Journal of Tropical Ecology http://journals.cambridge.org/TRO Additional services for Journal of Tropical Ecology: Email alerts: Click here Subscriptions: Click here Commercial reprints: Click here Terms of use : Click here Homerange size and territoriality in Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna William E. Magnusson, Angelo De Lima Francisco and Tania M. Sanaiotti Journal of Tropical Ecology / Volume 11 / Issue 02 / May 1995, pp 179 188 DOI: 10.1017/S0266467400008622, Published online: 10 July 2009 Link to this article: http://journals.cambridge.org/abstract_S0266467400008622 How to cite this article: William E. Magnusson, Angelo De Lima Francisco and Tania M. Sanaiotti (1995). Homerange size and territoriality in Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna. Journal of Tropical Ecology, 11, pp 179188 doi:10.1017/S0266467400008622 Request Permissions : Click here Downloaded from http://journals.cambridge.org/TRO, IP address: 155.97.150.99 on 18 Apr 2013

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Tropical Ecologyhttp://journals.cambridge.org/TRO
Additional services for Journal of Tropical Ecology:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Homerange size and territoriality in Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna
William E. Magnusson, Angelo De Lima Francisco and Tania M. Sanaiotti
Journal of Tropical Ecology / Volume 11 / Issue 02 / May 1995, pp 179 188DOI: 10.1017/S0266467400008622, Published online: 10 July 2009
Link to this article: http://journals.cambridge.org/abstract_S0266467400008622
How to cite this article:William E. Magnusson, Angelo De Lima Francisco and Tania M. Sanaiotti (1995). Homerange size and territoriality in Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna. Journal of Tropical Ecology, 11, pp 179188 doi:10.1017/S0266467400008622
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/TRO, IP address: 155.97.150.99 on 18 Apr 2013

Journal of Tropical Ecology (1995) 11:179-188. With 3 figures
Home-range size and territoriality in Bolomyslasiurus (Rodentia: Muridae) in an Amazoniansavanna
WILLIAM E. MAGNUSSON, ANGELO DE LIMA FRANCISCO andTANIA M. SANAIOTTI
Departamento de Ecologia, Institute Nacional de Pesquisas da Amazonia, Caixa Postal478, 69011-970 Manaus AM,, Brasil
ABSTRACT. Bolomys lasiurus is the only species of rodent commonly found in the savanna of Alterdo Chao, central Amazonia. Peak densities during 42 months were less than have been reportedfrom the cerrado biome of central Brazil. However, the population at Alter do Chao appears tobe more stable than those in other areas and we never recorded zero densities. Many animals werecaptured over periods greater than 7 months, indicating that either mortality or dispersal was lessthan in previous studies. Mean home-range size of females was inversely related to the proportionof females that were gravid or lactating. Female home ranges were mutually exclusive and averaged35% (SD = 8.2%) of male home ranges. Male home ranges overlapped extensively. Indices ofhome-range size varied seasonally and among years.
KEY WORDS: behaviour, mammal, sex, population dynamics
Bolomys lasiurus has a wide distribution in South America and is the most fre-quently captured rodent in many situations (Alho 1982). The species has beenstudied in the cerrados (savannas) of Goias (Melo 1980), Minas Gerais (Valleet al. 1982) and the Federal District (Pereira 1982, Souza & Alho 1980) ofcentral Brazil. These studies indicate that, although the species is abundant,populations fluctuate markedly and may disappear from trapping grids. Thereare no studies of the population dynamics of the species in Amazonian-typesavanna (Huber 1982), and no study covered more than two years.
Macedo & Mares (1987) reported that the population of B. lasiurus nearBelem in eastern Amazonia is morphologically distinct from populations furthersouth. Also, Alho (1982) reported that the species is primarily nocturnal,whereas our observations indicate that B. lasiurus is diurnal and crepuscular inan Amazonian savanna near Alter do Chao. (Lat. 2°31'S, Long.55° 00' W). In this paper, we report on aspects of the use of space andpopulation turnover of the population near Alter do Chao during a period of42 months. Information on growth and reproduction in this population hasbeen presented elsewhere (Francisco et al., in press).
179

180 WILLIAM E. MAGNUSSON ET AL.
METHODS
The study area has been described by Miranda (1991). It is in an area of sandysoils within an Amazonian savanna. The site does not flood and, even afterheavy rains, there is no standing water. Grassland, clumps of shrubs and treesare distributed fairly homogeneously through the savanna; any point in the areacan be expected to be within 10 m of open grassland (Paspalum carinatumFluegge), clumps of shrubs (mainly Myrcia spp.) and trees (mainly Qualea grandi-flora Mart.).
Rats were captured in Sherman (TM) live traps baited with Brazil nuts anda mixture of oat meal and peanut butter. From July 1985 to December 1985,
the traps were set on a 150 m by 200 m grid with a minimum spacing of 10 mbetween capture points. Traps were set uniformly over the whole area on 25%of the grid intersections for 3 or 4 days and then moved one trap position untilthe whole grid had been covered. The method is a modification of the movingtransect method described by Alho (1979a,b) and similar to that used by Pereira(1982). Moving the traps within sessions reduces their influence on the areaused by the rats. We believe that animals that became 'trap happy' may haveconcentrated their activities within part of their home range but it is unlikelythat traps enticed rats out of their home ranges as many other traps wereavailable within each home range.
As most animals were captured within the first two days at each trap position,and animals became trap happy and increased markedly in weight over thetrapping period, we reduced the number of days at each trap position to twoin subsequent trapping sessions. From February 1986 to June 1987, weincreased the grid size to 200 m by 200 m and increased the number of trapsto maintain 25% coverage of the grid per day. The grid was increased to 340 mby 300 m in February 1988 until the end of the study in January 1989. Thegeneral protocol remained the same but the traps were set on a grid with aminimum spacing of 20 m between capture points.
Traps were left open from 16:00-18:00 h and 06:00-09:00 h. During prelimin-ary studies, few rats entered traps after 19:00 h. Traps were not left open duringmost of the daylight hours because rats that were captured became heat stressedand some died.
Sampling sessions for the whole period are shown in Figure 1. Rats weremarked by toe clipping with sharp scissors. They did not appear stessed by theprocedure and usually re-entered traps the following morning or afternoon. Themethod has little effect on Microtus ochrogaster (Wood & Slade 1990) and noother method that would last more than 12 months was available. At the firstcapture in each session, the rats were weighed with 100 g spring balances accur-ate to 0.5 g.
The area had not been burned for several years before the study. However,local residents burned the area in September 1986 and September 1987. Weburned fire breaks around the area in September 1988 (Sanaiotti & Magnusson1995).
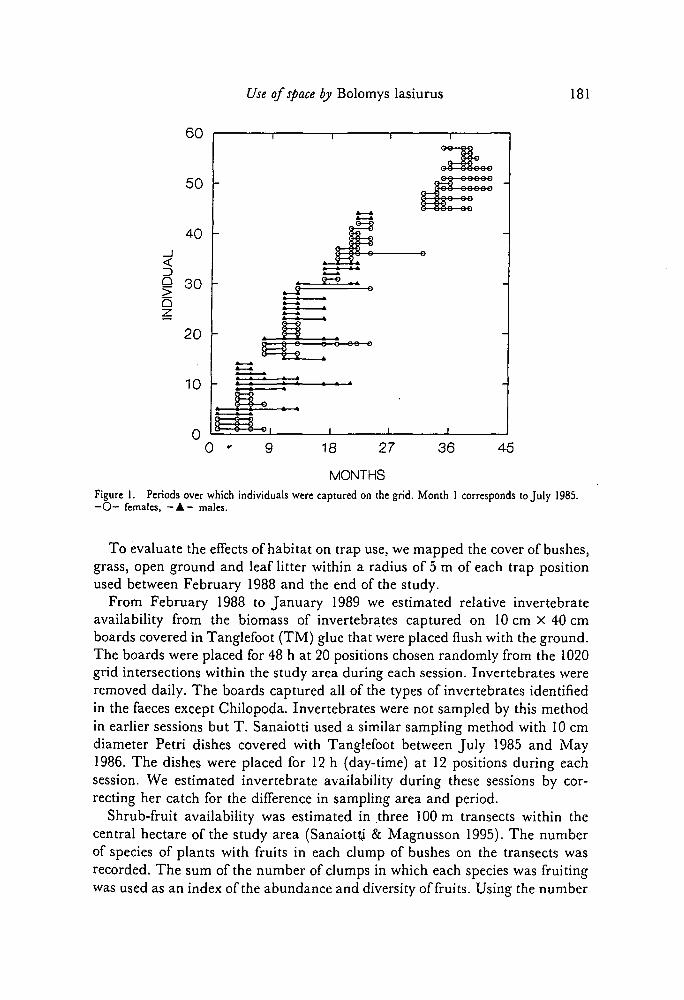
Use of space by Bolomys lasiurus 181
0 '
MONTHSFigure 1. Periods over which individuals were captured on the grid. Month 1 corresponds to July 1985.—O— females, — A - males.
To evaluate the effects of habitat on trap use, we mapped the cover of bushes,grass, open ground and leaf litter within a radius of 5 m of each trap positionused between February 1988 and the end of the study.
From February 1988 to January 1989 we estimated relative invertebrateavailability from the biomass of invertebrates captured on 10 cm X 40 cmboards covered in Tanglefoot (TM) glue that were placed flush with the ground.The boards were placed for 48 h at 20 positions chosen randomly from the 1020grid intersections within the study area during each session. Invertebrates wereremoved daily. The boards captured all of the types of invertebrates identifiedin the faeces except Chilopoda. Invertebrates were not sampled by this methodin earlier sessions but T. Sanaiotti used a similar sampling method with 10 cmdiameter Petri dishes covered with Tanglefoot between July 1985 and May1986. The dishes were placed for 12 h (day-time) at 12 positions during eachsession. We estimated invertebrate availability during these sessions by cor-recting her catch for the difference in sampling area and period.
Shrub-fruit availability was estimated in three 100 m transects within thecentral hectare of the study area (Sanaiotti & Magnusson 1995). The numberof species of plants with fruits in each clump of bushes on the transects wasrecorded. The sum of the number of clumps in which each species was fruitingwas used as an index of the abundance and diversity of fruits. Using the number

182 WILLIAM E. MAGNUSSON ET AL.
of species fruiting or the number of clumps with any species fruiting gave asimilar pattern of monthly variation. Most of the fruits in the clumps of shrubswere fleshy. Fleshy-fruited tree species followed a similar pattern (Miranda1991).
We used the number of individual trees with wind-dispersed fruits that werefruiting in the central hectare of the study area in each period as an index ofthe availability of dry tree fruits. Dry fruits of the grassland were not quantified.
Rainfall records for Santarem airport, 8 km distant, were supplied by theComandante do Destacamento da Aeronautica de Santarem, Para.
Analyses of the factors affecting home-range size are based on means forvariables in each collecting period. Therefore, there is no pseudoreplication dueto the inclusion of several animals whose home ranges were measured underthe same conditions. However, as many animals survived from one period tothe next (Figure 1), mean values of home-range size are not necessarily inde-pendent of each other among periods. The validity of the probability valuespresented depends on the validity of our assumption that the effect of individualvariation is small compared with seasonal variation. Readers should take thisinto account when making their own conclusions.
Statistical tests were carried out on the SYSTAT statistical program(Wilkinson 1990). Data for males from February 1988 on were lost in a laborat-ory accident. Therefore, analyses over extensive time periods could only bemade for females.
RESULTS
Bolomys lasiurus was the only small mammal captured regularly on the grid. Afew Proechimys sp. and juvenile Didelphis marsupialis were caught but they werenot recaptured. Both species are common in the forest patches around thesavanna. Only one juvenile B. lasiurus was captured in the forest despite extens-ive trapping.
We generally caught more animals per trap session before the first fire(X = 28.6, SE = 4.7, N = 5) than afterwards (X = 16.9, SE = 1.2, N = 14).The difference is statistically significant (FM7 = 12.0, P = 0.003) even thoughthe size of the grid was much larger for most of the second period.
Nine animals were recaptured over periods in excess of 7 months (Figure 1).For animals captured in more than one capture session, there was no significantdifference in the period between first and last capture for males and females(Fi,42 = 0.98, P = 0.32), nor was there any significant difference in the propor-tion of males (N = 29) to females (N = 26) that were captured in only onecapture session (X, = 0.16, P > 0.5). The longest residence period on the gridwas 17 months for males and 16 months for females.
Home ranges of animals captured in more than one trapping session, andwhich were not captured in traps on the perimeter of the grid, were estimatedfrom the area of a minimum convex polygon that included all the sites at which

Use of space by Bolomys lasiurus 183
they were captured. No correction was made for the distance between traps orthe number of captures (Figure 2).
Home-range size varied between years and between sexes (2-way ANOVA:Year: F,,14 = 19.35, P = 0.001; Sex: F2,14 = 3.81, P = 0.048). It was not pos-sible to include data for 1988 in this analysis because data for males were notavailable for that year. However, home ranges of females were significantlylarger in 1988 than in the other three years (Fisher's Least-Significant-Difference test, P < 0.05 in each case). There were insufficient years to teststatistically for the cause of the differences between years. However, variationin methodology did not appear to be significant. Mean yearly home-range sizedid not covary with the grid size, the number of days per capture session, orthe mean number of captures per animal. There was also no obvious relation-ship between home-range size and the presence of fire in that year.
The presence of large differences between years makes the mean home-rangesize of the population a nebulous concept. The mean home-range size for malesvaried from 4138 m2 in 1987 to 21,880 m2 in 1988 and for females varied from1317 m2 to 5480 m2 in the same years. The mean yearly home-range size washighly correlated between the sexes (r = 0.97, P = 0.029). Mean yearly femalehome ranges averaged 35% (SD = 8.2%) those of males but this probablyunderestimates the true difference as there was a high probability of the largesthome ranges overlapping the perimeter of the study area. The mean home-rangesize for males in 1988 was 21,880 m2 but one male, whose home range over-lapped the perimeter of the study area, used 51,000 m2 within the grid.
Home ranges varied within years (Figure 3). Between February 1988 andOctober 1988, five females disappeared from the grid (individuals B, C, G, Fand O), three retained their home ranges (E, D and A) and two occupied partsof home ranges that had been left vacant (C and G). To investigate seasonalvariation in home-range size, we analysed the mean home-range size of females.The means are not completely independent as females frequently survive fromone trapping period to the next (Figure 1). However, there is no other way toinvestigate seasonal trends with these data.
There was a significant interaction between the effects of the presence offire and population reproductive activity (% of females gravid or lactating)(ANCOVA: P = 0.026). Therefore, we analysed the data before and after thefirst fire separately. There was no relationship between the mean size of monthlyranges and reproductive activity before the first fire (r2 = 0.16, P = 0.333),but monthly-range size (HR - m2) was negatively correlated with reproductiveactivity (RA - %) after the first fire (HR = 16.6 - 0.14RA, r2 = 0.74,Fli9 = 10.5, P = 0.010).
Multiple regression indicated, that neither rainfall, fleshy-fruit diversity, avail-ability of dry tree fruits or insect availability contribute significantly to monthly-range size before or after the first fire or for the two periods combined (P > 0.1in each case). Analysis of residuals indicated that the assumptions of the analysishad been met and fishing for combinations of factors revealed no probability values<0.05 except for reproductive activity, so the result appears to be robust.

184 WILLIAM E. MAGNUSSON ET AL.
"'%'/' K-Y * " ' " " • • \ '
- J k ^ ^ ^ ^ v B\ * 1^ 0 0 0 1? 0 „
o ^ . ^ '
1 ' • • " ^ 1 1
}, B - b
C 0 1
0 |
« H
Q H
G
3-lf,1
'̂
' E
/
, 8
-cr -°-0 {?'"'f , ,
/ ' F ;'
^ / '\ . '
, \
v'
i
iiier
-s-0 ,c 1c c,
r
c
i
e -
0 B I
0 50 100 150 200 0 50 100 150 200
Figure 2. Home ranges of females (a) and males (b) between July and December 1985. During this periodthe grid was 150 m by 200 m. Home ranges of females (c) and males (d) between February and June 1986.During this period the grid was 200 m by 200 m. The broken line (c) indicates the home range of female ' Cin the month of June when female 'B' was not captured. Home ranges of females (e) and males (f) betweenNovember 1986 and April 1987; during this period the grid was 200 m by 200 m..
The large yearly variation in home-range size could mask variation in theproportion of the home-range used by individuals. However, analyses of themean proportion of the total home range used by females in each month gaveresults qualitatively identical to those for home-range size. We therefore

Use of space by Bolomys lasiurus 185
conclude that reproduction reduces home-range size in females within years butwe are unable to explain the changes in home-range sizes between years.
Trap sites with greater cover of grass and bushes tended to have highernumbers of captures (Table 1). However, the cover of vegetation within 10 mof the trap explained only 11 % of the variance in capture frequency. Analysesfor adult males, adult females and juveniles separately gave similar results. Ifthe rats respond to vegetation structure, it must be on a scale much less thanthe 10 m diameter plots that we used. This agrees with the apparently randomplacement of home ranges between years (Figure 2).
The distribution of home ranges indicates that females are strongly territorial.The home ranges of females that were captured in more than one capturesession (presumed residents) were generally exclusive (Figures 3, 2a,c,e). It isnot possible to compare home ranges over periods greater than 3-5 months
Table 1. Statistical summary of multiple regression relating the number of rats captured at a site to theproportion of grass (GR), open areas (OA), bushes (BU) and ground covered by leaves (LE) within a 10 mradius of the trap position (R? = 0.11, N = 280, F«,,75 = 8.25, P = 0.001).
Variable Coefficient Tolerance
ConstantOABUGRFOL
0.4780.7602.0281.0850.011
0.5880.8360.6940.629
1.661-0.956
4.5693.043
-0.014
0.0980.3400.0000.0030.989
300
200 -
100 -
200 300
Figure 3. Home ranges of females on the grid between February 1988 and June 1988 (a) and between July1988 and January 1989 (b). Animals 'A', 'D' and 'E' occupied home ranges on the grid in both periods.Other home ranges are specific to one period. During these periods the grid was 340 m by 300 m.

186 WILLIAM E. MAGNUSSON ET AL.
because of the high population turnover. Even within these short periods,females adjust their home ranges if a neighbour disappears. For example,between May and July 1986 female 'B' and ' C had mutually exclusive homeranges (Figure 2c, solid outlines). However, when female 'B' disappeared fromthe area in August 1986, female ' C increased her home range to include muchof the home range of 'B' (broken line).
The only exception was female 'O' (Fig. 3a, broken line). She was capturedover a period of 6 months in five capture sessions, always within the homeranges of other females. Why this female was tolerated within, or was able toinvade, other home ranges is not clear but it may be significant that she wasnever recorded as gravid or lactating.
In contrast, home ranges of males overlapped extensively in all years ofthe study (Figure 2b,d,f). Their use of space offers no evidence that they areterritorial.
DISCUSSION
Bolomys lasiurus is morphologically and behaviourally convergent on northernhemisphere voles of the genus Microtus (Microtinae). It inhabits grassland, hasa short tail and ears, small eyes and is mainly diurnal at Alter do Chao. How-ever, like other South American cricetines, it is much more insectivorous thanthe true voles (Glanz 1982). Males have large overlapping home ranges andfemales have smaller, largely exclusive, home ranges. Omnivorous voles showthe same differences between the sexes in the use of space (Ostfeld 1985).Females can probably maximize their reproductive potential by defending foodsources, whereas males maximize their reproductive potential by maintainingcontact with many more females than they can defend (Ostfeld 1985). Malehome ranges averaged almost three times the size of females, a difference consid-erably greater than the X 1.5-1.6 reported for Microtus ochrogaster (Swihart &Slade 1989) or X2 for M. californicus (Ostfeld 1986). As with voles whose use ofspace indicates that males are sexually promiscuous (Heske & Ostfeld 1990),male B. lasiurus reach larger sizes than non-gravid females (Francisco et al., inpress).
Pavone & Boonstra (1985) reported that toe clipping affected survival of malebut not female meadow voles (Microtus pennsylvanicus). However, our data donot indicate significant differences in survival between males and females.
This study used methods different from those used in central Brazil (Melo1980, Pereira 1982, Souza & Alho 1980, Valle et al. 1982). However, there aredistinct differences in the population dynamics between the two areas thattranscend differences due to methodology. The populations on the grids studiedby Melo (1980), Pereira (1982) and Souza & Alho (1980) went to very lowdensities (apparently zero) several times during their studies. Valle et al. (1982)recorded only two animals during the last month of their study. The maximumdensities recorded in central Brazil (21 animals on 1.5 ha (Melo 1980), 18

Use of space by Bolomys lasiurus 187
animals on 1 ha (Souza & Alho 1980), 20 animals on 1.2 ha (Valle et al. 1982)and ~200 animals on 16 ha (Pereira 1982) were far higher than the maximumdensity recorded in this study (42 animals on 10 ha). The density of rats on thegrid at Alter do Chao apparently was higher before the area was burned at themiddle of the study but the reduction in density was only about 40%. We neverrecorded an apparent density of zero. Possible reasons for these differencesinclude the number of syntopic rodents (zero at Alter do Chao; up to 12 inGoias), climate and differences in vegetation composition (Miranda 1991).However, we have no data to differentiate between these hypotheses. The inter-vals between first and last captures were short (4 and 7 months respectively)in the studies by Melo (1980) and Pereira (1982). In contrast, at Alter do Chao,many animals were captured over periods greater than 7 months and someremained on the grid for over a year. Either mortality or dispersal or both mustbe much lower at Alter do Chao than at the sites studied by Melo (1980) andPereira (1982).
Average home-range size varied between and within years. We were unableto associate these changes with the presence of fire, rainfall or fruit production.Home-range size of voles may be influenced by population density (Erlinge etal. 1990, Mazurkiewicz 1971). However, it was not related to the number ofrats resident on the grid in our study. Part of the seasonal variation appears tobe related to reproduction. Mean home-range size of females was inverselyrelated to the proportion of females that were gravid or lactating. Erlinge et al.(1990) found the opposite relationship for Microtus agrestis.
Bolomys lasiurus has a typically vole-like social system: females with smallexclusive home ranges and males with large overlapping home ranges. However,all other aspects of its behaviour and population dynamics that have beeninvestigated appear to vary geographically and/or temporally. Therefore, itwould appear unwise to try to make generalizations about its biology with thedata presently available.
LITERATURE CITED
ALHO, C. J. R. 1979a. Relative exclusiveness of space as a measure of spatial distribution of mammalpopulations. Rcvista BrasiUira de Biologia 39:275—299.
ALHO, C. J . R. 1979b. The application of a technique to survey small mammal populations underecological circumstances. Revista BrasiUira de Biologia 39:597-600.
ALHO, C. J. R. 1982. Brazilian rodents: their habitats and habits. Pp. 143-166 in Mares, M. A. &Genoways, H. H. (eds). Mammalian biology in South America. University of Pittsburgh, Pittsburgh. 539 pp.
ERLINGE, S., HOOGENBOOM, I., AGRELL, J., NELSON, J . & SANDELL, M. 1990. Densityrelated home-range size and overlap in adult field voles (Microtus agrestis) in southern Sweden. Journalof Mammalogy 71:597-603.
FRANCISCO, A. D. L., MAGNUSSON, W. E. & SANAIOTTI, T. M. In press. Variation in growthand reproduction of Bolomys lasiurus (Rodentia: Muridae) in an Amazonian savanna. Journal of TropicalEcology.
GLANZ, W. E. 1982. Adaptive zones of neotropical mammals: a comparison of some temperate andtropical patterns. Pp. 95-101 in Mares, M. A. & Genoways, H. H. (eds). Mammalian Biology in SouthAmerica. University of Pittsburgh, Pittsburgh. 539 pp.
HESKE, E. J . & OSTFELD, R. S. 1990. Sexual dimorphism in size, relative size of testes, and matingsystems in North American voles. Journal of Mammalogy 71(4):510-519.

188 W I L L I A M E. MAGNUSSON ET AL.
HUBER, O. 1982. Significance of savanna vegetation in the Amazon Territory of Venezuela. Pp. 221—224 in Prance, G. T. (ed.). Biological diversification in the tropics. Columbia University Press, New York.741 pp.
MACEDO, R. H. & MARES, M. A. 1987. Geographic variation in the South American cricetine rodentBolomys lasiurus. Journal of Mammalogy 68:578-594.
MAZURKIEWICZ, M. 1971. Shape, size and distribution of home ranges of Clethrionomys glareolus(Schreber 1780). Ada Theriologica 16:23-60.
MELO, D. A. 1980. Estudo populacional de algumas especies de roedores do cerrado (norte do munici'piode Formosa, Goias). Revista Brasileira de Biologia 40:843-860.
MIRANDA, I. DE S. 1991. Estrutura e Fenologia de uma Comunidade Arbdrca da Savana Amazonica de Alterdo Chao, Para. Master's Thesis, Instituto Nacional de Pesquisas da Amazonia/Universidade Federaldo Amazonas. Manaus. 129 pp.
OSTFELD, R. S. 1985. Limiting resources and territoriality in microtine rodents. American Naturalist1 2 6 ( 1 ) 5
OSTFELD, R. S. 1986. Territoriality and mating system of California voles. Journal of Animal Ecology55:691-706.
PAVONE, L. V. & BOONSTRA, R. 1985. The effects of toe clipping on the survival of the meadowvole (Microtus pennsylvanicus). Canadian Journal of Zoology 63:499-501.
PEREIRA, L. A. 1982. Uso ecologico do espaco de Zygodontomys lasiurus (Rodentia, Cricetinae) em habitatnatural de cerrado do Brasil central. Master's Thesis, Uniyersidade de Brasilia, Brasilia. 128 pp.
SANAIOTTI, T. M. &. MAGNUSSON, W. E. 1995. Effects of annual fires on the production of fleshyfruits used by birds in Brazilian Amazonian savanna. Journal of Tropical Ecology 11:53-65.
SOUZA, M. J. & ALHO, C. J . R. 1980. Distribuicao espacial do roedor silvestre Zygodontomys lasiurusem habitat natural do cerrado. Brasil Florestal 44:31-74.
SWIHART, R. K. & SLADE, N. A. 1989. Differences in home-range size between sexes of Microtusochrogaster. Journal of Mammalogy 70:816-820.
VALLE, C. M. C , ALVES, M. C , SANTOS, I. B. & VAREJAO, J. B. M. 1982. Obscrvacao sobredinamica de populacao de Zygodonlomys lasiurus (Lund, 1841), Colomys expulsus (Lund, 1841) e Oryzomyssubjkvus (Wagner, 1842) em vegetac,ao de cerrado no Vale do Rio das Velhas (Prudente de Morais,Minas Gerais, Brasil - 1981) Rodentia = Cricetidae. Lundiana 1982(2):70-83.
WILKINSON, L. 1990. SYSTAT. The system for statistics. SVSTAT Inc., Evanston, Illinois, USA.WOOD, M. D. & SLADE, N. A. 1990. Comparison of ear-tagging and toe-clipping in Prairie voles,
Microtus ochrogaster. Journal of Mammalogy 7:252-255.
Accepted 31 March 1994
Related Documents








![From Personalism to Territoriality - [email protected] Home](https://static.cupdf.com/doc/110x72/61fb40b32e268c58cd5bf937/from-personalism-to-territoriality-emailprotected-home.jpg)


