ORIGINAL ARTICLE doi:10.1111/evo.12270 HOST-SPECIALIST LINEAGES DOMINATE THE ADAPTIVE RADIATION OF REEF CORAL ENDOSYMBIONTS Daniel J. Thornhill, 1 Allison M. Lewis, 2 Drew C. Wham, 2 and Todd C. LaJeunesse 2,3 1 Department of Conservation Science and Policy, Defenders of Wildlife, 1130 17th Street NW, Washington, DC 20007 2 Department of Biology, Pennsylvania State University, 208 Mueller Laboratory, University Park, PA 16802 3 E-mail: [email protected] Received April 8, 2013 Accepted September 4, 2013 Data Archived: Dryad doi: 10.5061/dryad.2247c Bursts in species diversification are well documented among animals and plants, yet few studies have assessed recent adaptive radiations of eukaryotic microbes. Consequently, we examined the radiation of the most ecologically dominant group of endosym- biotic dinoflagellates found in reef-building corals, Symbiodinium Clade C, using nuclear ribosomal (ITS2), chloroplast (psbA ncr ), and multilocus microsatellite genotyping. Through a hierarchical analysis of high-resolution genetic data, we assessed whether ecologically distinct Symbiodinium, differentiated by seemingly equivocal rDNA sequence differences, are independent species lin- eages. We also considered the role of host specificity in Symbiodinium speciation and the correspondence between endosymbiont diversification and Caribbean paleo-history. According to phylogenetic, biological, and ecological species concepts, Symbiodinium Clade C comprises many distinct species. Although regional factors contributed to population-genetic structuring of these lineages, Symbiodinium diversification was mainly driven by host specialization. By combining patterns of the endosymbiont’s host speci- ficity, water depth distribution, and phylogeography with paleo-historical signals of climate change, we inferred that present-day species diversity on Atlantic coral reefs stemmed mostly from a post-Miocene adaptive radiation. Host-generalist progenitors spread, specialized, and diversified during the ensuing epochs of prolonged global cooling and change in reef-faunal assemblages. Our evolutionary reconstruction thus suggests that Symbiodinium undergoes “boom and bust” phases in diversification and extinction during major climate shifts. KEY WORDS: Climate change, coral symbiosis, dinoflagellate, ecological specialization, phylogeography, speciation, Symbio- dinium. Adaptive radiations occur when lineages diversify in response to a variety of ecological opportunities (Gavrilets and Losos 2009; Glor 2010). These bursts of diversification occurred repeatedly at different taxonomic, spatial, and temporal scales throughout the history of life. Iconic examples of adaptive radiations include the Cambrian explosion (Valentine et al. 1999), the Cenozoic diversi- fication of therian mammals (Janis 1993; Agadjanian 2003), and more recent radiations of Darwin’s finches of the Gal´ apagos is- lands (Grant and Grant 2007), African cichlids (Seehausen 2006), Caribbean island Anolis lizards (Losos 2011), and Hawaiian sil- verswords (Baldwin et al. 1991). By comparison, relatively little is known about the adaptive radiations of microorganisms, including marine protists (see Falkowski et al. 2004). Their size, impover- ished geologic record, and difficulty in acquiring uncontaminated specimens for genetic analysis have limited our understanding of the tempo and mode of their diversification. Indications of the nature of adaptive radiations in eukary- otic microbes can be derived from the major groups of mod- ern eukaryotic phytoplankton—the dinoflagellates, diatoms, and coccolithophorids—following the mass extinction of marine life 352 C 2013 The Author(s). Evolution C 2013 The Society for the Study of Evolution. Evolution 68-2: 352–367

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
doi:10.1111/evo.12270
HOST-SPECIALIST LINEAGES DOMINATETHE ADAPTIVE RADIATION OF REEF CORALENDOSYMBIONTSDaniel J. Thornhill,1 Allison M. Lewis,2 Drew C. Wham,2 and Todd C. LaJeunesse2,3
1Department of Conservation Science and Policy, Defenders of Wildlife, 1130 17th Street NW, Washington, DC 200072Department of Biology, Pennsylvania State University, 208 Mueller Laboratory, University Park, PA 16802
3E-mail: [email protected]
Received April 8, 2013
Accepted September 4, 2013
Data Archived: Dryad doi: 10.5061/dryad.2247c
Bursts in species diversification are well documented among animals and plants, yet few studies have assessed recent adaptive
radiations of eukaryotic microbes. Consequently, we examined the radiation of the most ecologically dominant group of endosym-
biotic dinoflagellates found in reef-building corals, Symbiodinium Clade C, using nuclear ribosomal (ITS2), chloroplast (psbAncr),
and multilocus microsatellite genotyping. Through a hierarchical analysis of high-resolution genetic data, we assessed whether
ecologically distinct Symbiodinium, differentiated by seemingly equivocal rDNA sequence differences, are independent species lin-
eages. We also considered the role of host specificity in Symbiodinium speciation and the correspondence between endosymbiont
diversification and Caribbean paleo-history. According to phylogenetic, biological, and ecological species concepts, Symbiodinium
Clade C comprises many distinct species. Although regional factors contributed to population-genetic structuring of these lineages,
Symbiodinium diversification was mainly driven by host specialization. By combining patterns of the endosymbiont’s host speci-
ficity, water depth distribution, and phylogeography with paleo-historical signals of climate change, we inferred that present-day
species diversity on Atlantic coral reefs stemmed mostly from a post-Miocene adaptive radiation. Host-generalist progenitors
spread, specialized, and diversified during the ensuing epochs of prolonged global cooling and change in reef-faunal assemblages.
Our evolutionary reconstruction thus suggests that Symbiodinium undergoes “boom and bust” phases in diversification and
extinction during major climate shifts.
KEY WORDS: Climate change, coral symbiosis, dinoflagellate, ecological specialization, phylogeography, speciation, Symbio-
dinium.
Adaptive radiations occur when lineages diversify in response to
a variety of ecological opportunities (Gavrilets and Losos 2009;
Glor 2010). These bursts of diversification occurred repeatedly at
different taxonomic, spatial, and temporal scales throughout the
history of life. Iconic examples of adaptive radiations include the
Cambrian explosion (Valentine et al. 1999), the Cenozoic diversi-
fication of therian mammals (Janis 1993; Agadjanian 2003), and
more recent radiations of Darwin’s finches of the Galapagos is-
lands (Grant and Grant 2007), African cichlids (Seehausen 2006),
Caribbean island Anolis lizards (Losos 2011), and Hawaiian sil-
verswords (Baldwin et al. 1991). By comparison, relatively little is
known about the adaptive radiations of microorganisms, including
marine protists (see Falkowski et al. 2004). Their size, impover-
ished geologic record, and difficulty in acquiring uncontaminated
specimens for genetic analysis have limited our understanding of
the tempo and mode of their diversification.
Indications of the nature of adaptive radiations in eukary-
otic microbes can be derived from the major groups of mod-
ern eukaryotic phytoplankton—the dinoflagellates, diatoms, and
coccolithophorids—following the mass extinction of marine life
3 5 2C© 2013 The Author(s). Evolution C© 2013 The Society for the Study of Evolution.Evolution 68-2: 352–367

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
during the end-Permian (Falkowski et al. 2004). This diversifi-
cation was likely facilitated by the vacancy of many niches, the
endosymbiotic acquisition of red algal plastids by previously het-
erotrophic lineages, and the competitive superiority of these phy-
toplankton in the low-nitrogen, low-trace-element oceans of the
Mesozoic (Falkowski et al. 2004). This example suggests that eco-
logical opportunities, including symbiotic interactions and major
changes in climate and ocean chemistry, drove adaptive radiations
(and extinctions) in unicellular eukaryotic life.
Symbiodinium spp. dinoflagellates, colloquially known as
zooxanthellae, are important microbes to coral–reef ecosys-
tems because their photosynthesis enhances animal calcifica-
tion and supports animal metabolism through the supply of or-
ganic nutrients (Muscatine et al. 1981; Barnes and Chalker 1990;
Colombo-Pallotta et al. 2010). The genus Symbiodinium com-
prises many phylogenetically divergent Clades1 and numerous
subcladal “types” (reviewed in Coffroth and Santos 2005). Anal-
yses of their diversity using nuclear ribosomal DNA, chloroplast
DNA, as well as microsatellites and their flanking sequences
revealed complex phylogeographic patterns and differences in
host specificity among subclade lineages (e.g., LaJeunesse et al.
2004, 2010; Finney et al. 2010; Silverstein et al. 2011; Thornhill
et al. 2013). Many past investigations used the internal transcribed
spacer regions (ITS1 and ITS2) to resolve putative Symbiodinium
species (e.g., LaJeunesse 2001; Sampayo et al. 2009; LaJeunesse
et al. 2010). Equating species designations to ITS “types” has
been met with skepticism (Correa and Baker 2009); however, evi-
dence from additional genetic markers affirms that closely related
ITS lineages are independent evolutionarily units (Sampayo et al.
2009; LaJeunesse et al. 2010; LaJeunesse and Thornhill 2011;
Pochon et al. 2012). Following this logic, some species lineages
were described formally with an emphasis on concordant data
from multiple DNA loci, complemented by ecological, physio-
logical, and morphological information (LaJeunesse et al. 2012).
Genetic evidence suggests that present day diversity in sev-
eral Symbiodinium Clades originates from adaptive radiations
during recent geological epochs (LaJeunesse 2005; Pochon et
al. 2006). One hypothesis posited that allopatry, major changes
in climate, and, most importantly, host specialization drove the
tempo and mode of Symbiodinium diversification (LaJeunesse
2005). To explain the dominance of Clade C, LaJeunesse (2005)
proposed that adaptive radiations occurred in this group as the
Atlantic and Pacific Ocean basins separated (∼4–6 Mya) with the
shoaling and eventual closure of the Central American Seaway
(Chaisson and Ravelo 2000; Haug et al. 2001). These major ge-
1Note that throughout the text, the capitalized term “Clade” will be used when
referring to one of the nine recognized subgroupings of Symbiodinium (i.e.,
Clades A–I), whereas the lower case term “clade” will be used in the typical
context to refer to an ancestor and all of its descendants.
ological changes coincided with the onset of global cooling that
eventually led to periodic cycles of northern hemisphere glaciation
(2–3 Mya; Haq et al. 1987; Ruddiman and Raymo 1988; Beerling
et al. 2002). This hypothesis suggested that hundreds of Symbio-
dinium lineages have evolved since the Miocene–Pliocene transi-
tion. Phylogenetic patterns based on ITS2 nrDNA indicated that
during this climatic upheaval, generalist Symbiodinium spp. ca-
pable of associating with many host taxa became widespread
(LaJeunesse 2005). Over time these lineages putatively speciated,
as numerous populations evolved habitat specificity for particu-
lar host taxa (LaJeunesse 2005). This model of evolution implies
that large numbers of Symbiodinium spp. are lost and gained dur-
ing and after major climate shifts. Furthermore, speciation and
extinction events appear to be considerably more dynamic in en-
dosymbiont lineages compared to their hosts.
To further investigate these initial inferences about Sym-
biodinium spp. diversification, we examined the phylogenetic
relationships of Symbiodinium Clade C from the Atlantic.
Although very low-density populations of “background,” cryptic,
or ephemeral types can exist (Mieog et al. 2007; LaJeunesse
et al. 2009), most colonies of symbiotic Cnidaria harbor a
dominant Symbiodinium whose in hospite population comprises
a single clonal genotype that may persist for years or more
(Goulet and Coffroth 2003; Thornhill et al. 2006, 2009, 2013;
Andras et al. 2011; Pettay et al. 2011). Therefore, these stable and
physiologically important symbionts are regarded as ecologically
and evolutionarily significant to the host. To examine the diversity
of these Symbiodinium and their evolutionary origins, we com-
bined ITS2 genotyping, partial sequences of the psbA minicircle
noncoding region (psbAncr), and microsatellite population genetic
markers (Moore et al. 2003; Babcock et al. 2006; LaJeunesse and
Thornhill 2011; Wham et al. 2013). Each of these approaches
differs in its resolution of evolutionary relationships among
Symbiodinium. Nuclear ribosomal DNA, for example, provides
coarse phylogenetic resolution and enables comparisons across
the entire genus (Rowan and Powers 1991; LaJeunesse 2001).
By contrast, the psbA minicircle noncoding region, which is
vertically inherited as one element of the unusual choloroplast
genomes of peridinin-containing dinoflagellates, allows for
high-resolution comparisons within Clades and among closely
related Symbiodinium lineages (Barbrook et al. 2006; LaJeunesse
and Thornhill 2011). Microsatellites similarly provide fine-scale
resolution of populations and strains within closely related Sym-
biodinium species, facilitating studies on dispersal, gene flow,
and allele exchange (e.g., Andras et al. 2011; Pettay et al. 2011;
Thornhill et al. 2013). Our objectives were to examine whether (1)
small albeit fixed sequence differences in rapidly evolving rDNA
(i.e., ITS2) approximate species lineages; (2) ecological special-
ization to different kinds of host led to species evolution; and (3)
high-resolution phylogeographic evidence supports the
EVOLUTION FEBRUARY 2014 3 5 3

DANIEL J. THORNHILL ET AL.
500 km
0˚
10˚S
35˚W55˚W
25˚W35˚W 15˚W
15˚N
25˚N
5˚N
10˚N
70˚W 60˚W80˚W
25˚N
20˚N 20˚N
15˚N
10˚N
70˚W80˚W90˚W
Figure 1. Map of the Caribbean and western Atlantic showing collection locations and prevailing surface currents.
hypothesis that major adaptive radiations occurred in Clade C
in response to post-Miocene climate cooling and loss in the
connectivity between the Atlantic and Pacific.
MethodsSAMPLING LOCATIONS AND METHODS
Symbiotic cnidarian samples analyzed in this study were collected
from various reef habitats in regions throughout the Caribbean
and Atlantic, including Africa (Cape Verde Island), the Bahamas
(Abaco and Lee Stocking Islands), Barbados, Belize (Carrie Bow
Cay), Brazil, Curacao, Florida (Upper Florida Keys), the Gulf
of Mexico (Flower Garden Banks), Mexico (Puerto Morelos),
Panama (San Blas), and the U.S. Virgin Islands (St. Croix; Fig. 1).
In an effort to maximize the diversity of Clade C Symbiodinium
genotypes recovered, samples were collected from a variety of
host taxa, reef habitats, and water depths with an emphasis on the
scleractinian genera Agaricia, Orbicella (formerly the Montas-
traea annularis species complex), and Siderastrea. For compari-
son, samples were also collected from two clonal cultures (rt-113
and rt-152) and select Indo-Pacific locations including Australia
(the Great Barrier Reef), Baja California, central Mexico (Ban-
deras Bay), Hawaii, and Japan (Zamami Island). Collections were
conducted using scuba equipment, with a several cm2 tissue biop-
sies taken from each coral colony using hammer and chisel, punch
core, pliers, or scissors. A supplementary table is provided that
details the host genus and species, sampling dates, source loca-
tion, and depth of collected specimens (Table S1). Differences
in water depth distributions between phylogenetic groupings of
Symbiodinium were evaluated statistically using a homoscedas-
tic two-sample T-test in StatPlus:mac LE2009 (AnalystSoft Inc.,
Vancouver, Canada).
MOLECULAR WORK
Skeletal fragments, clippings from oral discs, or concentrated
Symbiodinium pellets comprising approximately 5–10 mm2 of
preserved tissue were mechanically disrupted using a Biospec
Beadbeater (Bartlesville, OK) for 1 min at maximum speed. Nu-
cleic acids were then extracted using a modified Promega Wizard
DNA extraction protocol (Madison, WI) following LaJeunesse
et al. (2003, 2010). All samples were initially analyzed using de-
naturing gradient gel electrophoresis (DGGE) fingerprinting (i.e.,
genotyping) of the ribosomal internal transcribed spacer 2 (ITS2)
with 45–80% gradient gels on a C.B.S. ScientificTM DGGE-2001,
followed by excision and DNA sequencing of all discreet, promi-
nent bands in the DGGE profile as described by LaJeunesse
(2002). Profiles and sequences were assigned an alphanumeric
ITS2 type designation (sensu LaJeunesse 2001) representative of
that fingerprint and symbiont type. This technique targets the nu-
merically dominant sequence variants in the symbiont’s genome
(Thornhill et al. 2007) and is able to detect symbionts com-
prising approximately 10% or more of a symbiotic population
(Thornhill et al. 2006). Because this study focused on the
3 5 4 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
ITS2 fingerprinting with PCR-DGGE &
band sequencing
Removalfrom
dataset
psbAncr sequencing and phylogenetic analyses
Multilocus microsatellitegenotyping
Not included in microsatellite genotyping
Not included in population
genetic analysis
Population genetic analyses
Sample collection & DNA extraction
Member of the C7, C7a/C12, or C3
Siderastrealineages?
Retrieval of a complete multilocus genotype?
NoYes
Diagnosis of aClade C type?
NoYes
NoYes
Figure 2. Data collection pipeline for this study with decision
points highlighted in gray.
evolution of Symbiodinium Clade C, results from other Clades
are not reported here (Fig. 2).
Each Clade C Symbiodinium sample was further analyzed by
amplifying and sequencing the noncoding region of the psbA mini-
circle (psbAncr) following the amplification settings and primers
(7.4-Forw and 7.8-Rev) specified by Moore et al. (2003) and
LaJeunesse and Thornhill (2011). All successful reactions were
sequenced using Big Dye 3.1 reagents (Applied Biosystems, Fos-
ter City, CA) at the DNA Core Facility (Penn State University)
using the forward primer (7.4-Forw). Only samples with a single
unambiguous psbAncr sequence (in ∼90% of samples) were used,
indicating the existence a single predominant Symbiodinium geno-
type (LaJeunesse and Thornhill 2011). Chromatograms were vi-
sually inspected for accuracy in base calling (Sequence Navigator)
and the edited sequences aligned initially using the online appli-
cation of ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/)
and additional adjustments to the alignment made during visual in-
spection of the output file. Sequences were deposited in GenBank
(see Table S1 for Accession numbers) and the resulting alignments
are available at http://datadryad.org (doi: 10.5061/dryad.2247c).
Ten polymorphic microsatellite loci developed for Clade
C Symbiodinium were used (loci Sgr 34, SgrSpl 13, SgrSpl 22,
SgrSpl 24, SgrSpl 25, SgrSpl 26, SgrSpl 78, Spl 1, and Spl 16
[Wham et al. 2013] and locus C1.05 [Bay et al. 2009]) to produce
multilocus genotypes for 67 samples of type C3 from Siderastrea
siderea, as well as 83 samples of C7 and 43 samples of C7a/
C12 from Orbicella spp. (Fig. 2 and Table S2). Each locus was
amplified in 10 μL reaction volumes containing 1 μL of 10 mM
dNTPs, 0.2 U Taq DNA Polymerase (New England Biolabs), 1 μL
standard Taq Buffer (New England Biolabs, Ipswich, MA), and
1 μL 25 mM MgCl2, 1 μL of each forward and reverse primer
at 10 μM, and 1 μL of approximately 50 ng DNA template.
PCR amplification was preformed according to the specifications
given by Wham et al. (2013). Following amplification, fragment
sizes were analyzed on an ABI 3730 Genetic Analyzer (Applied
Biosystems) using a 500-bp standard (LIZ-labeled) at the Penn
State University Nucleic Acid Facility. Fragments sizes were an-
alyzed visually using GeneMarker version 1.91 (SoftGenetics,
State College, PA). Although Symbiodinium, like most dinoflag-
ellates, are haploid (Santos and Coffroth 2003), most Clade C
members posses a duplicated genome in which many loci pro-
duce two alleles (Wham et al. 2013). Samples with background
genotypes (<10%), defined by electropherograms that contained
additional allele peaks one-third or less than the intensity of the
main peaks, were scored based on the dominant allele peaks. In
cases where multiple genotypes were observed to codominate,
the sample was removed from the dataset before further analysis
(Fig. 2). This reduced the sample size of the data set, but enabled
us to confidently identify multilocus genotypes and conduct pop-
ulation genetic analyses with the remaining data. The multilocus
allele scores and geographic data for these samples are available
at http://datadryad.org (doi: 10.5061/dryad.2247c).
DATA ANALYSIS
Sequences of the psbAncr fell into two distinct clusters—
haplotypes from Symbiodinium with ITS2 sequences similar or
identical to type C3 and haplotypes with ITS2 sequences similar
or identical to type C1 (see Results). Hereafter these psbAncr lin-
eages will be referred to as the C3 and C1 groups, respectively.
Because of the high level of psbAncr variability among samples,
it was impossible to unambiguously align sequences from the C3
group with those from the C1 group. Therefore, these two lineages
or groups of Symbiodinium Clade C were analyzed separately as
follows.
Phylogenetic reconstructions from the complete ITS2 and
partial sequences of the psbAncr were conducted using Bayesian
EVOLUTION FEBRUARY 2014 3 5 5

DANIEL J. THORNHILL ET AL.
Inference using MrBayes version 3.2.1 (Huelsenbeck and
Ronquist 2001) implementing the Hasegawa–Kishino–Yano
(HKY; ITS2 data), general time reversible (GTR) + I + � (C3
group psbAncr data), or symmetrical (SYM) + I + � (C1 group
psbAncr data) models of substitution, as indicated by the Akaike In-
formation Criterion by MrModeltest version 2.3 (Nylander 2004).
Each MCMC analysis was run for 1.0 × 106 generations and sam-
pled every 100 generations. Because of convergence of chains
within 2.5 × 105 generations, the first 2500 trees were discarded
as burn-in, and a 50% majority-rule consensus tree was calcu-
lated from 7501 remaining trees. Symbiodinium types C3 and
C1 from Indo-Pacific hosts were used as outgroups. Posterior
probabilities (PP) were recorded to assess reliability of recovered
nodes.
Topologies based on Maximum Parsimony were also con-
structed under the default settings of PAUP∗ 4.0b10 (Swofford
2000). For calculating phylogenies based on Maximum Parsi-
mony, and due to the high frequency of informative insertions and
deletions (indels), sequence gaps were included as a fifth character
state with the entire insertion or deletion weighted as one charac-
ter. Outgroups were identical to those used in Bayesian Inference.
A total of 1000 bootstrap replicates were performed to assess
statistical significance of internal branching. For all Bayesian In-
ference and Maximum Parsimony phylogenetic analyses, Symbio-
dinium ITS2-DGGE genotype, host identity, and collection loca-
tion were mapped onto the resulting topologies as character states.
Uncorrected genetic distances (P) between haplotypes, in-
cluding all substitutions and with gaps considered via pairwise
deletion, were calculated with MEGA version 5.2.2 (Tamura
et al. 2011). Sequences that differed by P < 0.030 were grouped
and the mean genetic distances between groups were calculated
in MEGA version 5.2.2.
Because recurring multilocus microsatellite genotypes (i.e.,
clones) bias allelic frequencies, clones were removed from the
data set (i.e., each genotype was represented only once; Fig. 2)
with GenAlEx version 6.1 (Peakall and Smouse 2006). The re-
sulting unique multilocus genotypes were analyzed using the
Bayesian clustering program STRUCTURE (Pritchard et al.
2000). The analysis in STRUCTURE was performed without a
prior location, under an admixture and uncorrelated alleles model
with a burn-in of 100,000 iterations and 1,000,000 iterations of
analysis at K values ranging from 1 to 10. The optimum K value
was determined by the Delta-K method (Evanno et al. 2005).
The program BEAST version 1.7.4 (Drummond and
Rambaut 2007) was used to approximate divergence times among
representatives of the C3 group estimated under a relaxed molec-
ular clock using the HKY substitution model (base frequencies
estimated). The tree prior was set at Yule Process (Gernhard 2008)
and random starting tree for the tree model. The mean substi-
tution rate (ucld.mean) of 0.006 substitutions × site−1 × My−1
(± 0.003) was calculated from the divergence between the psbAncr
haplotypes associated with Eastern Pacific and Atlantic Porites
(assuming divergence occurred 3.1–4.6 Mya; LaJeunesse 2005).
Several calibration points for estimating timing of a most recent
common ancestor (tmrca) were determined from nodes in the
phylogeny where Pacific and Atlantic lineages converged. Monte
Carlo Markov Chain length was set at 100 million.
ResultsRESOLUTION AMONG SYMBIODINIUM CLADE C WITH
ITS rDNA
ITS2 and partial 5.8S sequence data were compiled from 309
field-collected samples of Symbiodinium Clade C, comprising 38
cnidarian genera from 12 western Atlantic and five Indo-Pacific
locations (Table S1). The aligned ITS2 sequence data set included
300 nucleotide positions, all of which (100%) could be unam-
biguously aligned. Phylogenetic reconstruction from these ITS2–
5.8S alignments resulted in a largely unresolved polytomy with
member lineages unique to either the Pacific or the Atlantic. The
limited divergence between these sequences produced few statis-
tically supported lineages.
HIGH-RESOLUTION PHYLOGENETIC ANALYSIS
OF THE C1-GROUP
In contrast to the ITS2 region, psbAncr sequences were highly vari-
able across samples; in fact, samples belonging to the C1 group
did not align to the psbAncr sequences of the C3 group. Therefore,
these data were analyzed separately. The C1 group psbAncr align-
ment of 69 sequences consisted of 863 nucleotide positions and
63 unique haplotypes. Conserved regions in the alignment were
punctuated by areas with frequent nucleotide substitutions as well
as numerous insertion–deletion polymorphisms (see also Moore
et al. 2003). Phylogenetic analyses of these data characterized sev-
eral well-supported, monophyletic psbAncr lineages (Fig. 3A) that
are poorly resolved with ITS2 sequences (Fig. 3B). Some of the
monophyletic lineages corresponded to a particular ITS2 type des-
ignation, associated with a particular host taxon, or both (Fig. 3A).
For instance, the host-specific C1 group lineages from the west-
ern Atlantic (Palythoa spp. [ITS2 type C1] and Porites spp. [types
C1aj and C80]) were distinct from host-specific Pacific lineages
(e.g., Porites panamensis [types C1 and C1v]). However, other
lineages within the C1 group exhibited little host or geographic
specificity and ITS2 type C1 occurred throughout the phylogeny.
For example, one of the largest sequence clusters, designated as
Symbiodinium goreaui (sensu stricto), contained psbAncr haplo-
types from both the Atlantic and Pacific (Fig. 3A). Symbiodinium
goreaui was characterized by short internal branch lengths and lit-
tle differentiation between psbAncr haplotypes associated with the
3 5 6 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
*
0.85* 0.77
0.76
0.88
*
*
*
*
0.58
Porites astreoides
C1
Palythoa
C1 (Symbiodinium goreaui)
Acropora
Protopalythoa
Siderastrea
Bartholomea
Lebrunia
P. astreoidesDiscosoma
PalythoaTridacna
Corallimorpharia
C1aj
C80
0.1
Siderastrea
0.89
*0.87
**
*
*0.88
0.75
0.72
0.58
0.56
0.61*
*
0.78
0.89
Porites panamensis (Eastern Pacific)
C1b-f
C1Bartholomea
Fungia
PsammocoraCyphastrea
Galaxea
Coscinarea
Siderastrea
Fungia
Siderastrea
Zoanthus
AcroporaGalaxea
Astreopora
Discosoma
CorallimorphariaIsaurus
*
C1
C1v
Psammocora
**
*
*
*
cultured isolateIndo-Pacific origin
culture rt-152culture rt-113
B
0.98
f
b
ajC80
C1b-f
C1vC1
C1a-j
1 base diff.
A
Figure 3. High-resolution phylogeny of the C1 group of Symbiodinium based on a Bayesian Inference of chloroplast psbAncr (A). Nodal
support indicated as posterior probabilities (PP) above 0.50 next to the relevant node or asterisks for a PP = 1.00. Colored symbols
indicate the collection location of each sample corresponding to Figure 1. The corresponding Symbiodinium ITS2 types (based on DGGE
fingerprinting), symbiotic host affiliations, and cultured isolate designations are indicated to the right of each sample on the topology.
By comparison, Maximum Parsimony analysis of nuclear ribosomal ITS2 (B) provides less genetic resolution among members of the C1
group.
cnidarians Acropora, Bartholomea, Corallimorpharia, Dicosoma,
Lebrunia, Palythoa, Porites astreoides, Protopalythoa, Sideras-
trea, and the giant clam Tridacna. Although no psbAncr haplo-
types were shared between locations, similar psbAncr haplotypes
of S. goreaui were identified throughout the western Atlantic in-
cluding Belize, Brazil, the Florida Keys, the Gulf of Mexico, and
Mexico, as well as from the Indo-Pacific. Thus, the C1 group con-
tained both host-specific lineages and geographically widespread
host-generalists.
HIGH-RESOLUTION PHYLOGENETIC ANALYSIS
OF THE C3 GROUP
The psbAncr nucleotide alignment of the C3 group consisted of
242 sequences comprising 181 unique psbAncr haplotypes and
826 aligned nucleotide positions (indels included). The Bayesian
Inference phylogenetic reconstruction of the C3 group produced
numerous divergent lineages that grouped mostly by host origin
(Fig. 4A). By comparison, these lineages were poorly resolved
with ITS2 sequences (Fig. 4B). Included in these analyses were
EVOLUTION FEBRUARY 2014 3 5 7

DANIEL J. THORNHILL ET AL.
A
* *
**
*
*
0.81
0.81
*
**
* *0.73
0.90 0.88
0.52
0.68
*0.96
0.98
0.67
0.67 0.98*
*
*0.81
*
**
0.98
0.840.60
0.72
*
*0.63
** *
**0.980.83
**
*
0.57
0.630.59
* 0.64
0.76
0.1
Indo-Pacific
C3h
C3 Scolymia
C3 Mussa
C3 Leptoseris
C3 Siderastrea siderea
C3 Ricordia
C3 StephanocoeniaC3 Erythropodium
Indo-Pacific
C3 Leptoseris
C3 AgariciaC3c Mycetophyllia reesi
C3 Isophyllastrea
C3 Siderastrea siderea
C3, C3b, C3q
Agaricia & Leptoseris
C3 Agaricia
Atla
ntic
Atla
ntic
C31, C31c
Montipora
Indo-Pacific
C3C16a
C16C87
Stephanocoenia
C3 SiderastreaC3 Diploria
C3 Mycetophyllia
C27
C3 Orbicella franksi
Agaricia & Leptoseris
C3 MycetophylliaC3 Scolymia
C3
C3k Acropora
C40 Indo
-Pac
ific
C7, C7a(C12), C7cOrbicella
annularis
complex
0.89
0.96
C40C3a
C87
C31/ C31c
C3
C3b
C21
C3s
C3q
C7C7a (C12)
C16
C16a
c
C17
C21a
C3k
C27
B
C3, C3a, C3b, C3s
a
1 base diff.
Figure 4. High-resolution phylogeny of the C3 group in Symbiodinium based on Bayesian inference of chloroplast psbAncr (A). Nodal
support indicated as posterior probabilities (PP) above 0.50 next to the relevant node or asterisks for a PP = 1.00. Dark grey arrowheads
(�) denote ancestral branches in the radiation with low support values. Symbiodinium ITS2 types (based on DGGE fingerprinting),
symbiotic host affiliation, and major ocean basin are indicated to the right of each clade on the topology. Star symbols (�) indicate branch
connections to subclades that are drawn in greater detail in Figures 5, S1, and S2. By comparison, Maximum Parsimony analysis of nuclear
ribosomal ITS2 (B) provides less genetic resolution among members of the C3 group.
3 5 8 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
several Indo-Pacific representatives (e.g., C3h, C27, and C31)
that shared a more recent common ancestry with western Atlantic
representatives than with each other, indicating that the initial
diversification of these lineages predated the closure of the Central
American Seaway. The putatively ancestral C3 ITS2 genotype was
found to represent distinct lineages throughout the phylogeny,
whereas more derived ITS2 genotypes occurred as monophyletic
(i.e., C3a, C3h, C3k, C3q, C7, C7c, C12, C16a, C27, C31/C31c,
C40), paraphyletic (i.e., C3c, C3s, C7a, C16, C87), or polyphyletic
(i.e., type C3b) groups. The evolutionary relatedness inferred via
Maximum Parsimony for many of these lineages was similar to
that estimated by Bayesian Inference (data not shown).
Statistical support was generally low for most branching
points, or ancestral nodes, that occurred deeper in the psbAncr
phylogeny of the C3 group (indicated with dark gray arrowheads
in Fig. 3; PP = 0.52–0.72). PsbAncr lineages from the Atlantic all
exhibited host-specific associations whereas many of the Indo-
Pacific lineages used in our analysis associated with various host
taxa. Furthermore, two or more distinct psbAncr lineages associ-
ated with the Atlantic coral genera Agaricia, Leptoseris, Myce-
tophyllia, Orbicella, Scolymia, Siderastrea, and Stephanocoenia
(Fig. 4A). Finally, members of the C3 group occurred predomi-
nantly in deeper environments (mean depth = 12.0 ± 0.5 m [SE];
Table S1; two-sample T-test, P < 0.0001) than the generally shal-
lower habitats of the C1 group (mean depth = 6.5 ± 1.0 m [SE];
Table S1).
GENETIC COHESION AMONG SYMBIODINIUM
ASSOCIATED WITH ORBICELLA
ITS2 and psbAncr genotyping revealed a large group of Symbio-
dinium (types C7, C7a, C7c, and C12) that associated specifi-
cally with O. annularis, Orbicella faveolata, and Orbicella franksi
throughout the Greater Caribbean. Phylogenetic analysis of the
psbAncr haplotypes showed that these Symbiodinium comprised
a well-supported monophyletic group that splits into two distinct
subclades (Figs. 4, 5). These lineages are further divided into less
resolved groupings that correlated with location and ITS2 type.
For example, psbAncr haplotypes of type C7 Symbiodinium oc-
curred in Orbicella spp. from the western Caribbean, including
Mexico and Belize. In contrast, psbAncr haplotypes of type C7a
occurred in Orbicella spp. from the eastern Caribbean province,
including St. Croix and Barbados; closely related C12 psbAncr
haplotypes were obtained from the Bahamas, whereas type C7c
was found only in Panama. Curacao represented a well-sampled
location where overlap between psbAncr haplotypes of C7 and C7a
occurred (Fig. 5). In all of these cases, psbAncr haplotypes were
distinct at a particular location and none were shared among local-
ities, further indicating that these Symbiodinium populations are
spatially structured. Similar fine-scale phylogeographic patterns
*
*
C7a*
C12
C7c
*
C7
B
C
D
5 changes
C3
*
*
**
*
*
*
*
A Symbiodinium Clade C associated with Orbicella sp.
Figure 5. (A) Maximum Parsimony topology based on the Sym-
biodinium chloroplast psbAncr haplotypes associated with mem-
bers of the bolder star coral, Orbicella annularis, species complex.
The basal node of this tree connects to the topology presented
in Figure 4 (note � symbol at the node labeled C7/C7a/C12 in
Fig. 4). Nodal support with bootstrap values greater that 95%
are indicated by asterisks. Symbiodinium nuclear ribosomal ITS2-
DGGE genotypes are indicated to the right of each clade on
the topology. Colored symbols indicate the collection location of
each sample corresponding to Figure 1. Panels B–D provide ex-
ample photographs of the O. annularis species complex, includ-
ing Orbicella faveolata (B), Orbicella annularis (C), and Orbicella
franksi (D).
were observed for Siderastrea siderea and the agaricid genera
Agaricia and Leptoseris (Figs. S1, S2 and Text S1, S2).
To investigate whether recent sexual recombination occurred
within and between the two major groupings of Orbicella sym-
bionts, 10 microsatellite markers were examined in Symbiodinium
C7, C7a/C12, and a third monophyletic lineage, the C3 type from
Siderastrea (i.e., C3Siderastrea). A total of 193 samples were geno-
typed. Of these, 26 were removed because they contained clear
mixtures of more than one genotype and another 89 were removed
because they constituted clone replicates. This left a total of 78
distinct genotypes (C7, n = 31; C7a/C12, n = 14; C3Siderastrea,
n = 33) for analysis of genetic similarity. For summary statistics
EVOLUTION FEBRUARY 2014 3 5 9
DANIEL J. THORNHILL ET AL.
Beliz
e
Mex
ico
Cura
çao
Baha
mas
Barb
ados
Mex
ico
Beliz
eBa
ham
as
Flor
ida
St. C
roix
Barb
ados
Cura
çao
St. C
roix
C7 C7a C3Siderastrea
0
50
100
0
50
100
K=3
K=5
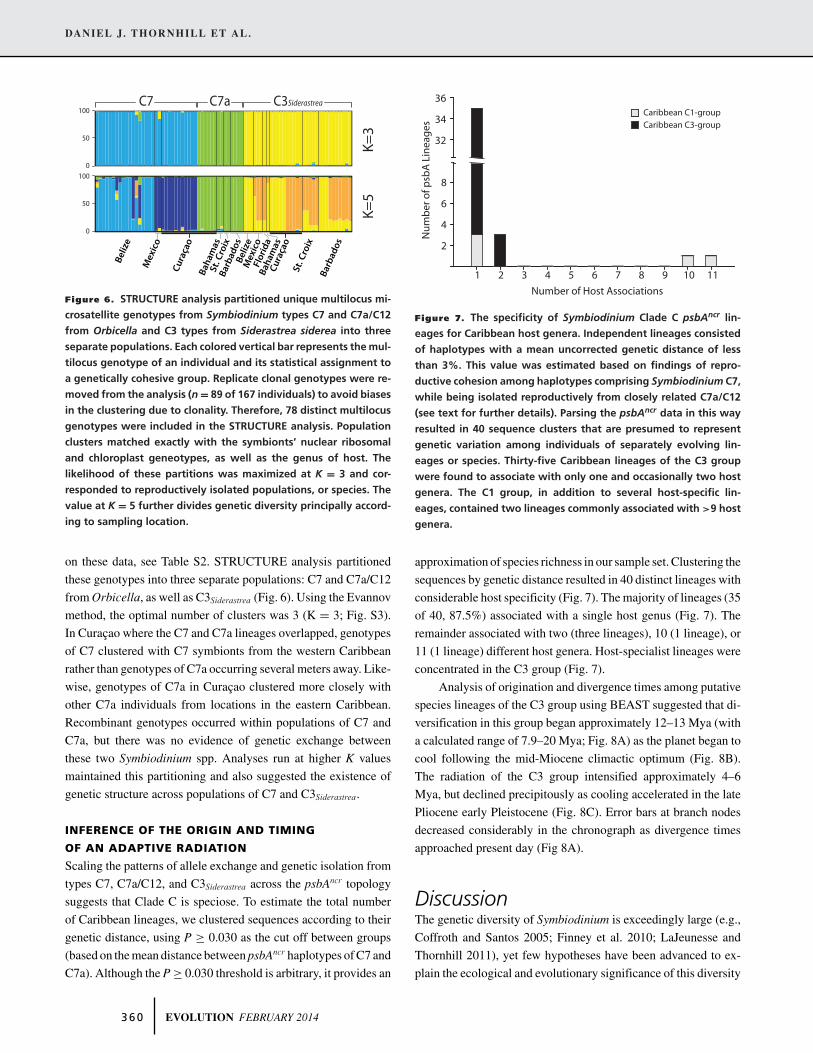
Figure 6. STRUCTURE analysis partitioned unique multilocus mi-
crosatellite genotypes from Symbiodinium types C7 and C7a/C12
from Orbicella and C3 types from Siderastrea siderea into three
separate populations. Each colored vertical bar represents the mul-
tilocus genotype of an individual and its statistical assignment to
a genetically cohesive group. Replicate clonal genotypes were re-
moved from the analysis (n = 89 of 167 individuals) to avoid biases
in the clustering due to clonality. Therefore, 78 distinct multilocus
genotypes were included in the STRUCTURE analysis. Population
clusters matched exactly with the symbionts’ nuclear ribosomal
and chloroplast geneotypes, as well as the genus of host. The
likelihood of these partitions was maximized at K = 3 and cor-
responded to reproductively isolated populations, or species. The
value at K = 5 further divides genetic diversity principally accord-
ing to sampling location.
on these data, see Table S2. STRUCTURE analysis partitioned
these genotypes into three separate populations: C7 and C7a/C12
from Orbicella, as well as C3Siderastrea (Fig. 6). Using the Evannov
method, the optimal number of clusters was 3 (K = 3; Fig. S3).
In Curacao where the C7 and C7a lineages overlapped, genotypes
of C7 clustered with C7 symbionts from the western Caribbean
rather than genotypes of C7a occurring several meters away. Like-
wise, genotypes of C7a in Curacao clustered more closely with
other C7a individuals from locations in the eastern Caribbean.
Recombinant genotypes occurred within populations of C7 and
C7a, but there was no evidence of genetic exchange between
these two Symbiodinium spp. Analyses run at higher K values
maintained this partitioning and also suggested the existence of
genetic structure across populations of C7 and C3Siderastrea.
INFERENCE OF THE ORIGIN AND TIMING
OF AN ADAPTIVE RADIATION
Scaling the patterns of allele exchange and genetic isolation from
types C7, C7a/C12, and C3Siderastrea across the psbAncr topology
suggests that Clade C is speciose. To estimate the total number
of Caribbean lineages, we clustered sequences according to their
genetic distance, using P ≥ 0.030 as the cut off between groups
(based on the mean distance between psbAncr haplotypes of C7 and
C7a). Although the P ≥ 0.030 threshold is arbitrary, it provides an
Number of Host Associations1 2 3 4 5 6 7 8 9 10 11
Num
ber o
f psb
A L
inea
ges
2
4
6
8
32
34
36Caribbean C1-groupCaribbean C3-group
Figure 7. The specificity of Symbiodinium Clade C psbAncr lin-
eages for Caribbean host genera. Independent lineages consisted
of haplotypes with a mean uncorrected genetic distance of less
than 3%. This value was estimated based on findings of repro-
ductive cohesion among haplotypes comprising Symbiodinium C7,
while being isolated reproductively from closely related C7a/C12
(see text for further details). Parsing the psbAncr data in this way
resulted in 40 sequence clusters that are presumed to represent
genetic variation among individuals of separately evolving lin-
eages or species. Thirty-five Caribbean lineages of the C3 group
were found to associate with only one and occasionally two host
genera. The C1 group, in addition to several host-specific lin-
eages, contained two lineages commonly associated with >9 host
genera.
approximation of species richness in our sample set. Clustering the
sequences by genetic distance resulted in 40 distinct lineages with
considerable host specificity (Fig. 7). The majority of lineages (35
of 40, 87.5%) associated with a single host genus (Fig. 7). The
remainder associated with two (three lineages), 10 (1 lineage), or
11 (1 lineage) different host genera. Host-specialist lineages were
concentrated in the C3 group (Fig. 7).
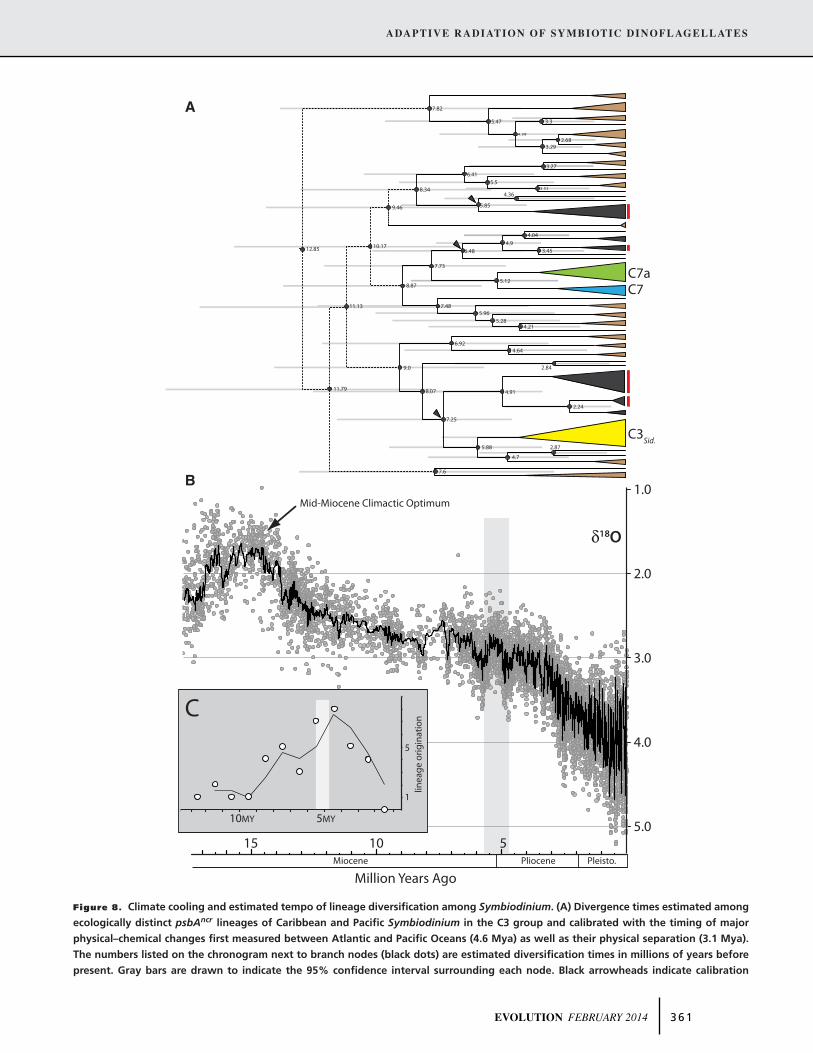
Analysis of origination and divergence times among putative
species lineages of the C3 group using BEAST suggested that di-
versification in this group began approximately 12–13 Mya (with
a calculated range of 7.9–20 Mya; Fig. 8A) as the planet began to
cool following the mid-Miocene climactic optimum (Fig. 8B).
The radiation of the C3 group intensified approximately 4–6
Mya, but declined precipitously as cooling accelerated in the late
Pliocene early Pleistocene (Fig. 8C). Error bars at branch nodes
decreased considerably in the chronograph as divergence times
approached present day (Fig 8A).
DiscussionThe genetic diversity of Symbiodinium is exceedingly large (e.g.,
Coffroth and Santos 2005; Finney et al. 2010; LaJeunesse and
Thornhill 2011), yet few hypotheses have been advanced to ex-
plain the ecological and evolutionary significance of this diversity
3 6 0 EVOLUTION FEBRUARY 2014
ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
A
B
51015
Million Years Ago
1.0
2.0
3.0
4.0
5.0
δ18O
Miocene Pliocene Pleisto.
Mid-Miocene Climactic Optimum
C3Sid.
C7C7a
12.85 10.17
11.13
11.79
8.87
9.0
8.07
7.25
7.6
4.7
2.87
2.24
4.91
5.88
6.92
5MY10MY
linea
ge o
rigin
atio
n
1
5
C
4.64
4.215.28
5.967.48
2.84
5.12
7.73
6.48
4.04
3.454.9
9.46 5.85
8.34
6.41
4.36
3.27
3.29
5.5
2.68
7.82
5.47 3.3
Figure 8. Climate cooling and estimated tempo of lineage diversification among Symbiodinium. (A) Divergence times estimated among
ecologically distinct psbAncr lineages of Caribbean and Pacific Symbiodinium in the C3 group and calibrated with the timing of major
physical–chemical changes first measured between Atlantic and Pacific Oceans (4.6 Mya) as well as their physical separation (3.1 Mya).
The numbers listed on the chronogram next to branch nodes (black dots) are estimated diversification times in millions of years before
present. Gray bars are drawn to indicate the 95% confidence interval surrounding each node. Black arrowheads indicate calibration
EVOLUTION FEBRUARY 2014 3 6 1

DANIEL J. THORNHILL ET AL.
(but see LaJeunesse 2005; Pochon et al. 2006; Hill and Hill 2012).
The recent availability of additional high-resolution phylogenetic
and population genetic (LaJeunesse and Thornhill 2011; Wham
et al. 2013) markers enables detailed examination of evolution-
ary relationships among Symbiodinium. Here, we focused on the
psbAncr, which exhibits levels of sequence evolution 10–20 times
greater than that of ITS2 and unambiguously differentiates ge-
netically cohesive lineages comprising distinct haplotypes (LaJe-
unesse and Thornhill 2011). When psbAncr data were compared
with ITS2 and multilocus microsatellite genotyping, concordant
patterns occurred across markers, indicative of species bound-
aries. Below we examine these results in the context of different
species concepts and consider abiotic and biotic factors poten-
tially underlying the diversification of Symbiodinium Clade C.
DELIMITING SPECIES LINEAGES WITH
HIGH-RESOLUTION GENETIC DATA
Although different species concepts emphasize different lines of
evidence, most share the idea that species are separately evolving
units (de Queiroz 2007; Hausdorf 2011). The phylogenetic species
concept, for example, identifies species using statistically signifi-
cant character differences resulting from evolutionary divergence
of populations over time (Donoghue 1985; de Queiroz 2007). Un-
der this concept, members of a species share more diagnostic ge-
netic or morphological characters with each other—and are hence
monophyletic—than with individuals from other species. Phylo-
genetic reconstructions based on the psbAncr revealed fine-scale
genetic diversity in Symbiodinium Clade C, including numerous
monophyletic clades that were differentiated from each other by
many nucleotide substitutions and indels. These phylogenetic pat-
terns are consistent with species-level distinctions, in that sister
lineages, as independently evolving metapopulations, appear to
have lost shared ancestral haplotypes (Wakeley 2008).
The frequent phylogenetic correspondence, or reciprocal
monophyly, of the independently sorting ITS (nuclear) and psbAncr
(chloroplast) provides strong support for the hypothesis that Clade
C comprises hundreds of species worldwide. Despite this, sev-
eral genetically and ecologically distinct psbAncr lineages shared
dominant ITS2 sequences (e.g., type C3). In such instances, the
unusual molecular evolution of the multicopy ribosomal array is
likely responsible for the discrepancy between markers. Evolu-
tion of rDNA begins as a mutation in one copy, which spreads
across copies via gene conversion or unequal crossing over, and
ultimately becomes numerically dominant within the genomes
of individuals undergoing genetic exchange in a population—
a process known as concerted evolution (i.e., Smith 1976;
Arnheim 1983; Dover 1986; Saito et al. 2002; Galluzzi et al.
2004). Concerted evolution exerts a stabilizing influence that can
prevent the spread of rare sequence variants, thereby maintaining
the numerically dominant rDNA sequence and masking lineage
divergence (Arnheim 1983; Dover 1986; Thornhill et al. 2007).
The shared C3 sequence that dominated the genomes of many
Symbiodinim Clade C lineages is likely a shared ancestral state, a
symplesiomorphy, which has not been replaced by a more derived
sequence.
Consequently, high-resolution genetic data indicate that
ITS2-nrDNA only approximates species richness within Clade
C. In cases such as Symbiodinium type C3 or type B1 from the
Caribbean, fine-scale genetic markers demonstrate that apparent
host-generalists actually consist of several specialized lineages
(Santos et al. 2004; Finney et al. 2010). As a result, populations
of other provisional host-generalists, including ITS2 types A1,
A3, C15, and D1a (D1–4), require reexamination with higher-
resolution genetic markers (see Pinzon et al. 2011; Wham et al.
2011). Future investigations may reveal that putative generalists
instead comprise several independent and ecologically specialized
species lineages, but others like S. goreaui may indeed associate
with many different host genera over a large geographic area, and
are thus true host-generalist symbionts (Fig. 3A).
The phylogenetic patterns of the psbAncr beg the questions:
do the discreet sequence clusters represent evolutionarily distinct
species? Or are they simply the accumulation of somatic muta-
tions through clonal evolution? Given the clonality of many in
hospite Symbiodinium populations (e.g., Thornhill et al. 2013),
clonal evolution and periodic selective sweeps (Palys et al. 1997)
might explain the concordant phylogenetic patterns of the chloro-
plast and nuclear loci. We therefore tested for evidence of ge-
netic recombination using multilocus population genetic data.
Three sympatric ITS2/psbAncr lineages were examined, includ-
ing C7 and C7a/C12 from the star corals, Orbicella spp., and
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 8. points for estimating timing of a most recent common ancestor (tmrca) between Pacific and Atlantic lineages. The Atlantic
Symbiodinium spp., examined by microsatellite analyses (Fig. 6) and associated with the reef corals Orbicella (C7 and C7a/C12) and
Siderastrea (C3Sid), are noted so as to delimit the ranges of interspecific versus interindividual sequence variation. Putative species
lineages are shaded according to their geographic origin (black = Indo-Pacific, brown = Atlantic). Stippled lines represent branches with
poor statistical support (Bayesian Inference). Red highlighting to the right of terminal branches indicates host-generalist lineages. (B)
Climate curve over the past 17 My based on oxygen-isotope measurements from deep-sea foraminifera (from Zachos et al. 2008). (C) Basic
estimation of the rate and tempo of lineage diversification in the chronogram (panel A). The gray shading in panels B and C encompasses
the time horizon of major climate shifts corresponding to the Miocene–Pliocene boundary.
3 6 2 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
C3Siderastrea from the massive starlet coral, Siderastrea siderea,
with 10 unlinked microsatellite loci. STRUCTURE analyses iden-
tified boundaries to sexual recombination between C7, C7a/C12,
and C3Siderastrea that corresponded to the symbionts’ phylogenetic
grouping and host affiliation (Fig. 6). Thus, host affiliation and
phylogenetic groupings superseded location as the primary fac-
tors in determining clustering. Furthermore, individuals belong-
ing to each ITS2/psbAncr lineage comprised recombinant (i.e.,
scrambled) allele combinations, indicating sexual recombination.
Members within each phylogenetic grouping, or lineage, appear
capable of engaging in sexual recombination, but are reproduc-
tively isolated from the constituencies of other lineages. These
three lineages therefore operate as distinct species according to
the biological species concept.
A common driver of monophyly, reproductive isolation, and
speciation is the selection of individuals with gene combinations
adapted to a resource or habitat (Ackerly et al. 2006; Losos 2009;
Glor 2010). Here, most psbAncr lineages contained individual hap-
lotypes recovered from only a single coral species or genus and
are thus functionally distinct species under the ecological species
concept. These clear ecological differences are consistent with the
adaption of many psbAncr clades to a particular host habitat, sug-
gesting the importance of distinct intracellular environments—
characteristic of different hosts—in the ecology and evolution of
these dinoflagellates. Indeed, host identity appears to be the pri-
mary ecological factor, or α-niche axis, of lineage diversification
in Symbiodinium (Figs. 3, 4, 6; LaJeunesse 2005; Ackerly et al.
2006; Finney et al. 2010; Glor 2010; LaJeunesse et al. 2010).
In addition to the Symbiodinium associated with hosts that verti-
cally transmit their symbionts, many host-specific lineages were
found associated with Caribbean Cnidaria that must horizontally
acquire symbionts from the environment at the start of each gen-
eration. This indicates that vertical transmission is unnecessary
for the evolution of host-specialized Symbiodinium spp. Further-
more, we found examples of hosts that harbored several phylo-
genetically independent and host-specific endosymbionts across
their range (e.g., Fig. S2), suggesting that endosymbionts evolved
along similar ecological axes in different locations (i.e., paral-
lel evolution) and that speciation through ecological selection is
a major recurrent theme throughout the evolutionary history of
these dinoflagellates.
Beyond host associations, many monophyletic and host-
specific psbAncr clades within the C3-group contained geograph-
ically segregated haplotypes (e.g., in symbionts of Orbicella,
Siderastrea, Agaricia), suggesting that spatial isolation acts as
a secondary or β-niche axis of divergence in Symbiodinium evo-
lution (Ackerly et al. 2006; Glor 2010). This spatial partitioning
mirrored the presence of distinct coral reef species assemblages
(Briggs 1974), as well as the genetic partitioning of several shal-
low water species in the Greater Caribbean (Fig. 1; Taylor and
Helberg 2003; Baums et al. 2005), that relate to theoretically
expected patterns of allopatry based on major surface currents
(Cowen et al. 2006) and long-standing environmental gradients
(Chollett Ordaz 2011). Moreover, these phylogeographic patterns
are consistent with previous findings of genetic structure in Sym-
biodinium populations at scales encompassing several hundred
kilometers, or less (e.g., Santos et al. 2003; Howells et al. 2009;
Thornhill et al. 2009; Andras et al. 2011). Limited connectivity
among Symbiodinium populations probably results from a combi-
nation of geographic barriers such as prevailing currents, limited
dispersal capacity of free-living cells, high local recruitment or
retention, or density-dependent barriers preventing long-distance
migrants from successfully penetrating habitats that are already
dominated by other Symbiodinium genotypes (reviewed in Thorn-
hill et al. 2009). Finally, different local and regional conditions
may also select for locally adapted genotypes (Chollett Ordaz
2011; Howells et al. 2012).
Taken together, these ecological, geographic, reproductive,
population genetic, and phylogenetic patterns suggest that Sym-
biodinium Clade C could consist of hundreds of species globally.
These findings also point to major biotic and abiotic factors—host
specialization and geographic isolation—shaping Symbiodinium
evolution. The following section considers these observations in
the context of the paleo-history of the Caribbean Basin in order
to make general inferences about their evolutionary history with
reef corals.
A SYNTHESIS OF THE INTERPLAY BETWEEN
PALEO-HISTORY, GEOGRAPHIC ISOLATION, AND
HOST SPECIALIZATION
Phylogenetic comparisons between Cnidaria and Symbiodinium
indicate a lack of co-speciation over tens of millions of years
(Rowan and Powers 1991; LaJeunesse 2005; Pochon et al. 2006;
this study). A more dynamic process of partner recombination
and coevolution must be at work to explain the existence of nu-
merous closely related endosymbionts associated with distantly
related hosts (LaJeunesse 2005). Evidence from our phylogeo-
graphic and molecular clock analyses (Figs. 4, 8) indicate that
an acceleration of lineage diversification occurred with the sep-
aration of the Atlantic from the Pacific during the late Miocene
and early Pliocene. Continental drift and tectonic uplift initiated
major changes in ocean circulation patterns that coincided with
the overall reduction in the planet’s temperature and atmospheric
CO2 concentration (Haug and Tiedemann 1998; Zachos et al.
2001, 2008; Pearson and Palmer 2000). Terrestrial changes dur-
ing this time included a drying of the climate, significant turnover
in terrestrial mammals, and expansion of C4 grasses (Haq et al.
1987; Ruddiman and Raymo 1988; Janis 1993; Cerling et al.
1997; Haug et al. 2001). In Caribbean marine environments, coral
species diversity initially increased with the complete closure of
EVOLUTION FEBRUARY 2014 3 6 3

DANIEL J. THORNHILL ET AL.
the Central American Seaway (Budd 2000). However, climate
shifts continued during the Plio–Pleistocene transition, notably
including the onset of northern hemisphere glaciation and asso-
ciated oscillations in temperature, pCO2, and eustasy, resulting
in continual ecological change (Zachos et al. 2001, 2008). These
effects were more pronounced in the Greater Caribbean, which ex-
perienced significant losses in coral diversity relative to the Pacific
(Budd 2000). Major changes in climate that induced reef faunal
turnover presumably affected Symbiodinium species assemblages
as well.
A once small number of species within Symbiodinium Clade
C, designated by ancestral ITS2 genotypes C3 and C1 that were
common to both ocean basins, served as the progenitors to the
radiation of the C1 and C3 groups. Presumably, these ancestors
or their descendants displaced Symbiodinium spp. that existed
before the changes in climate related to the Miocene–Pliocene
transition. According to the psbA data, a major burst in clado-
genesis began in the Atlantic during the late-Miocene (Fig. 8A).
By 3.1–4.6 Mya, genetic exchange between the Atlantic and Pa-
cific had ceased, resulting in independent evolutionary trajecto-
ries among Symbiodinium populations located in each ocean basin
(LaJeunesse 2005; LaJeunesse et al. 2010). The shifts in climate
conditions and the taxonomic composition of hosts that occurred
during and following this event ostensibly favored the diversi-
fication and specialization of Symbiodinium according to host
identity and geographic location, resulting in the diverse assem-
blage of endosymbiont species found on contemporary Caribbean
reefs (Figs. 4, 5, S1, S2). PsbAncr and ITS2 sequence data sug-
gest that Pacific lineages in the C3 group underwent a parallel
radiation during this time (LaJeunesse 2005); however, several
dominant host-generalist lineages remain in the Pacific C3 group
(Figs. 4, 8A).
There is precedent for the proliferation and shift in dom-
inance of other microbial groups during major environmental
change, perhaps as a result of adaptive innovation, reduced com-
petition, or extinctions that made available previously occupied
niches (Crouch et al. 2001; Falkowski et al. 2004). Surpris-
ingly, after the large number of extinctions associated the Plio-
Pleistocene transition, the Caribbean fauna of reef corals has re-
mained stable for approximately 1.5 My despite frequent, evenly
spaced oscillations in global temperatures (Jackson et al. 1996;
Jackson and Johnson 2000; Pandolfi and Jackson 2001). During
that period, reduced sea level and altered sea-surface currents
with each glacial cycle created oceanographic barriers to disper-
sal in the western Atlantic basin (Veron 1995) that hypothetically
provided opportunities for sequence coalescence and the evolu-
tion of numerous regionally localized psbAncr haplotypes (e.g.,
Figs. 5, S1, S2). Indeed, Pleistocene climates may have generated
regionally localized species radiations involving Symbiodinium in
Clades B (LaJeunesse 2005) and D (LaJeunesse et al. 2010).
Why was the C3 group of Clade C Symbiodinium more evo-
lutionarily successful in terms of the numbers of hosts and habitats
occupied in the Atlantic (and Pacific)? Although answers to this
question are speculative given the available data, some insight
may be gained by comparing the ecologies of extant species with
the historic climatic conditions during the radiation of this group
over the past 5 My. It is noteworthy that members of the C3 group,
usually occurred in colonies from deeper environments (>10 m),
where average temperatures and irradiance levels are lower and
less variable. By comparison, members of the C1 group occurred
in shallower habitats (∼5 m), where they would be subjected
to greater extremes in temperature and light conditions. Further-
more, type C1 examined in Australia is substantially more heat
tolerant than type C3 (referred to as type C2 in Berkelmans and
van Oppen 2006; Abrego et al. 2008; Jones et al. 2008; but see
Hume et al. 2013). This difference in diversity between the C1
and C3 groups in the Greater Caribbean is significant because
the Plio–Pliestocene was cold relative to the rest of the Ceno-
zoic (Fig. 8B; Weyl 1968; Burchardt 1978; Wolfe 1978; Haq
et al. 1987; Ruddiman and Raymo 1988). It is plausible, there-
fore, that a cool climate contributed to the diversification of the C3
group. Alternatively, fewer shallow water niches may have been
available to members of the C1 group in the Atlantic due to the
success of Symbiodinium Clade B (and to a lesser degree Clade
A) in animals from shallow Caribbean habitats (LaJeunesse 2002,
2005; Finney et al. 2010).
In summary, climate change, favoring the success of a few
ancestors that subsequently diversified principally through host
specialization in geographic isolation, likely drove the evolution
of Symbiodinium species diversity over the past 12 My. Under this
scenario, divergent selection, acting on genetic variation within
a population, favored adaptations that increased fitness in a par-
ticular habitat (Dieckmann and Doebeli 1999; Schluter 2001),
which is determined primarily by the intracellular host envi-
ronment. Disruptive selection likely opposed gene flow between
sympatric subpopulations adapting to different hosts. Genotypes
capable of associating with multiple host species (i.e., multiple
habitats) would be suboptimal because, under ecological the-
ory, these phenotypes experienced reduced fitness relative to a
specialist within any particular niche (Dieckmann and Doebeli
1999; Schluter 2001). Instead, reinforcement by disruptive se-
lection and assortative mating likely increased the prevalence of
genotypes that were competitively superior in symbioses with
a particular host taxon. This ongoing process would result in
reproductively isolated Symbiodinium populations, and eventu-
ally generate species assemblages partitioned mostly by host
identity (i.e., ecological speciation; Dieckmann and Doebeli 1999;
Schluter 2001, 2009).
Reef coral symbioses of the near future will encounter
a warmer climate and more acidic ocean than that of the
3 6 4 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
preceding 7 My (Burchardt 1978; Wolfe 1978; Ruddiman and
Raymo 1988; Pachauri and Reisinger 2007). Many Symbiodinium
spp. face displacement and extinction with increased sea-surface
temperatures and greater, or more erratic, seasonal fluctuations
in temperature that favor opportunistic species (LaJeunesse et al.
2009). Alternatively, some populations may acclimatize or adapt
to this environmental change or migrate with their host popula-
tions to more suitable locations. Although the specific outcomes
of these processes are uncertain, our reconstruction of evolution-
ary history suggests that Symbiodinium spp. respond to climate
change through “boom and bust” phases in diversification and
extinction. As with past changes in climate, the current warm-
ing may facilitate the rise and spread of host-generalists that are
competitively inferior under stable conditions but experience suc-
cess in a changing environment (e.g., Thornhill et al. 2006; Jones
et al. 2008; LaJeunesse et al. 2009).
ACKNOWLEDGMENTSThe authors sincerely thank K. K. Richter and K. M. Halanych for as-sistance with the phylogenetic analyses as well as J. Zachos for sharingthe most current adjusted oxygen-isotope data from deep-sea benthicforaminifera. The authors also thank E. Sampayo, A. Cecere, R. Smith,T. White, L. L. Walsh, H. Oxenford, and W. K. Fitt for contributing invarious ways to this work. This research was supported by the Pennsyl-vania State University, the IOC-UNESCO-World Bank targeted workinggroup on coral bleaching, and the National Science Foundation grants(IOB 544854 and OCE-09287664).
LITERATURE CITEDAbrego, D., K. E. Ulstrup, B. L. Willis, and M. J. H. van Oppen. 2008.
Species-specific interactions between algal endosymbionts and coralhosts define their bleaching response to heat and light stress. Proc. R.Soc. B 1648:2273–2282.
Ackerly, D. D., D. W. Schwilk, and C. O. Webb. 2006. Niche evolution andadaptive radiation: testing the order of trait divergence. Ecology 87:S50–S61.
Agadjanian, A. K. 2003. Adaptive radiation of mammals: major stages. Pale-ontolog. J. 37:179–186.
Andras, J. P., N. L. Kirk, and C. D. Harvell. 2011. Range-wide populationgenetic structure of Symbiodinium associated with the Caribbean Seafan coral, Gorgonia ventalina. Mol. Ecol. 20:2525–2542.
Arnheim, N. 1983. Concerted evolution of multigene families. Pp. 38–61 inM. Nei and R. K. Koehn, eds. Evolution of genes and proteins. SinauerAssociates, Sauderland, MA.
Baldwin, B. G., D. W. Kyhos, J. Dvorak, and G. D. Carr. 1991. ChloroplastDNA evidence for a North-American origin of the Hawaiian silverswordalliance (Asteraceae). Proc. Natl. Acad. Sci. USA 88:1840–1843.
Barbrook A. C., S. Visram, A. E. Douglas, and C. J. Howe. 2006. Moleculardiversity of dinoflagellate symbionts of Cnidaria: the psbA minicircle ofSymbiodinium. Protist 157:159–171.
Barnes, D. J., and B. E. Chalker. 1990. Calcification and photosynthesis inreef-building corals and algae. Pp. 109–131 in Z. Dubinski, ed. Ecosys-tems of the world: 25 coral reefs. Elsevier Science, Amsterdam.
Baums, I. B., M. W. Miller, and M. E. Hellberg. 2005. Regionally isolatedpopulations of an imperiled Caribbean coral, Acropora palmata. Mol.Ecol. 14:1377–1390.
Bay, L. K., E. J. Howells, and M. J. H. van Oppen. 2009. Isolation, char-acterisation and cross amplification of thirteen microsatellite loci forcoral endo-symbiotic dinoflagellates (Symbiodinium clade C). Conserv.Genet. Res. 1:199–203.
Beerling, D. J. 2002. Low atmospheric CO2 levels during the Permo-Carboniferous glaciation inferred from fossil lycopsids. Proc. Natl.Acad. Sci. USA 99:12567–12571.
Berkelmans, R., and M. J. H. van Oppen. 2006. The role of zooxanthellae inthe thermal tolerance of corals: a ‘nugget of hope’ for coral reefs in anera of climate change. Proc. R. Soc. Lond. B Biol. Sci. 273:2305–2312.
Briggs, J. C. 1974. Marine zoogeography. McGraw-Hill, New York.Budd, A. F. 2000. Diversity and extinction in the Cenozoic history of
Caribbean reefs. Coral Reefs 19:25–35.Burchardt, B. 1978. Oxygen isotope paleotemperatures from the Tertiary pe-
riod in the North Sea area. Nature 275:121–123.Cerling, T. E., J. M. Harris, B. J. MacFadden, M. G. Leakey, J. Quade,
V. Eisenmann, and J. R. Ehleringer. 1997. Global vegetation changethrough the Miocene/Pliocene boundary. Nature 389:153–158.
Chaisson, W. P., and A. C. Ravelo. 2000. Pliocene development of the east-west hydrographic gradient in the equatorial Pacific. Paleoceanography15:497–504.
Chollett Ordaz, I. C. 2011. Environmental regimes in the Caribbean andimplications for the dynamics and distribution of its coral reefs. Ph.D.diss. University of Exeter, Devonx, U.K.
Coffroth, M. A., and S. R. Santos. 2005. Genetic diversity of symbiotic di-noflagellates in the genus Symbiodinium. Protist 156:19–34.
Colombo-Pallotta, M. F., A. Rodriguez-Roman, and R. Iglesias-Prieto. 2010.Calcification in bleached and unbleached Montastraea faveolata: eval-uating the role of oxygen and glycerol. Coral Reefs 29:899–907.
Correa, A. M. S., and A. C. Baker. 2009. Understanding diversity in coral-algalsymbiosis: a cluster-based approach to interpreting fine-scale geneticvariation in the genus Symbiodinium. Coral Reefs 28:81–93.
Cowen, R. K., C. B. Paris, and A. Srinivasan. 2006. Scaling of connectivityin marine populations. Science 311:522–527.
Crouch, E. M., C. Heilmann-Clausen, H. Brinkhuis, H. E. G. Morgans, K.M. Rogers, H. Egger, and B. Schmitz. 2001. Global dinoflagellate eventassociated with the late Paleocene thermal maximum. Geology 29:315–318.
de Queiroz, K. 2007. Species concepts and species delimitation. Syst. Biol.56:879–886.
Dieckmann, U., and M. Doebeli. 1999. On the origin of species by sympatricspeciation. Nature 400:354–357.
Donoghue, M. J. 1985. A critique of the biological species concept and rec-ommendations for a phylogenetic alternative. Bryologist 88:172–181.
Dover, G. A. 1986. Molecular drive in multigene families: how biologicalnovelties arise, spread and are assimilated. Trends Genet. 2:159–165.
Drummond, A. J., and A. Rambaut. 2007. BEAST: Bayesian evolutionaryanalysis by sampling trees. BMC Evol. Biol. 7:214.
Evanno, G., S. Regnaut, and J. Goudet. 2005. Detecting the number of clustersof individuals using the software STRUCTURE: a simulation study. Mol.Ecol. 14:2611–2620.
Falkowski, P. G., M. E. Katz, A. H. Knoll, A. Quigg, J. A. Raven, O. Schofield,and F. J. R. Taylor. 2004. The evolution of modern eukaryotic phyto-plankton. Science 305:354–360.
Finney, J. C., D. T. Pettay, E. M. Sampayo, M. E. Warner, H. A. Oxenford,and T. C. LaJeunesse. 2010. The relative significance of host-habitat,depth, and geography on the ecology, endemism, and speciation ofcoral endosymbionts in the genus Symbiodinium. Microb. Ecol. 60:250–263.
Galluzzi, L., A. Penna, E. Bertozzini, M. Vila, E. Garces, and M. Magnani.2004. Development of a real-time PCR assay for rapid detection and
EVOLUTION FEBRUARY 2014 3 6 5

DANIEL J. THORNHILL ET AL.
quantification of Alexandrium minutum (a dinoflagellate). Appl. Envi-ron. Microbiol. 70:1199–1206.
Gavrilets, S., and J. B. Losos. 2009. Adaptive radiation: contrasting theorywith data. Science 323:732–737.
Gernhard, T. 2008. New analytic results for speciation times in neutral models.B. Math. Biol. 4:769–778.
Glor, R. E. 2010. Phylogenetic insights on adaptive radiation. Ann. Rev. Ecol.Evol. Syst. 41:251–270.
Goulet, T. L., and M. A. Coffroth. 2003. Stability of an octocoral-algal sym-biosis over time and space. Mar. Ecol. Prog. Ser. 250:117–124.
Grant, P. R., and B. R. Grant. 2007. How and why species multiply: radiationof Darwin’s finches. Princeton Univ. Press, Princeton, NJ. Pp. 256.
Haq, B. U., J. Hardenbol, and P. R. Vail. 1987. Chronology of fluctuating sealevels since the Triassic. Science 235:1156–1167.
Haug, G. H., and R. Tiedemann. 1998. Effect of the formation of the Isthmusof Panama on Atlantic thermohaline circulation. Nature 393:673–676.
Haug, G. H., R. Tiedemann, R. Zahn, and A. C. Ravelo. 2001. Role of Panamauplift on oceanic freshwater balance. Geology 29:207–210.
Hausdorf, B. 2011. Progress toward a general species concept. Evolution65:923–931.
Hill, M., and A. Hill. 2012. The magnesium inhibition and arrested phago-some hypotheses: new perspectives on the evolution and ecology ofSymbiodinium symbioses. Biol. Rev. 87:804–821.
Howells, E. J., M. J. H. van Oppen, and B. L. Willis. 2009. High geneticdifferentiation and cross-shelf patterns of genetic diversity among GreatBarrier Reef populations of Symbiodinium. Coral Reefs 28:215–225.
Howells, E. J., V. H. Beltran, N. W. Larsen, L. K. Bay, B. L. Willis, and M. J.H. van Oppen. 2012. Coral thermal tolerance shaped by local adaptationof photosymbionts. Nat. Clim. Change 2:116–120.
Huelsenbeck, J. P., and F. Ronquist. 2001. MRBAYES: Bayesian inference ofphylogenetic trees. Bioinformatics 17:754–755.
Hume, B., C. D’Angelo, J. Burt, A. C. Baker, B. Riegl, and J. Wiedenmann.2013. Corals from the Persian/Arabian Gulf as models for thermotolerantreef-builders: prevalence of clade C3 Symbiodinium, host fluorescenceand ex situ temperature tolerance. Mar. Poll. Bull. 72:313–322.
Jackson, J. B. C., and K. G. Johnson. 2000. Life in the last few millionyears. In D. H. Erwin and S. L. Wing, eds. Deep time: paleobiology’spersepective. Paleobiology 26:221–235.
Jackson, J. B. C., A. F. Budd, and J. M. Pandolfi. 1996. The shifting balanceof natural communities? Pp. 89–122 in D. Erwin, D. Jablonski, andJ. Lipps, eds. Evolutionary paleobiology. University of Chicago Press,Chicago.
Janis, C. M. 1993. Tertiary mammal evolution in the context of changingclimates, vegetation and tectonic events. Annu. Rev. Ecol. Syst. 24:467–500.
Jones, A. M., R. Berkelmans, M. J. H. van Oppen, J. C. Mieog, and W.Sinclair. 2008. A community change in the algal endosymbionts of ascleractinian coral following a natural bleaching event: field evidence ofacclimatization. Proc. Biol. Sci. 275:1359–1365.
LaJeunesse, T. C. 2001. Investigating the biodiversity, ecology, and phylogenyof endosymbiotic dinoflagellates in the genus Symbiodinium using theITS region: in search of a “species” level marker. J. Phycol. 37:866–880.
———. 2002. Diversity and community structure of symbiotic dinoflagellatesfrom Caribbean coral reefs. Mar. Biol. 141:387–400.
———. 2005. “Species” radiations of symbiotic dinoflagellates in the Atlanticand Indo-Pacific since the Miocene-Pliocene transition. Mol. Biol. Evol.22:570–581.
LaJeunesse, T. C., and D. J. Thornhill. 2011. Improved resolution of reef-coralendosymbiont (Symbiodinium) species diversity, ecology, and evolutionthrough psbA non-coding region genotyping. PLoS ONE 6:e29013.
LaJeunesse, T. C., W. K. W. Loh, R. van Woesik, O. Hoegh-Guldberg, G.W. Schmidt, and W. K. Fitt. 2003. Low symbiont diversity in southernGreat Barrier Reef corals relative to those of the Caribbean. Limnol.Oceanogr. 48:2046–2054.
LaJeunesse, T. C., D. J. Thornhill, E. F. Cox, F. G. Stanton, W. K. Fitt, andG.W. Schmidt. 2004. High diversity and host specificity observed amongsymbiotic dinoflagellates in reef coral communities from Hawaii. CoralReefs 23:596–603.
LaJeunesse, T. C., R. T. Smith, J. Finney, and H. Oxenford. 2009. Outbreak andpersistence of opportunistic symbiotic dinoflagellates during the 2005Caribbean mass coral ‘bleaching’ event. Proc. Biol. Sci. 276:4139–4148.
LaJeunesse, T. C., D. T. Pettay, E. M. Sampayo, N. Phongsuwan, B. Brown,D. O. Obura, O. Hoegh-Guldberg, and W. K. Fitt. 2010. Long-standingenvironmental conditions, geographic isolation and host-symbiont speci-ficity influence the relative ecological dominance and genetic diversifi-cation of coral endosymbionts in the genus Symbiodinium. J. Biogeogr.37:785–800.
LaJeunesse, T. C., J. E. Parkinson, and J. D. Reimer. 2012. A genetics-baseddescription of Symbiodinium minutum sp. nov. and S. psygmophilum
sp. nov. (Dinophyceae), two dinoflagellates symbiotic with cnidarian. J.Phycol. 48:1380–1391.
Losos, J. B. 2011. Lizards in an evolutionary tree: ecology and adaptiveradiation of Anoles. University of California Press, Berkeley, CA.
Losos, J. B., and R. E. Ricklefs. 2009. Adaptation and diversification onislands. Nature 457:830–836.
Mieog, J. C., M. J. H. van Oppen, N. E. Cantin, W. T. Stam, and J. L. Olsen.2007. Real-time PCR reveals a high incidence of Symbiodinium clade Dat low levels in four scleractinian corals across the Great Barrier Reef:implications for symbiont shuffling. Coral Reefs 26:449–457.
Moore, R. B., K. M. Ferguson, W. K. W. Loh, O. Hoegh-Guldberg, and D. A.Carter. 2003. Highly organized structure in the non-coding region of thepsbA minicircle from clade C Symbiodinium. Int. J. Syst. Evol. Micr.53:1725–1734.
Muscatine, L., L. R. McCloskey, and R. E. Marian. 1981. Estimating the dailycontribution of carbon from zooxanthellae to coral animal respiration.Limnol. Oceanogr. 26:601–611.
Nylander, J. A. A. 2004. MrModeltest v2. Program distributed by the author.Evolutionary Biology Centre, Uppsala University, Sweden.
Pachauri, R. K., and A. Reisinger, eds. 2007. Climate Change 2007: SynthesisReport. Contribution of Working Groups I, II and III to the FourthAssessment Report of the Intergovernmental Panel on Climate ChangeCore Writing Team, IPCC, Geneva, Switzerland.
Palys, T., L. K. Nakamura, and F. M. Cohan. 1997. Discovery and classificationof ecological diversity in the bacterial world: the role of DNA sequencedata. Int. J. Syst. Bacteriol. 47:1145–1156.
Pandolfi, J. M., and J. B. C. Jackson. 2001. Community structure of Pliestocenecoral reefs of Curacao, Netherlands Antilles. Ecol. Monogr. 71:49–67.
Pinzon, J. H. 2011. Phylogenetics, population genetics and ecology to under-stand the evolution of coral-algal mutualisms. Ph.D. diss. PennsylvaniaState University, Pennsylvania.
Peakall, R., and P. E. Smouse. 2006. GENALEX 6: genetic analysis in Excel.Population genetic software for teaching and research. Mol. Ecol. Notes6:288–295.
Pearson, P. N., and M. R. Palmer. 2000. Atmospheric carbon dioxide concen-trations over the past 60 million years. Nature 406:695–699.
Pettay, D. T., D. C. Wham, J. H. Pinzon and T. C. LaJeunesse 2011. Genotypicdiversity and spatial–temporal distribution of Symbiodinium clones inan abundant reef coral. Mol. Ecol. 20:5197–5212.
Pochon, X., J. I. Montoya-Burgos, B. Stadelmann, and J. Palowski. 2006.Molecular phylogeny, evolutionary rates, and divergence timing of the
3 6 6 EVOLUTION FEBRUARY 2014

ADAPTIVE RADIATION OF SYMBIOTIC DINOFLAGELLATES
symbiotic dinoflagellate genus Symbiodinium. Mol. Phyl. Evol. 38:20–30.
Pochon, X., H. M. Putnam, F. Burki, and R. D. Gates. 2012. Identifyingand characterizing alternative molecular markers for the symbiotic andfree-living dinoflagellate genus Symbiodinium. PLoS ONE 7:e29816.
Pritchard, J. K., M. Stephens, and P. Donnelly. 2000. Inference of populationstructure using multilocus genotype data. Genetics 155:945–959.
Rowan, R., and D. A. Powers. 1991. A molecular genetic classification ofzooxanthellae and the evolution of animal-algal symbiosis. Science251:1348–1351.
Ruddiman, W. F., and M. E. Raymo. 1988. Northern Hemisphere climateregimes during the past 3 Ma: possible tectonic connections. Philos.Trans. R. Soc. B 318:411–430.
Saito, K., T. Drgon, J. A. Robledo, D. N. Krupatkina, and G. R. Vasta. 2002.Characterization of the rRNA locus of Pfiesteria piscicida and develop-ment of standard and quantitative PCR-based detection assays targetedto the nontranscribed spacer. Appl. Environ. Microbiol. 68:5394–5407.
Sampayo, E. M., S. Dove, and T. C. LaJeunesse. 2009. Cohesive molecu-lar genetic data delineate species diversity in the dinoflagellate genusSymbiodinium. Mol. Ecol. 18:500–519.
Santos, S. R., and M. A. Coffroth. 2003. Molecular genetic evidence thatdinoflagellates belonging to the genus Symbiodinium Freudenthal arehaploid. Biol. Bull. 241:10–20.
Santos, S. R., T. L. Shearer, A.R. Hannes, and M.A. Coffroth. 2004. Fine-scale diversity and specificity in the most prevalent lineage of symbioticdinoflagellates (Symbiodinium, Dinophyceae) of the Caribbean. Mol.Ecol. 13:459–469.
Schluter, D. 2001. Ecology and the origin of species. Trends Ecol. Evol.16:372–380.
———. 2009. Evidence for ecological speciation and its alternative. Science323:737–741.
Seehausen, O. 2006. African cichlid fish: a model system in adaptive radiationresearch. Proc. Biol. Soc. 273:1987–1998.
Silverstein, R. N., A. M. S. Correa, T. C. LaJeunesse, and A. C. Baker, 2011.Novel algal symbiont (Symbiodinium spp.) diversity in reef corals ofWestern Australia. Mar. Ecol. Prog. Ser. 422:63–75.
Smith, G. P. 1976. Evolution of repeated DNA sequences by unequalcrossover. Science 191:528–535.
Swofford, D. L. 2000. PAUP∗, Phylogenetic analysis using parsimony (∗andother methods), version 4.0b10. Sinauer, Sunderland, MA.
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei, and S. Kumar. 2011.MEGA5: molecular evolutionary genetics analysis using maximum like-lihood, evolutionary distance, and Maximum Parsimony methods. Mol.Biol. Evol. 28:2731–2739.
Taylor, M. S., and M. E. Hellberg. 2003. Genetic evidence for local retentionof pelagic larvae in a Caribbean reef fish. Science 299:107–109.
Thornhill, D. J., T. C. LaJeunesse, D. W. Kemp, W. K. Fitt, and G. W.Schmidt. 2006. Multi-year, seasonal genotypic surveys of coral-algalsymbioses reveal prevalent stability or post-bleaching reversion. Mar.Biol. 148:711–722.
Thornhill, D. J., T. C. LaJeunesse, and S. R. Santos. 2007. Measuring rDNAdiversity in eukaryotic microbial systems: how intragenomic variation,pseudogenes, and PCR artifacts confound biodiversity estimates. Mol.Ecol. 16:5326–5340.
Thornhill, D. J., Y. Xiang, W. K. Fitt, and S. R. Santos. 2009. Reef endemism,host specificity and temporal stability in populations of symbiotic di-noflagellates from two ecologically dominant Caribbean corals. PLoSONE 4:e6262.
Thornhill, D. J., Y. Xiang, D. T. Pettay, M. Zhong, and S. R. Santos. 2013.Population genetic data of a model symbiotic cnidarian system reveal re-markable symbiotic specificity and vectored introductions across oceanbasins. Mol. Ecol. 22:4499–4515.
Valentine, J. W., D. Jablonski, and D. H. Erwin. 1999. Fossils, molecules andembryos: new perspectives on the Cambrian explosion. Development126:851–859.
Veron, J. E. N. 1995. Corals in space and time: the biogeography and evolutionof the Scleractinia. UNSW Press, Sydney, Australia.
Wakeley, J. 2008. Coalescent theory: an introduction. Roberts and CompanyPublishers, Greenwood Village, Colorado.
Weyl, P. K. 1968. The role of the oceans in climate change; a theory of the iceages. Meteorol. Mon. 8:27–62.
Wham, D. C., D. T. Pettay, and T. C. LaJeunesse. 2011. Microsatellite locifor the host-generalist “zooxanthella” Symbiodinium trenchi and otherClade D Symbiodinium. Conserv. Genet. Res. 3:541–544.
Wham, D. C., M. Carmichael, and T. C. LaJeunesse. 2013. Microsatellite locifor Symbiodinium goreaui and other clade C Symbiodinium. Conserv.Genet. Res. doi: 10.1007/s12686-013-0023-5.
Wolfe, J. A. 1978. A paleobotanical interpretation of Tertiary climates in theNorthern Hemisphere. Am. Sci. 66:694–703.
Zachos, J. C., M. Pagani, L. Sloan, E. Thomas, and K. Billups 2001. Trends,rhythms, and aberrations in global climate 65 Ma to present. Science292:686–693.
Zachos, J. C., G. R. Dickens, and R. E. Zeebe. 2008. An early Cenozoicperspective on greenhouse warming and carbon-cycle dynamics. Nature451:279–283.
Associate Editor: J. Wares
Supporting InformationAdditional Supporting Information may be found in the online version of this article at the publisher’s website:
Figure S1. Maximum Parsimony topology based on the psbAncr for haplotypes associated with Siderastrea siderea (ITS2
type C3).Figure S2. Maximum Parsimony topology based on the psbAncr for haplotypes associated with Agaricia and Leptoseris (ITS2 types C3, C3a, C3b, C3s,and C3q).Figure S3. Evanno’s Delta-K showing the change in likelihood between K = 3 and corresponds to populations designated by the two sister lineages (C7and C7a/C12) associated with Orbicella and the Symbiodinium C3 Siderastrea with Siderastrea siderea.Table S1. Symbiodinium samples, their ITS2 designations along with host species, depth of collection, and geographic origin.Table S2. Summary statistics on allele number and evenness among unique multilocus genotypes representing Symbiodinium lineages associated withOrbicella and Siderastrea from the Greater Caribbean of the western Atlantic.
EVOLUTION FEBRUARY 2014 3 6 7

1
Supplemental for Siderastrea siderea associated Symbiodinium.−
PsbAncr haplotypes of Symbiodinium C3 associated with Siderastrea in the Greater Caribbean form a well-supported, monophyletic clade (Fig. S1). Phylogeographic patterns show that some populations are regionally localized. Within this clade there were various locally-endemic sub-clades and haplotypes, including lineages from the Bahamas, Barbados, Brazil and Curacao. A particular haplotype was only found in one sampling location. Figure S1, Maximum Parsimony topology based on the psbAncr for haplotypes associated with Siderastrea siderea (ITS2 type C3). The basal node of this tree connects to the topology presented in Fig. 4 (note «symbol at the node labeled C3 Siderastrea siderea in Fig. 4). Nodal support indicated as posterior probabilities (PP) above 0.85 next to the relevant node or asterisks for a PP = 1.00 (recovered from separate analyses using Mr. Bayes software, which produced a concordant tree topology). Colored symbols indicate the collection location of each haplotype corresponding to Fig. 1. The branch relating Brazil haplotypes with those of the Greater Caribbean is interrupted by slash marks to indicate that this lineage may be polyphyletic. Supplemental for Agaricidae associated Symbiodinium −
Multiple phylogenetically independent, or
paraphyletic, PsbAncr clades were found associated with Caribbean agaricidae: Agaricia and Leptoseris spp. (Figs. 4, S2). Many of these correlated with geography and corresponded to diagnostic intragenomic ITS2 sequence variants common in the genome. The two larger clades are abbreviated in Fig. 4 with a star symbol.
Samples designated as type C3b by ITS2 DGGE fingerprinting (data not shown) occurred at two positions in the phylogeny (designated by the superscripts 1 and 2) and represents the only case were ITS2 and psbA sequences did not correspond. This unusual finding was the only example of a polyphyletic ITS2 type in the psbAncr dataset and possibly resulted from homoplasy at the ITS2 level (or a highly unusual recombinant genotype). The C3b samples belonging to

2
separate groups could be differentiated according to minor differences in banding pattern upon close inspection of their ITS2-DGGE profiles (data not shown).
Figure S2 Legend, Maximum Parsimony topology based on based on the psbAncr for haplotypes associated with Agaricia and Leptoseris (ITS2 types C3, C3a, C3b, C3s, and C3q). Separate lineages insert at various locations in the phylogeny depicted in Figure 4 (note «symbol at the nodes in Fig. 4). Nodal support based on > 0.95 are indicated by asterisks. Colored symbols indicate the collection location of each haplotype corresponding to Fig. 1.
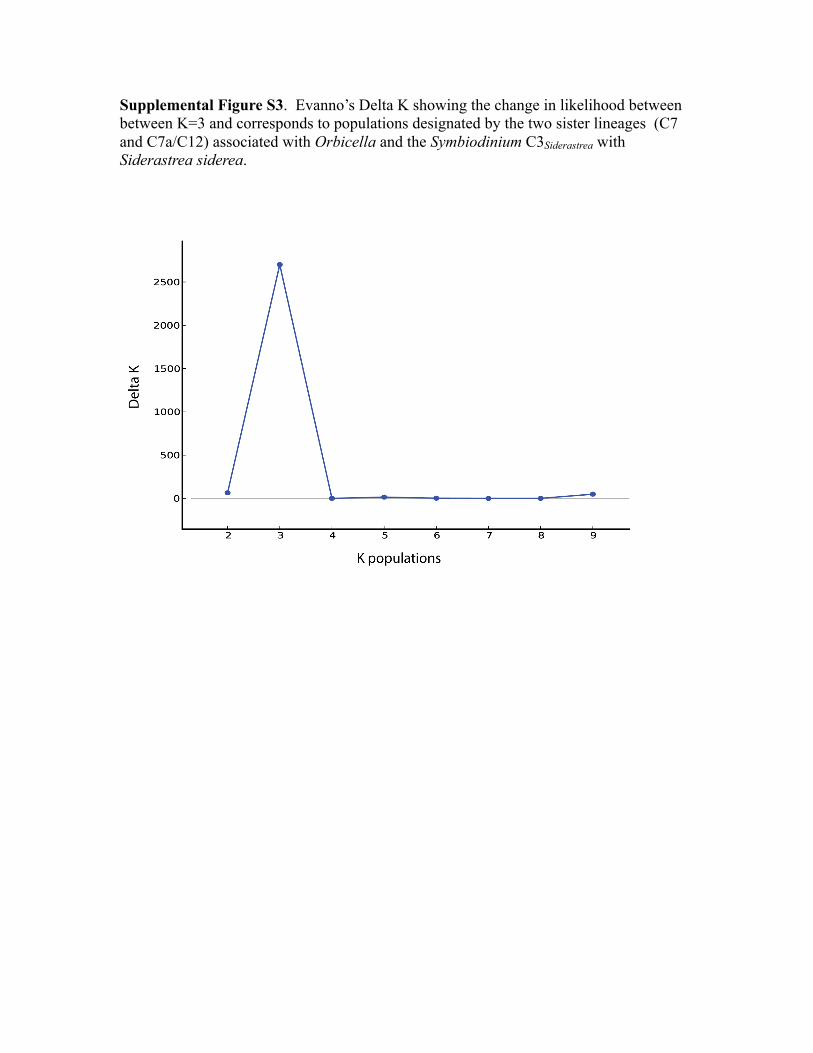
Supplemental Figure S3. Evanno’s Delta K showing the change in likelihood between between K=3 and corresponds to populations designated by the two sister lineages (C7 and C7a/C12) associated with Orbicella and the Symbiodinium C3Siderastrea with Siderastrea siderea.

1
Table S1. Symbiodinium samples, their ITS2 designations along with host species, depth of collection, and geographic origin. GenBank accession numbers for psbAncr sequences are provides and were used to reconstruct the phylogenies in Figures 3–5, S1-S2. Shallow, intermediate and deep refers to collection depths ranging from 1-5, 6-10, and > 10 meters, respectively.
Sample I.D ITS-DGGE type
psbAncr Genbank accession number
Host Genus Host Species Region Location Site Collection Depth
27B_mexico_Montastraea_C7 C7 KF572268 Orbicella Orbicella faveolata Caribbean Mexico Puerto Morelos Shallow 30B_mexico_Montastraea_C7 C7 KF572248 Orbicella Orbicella faveolata Caribbean Mexico Puerto Morelos Shallow A02_10_Acropora_C3 C3 JQ043639 Acropora Acropora cerealis West Pacific GBR Heron Island Shallow A02_105_Platygyra_C3 C3 JQ043635 Platygrya Platygra daedalea West Pacific GBR Heron Island Shallow A02_12_Acropora_C3 C3 JQ043634 Acropora Acropora clathroata West Pacific GBR Heron Island Shallow A02_136_Coralimorph_C1 C1 KF572204 Coralimorpharia Coralimorpharia sp. West Pacific GBR Heron Island Intermediate A02_20_Acropora_C3 C3 JQ043640 Acropora Acropora gemmifera West Pacific GBR Heron Island Intermediate A02_27_Acropora_C3 C3 JQ043641 Acropora Acropora microclados West Pacific GBR Heron Island Shallow A02_30_Acropora_C3 C3 JQ043642 Acropora Acropora nasuta West Pacific GBR Heron Island Shallow A02_34_Acropora_C3 C3 JQ043643 Acropora Acropora palifera West Pacific GBR Heron Island Shallow A02_36_Acropora_C3 C3 KF572372 Acropora Acropora polystoma West Pacific GBR Heron Island Shallow A02_44_Acropora_C3 C3 JQ043644 Acropora Acropora tenuis West Pacific GBR Heron Island Shallow A02_47_Astreopora_C1 C1 KF572203 Astreopora Astreopora myriophthalana West Pacific GBR Heron Island Shallow A02_63_Favia_C3 C3 JQ043636 Favia Favia stellegera West Pacific GBR Heron Island Shallow A02_64_Favia_C3 C3 JQ043637 Favia Favia abdita West Pacific GBR Heron Island Shallow A02_70_Fungia_C1 C1 KF572205 Fungia Fungia sp. West Pacific GBR Heron Island Shallow A02_71_Galaxea_C1 C1 KF572195 Galaxea Galaxea fascicularis West Pacific GBR Heron Island Shallow A02_74_Goniastrea_C3 C3 JQ043638 Goniastrea Goniastrea aspera West Pacific GBR Heron Island Shallow A03_11A_Turbinaria_C3h C3h JQ043628 Turbinaria Turbinaria frondens West Pacific GBR Feather Reef North Deep A03_127_Symphyllia_C40 C40 KF572360 Symphyllia Symphyllia radians West Pacific GBR Feather Reef North Shallow A03_138_Coscinarea_C1 C1 KF572190 Coscinarea Coscinarea columna West Pacific GBR Feather Reef North Shallow A03_198_Tridacna_C1 C1 KF572172 Tridacna Tridacna maxima West Pacific GBR Rib reef south Shallow A03_2_Pachyseris_C3h C3h KF572373 Pachyseris Pachyseris speciosa West Pacific GBR Feather Reef North Deep A03_221_Podabacia_C3h C3h JQ043630 Podabacia Podabacia crustacea West Pacific GBR Rib Reef North Deep A03_238_Acropora_C3K C3K JQ043648 Acropora Acropora nobilis West Pacific GBR Rib Reef North Shallow A03_326_Turbinaria_C40 C40 KF572357 Turbinaria Turbinaria frondens West Pacific GBR Rib Reef South Intermediate A03_331_Turbinaria_C40 C40 KF572361 Turbinaria Turbinaria reniformis West Pacific GBR Rib Reef South Intermediate A03_333_Turbinaria_C40 C40 KF572358 Turbinaria Turbinaria frondens West Pacific GBR Rib Reef South Intermediate A03_337_Turbinaria_C40 C40 KF572359 Turbinaria Turbinaria reniformis West Pacific GBR Rib Reef South Intermediate A03_379_Echinopora C3 KF572363 Echinopora Echinopora lamellosa West Pacific GBR Curacao Island Intermediate A03_44_Pectinia_C3h C3h JQ043631 Pectinia Pectinia paeonia West Pacific GBR Feather Reef North Deep A03_50_Acropora_C1 C1 KF572189 Acropora Acropora tenuis West Pacific GBR Feather Reef North Deep A03_52_Leptoseris_C3h C3h JQ043627 Leptoseris Leptoseris yabei West Pacific GBR Feather Reef North Deep A03_56_Corallomorpharia_C1 C1 KF572173 Discosoma Discosoma rhodostoma West Pacific GBR Feather Reef North Deep

2
A03_6_Fungia_C3h C3h JQ043629 Fungia Fungia sp. West Pacific GBR Feather Reef North Deep A03_64_Fungia_C3h C3h KF572374 Fungia Fungia granulosa (?) West Pacific GBR Feather Reef North Deep A03_71_Pavona_C3h C3h JQ043630 Podabacia Podabacia crustacea West Pacific GBR Rib Reef North Deep A03_81_Acropora_C3K C3K JQ043645 Acropora Acropora humilis West Pacific GBR Feather Reef North Shallow A03_82_Acropora_C3K C3K JQ043646 Acropora Acropora nasuta West Pacific GBR Feather Reef North Shallow A03_83_Acropora_C3K C3K JQ043647 Acropora Acropora florida West Pacific GBR Feather Reef North Shallow A03_98_Galaxea C1 KF572194 Galaxea Galaxea fascicularis West Pacific GBR Feather Reef North Shallow ABA06_26_Siderastrea_C3 C3 KF572331 Siderastrea Siderastrea siderea Caribbean Bahamas Guana Cay, Abaco Shallow ABA06_28_Siderastrea_C3 C3 KF572332 Siderastrea Siderastrea siderea Caribbean Bahamas Guana Cay, Abaco Shallow B701_NNP1_Siderastrea_C3 C3 KF572319 Siderastrea Siderastrea siderea Caribbean Bahamas Lee Stocking Shallow B704_NNP4_Siderastrea_C3 C3 KF572318 Siderastrea Siderastrea siderea Caribbean Bahamas Lee Stocking Shallow B705_NNP5_Siderastrea_C3 C3 KF572317 Siderastrea Siderastrea siderea Caribbean Bahamas Lee Stocking Shallow Bah_428_M_faveolata_C12 C12 KF572239 Orbicella Orbicella faveolata Caribbean Bahamas Lee Stocking Deep Bah_441_M_franksi_C12 C12 KF572238 Orbicella Orbicella franksi Caribbean Bahamas Lee Stocking Deep Bah_442_M_franksi_C12 B1, C12 KF572240 Orbicella Orbicella franksi Caribbean Bahamas Lee Stocking Deep Bah_725_Montastraea_C12 C12 KF572237 Orbicella Orbicella franksi Caribbean Bahamas Lee Stocking Deep Bah01_10_Agaricia_C3 C3 KF572378 Agaricia Agaricia tenuifolia Caribbean Bahamas Lee Stocking Shallow Bah01_08_Agaricia_C3b C3b KF572385 Agaricia Agaricia agaricites Caribbean Bahamas Lee Stocking Shallow Bah02_10_C3_Leptoseris_C3 C3 KF572292 Leptoseris Leptoseries cuculata Caribbean Bahamas Lee Stocking Deep Bah02_43_C3_Leptoseris_C3 C3 KF572294 Leptoseris Leptoseries cuculata Caribbean Bahamas Lee Stocking Deep Bah02_44_C3_Leptoseris_C3 C3 KF572295 Leptoseris Leptoseries cuculata Caribbean Bahamas Lee Stocking Deep Bah02_62_Agaricia_C3 C3 KF572379 Agaricia Agaricia tenuifolia Caribbean Bahamas Lee Stocking Shallow Bah02_77_Siderastrea_C3 C3 KF572301 Siderastrea Siderastrea siderea Caribbean Bahamas Lee Stocking Shallow Baja04_124_Porites_panamensis C1 KF572208 Porites Porites panamensis Eastern Pacific Gulf of California La Paz Shallow Baja04_135_Psammocora_C1bf C1b-f KF572213 Psammocora Psammocora profundacella Eastern Pacific Gulf of California La Paz Intermediate Baja04_14_Porites_panamensis C1 KF572210 Porites Porites panamensis Eastern Pacific Gulf of California La Paz Shallow Baja04_143_Psammocora_C1bf C1b-f KF572214 Psammocora Psammocora superficialis Eastern Pacific Gulf of California La Paz Intermediate Baja04_30_Porites_panamensis C1 KF572209 Porites Porites panamensis Eastern Pacific Gulf of California La Paz Shallow Baja04_34_Porites_panamensis C1 KF572207 Porites Porites panamensis Eastern Pacific Gulf of California La Paz Shallow Bar05_100_Diploria_C3 C3 KF572356 Diploria Diploria strigosa Caribbean Barbados Atlantis Bank Reef Deep Bar05_102_Erythropodium_C3 C3 KF572288 Erythropodium Erythropodium caribaeorum Caribbean Barbados Atlantis Bank Reef Deep Bar05_104_Montastraea_C7a C7a KF572222 Orbicella Orbicella faveolata Caribbean Barbados Atlantis Bank Reef Deep Bar05_113_Agaricia_C3 C3 KF572413 Agaricia Agaricia lamarcki Caribbean Barbados Atlantis Bank Reef Deep Bar05_114_Leptoseris C3 KF572293 Leptoseris Leptoseris cucullata Caribbean Barbados Atlantis Bank Reef Deep Bar05_119_Agaricea_C3b C3b KF572337 Agaricia Agaricia agaricites Caribbean Barbados Atlantis Bank Reef Deep Bar05_133_Montastraea_C7a C7a KF572223 Orbicella Orbicella faveolata Caribbean Barbados Atlantis Bank Reef Deep Bar05_134_Montastraea_C7a C7a KF572224 Orbicella Orbicella faveolata Caribbean Barbados Atlantis Bank Reef Deep Bar05_135_Montastraea_C7a C7a KF572225 Orbicella Orbicella faveolata Caribbean Barbados Atlantis Bank Reef Deep Bar05_45_Palythoa_C1 C1 KF572179 Palythoa Palythoa caribaeorum Caribbean Barbados Bachelors Hall Shallow Bar05_75_Montastraea_C7a C7a KF572226 Orbicella Orbicella franksi Caribbean Barbados Atlantis Bank Reef Deep Bar05_80_Montastraea_C7a C7a KF572227 Orbicella Orbicella annularis Caribbean Barbados Atlantis Bank Reef Deep Bar05_84_Palythoa_C1 C1 KF572182 Palythoa Palythoa caribaeorum Caribbean Barbados Atlantis Bank Reef Deep

3
Bar05_89_Agaricea_C3b C3b KF572336 Agaricia Agaricia agaricites Caribbean Barbados Atlantis Bank Reef Deep Bar05_91_Stephanocoenia_C3 C3 KF572414 Stephanocoenia Stephanocoenia intersepta Caribbean Barbados Atlantis Bank Reef Deep Bar07_03_Siderastrea_C3 C3 KF572303 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Bar07_10_Siderastrea_C3 C3 KF572302 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Bar07_130_Agaricia_Cq Cq KF572338 Agaricia Agaricia humilis Caribbean Barbados Bachelors Hall Shallow Bar07_131_Agaricia_Cq Cq KF572339 Agaricia Agaricia humilis Caribbean Barbados Bachelors Hall Shallow Bar07_132_Agaricia_Cq Cq KF572340 Agaricia Agaricia humilis Caribbean Barbados Bachelors Hall Shallow Bar07_133_Agaricia_Cq Cq KF572341 Agaricia Agaricia humilis Caribbean Barbados Bachelors Hall Shallow BAR07_20_Siderastrea_C3 C3 KF572305 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Bar07_23_Siderastrea_C3 C3 KF572304 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Barbados_AB27_sideratrea_C3 C3 KF572307 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Barbados_AB28_Siderastrea_C3 C3 KF572306 Siderastrea Siderastrea siderea Caribbean Barbados Atlantis Bank Reef Deep Barbados_AT16_Porites_C80 C80 KF572188 Porites Porites astreoides Caribbean Barbados Atlantis Bank Reef Deep Barbados_BR16_Scolymia_C3 C3 KF572285 Scolymia Scolymia cubensis Caribbean Barbados Batt's Rock Intermediate Barbados_R14_Agaricea_C3b C3b KF572388 Agaricia Agaricia agaricites Caribbean Barbados Reef Balls Deep Barbados_RB31_Agaricea_C3b C3b KF572387 Agaricia Agaricia agaricites Caribbean Barbados Reef Balls Deep Barbados_RB9_Agaricea_C3b C3b KF572386 Agaricia Agaricia humilis Caribbean Barbados Reef Balls Deep BB07_190_C1bf C1bf KF572215 Psammocora Psammocora stelata Eastern Pacific Banderas Bay Punta Minta Shallow BB07_201_C1bf C1bf KF572216 Psammocora Psammocora stelata Eastern Pacific Banderas Bay Punta Minta Shallow BB07_240_Porites_panamensis C1v KF572212 Porites Porites panamensis Eastern Pacific Banderas Bay Pocitos Shallow BB07_250_Porites_panamensis C1v KF572211 Porites Porites panamensis Eastern Pacific Banderas Bay Pocitos Shallow Be02_110_Leptoseris_C3 C3 KF572405 Leptoseris Leptoseris cucullata Caribbean Belize Carrie Bow Deep Be02_111_Montastraea_C7 C7 KF572254 Orbicella Orbicella faveolata Caribbean Belize Carrie Bow Deep Be02_116_C3_Erythropodium C3 KF572289 Erythropodium Erythropodium caribaeorum Caribbean Belize Carrie Bow Deep Be02_118_Lebrunia C1 KF572164 Lebrunia Lebrunia danae Caribbean Belize Carrie Bow Deep BE02_135_Bartholomea_C1 C1 KF572193 Bartholomea Bartholomea annulata Caribbean Belize Carrie Bow Shallow Be02_139_Bartholomea C1 KF572163 Bartholomea Bartholomea annulata Caribbean Belize Carrie Bow Shallow Be02_170_Stephanocoenia_C3 C3 KF572344 Stephanocoenia Stephanocoenia intercepta Caribbean Belize Carrie Bow Deep Be02_171_Stephanocoenia_C3 C3 KF572345 Stephanocoenia Stephanocoenia intercepta Caribbean Belize Carrie Bow Deep Be02_178_Ricordia_C3 C3 KF572286 Ricordia Ricordia florida Caribbean Belize Carrie Bow Deep Be02_185_Protopalythoa C1 KF572155 Protopalythoa Protopalythoa grandis Caribbean Belize Carrie Bow Shallow Be02_187_C3_Erythropodium C3 KF572290 Erythropodium Erythropodium caribaeorum Caribbean Belize Carrie Bow Shallow Be02_192_Mreesi_C3c C3c KF572299 Mycetophyllia Mycetophyllia reesi Caribbean Belize Carrie Bow Deep Be02_193_Mreesi_C3c C3c KF572300 Mycetophyllia Mycetophyllia reesi Caribbean Belize Carrie Bow Deep Be02_25_Agaricia_C3a C3a KF572402 Agaricia Agaricia tenuifolia Caribbean Belize Carrie Bow Shallow Be02_26_Agaricia_C3a C3a KF572400 Agaricia Agaricia tenuifolia Caribbean Belize Carrie Bow Shallow Be02_29_Palythoa C1 KF572166 Palythoa Palythoa caribbeorum Caribbean Belize Carrie Bow Shallow Be02_30_Lebrunia C1 KF572165 Lebrunia Lebrunia danae Caribbean Belize Carrie Bow Shallow Be02_36_Stephanocoenia_C3 C3 KF572346 Stephanocoenia Stephanocoenia intercepta Caribbean Belize Carrie Bow Deep Be02_51_Leptoseris_C3 C3 KF572406 Leptoseris Leptoseris cucullata Caribbean Belize Carrie Bow Intermediate Be02_61_Agaricia_C3a C3a KF572401 Agaricia Agaricia agaricites Caribbean Belize Carrie Bow Intermediate Be02_74_Agaricia_C3a C3a KF572403 Agaricia Agaricia tenuifolia Caribbean Belize Carrie Bow Intermediate
4
Be02_81_Agaricia_C3 C3 KF572404 Agaricia Agaricia agaricites Caribbean Belize Carrie Bow Intermediate Be02_90_Palythoa_C1 C1 KF572180 Palythoa Palythoa caribbeorum Caribbean Belize Carrie Bow Intermediate Be02_91_Agaricia_C3 C3 KF572342 Agaricia Agaricia agaricites Caribbean Belize Carrie Bow Deep Be02_97_Agaricia_C3 C3 KF572343 Agaricia Agaricia agaricites Caribbean Belize Carrie Bow Deep Be02_98_Porites_C1a_j C1aj KF572187 Porites Porites astreoides Caribbean Belize Carrie Bow Deep Be02_99_Stephanocoenia_C3 C3 KF572347 Stephanocoenia Stephanocoenia intersepta Caribbean Belize Carrie Bow Deep Be03_21_Stephanocoenia_C16 C16 KF572350 Stephanocoenia Stephanocoenia intersepta Caribbean Belize Carrie Bow Intermediate Be03_22_Stephanocoenia_C16 C16 KF572351 Stephanocoenia Stephanocoenia intersepta Caribbean Belize Carrie Bow Intermediate Be03_25_Stephanocoenia_C16 C16 KF572352 Stephanocoenia Stephanocoenia intersepta Caribbean Belize Carrie Bow Intermediate Belize2_483_M_faveolata_C7 C7 KF572270 Orbicella Orbicella faveolata Caribbean Belize Carrie Bow Deep Belize2_487M_faveolata_C7 C7 KF572271 Orbicella Orbicella faveolata Caribbean Belize Carrie Bow Deep Belize3_224_M_faveolata_C7 C7 KF572269 Orbicella Orbicella faveolata Caribbean Belize Carrie Bow Deep Belize3_303M_faveolata_C7 C7 KF572247 Orbicella Orbicella faveolata Caribbean Belize Carrie Bow Deep CB13_02_Belize_Siderastrea_C3 C3 KF572316 Siderastrea Siderastrea siderea Caribbean Belize Carrie Bow Intermediate CB14_02_Belize_Siderastrea_C3 C3 KF572313 Siderastrea Siderastrea siderea Caribbean Belize Carrie Bow Intermediate CB32_02_Belize_Siderastrea_C3 C3 KF572314 Siderastrea Siderastrea siderea Caribbean Belize Carrie Bow Intermediate CB33_02_Belize_Siderastrea_C3 C3 KF572315 Siderastrea Siderastrea siderea Caribbean Belize Carrie Bow Intermediate Chagos_Isopora21_C40 C40 KF572370 Isopora Isopora palifera Indian Ocean Chagos Archipelago Shallow Culture_113 C1 KF572161 N.A. N.A. N.A. N.A. N.A. N.A. Culture_152 C1 KF572162 N.A. N.A. N.A. N.A. N.A. N.A. Cur09_106_Agaricea_C C3 KF572397 Agaricia Agaricia tenuifolia Caribbean Curacao Slangenbaai Intermediate Cur09_107A_Siderastrea_C3 C3 KF572320 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_109_Agaricea_C C3 KF572395 Agaricia Agaricia tenuifolia Caribbean Curacao Slangenbaai Intermediate Cur09_111_Agaricea_C C3 KF572396 Agaricia Agaricia tenuifolia Caribbean Curacao Slangenbaai Intermediate Cur09_114_Siderastrea_C3 C3 KF572322 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_23A_Mycetophyllia C3 KF572423 Mycetophyllia Mycetophyllia sp. Caribbean Curacao Slangenbaai Deep Cur09_25_Scolymia C3 KF572298 Scolymia Scolymia cubensis Caribbean Curacao Slangenbaai Deep Cur09_26_Agaricia C3 KF572409 Agaricia Agaricia graheami Caribbean Curacao Slangenbaai Deep Cur09_27A_Siderastrea_C3 C3 KF572323 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_27C_Siderastrea_C3 C3 KF572324 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_28_Agaricia C3 KF572408 Agaricia Agaricia graheami Caribbean Curacao Slangenbaai Deep Cur09_31A_Siderastrea_C3 C3 KF572325 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_31B_Siderastrea_C3 C3 KF572326 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_31C_Siderastrea_C3 C3 KF572327 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_32_Mycetophyllia C3 KF572424 Mycetophyllia Mycetophyllia sp. Caribbean Curacao Slangenbaai Deep Cur09_34A_Mycetophyllia_C3 C3 KF572354 Mycetophyllia Mycetophyllia sp. Caribbean Curacao Slangenbaai Deep Cur09_40C_Mycetophyllia_C3 C3 KF572355 Mycetophyllia Mycetophyllia sp. Caribbean Curacao Slangenbaai Deep Cur09_45A_Siderastrea_C3 C3 KF572328 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_45B_Siderastrea_C3 C3 KF572329 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_45C_Siderastrea_C3 C3 KF572330 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_47B_Siderastrea_C3 C3 KF572321 Siderastrea Siderastrea siderea Caribbean Curacao Slangenbaai Intermediate Cur09_93_Agaricia_C3 C3 KF572398 Agaricia Agaricia tenuifolia Caribbean Curacao Slangenbaai Intermediate

5
Cur11_20_Montastraea C7a KF572242 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_22_Montastraea C7a KF572243 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_23_Montastraea C7a KF572246 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_25_Montastraea C7a KF572241 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_27a_Montastraea C7a KF572244 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_27b_Montastraea C7a KF572245 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_44_Montastraea C7a KF572233 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_46_Montastraea C7a KF572234 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate Cur11_47_Montastraea C7a KF572232 Orbicella Orbicella faveaolata Caribbean Curacao Intermediate CVX09_01_Palythoa_C1 C1 KF572184 Palythoa Palythoa caribaeorum East Atlantic Cape Verde Intermediate F1362_AG_Mfranksi2_C3 C3 KF572274 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep F1363_AG_M_franksi_C3 B1, C3 KF572276 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep F1377_AG_Mfranksi2_C3 C3 KF572275 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep F1378_AG_Mfranksi3_C3 C3 KF572272 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep F1379_AG_Mfranksi4_C3 C3 KF572273 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep F1380_AG_M_franksi_C3 C3 KF572277 Orbicella Orbicella franksi Caribbean Florida Keys Alligator Reef Deep FGB_70_Siderastrea C1 KF572160 Siderastrea Siderastrea siderea Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_17_Mussa_C3 C3 KF572281 Mussa Mussa angulosa Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_18_Mussa_C3 C3 KF572280 Mussa Mussa angulosa Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_30_C3_Leptoseris_C3 C3 KF572335 Leptoseris Leptoseris cucullata Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_31_C3_Leptoseris_C3 C3 KF572334 Leptoseris Leptoseris cucullata Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_32_C3_Leptoseris_C3 C3 KF572333 Leptoseris Leptoseris cucullata Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_34_Siderastrea C1 KF572157 Siderastrea Siderastrea siderea Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_35_Agaricia_C3s C3s KF572384 Agaricia Agaricia agaricites Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_39_Stephanocoenia_C87 C87 KF572416 Stephanocoenia Stephanocoenia intersepta Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_40_Stephanocoenia_C87 C87 KF572417 Stephanocoenia Stephanocoenia intersepta Caribbean Flower Garden Banks West Bank-Bouy Deep FGB05_53_Agaricia_C3s C3s KF572383 Agaricia Agaricia agaricites Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_54_Agaricia_C3s C3s KF572382 Agaricia Agaricia agaricites Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_55_Agaricia_C3s C3s KF572381 Agaricia Agaricia agaricites Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_64a_Mussa_C3 C3 KF572282 Mussa Mussa angulosa Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_65_Mussa_C3 C3 KF572279 Mussa Mussa angulosa Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_66_Mussa_C3 C3 KF572278 Mussa Mussa angulosa Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_67_Scolemia_C3 C3 KF572297 Scolemia Scolemia cubensis Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_69_Siderastrea C1 KF572158 Siderastrea Sideratrea siderea Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_76_Scolemia_C3 C3 KF572296 Scolemia Scolemia cubensis Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_78_Stephanocoenia_C87 C87 KF572418 Stephanocoenia Stephanocoenia intersepta Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_81_Agaricia_C3s C3s KF572380 Agaricia Agaricia agaricites Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_77_Stephanocoenia_C87 C87 KF572415 Stephanocoenia Stephanocoenia intersepta Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_89_Siderastrea C1 KF572156 Siderastrea Siderastrea siderea Caribbean Flower Garden Banks East Bank-Bouy Deep FGB05_90_Acropora C1 KF572159 Acropora Acropora palmata Caribbean Flower Garden Banks Deep FL01_31_C3_Erythropodium_C3 C3 KF572291 Erythropodium Erythropodium caribaeorum Caribbean Florida Keys Key Largo Intermediate FL01_35_Palythoa C1 KF572178 Palythoa Palythoa caribaeorum Caribbean Florida Keys Key Largo Shallow
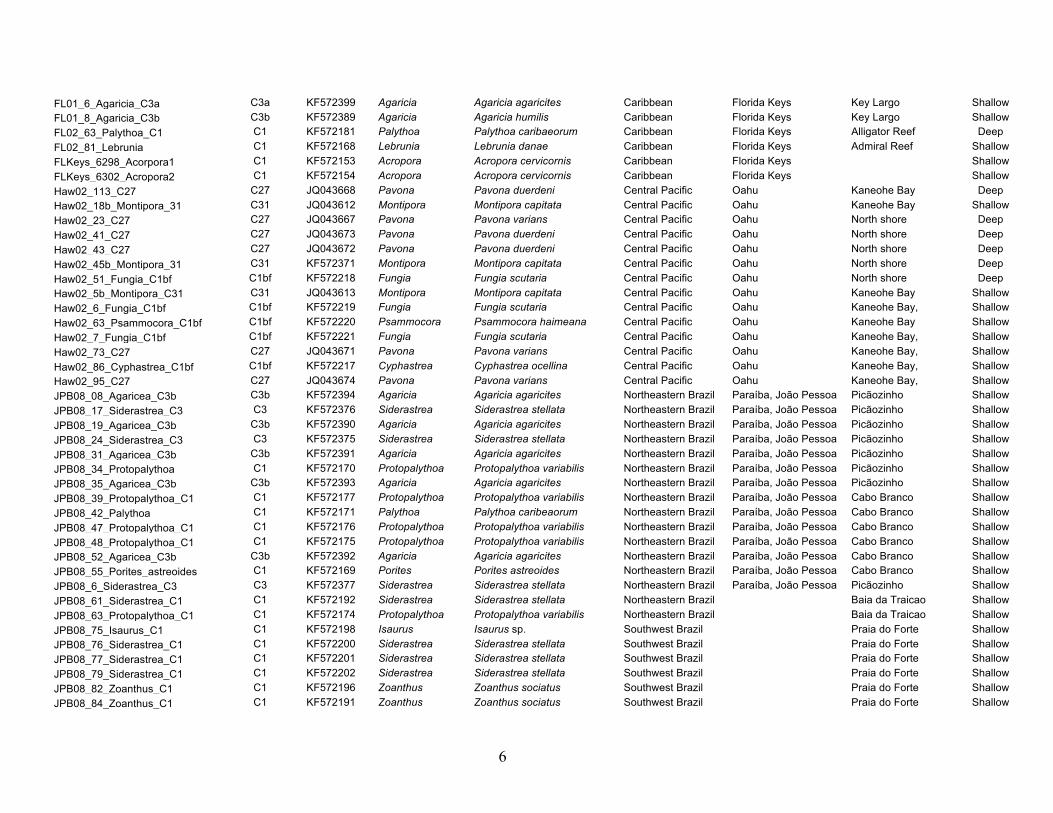
6
FL01_6_Agaricia_C3a C3a KF572399 Agaricia Agaricia agaricites Caribbean Florida Keys Key Largo Shallow FL01_8_Agaricia_C3b C3b KF572389 Agaricia Agaricia humilis Caribbean Florida Keys Key Largo Shallow FL02_63_Palythoa_C1 C1 KF572181 Palythoa Palythoa caribaeorum Caribbean Florida Keys Alligator Reef Deep FL02_81_Lebrunia C1 KF572168 Lebrunia Lebrunia danae Caribbean Florida Keys Admiral Reef Shallow FLKeys_6298_Acorpora1 C1 KF572153 Acropora Acropora cervicornis Caribbean Florida Keys Shallow FLKeys_6302_Acropora2 C1 KF572154 Acropora Acropora cervicornis Caribbean Florida Keys Shallow Haw02_113_C27 C27 JQ043668 Pavona Pavona duerdeni Central Pacific Oahu Kaneohe Bay Deep Haw02_18b_Montipora_31 C31 JQ043612 Montipora Montipora capitata Central Pacific Oahu Kaneohe Bay Shallow Haw02_23_C27 C27 JQ043667 Pavona Pavona varians Central Pacific Oahu North shore Deep Haw02_41_C27 C27 JQ043673 Pavona Pavona duerdeni Central Pacific Oahu North shore Deep Haw02_43_C27 C27 JQ043672 Pavona Pavona duerdeni Central Pacific Oahu North shore Deep Haw02_45b_Montipora_31 C31 KF572371 Montipora Montipora capitata Central Pacific Oahu North shore Deep Haw02_51_Fungia_C1bf C1bf KF572218 Fungia Fungia scutaria Central Pacific Oahu North shore Deep Haw02_5b_Montipora_C31 C31 JQ043613 Montipora Montipora capitata Central Pacific Oahu Kaneohe Bay Shallow Haw02_6_Fungia_C1bf C1bf KF572219 Fungia Fungia scutaria Central Pacific Oahu Kaneohe Bay, Shallow Haw02_63_Psammocora_C1bf C1bf KF572220 Psammocora Psammocora haimeana Central Pacific Oahu Kaneohe Bay Shallow Haw02_7_Fungia_C1bf C1bf KF572221 Fungia Fungia scutaria Central Pacific Oahu Kaneohe Bay, Shallow Haw02_73_C27 C27 JQ043671 Pavona Pavona varians Central Pacific Oahu Kaneohe Bay, Shallow Haw02_86_Cyphastrea_C1bf C1bf KF572217 Cyphastrea Cyphastrea ocellina Central Pacific Oahu Kaneohe Bay, Shallow Haw02_95_C27 C27 JQ043674 Pavona Pavona varians Central Pacific Oahu Kaneohe Bay, Shallow JPB08_08_Agaricea_C3b C3b KF572394 Agaricia Agaricia agaricites Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_17_Siderastrea_C3 C3 KF572376 Siderastrea Siderastrea stellata Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_19_Agaricea_C3b C3b KF572390 Agaricia Agaricia agaricites Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_24_Siderastrea_C3 C3 KF572375 Siderastrea Siderastrea stellata Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_31_Agaricea_C3b C3b KF572391 Agaricia Agaricia agaricites Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_34_Protopalythoa C1 KF572170 Protopalythoa Protopalythoa variabilis Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_35_Agaricea_C3b C3b KF572393 Agaricia Agaricia agaricites Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_39_Protopalythoa_C1 C1 KF572177 Protopalythoa Protopalythoa variabilis Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_42_Palythoa C1 KF572171 Palythoa Palythoa caribeaorum Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_47_Protopalythoa_C1 C1 KF572176 Protopalythoa Protopalythoa variabilis Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_48_Protopalythoa_C1 C1 KF572175 Protopalythoa Protopalythoa variabilis Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_52_Agaricea_C3b C3b KF572392 Agaricia Agaricia agaricites Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_55_Porites_astreoides C1 KF572169 Porites Porites astreoides Northeastern Brazil Paraíba, João Pessoa Cabo Branco Shallow JPB08_6_Siderastrea_C3 C3 KF572377 Siderastrea Siderastrea stellata Northeastern Brazil Paraíba, João Pessoa Picãozinho Shallow JPB08_61_Siderastrea_C1 C1 KF572192 Siderastrea Siderastrea stellata Northeastern Brazil Baia da Traicao Shallow JPB08_63_Protopalythoa_C1 C1 KF572174 Protopalythoa Protopalythoa variabilis Northeastern Brazil Baia da Traicao Shallow JPB08_75_Isaurus_C1 C1 KF572198 Isaurus Isaurus sp. Southwest Brazil Praia do Forte Shallow JPB08_76_Siderastrea_C1 C1 KF572200 Siderastrea Siderastrea stellata Southwest Brazil Praia do Forte Shallow JPB08_77_Siderastrea_C1 C1 KF572201 Siderastrea Siderastrea stellata Southwest Brazil Praia do Forte Shallow JPB08_79_Siderastrea_C1 C1 KF572202 Siderastrea Siderastrea stellata Southwest Brazil Praia do Forte Shallow JPB08_82_Zoanthus_C1 C1 KF572196 Zoanthus Zoanthus sociatus Southwest Brazil Praia do Forte Shallow JPB08_84_Zoanthus_C1 C1 KF572191 Zoanthus Zoanthus sociatus Southwest Brazil Praia do Forte Shallow
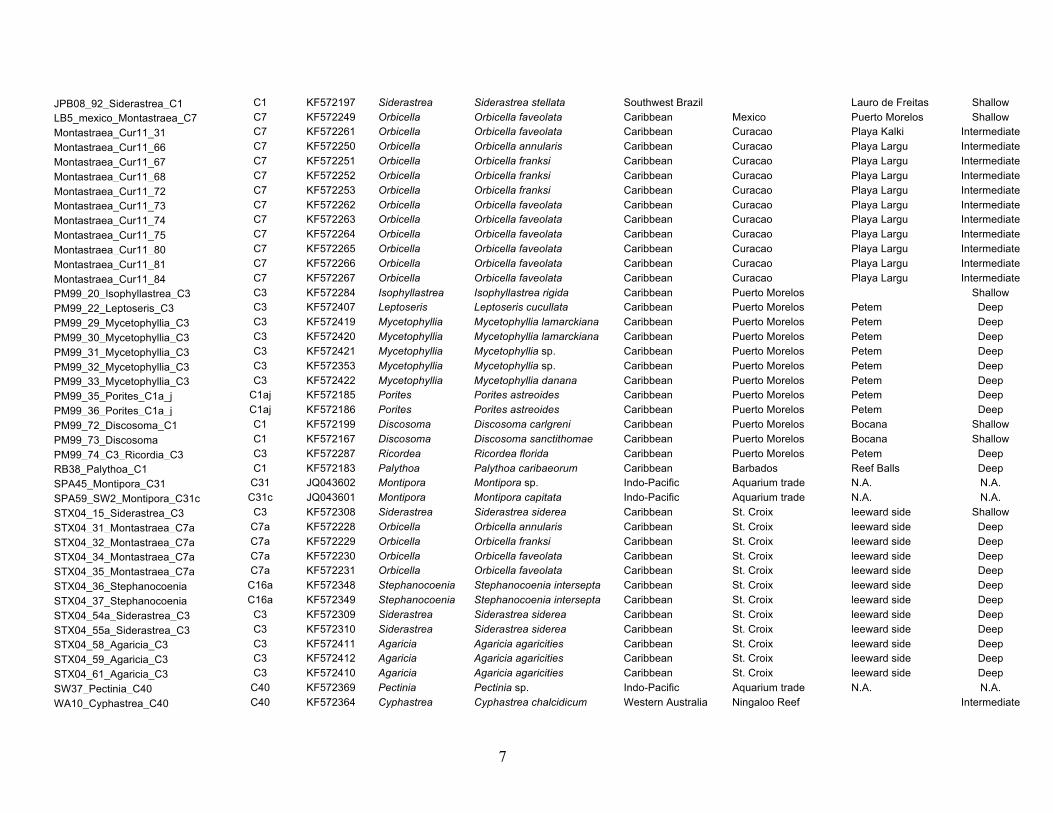
7
JPB08_92_Siderastrea_C1 C1 KF572197 Siderastrea Siderastrea stellata Southwest Brazil Lauro de Freitas Shallow LB5_mexico_Montastraea_C7 C7 KF572249 Orbicella Orbicella faveolata Caribbean Mexico Puerto Morelos Shallow Montastraea_Cur11_31 C7 KF572261 Orbicella Orbicella faveolata Caribbean Curacao Playa Kalki Intermediate Montastraea_Cur11_66 C7 KF572250 Orbicella Orbicella annularis Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_67 C7 KF572251 Orbicella Orbicella franksi Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_68 C7 KF572252 Orbicella Orbicella franksi Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_72 C7 KF572253 Orbicella Orbicella franksi Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_73 C7 KF572262 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_74 C7 KF572263 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_75 C7 KF572264 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_80 C7 KF572265 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_81 C7 KF572266 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate Montastraea_Cur11_84 C7 KF572267 Orbicella Orbicella faveolata Caribbean Curacao Playa Largu Intermediate PM99_20_Isophyllastrea_C3 C3 KF572284 Isophyllastrea Isophyllastrea rigida Caribbean Puerto Morelos Shallow PM99_22_Leptoseris_C3 C3 KF572407 Leptoseris Leptoseris cucullata Caribbean Puerto Morelos Petem Deep PM99_29_Mycetophyllia_C3 C3 KF572419 Mycetophyllia Mycetophyllia lamarckiana Caribbean Puerto Morelos Petem Deep PM99_30_Mycetophyllia_C3 C3 KF572420 Mycetophyllia Mycetophyllia lamarckiana Caribbean Puerto Morelos Petem Deep PM99_31_Mycetophyllia_C3 C3 KF572421 Mycetophyllia Mycetophyllia sp. Caribbean Puerto Morelos Petem Deep PM99_32_Mycetophyllia_C3 C3 KF572353 Mycetophyllia Mycetophyllia sp. Caribbean Puerto Morelos Petem Deep PM99_33_Mycetophyllia_C3 C3 KF572422 Mycetophyllia Mycetophyllia danana Caribbean Puerto Morelos Petem Deep PM99_35_Porites_C1a_j C1aj KF572185 Porites Porites astreoides Caribbean Puerto Morelos Petem Deep PM99_36_Porites_C1a_j C1aj KF572186 Porites Porites astreoides Caribbean Puerto Morelos Petem Deep PM99_72_Discosoma_C1 C1 KF572199 Discosoma Discosoma carlgreni Caribbean Puerto Morelos Bocana Shallow PM99_73_Discosoma C1 KF572167 Discosoma Discosoma sanctithomae Caribbean Puerto Morelos Bocana Shallow PM99_74_C3_Ricordia_C3 C3 KF572287 Ricordea Ricordea florida Caribbean Puerto Morelos Petem Deep RB38_Palythoa_C1 C1 KF572183 Palythoa Palythoa caribaeorum Caribbean Barbados Reef Balls Deep SPA45_Montipora_C31 C31 JQ043602 Montipora Montipora sp. Indo-Pacific Aquarium trade N.A. N.A. SPA59_SW2_Montipora_C31c C31c JQ043601 Montipora Montipora capitata Indo-Pacific Aquarium trade N.A. N.A. STX04_15_Siderastrea_C3 C3 KF572308 Siderastrea Siderastrea siderea Caribbean St. Croix leeward side Shallow STX04_31_Montastraea_C7a C7a KF572228 Orbicella Orbicella annularis Caribbean St. Croix leeward side Deep STX04_32_Montastraea_C7a C7a KF572229 Orbicella Orbicella franksi Caribbean St. Croix leeward side Deep STX04_34_Montastraea_C7a C7a KF572230 Orbicella Orbicella faveolata Caribbean St. Croix leeward side Deep STX04_35_Montastraea_C7a C7a KF572231 Orbicella Orbicella faveolata Caribbean St. Croix leeward side Deep STX04_36_Stephanocoenia C16a KF572348 Stephanocoenia Stephanocoenia intersepta Caribbean St. Croix leeward side Deep STX04_37_Stephanocoenia C16a KF572349 Stephanocoenia Stephanocoenia intersepta Caribbean St. Croix leeward side Deep STX04_54a_Siderastrea_C3 C3 KF572309 Siderastrea Siderastrea siderea Caribbean St. Croix leeward side Deep STX04_55a_Siderastrea_C3 C3 KF572310 Siderastrea Siderastrea siderea Caribbean St. Croix leeward side Deep STX04_58_Agaricia_C3 C3 KF572411 Agaricia Agaricia agaricities Caribbean St. Croix leeward side Deep STX04_59_Agaricia_C3 C3 KF572412 Agaricia Agaricia agaricities Caribbean St. Croix leeward side Deep STX04_61_Agaricia_C3 C3 KF572410 Agaricia Agaricia agaricities Caribbean St. Croix leeward side Deep SW37_Pectinia_C40 C40 KF572369 Pectinia Pectinia sp. Indo-Pacific Aquarium trade N.A. N.A. WA10_Cyphastrea_C40 C40 KF572364 Cyphastrea Cyphastrea chalcidicum Western Australia Ningaloo Reef Intermediate

8
WA112_Hydnophora_C40 C40 KF572366 Hydnophora Hydnophora rigida Western Australia Ningaloo Reef Intermediate WA29_Echinophora_C40 C40 KF572367 Echinopora Echinopora lamellosa Western Australia Ningaloo Reef Intermediate WA908_Acropora_C40 C40 KF572362 Acropora Acropora latistella Western Australia Dampier Intermediate WA93_Hydnophora_C40 C40 KF572368 Hydnophora Hydnophora rigida Western Australia Ningaloo Reef Intermediate WA99_Cyphastrea_C40 C40 KF572365 Cyphastrea Cyphastrea chalcidicum Western Australia Ningaloo Reef Intermediate YBD3_Montastraea_C7 C7 KF572259 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate YBD4_Montastraea_C7 C7 KF572260 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate YBD59_Montastraea_C7 C7 KF572258 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate 98_78_Montastraea_C7 C7 KF572255 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate 98_79_Montastraea_C7 C7 KF572256 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate 98_80_Montastraea_C7 C7 KF572257 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate 98_83_Montastraea_C7c C7c KF572236 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate 98_84_Montastraea_C7c C7c KF572235 Orbicella Orbicella faveolata Caribbean Panama San Blas Intermediate Zam03_10M_28_Alveopora_C27 C27 JQ043670 Pachyseris Pachyseris speciosa West Pacific Ryukyus Isls. Zamami Island Intermediate Zam03_10M_33_Alveopora_C27 C27 JQ043669 Alveopora Alveopora sp. West Pacific Ryukyus Isls. Zamami Island Intermediate Zam03_10m_68_Montipora_C31 C31 JQ043599 Montipora Montipora sp. West Pacific Ryukyus Isls. Zamami Island Intermediate Zam03_10m_87_Montipora_C31 C31 JQ043604 Montipora Montipora venosa West Pacific Ryukyus Isls. Zamami Island Intermediate Zam03_3m_87_Montipora_C31 C31 JQ043603 Montipora Montipora sp. West Pacific Ryukyus Isls. Zamami Island Shallow Zam03_3m_92_Montipora_C30 C30 JQ043605 Montipora Montipora efflorescens West Pacific Ryukyus Isls. Zamami Island Shallow Zam03_3m_95_Montipora_C31 C31 JQ043600 Montipora Montipora capitata West Pacific Ryukyus Isls. Zamami Island Shallow Zam03_3m_25_Astreopora_C1 C1 KF572206 Astreopora Astreopora myriophthalma West Pacific Ryukyus Isls. Zamami Island Shallow Zam03_41_Fungia_C27 C27 JQ043676 Fungia Fungia sp. West Pacific Ryukyus Isls. Zamami Island Shallow

Table S2. Summary statistics on allele number and evenness among unique multilocus genotypes representing Symbiodinium lineages associated with Orbicella and Siderastrea from the Greater Caribbean of the western Atlantic. Standard error (numbers in smaller font) is given below each mean. C7 C7a/C12 C3Siderastrea Nunique 31 14 33 Na 3.200 3.400 7.100 0.573 0.600 0.767 Ae 2.055 2.356 3.697 0.347 0.412 0.442 Ho 0.000 0.000 0.599 0.000 0.000 0.077 He 0.377 0.445 0.673 0.094 0.090 0.057
Na = Average no. of alleles per locus Ae = No. of effective alleles per locus [1 / (Sum pi^2)] Ho = Observed heterozygosity [No. of Hets / N] He = Expected heterozygosity [1 - Sum pi^2]
Related Documents











