (in Portuguese).
Newell, N.D., Kummel, B., 1942. Lower Eo-Triassic stratigraphy,western Wyoming and southeast Idaho. Geological Society ofAmerica Bulletin 53, 937–995.
Oliveira, E.P., 1930. Fósseis marinhos na série Itararé do Estado de SantaCatarina. Anais da Academia Brasileira de Ciências 2, 17–21.
Payne, J.L., 2005. Evolutionary dynamics of gastropod size across theend-Permian extinction and through the Triassic recovery interval.Paleobiology 31, 269–290.
Payne, J.L., Lehrmann, D.J., Wei, J.Y., Orchard, M.J., Schrag, D.P.,Knoll, A.H., 2004. Large perturbations of the carbon cycle duringrecovery from the end-Permian extinction. Science 305, 506–509.
Peck, L.S., Rhodes, M.C., Curry, G.B., Ansell, A.D., 1997.Physiology. In: Kaesler, R.L. (Ed.), Treatise on invertebratepaleontology, Part H, Brachiopoda. The Geological Society ofAmerica, pp. 213–242 (revised).
Peng, Y.Q., Shi, G.R., submitted for publication. New Early TriassicLingulidae (Brachiopoda) genera and species from South China.Alcheringa.
Pickerill, R.K., Harland, T.L., Fillion, D., 1984. In situ lingulids fromdeep-water carbonates of the Middle Ordovician Table HeadGroup of Newfoundland and the Trenton Group of Quebec.Canadian Journal of Earth Sciences 21, 194–199.
Price-Lloyd, N., Twitchett, R.J., 2001. The Lilliput effect in theaftermath of the end-Permian mass extinction event. GSAAbstracts with Programs 34 (6), 355.
Retallack, G.J., Veevers, J.J., Morante, R., 1996. Global coal gapbetween Permian–Triassic extinction and middle Triassic recoveryof peat forming plants. Geological Society of America Bulletin108, 195–207.
Rodland, D.L., Bottjer, D.J., 2001. Biotic recovery from the end-Permian mass extinction: behavior of the inarticulate brachiopodLingula as a disaster taxon. Palaios 16, 95–101.
Rowell, A.J., 1970. Lingula from the basal Triassic Kathwai Member,Mianwali Formation, Salt Range and Surghar Range, WestPakistan. In: Kummel, B., Teichert, C. (Eds.), StratigraphicBoundary Problems: Permian and Triassic of West Pakistan. TheUniversity Press of Kansas, Lawrence, pp. 111–116.
Scotese, C.R., McKerrow, W.S., 1990. Revised world maps andintroduction. In: McKerrow, W.S., Scotese, C.R. (Eds.), PalaeozoicPalaeogeography and Biogeography. Geological Society Memoir,vol. 12, pp. 1–21.
Shen, S.Z., Archbold, N.W., 2002. Chonetoidea (Brachiopoda) fromthe Lopingian (Late Permian) of South China. Alcheringa 25,327–349.
Shen, S.Z., Shi, G.R., 2002. Paleobiogeographical extinction patternsof Permian brachiopods in the Asian-western Pacific region.Paleobiology 28, 449–463.
Shi, G.R., Shen, S.Z., 2000. Asian-western Pacific Permianbrachiopoda in space and time: biogeography and extinctionpatterns. In: Yin, H.F., Dickins, J.M., Shi, G.R., Tong, J.N.(Eds.), Permian–Triassic Evolution of Tethys and WesternCircum-Pacific. Elsevier, Amsterdam, pp. 327–352.
Smirnova, T.N., Ushatinskaya, G.T., 2001. New lingulids (Brachio-poda) from the Lower Cretaceous of European Russia, with notesin the microstructure of their shells. Paleontologicheskii Zhurnal 4,51–59 (in Russian).
Spath, L.F., 1930. The Eotriassic invertebrate fauna of East Greenland.Meddelelser om Grøland 83, 1–90.
Tasch, P., Jones, P.J., 1979. Carboniferous, Permian and Triassicconchostracans of Australia, three new studies. Bulletin of the

131Y. Peng et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 252 (2007) 118–131
Bureau of Mineral Resources, Geology and Geophysics, Australia185, 1–47.
Twitchett, R.J., 1999. Palaeoenvironments and faunal recovery afterthe end-Permian mass extinction. Palaeogeography, Palaeoclima-tology, Palaeoecology 154, 27–37.
Twitchett, R.J., 2001. Incompleteness of the Permian–Triassic fossilrecord: a consequence of productivity decline? Geological Journal36, 341–353.
Twitchett, R.J., 2006. The palaeoclimatology, palaeoecology andpalaeoenvironmental analysis of mass extinction events. Palaeo-geography, Palaeoclimatology, Palaeoecology 232, 190–213.
Twitchett, R.J., Barras, C.G., 2004. Trace fossils in the aftermathof mass extinction events. In: McIlroy, D. (Ed.), The Applicationof Ichnology to Palaeoenvironmental and StratigraphicAnalysis. Geological Society, London, Special Publications,vol. 228, pp. 397–418.
Urbanek, A., 1993. Biotic crises in the history of Upper Siluriangraptoloids: a palaeobiological model. Historical Biology 7, 29–50.
Wignall, P.B., Hallam, A., 1992. Anoxia as a cause of the Permian/Triassic mass extinction: facies evidence from northern Italy andthe western United States. Palaeogeography, Palaeoclimatology,Palaeoecology 93, 21–46.
Williams, A.J., 1977. Insight into lingulid evolution from the LateDevonian. Alcheringa 1, 401–406.
Williams, A., Cusack, M., Mackay, S., 1994. Collagenous chitinopho-sphatic shell of the brachiopod Lingula. Philosophical Transactionsof the Royal Society of London. Series B 346, 223–266.
Xu, G.R., Grant, R.E., 1994. Brachiopods Near the Permian–TriassicBoundary in South China. Smithsonian Institution Press, Washing-ton D.C. 68 pp.
Yang, Z.Y., Wu, S.B., Yin, H.F., Xu, G.R., Zhang, K.X., Bi, X.M.,1993. Permo-Triassic Events of South China. Geological Publish-ing House, Beijing. 153 pp.
Yang, F.Q., Peng, Y.Q., Gao, Y.Q., 2001. Study on Late PermainClaraia in South China. Science in China, Series D 44, 797–807.
Yegorov, A.N., Popov, L.Ye, 1990. A new Lower Permian lingulidfrom the Siberian Platform. Paleontologicheskii Zhurnal 4,111–115 (in Russian).
Yin, H.F., 1983. Uppermost Permian (Changxingian) pectinacea fromSouth China. Revista Italiana di Paleontologia 88, 337–386.
Yin, H.F., 1985. Bivalves near the Permian–Triassic boundary inSouth China. Journal of Paleontology 59, 572–600.
Zehnder, A., 1988. Biology of Anaerobic Organisms. John Wiley,New York. 872 pp.
Related Documents











