41 Number 71, December 2010 Abstract Historicising the emergence of ethnographic activities provides insights into the reliability of ethnographic analogies to aid archaeological understandings of past human societies, as well as allowing us to explore the historical emergence of ethnographically contextualised cultural traits. Epe Amoho is the largest hunting camp rockshelter used by the Himaiyu clan (Rumu people) of the Kikori River region, southern Papua New Guinea. Contemporary ethnographic information indicates dry season site use with subsistence practices directed towards riverine fishing and shellfishing, mammal hunting and gardening in the surrounding rainforest. But how long has the site been used and when in the past did activities start to resemble those known ethnographically? Archaeological excavations revealed three pulses of activity: Recent Phase (0-500 cal BP), Middle Phase (900-1200 cal BP) and Early Phase (2500-2850 cal BP). Pollen data reveal increasing rainforest disturbance by people through time. While the best match between ethnographic and archaeological practices occurs during the Recent Phase, selected aspects of Rumu subsistence extend back to the Early Phase. As the temporal depth of ethnographically-known practices differs between archaeological sites, a complex picture emerges where Rumu cultural practices unfolded at differing points in time and space over a period of at least 3000 years. Introduction A cornerstone of Indigenous archaeology in Australia and New Guinea is the availability of ethnographic information (Lilley 2006; Lourandos 1997; Mulvaney 1969; White and O’Connell 1982). The first scientific and controlled excavation of a rockshelter site in Australia associated upper stratigraphic layers with local ethnographic practices (Hale and Tindale 1930). Yet amongst archaeologists debate continues over the usefulness of ethnographic analogy and the relevance of historically-recorded information from the past two centuries to aid understanding of archaeological materials that in some cases are one and two orders of magnitude older in age (Hiscock 2008; Meehan and Jones 1988; Spriggs 2008). We advocate an approach that attempts to demonstrate the relevance of ethnographic information in archaeological interpretation by directly investigating the history and antiquity of ethnographically- known practices. That is, through archaeological investigation we historicise ethnographic sites to reveal when in the past they took their ethnographically-known form. Our approach takes activities as expressed materially in the surface levels of ethnographically-documented sites and tracks these back into the past and down through deposits (see David et al. 1994; McNiven et al. 2009). In this paper, we apply our approach to a rainforest rockshelter hunting camp named Epe Amoho that continues to be used by Himaiyu clan members of the Rumu people of the Kikori River of southern Papua New Guinea. Our research contributes to a small but growing ethnoarchaeological literature on human occupation of caves and rockshelters (e.g. Galanidou 2000; Gorecki 1991; Nicholson and Cane 1991; Pannell and O’Connor 2005; Pétrequin and Pétrequin 2006) and discusses the complexities of using ethnographic information in archaeological interpretation. Kikori Region – Archaeology and Ethnographic Analogy In the late 1970s, Jim Rhoads pioneered ethnoarchaeological and archaeological research in the western Gulf of Papua, centring on the middle reaches of the Kikori River (Rhoads 1980) (Figure 1). As ethnographic information for the Kikori region was rudimentary, Rhoads undertook his own ethnoarchaeological HISTORICISING THE PRESENT: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea Ian J. McNiven 1 , Bruno David 1 , Ken Aplin 2 , Max Pivoru 3 , William Pivoru 3 , Alex Sexton 4 , Jonathan Brown 1 , Chris Clarkson 5 , Kate Connell 5 , John Stanisic 6 , Marshall Weisler 5 , Simon Haberle 7 , Andrew Fairbairn 5 and Noel Kemp 8 1 School of Geography and Environmental Science, Monash University, Clayton, VIC 3800, Australia [email protected], bruno. [email protected], [email protected] 2 Australian National Wildlife Collection, CSIRO Division of Ecosystem Sciences, PO Box 284, Canberra, ACT 2601, Australia [email protected] 3 Kopi Village, Kikori District, Gulf Province, Papua New Guinea 4 PO Box 624, Mt Isa, QLD 4825, Australia [email protected] 5 School of Social Science, The University of Queensland, Brisbane, QLD 4072, Australia [email protected], [email protected], [email protected], [email protected] 6 Queensland Museum, South Brisbane, QLD 4101, Australia [email protected] 7 Research School of Pacific and Asian Studies, The Australian National University, Canberra, ACT 0200, Australia [email protected] 8 50 Derwent Avenue, Lindisfarne, TAS 7015, Australia [email protected] Aird Hills Ivi River Sab o r o R i v e r P in ini C re e k Emo Wokoi Amoho Fly R. D ig u l R . WEST PAPUA PAPUA NEW GUINEA Torres Strait Kilometres N Study area 200 0 Veir u C r e e k K i k o r i R i v e r Mati Ck U t i t i C k Kikiniu Puriau Kikori Ibira Herekuna Ouloubomoto Waira Kopi/Mampaiu Rupo Epe Amoho kilometres 10 0 Figure 1 Study area.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

41Number 71, December 2010
AbstractHistoricising the emergence of ethnographic activities provides insights into the reliability of ethnographic analogies to aid archaeological understandings of past human societies, as well as allowing us to explore the historical emergence of ethnographically contextualised cultural traits. Epe Amoho is the largest hunting camp rockshelter used by the Himaiyu clan (Rumu people) of the Kikori River region, southern Papua New Guinea. Contemporary ethnographic information indicates dry season site use with subsistence practices directed towards riverine fishing and shellfishing, mammal hunting and gardening in the surrounding rainforest. But how long has the site been used and when in the past did activities start to resemble those known ethnographically? Archaeological excavations revealed three pulses of activity: Recent Phase (0-500 cal BP), Middle Phase (900-1200 cal BP) and Early Phase (2500-2850 cal BP). Pollen data reveal increasing rainforest disturbance by people through time. While the best match between ethnographic and archaeological practices occurs during the Recent Phase, selected aspects of Rumu subsistence extend back to the Early Phase. As the temporal depth of ethnographically-known practices differs between archaeological sites, a complex picture emerges where Rumu cultural practices unfolded at differing points in time and space over a period of at least 3000 years.
IntroductionA cornerstone of Indigenous archaeology in Australia and New
Guinea is the availability of ethnographic information (Lilley
2006; Lourandos 1997; Mulvaney 1969; White and O’Connell
1982). The first scientific and controlled excavation of a
rockshelter site in Australia associated upper stratigraphic layers
with local ethnographic practices (Hale and Tindale 1930). Yet
amongst archaeologists debate continues over the usefulness of
ethnographic analogy and the relevance of historically-recorded
information from the past two centuries to aid understanding
of archaeological materials that in some cases are one and
two orders of magnitude older in age (Hiscock 2008; Meehan
and Jones 1988; Spriggs 2008). We advocate an approach
that attempts to demonstrate the relevance of ethnographic
information in archaeological interpretation by directly
investigating the history and antiquity of ethnographically-
known practices. That is, through archaeological investigation
we historicise ethnographic sites to reveal when in the past
they took their ethnographically-known form. Our approach
takes activities as expressed materially in the surface levels of
ethnographically-documented sites and tracks these back into
the past and down through deposits (see David et al. 1994;
McNiven et al. 2009). In this paper, we apply our approach to
a rainforest rockshelter hunting camp named Epe Amoho that
continues to be used by Himaiyu clan members of the Rumu
people of the Kikori River of southern Papua New Guinea. Our
research contributes to a small but growing ethnoarchaeological
literature on human occupation of caves and rockshelters (e.g.
Galanidou 2000; Gorecki 1991; Nicholson and Cane 1991;
Pannell and O’Connor 2005; Pétrequin and Pétrequin 2006) and
discusses the complexities of using ethnographic information in
archaeological interpretation.
Kikori Region – Archaeology and Ethnographic AnalogyIn the late 1970s, Jim Rhoads pioneered ethnoarchaeological and
archaeological research in the western Gulf of Papua, centring
on the middle reaches of the Kikori River (Rhoads 1980) (Figure
1). As ethnographic information for the Kikori region was
rudimentary, Rhoads undertook his own ethnoarchaeological
HISTORICISING THE PRESENT:Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New GuineaIan J. McNiven1, Bruno David1, Ken Aplin2, Max Pivoru3, William Pivoru3, Alex Sexton4,
Jonathan Brown1, Chris Clarkson5, Kate Connell5, John Stanisic6, Marshall Weisler5, Simon
Haberle7, Andrew Fairbairn5 and Noel Kemp8
1 School of Geography and Environmental Science, Monash University, Clayton, VIC 3800, Australia [email protected], [email protected], [email protected]
2 Australian National Wildlife Collection, CSIRO Division of Ecosystem Sciences, PO Box 284, Canberra, ACT 2601, Australia [email protected]
3 Kopi Village, Kikori District, Gulf Province, Papua New Guinea4 PO Box 624, Mt Isa, QLD 4825, Australia [email protected] School of Social Science, The University of Queensland, Brisbane,
QLD 4072, Australia [email protected], [email protected], [email protected], [email protected]
6 Queensland Museum, South Brisbane, QLD 4101, Australia [email protected]
7 Research School of Pacific and Asian Studies, The Australian National University, Canberra, ACT 0200, Australia [email protected]
8 50 Derwent Avenue, Lindisfarne, TAS 7015, Australia [email protected]
AirdHills
Ivi River
Saboro River
Pinini Creek
Emo
Wokoi Amoho
Fly R.Digul R
.
WESTPAPUA PAPUA NEW
GUINEA
TorresStraitKilometres
NStudy area
2000
Veiru Creek
Kikori R
iver
Mati Ck
Utiti Ck
Kikiniu
Puriau
Kikori
Ibira HerekunaOuloubomoto
Waira
Kopi/Mampaiu
RupoEpe Amoho
kilometres100
Figure 1 Study area.

42 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
investigations to develop a detailed settlement-subsistence model
for the local Rumu (Kairi) people. This model and historical
analogue formed the basis of a series of hypotheses which were
tested against archaeological data derived from excavation of
a range of local open and rockshelter sites. Rhoads (1980:iv)
concluded that while occupation extended back to 3000 years ago,
‘subsistence strategies appear to have changed little during that
time from what is known today’. However, 1200–1500 years ago
settlement patterns changed with the development of permanent
or semi-permanent villages along major waterways, increased
use of temporary, specialised inland encampments (used mostly
during the dry season), and increased use of exotic commodities,
especially pottery and chert, associated with elaborated prestige-
goods exchange systems (Rhoads 1980:iv, 1982a:140-141).
Although Rhoads (1980:243) suggested ‘a general continuity
in land-use patterns over roughly the last 1500 years’, he
concluded that ethnographically-known settlement-subsistence
arrangements best match the Proto-historic period of the past
400 years. These changes were seen to ‘represent intense periods
of coastal trade linking sago-producing peoples of the Papuan
Gulf with more easterly sited pottery-making communities’
associated with the ethnographically-known Motuan hiri trade
system (see Dutton 1982; Rhoads 1982a). None of these cultural
changes was seen to reflect environmental change, as Rhoads
(1980:97) assumed that the natural environmental context of
the Rumu had ‘remained substantially unchanged over the past
6000 years’ (following the work of Nix and Kalma 1972 and
Powell 1976).
In part we endorse Rhoads’ use of ethnographic information
in archaeological interpretation. That is, broadly framed by the
direct-historical approach to analogy (Lyman and O’Brien 2001),
continuity between the ethnographic present and archaeological
past needs to be demonstrated and not simply assumed. This
demonstration is undertaken by examining how far back in time
archaeological correlates of ethnographic practices extend. By
historicising present (ethnographic) practices and investigating
when and how such practices developed, a dynamic view of the
past is achieved. The key is the investigation of cultural similarities
(continuities) and differences (discontinuities) between past and
present (Gould 1980). This diachronic approach to ethnographic
information circumvents the problematic use of ethnographic
analogies which simply assume continuities between past and
present and promote static views of the past (see Hiscock 2008;
Spriggs 2008; Wobst 1978). Yet a potential problem with many
applications of ethnographic analogy is that the level of generality
of ethnographic information potentially allows the variability
of a wide range of archaeological data to be overlooked and
subsumed. As such, actual differences between the ethnographic
present and archaeological past may become lost and masked as
continuities (see Gould and Watson 1982:375). Our approach
elaborates Rhoads’ modelling of Rumu settlement-subsistence
practices by focusing on the ethnography and archaeology of a
single Rumu site – namely a rockshelter hunting camp known as
Epe Amoho. As we have demonstrated elsewhere (e.g. David et
al. 1994; McNiven et al. 2009), documenting ethnographic use
of a single site allows detailed and specific investigation of the
temporal depth of the ethnographic present. Such fine-grained
understanding needs to be matched by fine-grained analysis
and dating of a broad range of activities contained within the
archaeological record of the site and detailed understanding
of the site’s palaeoenvironmental context. Although Rhoads
assumed environmental stability during human occupation of
the region, such an assumption also needs to be tested as much
as the ethnographic record needs to be tested. Furthermore,
Rhoads’ chronology for Rumu sites needs to be tested and refined
as many of his radiocarbon dates can be considered unreliable
(see David 2008 for details). This paper documents the broader
and specific ethnographic context of Epe Amoho and details the
results of complementary and comparative archaeological and
palynological analyses of excavated deposits from the site.
Cultural Backdrop – Rumu Hunters and FishersRumu number around 700 people and inhabit the mid-Kikori
River region of the Gulf of Papua (Busse et al. 1993; David et
al. 2008; Petterson and Petterson 2008) (Figure 1). Although
the time depth of the Rumu as a distinctive cultural group is
unknown, archaeological research indicates a human presence
in the Kikori region dating back to at least 11,000 years ago, with
major increases in occupation occurring within the past 2500
and especially the past 500 years (David 2008; David et al. 2007,
2010; Rhoads 1980). Sustained European contact with the Rumu
began in 1912 with the establishment of a government station
at Kikori located 26km inland from the coast (Hope 1979:43).
The geographical world of the Rumu is riddled with small and
major waterways weaving through karst plains with a veneer of
alluvium and isolated limestone hill remnants flanked by rugged
limestone country in the form of polygonal karst (limestone
towers and associated depressions) and limestone plateau with
karst corridors (Löffler 1974:12, 1977:54-57). This lowland
landscape is mostly 10–40m asl and is covered in dense tropical
rainforest and sago swamps. Annual rainfall is 5000–8000mm,
falling mostly between May and September. Flooding occurs
annually and, indeed, today all houses, whether river-bordering
or not, are elevated on high stilts to avoid the swampy and often
flooded terrain. Movement across this fluid terrestrial landscape
is usually by foot and dugout canoe.
As with many coastal and lowland hinterland groups of
the Papuan Gulf region, the Rumu are hunter-gatherer-fisher-
cultivators who process sago palms into a starch staple (Busse
et al. 1993; Knauft 1993; Rhoads 1980, 1982b). Rhoads’ (1980)
ethnoarchaeological research remains the most detailed
investigation of lowland rainforest settlement-subsistence
patterns in the Kikori River basin. Terrestrial and aquatic animals
are the main source of protein. Domesticated pigs, chickens and
cassowaries have minor dietary significance and are ‘eaten only
at important feasts, which occur sporadically’ (Rhoads 1980:33).
Stands of sago (Metroxylon sp.) are planted in areas of cleared
or managed rainforest and the labour-intensive process of
pounding felled palm trunks and washing the macerated pith to
isolate starch is undertaken mainly by women. In the 1970s, many
Rumu women continued using hafted stone sago pounders in
preference to introduced metal pipe-tipped pounders (Rhoads
1980:27). At least 129 ‘ethnobotanical’ varieties of plants are
cultivated by the Rumu, including subsistence plants – pandanus
(9 varieties), sago palm (26 varieties), banana (11 varieties) and
sweet potato (3 varieties) – and utility plants such as bamboo
(8 varieties) for cooking and water storage vessels, yaemi (in
Rumuhei, the language of the Rumu) trees for canoe logs, Areca

43Number 71, December 2010
Ian J. McNiven et al.
sp. (black palm) for bows and house flooring, and the fish poison
Derris spp. (3 varieties) (Rhoads 1980:Table III-4). Significantly,
‘the contemporary subsistence and settlement modes of the
Kairi [Rumu] bear little evidence of over one hundred years of
European contact’ (Rhoads 1980:21).
Rainforest HuntingHunting is mostly a male activity among the Rumu. Rhoads
(1980:Table III-7) recorded 29 ‘ethnozoological species’
hunted in the study area. Ground-dwelling species include pig,
cassowary, wallaby (Dorcopsis veterum and Thylogale brunii),
cuscus (?Phalanger gymnotis), bandicoot (Echymipera rufescens),
rats (e.g. Uromys caudimaculatus), monitor lizards (Varanus
spp.), snail, frog, and birds such as pigeon and bush hen
(Megapodiidae). Lower canopy species comprise cuscuses and
snakes, and high canopy species include flying fox and birds such
as hornbill, cockatoo and parrot. Of these, pig and cassowary are
‘the most eagerly sought’ (Rhoads 1980:39). Hunting techniques
focus on the use of bows and arrows, shotguns and clubs, with
dead fall and snare traps also used by previous generations
(Rhoads 1980:39-40). Dogs assist with pig, cassowary and wallaby
hunting in particular (e.g. Rhoads 1980:34). The introduction of
shotguns ‘does not appear to have greatly altered general hunting
strategies’ (Rhoads 1980:46). Small bats (Microchiroptera) and
flying foxes (Dobsonia spp.) are obtained from limestone caves.
Riverine Fishing and ShellfishingThe Kikori River basin has the second highest range of fishes for
any river system in New Guinea (Allen et al. 2008:7). Rhoads
(1980:35) observed that along with sago starch, ‘riverine
resources supply the most consistent source of food’. Forty
‘ethnotaxonomic species’ are identified including water rat,
crocodile, turtles and birds (Rhoads 1980:Table III-7). This list
also includes 24 types of fishes (e.g. catfish, barramundi, mullet,
gudgeon, mackerel and eel) and four types of crustaceans (e.g.
crab, prawn and crayfish), and six types of molluscs. Apart from
major waterways such as the Kikori River, areas of rainforest
seasonally inundated by flood waters provide important dry
season fishing locations. ‘Large doline ponds’, ‘ephemeral
ponds’ and ‘bush creeks’ restocked with fishes by the flooding
of waterways ‘provide an important dry season focus for fishing’
by men and women, and represent ‘extremely lucrative resource
localities’ (Rhoads 1980:35-38). Local fishing methods employed
variously by both men and women include bows and arrows,
fishing lines and hooks, nets, weirs and drugging using Derris sp.
(pers. obs. 2005-2009; Georges et al. 2008; Rhoads 1980:35-36).
Freshwater turtles are ‘an important source of protein’ in
the Kikori region (Georges et al. 2008:709). Georges et al.
(n.d.:3) state that pig-nosed turtles (Elseya novaeguineae) are
‘commonly caught on fishing lines and in fishing nets as part
of general hunting and fishing activity, but we were told that
the best time to catch them was in the dry season, when they
retreat into tunnels in the riverbanks’. Crustaceans and molluscs
are generally collected by women by hand ‘mainly during the
dry season’ when the lower banks of waterways – the preferred
habitat of these animals – become exposed and easier to access
due to receding water levels (Rhoads 1980:37, 43, 225). Shellfish,
available only during the dry season, include Batissa violacea,
Hyeidella sp., Melanoides sp. and Vivaparidae (Sam Kaiwari and
Kupere Karamavi, pers. comm., 2009; Rhoads 1980:68). Neritina
sp. – known as kipiri in Rumuhei – and the locally identified
towoi (species unidentified) are the only gastropods ‘easily found’
during the wet season (Rhoads 1980:43).
Settlement Patterns and Hunting CampsRumu settlement patterns revolve around permanently settled
nuclear villages along major rivers and a complex arrangement
of temporary camps (often rockshelters) used mostly during
Edge of cleared area
N
60metres
Overhang
Rainforest
Rainforest
A
Bamboo grove
Limestone karst Tree trunk >0.5m diameter
Black palm
Figure 2 Plan of Epe Amoho.

44 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
the dry season (October to April) located close to ‘inland’ water
sources. Both site types are used by men, women and children.
Temporary camps generally fall into one of two groups – those
associated with generalised land-use and social refuges away
from village life (usually located more than 2.5-3km from a
village) and specialised sites associated with sago production and
seasonal procurement of aquatic fauna (e.g. spawning mullet
and turtle eggs in the wet season and doline pond fishes in the
dry season) (Rhoads 1980:45, 62, 65-76).
Epe Amoho
Site DescriptionEpe Amoho is a limestone rockshelter designated KG14 by David
et al. (2005). The site is named after Epe Creek located 300m
to the north. The site name derives from Epe (a place name)
and amoho (‘rockshelter’ or ‘cave’ in Rumuhei). It is located on
the margins of a heavily-forested swampy alluvial plain adjacent
to limestone polygonal karst country at the headwaters of Utiti
Creek 5km west of the Kikori River and 46km inland from the
coast (Figure 1). Epe Amoho is positioned at the base of a c.5–
10m-high rugged limestone cliff-line draped in ferns and vines
and set amongst dense tropical rainforest (Figures 2-3).
The curvilinear rockshelter has a length of c.60m and is
divided into two sections, a c.30m-long western section with
a northwest aspect, and a c.30m-long eastern section with a
northeast aspect. The two sections are separated by a northeast-
southwest oriented large tunnel cave that extends into the
darkness. The western end of the site is the driest owing to the
greater extent of overhang. Along its length, the area immediately
in front of the rockshelter is relatively open, being repeatedly
cleared of vegetation upon its occupation or visitation. However,
the area more than 5–10m in front of the shelter is invariably
covered by thick rainforest, especially stands of bamboo and
black palm. The site is flooded or soggy during the wet season.
Cultural ContextEpe Amoho has economic and spiritual significance to the
Himaiyu clan. The following information and quotes were
provided by Max and William Pivoru, two senior members of the
Himaiyu clan during the period of excavation of Epe Amoho and
before the results of our excavations were apparent. Subsequently,
further interviews on Himaiyu settlement-subsistence activities
with a larger number of clan members (men and women) were
undertaken following completion of an early draft of this paper
to shed further light on relevant points. Himaiyu clans-people
are the traditional owners of the site which is considered ‘one
of the historical places in the history of the clan’. Epe Amoho
is the largest hunting camp used by the Himaiyu clan. Hunting
camps are known as iapu in Rumuhei (and sometimes by the
Motu term kombati). The site was used often in association
with smaller hunting camps such as Wokoi Amoho, Inorave and
Tirimu. In this situation, men would visit other hunting camps
while women would stay back and ‘look after’ Epe Amoho. In the
past access to the site from villages was by canoe up Utiti Creek
followed by a 5–10 minute walk.
Animal food resources obtained while camping at Epe Amoho
include pigs and cassowaries hunted from surrounding forests,
bats from a nearby cave, and fishes and prawns captured by hand
from nearby Epe Creek. Cultivated plant resources located in
front of Epe Amoho include bamboo, pandanus, poison roots,
sago palm, and black palms.
Camping at Epe Amoho occurs every year and is a dry season
activity (Figure 3). Use of the site occurs ‘when people run out of
food, especially meat’ within the village. ‘If we don’t go there then
we don’t have enough protein to eat’. As various clan members
pointed out, Rumu clans-people undertake both wet and dry
season hunting. Wet season hunting takes place at a time when
much of the land is flooded or boggy, and at this time terrestrial
animals take refuge on high ground. Clan members travel in
multiclan groups by canoe to elevated areas, surrounding high
areas where animals are concentrated; ‘kill what you can there,
then go to another high ground’. During expeditions, hunters
travel for the day to return to their home village at the end of
the day. Dry season hunting takes on a different hue – extended
periods are spent at camps located within a clan’s hunting
ground. Hunters depart early in the morning and return to the
hunting camp at the end of the day. Sometimes families and
hunting groups use major hunting camps such as Epe Amoho
in association with specialised camps, such as sago camps. Prior
to the extra commitments of village life after contact (such as
schooling, church), people camped at Epe Amoho ‘for maybe six
Figure 3 Remains of recent camping at Epe Amoho, 16 April 2005. Note planted bamboo groves in background (Photograph: Ian J. McNiven).
Figure 4 Square A after excavation, Epe Amoho. Looking southwest, 9 April 2006 (Photograph: Ian J. McNiven).

45Number 71, December 2010
Ian J. McNiven et al.
months or four months, like it was their home’. Camping could
take place anywhere along the length of the rockshelter.
ExcavationA single 1m x 1m pit (Square A) was excavated within Epe
Amoho. The location of Square A was selected for excavation
as it is higher and drier than other parts of the site and is thus
considered to have been attractive for occupation throughout the
site’s history of use (Figure 4). Furthermore, this location exhibits
a larger overhang providing more opportunity for a wider range
of activities (e.g. cooking, sleeping, tool preparation etc). While
Square A represents a small proportion of Epe Amoho, it is
located within the confined area of the prime camping location
at the site. As such, Square A is considered to provide a reliable
record of broad usage patterns of Epe Amoho though time.
Excavation was stopped at a depth of 109cm owing to
problems of accessibility and the apparent lack of cultural
materials in basal XUs. The excavation was undertaken in 33
excavation units (XUs) (which were often subdivided into sub-
XU parts ‘a’, ‘b’ and ‘c’ to avoid mixing of sediments of different
colour and texture) with a mean thickness of 3.2cm (range: 0.4-
5.3cm). A single sloping slab of limestone roof fall was uncovered
across the northern half of the pit. Weight (to nearest 100g) and
volume (to nearest 500ml) was taken for excavated sediments
for each XU. At least five depth elevations (to nearest mm) were
taken at the beginning and end of each XU. Sediments were wet-
sieved (freshwater) through 2.1mm mesh. Small bulk sediment
samples were collected from each XU prior to sieving. In addition,
22 small samples of sediment for pollen analysis were collected
in 1cm-wide vials pressed into the south section of the pit at
5cm depth intervals (Figure 5). The pit was backfilled with local
sediments taken well away from the site. Excavation took place
between 4 and 9 April 2006.
Stratigraphy and SedimentsSquare A sediments were divided into three major stratigraphic
units (SUs) (Figure 5). SUs 1 to 3a extend from the ground
surface to a depth of between 7 and 22cm and grade with depth
from dark grey/brown loose crumbly clay (SUs 1-2) to brown
sticky clay (SU3a). They contain most cultural materials (e.g.
charcoal, stone artefacts, bone and shell) recovered from the site
with items of European origin (e.g. glass and metal) restricted
to SUs 1 and 2 located at a maximum depth of 18cm below the
ground surface. SUs 3b and 3c represent over 90% of excavated
sediments and comprise mostly brown sticky clays with very
small amounts of cultural materials (charcoal, bone and shell).
In addition to the large limestone slab, fragments of limestone
roof fall were encountered across the lower sections of the square
in SU3c mostly between 82 and 97cm below the ground.
An intrusive ‘pit’ feature was sectioned by the east wall of the
square. The pit averages around 17cm in diameter and extends
from the top of SU3a to maximum depth of 62cm below the
ground surface in SU3c. The feature was created at a time when
the top of SU3a was the ground surface. A fragment of glass
recovered from the base of the pit confirms that the feature was
filled by sediments that included SU2 (which contains a range of
European items including glass). The pit is interpreted as a post-
hole, and is consistent with the Himaiyu practice of building stilt
structures against the rockshelter during periods of encampment.
Bulk sediment samples for each XU were subjected to Low
Angle Laser Scattering (LALS) particle size analysis using a
Coulter 100LS particle size analyser. Samples were dispersed
in 5%v sodium tetra-pyrophosphate (Na4P2O7) and agitated
in an ultrasonic bath for 30 minutes. Prior to analysis samples
were vigorously dispersed using a vortex mixer and analysed.
Percentages of organic carbon and CaCO3 were determined by
weighing dry samples, treating with 30% H2O2 over a period of
3 months, until reaction ceased, additional treatment with HCl,
and weighing of the remaining sample.
Sediments are dominated by very fine particles. Particles
finer than 20µm comprise over 60% by volume of all samples,
and over 70% for the majority. The modal particle size of all
samples was between 2 and 10µm (clay and fine silt). Overall,
particle size distributions are consistent with an alluvial
depositional environment. The proximity of the rockshelter to
swampland and waterways, combined with high rainfall make
suspended sediment in flood water a primary source of sediment
entering the rockshelter. Visual stratigraphic layers within
the profile identified at the time of excavation do not exhibit
distinct particle size characteristics. Furthermore, within the
sediment profile there is limited soil development which is not
unexpected given the limited age of the deposit (<2850 years).
Figure 5 Stratigraphy, Square A, Epe Amoho.

46 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
There is evidence for translocation of fine clays from the upper
part of the sequence (0-20cm) into the lower profile (20-65cm),
indicative of illuviation as a result of downward percolation of
water. The existence of coarse particles within the soil is probably
evidence of rock fall from the roof of the rockshelter, either in
the dramatic fashion shown at the base of the profile (with large
limestone rocks), or less abrupt additions from water movement
over the surface of the rockshelter roof, or disintegration of the
limestone as a result of human activity (e.g. heat from fires).
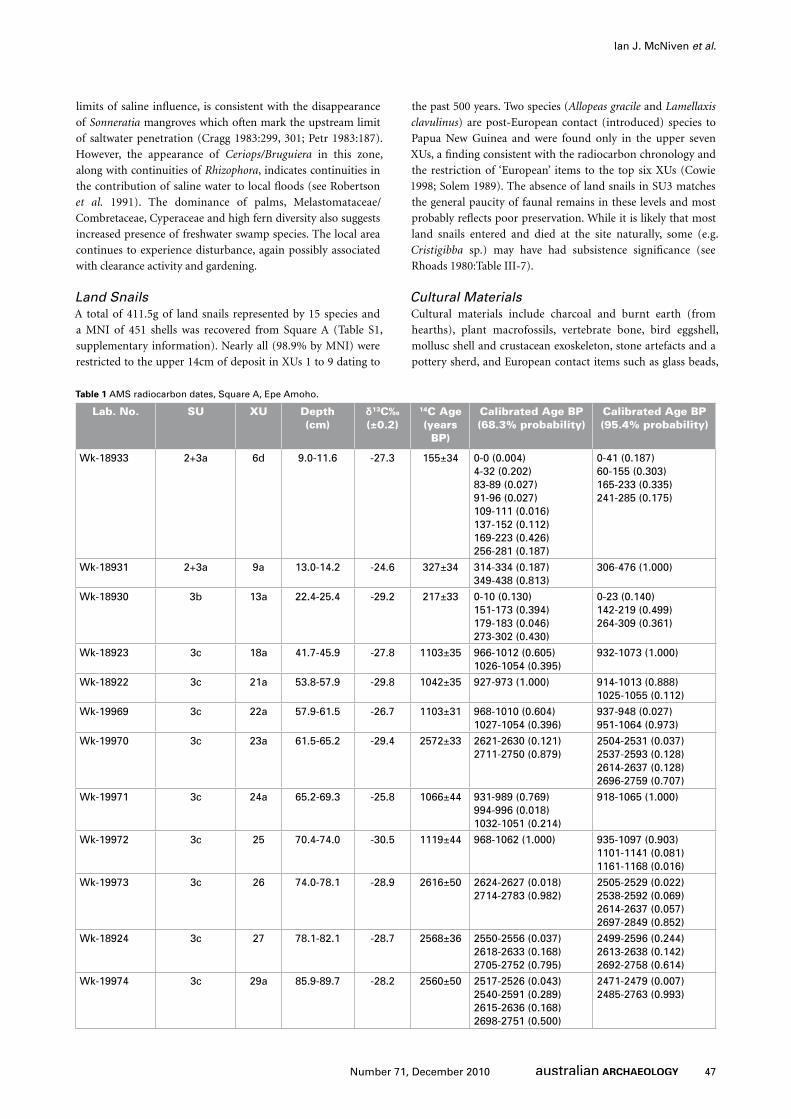
Radiocarbon Dating and ChronologyTwelve AMS radiocarbon dates were obtained for Epe Amoho
(Table 1). All dates were obtained on single fragments of
wood charcoal and determined by the University of Waikato
Radiocarbon Dating Laboratory. Radiocarbon dates were
calibrated into calendar years using the online calibration
program Calib 6.0 (Stuiver and Reimer 1993) and the IntCal09
dataset (Reimer et al. 2009).
Dates cluster into three age groupings separated by
considerable periods of time (Figure 6). The oldest grouping
(Early Phase) comprises four almost identical dates ranging
from 2616±50 BP to 2560±50 BP calibrating to between
c.2500 and c.2850 cal BP (years before AD 1950). These
dates take in the lower half of SU3c over a vertical distance
of 28cm between 62 and 90cm below the ground. The single
dating inversion within the Early Phase (Wk-19970) probably
represents post-depositional upward movement of the single
charcoal fragment upon which the date was obtained. The next
grouping of dates (Middle Phase) comprises five similar dates
ranging from 1119±44 BP to 1042±35 BP and calibrating to
between c.900 and c.1200 cal BP. These dates span the upper
half of SU3c over a vertical distance of 32cm between 42 and
74cm below the ground surface. The third and final grouping
of dates (Recent Phase) consists of three dates ranging between
327±34 BP and 155±34 BP which calibrate to the past 500
years. This group takes in the upper 25cm of deposit and SU1
to the upper sections of SU3a. That the third group of dates
takes in the past 150 years is consistent with the recovery of
items of European origin (e.g. glass and metal) and ‘introduced’
fauna (see below) in SUs 1 and 2. While the Early and Middle
Phases reveal some stratigraphic overlap and mixing (one
radiocarbon dating inversion – Wk-19970) between XUs 23a
and 25 spanning a vertical distance of 12cm between 62 and
74cm below the ground surface, overall the deposit exhibits
good chronostratigraphic integrity.
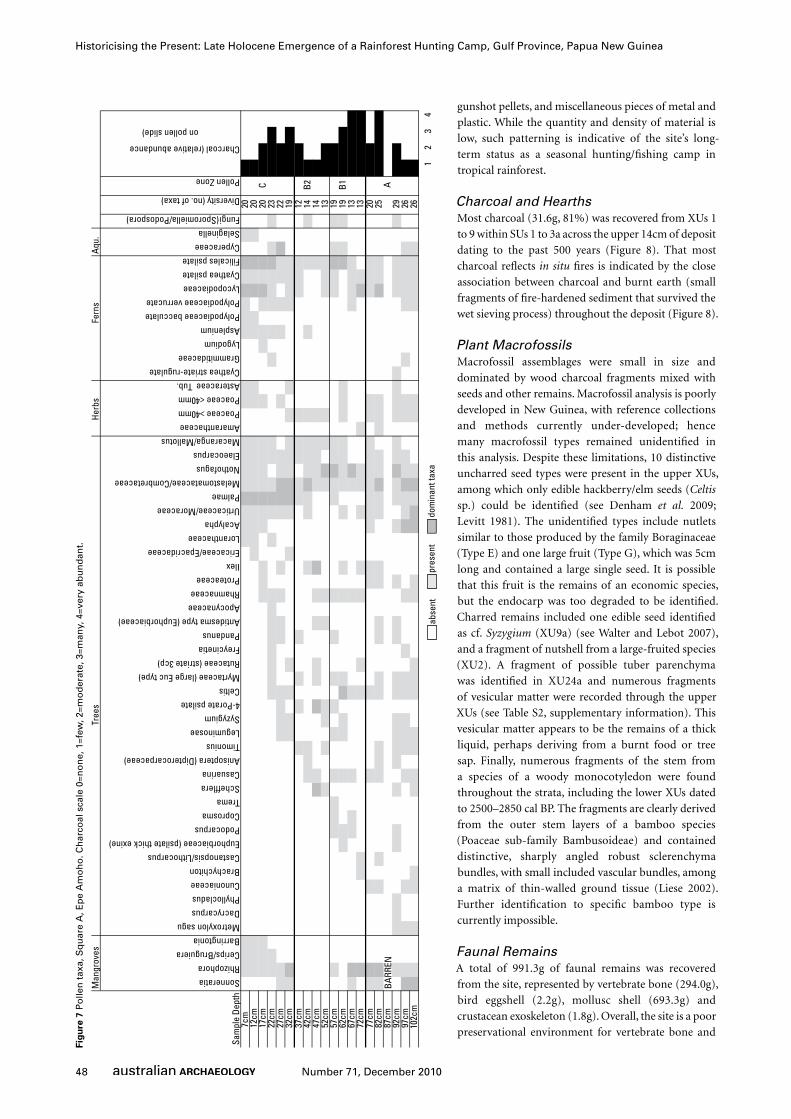
Pollen and PalaeoenvironmentsPreparation of the pollen samples removed from the south
section followed standard procedures described in Bennett
and Willis (2001) with the addition of a Lycopodium spike to
determine relative concentrations of palynomorphs. Pollen
concentrations and preservation were generally low in most
slides and was considered too low for standard percentage
pollen analysis. Each slide was counted to a minimum of 100
pollen grains and then scanned for the presence of further taxa.
A presence/absence diagram was then constructed (Figure 7).
Charcoal concentration (relative to the Lycopodium spike) was
estimated on a 5-point scale (0=none to 4=very abundant).
Pollen zones where determined using a Constrained Incremental
Sum of Squares (CONISS) cluster analysis within the programme
Psimpoll (Bennett and Willis 2001). The diversity of taxa within
each sample is determined by simple addition of the number
of taxa encountered in any one sample. The pollen diagram is
divided into three significant zones (based on the pollen CONISS
statistics) which correspond stratigraphically to the temporal
zones (phases) identified in the archaeological analysis.
Zone A (77-102cm depth, c.2500-2850 cal BP): The saline-
tolerant mangroves Sonneratia and Rhizophora dominate this zone
with generally low levels of charcoal and fern spores. Sonneratia
prefers brackish to near-freshwater habitats and ‘recent mud banks
and accreting inner river curves’ and often allows development of
sediment conditions favourable to the establishment of Rhizophora
(Cragg 1983:309; Paijmans 1983:229). This mixture of lower
salinity (Sonneratia) and higher salinity (Rhizophora) mangrove
species suggests floodwaters deriving from diverse fresh and saline
waterways. A high diversity of rainforest tree taxa is recorded with
a range of local and long-distance dispersal taxa present such
as Nothofagus, Dacrycarpus and Phyllocladus (derived from the
Highlands of PNG). These taxa may have been dispersed by wind
or more likely through river floodwaters carrying upland taxa
into the lowlands. Metroxylon sagu (sago) is present in this zone
suggesting that sago was present near the site. This may represent
a low level of exploitation as intense harvesting of sago palms
will prevent flowering of the plant and reduce or eliminate the
production of pollen. Grasses and the edible herb Amaranthaceae
are also present.
Zone B (37-72cm, c.900-1200 cal BP): Mangrove pollen is
less important although saline-inhabiting Rhizophora dominates
the pollen assemblage in the lower part of this zone. Low diversity
of pollen taxa suggests that the local area has been disturbed,
possibly through human agency, and primary rainforest cover
has been reduced. This is further supported by the presence of
pioneer woody plants such as Trema, Schefflera and Casuarina.
Grasses and the edible herb Amaranthaceae continue to be
present. The pollen assemblage in this zone reflects high levels
of forest disturbance, most likely clearance activity near the site,
possibly associated with site maintenance and gardening. Higher
levels of charcoal in this zone probably reflect hearths within
the rockshelter and not in-washing of environmental charcoal as
firing of local rainforest is unlikely given high levels of moisture
and rainfall.
Zone C (7-32cm, c.0-500 cal BP): Mangrove pollen has
greater diversity with the appearance of the freshwater-
tolerant taxa Barringtonia and the aquatic fern Selaginella. The
appearance of Barringtonia, which is found often beyond the
0 500 1000 1500 2000 2500 3000
Wk-19969 Wk-19971 Wk-18922 Wk-18931 Wk-18930 Wk-18933
Wk-19973 Wk-19970 Wk-18924 Wk-19974 Wk-19972 Wk-18923
Age (cal BP)
Figure 6 Calibrated AMS radiocarbon dates, Square A, Epe Amoho (white=95.4% probability, black=68.3% probability).
47Number 71, December 2010
Ian J. McNiven et al.
limits of saline influence, is consistent with the disappearance
of Sonneratia mangroves which often mark the upstream limit
of saltwater penetration (Cragg 1983:299, 301; Petr 1983:187).
However, the appearance of Ceriops/Bruguiera in this zone,
along with continuities of Rhizophora, indicates continuities in
the contribution of saline water to local floods (see Robertson
et al. 1991). The dominance of palms, Melastomataceae/
Combretaceae, Cyperaceae and high fern diversity also suggests
increased presence of freshwater swamp species. The local area
continues to experience disturbance, again possibly associated
with clearance activity and gardening.
Land SnailsA total of 411.5g of land snails represented by 15 species and
a MNI of 451 shells was recovered from Square A (Table S1,
supplementary information). Nearly all (98.9% by MNI) were
restricted to the upper 14cm of deposit in XUs 1 to 9 dating to
the past 500 years. Two species (Allopeas gracile and Lamellaxis
clavulinus) are post-European contact (introduced) species to
Papua New Guinea and were found only in the upper seven
XUs, a finding consistent with the radiocarbon chronology and
the restriction of ‘European’ items to the top six XUs (Cowie
1998; Solem 1989). The absence of land snails in SU3 matches
the general paucity of faunal remains in these levels and most
probably reflects poor preservation. While it is likely that most
land snails entered and died at the site naturally, some (e.g.
Cristigibba sp.) may have had subsistence significance (see
Rhoads 1980:Table III-7).
Cultural MaterialsCultural materials include charcoal and burnt earth (from
hearths), plant macrofossils, vertebrate bone, bird eggshell,
mollusc shell and crustacean exoskeleton, stone artefacts and a
pottery sherd, and European contact items such as glass beads,
Table 1 AMS radiocarbon dates, Square A, Epe Amoho.
Lab. No. SU XU depth (cm)
δ13C‰ (±0.2)
14C Age (years
BP)
Calibrated Age BP (68.3% probability)
Calibrated Age BP (95.4% probability)
Wk-18933 2+3a 6d 9.0-11.6 -27.3 155±34 0-0 (0.004)4-32 (0.202)83-89 (0.027)91-96 (0.027)109-111 (0.016)137-152 (0.112)169-223 (0.426)256-281 (0.187)
0-41 (0.187)60-155 (0.303)165-233 (0.335)241-285 (0.175)
Wk-18931 2+3a 9a 13.0-14.2 -24.6 327±34 314-334 (0.187)349-438 (0.813)
306-476 (1.000)
Wk-18930 3b 13a 22.4-25.4 -29.2 217±33 0-10 (0.130)151-173 (0.394)179-183 (0.046)273-302 (0.430)
0-23 (0.140)142-219 (0.499)264-309 (0.361)
Wk-18923 3c 18a 41.7-45.9 -27.8 1103±35 966-1012 (0.605)1026-1054 (0.395)
932-1073 (1.000)
Wk-18922 3c 21a 53.8-57.9 -29.8 1042±35 927-973 (1.000) 914-1013 (0.888)1025-1055 (0.112)
Wk-19969 3c 22a 57.9-61.5 -26.7 1103±31 968-1010 (0.604)1027-1054 (0.396)
937-948 (0.027)951-1064 (0.973)
Wk-19970 3c 23a 61.5-65.2 -29.4 2572±33 2621-2630 (0.121)2711-2750 (0.879)
2504-2531 (0.037)2537-2593 (0.128)2614-2637 (0.128)2696-2759 (0.707)
Wk-19971 3c 24a 65.2-69.3 -25.8 1066±44 931-989 (0.769)994-996 (0.018)1032-1051 (0.214)
918-1065 (1.000)
Wk-19972 3c 25 70.4-74.0 -30.5 1119±44 968-1062 (1.000) 935-1097 (0.903)1101-1141 (0.081)1161-1168 (0.016)
Wk-19973 3c 26 74.0-78.1 -28.9 2616±50 2624-2627 (0.018)2714-2783 (0.982)
2505-2529 (0.022)2538-2592 (0.069)2614-2637 (0.057)2697-2849 (0.852)
Wk-18924 3c 27 78.1-82.1 -28.7 2568±36 2550-2556 (0.037)2618-2633 (0.168)2705-2752 (0.795)
2499-2596 (0.244)2613-2638 (0.142)2692-2758 (0.614)
Wk-19974 3c 29a 85.9-89.7 -28.2 2560±50 2517-2526 (0.043)2540-2591 (0.289)2615-2636 (0.168)2698-2751 (0.500)
2471-2479 (0.007)2485-2763 (0.993)
48 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
gunshot pellets, and miscellaneous pieces of metal and
plastic. While the quantity and density of material is
low, such patterning is indicative of the site’s long-
term status as a seasonal hunting/fishing camp in
tropical rainforest.
Charcoal and HearthsMost charcoal (31.6g, 81%) was recovered from XUs 1
to 9 within SUs 1 to 3a across the upper 14cm of deposit
dating to the past 500 years (Figure 8). That most
charcoal reflects in situ fires is indicated by the close
association between charcoal and burnt earth (small
fragments of fire-hardened sediment that survived the
wet sieving process) throughout the deposit (Figure 8).
Plant MacrofossilsMacrofossil assemblages were small in size and
dominated by wood charcoal fragments mixed with
seeds and other remains. Macrofossil analysis is poorly
developed in New Guinea, with reference collections
and methods currently under-developed; hence
many macrofossil types remained unidentified in
this analysis. Despite these limitations, 10 distinctive
uncharred seed types were present in the upper XUs,
among which only edible hackberry/elm seeds (Celtis
sp.) could be identified (see Denham et al. 2009;
Levitt 1981). The unidentified types include nutlets
similar to those produced by the family Boraginaceae
(Type E) and one large fruit (Type G), which was 5cm
long and contained a large single seed. It is possible
that this fruit is the remains of an economic species,
but the endocarp was too degraded to be identified.
Charred remains included one edible seed identified
as cf. Syzygium (XU9a) (see Walter and Lebot 2007),
and a fragment of nutshell from a large-fruited species
(XU2). A fragment of possible tuber parenchyma
was identified in XU24a and numerous fragments
of vesicular matter were recorded through the upper
XUs (see Table S2, supplementary information). This
vesicular matter appears to be the remains of a thick
liquid, perhaps deriving from a burnt food or tree
sap. Finally, numerous fragments of the stem from
a species of a woody monocotyledon were found
throughout the strata, including the lower XUs dated
to 2500–2850 cal BP. The fragments are clearly derived
from the outer stem layers of a bamboo species
(Poaceae sub-family Bambusoideae) and contained
distinctive, sharply angled robust sclerenchyma
bundles, with small included vascular bundles, among
a matrix of thin-walled ground tissue (Liese 2002).
Further identification to specific bamboo type is
currently impossible.
Faunal RemainsA total of 991.3g of faunal remains was recovered
from the site, represented by vertebrate bone (294.0g),
bird eggshell (2.2g), mollusc shell (693.3g) and
crustacean exoskeleton (1.8g). Overall, the site is a poor
preservational environment for vertebrate bone and
Man
grov
esTr
ees
Her
bsFe
rns
Aqu
.
Sam
ple
Dep
th
SonneratiaRhizophoraCerips/BruguieraBarringtoniaMetroxylon saguDacrycarpusPhyllocladusCunoniaceaeBrachychitonCastanopsis/LithocarpusEuphorbiaceae (psilate thick exine)PodocarpusCoprosmaTremaScheffleraCasuarinaAnisoptera (Dipterocarpaceae)TimoniusLeguminosaeSyzygium4-Porate psilateCeltisMyrtaceae (large Euc type)Rutaceae (striate 3cp)FreycinetiaPandanusAntidesma type (Euphorbiaceae)ApocynaceaeRhamnaceaeProteaceaeIlexEricaceae/EpacridaceaeLoranthaceaeAcalyphaUrticaceae/MoraceaePalmaeMelastomataceae/CombretaceaeNothofagusElaeocarpusMacaranga/MallotusAmaranthaceaePoaceae >40mmPoaceae <40mmAsteraceae Tub.Cyathea striate-rugulateGrammitidaceaeLygodiumAspleniumPolypodiaceae bacculatePolypodiaceae verrucateLycopodiaceaeCyathea psilateFilicales psilateCyperaceaeSelaginellaFungi (Sporomiella/Podospora)
Diversity (no. of taxa)
Pollen Zone
Charcoal (relative abundance
on pollen slide)
7cm
2012
cm20
17cm
20C
22cm
2327
cm22
32cm
1937
cm12
42cm
14B
247
cm14
52cm
1357
cm19
62cm
19B
167
cm13
72cm
1377
cm20
82cm
2587
cmB
ARR
ENA
92cm
2997
cm26
102c
m26
pres
ent
abse
ntdo
min
ant t
axa
12
34
Fig
ure
7 P
olle
n t
axa,
Sq
uar
e A
, Ep
e A
mo
ho
. Ch
arco
al s
cale
0=n
on
e, 1
=few
, 2=m
od
erat
e, 3
=man
y, 4
=ver
y ab
un
dan
t.

49Number 71, December 2010
Ian J. McNiven et al.
crustacean exoskeleton. The largest quantities of bone (>20g/
XU) were recovered from XUs 1 to 6 (SUs 1 to 3a), while below
XU9 the quantities drop away to miniscule amounts (<2g/XU).
Even in the uppermost layers of the deposit, the bone is visibly
degraded, with most teeth represented only by enamel caps (i.e.
dentine rotted away). A further indicator of the degraded state of
the assemblage is the fact that, in most samples, more than 50%
of the surviving bone is burnt to varying degrees. Burnt bone
tends to persist longer than unburnt bone in many sedimentary
contexts due to its lower organic content and slower rate of
breakdown by soil microbial organisms (e.g. Buzon et al. 2005;
Lubinski 1996). Relatively few specimens are complete enough
for identification below family level.
Several features of the bone assemblage point to it being almost
entirely cultural in origin, as distinct from non-human predator
accumulation. One is the absence of any small murid remains –
these and similarly-sized mammals usually make up the bulk of
bone assemblages in rockshelter and cave deposits accumulated
by avian predators (Kusmer 1990; Marshall 1986). Another key
indicator is the high intensity of burning of much of the remains,
consistent with secondary burning in the intense heat within or
immediately beneath a hearth (Asmussen 2009; Stiner et al. 1995).
The few microchiropteran bat bones might be a result of natural
deaths. However, these are also frequently captured and consumed
today, and may well be part of the cultural assemblage.
Mammal: Isolated teeth permitted reliable identification of
two species of wallaby (Dorcopsis sp., Thylogale sp.), two cuscuses
(Spilocuscus sp., Phalanger gymnotis), one bandicoot (Echymipera
rufescens), a fruit bat (Dobsonia sp.), two murid rodents (Uromys
sp., Xenuromys barbatus) and pig (Sus scrofa) (Table 2). One
upper molar of a bandicoot from XU2 is smaller-toothed than
Echymipera rufescens and cannot be allocated with certainty to
any known taxon. With the exception of a wallaby bone fragment
in XU25, all identifiable mammal bones were recovered from
XUs 1 to 10, dating to within the past 500 years. The fragments
of pig teeth are too small to allow assessment of individual ages.
All of these taxa are available locally and could have been hunted
from lowland rainforests of the region.
Bird: The only bird bone recorded was a cassowary sternum
of recent age cached on a limestone ledge 4m southwest of
Square A. However, excavations revealed a small quantity (2.2g)
of bird eggshell in XUs 1 to 9 dating to within the past 500 years.
The form (curvature and thin walls) of the shell is consistent
with megapode eggs. Known megapodes of this rainforest region
are the orange-footed megapode (Megapodius reinwardti) and
black-billed brush turkey (Talegalla fuscirostris) and their eggs
can be easily removed from mounded ground nests.
Lizards and Snake: Snakes and agamid lizards form a
consistent part of subsistence practices at the site over the past
500 years. Evidence of monitor lizards (Varanus spp.) is restricted
to XU4a near the surface (Table 2). These reptiles are available
from local rainforests.
Freshwater Turtle: Freshwater turtle bones (18.9g) were
recovered from XUs 1 to 16. While they represent only 6%
(by weight) of the bone assemblage, they formed a relatively
consistent part of the diet over the past 500 years. Four species
of freshwater turtle have been recorded in the Kikori region from
local tidal freshwater creeks and streams, flooded sinkholes and
swamps (Georges et al. 2008).
0.1 1 10 100 1000
1
2
3a
3b
4a
4b
5a
5b
5c
6a
6b
6c
6d
7
8
9a
10a
10b
11a
11b
12a
12b
12c
13a
13b
14a
14b
15a
16a
17a
18a
19a
20a
21a
22a
23a
23b
24a
24b
25
26
27
28a
28b
29a
29b
30a
30b
31
32
33
g / 10kg of deposit
Charcoal
Burnt earth
Exca
vatio
n U
nit
Figure 8 Vertical changes in charcoal and burnt earth density, Square A, Epe Amoho.

50 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
Fish: Fish bones totaling 47.4g and representing at least eight
taxa were found intermittently throughout the Epe Amoho
deposits spanning the past 2500–2850 years, with most bones
(99%) within XUs 1 to 15, dating to the past 900–1200 years (Table
2). Analysis was undertaken using a 10x binocular microscope
due to the small size of bones. Where possible, bones were sorted
to element and side then the presence/absence of burning was
noted. The most common identifiable element was the saccular
otolith (n=17, from at least 11 individuals), the majority of
which were derived from catfish (Ariidae). Cranial bones of ariid
catfish are also present in most samples, distinguished by their
finely granular ectocranial surfaces. In addition, many samples
also contained one or more of the rounded, grinding teeth that
typify members of another family of catfish, the Plotosidae.
Dentigerous elements of a medium-sized member of the family
Lutjanidae (in XU1 and XU8) were the only other identifiable
elements. The maximum diameter of 188 fish vertebrae averaged
only 3.29±1.27mm (range: 1.35-9.23mm) pointing to the small
size of most individual fish. However, some of the plotosid teeth
are probably derived from larger fish. The predominance of ariid
and plotosid bones in the fish bone sample is consistent with
modern fish communities in the large rivers of southern New
Guinea (Allen et al. 2008; Haines 1979; Roberts 1978).
Shark: Ten shark teeth were recorded in XUs 12c to 27 in the
lower levels of the deposit (Table 2). This finding is consistent
with more saltwater-tolerant mangrove pollens in lower levels
(see above). As such, the shark teeth date from around 300–
500 years ago back to 2500–2850 years ago. Of the nine teeth
identified taxonomically, all are Carcharhinus sp. with one tooth
similar to a bronze whaler (Carcharhinus brachyurus) (XU12c)
and two teeth similar to silky shark (Carcharhinus falciformis)
(XUs 15a and 27). The dietary significance of the shark teeth
is difficult to ascertain as sharks have been documented in the
literature only for the lower Kikori River (Enesar Consulting
2005:9.54). However, Himaiyu clans-people today state that saw-
sharks and smaller sharks come up the Kikori River with the tides.
These sharks are not hunted by the Rumu but may be eaten if
caught accidentally in nets.
Shellfish: A total of 693.3g of mollusc shell comprising
six taxa and a MNI of 308 shells was recovered from Square
A (Table S3, supplementary information). The most common
shellfish taxa are Melanoides sp. (50%), Thiara sp. (27%) and
Batissa violacea (17%). As with bone, most mollusc shell (97.7%
by MNI) was restricted to the upper 14cm of deposit in XUs 1
to 9 and SUs 1 to 3a dating to the past 500 years. However, small
quantities of taxonomically-identifiable shell down to XU22b
indicate collection of molluscs back to 900–1200 years ago.
All six types of molluscs are freshwater species available from
local waterways.
Crustacea: The small amount (1.8g) of crustacean exoskeleton
was concentrated in two zones – XUs 4 to 7, dating to the past
500 years, and XUs 24 to 27 dating to between 900–1200 and
2500–2850 years ago. Most surviving fragments of exoskeleton
are burnt, probably as a result of cooking. All fragments are too
small to allow differentiation between crab and crayfish.
Material CultureStone Artefacts: Twenty stone artefacts (including likely stone
artefacts) weighing 6.3g were recovered from XUs 3a to 12a
dating to the past 500 years. The assemblage was divided into
four technological types: flakes (n=10), flaked pieces (n=6), heat
spalls (n=3) and potlids (n=1). The majority of pieces are broken
transversely (68%), with flake fragments (36%) and flaked pieces
(27%) dominating the assemblage. Artefacts show evidence of
uncontrolled heating in 27% of cases, resulting in the presence
of pot lids, heat fractures and heat spalls. Grey chert dominates
the assemblage (90% by number), with the remaining 10% of
artefacts made from a coarse-grained stone, possibly quartzite
or volcanic rock.
The assemblage comprises very small artefacts with a mean
weight of only 0.32±0.6g and a mean length of 8±5mm. The four
complete flakes are smaller than the average for the assemblage
with a mean weight of 0.1g and a mean length of 5mm. Most
flakes are squat, with widely varying marginal contraction or
expansion along the length of the flake. Platforms are quite
small compared to ventral area, with overhang removal found
on all complete flakes and proximal fragments, suggesting blows
were struck close to the edge of prepared platforms. All complete
flakes and distal portions of flakes show feather terminations.
Dorsal scar orientations are all from the proximal to the distal,
consistent with unidirectional flaking from single platform
cores. Cortex is absent from all specimens, consistent with late
stage reduction.
A use-wear and residue study was performed on 16 lithic
pieces to examine artefact life history and function (Table S4,
supplementary information). The three heat spalls and the pot
lid that make up the remainder of the stone artefact assemblage
were not examined. Of the 16 specimens examined, functional
information was obtained for five. Seven pieces were not used
at all, while evidence for use for the remaining four specimens
was inconclusive. Despite the small size of the artefacts, it is
clear that flakes could be used for a variety of tasks, or were
detached from larger tools. The majority of use-traces indicate
a role in wood-working, including working of resinous wood.
One artefact also shows signs of bone-working. Although no
traces of hafting could be detected, it seems likely that these
small tools would have been hafted to have been effective in
hardwood- and bone-working (see Hardy and Sillitoe 2003;
Watson 1995).
Tooth artefact: A dog canine with a biconically drilled hole
through the root was excavated from XU3a at a depth of 2–4cm
and dates to the past 150 years given stratigraphic associations
with items of European origin (Figure 9). The tooth probably
formed part of a body adornment. Drilled dog teeth were also
recovered by Rhoads (1980:A106) from the nearby rockshelters
of Rupo and Herekuna. Dog tooth necklaces and headbands
are well-known ethnographically as highly-valued elements of
Papuan Gulf material culture (Specht and Fields 1984:148-151;
Williams 1924:47; Young and Clark 2001:75) and dog teeth were
traditional items of wealth among the Rumu (Petterson and
Petterson 2008:42). It is also possible some of the excavated shark
teeth were used as artefacts given shark tooth ‘borers’ were used
by other Papuan Gulf groups (Williams 1924:41).
Pottery: A single, tiny, undecorated, earthenware pottery
sherd (0.06g) recovered from XU6a at a depth of 8–11cm
dates to within the past 500 years based on radiocarbon dates
and probably within the past 150 years given its stratigraphic
association with European items. There is no known history

51Number 71, December 2010
Ian J. McNiven et al.
Table 2 Presence/absence of vertebrate taxa, Square A, Epe Amoho.
XU Mean
dep
th b
elo
wsu
rface (
cm
)
Tele
ost
fish
Sh
ark
(te
eth
)
Turt
le
Sn
ak
e
Ag
am
idae
Vara
nu
s sp
.
Macro
po
did
ae
Sp
ilo
cu
scu
s sp
.
Ph
ala
ng
er
sp.
Pero
rycti
dae
Larg
e m
uri
d
Meg
ach
iro
pte
ra
Mic
roch
iro
pte
ra
Su
s sc
rofa
1 0.4 + + + + + +2 2.0 + + + + + + + + +
3A 3.9 + + + + + + + + + +3B 4.0 + + + + +4A 5.7 + + + + + + +4B 6.3 + + + + + + + + + +5A 7.8 + + + + + + + + +5B 9.0 + + + + + +5C 7.7 + +6A 10.7 + + + + + + + +6B 10.5 + + + +6C 11.4 +6D 11.6 + + + + + +7 11.8 + + + +8 13.0 + + + + + +
9A 14.2 + + + +9B 16.0 +
10A 17.1 + + +10B 15.2 +11A 19.6 +11B 17.9 + +12A 23.0 +12B 21.1 +12C 21.8 + +12D 22.313A 25.4 +13B 23.9 +13C 26.2 +14A 29.6 +14B 28.6 +14C 30.815A 33.3 + +15B 34.016A 37.016B 38.1 +17A 41.717B 41.518A 45.918B 46.519A 49.719B 50.1 +20A 53.8 + +20B 54.821A 57.921B 58.722A 61.5 +22B 63.123A 65.2 + +23B 66.124A 69.324B 71.425 74.0 + +26 78.1 +27 82.1 + +
28A 85.928B 85.329A 89.729B 88.530A 93.830B 93.531 96.9 +32 102.233 105.4

52 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
of pottery manufacture in the region; all known ceramics were
(directly or indirectly) imported through hiri and ancestral hiri
trade (Rhoads 1980, 1982a, 1983).
Glass Beads: Nine glass beads weighing 0.4g were recovered
from XUs 3a to 6d between 2 and 12cm below the surface
(Figure 10). All beads are white except for a red bead from XU6a.
Although glass beads dating back possibly to AD 1650–1750
were excavated by Rhoads (1980:A76-A80, 1984) from other
parts of Rumu territory, parsimony suggests most of the beads
from Epe Amoho date to the time of increased direct contact
with Europeans during the late nineteenth and/or twentieth
centuries. As with drilled dog teeth, glass beads are a known
body adornment amongst Papuan Gulf peoples (Specht and
Fields 1984:148-151).
Glass Sherds: Three fragments of glass were recovered from
XU3a (n=2) and XU22b (n=1) at a depth of 2–4cm and 59–63cm
(base of ‘pit’ feature) below the surface respectively. The XU3a
sherds have maximum lengths of 42mm and 8mm with mean
thicknesses of 2.3mm and 2.1mm respectively. The XU22b sherd
has a maximum length of 67mm and a mean thickness of 2.2mm.
All fragments are flat tabular pieces of clear glass. Although the
fragments do not conjoin, the similarity of colour, form and
thickness suggests that they may derive originally from a single
piece of glass. The form and colour of the glass suggests they date
to the time of European contact either in the late nineteenth and/
or twentieth centuries.
Shotgun Pellets: Nine shotgun pellets weighing 1.5g were
recovered from XUs 1 to 5a within the upper 8cm of deposit.
Weights and diameters indicate #4 and #2 or #3 shot. These
pellets clearly date to the time of European contact in either
the late nineteenth and/or twentieth centuries. Shotgun pellets
probably reflect use of shotguns for hunting local game by
rockshelter occupants; shotguns are sometimes used by clans-
people today while hunting. The pellets were probably discarded
after being removed from game brought back to the site during
food preparation and consumption.
Other Metal: Numerous small fragments of rusty metal,
a section of copper wire and a tiny copper/brass oval ring
with a total weight of 10.3g were obtained from XUs 3a to 6a.
The rusty metal items include a stud (XU6a) and a looped
section of metal that may be a key winder from a canned
beef tin (XU3a). All of these items probably date to the time
of European contact either in the late nineteenth and/or
twentieth centuries.
Plastic: An 8mm-long, 1.5mm-diameter, section of red
plastic electrical wire insulation was recovered from XU5b. The
item has been cut at both ends and shows considerable signs of
rodent gnawing. The function of the item is unknown but may
have been used as a body adornment.
Epe Amoho: Comparing Past and Present Site Use
Environmental ContextRhoads’ (1980) assumption of relatively stable local
environmental conditions for the past 3000 years is only borne
out in part by palaeoenvironmental analyses. Sediment, pollen
and plant macrofossil data indicate that the rainforest and
riverine environmental context of Epe Amoho underwent a
series of minor changes over the past 3000 years. For example,
pollen analysis reveals increasing rainforest disturbance through
time (most likely associated with gardening activity) and the
presence of freshwater tolerant mangroves within the past 500
years. This change in mangroves may have multiple causes,
including increased freshwater input (possibly from development
of new drainage regimes), increased sedimentation (possibly
elevating many lowland areas above saline influence) and/or
decreasing saline intrusions (possibly indicating a downstream
movement of saltwater penetration of the Kikori River due to
coastal progradation of the river delta) (see Crowley and Gagan
1995; Wang and Chappell 2001; Woodroffe et al. 1993). To what
extent decreasing sea-level over the past 3000 years, as argued
recently for northeast Australia (Lewis et al. 2008; Woodroffe
2006), influenced saltwater intrusions and coastal progradation
along the Gulf of Papua remains to be determined. Whatever the
case, coastal progradation would have resulted in Epe Amoho
becoming located increasingly inland from the coast. At this
stage, the only known possible impact of coastal progradation
on Epe Amoho is the absence of shark teeth in recent levels of
the site. This absence may reflect decreased local availability of
sharks associated with increasing freshwater conditions of the
past 500 years. In this connection, the Kamoro people of West
Papua associate decreased availability of sharks upstream on
Figure 9 Drilled dog tooth, Square A, Epe Amoho (Photograph: Ken Aplin).
Figure 10 Glass beads, Square A, Epe Amoho (Photograph: Steve Morton).

53Number 71, December 2010
Ian J. McNiven et al.
the Wania River with extensive coastal progradation at the river
mouth (Pickell 2002:106).
HuntingWallabies, bats, cuscuses and large murids and perhaps to a lesser
extent bandicoots and pigs were components of subsistence
activities at Epe Amoho. Aplin’s analysis of faunal remains from
Rhoads’ (1980) excavations at open village sites (Kulupuari
[Kikiniu hereafter], Mampaiu, Ibira, Bageima [Puriau hereafter]
and Waira) and rockshelter sites (Rupo, Ouloubomoto and
Herekuna) revealed a range of terrestrial vertebrate remains with
broad similarities to the Epe Amoho faunal assemblage. All of the
sites with sizable samples of bone included most of the following
suite of terrestrial vertebrates – pig, wallaby, tree kangaroo, cuscus,
possum, bandicoot, bat, rodent, dog and cassowary. Overall,
hunting patterns reported by Rhoads (1980) and the present
study reveal comprehensive hunting of most families of medium-
and large-sized mammals from local rainforest habitats. As such,
both archaeological and ethnographic data on hunting strategies
show close agreement. The only major exception is cassowary
which was not excavated at Epe Amoho. While Rhoads found
evidence of mammal hunting back to 2000 years ago at Rupo
(dating based on David 2008) and 1500 years ago for cassowary
at Kikiniu (chronology confirmed by David 2008), at Epe Amoho
evidence for hunting of mammals and cassowary was restricted
to the past 500 years. As such, the temporal depth of broad-scale
ethnographically-known hunting practices differs between sites.
FishingThe occurrence of at least eight fish taxa at Epe Amoho compares
well with the diversity of freshwater fish recorded at sites excavated
by Rhoads (1980). In both cases, fish assemblages are dominated
by catfish (Ariidae) which matches well with ethnographically-
documented Rumu fishing practices. However, whereas Rhoads
(1980) recovered fish bones back to 1500 years ago at Kikiniu and
2000 years ago at Rupo, fish bones at Epe Amoho extend back
to 2500–2850 years ago. This broader temporal depth includes
sharks which appear to drop out of Epe Amoho deposits within
the past 300 years. As with hunting practices, the temporal depth
of broad-scale ethnographically-known fishing practices differs
between sites.
ShellfishingFreshwater shellfish from Epe Amoho match evidence for
shellfishing documented at other excavated sites in the mid-
Kikori area. Rhoads (1980) found that the bivalve Batissa violacea
and the gastropod Neritina sp. dominate open village sites,
while the range of shellfish at rockshelter sites was wider and
included sizable representations of the gastropods Melanoides sp.
and Viviparidae. While shellfish at Epe Amoho are temporally
restricted to the past 900–1200 years, Rhoads (1980:Table VIII-
9) recovered shellfish throughout the 2000 year-long Rupo
rockshelter sequence. Thus, the antiquity of shellfishing practices
as documented ethnographically varies from site to site.
Plant GatheringBamboo is the most consistent macrofossil plant type recovered
from Epe Amoho. This consistency suggests that use of bamboo
(e.g. cooking utensils as recorded ethnographically within the
rockshelter), is a local tradition extending back to at least 2500–
2850 years ago. Macrofossil plant remains were not discussed
by Rhoads (1980). The absence of Celtis seeds in pre-500 cal BP
levels of the site is most likely indicative of a change in economic
activity as pollen analyses indicate the presence of Celtis in the
region for the past 2400–2850 years and its absence is thus not
explained by vegetation change.
Specialised TechnologyNo plant food processing implements (e.g. sago pounders)
were recovered from Epe Amoho. Rhoads (1980:229) similarly
found that ‘artefacts representing the processing of palm starch
are not present in any rockshelter deposits’ but ‘sago choppers’
were excavated from the village sites of Kikiniu, Mampaiu,
Puriau and Waira. This pattern is consistent with ethnographic
data indicating that food processing technology at rockshelter
sites focused on hunting and collection of fauna. In this
connection, use-wear and residue analyses of stone artefacts
from Epe Amoho indicate wood-working, a ubiquitous activity
in hunting camps (e.g. tool manufacture and maintenance).
The restriction of stone artefacts to the past 500 years at Epe
Amoho matches the temporal depth for hunting of terrestrial
vertebrates at the site.
All stone artefacts excavated from Epe Amoho are exotic
to the region and probably reveal importation through
ethnographically-known exchange relationships with peoples
further inland (see Busse et al. 1993; Rhoads 1980, 1982a, 1983;
Rhoads and MacKenzie 1991). However, such inter-regional
exchange systems extend well beyond the restricted timeframe of
500 years ago at Epe Amoho as Rhoads (1980, 1982a) excavated
exotic stone artefacts dating back to at least 1500 years ago at
Kikiniu. Recently, David et al. (2010:51) report that use of
‘imported, curated chert artefacts’ increases ‘noticeably’ at Emo
after 1530 years ago, ‘suggesting heightened levels of trade with
stone tool manufacturers in the highlands foothills to the north’.
SeasonalityArchaeological data are available to discern seasonal patterns of
occupation at Epe Amoho and at the sites previously excavated
by Rhoads (1980). Shellfish indicate dry season occupation of
Epe Amoho, with relatively high numbers of B. violacea and
a near-total absence of Neritina sp. in deposits. B. violacea is
predominantly a dry season shellfish (because it is said by Rumu
clan members that this species does not have much flesh on it
during the wet season), while Neritina is a wet season species
recovered mostly from year-round occupied open village sites. In
addition, fish, turtle and crustacean remains suggest harvesting
from low water level sources (creeks and ponds) during the
dry season. While shark fishing is poorly documented amongst
the Rumu, people of the lower Kikori River delta catch sharks
(various species, mainly Carcharhinus spp.) using ‘seine fences’
across small creeks only during the dry season (Frusher and
Subam 1984:82). Similarly, megapode eggs are collected by
Himaiyu clans-people today only during the dry season. Dry
season use of Epe Amoho is consistent with local hydrology,
as much of the site is wet to moist and an unattractive place
for camping during the wet season. Sediment particle size
analysis indicates periodic flooding of the site during the wet
season. The long-term history of Epe Amoho as a dry season

54 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
camp is consistent with the ethnographic record of this site as
the Himaiyu clan’s most important dry season hunting camp.
Rhoads (1980:232, 235) concluded also that rockshelters were
inhabited during the dry season with minimal use during the
wet season.
Regional Chronology: Pulsed Occupation and HiatusesHow does the pulsed occupation of Epe Amoho – low intensity
occupation during the Early (2500-2850 cal BP) and Middle
(900-1200 cal BP) Phases followed by more intensive occupation
during the Recent Phase (the past 500 years) – compare with
regional chronological trends for the Kikori River basin and the
broader Papuan Gulf region? David (2008) provides a major
overview and reassessment of chronological trends for the Kikori
River based on 99 radiocarbon dates from 16 sites (including
the 12 radiocarbon dates from Epe Amoho). It is clear from this
overview that the pulsed occupation of Epe Amoho is related to
a series of ‘major’ and ‘minor’ pulses of occupation across the
Kikori region over the past 3000 years. The two minor pulses
occur at approximately 2800 cal BP (‘minor pulse 1’) and 2000
cal BP (‘minor pulse 2’), while the two major pulses occur at
approximately 950–1450 cal BP (‘early major pulse’) and 0–500
cal BP (‘late major pulse’). ‘Minor pulse 1’ is represented by initial
occupation of two sites – Early Phase occupation of Epe Amoho
and a basal occupation date from a nearby cave site (KG141).
‘Minor pulse 2’ is not represented in the Epe Amoho excavations.
The ‘early major pulse’ includes Middle Phase occupation of
Epe Amoho while the ‘late major pulse’ corresponds to Recent
Phase occupation of Epe Amoho. Recent excavation of sites
in the Aird Hills, Emo in particular, has revealed radiocarbon
determinations during the hiatus between the ‘early major pulse’
and the ‘minor pulse 2’ (David et al. 2010), supporting in part
Rhoads’ (1982a:143) view that the radiocarbon age pulses for
the Kikori River region relate to changes in settlement systems
rather than to periods of human presence separated by regional
abandonment. General correspondence in the pulsed occupation
of Epe Amoho and the broader Kikori region suggests that
cultural and historical processes influencing occupation and
use of Epe Amoho were part of cultural and historical processes
operating across the Kikori region over the past 3000 years. This
correspondence extends in part to the broader Papuan Gulf
region where intensive occupation of Epe Amoho over the past
500 years, including use of imported chert and pottery through
regional trade networks, occurred concurrently with intensified
village use and the development of large-scale trade systems
known ethnographically as the hiri across the Papuan Gulf over
the past 400–500 years (see Frankel and Rhoads 1994; Rhoads
1982a; Rhoads and Mackenzie 1991).
ConclusionEpe Amoho provides a rare case study of the archaeology of a
rockshelter that continues to be used today. Both ethnographic
and archaeological evidence supports the view that Epe Amoho
has been a temporary camp associated with exploitation of local
rainforest resources over the past 2500–2850 years. However,
while evidence for fishing and use of bamboo utensils occurs
throughout the site’s history, exploitation of terrestrial faunal
resources and use of stone tools is restricted mostly to the Recent
Phase of the past 500 years. The fact that fragile fish bones were
recovered from the Early and Middle Phases of the site dating to
between 2500–2850 and 900–1200 cal BP respectively suggests
the paucity of terrestrial bone in these lower levels is not due to
taphonomic factors. Over the past 2500–2850 years, Epe Amoho
appears to have witnessed a broadening of subsistence strategies
such that the status of the site changed from a specialised fishing
camp between 900–1200 and 2500–2850 cal BP to a fishing and
hunting camp of ethnographic form during the past 500 years.
While these developments took place in the context of local
environmental changes, further research is required to determine
the degree to which such changes influenced settlement-
subsistence arrangements. At this point, pollen analysis suggests
that increasing settlement intensity of the region over the past
1500 years is matched by increasing rainforest disturbance
(probably associated with increased gardening and exploitation
of sago palms).
It would be misleading to conclude that because ethnographic
use of Epe Amoho emerged within the past 500 years that use of
ethnographic analogies for Rumu sites have a similar restricted
temporal relevance. Fishing and bamboo use at Epe Amoho
indicates that certain key elements of the Rumu economy
have historical continuity extending back 2500–2850 years.
Furthermore, excavations by Rhoads (1980) reveal that Rumu
hunting practices extend back at least 2000 years and importation
of exotic stone for artefacts extends back at least 1500 years ago.
Clearly, different Rumu sites provide different insights into Rumu
history and the temporal depth of ethnographically-known
practices. While excavations at Epe Amoho reveal that certain
Rumu subsistence practices such as freshwater fishing extend
back earlier than that recorded by Rhoads, this study concurs
with Rhoads’ research that the best match between Rumu
settlement-subsistence practices documented ethnographically
and archaeologically date to the past 400–500 years. Furthermore,
this study also supports Rhoads’ contention that settlement
patterns were more dynamic than subsistence practices through
time with Epe Amoho showing pulsed occupation over the past
3000 years. For reasons not currently understood, Epe Amoho
shows no signs of occupation 1200–2400 and 500–900 years ago,
presumably when activities shifted to other sites in the region. As
ethnographic practices are expressed in a punctuated, patchy and
mostly cumulative fashion across the region, it is apparent that
no simple temporal threshold exists for the reliable application
of ethnographic analogies in the region. A more complex
and nuanced picture emerges where Rumu cultural practices
unfolded at differing points in time at different sites over a
period of at least 3000 years.
Supplementary InformationSupplementary information for this article is available online at
www.australianarchaeologicalassociation.com.au.
AcknowledgementsOur research was made possible by the hospitality of the
Rumu people and members of the Himaiyu clan in particular.
We would especially like to thank Max Pivoru and his brother
William Pivoru for introducing us to the world of Epe Amoho
and for permission to excavate in their backyard. Excavations at
Epe Amoho were made possible by the hard work and assistance

55Number 71, December 2010
Ian J. McNiven et al.
of Lisa Araho, Jeremy Ash, Alice Bedingfield, Thomas Kokents,
David Kupere (Jr), Hale Lahui, Paul Opai, Aubau Tami and
Duncan Wright. Subsequent interviews were also undertaken
at Kopi village with Rumu clans-people, in particular Kupere
Karamavi, Margaret Murepe, Max and Grace Pivoru, Frank
Pivoru and Sam and Hilda Kaiwari. Lynden McGregor undertook
survey mapping of the site. Kara Rasmanis and Toby Wood
(School of Geography and Environmental Science, Monash
University) kindly drafted the figures. Noel Kemp thanks Peter
Last and Al Graham and CSIRO (Hobart) for allowing access
to shark tooth collections. We thank also Carla Eisemberg and
Robert and Deborah Petterson for permission to cite their
unpublished manuscripts. This project was funded in part
by an ARC Discovery grant and QEII Fellowship DP0877782.
Helpful comments on earlier drafts of this paper were kindly
provided by Peter White, Jim O’Connell, Nicky Horsfall and
anonymous referees.
ReferencesAllen, G.R., A.W. Storey and M. Yarrao 2008 Freshwater Fishes of the Fly River, Papua
New Guinea. Tabubil: Ok Tedi Mining.
Asmussen, B. 2009 Intention or incidental thermal alteration? Analysing site
occupation through burnt bone. Journal of Archaeological Science 36:528-536.
Bennett, K. and K. Willis 2001 Pollen. In J.P. Smol, H.J.B. Birks and W.M. Last (eds),
Tracking Environmental Change Using Lake Sediments: Volume 3: Terrestrial,
Algal and Siliceous Indicators, pp.5-32. Dordrecht, The Netherlands: Kluwer
Academic Publishers.
Busse, M., S. Turner and N. Araho 1993 The People of Lake Kutubu and Kikori:
Changing Meanings of Daily Life. Boroko: Trustees of the Papua New Guinea
National Museum and Art Gallery.
Buzon, M.R., J.T. Eng, P.M. Lambert and P.L. Walker 2005 Bioarchaeological
methods. In H.D.G. Maschner and C. Chippindale (eds), Handbook of
Archaeological Methods, pp.871-918. Vol. 2. Lanham: AltaMira Press.
Cowie, R.H. 1998 Catalog of the non-marine snails and slugs of the Samoan Islands.
Bishop Museum Bulletin in Zoology 3:1-122.
Cragg, S. 1983 The mangrove ecosystem of the Purari delta. In T. Petr (ed.), The
Purari: Tropical Environment of a High Rainfall River Basin, pp.295-324.
Monographiae Biologicae 51. The Hague: Dr W. Junk Publishers.
Crowley, G.M. and M.K. Gagan 1995 Holocene evolution of coastal wetlands in wet-
tropical northeastern Australia. The Holocene 5(4):385-399.
David, B. 2008 Rethinking cultural chronologies and past landscape engagement
in the Kopi region, Gulf Province, Papua New Guinea. The Holocene
18(3):463-479.
David, B., A. Fairbairn, K. Aplin, L. Murepe, M. Green, J. Stanisic, M. Weisler, D.
Simala, T. Kokents, J. Dop and J. Muke 2007 OJP, a terminal Pleistocene
archaeological site from the Gulf Province lowlands, Papua New Guinea.
Archaeology in Oceania 42(1):31-33.
David, B., J-M. Geneste, K. Aplin, J-J. Delannoy, N. Araho, C. Clarkson, K. Connell,
S. Haberle, B. Barker, L. Lamb, J. Stanisic, A. Fairbairn, R. Skelly and C. Rowe
2010 The Emo Site (OAC), Gulf Province, Papua New Guinea: Resolving
long-standing questions of antiquity and implications for the history of the
ancestral hiri maritime trade. Australian Archaeology 70:39-54.
David, B., I.J. McNiven, V. Attenbrow, J. Flood and J. Collins 1994 Of Lightning
Brothers and white cockatoos: Dating the antiquity of signifying systems in
the Northern Territory, Australia. Antiquity 68: 241-251.
David, B., I.J. McNiven, T. Denham and J. Dop 2005 Cultural Heritage Sites Survey
along the Kaiam-Goaribari Section of the Papua New Guinea Gas Pipeline
Route. Monash University Cultural Heritage Report Series 14. Clayton, VIC:
School of Geography and Environmental Science, Monash University.
David, B., M. Pivoru, W. Pivoru, M. Green, B. Barker, J.F. Weiner, D. Simala, T. Kokents,
L. Araho and J. Dop. 2008 Living landscapes of the dead: Archaeology of the
afterworld among the Rumu of Papua New Guinea. In B. David and J. Thomas
(eds), Handbook of Landscape Archaeology, pp.158-166. Walnut Creek, CA:
Left Coast Press.
Denham, T., R. Fullagar and L. Head 2009 Plant exploitation on Sahul: From
colonisation to the emergence of regional specialisation during the Holocene.
Quaternary International 202:29-40.
Dutton, T. (ed.) 1982 The Hiri in History: Further Aspects of Long Distance Motu
Trade in Central Papua. Pacific Research Monograph 8. Canberra: Australian
National University.
Enesar Consulting 2005 PNG Gas Project. Environmental Impact Statement
Volume 1. Main Report Text. Report prepared by Enesar Consulting Pty Ltd
for Esso Highlands Ltd.
Frankel, D. and J. Rhoads (eds) 1994 Archaeology of a Coastal Exchange System:
Sites and Ceramics of the Papuan Gulf. Research Papers in Archaeology and
Natural History 25. Canberra: Division of Archaeology and Natural History,
Research School of Pacific and Asian Studies, Australian National University.
Frusher, S.D. and S. Subam 1984 Traditional fishing methods and practices in
the northern Gulf of Papua. In N.J. Quinn, B. Kojis and P.R. Warpeha (eds),
Subsistence Fishing Practices of Papua New Guinea, pp.80-87. Lae: Appropriate
Technology Development Institute, Liklik Buk Information Centre.
Galanidou, N. 2000 Patterns in caves: Foragers, horticulturalists, and the use of
space. Journal of Anthropological Archaeology 19(3):243-275.
Georges, A., E. Alacs, M. Pauza, F. Kinginapi, A. Ona and C. Eisemberg 2008
Freshwater turtles of the Kikori Drainage, Papua New Guinea, with special
reference to the pig-nosed turtle, Carettochelys insculpta. Wildlife Research
35(7):700-711.
Georges, A., C. Eisemberg and L. Bauer n.d. In search of Keheko. Unpublished
manuscript in author’s possession.
Goreki, P.P. 1991 Horticulturalists as hunter-gatherers: Rockshelter usage in Papua
New Guinea. In C.S. Gamble and W.A. Boismier (eds), Ethnoarchaeological
Approaches to Mobile Campsites, pp.237-262. Ann Arbor: International
Monographs in Prehistory.
Gould, R.A. 1980 Living Archaeology. Cambridge: Cambridge University Press.
Gould, R.A. and P.J. Watson 1982 A dialogue on the meaning and use of analogy in
ethnoarchaeological reasoning. Journal of Anthropological Archaeology 1:355-
381.
Haines, A.K. 1979 An Ecological Survey of Fish of the Lower Purari River System,
Papua New Guinea. Department of Minerals and Energy, Papua New Guinea,
Purari River Hydroelectric Scheme Environmental Studies 6.
Hale, H.M. and N.B. Tindale 1930 Notes on some human remains in the lower
Murray River valley, South Australia. Records of the South Australian Museum
4(2):145-218.
Hardy, K. and P. Sillitoe 2003 Material perspectives: Stone tool use and material
culture in Papua New Guinea. Internet Archaeology 14.
Hiscock, P. 2008 Archaeology of Ancient Australia. London: Routledge.
Hope, P. 1979 Long Ago is Far Away: Accounts of the Early Exploration and Settlement
of the Papuan Gulf Area. Canberra: Australian National University Press.
Knauft, B.M. 1993 South Coast New Guinea Cultures. Cambridge: Cambridge
University Press.
Kusmer, K.D. 1990 Taphonomy of owl pellet deposition. Journal of Paleontology
64(4):629-637.
Levitt, D. 1981 Plants and People: Aboriginal Uses of Plants on Groote Eylandt.
Canberra: Australian Institute of Aboriginal Studies.
Lewis, S.E., R.A.J. Wüst, J.M. Webster and G.A. Shields 2008 Mid-late Holocene
sea-level variability in eastern Australia. Terra Nova 20(1):74-81.
Liese, W. 2002 The Anatomy of Bamboo Culms. New York: International Network
for Bamboo & Rattan.

56 Number 71, December 2010
Historicising the Present: Late Holocene Emergence of a Rainforest Hunting Camp, Gulf Province, Papua New Guinea
Lilley. I. (ed.) 2006 Archaeology of Oceania: Australia and the Pacific Islands.
Malden, MA: Blackwell.
Löffler, E. 1974 Explanatory Notes to the Geomorphological Map of Papua New
Guinea. Melbourne: CSIRO.
Löffler, E. 1977 Geomorphology of Papua New Guinea. Canberra: CSIRO and ANU
Press.
Lourandos, H. 1997 Continent of Hunter-Gatherers. Cambridge: Cambridge
University Press.
Lubinski, P.M. 1996 Fish heads, fish heads: An experiment of differential bone
preservation in a salmonid fish. Journal of Archaeological Science 23:175-
181.
Lyman, R.L. and M.J. O’Brien 2001 The direct historical approach, analogical
reasoning, and theory in Americanist archaeology. Journal of Archaeological
Method and Theory 8(4):303-342.
Marshall, B. 1986 An experimental evaluation of the criteria used to distinguish
owl-deposited bone in archaeological cave deposits in Australia. Australian
Archaeology 22:104-121.
McNiven, I.J., B. David, Goemulgau Kod and J. Fitzpatrick 2009 The great kod of
Pulu: Mutual historical emergence of ceremonial sites and social groups in
Torres Strait, NE Australia. Cambridge Archaeological Journal 19(3):291-317.
Meehan, B. and R. Jones (eds) 1988 Archaeology with Ethnography: An Australian
Perspective. Canberra: Department of Prehistory, Research School of Pacific
Studies, Australian National University.
Mulvaney, D.J. 1969 The Prehistory of Australia. New York: Frederick A. Praeger.
Nicholson, A. and S. Cane 1991 Desert camps: Analysis of Australian Aboriginal
proto-historic campsites. In C.S. Gamble and W.A. Boismier (eds),
Ethnoarchaeological Approaches to Mobile Campsites, pp.263-354. Ann Arbor:
International Monographs in Prehistory.
Nix, H.A. and J.D. Kalma 1972 Climate as a dominant control in the biogeography
of northern Australia and New Guinea. In D. Walker (ed.), Bridge and Barrier:
A Natural and Cultural History of the Torres Strait, pp.61-91. Canberra:
Department of Biogeography and Geomorphology, Research School of Pacific
Studies, Australian National University.
Paijmans, K. 1983 The vegetation of the Purari catchment. In T. Petr (ed.), The
Purari: Tropical Environment of a High Rainfall River Basin, pp.227-252.
Monographiae Biologicae 51. The Hague: Dr W. Junk Publishers.
Pannell, S. and S. O’Connor 2005 Toward a cultural topography of cave use in east
Timor: A preliminary study. Asian Perspectives 44(1):193-206.
Petr, Y. 1983 Limnology of the Purari basin. In T. Petr (ed.), The Purari: Tropical
Environment of a High Rainfall River Basin, pp.179-203. Monographiae
Biologicae 51. The Hague: Dr W. Junk Publishers.
Pétrequin, A.-M. and P. Pétrequin 2006 Objets de Pouvoir en Nouvelle-Guinée:
Approche Ethnoarchéologique d’un Système de Signe Sociaux: Catalogue de
la donation Anne-Marie et Pierre Pétrequin. Paris: Musée d’Archéologie
Nationale de Saint-Germain-en-Laye.
Petterson, R. and D. Petterson 2008 Rumu Ethnography. Unpublished report in
authors’ possession.
Pickell, D. 2002 Between the Tides: A Fascinating Journey among the Kamoro of New
Guinea. Hong Kong: Periplus.
Powell, J.M. 1976 Ethnobotany. In K. Paijmans (ed.), New Guinea Vegetation,
pp.106-183. Canberra: Australian National University Press.
Reimer, P.J., M.G.L. Baillie, E. Bard, A. Bayliss, J.W. Beck, P.G. Blackwell, C. Bronk
Ramsey, C.E. Buck, G.S. Burr, R.L. Edwards, M. Friedrich, P.M. Grootes, T.P.
Guilderson, I. Hajdas, T.J. Heaton, A.G. Hogg, K.A. Hughen, K.F. Kaiser, B. Kromer,
F.G. McCormac, S.W. Manning, R.W. Reimer, D.A. Richards, J.R. Southon, S.
Talamo, C.S.M. Turney, J. van der Plicht and C.E. Weyhenmeyer 2009 IntCal09
and Marine09 radiocarbon age calibration curves, 0-50,000 years cal BP.
Radiocarbon 51(4):1111-1150.
Rhoads, J.W. 1980 Through a Glass Darkly: Present and Past Land-Use Systems
of Papuan Sagopalm Users. Unpublished PhD thesis, Australian National
University, Canberra.
Rhoads, J.W. 1982a Prehistoric Papuan exchange systems: The hiri and its
antecedents. In T. Dutton (ed.), The Hiri in History: Further Aspects of
Long Distance Motu Trade in Central Papua, pp.131-151. Pacific Research
Monograph 8. Canberra: Australian National University.
Rhoads, J.W. 1982b Sagopalm management in Melanesia: An alternative
perspective. Archaeology in Oceania 17(1):20-27.
Rhoads, J.W. 1983 Prehistoric sites from the Kikori region, Papua New Guinea.
Australian Archaeology 16:96-114.
Rhoads, J.W. 1984 Pre-contact glass artefacts from Papua New Guinea. South-East
Asian Studies Newsletter 14:1-5.
Rhoads, J.W. and D.E. Mackenzie 1991 Stone axe trade in prehistoric Papua: The
travels of python. Proceedings of the Prehistoric Society 57:35-49.
Roberts, T.R. 1978 An Ichthyological Survey of the Fly River in Papua New Guinea
with Descriptions of New Species. Smithsonian Contributions to Zoology 281.
Robertson, A.I., P.A. Daniel and P. Dixon 1991 Mangrove forest structure and
productivity in the Fly River estuary, Papua New Guinea. Marine Biology
111:147-155.
Solem, A. 1989 Non-camaenid land snails of the Kimberley and Northern Territory.
Invertebrate Taxonomy 2:455-604.
Specht, J. and J. Fields 1984 Frank Hurley in Papua: Photographs of the 1920-1923
Expeditions. Bathurst: Robert Brown and Associates in association with the
Australian Museum Trust.
Spriggs, M. 2008 Ethnographic parallels and the denial of history. World
Archaeology 40(4):538-552.
Stiner, M.C., S.L. Kuhn, S. Weiner and O. Bar-Yosef 1995 Differentiating burning,
recrystallization, and fragmentation of archaeological bone. Journal of
Archaeological Science 22:223-237.
Stuiver, M. and P.J. Reimer 1993 Extended 14C database and revised CALIB
radiocarbon calibration program. Radiocarbon 35:215-230.
Walter, A. and V. Lebot 2007 Gardens of Oceania. ACIAR Monograph 122.
Wang, P. and J. Chappell 2001 Foraminifera as Holocene environmental indicators
in the South Alligator River, northern Australia. Quaternary International 83-
85:47-62.
Watson, V.D. 1995 Simple and significant: Stone tool production in Highland New
Guinea. Lithic Technology 20(2):89-99.
White, J.P. and J.F. O’Connell 1982 A Prehistory of Australia, New Guinea and Sahul.
Sydney: Academic Press.
Williams, F.E. 1924 The Natives of the Purari Delta. Territory of Papua, Anthropology
Report 5. Port Moresby: Government Printer.
Wobst, H.M. 1978 The archaeo-ethnography of hunter-gatherers or the tyranny of
the ethnographic record in archaeology. American Antiquity 43:303-309.
Woodroffe, C.D., M.E. Mulrennan and J. Chappell 1993 Estuarine infill and coastal
progradation, southern Van Diemen Gulf, northern Australia. Sedimentary
Geology 83:257-275.
Woodroffe, S.A. 2006 Holocene Relative Sea-Level Changes in Cleveland Bay, North
Queensland, Australia. Unpublished PhD thesis, Department of Geography,
Durham University.
Young, M.W. and J. Clark 2001 An Anthropologist in Papua: The Photography of F.E.
Williams, 1922-39. Adelaide: Crawford House Publishing.
Related Documents











