Host pigments: potential facilitators of photosynthesis in coral symbioses SOPHIE G. DOVE 1 , CARLI LOVELL 1 , MAOZ FINE 1 , JEFFRY DECKENBACK 1 , OVE HOEGH-GULDBERG 1 , ROBERTO IGLESIAS-PRIETO 2 & KENNETH R.N. ANTHONY 1 1 Centre for Marine Studies, Australian Research Council Centre of Excellence for Coral Reef Studies, University of Queensland, St Lucia, Qld 4072 Australia and 2 Unidad Académica Puerto Morelos, Instituto de Ciencias del Mary Limnología, Universidad Nacional Autónoma de México Apdo, Postal (1152) Cancún, 77500, Q. Roo, Mexico ABSTRACT Reef-building corals occur as a range of colour morphs because of varying types and concentrations of pigments within the host tissues, but little is known about their physi- ological or ecological significance. Here, we examined whether specific host pigments act as an alternative mecha- nism for photoacclimation in the coral holobiont. We used the coral Montipora monasteriata (Forskål 1775) as a case study because it occurs in multiple colour morphs (tan, blue, brown, green and red) within varying light-habitat distributions. We demonstrated that two of the non- fluorescent host pigments are responsive to changes in external irradiance, with some host pigments up-regulating in response to elevated irradiance. This appeared to facili- tate the retention of antennal chlorophyll by endosym- bionts and hence, photosynthetic capacity. Specifically, net Pmax Chl a -1 correlated strongly with the concentration of an orange-absorbing non-fluorescent pigment (CP-580). This had major implications for the energetics of bleached blue- pigmented (CP-580) colonies that maintained net Pmax cm -2 by increasing Pmax Chl a -1 . The data suggested that blue morphs can bleach, decreasing their symbiont populations by an order of magnitude without compromising symbiont or coral health. Key-words: all-protein chromophore; bleaching; dinoflagel- late; host pigment; photoacclimation; photosynthesis. INTRODUCTION Coral reefs are one of the most productive systems on the planet, which is in part because of the photosynthetic activ- ity of their endosymbiotic dinoflagellates (Symbiodinium sp.) and the efficient transfer of energy to the host in return for the host metabolic waste products (Hatcher 1988; Mus- catine 1990). This exchange results in a tight recycling of nutrients that partially explains the occurrence of coral reefs in the oligotrophic waters of the tropics (Lewis & Smith 1971; Muscatine 1990). Photosynthesis, however, is not merely dependant on an adequate supply of CO2 and nutrients, but also on the internal light environment experienced by their symbionts. Recently, the role played by the coral host in trapping available photons based on the diffuse reflective surface offered by the skeleton (Enriquez, Mendez & Iglesias-Prieto 2005) has been used to explain the observations of twofold or more increases in photon flux density (PFD) within normally pigmented host tissue over external PFD (Kuhl et al. 1995), and the paradoxically low number of external photons required by corals to evolve a molecule of oxygen given the light absorption properties calculated for isolated Symbiodinium in the lit- erature (Dubinsky et al. 1984, 1990). Organisms that have evolved to trap light have usually evolved mechanisms to counteract its negative consequences. In this study, we hypothesized that some corals use host pigments to reduce the negative consequences of high light while allowing sym- bionts to maintain high photosynthetic pigment densities. The efficiency with which corals are able to trap light suggests that maximizing light absorption is an important strategy even in the clear waters of tropical oceans (Enriquez et al. 2005). Corals display a wide range of skeletal structures (Veron 1986; Muscatine 1990), exhibit differential host tissue thickness (Anthony, Connolly & Willis 2002) and may harbour symbiont phylotypes with differential abilities to acclimate to changing light environ- ments (Iglesias-Prieto & Trench 1997; Rowan et al. 1997; Iglesias-Prieto et al. 2004). These differences suggest that conspecifics may adopt different strategies for maximizing photosynthesis and extending their range into a broad spec- trum of environments (light niches). Somewhat controver- sial is the role that host pigments play in modulating the internal light field for the endosymbionts with the literature providing articles that both support (Salih et al. 2000; Dove, Hoegh-Guldberg & Ranganathan 2001; Enriquez et al. 2005) and oppose (Gilmore et al. 2003; Mazel et al. 2003) the hypothesis that host pigments play either photo- protective or photo-enhancing roles in the symbiosis. Organisms that have evolved to capture light energy effi- ciently have also evolved mechanisms for dealing with its negative aspects (Niyogi 1999; Sparrow, Nakanishi & Parish 2000). The negative effects of ultraviolet radiation are well documented and include direct and indirect damage.Visible light, however, can be equally damaging through the same indirect pathway. In the indirect pathway, the interaction of light with a photosensitizer leads to the generation of Correspondence: S. G. Dove. Fax: +61 (7) 3365 4755; e-mail: [email protected] Plant, Cell and Environment (2008) 31, 1523–1533 doi: 10.1111/j.1365-3040.2008.01852.x © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd 1523

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Host pigments: potential facilitators of photosynthesis incoral symbioses
SOPHIE G. DOVE1, CARLI LOVELL1, MAOZ FINE1, JEFFRY DECKENBACK1, OVE HOEGH-GULDBERG1,ROBERTO IGLESIAS-PRIETO2 & KENNETH R.N. ANTHONY1
1Centre for Marine Studies, Australian Research Council Centre of Excellence for Coral Reef Studies, University ofQueensland, St Lucia, Qld 4072 Australia and 2Unidad Académica Puerto Morelos, Instituto de Ciencias del MaryLimnología, Universidad Nacional Autónoma de México Apdo, Postal (1152) Cancún, 77500, Q. Roo, Mexico
ABSTRACT
Reef-building corals occur as a range of colour morphsbecause of varying types and concentrations of pigmentswithin the host tissues, but little is known about their physi-ological or ecological significance. Here, we examinedwhether specific host pigments act as an alternative mecha-nism for photoacclimation in the coral holobiont. We usedthe coral Montipora monasteriata (Forskål 1775) as a casestudy because it occurs in multiple colour morphs (tan,blue, brown, green and red) within varying light-habitatdistributions. We demonstrated that two of the non-fluorescent host pigments are responsive to changes inexternal irradiance, with some host pigments up-regulatingin response to elevated irradiance. This appeared to facili-tate the retention of antennal chlorophyll by endosym-bionts and hence, photosynthetic capacity. Specifically, netPmax Chl a-1 correlated strongly with the concentration of anorange-absorbing non-fluorescent pigment (CP-580). Thishad major implications for the energetics of bleached blue-pigmented (CP-580) colonies that maintained net Pmax cm-2
by increasing Pmax Chl a-1. The data suggested that bluemorphs can bleach, decreasing their symbiont populationsby an order of magnitude without compromising symbiontor coral health.
Key-words: all-protein chromophore; bleaching; dinoflagel-late; host pigment; photoacclimation; photosynthesis.
INTRODUCTION
Coral reefs are one of the most productive systems on theplanet, which is in part because of the photosynthetic activ-ity of their endosymbiotic dinoflagellates (Symbiodiniumsp.) and the efficient transfer of energy to the host in returnfor the host metabolic waste products (Hatcher 1988; Mus-catine 1990). This exchange results in a tight recycling ofnutrients that partially explains the occurrence of coralreefs in the oligotrophic waters of the tropics (Lewis &Smith 1971; Muscatine 1990). Photosynthesis, however, isnot merely dependant on an adequate supply of CO2 andnutrients, but also on the internal light environment
experienced by their symbionts. Recently, the role played bythe coral host in trapping available photons based on thediffuse reflective surface offered by the skeleton (Enriquez,Mendez & Iglesias-Prieto 2005) has been used to explainthe observations of twofold or more increases in photonflux density (PFD) within normally pigmented host tissueover external PFD (Kuhl et al. 1995), and the paradoxicallylow number of external photons required by corals toevolve a molecule of oxygen given the light absorptionproperties calculated for isolated Symbiodinium in the lit-erature (Dubinsky et al. 1984, 1990). Organisms that haveevolved to trap light have usually evolved mechanisms tocounteract its negative consequences. In this study, wehypothesized that some corals use host pigments to reducethe negative consequences of high light while allowing sym-bionts to maintain high photosynthetic pigment densities.
The efficiency with which corals are able to trap lightsuggests that maximizing light absorption is an importantstrategy even in the clear waters of tropical oceans(Enriquez et al. 2005). Corals display a wide range ofskeletal structures (Veron 1986; Muscatine 1990), exhibitdifferential host tissue thickness (Anthony, Connolly &Willis 2002) and may harbour symbiont phylotypes withdifferential abilities to acclimate to changing light environ-ments (Iglesias-Prieto & Trench 1997; Rowan et al. 1997;Iglesias-Prieto et al. 2004). These differences suggest thatconspecifics may adopt different strategies for maximizingphotosynthesis and extending their range into a broad spec-trum of environments (light niches). Somewhat controver-sial is the role that host pigments play in modulating theinternal light field for the endosymbionts with the literatureproviding articles that both support (Salih et al. 2000;Dove, Hoegh-Guldberg & Ranganathan 2001; Enriquezet al. 2005) and oppose (Gilmore et al. 2003; Mazel et al.2003) the hypothesis that host pigments play either photo-protective or photo-enhancing roles in the symbiosis.
Organisms that have evolved to capture light energy effi-ciently have also evolved mechanisms for dealing with itsnegative aspects (Niyogi 1999; Sparrow, Nakanishi & Parish2000). The negative effects of ultraviolet radiation are welldocumented and include direct and indirect damage.Visiblelight, however, can be equally damaging through the sameindirect pathway. In the indirect pathway, the interaction oflight with a photosensitizer leads to the generation of
Correspondence: S. G. Dove. Fax: +61 (7) 3365 4755; e-mail:[email protected]
Plant, Cell and Environment (2008) 31, 1523–1533 doi: 10.1111/j.1365-3040.2008.01852.x
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd 1523

singlet oxygen (1O2) that oxidizes lipids, proteins and DNA,leading to damage and, ultimately, to either cell apoptosis ornecrosis depending on the extent to which the cell machin-ery is damaged (Suto et al. 2005). The blue light-absorbingfluorescent pigment A2E, for example, acts as a photosen-sitizer causing apoptosis of photoreceptors of the humanretina (Sparrow et al. 2000). This particular effect is miti-gated by the deployment of the plant pigment zeaxanthinthat not only effectively attenuates blue light but also scav-enges singlet oxygen (1O2) (Sparrow et al. 2000). Similarly,photosynthesis involves the capture of light energy by thephotosensitizer, chlorophyll (Chl), and generates a flow ofelectrons at the same time as it generates oxygen (O2),which readily accepts those electrons.The creation of 1O2 asa result of the interaction of Chls that are unable to offloadtheir excitation energy with O2, as well as the creation ofother forms of reactive oxygen species (ROS) such as super-oxide anions (O2
-) as O2 accepts electrons that are unable tobe processed by electron transport to NADPH, can be del-eterious to the photosynthetic apparatus (Niyogi 1999). Inthis context, pigments act both positively and negatively.Positively, because they are essential for capturing lightenergy (e.g. Chl), dissipating energy captured to heat (e.g.xanthophylls) and quenching ROS (e.g. carotenoids, inclu-sive of xanthophylls), and negatively, because they can alsobe a source of the harmful ROS.
Dinoflagellates, like other phototrophs, use Chl to traplight and carotenoids to mitigate the deleterious sideeffects. Corals that host dense populations of Symbiod-inium use their carbonate skeletons to trap light and con-sequently enhance the specific absorption of Chl underdownwelling irradiance (Enriquez et al. 2005). We hypoth-esize that reef-building corals use some of their host pig-ments [e.g. chromoproteins like the pocilloporins describedby Dove et al. (2001)] not only to mitigate the undesirableside effects to their own cells, but also to protect their criti-cally important resident Symbiodinium populations. Mon-tipora monasteriata is an abundant plating coral species onoffshore reefs of the southern Great Barrier Reef (GBR).The species occurs in a range of distinct colour morphs thatvary according to light habitat, suggesting that these chro-moproteins play an important ecological role (Dove et al.2006). Specifically, red, dark brown and green morphs occurin low-light habitats such as caves and under overhangs,whereas purple (blue) and tan morphs occur in open high-light habitats. The occurrence of morphs with distinctivecolouration within the same and distinct light habitats pre-sents an opportunity to study the effects of host pigmentson endosymbiont photophysiology.
In this study, we investigated how host and symbiontpigments respond to reciprocal transplantation betweencontrasting light environments at the same depth.We deter-mined whether host photoacclimation mitigated dinoflagel-late acclimation classically exemplified as decreases in Chlper symbiont cell in response to an increase in light(Falkowski & Dubinsky 1981; Hoegh-Guldberg & Smith1989). As part of this initial question, we characterized thepigments within the different morphs that were responsible
for their visually distinct colouration. Secondly, we investi-gated whether host chromoproteins reduced the efficiencyof O2 evolution by endosymbionts for coral surfacesexposed to a wavelength band that includes the pigment’swavelength of maximum absorption (lmax). Finally, weexplored the role of chromoproteins in situations wheredinoflagellate pigmentation is reduced as a function of lostdinoflagellate cells.
MATERIALS AND METHODS
Study sites and colony abundance
The study was conducted in reef grooves along the northernfront of Wistari Reef (23°27′S, 151°54′E) located nearHeron Island in the southern section of the GBR,Australia.Wistari Reef has a spur-and-groove formation, providing adiversity of natural light environments with shaded andexposed areas. At a depth range of between 5 and 7 m, theplate coral M. monasteriata is found in a variety of colourmorphs on the upper reef slope, along walls and underoverhangs (Anthony & Hoegh-Guldberg 2003b). Largenumbers of green, brown and red colonies are found in theshaded areas, while blue and tan colonies are found on thewell-lit parts of the reef slope. Corals were sampled in 2001,2002 and 2004.
Extraction and isolation of host pigment
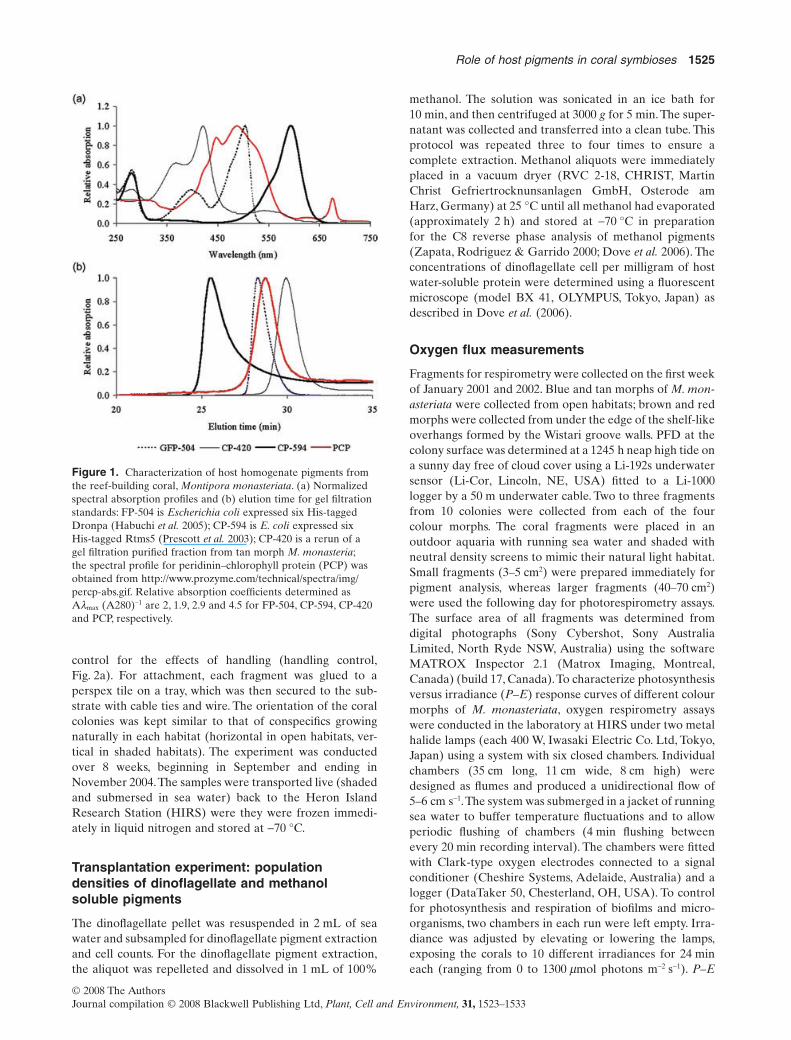
The tissue of each coral sample was removed from theskeleton by airbrushing with 10 mL 0.065 M phosphatebuffer solution (pH 6.65). This coral tissue slurry was thencentrifuged at 4500 g for 10 min at 4 °C, and the supernatantcontaining water-soluble host pigments was isolated fromthe dinoflagellate pellet and filtered (polyvinylidene difluo-ride, 0.45 mm). Water-soluble pigment proteins were sepa-rated by gel filtration chromatography as described by Doveet al. (2006) and identified on the basis of elution time[co-elution with CP-594 aka Rtms5-6His (Prescott et al.2003) and FP-504 aka Dronpa-6His (Habuchi et al. 2005)]and spectral signatures determined by the photodiode arraydetector (Fig. 1a,b). The remaining supernatant was trans-ferred to a clean tube and protein concentration estimatedusing the equation proposed byWhitaker & Granum (1980).
Reciprocal transplant experiment
To investigate photoacclimative responses of differentcolour morphs of M. monasteriata from contrasting lighthabitats, a reciprocal transplantation experiment betweenshaded (overhang) habitats and open groove habitats wasconducted. Two plate fragments, approximately 5 ¥ 5 cm insize, were cut from each of six colonies belonging to fivecolour morphs (providing a total of 60 fragments). Blue andtan colonies were initially collected from open habitats, andgreen, red and brown colonies from shaded habitats. Frag-ments were transplanted to two habitats: a contrasting lighthabitat (relocated, Fig. 2a) and their native habitat to
1524 S. G. Dove et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533
control for the effects of handling (handling control,Fig. 2a). For attachment, each fragment was glued to aperspex tile on a tray, which was then secured to the sub-strate with cable ties and wire. The orientation of the coralcolonies was kept similar to that of conspecifics growingnaturally in each habitat (horizontal in open habitats, ver-tical in shaded habitats). The experiment was conductedover 8 weeks, beginning in September and ending inNovember 2004.The samples were transported live (shadedand submersed in sea water) back to the Heron IslandResearch Station (HIRS) were they were frozen immedi-ately in liquid nitrogen and stored at -70 °C.
Transplantation experiment: populationdensities of dinoflagellate and methanolsoluble pigments
The dinoflagellate pellet was resuspended in 2 mL of seawater and subsampled for dinoflagellate pigment extractionand cell counts. For the dinoflagellate pigment extraction,the aliquot was repelleted and dissolved in 1 mL of 100%
methanol. The solution was sonicated in an ice bath for10 min, and then centrifuged at 3000 g for 5 min. The super-natant was collected and transferred into a clean tube. Thisprotocol was repeated three to four times to ensure acomplete extraction. Methanol aliquots were immediatelyplaced in a vacuum dryer (RVC 2-18, CHRIST, MartinChrist Gefriertrocknunsanlagen GmbH, Osterode amHarz, Germany) at 25 °C until all methanol had evaporated(approximately 2 h) and stored at -70 °C in preparationfor the C8 reverse phase analysis of methanol pigments(Zapata, Rodriguez & Garrido 2000; Dove et al. 2006). Theconcentrations of dinoflagellate cell per milligram of hostwater-soluble protein were determined using a fluorescentmicroscope (model BX 41, OLYMPUS, Tokyo, Japan) asdescribed in Dove et al. (2006).
Oxygen flux measurements
Fragments for respirometry were collected on the first weekof January 2001 and 2002. Blue and tan morphs of M. mon-asteriata were collected from open habitats; brown and redmorphs were collected from under the edge of the shelf-likeoverhangs formed by the Wistari groove walls. PFD at thecolony surface was determined at a 1245 h neap high tide ona sunny day free of cloud cover using a Li-192s underwatersensor (Li-Cor, Lincoln, NE, USA) fitted to a Li-1000logger by a 50 m underwater cable. Two to three fragmentsfrom 10 colonies were collected from each of the fourcolour morphs. The coral fragments were placed in anoutdoor aquaria with running sea water and shaded withneutral density screens to mimic their natural light habitat.Small fragments (3–5 cm2) were prepared immediately forpigment analysis, whereas larger fragments (40–70 cm2)were used the following day for photorespirometry assays.The surface area of all fragments was determined fromdigital photographs (Sony Cybershot, Sony AustraliaLimited, North Ryde NSW, Australia) using the softwareMATROX Inspector 2.1 (Matrox Imaging, Montreal,Canada) (build 17, Canada).To characterize photosynthesisversus irradiance (P–E) response curves of different colourmorphs of M. monasteriata, oxygen respirometry assayswere conducted in the laboratory at HIRS under two metalhalide lamps (each 400 W, Iwasaki Electric Co. Ltd, Tokyo,Japan) using a system with six closed chambers. Individualchambers (35 cm long, 11 cm wide, 8 cm high) weredesigned as flumes and produced a unidirectional flow of5–6 cm s-1.The system was submerged in a jacket of runningsea water to buffer temperature fluctuations and to allowperiodic flushing of chambers (4 min flushing betweenevery 20 min recording interval). The chambers were fittedwith Clark-type oxygen electrodes connected to a signalconditioner (Cheshire Systems, Adelaide, Australia) and alogger (DataTaker 50, Chesterland, OH, USA). To controlfor photosynthesis and respiration of biofilms and micro-organisms, two chambers in each run were left empty. Irra-diance was adjusted by elevating or lowering the lamps,exposing the corals to 10 different irradiances for 24 mineach (ranging from 0 to 1300 mmol photons m-2 s-1). P–E
Figure 1. Characterization of host homogenate pigments fromthe reef-building coral, Montipora monasteriata. (a) Normalizedspectral absorption profiles and (b) elution time for gel filtrationstandards: FP-504 is Escherichia coli expressed six His-taggedDronpa (Habuchi et al. 2005); CP-594 is E. coli expressed sixHis-tagged Rtms5 (Prescott et al. 2003); CP-420 is a rerun of agel filtration purified fraction from tan morph M. monasteria;the spectral profile for peridinin–chlorophyll protein (PCP) wasobtained from http://www.prozyme.com/technical/spectra/img/percp-abs.gif. Relative absorption coefficients determined asAlmax (A280)-1 are 2, 1.9, 2.9 and 4.5 for FP-504, CP-594, CP-420and PCP, respectively.
Role of host pigments in coral symbioses 1525
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533
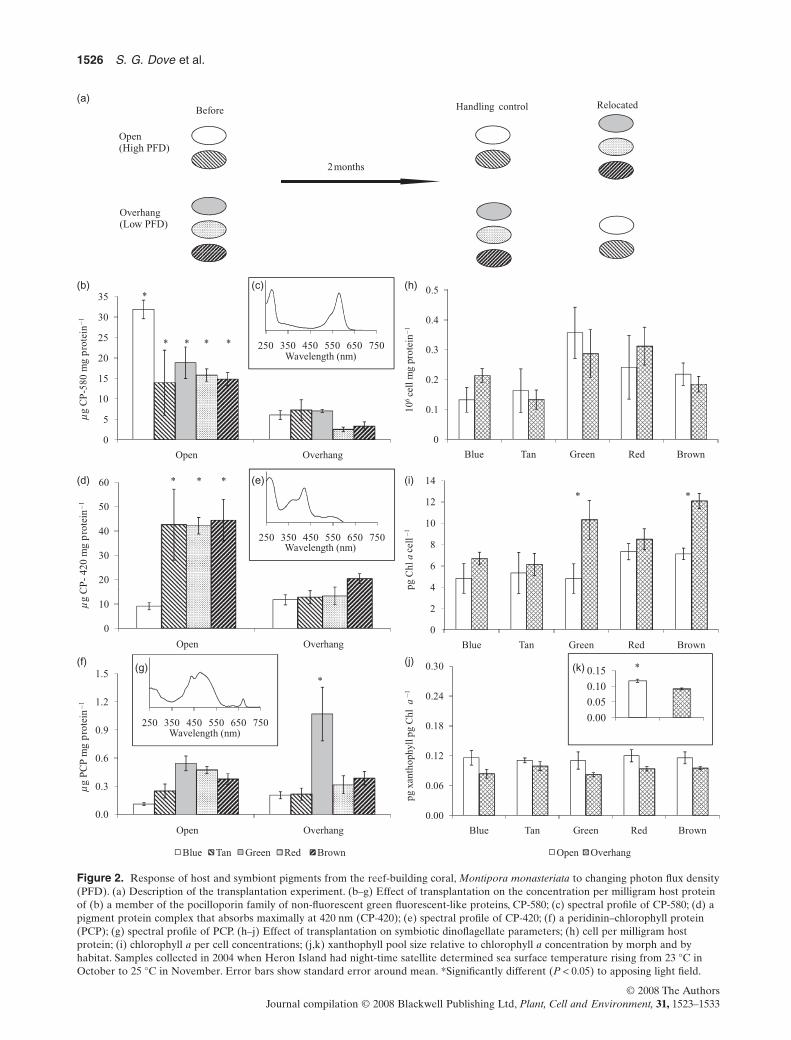
Figure 2. Response of host and symbiont pigments from the reef-building coral, Montipora monasteriata to changing photon flux density(PFD). (a) Description of the transplantation experiment. (b–g) Effect of transplantation on the concentration per milligram host proteinof (b) a member of the pocilloporin family of non-fluorescent green fluorescent-like proteins, CP-580; (c) spectral profile of CP-580; (d) apigment protein complex that absorbs maximally at 420 nm (CP-420); (e) spectral profile of CP-420; (f) a peridinin–chlorophyll protein(PCP); (g) spectral profile of PCP. (h–j) Effect of transplantation on symbiotic dinoflagellate parameters; (h) cell per milligram hostprotein; (i) chlorophyll a per cell concentrations; (j,k) xanthophyll pool size relative to chlorophyll a concentration by morph and byhabitat. Samples collected in 2004 when Heron Island had night-time satellite determined sea surface temperature rising from 23 °C inOctober to 25 °C in November. Error bars show standard error around mean. *Significantly different (P < 0.05) to apposing light field.
1526 S. G. Dove et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533
curve parameters and associated standard errors wereestimated by fitting the model [P = Pmax tanh (E/Ek) - R](Jassby & Platt 1976) to the net photosynthesis-irradiancedata set using iterative non-linear estimation (STATIS-TICA version 7; StatSoft Inc., Tulsa, OK, USA). The pho-toinhibition model of Platt, Gallegos & Harrison (1980) wastried but found to fit the data less well. For all morphs, thecoefficient of photoinhibition, b, was found not to be signifi-cantly different from 0. Alpha, oxygen flux per unit irradi-ance was determined directly from the linear slope of the3–5 points taken at extremely low illumination (Iglesias-Prieto & Trench 1994).
To determine the responses of different colour morphs tochanges in the spectral quality, a second set of respirometryassays was run to determine the efficiency of O2 evolution(aw) for fragments exposed to sub-saturating light intensitiesof white light, then subsequently red (ar), green (ag) or blue(ab) light derived by covering the chambers with LEE filters(Andover, UK). Red Lee filters (Andover, Hampshire, UK)(number 026) transmit photosynthetically active radiation(PAR) in excess of 575 nm (Fig. 3a); green filters (number124) transmit PAR between 450 and 600 nm (Fig. 3b), andblue filters (number 715) transmit PAR less than 550 nm andgreater than 625 nm (Fig. 3c). Corrections were made toinsure that PAR exposure was the same under all treatments,leaving only spectral quality to vary. aw, ar, ag and ab were
determined for five colonies from each of the four morphs.ar, ag and ab were normalized to aw in order to provide aresponse that is independent of differential O2 productionthat are unrelated to spectral quality.
Rates of photosynthesis and the values of all key P–Emodel parameters (except Ek) were normalized to concen-trations of Chl a and coral surface area.At the conclusion ofthe oxygen flux assays, coral subcolonies were waterpikedwith filtered sea water, and the slurry was centrifuged at4000 g for 5 min and then homogenized. Pellets containingsymbionts were resuspended in sea water and divided intotwo equal portions. The number of cells was estimated bycounting six independent subsamples of one portion, theremaining portion was centrifuged at 4000 g for 5 min, withthe pellet resuspended in 90% acetone. Three separateacetone extractions were combined and used for spectro-scopic determination of Chl a concentrations (Jeffrey &Humphrey 1975).
Statistical analysis of transplantation data
Because colony fragments were halved, with one-halfplaced back into the original light environment and theother half placed into the opposing light environment, datawere analysed using a factorial (morph, treatment – PFD)within-subject analysis of variance (anova). A StudentNewman Keul (SNK) post hoc analysis was applied to inter-pret statistically significant interactions.
Statistical analysis of data of oxygen flux data
Effects of habitat (high light versus low light) and colourmorph on a variety of biophysiological variables were testedseparately using two one-way anovas because the colourmorphs are not present in both habitats. Cochran’s C, Bar-tlett’s chi-squared and Hartley’s F-max tests were used toexamine the assumption of homogeneity of variance.
RESULTS
The four colour morphs of M. monasteriata contained anarray of host chromoproteins with different optical proper-ties, explaining their colour expression in situ. The bluemorph of M. monasteriata from open high-light habitats isblue at depth (where red light is preferentially lost) andpurple at the surface. Blue morph contains an orange-absorbing non-fluorescent pigment (CP-580), with an aminoacid sequence that is 98% identical to the tetramer Rtms5(CP-594) and belongs to a group of proteins referred to aspocilloporins that are either blue (lmax = 594 nm), blue-purple (lmax = 586 nm),purple (lmax = 580 nm) or clear whenexpressed in bacteria (Dove et al. 2002). The tan morph,also from open habitats, contains a novel non-fluorescentviolet-absorbing (lmax = 420 nm) yellow pigment (CP-420)that, at approximately 20 kD, is smaller than either aHis-tag monomeric green florescent-like protein (GFP)(engineered Dronpa = FP-504) or a peridinin–chlorophyllprotein [(PCP) Fig. 1b].While FP-504 and pocilloporins (e.g.
(a) (b) (c)
0.8
1.0
effi
cien
cy
*
(d)
0.0
0.2
0.4
0.6
Blue morph
Rel
ativ
e ph
otos
ynth
etic
*
Open Overhang
Tan morph Red morph Brown morph
60
80
100
60
80
100
60
80
100
0
20
40
0
20
40
0
20
40
400 500 600 700 400 500 600 700 400 500 600 700
% T
rans
mis
sion
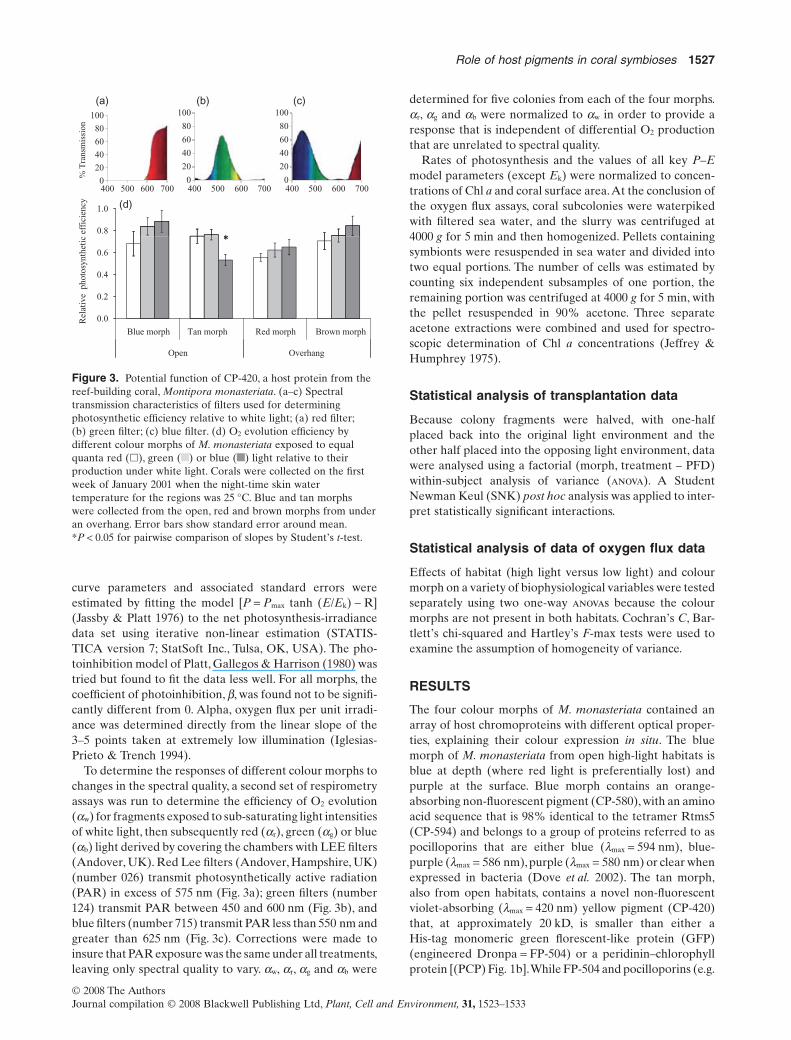
Figure 3. Potential function of CP-420, a host protein from thereef-building coral, Montipora monasteriata. (a–c) Spectraltransmission characteristics of filters used for determiningphotosynthetic efficiency relative to white light; (a) red filter;(b) green filter; (c) blue filter. (d) O2 evolution efficiency bydifferent colour morphs of M. monasteriata exposed to equalquanta red (�), green ( ) or blue ( ) light relative to theirproduction under white light. Corals were collected on the firstweek of January 2001 when the night-time skin watertemperature for the regions was 25 °C. Blue and tan morphswere collected from the open, red and brown morphs from underan overhang. Error bars show standard error around mean.*P < 0.05 for pairwise comparison of slopes by Student’s t-test.
Role of host pigments in coral symbioses 1527
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533

CP-580) have very similar absorption coefficients at lmax, therelative absorption coefficient of CP-420 is approximately1.6 times than that of these all-protein chromophores(APCs) but less than that of PCP at lmax (cf. relative absorp-tion coefficients, Fig. 1a). Elevated concentrations of PCPwere found in the host homogenate of green morphscollected from the overhangs [Fig. 2f,g; morph ¥ PFD,F(4,16) = 4.06, P = 0.019; SNK]. Also present in this greenmorph at low concentrations [<2.5 mg mg (protein)-1, assum-ing similar extinction coefficient to FP-504 and CP-580] wasa green fluorescent pigment protein that eluted in the samefraction as the tetrameric GFP-like protein pocilloporin, butwith a maximal absorbance at 465 nm. Despite their visuallydistinct colouration at depth, the red and brown morphsfrom the shaded overhangs had similar spectral signatures,both revealing low concentrations of CP-580, CP-420 andPCP in host homogenate.
In the transplantation experiment, host pigment proteinsresponded to changing PFD although not all colour morphsresponded in the same way. The amount of blue pigmenta-tion (CP-580) was reduced in all corals moved from high-light to low-light environments, and increased for coralsmoving from low light to high light (Fig. 2b,c; morph ¥ PFD:F(4,16) = 6.63, P = 0.002). The same was true for the yellowpigment CP-420, with the exception of the green and bluemorphs that appeared unable to express large quantitiesof CP-420 when placed in high light fields (Fig. 2d,e;morph ¥ PFD: F(3,14) = 3.51, P = 0.04). The blue morphexpressed more CP-580 in high-light fields than any othermorph (Fig. 2a,b; SNK, blue > others).The concentration ofCP-580 and CP-420 in the tan, brown and red morphs,however, appeared to be solely a function of the PFD(Fig. 2b–e).
The number of dinoflagellate cells per milligram hostprotein was not significantly different between light habitats(Fig. 2h; PDF, F(1,16) = 0.22, P = 0.89), which is a similarresult to that reported by Anthony & Hoegh-Guldberg(2003a). In addition, there was no effect of colour morph[F(4,16) = 1.95, P = 0.15] on this variable, and themorph ¥ treatment interaction term was non-significant.There was, however, a significant morph ¥ treatment inter-action for the concentration of Chl a per cell. Onlydinoflagellates from brown and green morphs had higherconcentrations of Chl a per cell for fragments held in thelow-light environment compared with the high-light envi-ronment (Fig. 2i; morph ¥ PFD, F(4,17) = 2.98, P = 0.049).Peridinin to Chl a ratios were not significantly differentfor any factor [morph, F(4,17) = 1.50, P = 0.25; PFD,F(1,17) = 1.02,P = 0.33; interaction,F(4,17) = 0.77,P = 0.56].The xanthophyll pool (diadinoxanthin + diatoxanthin) toChl a ratios were,however,affected by PFD,with a relativelylarger xanthophyll pool in corals exposed to high-light com-pared with low-light regimes (Fig. 2j,k; PFD, F(1,17) = 15.25,P = 0.001). Within light environments, the xanthophyll poolto Chl a ratios were constant across colour morphs (Fig. 2j).The presence of large quantities of PCP in the host homo-genate of shaded green morphs may be explained by agreater tendency for Symbiodinium lysis in these samples.
Alternatively, Chl a in green morphs may be associatedwith the more water-soluble peripheral antennae as opposedto the membrane bound Chl a-Chl c2-peridinin-proteincomplexes (acpPCs) (Iglesias-Prieto & Trench 1997).
The relative efficiency of O2 evolution by symbiontswithin the tan colour morph of M. monasteriata was signifi-cantly reduced when coral surfaces were irradiated withsame intensity blue as opposed to green or red light (pair-wise comparison of slopes by Student’s t: green versus bluefilter, t(32,2) = 2.77, P = 0.009; red versus blue filter, t(32,2)
= 2.59, P = 0.015; Fig. 3d). This result is consistent with thehigh abundance of CP-420 in the tan morph (cf. Fig. 4 &Table 1), which acts as an internal violet-blue filter. Inter-estingly, none of the other morphs displayed variation inphotosynthetic efficiency among light qualities (cf. Fig. 4 &Table 1).
Morphs of M. monasteriata began to exhibit coral bleach-ing in early January of 2002, with all morphs showing reduc-tions in areal Symbiodinium densities (Fig. 4e, cf. Anthony
500
–1
0.06
anta
)
0
100
200
300
400
Tan Blue Brown Red
mmol
qua
nta
m–2
s
0.00
0.02
0.04
tan blue brown red
(mol
O2
mol
qua
3.0
4.0
)
C
1 0
1.5
a -1
)
D
3.0
4.0
–2)
1.0
1.5 *
*
*
0.0
1.0
2.0
tan blue brown red
Pm
ax (c
m-2
0.0
0.5
1.0
tan blue brown red
Pm
ax (m
g C
hl
10
15
20
5cm
-2 59
77
93
E
10
15
a ce
ll -1
F
0.0
1.0
2.0
tan blue brown red
Pm
ax (c
m
0.0
0.5
1.0
tan blue brown red
Pm
ax (mg
Chl
a –
1 )
6
9
12
15
105
cm–2
6
9
12
h
*
*
0
5
10
tan blue brown red
cell
x10
14
0
5
tan blue brown red
pg C
hl
0
3
6
Tan Blue Brown Red
cell
×
0
3
Tan Blue Brown Red
pg C
hl a
cel
l –1
(b)
(d)
(f)
(a)
(c)
(e)
Figure 4. Photosynthetic response of different morphs ofthe reef-building coral, Montipora monasteriata at the onsetof thermal bleaching in the summer of 2002. (a) Lightintensity at colony surface taken at midday on a neap tide;(b) photosynthetic efficiency – O2 evolved per quanta incidenton coral surface area, aarea (mol O2 mol-1 quanta); (c) arealphotosynthetic maxima – Pmax cm-2 (mmol O2 h-1 cm-2);(d) photosynthetic maxima per dinoflagellate chlorophylla – Pmax mg Chl a-1 (mmol O2 h-1 mg-1 Chl a); (e) areal density ofdinoflagellates (cell cm-2); (f) chlorophyll a concentration perdinoflagellate cell (pg Chl a cell-1). Corals were collected on thefirst week of January 2002 when the night-time skin watertemperature for the regions attained 27.5 °C. Error bars showstandard error around mean. *Significantly different (P < 0.05) bypost hoc Student Neuman Keul (SNK), following a significanteffect in a two-factorial analysis of variance (anova). This coversboth simultaneously.
1528 S. G. Dove et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533
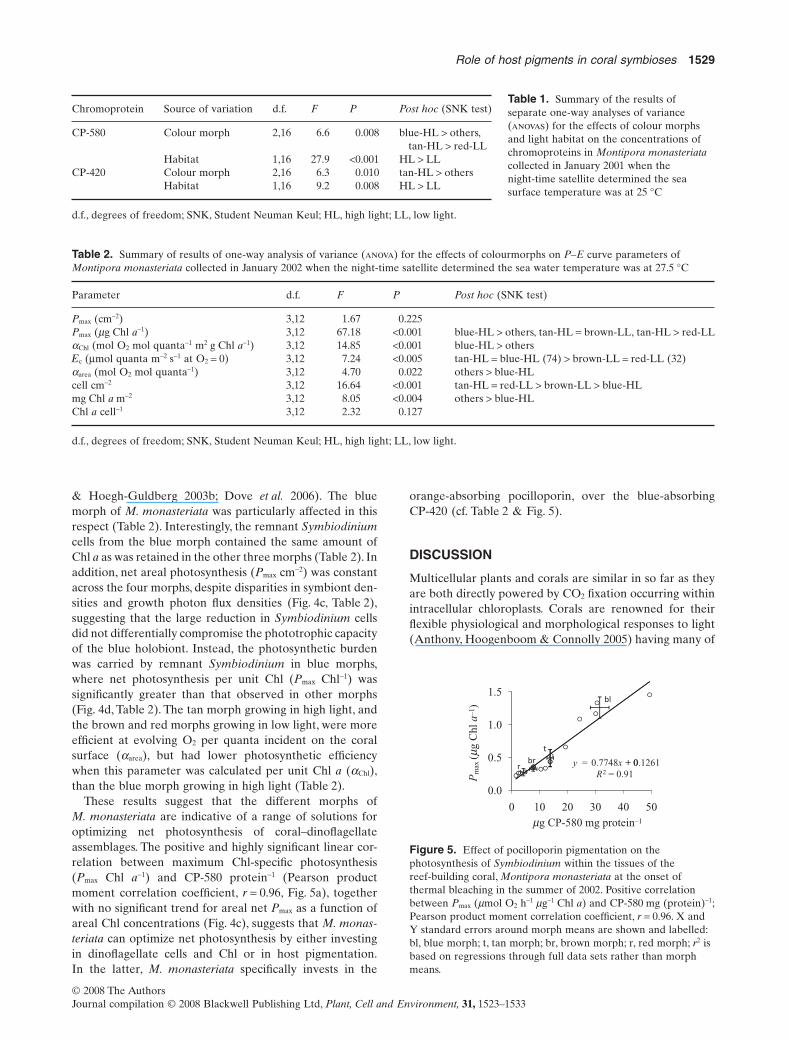
& Hoegh-Guldberg 2003b; Dove et al. 2006). The bluemorph of M. monasteriata was particularly affected in thisrespect (Table 2). Interestingly, the remnant Symbiodiniumcells from the blue morph contained the same amount ofChl a as was retained in the other three morphs (Table 2). Inaddition, net areal photosynthesis (Pmax cm-2) was constantacross the four morphs, despite disparities in symbiont den-sities and growth photon flux densities (Fig. 4c, Table 2),suggesting that the large reduction in Symbiodinium cellsdid not differentially compromise the phototrophic capacityof the blue holobiont. Instead, the photosynthetic burdenwas carried by remnant Symbiodinium in blue morphs,where net photosynthesis per unit Chl (Pmax Chl-1) wassignificantly greater than that observed in other morphs(Fig. 4d, Table 2). The tan morph growing in high light, andthe brown and red morphs growing in low light, were moreefficient at evolving O2 per quanta incident on the coralsurface (aarea), but had lower photosynthetic efficiencywhen this parameter was calculated per unit Chl a (aChl),than the blue morph growing in high light (Table 2).
These results suggest that the different morphs ofM. monasteriata are indicative of a range of solutions foroptimizing net photosynthesis of coral–dinoflagellateassemblages. The positive and highly significant linear cor-relation between maximum Chl-specific photosynthesis(Pmax Chl a-1) and CP-580 protein-1 (Pearson productmoment correlation coefficient, r = 0.96, Fig. 5a), togetherwith no significant trend for areal net Pmax as a function ofareal Chl concentrations (Fig. 4c), suggests that M. monas-teriata can optimize net photosynthesis by either investingin dinoflagellate cells and Chl or in host pigmentation.In the latter, M. monasteriata specifically invests in the
orange-absorbing pocilloporin, over the blue-absorbingCP-420 (cf. Table 2 & Fig. 5).
DISCUSSION
Multicellular plants and corals are similar in so far as theyare both directly powered by CO2 fixation occurring withinintracellular chloroplasts. Corals are renowned for theirflexible physiological and morphological responses to light(Anthony, Hoogenboom & Connolly 2005) having many of
Table 1. Summary of the results ofseparate one-way analyses of variance(anovas) for the effects of colour morphsand light habitat on the concentrations ofchromoproteins in Montipora monasteriatacollected in January 2001 when thenight-time satellite determined the seasurface temperature was at 25 °C
Chromoprotein Source of variation d.f. F P Post hoc (SNK test)
CP-580 Colour morph 2,16 6.6 0.008 blue-HL > others,tan-HL > red-LL
Habitat 1,16 27.9 <0.001 HL > LLCP-420 Colour morph 2,16 6.3 0.010 tan-HL > others
Habitat 1,16 9.2 0.008 HL > LL
d.f., degrees of freedom; SNK, Student Neuman Keul; HL, high light; LL, low light.
Table 2. Summary of results of one-way analysis of variance (anova) for the effects of colourmorphs on P–E curve parameters ofMontipora monasteriata collected in January 2002 when the night-time satellite determined the sea water temperature was at 27.5 °C
Parameter d.f. F P Post hoc (SNK test)
Pmax (cm-2) 3,12 1.67 0.225Pmax (mg Chl a-1) 3,12 67.18 <0.001 blue-HL > others, tan-HL = brown-LL, tan-HL > red-LLaChl (mol O2 mol quanta-1 m2 g Chl a-1) 3,12 14.85 <0.001 blue-HL > others�c (mmol quanta m-2 s-1 at O2 = 0) 3,12 7.24 <0.005 tan-HL = blue-HL (74) > brown-LL = red-LL (32)aarea (mol O2 mol quanta-1) 3,12 4.70 0.022 others > blue-HLcell cm-2 3,12 16.64 <0.001 tan-HL = red-LL > brown-LL > blue-HLmg Chl a m-2 3,12 8.05 <0.004 others > blue-HLChl a cell-1 3,12 2.32 0.127
d.f., degrees of freedom; SNK, Student Neuman Keul; HL, high light; LL, low light.
0 7748 + 0 12610.5
1.0
1.5
y = . x + 0.R2 = 0.91
0.0
0 10 20 30 40 50
Pm
ax (m
g C
hl a
–1)
mg CP-580 mg protein–1
Figure 5. Effect of pocilloporin pigmentation on thephotosynthesis of Symbiodinium within the tissues of thereef-building coral, Montipora monasteriata at the onset ofthermal bleaching in the summer of 2002. Positive correlationbetween Pmax (mmol O2 h-1 mg-1 Chl a) and CP-580 mg (protein)-1;Pearson product moment correlation coefficient, r = 0.96. X andY standard errors around morph means are shown and labelled:bl, blue morph; t, tan morph; br, brown morph; r, red morph; r2 isbased on regressions through full data sets rather than morphmeans.
Role of host pigments in coral symbioses 1529
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533

the characteristics seen in terrestrial plant communitiesinhabiting rain forests (Anthony & Hoegh-Guldberg2003b). The architecture of mature plants; the size and dis-position of internodes and leaves; the balance betweenmain stem and lateral branches; and the angles of petiolesand leaf laminae are all aimed at placing these chloroplastsin the optimal light environment (Pearcy & Yang 1998).Similarly, the solid support provided by a skeleton enablesgrowing corals to seek regions of relatively unattenuatedlight. In plants, non-chloroplast pigments–proteins, the phy-tochromes, play key roles in this process. In this study, ouraim was to determine the significance of non-chloroplast-based pigments in corals.
The pigmented cofactors of phytochromes undergophotoisomerization in response to different wavelengthsof light and are thereby able to distinguish between theamount of red and far-red light in the environment, and actas proximity sensors directing branch projection into unat-tenuated light fields (Smith 2000). Some APCs (also knownas GFP homologs, inclusive of blue-purple non-fluorescentpocilloporins) have been shown to undergo photoisomer-ization in response to blue and green wavelengths of light(Lukyanov et al. 2000; Turcic et al. 2006). These photoacti-vated host pigments, therefore, have the potential to beinvolved in photoreception in the marine environment,where the dominance of blue over green light is a functionof depth and the number of photosynthetic organismsin the water column. Phytochromes, however, tend tobe expressed more in low- than high-light environments(Sharrock & Clack 2002), which is the opposite of what wasobserved for the two dominant host pigments (CP-580 andCP-420) in this study. The increased expression of thesepigments upon transfer to higher-light fields is suggestive ofa function in photoprotection.
The array of light niches inhabited by corals, or foundwithin any coral colony, is highly diverse. Hermatypic coralsappear to have evolved an effective means for trapping lightwhere little light is available (Enriquez et al. 2005). In thelow-light environments, simply increasing Chl content toincrease the efficiency of light harvesting is counterproduc-tive as increasing pigment density simultaneously serves tofurther attenuate light. Corals mitigate this effect by placingdinoflagellates in a highly reflective box (the polyp calyxwithin coral skeletons). The enhanced ability to capturephotons, however, is problematic for well-illuminated posi-tions within the coral colony. In this study, we have demon-strated that host pigments are responsive to increases inexternal PFD and may therefore be used to dampen theinternal light environment. In line with this, some highlypigmented morphs of Pocillopora damicornis have beenshown to also increase host pigmentation upon transfer tohigh-light environments and to decrease it upon transfer tolow-light environments (Takabayashi & Hoegh-Guldberg1995). Furthermore, recent work has demonstrated thathosts can double pigmentation within a week, with anincrease in pocilloporin mRNA expression detected within24 h (J. Deckenback, personal observation).This time framefor photoacclimation is similar to that observed for
free-living phytoplankton, higher plants and some symbi-otic dinoflagellates, where the protein template of theantennae is altered to accommodate a greater xanthophyllpool and reduced Chl concentrations (Falkowski & Dubin-sky 1981; Niyogi 1999;Anthony & Hoegh-Guldberg 2003a).
Corals have evolved to effectively harvest light energy,and it is likely that host pigmentation represents a mecha-nism for controlling the ensuing negative consequences.Host pigments (APCs) are found in a variety of organisms,some of which lack limestone skeletons and photosyntheticsymbionts (Wilmann et al. 2006). It is clear, therefore, thatthe proposed photoprotective role does not have to includethe provision of an optimal light environment for photosyn-thesis. Despite this fact, corals that contain dinoflagellatesas symbionts are more effective at depositing the extensiveand elaborate calcium carbonate skeletons that serve totrap light (Barnes & Chalker 1990; Enriquez et al. 2005).
Only two pigments, the as yet undescribed, CP-420, andthe pocilloporin APC, CP-580, dominated the water-solublespectral profile of morphs of M. monasteriata grown underhigh light. In vitro, these pigments had absorption spectrathat cover the troughs within (e.g. CP-580), or substantiallyoverlap (e.g. CP-420) the absorption spectra of the majordinoflagellate light-harvesting pigment, PCP (Fig. 1a). Theexpression of threefold more CP-420 milligram per milli-gram protein in the tan morph corresponded with the onlycase observed where sub-saturation intensities of blue light(lmax = 440 nm) were less efficient at evolving oxygen thanthe same intensity green or red light. These results suggestthat the presence of a non-fluorescent blue-absorbingpigment in the host may intercept photons otherwise des-tined for photochemical quenching by the endosymbionts.
Dinoflagellate densities for M. monasteriata (inclusive ofblue morph) collected in April of 2003 were approximately0.4 ¥ 106 cell mg (protein)-1 or 2.0 ¥ 106 cell cm-2 (Dove et al.2006). This is approximately double the values obtained forblue morphs in late spring of 2004 (ª 0.2 ¥ 106 cell mg (pro-tein)-1 and is an order of magnitude greater than the valuesobtained for this morph in the summer of 2002 (ª 2 ¥ 105
cell cm-2). There is, therefore, a suggestion of a temporalshift in symbiont densities within this morph of blue M.monasteriata, potentially dictated by temperature. At thevery least, observed shifts in Symbiodinium populationsbetween studies justifies the collection of a much morecomprehensive data set to determine actual seasonal trends(Fitt et al. 2000; Warner et al. 2002). In early January 2002,temperature records show that the thermal threshold forthe region that coincide with a detrimental mass bleachingevent were not yet attained (Hoegh-Guldberg 1999; Berkel-mans 2002). Two options thus present: (1) blue morph M.monasteriata seasonally bleached to this extent with nonegative consequences at the colony level or (2) bluemorph M. monasteriata have relatively low thermal thresh-olds. Colony health is determined not only by the totalquantity of carbon fixed, but also by the proportion of fixedcarbon that is translocated to the host (Muscatine 1990).For some corals, the proportion of carbon translocatedvaries with environmental variables such as temperature
1530 S. G. Dove et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533

(Loram, Trapido-Rosenthal & Douglas 2007). An increasein the proportion of fixed carbon translocated to the host,however, may compromise symbiont population densitiesin favour of host growth (Smith 1987). Therefore, bleachingassociated with a thermal increase that does not compro-mise carbon fixation need not negatively affect host health.
Endosymbionts within blue morph M. monasteriata expe-riencing significant reductions in population densities inearly 2002 did not reduce the concentration of Chl per cell.Predicted elevated internal light fields (Adamska, Ohad &Kloppstech 1992; Enriquez et al. 2005) did not lead to eitheruncontrollable photobleaching of the antennae (Dove et al.2006) or a restructuring of the antennae that results in fewerChls appended to light-harvesting proteins (Niyogi 1999).The response is notably distinct from that obtained forthe blue morph of M. monasteriata exposed to 32 °C for6 h, where photo-oxidation of the endosymbionts’ light-harvesting antennae co-occurred with almost a completeloss of CP-580 (Dove et al. 2006).
In the period leading into the 2002 GBR mass bleachingevent, net O2 production under saturating light intensitiesper unit investment in Chl a was unrelated to CP-420 con-centration, but showed an almost perfect correlation withthe concentration of CP-580 present in the host tissue. Theresponse in net Pmax Chl a-1 (equally in this instance, netPmax cell-1) appears to be a response to the fact that the hosttissue of the blue morph has become optically thinner as aresult of significant bleaching, potentially increasing theexcitation pressure on photosystem II (PSII) as a conse-quence (Iglesias-Prieto et al. 2004). A similar response wasnoted by Maxwell, Falk & Huner (1995) who found thatcultures of Chorella vulgaris exposed to a sevenfoldincrease in PSII excitation pressure trebled light-saturatedrates of O2 evolution.
The results of the study, therefore, question someassumptions currently prevalent in the coral literature.These are: (1) that thermal tolerance necessarily equates tobleaching tolerance (aka resistance to reductions in sym-biont densities) and (2) that the loss of dinoflagellates fromcorals necessarily correlates with reduced carbon fixationby remnant dinoflagellates (Rowan 2004). Arguably, acoral is not detrimentally bleached until it is substantiallydepleted in both host and symbiont pigmentation. This hasthe consequence that the current operational definition ofthermal tolerance, which is defined in terms of the resilienceof corals to bleaching (and which is used to determinethermal thresholds for coral health), is not universallyapplicable to all coral morphs. The problems with thisoperational definition are perhaps best illustrated by thefact that the growing margins of fast-growing corals (fre-quently blue in colouration) are typically bleached in com-parison with the rest of the colony (Gladfelter 1983), andthe converse fact that corals that host bleaching resistantsymbionts of clade D grow at a slow rate (Little, van Oppen& Willis 2004; Berkelmans & van Oppen 2006).
Unlike CP-420, CP-580 principally absorbs where thephotochemical pigments of Symbiodinium do not. For thisreason, any photoprotection offered by CP-580 towards
symbiont photosynthesis is likely to be indirect. Inthis regard, skeletal light amplification is greater forwavelengths that are not attenuated by the presence ofpigments (Kuhl et al. 1995). High densities of CP-580 poten-tially prevents the amplification of light between 550 and650 nm, thereby blue enriching internal light fields experi-enced by endosymbionts. Blue-enriched spectra have beenargued to promote Chl retention and large antioxidantpools (Levy et al. 2006). Additionally, blue light is alsolinked to the mRNA induction of early light-inducible pro-teins (ELiPs) that are significantly repressed by light in the550–600 nm wavelength band even in the presence of bluelight (Adamska et al. 1992). ELiPs are expressed in a rangeof photosynthetic organisms in response to increased lightand/or to facilitate chloroplast formation or chloroplastrestructuring (Bruno & Wetzel 2004). They are believed tobe essential to limiting the photo-oxidation of the antennaeresultant from the excitation pressure on PSII, either byproviding an alternative structure upon which to bind oth-erwise uncoupled but photoactive Chls (Adamska 2001)or by directly facilitating reaction centre quenching (RCquenching) (Montane & Kloppstech 2000; Matsubara &Chow 2004). The retention of Chl antennae by remnantSymbiodinium may relate to the expression of ELiP ana-logues facilitated by the presence of CP-580. ELiPs areancient proteins present in higher plants and algae, withclosely related functional analogues in cyanobacteria (highlight inducible proteins; Montane & Kloppstech 2000). BothELiPs and high light inducible proteins are related to theproteins (light harvesting chlorophyll binding proteins ofII) in the light-harvesting antennae of algae and plants,and to the membrane-bound antennae proteins (acpPC)present in dinoflagellates (Green & Durnford 1996;Montane & Kloppstech 2000). Arguably, ELiP evolutionpredates the evolution of peripheral light-harvesting com-plexes, with the implication that ELiP variants may beretained in most photosynthetic organisms inclusive ofSymbiodinium (Montane & Kloppstech 2000).
In summary, photoacclimation of a coral involves bothhost and symbiont responses. A coral that grows quickly,post-filling growing margins with dinoflagellates, or oper-ates in some seasons with few internal symbiont, facessimilar problems associated with unattenuated light fieldsto that experienced by a greening plant or a developingseedling (Adamska et al. 1992). Host pigments that areup-regulated in response to elevated light fields can facili-tate the establishment of new or restructured dinoflagellatechloroplasts by modulating the internal light field. In thisstudy, we showed that elevated concentrations of CP-420provide photoprotection for resident Symbiodinium byintercepting photons destined for photochemical quenchingby dinoflagellates. This aspect of CP-580 was not investi-gated because of an absence of the appropriate filter; it was,however, shown that extensively bleached corals enrichedwith CP-580 contained remnant highly productivedinoflagellates rich in Chl. It is hypothesized that this ispossible because of the up-regulation of dinoflagellate ELiPanalogues and increased use of RC quenching to link active
Role of host pigments in coral symbioses 1531
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533

RCs to inactive RCs and transfer a large portion of theenergy harvested in the antennae to heat. As demonstratedby Pocock et al. (2007), RC quenching, despite the loss ofenergy to heat, is consistent with increased algal produc-tion. The identification and description of the Symbiod-inium equivalents of ELiP will be an important further stepin understanding the intricate photosynthetic behaviour ofreef-building corals and their dinoflagellate symbionts.
ACKNOWLEDGMENT
We are grateful to the Australian Research Council Centreof Excellence in Coral Reef Studies for funding this study.
REFERENCES
Adamska I. (2001) The Elip family of stress proteins in the thyla-kold membranes of pro- and eukaryota. In Advances in Photo-synthesis and Respiration; Regulation of Photosynthesis (edsE.M. Aro & B. Andersson) Vol. 11, pp. 487–515. Kluwer Aca-demic Publishers, Dordrecht, Boston, London.
Adamska I., Ohad I. & Kloppstech K. (1992) Synthesis of the earlylight-inducible protein is controlled by blue light and related tolight stress. Proceedings of the National Academy of Sciences ofthe United States of America 89, 2610–2613.
Anthony K.R.N. & Hoegh-Guldberg O. (2003a) Kinetics ofphotoacclimation in corals. Oecologia 134, 23–31.
Anthony K.R.N. & Hoegh-Guldberg O. (2003b) Variation in coralphotosynthesis, respiration and growth characteristics in con-trasting light microhabitats: an analogue to plants in forest gapsand understoreys? Functional Ecology 17, 246–259.
Anthony K.R.N., Connolly S.R. & Willis B.L. (2002) Comparativeanalysis of energy allocation to tissue and skeletal growth incorals. Limnology and Oceanography 47, 1417–1429.
Anthony K.R.N., Hoogenboom M.O. & Connolly S.R. (2005)Adaptive variation in coral geometry and the optimization ofinternal colony light climates. Functional Ecology 19, 17–26.
Barnes D.J. & Chalker B.E. (1990) Calcification and photosynthesisin reef-building corals and algae. In Coral Reefs, Ecosystem of theWorld 25 (ed. Z. Dubinsky) pp. 109–131. Elsevier, Amsterdam.
Berkelmans R. (2002) Time-integrated thermal bleaching thresh-olds of reefs and their variation on the Great Barrier Reef.Marine Ecology Progress Series 229, 73–82.
Berkelmans R. & van Oppen M.J.H. (2006) The role of zooxan-thellae in the thermal tolerance of corals: a ‘nugget of hope’ forcoral reefs in an era of climate change. Proceedings of the RoyalSociety B-Biological Sciences 273, 2305–2312.
Bruno A.K. & Wetzel C.M. (2004) The early light-inducible protein(ELIP) gene is expressed during the chloroplast-to-chromoplasttransition in ripening tomato fruit. Journal of ExperimentalBotany 55, 2541–2548.
Dove S.G., Hoegh-Guldberg O. & Ranganathan S. (2001) Majorcolour patterns of reef-building corals are due to a family ofGFP-like proteins. Coral Reefs 19, 197–204.
Dove S.G., Hoegh-Guldberg I.O., Prescott M., Jones E.L., KaranM., Brugliera F. & Mason J. (2002) Novel color-facilitating mol-ecule for producing a biomatrix, has a polypeptide which alone/along with molecules imparts altered visual characteristics tocells in the absence of excitation by extraneous non-white light.ed. W.I.P. Organisation p. 510. Patent No. WO200270703-A.
Dove S., Ortiz J.C., Enriquez S., Fine M., Fisher P., Iglesias-PrietoR., Thornhill D. & Hoegh-Guldberg O. (2006) Response of holo-symbiont pigments from the scleractinian coral Montipora
monasteriata to short-term heat stress. Limnology and Oceanog-raphy 51, 1149–1158.
Dubinsky Z., Falkowski P.G., Porter J.W. & Muscatine L. (1984)Absorption and utilization of radiant energy by light-adaptedand shade-adapted colonies of the hermatypic coral Stylophorapistillata. Proceedings of the Royal Society of London SeriesB-Biological Sciences 222, 203–214.
Dubinsky Z., Stambler N., Benzion M., McCloskey L.R., MuscatineL. & Falkowski P.G. (1990) The effect of external nutrientresources on the optical properties and photosynthetic efficiencyof Stylophora pistillata. Proceedings of the Royal Society ofLondon Series B-Biological Sciences 239, 231–246.
Enriquez S., Mendez E.R. & Iglesias-Prieto R. (2005) Multiplescattering on coral skeletons enhances light absorption by sym-biotic algae. Limnology and Oceanography 50, 1025–1032.
Falkowski P.G. & Dubinsky Z. (1981) Light-shade adaptation ofStylophora pistillata, a hermatypic coral from the Gulf of Eilat.Nature 289, 172–174.
Fitt W.K., McFarland F.K., Warner M.E. & Chilcoat G.C. (2000)Seasonal patterns of tissue biomass and densities of symbioticdinoflagellates in reef corals and relation to coral bleaching.Limnology and Oceanography 45, 677–685.
Gilmore A.M., Larkum A.W.D., Salih A., Itoh S., Shibata Y., BenaC., Yamasaki H., Papina M. & Van Woesik R. (2003) Simulta-neous time resolution of the emission spectra of fluorescentproteins and zooxanthellar chlorophyll in reef-building coral.Photochemistry and Photobiology 77, 515–523.
Gladfelter E.H. (1983) Spatial and temporal patterns of mitosis inthe cells of the axial polyp of the reef coral Acropora cervicornis.Biological Bulletin 165, 811–815.
Green B.R. & Durnford D.G. (1996) The chlorophyll-carotenoidproteins of oxygenic photosynthesis. Annual Review of PlantPhysiology and Plant Molecular Biology 47, 685–714.
Habuchi S., Ando R., Dedecker P., Verheijen W., Mizuno H.,Miyawaki A. & Hofkens J. (2005) Reversible single-moleculephotoswitching in the GFP-like fluorescent protein Dronpa. Pro-ceedings of the National Academy of Sciences of the United Statesof America 102, 9511–9516.
Hatcher B.G. (1988) Coral reef primary productivity: a beggar’sbanquet. Tree 3, 106–111.
Hoegh-Guldberg O. (1999) Climate change, coral bleaching andthe future of the world’s coral reefs. Marine and FreshwaterResearch 50, 839–866.
Hoegh-Guldberg O. & Smith G.J. (1989) The effect of suddenchanges in temperature, light and salinity on the population-density and export of zooxanthellae from the reef corals Stylo-phora pistillata Esper and Seriatopora hysterix Dana. Journal ofExperimental Marine Biology and Ecology 129, 279–303.
Iglesias-Prieto R. & Trench R.K. (1994) Acclimation and adapta-tion to irradiance in symbiotic dinoflagellates. 1. Responses ofthe photosynthetic unit to changes in photon flux-density.Marine Ecology Progress Series 113, 163–175.
Iglesias-Prieto R. & Trench R.K. (1997) Acclimation and adapta-tion to irradiance in symbiotic dinoflagellates. II. Response ofchlorophyll-protein complexes to different photon-flux densi-ties. Marine Biology 130, 23–33.
Iglesias-Prieto R., Beltran V.H., LaJeunesse T.C., Reyes-Bonilla H.& Thome P.E. (2004) Different algal symbionts explain the ver-tical distribution of dominant reef corals in the eastern Pacific.Proceedings of the Royal Society of London Series B-BiologicalSciences 271, 1757–1763.
Jassby A.D. & Platt T. (1976) Mathematical formulation of rela-tionship between photosynthesis and light for phytoplankton.Limnology and Oceanography 21, 540–547.
Jeffrey S.W. & Humphrey G.F. (1975) New spectrophotometricequations for determining chlorophylls a, B, C1 and C2 in higher
1532 S. G. Dove et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533

plants, algae and natural phytoplankton. Biochemie Und Physi-ologie Der Pflanzen 167, 191–194.
Kuhl M., Cohen Y., Dalsgaard T., Jorgensen B.B. & Revsbech N.P.(1995) Microenvironment and photosynthesis of zooxanthellaein scleractinian corals studied with microsensors for O2, pH andlight. Marine Ecology Progress Series 117, 159–172.
Levy O., Achituv Y., Yacobi Y.Z., Stambler N. & Dubinsky Z.(2006) The impact of spectral composition and light periodicityon the activity of two antioxidant enzymes (SOD and CAT) inthe coral Favia favus. Journal of Experimental Marine Biologyand Ecology 328, 35–46.
Lewis D.H. & Smith D.C. (1971) Autotrophic nutrition of symbi-otic marine coelenterates with special reference to hermatypiccorals. 1. Movement of photosynthetic products between sym-bionts. Proceedings of the Royal Society of London SeriesB-Biological Sciences 178, 111–129.
Little A.F., van Oppen M.J.H. & Willis B.L. (2004) Flexibility inalgal endosymbioses shapes growth in reef corals. Science 304,1492–1494.
Loram J.E., Trapido-Rosenthal H.G. & Douglas A.E. (2007)Functional significance of genetically different symbiotic algaeSymbiodinium in a coral reef symbiosis. Molecular Ecology 16,4849–4857.
Lukyanov K.A., Fradkov A.F., Gurskaya N.G., et al. (2000) Naturalanimal coloration can be determined by a nonfluorescent greenfluorescent protein homolog. The Journal of Biological Chemis-try 275, 25879–25882.
Matsubara S. & Chow W.S. (2004) Populations of photo inactivatedphotosystem II reaction centers characterized by chlorophylla fluorescence lifetime in vivo. Proceedings of the NationalAcademy of Sciences of the United States of America 101, 18234–18239.
Maxwell D.P., Falk S. & Huner N.P.A. (1995) Photosystem-II exci-tation pressure and development of resistance to photoinhibi-tion. 1. Light-harvesting complex-II abundance and zeaxanthincontent in Chlorella vulagaris. Plant Physiology 107, 687–694.
Mazel C.H., Lesser M.P., Gorbunov M.Y., Barry T.M., Farrell J.H.,Wyman K.D. & Falkowski P.G. (2003) Green-fluorescent pro-teins in Caribbean corals. Limnology and Oceanography 48,402–411.
Montane M.H. & Kloppstech K. (2000) The family of light-harvesting-related proteins (LHCs, ELIPs, HLIPs): was the har-vesting of light their primary function? Gene 258, 1–8.
Muscatine L. (1990) The role of symbiotic algae in carbon andenergy flux in reef corals. In Ecosystems of the World: Coral Reefs(ed. Z. Dubinsky) pp. 75–87. Elsevier, Amsterdam.
Niyogi K.K. (1999) Photoprotection revisited: genetic and molecu-lar approaches. Annual Review of Plant Physiology and PlantMolecular Biology 50, 333–359.
Pearcy R.W. & Yang W. (1998) The functional morphology of lightcapture and carbon gain in the Redwood forest understoreyplant Adenocaulon bicolor Hook. Functional Ecology 12, 543–552.
Platt T., Gallegos C.L. & Harrison W.G. (1980) Photoinhibition ofphotosynthesis in natural assemblages of marine phytoplankton.Journal of Marine Research 38, 687–701.
Pocock T., Sane P.V., Falk S. & Ner N.P.A. (2007) Excitation pres-sure regulates the activation energy for recombination events in
the photosystem II reaction centres of Chlamydomonas rein-hardtii. Biochemistry and Cell Biology 85, 721–729.
Prescott M., Ling M., Beddoe T., Oakley A.J., Dove S., Hoegh-Guldberg O., Devenish R.J. & Rossjohn J. (2003) The 2.2 Acrystal structure of a pocilloporin pigment reveals a nonplanarchromophore conformation. Structure 11, 275–284.
Rowan R. (2004) Coral bleaching – thermal adaptation in reefcoral symbionts. Nature 430, 742–742.
Rowan R., Knowlton N., Baker A. & Jara J. (1997) Landscapeecology of algal symbionts creates variation in episodes of coralbleaching. Nature 388, 265–269.
Salih A., Larkum A., Cox G., Kuhl M. & Hoegh-Guldberg O.(2000) Fluorescent pigments in corals are photoprotective.Nature 408, 850–853.
Sharrock R.A. & Clack T. (2002) Patterns of expression andnormalized levels of the five arabidopsis phytochromes. PlantPhysiology 130, 442–456.
Smith D.C. (1987) Regulation and change in symbiosis. Annals ofBotany 60, 115–127.
Smith H. (2000) Phytochromes and light signal perception byplants – an emerging synthesis. Nature 407, 585–591.
Sparrow J.R., Nakanishi K. & Parish C.A. (2000) The lipofuscinfluorophore A2E mediates blue light-induced damage to retinalpigmented epithelial cells. Investigative Ophthalmology & VisualScience 41, 1981–1989.
Suto D., Sato K., Ohba Y., Yoshimura T. & Fugii J. (2005) Suppres-sion of the pro-apoptotic function of cytochrome c by singletoxygen via a haem redox state-independent mechanism. TheBiochemical Journal 392, 399–406.
Takabayashi M. & Hoegh-Guldberg O. (1995) Ecological andphysiological differences between 2 color morphs of the coralPocillopora damicornis. Marine Biology 123, 705–714.
Turcic K., Pettikiriarachchi A., Battad J., Wilmann P.G., Rossjohn J.,Dove S.G., Devenish R.J. & Prescott M. (2006) Amino acidsubstitutions around the chromophore of the chromoproteinRtms5 influence polypeptide cleavage. Biochemical and Bio-physical Research Communications 340, 1139–1143.
Veron J.E.N. (1986) Corals of Australia and the Indo-Pacific. Angusand Robertson, Sydney.
Warner M.E., Chilcoat G.C., McFarland F.K. & Fitt W.K. (2002)Seasonal fluctuations in the photosynthetic capacity of photo-system II in symbiotic dinoflagellates in the Caribbean reef-building coral Montastraea. Marine Biology 141, 31–38.
Whitaker J.R. & Granum P.E. (1980) An absolute method forprotein determination based on difference in absorbance at 235and 280 nm. Analytical Biochemistry 109, 156–159.
Wilmann P.G., Battad J., Petersen J., Wilce M.C.J., Dove S.,Devenish R.J., Prescott M. & Rossjohn J. (2006) The 2.1 ang-strom crystal structure of copGFP, a representative member ofthe copepod clade within the green fluorescent protein super-family. Journal of Molecular Biology 359, 890–900.
Zapata M., Rodriguez F. & Garrido J.L. (2000) Separation of chlo-rophylls and carotenoids from marine phytoplankton: a newHPLC method using a reversed phase C-8 column and pyridine-containing mobile phases. Marine Ecology Progress Series 195,29–45.
Received 1 May 2008; revised in revised form 3 July 2008; acceptedfor publication 5 July 2008
Role of host pigments in coral symbioses 1533
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Plant, Cell and Environment, 31, 1523–1533
Related Documents











