ELSEVIER HLA Allele and Haplotype Frequencies in Algerians Relatedness to Spaniards and Basques Antonio Arnaiz-Villena, Djamal Benmamar, Miguel Alvarez, Nieves Diaz-Campos, Pilar Varela, Eduardo Gomez-Casado, and Jorge Martinez-Las0 ABSTRACT: The powerful genetic polymorphism of the HLA system has been used to identify individuals and populations. Ethnic groups may be characterized by spe- cific HLA allele frequencies and particular extended HLA haplotypes; also, genetic relationships among these groups may be deduced. In the present study, serology and DNA typing were used to detect HLA-A, -B, -C, -DR, and -DQ alleles in each individual and to calculate characteristic haplotypes in Algerians. These results were compared to those previously obtained in other popula- tions, particularly northern Mediterraneans; genetic dis- tances and their respective dendrograms place Basques and Spaniards closer to Algerians than to other Europeans. Also, characteristic Basque and/or Spanish haplotypes are found in Algerians; i.e., A3O-Bl8-CwT-DR3-DQ2 and A 1 -B57-Cw7-DR7-DQ2. This supports the evidence that the Algerian population, mainly its paleo-North African component (Berbers), has a common descent with Basques and Spaniards, probably reflecting a preneolithic relation- ship between Iberians and paleo-North Africans. Human Immunology 43) 259-268 (1995) ABBREVIATIONS :; haplotype frequency neighbor-joining RLD SGD relative linkage disequilibria standard genetic distance INTRODUCTION The population of Africa in pteneolithic times (about 7000-3000 years BC) has been classified into live main ethnic groups: (1) Hamites (Hamite-speaking people) at the Mediterranean and Red Sea coasts, they were white people who have been further subdivided into Berbers and Egyptians (approximately n = 100,000) and whitc- black Ethiopians (n = 100,000); (2) blacks at the Guinea Golf (n = 250,000); (3) black Nilo-Saharians (n = 250,000), surrounding the present-day Sahara Desert and Nilo River; (4) black Pygmies at southwestern Af- rican coasts (n = 200,000); and (5) black Bushmen at south and southeast coasts (n = 350,000) (see Fig. 1A) El}. In Pharaonic Africa (3000 years BC) population numbers had suffered drastic changes. Agricultural Egypt had 1 million people and the rest of Africa (non- agricultural hunters) had 1,200,OOO altogether. By 400 years AD, blacks from Guinea Golf had overwhelmed central and southern Africa, reduced Bushmen, and al- most eliminated Pygmies (see Fig. 1B) El]. From the Department of Immunology, University Complutense, Hospital 12 de Octubre, Cawetera Anablucia, Madrid, Spain. Address reprint requests to A. Arnaiz-Villena, Departamento de Inmu- nologia, Universihd Comphtense, Hospital I2 de Octthe, Cawetera An- Llucia, 28041 Madrid, Spain. Drastic climatic changes had dried northern Africa by around 8000-4000 years BC; the forest line had gone down towards the Equator from about parallel 16 by 3000 years BC (Fig. 1C) 121. By then the Sahara Desert had similar characteristics to the present-day Sahara and strong migrations towards the north sent people to the Iberian Peninsula and Mediterranean and Canary Islands; the first Iberians and Canary Islands “guanches” may in part come from these displaced paleo-North African in- habitants (Hamite-Berbers) or be closely related (33. Receivedjuly 19, 1994; accepted February 6, 1995. The population of Algeria has followed the general Maghteb demographic movements El). In neolithic Human Immunology 43, 259-268 (1995) 0198-8859l95lS9.50 0 American Society for Histocompatibility and Immunogenetics, 1995 SSDI 0198-8859(95)00024-X

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER
HLA Allele and Haplotype Frequencies in Algerians Relatedness to Spaniards and Basques
Antonio Arnaiz-Villena, Djamal Benmamar, Miguel Alvarez, Nieves Diaz-Campos, Pilar Varela, Eduardo Gomez-Casado, and Jorge Martinez-Las0
ABSTRACT: The powerful genetic polymorphism of the HLA system has been used to identify individuals and populations. Ethnic groups may be characterized by spe- cific HLA allele frequencies and particular extended HLA haplotypes; also, genetic relationships among these groups may be deduced. In the present study, serology and DNA typing were used to detect HLA-A, -B, -C, -DR, and -DQ alleles in each individual and to calculate characteristic haplotypes in Algerians. These results were compared to those previously obtained in other popula- tions, particularly northern Mediterraneans; genetic dis-
tances and their respective dendrograms place Basques and Spaniards closer to Algerians than to other Europeans. Also, characteristic Basque and/or Spanish haplotypes are found in Algerians; i.e., A3O-Bl8-CwT-DR3-DQ2 and A 1 -B57-Cw7-DR7-DQ2. This supports the evidence that the Algerian population, mainly its paleo-North African component (Berbers), has a common descent with Basques and Spaniards, probably reflecting a preneolithic relation- ship between Iberians and paleo-North Africans. Human Immunology 43) 259-268 (1995)
ABBREVIATIONS
:; haplotype frequency neighbor-joining
RLD SGD
relative linkage disequilibria standard genetic distance
INTRODUCTION
The population of Africa in pteneolithic times (about
7000-3000 years BC) has been classified into live main ethnic groups: (1) Hamites (Hamite-speaking people) at
the Mediterranean and Red Sea coasts, they were white
people who have been further subdivided into Berbers
and Egyptians (approximately n = 100,000) and whitc- black Ethiopians (n = 100,000); (2) blacks at the
Guinea Golf (n = 250,000); (3) black Nilo-Saharians (n
= 250,000), surrounding the present-day Sahara Desert and Nilo River; (4) black Pygmies at southwestern Af- rican coasts (n = 200,000); and (5) black Bushmen at
south and southeast coasts (n = 350,000) (see Fig. 1A)
El}. In Pharaonic Africa (3000 years BC) population
numbers had suffered drastic changes. Agricultural Egypt had 1 million people and the rest of Africa (non-
agricultural hunters) had 1,200,OOO altogether. By 400
years AD, blacks from Guinea Golf had overwhelmed
central and southern Africa, reduced Bushmen, and al-
most eliminated Pygmies (see Fig. 1B) El].
From the Department of Immunology, University Complutense, Hospital 12 de Octubre, Cawetera Anablucia, Madrid, Spain.
Address reprint requests to A. Arnaiz-Villena, Departamento de Inmu- nologia, Universihd Comphtense, Hospital I2 de Octthe, Cawetera An- Llucia, 28041 Madrid, Spain.
Drastic climatic changes had dried northern Africa by
around 8000-4000 years BC; the forest line had gone down towards the Equator from about parallel 16 by
3000 years BC (Fig. 1C) 121. By then the Sahara Desert had similar characteristics to the present-day Sahara and
strong migrations towards the north sent people to the Iberian Peninsula and Mediterranean and Canary Islands; the first Iberians and Canary Islands “guanches” may in part come from these displaced paleo-North African in-
habitants (Hamite-Berbers) or be closely related (33.
Receivedjuly 19, 1994; accepted February 6, 1995.
The population of Algeria has followed the general Maghteb demographic movements El). In neolithic
Human Immunology 43, 259-268 (1995) 0198-8859l95lS9.50 0 American Society for Histocompatibility and Immunogenetics, 1995 SSDI 0198-8859(95)00024-X

260
FIGURE 1 African Population changes between 3000 years BC and 400 years AD. (A) Ethnic groups in Africa 3000 years BC. (B) Ethnic groups in Africa 400 years AD. (C) Line of jungle displacement between 8000 and 1000 years BC.
times a scattering of Berber pastoralists and cultivators (a
few hundred thousand) existed there. However, they re- mained stuck at a simple neolithic level while other Mediterraneans were evolving through the Bronze and Iron Ages. By 1000 years BC, Phoenicians from Lebanon found a stone age neolithic culture in Maghreb. They set up Carthage (Tunisia) and were overthrown by the Ro- mans in 146 BC. At this time there were 100,000 Phoe-
nicians and 500,000 Berbers in Tunisia plus another 2.5 million Berbers in the rest of North Africa. By 200 AD
Algeria had no more than 2 million people, which were slowly going into agriculture due to Roman rule. During the seventh century Arabs invaded Algeria and mixed
A. Arnaiz-Villena et al.
with Berbers at a rate of about 40% Arab and 60%
Berber [4}. Culturally and demographically Algeria (and
Maghreb) has been stuck until very recently, when France and Spain left their colonies. The Algerian pop-
ulation was 9 million in 1950 and is more than 25
million at present.
One of the problems of Mediterranean ethnography has been the lack of genetic marker data coming from
southern Mediterranean populations [5]. The powerful and discriminating HLA polymorphism [3} has not been
studied in the Algerian population even in the last In-
ternational Workshop anthropological section 161, where
a few French Algerians were studied only by serology and
complete data were not shown. This has led to a lack of comparative genetic studies between northern and south-
ern Mediterraneans.
In the present work, we tried to fulfill this gap by
collecting blood samples from unrelated Algerians at Al- giers and testing them for HLA-A, -B, -DR, and -DQ
antigens both by serology and DNA sequencing. We also compared our data with that of Europeans, partic-
ularly Spaniards, Basques, and Sardinians, obtaining the corresponding genetic distances and a relatedness den-
drogram .
MATERIALS AND METHODS
Population Sample
One hundred six unrelated healthy Algerian blood do- nors were randomly selected as a representative panel of
this population. Blood was collected in Algiers, Hopital Central de I’ArmCe, Blood Bank. Peripheral blood lym-
phocytes were isolated by Ficoll gradient, frozen down,
and kept in liquid nitrogen until use in Madrid. Subjects did not have recorded admixture with Europeans over at
least three generations.
HLA Class I and II Serotyping
HLA class I (A, B, C) and class II (DR, DQ) typing was done by a two-step microlymphocytotoxicity technique
on T or B lymphocytes E71, respectively, using the 10th and 1 lth International Histocompatibility Workshop
and local reagents.
HLA-DRBl, DRB3, DRB4, DQAl, and DQBl Exon-2 Sequences
DNA extraction. Genomic DNA from whole blood con- taining EDTA was extracted with a 340A Nucleic Acid
Extractor (Applied Biosystems, Foster City, CA, USA) with the use of ABI reagents and protocols 181.
Amplification of genomic DNA. A generic HLA-DRB am- plification as well as group-specific amplifications of the DR 1, DR2, DR4, DRB 1 alleles associated with DR52 and DRB3 were made. The DRB 1, DRB3, DQAl, and

HLA in Algerians 261
DQBl exon-2 DNA was amplified by polymerase chain
reaction (PCR) using Taq polymerase (Cetus, USA), as
previously described 19). Primers and oligonucleotides for DRBl, DRB3, DRB4, DQAl, and DQBl were
those described by Kimura and Sasatuki El01 for the 1 lth International Histocompatibility Workshop. They
were synthesized in a DNA-SM automated synthesizer (Beckman, Palo Alto, CA, USA).
Dot-blot hybridization. Five percent of the amplified
DNA was denatured in 0.4 mol/l NaOH for 10 minutes,
neutralized in 1 mol/l ammonium acetate, and trans-
ferred to a Hybond-N membrane (Amersham, Bucks,
UK). The filters were prehybridized for 1 hour in a solution containing 50 mmol/l Tris-HCI (pH = 8.0), 3
mol/l tetramethylammonium chloride, 2 mmolll EDTA
(pH = 8.0), 5X Denhart’s solution (2% bovine serum
albumin, 2% polyvinyl pyrrolidone 40, 2% ficoll 4OO),
and 0.1 mg/ml salmon sperm DNA. Then the radiola- beled oligonucleotide was added and hybridized at 54°C
for 3 hours. The filters were washed twice in 2X SSPE
(30X SSPE: 4.5 mol/l NaCl, 0.3 mol/l NaH,P04, 30 mmol/l EDTA, pH = 7.7) and 0.1% SDS at room
temperature for 10 minutes, once in TMAC solution (50
mmol/l Tris-HCl, pH = 8.0, 3 mol/l tetramethylam- monium chloride, and 2 mmol/l EDTA, 0.1% SDS) at
room temperature for 10 minutes, and twice in TMAC
solution at 58°C for 10 minutes. The dots were visual-
ized after 8 hours of exposure to Kodak XAR5 film at - 80°C with intensifying screens.
Oligonucleotide probes. Information about the sequences
and specificities of the DRB 1, DRB3, DQA 1, and DQB 1 oligonucleotides are from the 1 lth International
Histocompatibility Workshop [ 101. Oligonucleotide
synthesis was done using the cyanoethyl phosphora-
midite technique in a Beckman DNA-SM automated
DNA synthesizer, following the manufacturer’s proto- col. The oligonucleotides were labeled and purified ac-
cording to the methods of Nepom et al. Ill].
Cloning of amplified DNA and automated dideoxy sequenc- ing. Amplification products were purified and inserted
into the Replicative Form of SmaI-digested M13mp18 virus DNA (Boehringer Mannheim, Germany). Trans- formation of competent Eschericbia coli TG 1 cells, screen-
ing for HLA-DRB inserts, and purification of the recom-
binants was done as previously described [9]. Single- stranded DNA templates were sequenced using Sanger’s dideoxy chain termination method on an Applied Bio- systems 370A automated DNA sequencer {12) as de-
scribed [91. DNA from individuals with newly found DR-DQ associations was cloned and sequenced (three clones at least) in order to confirm associations.
Statistic Analysis
The linkage disequilibria between two alleles and the
level of significance (p) for 2 X 2 comparisons were
computed using the formulas of Mattiuz et al. { 131. The linkage disequilibria for comparison of more than two
alleles were calculated using the 1 lth International His-
tocompatibility Workshop methodology E 141. Also, the relative linkage disequilibria (RLD) were calculated as described in Imanishi et al. f 141 and Degos (151. Stan-
dard errors are worked out as detailed in Pickbourne et
al. { 161. The HLA-A, -B, -C, -DRBl, -DRB3, -DQAl, and -DQB 1 allelic frequencies and linkage disequilibria
were calculated by using two Fortram IV computer pro- grams 117). In addition (see Table 6), most frequent
complete haplotypes were tentatively deduced from (a)
the 2, 3, and 4 HLA loci haplotype frequencies depicted in Tables 3, 4, and 5; (b) the previously described hap-
lotypes in other populations 118, 191; and (c) it was
helpful to define haplotypes, if they appear in 2 or more individuals and the alternative haplotype was well de-
fined. This logical combination is more accurate for the
class II loci DRB 1, DRB3, DQA 1, and DQB 1, but is
somewhat less dependable for HLA-A-B, DR or HLA-
A,B. For comparison of phenotype and haplotype fre- quencies the reference tables of the 11th International
Histocompatibility Workshop were used { 18).
Dendrograrns
Phylogenetic trees (dendrograms) were constructed with
the genotype frequencies by using the neighbor-joining (NJ) method E201 with the standard genetic distances
(SGDs) between populations 1211 using the software DISPAN containing the programs GNKDST and
TREEVIEW.
RESULTS AND DISCUSSION
Characteristic HLA Alleles That Identify the Algerian Population (see Tables 1 and 2)
The HLA system has been used to identify individuals in
order to solve paternity disputes and also in forensic problems [22, 231. The discriminative strength of the
HLA system stems from the fact that the number of alleles for each loci is very high. HLA strength for iden-
tifying individuals for paternity testing is higher than the additive strength of many others enzymes and blood group polymorphic systems. Presently, DNA-typed
HLA class II alleles and the discovery of new loci even increase the HLA strength for distinguishing individuals (241. This HLA discriminating ability may also be used to identify populations; particular alleles only appear or
262 A. Amaiz-Villena et al.
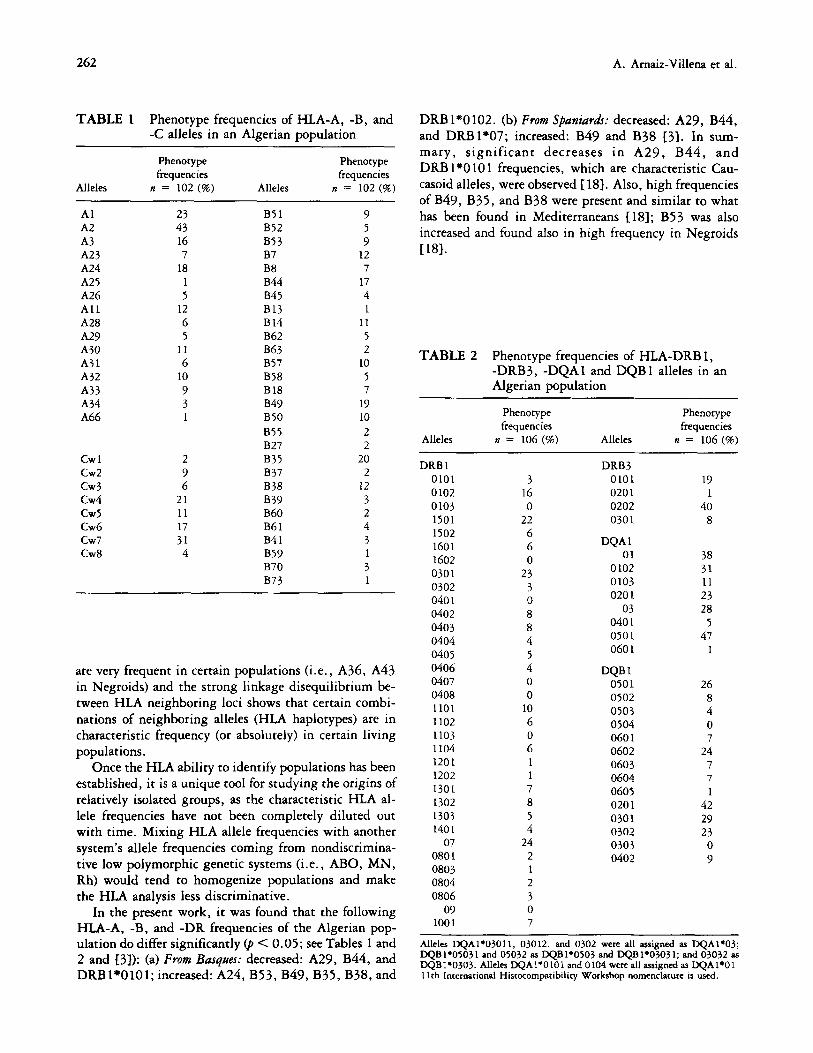
TABLE 1 Phenotype frequencies of HLA-A, -B, and DRB1*0102. (b) From Spaniudr: decreased: A29, B44, -C alleles in an Algerian population and DRB1’07; increased: B49 and B38 137. In sum-
Phenotype Phenotype mary, significant decreases in A29, B44, and
frequencies frequencies DRB lx0 10 1 frequencies, which are characteristic Cau-
Alleles n = 102 (%) Alleles n = 102 (%) casoid alleles, were observed 1187. Also, high frequencies
of B49, B35, and B38 were present and similar to what 23 9 43
has been found in Mediterraneans [18); B53 was also 5
16 9 increased and found also in high frequency in Negroids
12 1181.
Al A2 A3 A23 A24 A25 A26 All A28 A29 A30 A31 A32 A33 A34 A66
B51 B52 B53 B7 B8 B44 B45 B13 B14 B62 B63 B57 B58 B18 B49 B50
855 B27
Cwl 2 cw2 9 cw3 6 cw4 21 cw5 11 Cw6 17 cw7 31 Cw8 4
18 1 5
12 6 5
II 6
10 9 3 1
17 4
11 5 2
10 5
19 10 2 2
B35 20 B37 2 B38 12 B39 3 B60 2 B61 4 B41 3 B59 1 B70 3 B73 1
are very frequent in certain populations (i.e., A36, A43
in Negroids) and the strong linkage disequilibrium be- tween HLA neighboring loci shows that certain combi-
nations of neighboring alleles (HLA haplotypes) are in characteristic frequency (or absolutely) in certain living
populations. Once the HLA ability to identify populations has been
estabiished, it is a unique tool for studying the origins of
relatively isolated groups, as the characteristic HLA al- lele frequencies have not been completely diluted out with time. Mixing HLA allele frequencies with another
system’s allele frequencies coming from nondiscrimina-
tive low polymorphic genetic systems (i.e., ABO, MN, Rh) would tend to homogenize populations and make the HLA analysis less discriminative.
In the present work, it was found that the following HLA-A, -B, and -DR frequencies of the Algerian pop- ulation do differ significantly @ C 0.05; see Tables 1 and 2 and {37): (a) From Baqza: decreased: A29, B44, and DRBl*OlOl; increased: A24, B53, B49, B35, B38, and
TABLE 2 Phenotype frequencies of HLA-DRB 1, -DRB3, -DQAl and DQB 1 alleles in an Algerian population
Alleles
Phenotype frequencies
n = 106 (%) Alleles
Phenotype frequencies
n = 106 (%)
DRB 1 0101 0102 0103 1501 1502 1601 1602 0301 0302 0401 0402 0403 0404 0405 0406 0407 0408 1101 1102 1103 1104 1201 1202 1301 1302 1303 1401
07 0801 0803 0804 0806
09 1001
3 16 0
22 6 6 0
23 3 0 8 8 4 5 4 0
0
10 6 0 6
8 5 4
24 2
2 3 0
DRB3 0101 0201 0202 0301
DQA 1 01
0102 0103 0201
03 040 1 050 1 060 1
DQB 1 0501 0502 0503 0504 060 1 0602 0603 0604 0605 0201 0301 0302 0303 0402
19
40 8
38 31 11 23 28
47 1
26 8 4 0
24
7
42 29 23
0 9
Alleles DQA1*03011, 03012, and 0302 were all assigned as DQA1+03; DQB1.05031 and 05032 as DQB1+0503 and DQBP03031; and 03032 as DQB190303. Alleles DQAl+OlOl and 0104 were ail assigned as DQAl*Ol. 1 Ich International Histocompatibility Workshop nomemlacure is used.
HLA in Algerians 263
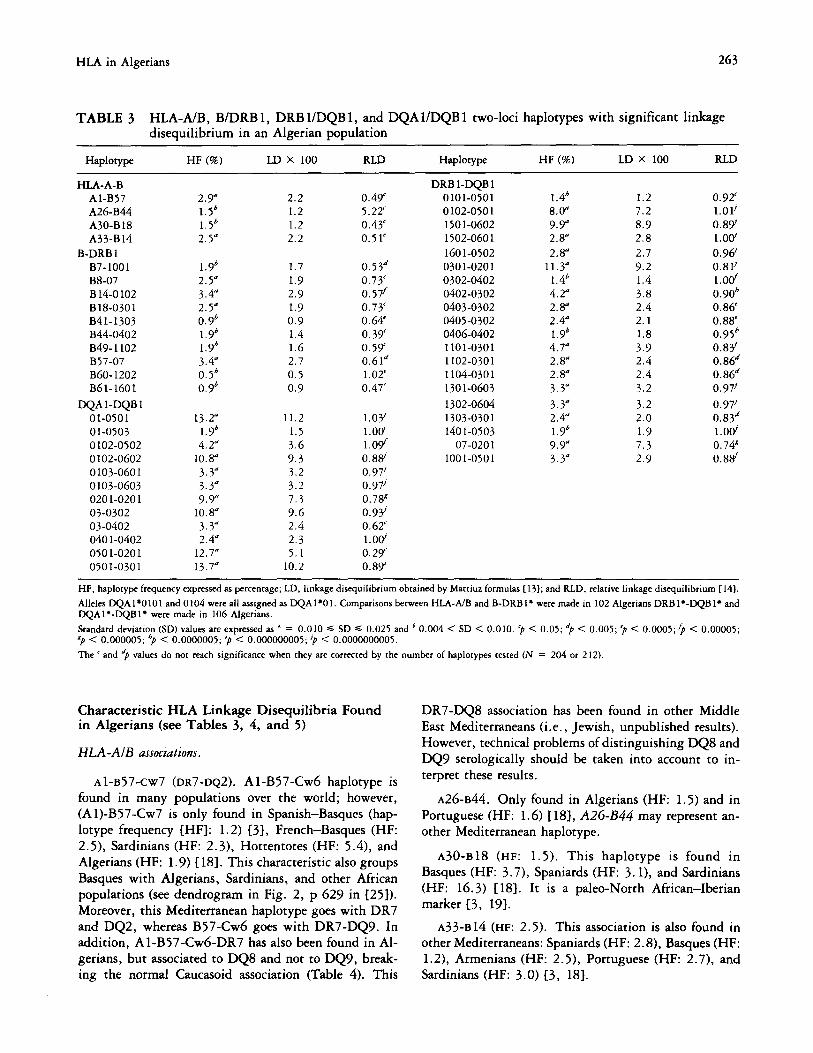
TABLE 3 HLA-A/B, B/DRB 1, DRB l/DQB 1, and DQAl/DQB 1 two-loci haplotypes with significant linkage disequilibrium in an Algerian population
HF (%) LD x 100 RLD Haplotype HF (%) LD X 100 RLD
HLA-A-B Al-B57 A26-B44 A30-B 18 A33-B 14
B-DRB 1
B7-1001 B8-07
B 14-O 102 B18-0301 B41-1303 B44-0402 B49- 1102
B57-07 B60- 1202 Bbl-1601
DQA 1-DQB 1
01-0501
01-0503 0102-0502 0 102-0602
0103-0601 0103-0603 0201-0201
03-0302 03-0402
040 l-0402 0501-0201
0501-0301
2.9” 1.5b 1.5b
2.5”
1.9b
2.5” 3.4”
2.5” o.9b 1.9b 1.9b 3.4”
o.5b o.9b
13.2”
1.9b 4.2”
10.8”
3.3” 3.3”
9.3” 10.8”
3.3” 2.4”
12.7“ 13.7”
2.2 0.49’ 1.2 5.22’ 1.2 0.43’
2.2 0.51’
1.7 0.53d
1.9 0.73’
2.9 0.57f
1.9 0.73‘
0.9 0.64 1.4 0.3Y 1.6 0.59’ 2.7 0.61d
0.5 1.02’
0.9 0.47’
11.2 l.OY
1.5 1.00‘
3.6 l.O$f
9.3 0.88
3.2 0.97’
3.2 0.97’
7.3 0.78”
9.6 0.9Y 2.4 0.62‘
2.3 l.OV
5.1 0.29’ 10.2 0.89’
DRB l-DQB 1
0101-0501 0 102-050 1 1501-0602 1502-0601
160 l-0502
030 l-020 1 0302-0402 0402-0302
0403-0302 0405-0302 0406-0402 1101-0301 1102-0301
1104-030 1
130 l-0603
1302-0604 1303-0301
1401-0503 07-020 1
1001-0501
l.4b 1.2 0.92’ 8.0” 7.2 1.01’
9.9” 8.9 0.89’ 2.8” 2.8 l.O(Y
2.8” 2.7 0.98 11.3” 9.2 0.81’
l.4b 1.4 l.Od 4.2” 3.8 0.90h 2.8” 2.4 0.86 2.4” 2.1 0.88’ 1.9b 1.8 0.95h 4.7” 3.9 0.8g 2.8” 2.4 0.86d 2.8” 2.4 0.86d
3.3” 3.2 0.971
3.3” 3.2 0.971 2.4” 2.0 0.83d 1.9b 1.9 l.O(Y
9.9” 7.3 0.74x
3.3” 2.9 0.8s’
HF, haplotype frequency expressed ar percentage; LD, linkage disequilibrium obtained by Mattiuz formulas (131; and RLD, relative linkage disequilibrium [IdI.
Alleles DQA 1+0 10 1 and 0 104 were all assigned as DQA l*O 1. Comparisons between HLA-A/B and B-DRB l* were made in 102 Algerians DRB 1 l -DQB 1’ and DQAl*-DQBI’ were made in 106 Algerians.
Standard deviation (SD) values ate expressed as ’ = 0.010 G SD G 0.025 and 6 0.004 < SD < 0.010. ‘p < 0.05; “p < 0.005; ‘p < 0.0005;‘p < 0.00005; "p < 0.000005; "p < 0.0000005; 'p < 0.000000005; ‘p < 0.0000000005.
The < and "p V&RS do not reach significance when they ate corrected by the number of haplotypes tested 0’ = 204 or 212).
Characteristic HLA Linkage Disequilibria Found in Algerians (see Tables 3, 4, and 5)
HLA-AIB associations.
Al-B57-Cw7 (DR7-DQ2). A 1-B57-Cw6 haplotype is found in many populations over the world; however, (Al)-B57-Cw7 is only found in Spanish-Basques (hap-
lotype frequency [HF]: 1.2) 131, French-Basques (HF: 2.5), Sardinians (HF: 2.3), Hottentotes (HF: 5.4), and
Algerians (HF: 1.9) 118). This characteristic also groups
Basques with Algerians, Sardinians, and other African populations (see dendrogram in Fig. 2, p 629 in 1251).
Moreover, this Mediterranean haplotype goes with DR7 and DQ2, whereas B57-Cw6 goes with DR7-DQ9. In addition, Al-B57-Cwb-DR7 has also been found in Al-
gerians, but associated to DQS and not to DQ9, break- ing the normal Caucasoid association (Table 4). This
DR7-DQ8 association has been found in other Middle East Mediterraneans (i.e., Jewish, unpublished results).
However, technical problems of distinguishing DQS and DQ9 serologically should be taken into account to in-
terpret these results.
~26-~44. Only found in Algerians (HF: 1.5) and in
Portuguese (HF: 1.6) 1181, A26-B44 may represent an-
other Mediterranean haplotype.
A30-B18 (HF: 1.5). This haplotype is found in
Basques (HF: 3.7), Spaniards (HF: 3. l), and Sardinians (HF: 16.3) IlS]. It is a paleo-North African-Iberian
marker 13, 191.
~33~14 (HF: 2.5). This association is also found in
other Mediterraneans: Spaniards (HF: 2.8), Basques (HF: 1.2), Armenians (HF: 2.5), Portuguese (HF: 2.7), and Sardinians (HF: 3.0) [3, 181.
264 A. Amaiz-Villena et al.
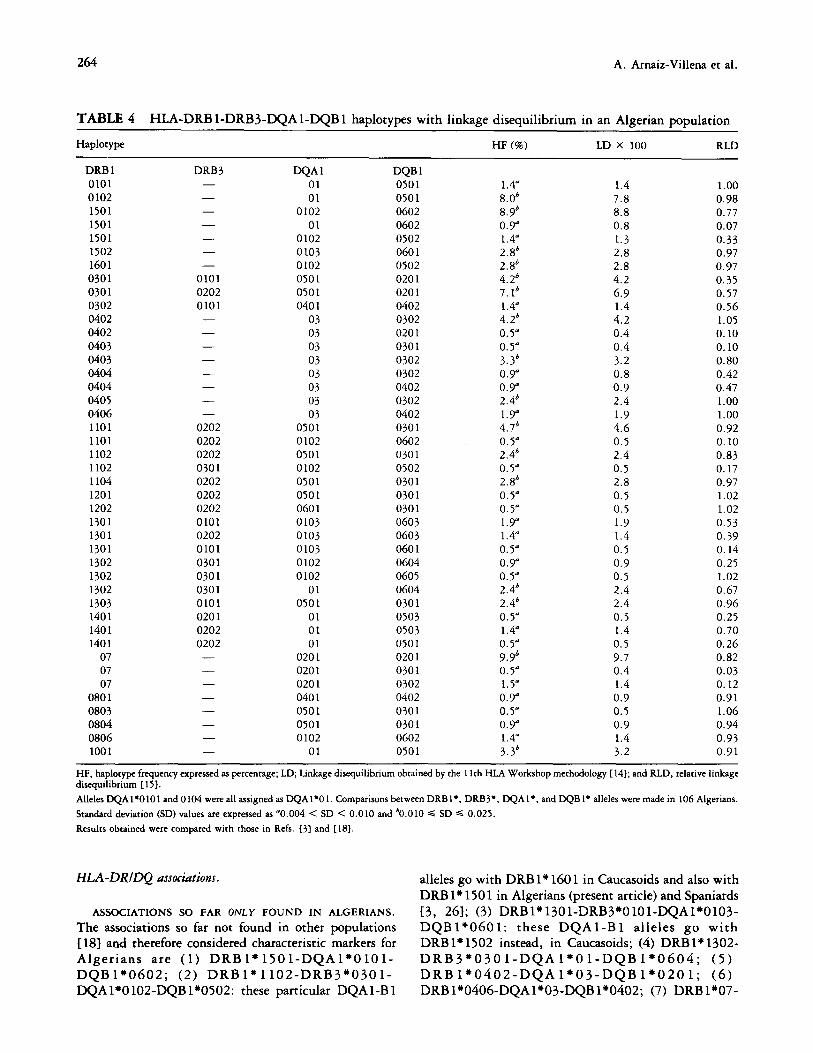
TABLE 4 HLA-DRB l-DRB3-DQA l-DQB 1 haplotypes with linkage disequilibrium in an Algerian population
Haplotype HF (%) LD X 100 RLD
DRB 1 0101 0102 1501 1501 1501 1502 1601 0301 0301 0302 0402 0402 0403 0403 0404 0404 0405 0406 1101 1101 1102 1102 1104 1201 1202 1301 1301 1301 1302 1302 1302 1303 1401 1401 1401
07 07 07
0801 0803 0804 0806 1001
DRB3 - - - - - - -
0101 0202 0101 - - - - - - - -
0202 0202 0202 0301 0202 0202 0202 0101 0202 0101 0301 0301 0301 0101 0201 0202 0202 - - - - - - - -
WA1 DQB 1 01 0501 01 0501
0102 0602 01 0602
0102 0502 0103 0601 0102 0502 0501 0201 0501 0201 040 1 0402
03 0302 03 0201 03 0301 03 0302 03 0302 03 0402 03 0302 03 0402
0501 0301 0102 0602 0501 0301 0102 0502 0501 0301 0501 0301 0601 0301 0103 0603 0103 0603 0103 0601 0102 0604 0102 0605
01 0604 0501 0301
01 0503 01 0503 01 0501
0201 0201 0201 0301 0201 0302 040 1 0402 0501 0301 0501 0301 0102 0602
01 0501
1.4” 1.4 1.00
8.0” 7.8 0.98 8.9’ 8.8 0.77 0.9” 0.8 0.07 1.4” 1.3 0.33 2.8’ 2.8 0.97 2.8’ 2.8 0.97 4.2’ 4.2 0.35 7.1b 6.9 0.57 1.4” 1.4 0.56 4.2’ 4.2 1.05 0.5” 0.4 0.10 0.5” 0.4 0.10 3.3b 3.2 0.80 0.3” 0.8 0.42 0.9” 0.9 0.47 2.4’ 2.4 1.00 1.9” 1.9 1.00 4.7b 4.6 0.92 0.5” 0.5 0.10 2.4’ 2.4 0.83 0.5” 0.5 0.17 2.8’ 2.8 0.97 0.5” 0.5 1.02 0.5” 0.5 1.02 1.9” 1.9 0.53 1.4” 1.4 0.39 0.5” 0.5 0.14 0.3” 0.9 0.25 0.5” 0.5 1.02 2.4b 2.4 0.67 2.4’ 2.4 0.96 0.5” 0.5 0.25 1.4” 1.4 0.70 0.5” 0.5 0.26 9.9b 9.7 0.82 0.5” 0.4 0.03 1.5” 1.4 0.12 0.9” 0.9 0.91 0.5” 0.5 1.06 0.Y 0.9 0.94 1.4” 1.4 0.93 3.3b 3.2 0.91
HF, haplotype frequency expressed as percentage; LD; Linkage disequilibrium obtained by the 1 lth HLA Workshop methodology {141; and RLD, relative linkage disequilibrium [ 151.
Alleles DQA 1’010 1 and 0 104 were all assigned as DQAI’O 1. Comparisons between DRB 1 *, DRB3*, DQAl*, and DQBl* alleles were made in 106 Algerians.
Standard deviation (SD) values are expressed as “0.004 < SD < 0.010 and ‘0.010 s SD s 0.025.
Results obtained were compared with chose in Refs. 131 and [ 181.
HLA-DRIDQ associations. alleles go with DRB 1” 160 1 in Caucasoids and also with
DRB lx 150 1 in Algerians (present article) and Spaniards
ASSOCIATIONS SO FAR ONLY FOUND IN ALGERIANS. [3, 261; (3) DRB1”1301-DRB3”0101-DQA1’0103- The associations so far not found in other populations DQBl”0601: these DQAl-Bl alleles go with { 181 and therefore considered characteristic markers for DRB 1” 1502 instead, in Caucasoids; (4) DRB l+ 1302- Algerians are (1) DRB 1’ 150 l-DQA l*O 10 l- DRB3’0301-DQAl”Ol-DQB1’0604; (5) DQB 1*0602; (2) DRB 1” 1102-DRB3*030 l- DRB1*0402-DQAl*03-DQB1*0201; (6) DQA 1’0 102-DQB l”O502: these particular DQAl-B 1 DRB 1”0406-DQA1’03-DQB 1+0402; (7) DRB 1*07-

HLA in Algerians 265
TABLE 5 HLA-B, DR, DQ three-loci with linkage disequilibrium Algerian population
haplotypes in an
is included in the A30-B18-DR3 haplotype, a classical
Iberian and paleo-North African haplotype 119,291; (5)
DRB 1*0405-DQA 1*03-DQB 1*0302 is also described
in Spaniards 13 11 and considered a rarity in European and American Caucasoids {32-341, DRB l”O405 allele has
been found in other Mediterraneans such as Sardinians
IlSl, Ethiopians 1351, and Greeks [361 but associated with other DQ alleles; (6) DRB lx 140 l-DRB3*020 l-
DQAl*O l-DQB lx0503 is also described in Sardinians
E291; and (7) DRB 1*0402-DQA1*03-DQB 1*0302 is also found in high frequency in Israeli Jews 1371.
Haplotype HF (%) LD X 100
B-DRB I-DQA l-DQB 1 B 14-O 102-O l-050 1 3.4
B7-1501-0102-0602 1.9
B44-1502-0103-0601 0.9
B35-0301(DRB3’0101)-0501-0201 2.5
B IS-0301(DRB3+0202)-0501-020 1 2.5
B44-0402-03-0302 1.9
B49-1102(DRB3f0202)-050 L-0301 1.9
B35-1104(DRB3*0202)-0501-0301 1.5
B57-07-0201-0201 2.5
B57-07-020 l-0302 1.5
B38-0801-0401-0402 0.9
B7-1001-01-0501 1.9
3.2 1.6
0.9 1.9 2.2 1.8 1.8
1.5 2.2 1.2
0.9 1.8
HF. haplotype frequency expressed as percentage; LD, linkage disequilibrium obtained by the 1 lth HLA Workshop methodology [141.
Alleles DQAl*OlOl and 0104 were all assigned as DQAl*Ol.
Comparisons between HLA-B, DRBl+, DRB3*, DQAl*, and DQBl’ Al- leles were made in 102 Algerians.
Results obtained were compared with those in Refs. [31 and [181.
DQAl”020 l-DQB lx030 1; and (8) DRB 1*07- DQA l”O20 l-DQB 1’0302. These new-found associa-
tions were confirmed by automatic DNA sequencing (see
Materials and Methods).
ASSOCIATIONS FOUND IN ALGERIANS AND ALSO IN
OTHER POPULATIONS. Meditewaneans: (1) DRB 1*1502- DQA1*0102-DQB 1*0502 is also found in Spaniards
{26]; (2) DRB 1*0806-DQAl*O 102-DQB 1*0602 is the
most common DR8 subtype in Algerians [27] and is
found in Spaniards [3] and African Americans 1281; (3) DRB l+O 102-DQA lx0 l-DQB lx050 1 is present in high
frequency in Spaniards [3], Sardinians [29], Israeli Jews lb], and in North American Negroids (301; (4)
DRB 1*030 l-DRB3*0202-DQA lx050 1-DQB 1+020 1
FIGURE 2 Dendrogram showing relatedness between Al- gerians and other populations. Standard genetic distances were calculated by using HLA-A and -B (serologic) and -DRB 1, DQAl, and DQB 1 (DNA sequence) frequencies.
_ ALGERIANS
SVStlMEN
African Negroids: (8) DRB 1” 110 l-DRB3*0202- DQA1*0102-DQBl”0602 association is found in Ne-
groids 130, 381 and (9) DRB1#0404-DQA1”03- DQBl”0402 is also present in Bushmen and Hottentots
{18}. The latter DR/DQ association goes in favor of the
postulated Negroid component of Algerians 141.
Tentative Assignment of HLA Extended Haplotypes to the Different Original Ethnic Groups (Table 6)
Extended haplotypes in Algerians had been poorly stud- ied previously; this was in part due to a lack of HLA
DNA sequencing technology E6, 39, 401. However, pre- vious data have been confirmed and new haplotypes have
been described (see Table 6). An exhaustive study has
been made in order to compare Algerian population with
Spaniards, Sardinians, Basques, and others E6, 18, 29,
38, 411. The data presently available will test the hy- pothesis of a common Iberian paleo-North African origin
of Spaniards, Basques, Sardinians, and Algerians 119,
27, 3 11. HLA data are accompanied with recorded his-
toric origins and migrations of North African, Iberian, and European populations.
The most representative haplotypes found in Algeri-
ans and their HFs are shown in Table 6. In addition, the following haplotypes have only been
found in Algerians: A2-Cblank-B38-DRS-
(DRB I”080 l)-DQ4(DQA lx040 l-DQB 1*0402) and
A2-Cw7-B7-DR lO(DRB lx 100 l)-DQ l(DQA 1”O l-
DQBl*0501) (HF: 0.9 and 0.9, respectively) { 18).
HLA Relatedness of Algerians with Spaniards and Basques: Paleo North Africans, Iberians, and Basques (Fig. 2)
In the last 1 lth HLA Workshop a dendrogram was con- structed by using only HLA-A and -B frequencies and
the NJ method was done to calculate the genetic rela- tionships among worldwide populations [25]. Spaniards, Basques, and Sardinians clustered together with African populations and were separated from the main European
branch of ethnic groups. Maghreb and Middle East pop- ulations could not be included in the analysis, because of the lack of data.
In the present work, not only HLA-A and -B frequen-

266 A. Amaiz-Villena et al.
TABLE 6 The nine most frequent estimated extended haplotypes in Algerians and tentative assignment to the original contributing ethnic groups
HF Possible HLA-A,B,C HLA-DRB I-DRB3/4 HLA-DQAl-DQB 1 (%) origin
Al Cw7 B57 DR7(DRB1*07-DRB4*0101) DQ2(DQAl*O201-DQB1’0201)” 1.9 Iberian-paleo-North African (Basque-Algerian)
A30 Cw5 B18 DR3(DRB1*0301-DRB3+0202) DQ2(DQA1*0501-DQB1*0201)b 1.5 Iberian-paleo-North African
A33 Cw8 B14 DRl(DRBl*0102) DQ5(DQA1*Ol-DQBlf0501)r 1.5 Mediterranean
A2 cw4 B35 DRl1(DRBl’l104-DRB3’0202) DQ7(DQAIf0501-DQB1*0301)d 1.5 Mediterranean
A2 Cw4 B35 DR3(DRB1*0301-DRB3’0101) DC2(DQA1*0501-DQB1’0202)’ 1.5 Mediterranean (also in Sardinians)
A3 Cw7 B49 DRl l(DRB l* 1102.DRB3*0202) DQ7(DQA1*0501-DQB1*030ly 1.5 Autochthonous
A3 Cwl B7 DRlS(DRB1*1501) DQ6(DQAl*0102-DQBl*OGOZ)p 1.5 ?
A26 Cbl 844 DRlS(DRB1*1502) DQ6@QAlr0103-DQB1*0601)’ 0.9 Autochthonous
A2 Cw5 B44 DR4(DRB 1+0402-DRB4+0 10 1) DQ8(DQA1*03DQB1*0302) 0.9 Autochthonous
A2 Cw7 B7 DRlS(DRB1*1501) DQ6(DQA1*0102-DQBl*ObOZ)’ 0.9 ?
HF, haplotype frequency expressed as percentage. Alleles DQAl*OlOl and 0104 were all assigned as DQAl+Ol.
a Only common to French-Basques (HF: 2.7) and Spanish-Basques (HF: 1.2) I3.181. It is probably a haplotype of a paleo-North African-Iberian origin; it is different from the worldwide distributed Al-B57-DR7 haplotype which goes with Cw6 and DQ9. In a study done in 1981 in the AlgerianKabyle population, this haplotype was also found to be the second most frequent [39]. It was described as A I-Bw 17.
b Also found in Spaniards (HF: 3. I), Sardinians (HF: 11.4), and French-Basques (HF: 4.7) (18). Its frequency is much lower or absent in other Caucasoid populations. This haplotype was defined as Iberian-paleo-North African [ 19]. Presently, reported data on Algerians support this hypothesis. Bouali et al. 1391 found this haplotype to be the first most frequent in Algerians; it was described as B18-BfFl.
’ Shows the highest frequency in Armenians (HF: 3.1) and is also high in frequency or present in other Medirerranean Caucasoid populations and as Sardinians (HF: 2.7). Spaniards (HF: 2.6). French (HF: 1.4). Greeks (HF: 1. l), and Italians (HF: 0.7) [ 181. Bouali et al. [39] found it in Algerians. This haplotype does not appear in Basques and has a much lower frequency in northern European Caucasoid populations [3]. The fact that Armenians (and not Basques) bear this marker does not support a theory that Basques and Armenians are related and with common descent [3]. Armenians may have acquired this haplotype during their historical Middle East Mediterranean setrlements in the middle ages [451. Alternatively, this haplotype may be of Armenian origin.
’ Also found in Italians (HF: 2.3) and Greeks (HF: 2.0) [18]. B35-DRI l-DQ7 is also present in non-lberian Mediterranean areas such as Albania (HF: 3.3), Armenia (HF: 5.0). and France (HF: 1.1) and in other populations such as Germans (HF: 3.4), Indians (HF: 3.1). and Hungarians (HF: 3.9) [ 181.
’ Common to Sardinians (HF: 2.7) and French (HF: 1.0) (181. HLA-B35-DR3-DQ2 is also present in Greeks (HF: 1.9) and Uralic populations (HF: 3.1) [18]. It could be a non-Iberian Mediterranean haplotype.
’ Only present in Algerians.
8 Appears in Spaniards (HF: 1.4) [3] and has been defined as a northern European haplotype [29] due to its high frequency in Danes (HF: 3.6). Austrians (HF: 3.2), Czecks (HF: 2.5). Germans (HF: 2.5), Yugoslavians (HF: 2.4). and also in French (HF: 1.8) and Italians (HF: 0.6) [18].
b Common to French-Basques (HF: 3.6). Cornish (HF: 3.6). Austrians (HF: 2.6). Britons (HF: 2.3), and Spaniards (HF: 1.9) I3,18] and does not appear in high frequency in northern Cxmssoids, such as Scandinavian, or in other Mediterraneans. This haplotype is extended throughout England, Spain, and northern Africa.
ties, but also DR ones, have been used to test relatedness between Algerians and other Europeans, particularly
northern Mediterraneans such as Spaniards, Basques, and
Sardinians. The same 1 lth Workshop methodology was
used (see Materials and Methods). Our results fully con-
firm the 11th Workshop data; Basques and Spaniards
cluster together with an African population, Algerians.
This also supports the theory that Iberians and paleo- North African Hamites (mainly represented by Berbers at present) are genetically related populations with com-
mon ancestors and that Basques may be a relative isolate of these primitive Iberian populations 13, 191. The ge-
netic system used (HLA-A and -B serology and DRB 1,
DQAl, DQB 1 exon-2 DNA sequences) is one of the most powerful and polymorphic genetic systems which may be utilized to discriminate populations (see above). In addition, results obtained analyzing haplotype fre- quencies, particularly of the characteristic haplotype A30-B18 and Al-B57-Cw7, which are probably markers
of neolithic Iberian people (Table 6) {31, further support a relatedness between paleo-North Africans and present- day Spaniards and Basques (Iberians). Other genetic data that apparently identified Basques from surrounding populations were mostly based on blood groups. Rh-
negative phenotype high frequency and B group low
frequency, once regarded as distinctive of Basque popu- lations [42], has been found to be a general characteristic
of other neighboring western European populations. Rh-
negative gene frequencies (cde]: Spanish-Basques (0.44),
Portuguese (0.54), Isle of Man (0.43), Spaniards (0.38),
French (0.41), Irish (0.42), and English (0.39). B-gene frequencies: Basques (0.03), Portuguese (0.05), Irish
(0.07), Spaniards (O.O6), French (O.O6), and English (0.06) 151. Thus frequencies of Basque Rh-negative and B phenotype are not significantly different from sur-
rounding populations, although a distinctive effect
might be expected due to the historic Basque relative
lack of substantial admixture with neighbors and invad-
ers 143, 441. In summary, in the present work complete HLA data
on Basques and Spaniards was compared for the first time with similar HLA data on a North African population (Algerian) with a high paleo-North African (Berber)
component [4, 271. Figure 2 (dendrogram) shows that Basques and Spaniards are closer to Algerians than to any other Caucasoid, Negroid, or Mongoloid population. In addition, the specific highly discriminative HLA haplo- type, HLA-A30-B 1%DR3, is a marker common to Sar-

HLA in Algerians
dinians, Basques, Spaniards, and Algerians. Taken to- gether, these data are the strongest reported genetic sup- port for a common descent of Basques, Spaniards, and Algerians, probably reflecting the preneolithic relation- ship between Iberians and paleo-North Africans.
ACKNOWLEDGMENTS
This work was supported by Fondo de Investigaciones Sani-
tar&, Ministerio de Sanidad, and Fundacion Ramon Areces, Spain. We are grateful to Manuel Martin-Villa, Foutia
Bekhoucha, and Fatima Belhani for their collaboration. We
thank to Dr. Nei, Dr. Kumar, and Dr. Ota for the computer programs kindly provided.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
McEvedy C, Jones R: Atlas of World Population History.
Harmondsworth, Penguin Books, 1978.
Bodmer WF, Cavalli-Sforza LL: Genetics, Evolution and
Man. San Francisco, WH Freeman 1976, p 573.
Martinez-Las0 J, De Juan D, Martinet-Quiles N, Gomez- Casado E, Cuadrado E, Arnaiz-Villena A: The contribu-
tion of the HLA-A, -B, -C and -DR, -DQ DNA typing to the study of Spaniards and Basque origins. Tissue An-
tigens 45:237.
Julien CA: Historie de I’Afrique du Nord. Paris,
PAYOT, 196 1.
Roychouldhury AK, Nei M: Human Polymorphic Genes World Distribution. New York, Oxford University
Press, 1988.
Brautbar C, Friedman A, Battat S, Tambur AR, Cam-
bon-Thomsen A, Ohayon E, Tongio M, DuPont E, Es- sakali-Gharbi M: HLA in Israeli Jew, Moroccans and Al-
gerians. In Tsuji K, Aizawa M, Sasatuki T (eds). HLA
1991, vol 1 Oxford, Oxford University press, 1992, p
656.
Danilovs J, Terasaki PI, Park MS, Ayoub G: B lympho- cyte isolation by thrombin-nylon wool. In Terasaki PI
(ed): Histocompatibility Testing 1980. Los Angeles, UCLA Tissue Typing Laboratory, 1980, p 287.
Davis RW, Thomas M, Cameron J, St John TP, Padgett RA: Rapid isolation for enzymatic and hybridization anal- ysis. Methods Enzymol 65:404, 1980.
Core11 A, Martin-Villa JM, Varela P, Vicario JL, Mar- tinez-Laso J, Arnaiz-Villena A: Exon 2 DNA sequence of the DRw 13b allele obtained from genomes of five differ- ent individuals. Mol Immunol 27:313, 1990.
Kimura A, Sasazuki T: Eleventh International Histocom- patibility Workshop Reference Protocol for the HLA-
DNA-Typing Technique. In Tsuji K, Aizawa M, Sasa- zuki T (eds): HLA 199 1, vol 1. Oxford, Oxford Univer- sity Press, 1992, p 397.
Nepom GT, Byers P, Seyfried C, Healey LA, Wilske KR, Stage D, Nepom BS: HLA genes associated to rheu-
12. Gocayne J, Robinson DA, Fitzgerald MG, Chung FZ, Kerlavage AR, Lentes KU, Lai J, Wang CD, Fraser CM, Venter FG: Primary structure at rat cardiac P-adrenergic and muscarinic cholinergic receptors obtained by auto- mated DNA sequence analysis: further evidence for a
multigene family. Proc Nat1 Acad Sci USA 84:8296,
1987.
13.
14.
Mattiuz PL, Ihde D, Piazza A, Ceppelini R, Bodmer WF:
New approaches to the population genetics and segrega- tion analysis of the HLA system. In Histocompatibility
Testing 1970. Copenhagen, Munskaard, 1970, p 193.
Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojorobi T: Estimation of allele and haplotype frequencies for HLA and complement loci. In Tsuji K, Aizawa M, Sasazuki T
(eds): HLA 199 1, vol 1. Oxford, Oxford University Press,
1992, p 76.
15.
16.
Degos L: La repartition anthropologique des genes HLA et dynamique des populations. In Dausset (ed): HLA
1982. Complexe majeur d’histocompatibilite de I’homme. Flammarian Medicine Sciences, 1982, p 13 1.
Pickbourne P, Piazza A, Bodmer WF: Population analy-
sis. In Bodmer WF, Batchelor JR, Bodmer JG, Festesn- tein H, Morris PJ (eds): Histocompatibility Testing
1977. Copenhagen, Munksgaard, 1978, p 259.
17. Vicario JL, Martinez-Laso J, Core11 A, Regueiro JR, Ar- naiz-Villena A: Both HLA class II and class III restriction fragment length polymorphism factors linked to rheuma- toid juvenile arthritis. Clin Immunol Immunopathol 56:
22, 1990.
18. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojorobi
T: Allele and haplotype frequencies for HLA and comple- ment loci in various ethnic groups. In Tsuji K, Aizawa M, Sasazuki T (eds): HLA 1991, vol 1. Oxford, Oxford
University Press, 1992, p 1065.
19. Arnaiz-Villena A, Rodriguez de Cordoba S, Vela F, Pas- cual JC, Cervero J, Bootello A: HLA antigens in a sample
of the Spanish population: common features among Span-
iards, Basques and Sardinians. Hum Genet 58:344, 1981.
20. Saitou N, Nei M: The neighbor-joining method: a new method for reconstructing phylogenetic tree. Mol Biol Evol 4:406, 1987.
21. Nei M: Genetic distances between populations. Am Nat 106:283, 1972.
22.
23.
Rose NR, Macario EC, Fahey JL, Friedman H, Pen GM (eds): Manual of Clinical Laboratory Immunology. Wash-
ington, American Society for Microbiology, 1992, p 901.
Snell GD, Dausset J, Nathenson S (eds): Histocompati- bility: HLA Polymorphism. New York, Academy Press, 1976, p 210.
24. Tsuji K, Aizawa M, Sasazuki T (eds): HLA 1991: No-
267
matoid arthritis: identification of susceptibility alleles us-
ing specific oligonucleotide probes. Arthritis Rheum 32:
15, 1989.

268 A. Arnaiz-Villena et al.
25.
26.
27.
28.
29.
30.
31.
32.
33.
menclature for Factors of the HLA System, 1991, vol 1. oxford, Oxford University Press, 1992, p 17.
Imanishi T, Wakisaka A, Gojorobi T: Genetic relation-
ships among various human populations indicated by MHC polymorphisms. In Tsuji K, Aizawa M, Sasazuki T
(eds): HLA 199 1, vol 1. Oxford, Oxford University Press,
1992, p 627.
Vilches C, De Pablo R, Moreno M, Solis R, Kreisler M: Characterization of an HLA-DR15 DQS haplotype found
in the Spanish Caucasoid population. Hum Immunol 35:
223, 1992.
Benmamar D, Martinez-Las0 J, Varela P, Bekhoucha F, Morales P, Arnaiz-Villena A: Evolutionary relationships
of HLA DR8 alleles and description of a new subtype (DRB1*0806) in the Algerian population. Hum Immu-
no1 36:172, 1993.
Apple RJ, Erlich HA: Two new HLA-DRB 1 alleles found
in African Americans: implications for balancing selection
at positions 57 and 86. Tissue Antigens 40:69, 1992.
Piazza A, Fauchet R, Richiardi P, Carcassi C, Contu L:
Anthropology report: Belgian, French, Italian, Portu-
guese, Sardinian and Spanish population. In Tsuji K,
Aizawa M, Sasazuki T (eds): HLA 1991, vol 1. Oxford,
Oxford University Press, 1992, p 648.
Fernandez-Viiia MA, Gao X, Moraes ME, Moraes JR,
Salatiel I, Miller S, Tsai J, Sun Y, An J, Layrisse Z, Gazit E, Brautbar C, Stastny P: Alleles at four HLA class II loci determined by oligonucleotide hybridization and their as-
sociations in five ethnic groups. Immunogenetics 34:299,
1991.
Morales P, Martinez-Las0 J, Martin-Villa JM, Core11 A, Vicario JL, Varela P, Arnaiz-Villena A: High frequency of
the DRB 1*0405-(Dw 15)-DQw8 haplotype in Spaniards
and its relationship to diabetes susceptibility. Hum Im-
munol 32: 170, 199 1.
Lanchbury JSS, Hall MA, Welsh KI, Panayi GS: Se-
quence analysis of HLA-DR4B 1 subtypes: additional first domain variability is detected by oligonucleotide hybrid-
ization and nucleotide sequencing. Hum Immunol 27:
136, 1990.
Gao XJ, Fernandez-Vina M, Shumway W, Stastny P: DNA typing for class II HLA antigens with allele specific
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
or group specific amplification: typing for subsets of DR4. Hum Immunol 27:40, 1990.
Celis E, Karr RW, Gregersen PK, Reismoen NL: Func- tional analysis of MHC class II-restricted T cells derived
from a Caucasian with a DR4, Dw15, DQw8 haplotype. Hum Immunol 29:31, 1990.
Stastny P, Kimura A: DNA typing analysis of the DR4 group. In Tsuji K, Aizawa M, Sasazuki T (eds): HLA
1991, vol 1. Oxford, Oxford University Press, 1992, p 465.
Cutbush SD, Sachas JA, Papasteriades C, Boki K, Biro PA: HLA associations in ethnic populations: HLA-Dw 15
is associated with HLA-DR4 in Greek population. Pro- ceedings of the British Society for Immunology, Spring
Meeting, London, April 10-12, 1991, p 41.
Cohen N, Mickelson E, Amar A, Battat S, Hansen JA, Brautbar C: HLA-Dw clusters associated with DR4 in Israeli Jews and the definition of a new DR4 associated
Dw subtype: Dw ‘SHA.’ Hum Immunol 19:179, 1987.
Stastny P, Kalil J: HLA in North American and South
American Negroids. In Tsuji K, Aizawa M, Sasatuki T
(eds): HLA 199 1, vol 1. Oxford, Oxford University Press,
1992, p 638.
Bouali M, Dehay C, Benajam A, Poirier JC, Degos L, Marcelli-Barge A: HLA-A, B, C, Bf and glyoxalase I polymorphisms in a sample of the Kabyle population (Al-
geria). Tissue Antigens 17:501, 1981.
Mercier P, Vallo JJ, Vialettes B, Vague Ph: HLA-A, B, DR antigens and insulin-dependent diabetes in Algeri-
ans. Tissue Antigens 26:20, 1985.
Lindblom B, Svejaard A: HLA genes and haplotypes in the Scandinavian populations. In Tsuji K, Aizawa M,
Sasazuki T (eds): HLA 199 1, vol 1. Oxford, Oxford Uni-
versity Press, 1992, p 65 1.
Mourant AE: The blood groups of the Basques. Nature
160:505, 1947.
Caro Baroja J: Los Pueblos de Espaiia, vol 2. Madrid,
Ediciones Itsmo, 1981, p 33.
Roman de1 Cerro JL: El Origen Iberico de la Lengua Vasca. Alicante, Editorial Aguaclara, 1993, p 33.
Duby G: Atlas Historique. Librairie Larousse, 1987.
Related Documents











