Fisheries Research 99 (2009) 7–16 Contents lists available at ScienceDirect Fisheries Research journal homepage: www.elsevier.com/locate/fishres Historical analysis of Cuban commercial fishing effort and the effects of management interventions on important reef fishes from 1960–2005 Rodolfo Claro a , Yvonne Sadovy de Mitcheson b,∗ , Kenyon C. Lindeman c , Alida R. García-Cagide a a Instituto de Oceanología, Cuba b University of Hong Kong, Hong Kong and Society for the Conservation of Reef Fish Aggregations, Hong Kong c Florida Institute of Technology, United States article info Article history: Received 29 September 2008 Received in revised form 10 April 2009 Accepted 14 April 2009 Keywords: Spawning aggregation Reef fish Snappers Groupers Management Cuba abstract Long-term fishery datasets can provide valuable insights into fishing histories, and represent a base- line against which to examine current status and plan for future management. For coral reef-associated fisheries, however, such datasets are extremely rare. We analyse a unique 45-year set of landings data on six reef fishes of commercial importance in Cuba’s coastal fisheries, together with information on management interventions, and examine the likely implications of over-fishing and management on the long-term condition of one grouper (Serranidae) and five snapper (Lutjanidae) species. The data clearly demonstrate differential responses to fishing and management according to the biology of the different species examined. In particular, those species that aggregate most predictably, and at a small number of spawning sites, are more prone to uncontrolled fishing and in need of effective management. The increase in the number and severity of management measures over the last 20 years reflects attempts to address declines in the fishery following its development prior to the 1980s. The data suggest short-lived successes, and challenges to successful management from changing socio-economic circumstances and environmental impacts. The importance of considering, in addition to landings information, social and economic factors, management history, the biology of targeted species and the implications of environ- mental factors in relation to resource use, trends, and management is clearly illustrated in this study. Managers should strive to document species-specific data over the long-term and aim for robust and adaptive fishery management that constantly adjusts to the range of factors that affect fishing activity, exploited species, and responses to management. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Within the tropics, our understanding of the history of coastal fisheries and the impacts of management interventions is seri- ously undermined by an almost complete absence of the type of long-term species-specific data available for many temperate fish- eries. One of the very few exceptions is the detailed multi-decadal database on the coastal fishery of Cuba, a government-owned and managed fishing industry. Because of natural variability in fish pop- ulations over time and how this might interact with management actions, especially in the case of multi-species fisheries, long-term fishery data are particularly for providing important insights into the actual or probable effects of management and other biotic and abiotic factors on landings. The lessons learned from such informa- tion provide valuable guidance for future action that can be framed in more precautionary and practical terms than is possible in the absence of such a fishery history. The coastal fishery of Cuba repre- ∗ Corresponding author. Tel.: +852 2299 0603; fax: +852 2517 6082. E-mail address: [email protected] (Y.S.d. Mitcheson). sents a unique long-term record of a tropical, multi-species, fishery that promises to teach much about the interplay between fish- ery management and other social factors, and the biology of key exploited species. Traditionally, the catches of reef-associated finfishes in Cuba have shown strong seasonal trends, mainly associated with the reproductive periods of the most economically important species, among them snappers (Lutjanidae) and groupers (Serranidae). This seasonality in catch trends is determined by the increased vulner- ability of aggregating species to fishing during the reproductive period and the resulting focus of fishing activity on spawning individuals which yield a large proportion of annual catches of such species (Baisre, 1985, 2000, 2004; Claro et al., 2001). Since the responses of aggregating species to fishing vary according to the biology of different targeted species, an understanding of the impacts of fishing and the consequences of management can only be understood by species-specific analyses in the context of the coastal fishery and its management history as a whole. The history of the coastal, reef-associated, fishery of Cuba is one of increasing and decreasing fishing pressure and variable man- agement effectiveness that ultimately led to substantial declines 0165-7836/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2009.04.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hm
Ra
b
c
a
ARRA
KSRSGMC
1
fioledmuafitatia
0d
Fisheries Research 99 (2009) 7–16
Contents lists available at ScienceDirect
Fisheries Research
journa l homepage: www.e lsev ier .com/ locate / f i shres
istorical analysis of Cuban commercial fishing effort and the effects ofanagement interventions on important reef fishes from 1960–2005
odolfo Claroa, Yvonne Sadovy de Mitchesonb,∗, Kenyon C. Lindemanc, Alida R. García-Cagidea
Instituto de Oceanología, CubaUniversity of Hong Kong, Hong Kong and Society for the Conservation of Reef Fish Aggregations, Hong KongFlorida Institute of Technology, United States
r t i c l e i n f o
rticle history:eceived 29 September 2008eceived in revised form 10 April 2009ccepted 14 April 2009
eywords:pawning aggregationeef fishnappersroupersanagement
uba
a b s t r a c t
Long-term fishery datasets can provide valuable insights into fishing histories, and represent a base-line against which to examine current status and plan for future management. For coral reef-associatedfisheries, however, such datasets are extremely rare. We analyse a unique 45-year set of landings dataon six reef fishes of commercial importance in Cuba’s coastal fisheries, together with information onmanagement interventions, and examine the likely implications of over-fishing and management on thelong-term condition of one grouper (Serranidae) and five snapper (Lutjanidae) species. The data clearlydemonstrate differential responses to fishing and management according to the biology of the differentspecies examined. In particular, those species that aggregate most predictably, and at a small numberof spawning sites, are more prone to uncontrolled fishing and in need of effective management. Theincrease in the number and severity of management measures over the last 20 years reflects attempts toaddress declines in the fishery following its development prior to the 1980s. The data suggest short-lived
successes, and challenges to successful management from changing socio-economic circumstances andenvironmental impacts. The importance of considering, in addition to landings information, social andeconomic factors, management history, the biology of targeted species and the implications of environ-mental factors in relation to resource use, trends, and management is clearly illustrated in this study.Managers should strive to document species-specific data over the long-term and aim for robust andentpons
adaptive fishery managemexploited species, and res
. Introduction
Within the tropics, our understanding of the history of coastalsheries and the impacts of management interventions is seri-usly undermined by an almost complete absence of the type ofong-term species-specific data available for many temperate fish-ries. One of the very few exceptions is the detailed multi-decadalatabase on the coastal fishery of Cuba, a government-owned andanaged fishing industry. Because of natural variability in fish pop-
lations over time and how this might interact with managementctions, especially in the case of multi-species fisheries, long-termshery data are particularly for providing important insights intohe actual or probable effects of management and other biotic and
biotic factors on landings. The lessons learned from such informa-ion provide valuable guidance for future action that can be framedn more precautionary and practical terms than is possible in thebsence of such a fishery history. The coastal fishery of Cuba repre-∗ Corresponding author. Tel.: +852 2299 0603; fax: +852 2517 6082.E-mail address: [email protected] (Y.S.d. Mitcheson).
165-7836/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2009.04.004
that constantly adjusts to the range of factors that affect fishing activity,es to management.
© 2009 Elsevier B.V. All rights reserved.
sents a unique long-term record of a tropical, multi-species, fisherythat promises to teach much about the interplay between fish-ery management and other social factors, and the biology of keyexploited species.
Traditionally, the catches of reef-associated finfishes in Cubahave shown strong seasonal trends, mainly associated with thereproductive periods of the most economically important species,among them snappers (Lutjanidae) and groupers (Serranidae). Thisseasonality in catch trends is determined by the increased vulner-ability of aggregating species to fishing during the reproductiveperiod and the resulting focus of fishing activity on spawningindividuals which yield a large proportion of annual catches ofsuch species (Baisre, 1985, 2000, 2004; Claro et al., 2001). Sincethe responses of aggregating species to fishing vary according tothe biology of different targeted species, an understanding of theimpacts of fishing and the consequences of management can only
be understood by species-specific analyses in the context of thecoastal fishery and its management history as a whole.The history of the coastal, reef-associated, fishery of Cuba is oneof increasing and decreasing fishing pressure and variable man-agement effectiveness that ultimately led to substantial declines

8 es Res
ioewsufds
oisfiiltwfcser
ampfitahbeutaoaiet(mia
chldeosfizp
bdbf((sia
R. Claro et al. / Fisheri
n most key commercial species. Until 1955, commercial catchesf fishes and invertebrates on the Cuban coastal platform did notxceed 10,000 tonnes annually, but, by the 1970s several speciesere over-fished, despite the introduction of management mea-
ures (Baisre, 1985; Baisre and Páez, 1981; Claro et al., 2001). Upntil the 1960s, catches increased to 21,000 tonnes (18,000 tonnesrom national waters, the remainder from the Gulf of Mexico) andemand for fish was relatively low with a focus on high ‘quality’pecies and larger sizes of fish (Baisre, 2004).
Subsequently, demand for seafood increased due to shortages inther food products as a result of trade embargoes, resulting in thentroduction of large-scale fishing gears, such as bottom trawls, longet nets and small-meshed fish traps. Until the mid 1960s the coastalsheries in Cuba were considered under-exploited, but an increase
n fishing effort, fueled by the introduction of motorized boats andarge-scale fishing gears and a relaxation of fishing regulations ledo over-fishing of several species by the 1970s. Particularly affectedere the lane snapper (Lutjanus synagris) in the main fishing area
or this species, the Gulf of Batabano, mullets (Mugilidae) in theoastal lagoons of the southeast, the Nassau grouper (Epinephelustriatus) in almost all areas, the queen conch (Strombus gigas) in sev-ral regions, and shrimp in Ensenada of the Broa and the southeastegion (Claro et al., 2001).
To address over-fishing an administrative process was initi-ted in the 1980s to reduce commercial fishing effort on theost intensively exploited resources, although overall fishing
ressure was not reduced until 2000. To reduce commercialshing effort in the 1980s, some of the effort was redirectedowards hitherto under-fished resources, generally of smaller valuend quality, such as stingrays, mojarras, porgies, Atlantic threaderring (Opisthonema oglinum), blackedge cusk-eel (Lepophidiumrevibarbe), clams, swimming crabs and others (Baisre, 2000; Clarot al., 2001). Regulations were introduced to protect some fish pop-lations, including closures, limits to fishing effort and quotas, andhese are likely to have favoured the persistence of the spawningggregations of the most important species (snappers). Althoughverall fishing effort decreased in terms of number of fishing boatsnd fishing gears, the organization and efficiency of the fisheryncreased due to several factors: (1) catch reception points werestablished at sea, close to fishing grounds, to reduce navigationime (and fuel), so that fishermen could spend more time fishing;2) administrative measures were established to encourage fisher-
en to work 20 days every month (some fishermen do not workf they have sufficient money); and (3) economic incentives werevailable for boats to develop annual catch plans.
While the combined effects of these and other changes in theoastal fishery stabilized overall catch during the 1980s, speciesad evidently reached maximum sustainable yield (Baisre, unpub-
ished; Baisre, 1985; Baisre and Páez, 1981). By the early 1990s,espite an overall reduction in commercial fishing effort due to anconomic crisis that reduced fishing, recovery in landings failed toccur. In the second half of the 1990s both commercial and sub-istence fishing increased and only by the end of the 1990s wasshing effort reduced with the creation of the National Organi-ation for Fishing Inspection. “Subsistence” fishing includes bothoaching and licensed recreational, or sport, fishing.
Snappers and the Nassau grouper are traditionally considered toe major finfish resources in Cuba but many of these species haveeclined over the last 4 decades. The existence of extensive seagrasseds (feeding areas) close to reefs, and mangroves (refuge areas) areavourable conditions for supporting large populations of mutton
Lutjanus analis), lane, L. synagris, cubera (Lutjanus cyanopterus) greyL. griseus) and yellowtail snappers (Ocyurus chrysurus). The Nas-au grouper was an important fishery resource until the 1970s andnhabited coral reefs along the insular shelf. The fishery of snappersnd groupers in Cuba typically concentrated on seasonal spawningearch 99 (2009) 7–16
aggregations. In addition to fishing activities, habitat degradationhas occurred in many near-shore areas over the last two decades(Claro et al., 2006).
The objectives of this paper are (1) to describe the history andchanges in catches over 45 years in the commercial Cuban fish-ery, with a focus on five species of snapper and a single species ofgrouper, the Nassau grouper, (2) to analyse the various factors thathave likely influenced landings over the same time period, inclu-sive of fishery management, social conditions, fishing practices andecological characteristics of the six species in (1) above, and (3) toevaluate the probable implications of the concentration of fishingeffort on fish spawning aggregations, with a view to improving themanagement of aggregating species.
2. Methods
Total coastal finfish landings for six targeted aggregating speciesand overall fishing effort in the Cuban fishery were compiled froma range of sources. Total finfish catch since 1955, and monthlycatches from the Cuban shelf between 1962 and 2005 for each offive snapper species and for the Nassau grouper, were obtained frommonthly and annual fisheries statistical reports elaborated by theDirectorate of Fisheries Statistics of the Ministry of Cuba and avail-able at regional fisheries agencies of the Ministry. Average monthlycatches for sequential five-year periods were estimated for eachspecies and data recorded by region. From the 1960s onwards, theCuban shelf was divided into four fishing regions for reporting pur-poses; southeast, southwest, northwest and northeast. Data on totalannual fishing effort (boat-days fishing) were also obtained. Due tothe multi-species character of the fisheries, it was not possible toestimate fishing effort at the species level. However, total fishingeffort in the finfish fishery overall was available from 1980 to 2005,and likely affected all species similarly.
For most species studied, commercial catches were recordedat the species level. For cubera and grey snappers, catches wereregistered together until 1982 and later separately, but species iden-tification is not considered to be reliable (RC personal observation),i.e., small fishes (<40 cm) were typically treated as grey snapper,medium or large fishes as cubera snapper. Therefore, data werecombined for these two species and treated as the grey-cuberafishery.
Supplementary information was collected from fishers, fish biol-ogists and the published literature to identify possible biases in thedata and to describe biological characteristics of the species, includ-ing spawning season, spawning and nursery habitats and otherenvironmental factors. Over 30 interviews of fishing boat captainsand crew were conducted in western and northern Cuba between1994 and 1996, and in 2006. In addition to the fishery data andanecdotal information, published and grey literature were summa-rized to construct an overall history of the fishery for each of the sixspecies, inclusive of management interventions. Approximate loca-tions of exploited spawning aggregation sites of the study speciesare shown in Fig. 1.
3. Results
Fishing activity consists of both commercial and subsistencefishing sectors. While these two sectors vary over time in terms oftheir relative importance and in the reliability of associated catchdata, commercial fishing accounts for the majority of the landings
and for the most reliable landings information. It is important tobriefly consider the different fishery sectors for possible biases inthe database examined in this study. Until the mid 1990s, both fish-ery landings and fishing effort data were considered to be reliableindicators of commercial fishing activity because the only autho-
R.Claro
etal./Fisheries
Research99
(2009)7–16
9
Fig. 1. Approximate locations of exploited spawning aggregation sites for five snappers and the Nassau grouper on the Cuban shelf, showing the species that aggregate at each one, and the peak months of spawning for eachspecies (modified from Claro and Lindeman, 2003).

10 R. Claro et al. / Fisheries Research 99 (2009) 7–16
F effortd
rfie1iikgolSapszgmt
(auftsi(ctiaaslslwi2
3
1d
in the different countries was not known because landings infor-mation was not distinguished by fishing area across countries.Particularly notable was the sharp decline in Cuban national catchesafter 1969. Fig. 2 shows a remarkable drop in the national catches
ig. 2. Total finfish catches from the Cuban shelf between 1959 and 2005, total finfishays) since 1980.
ized entity in the country allowed to buy commercial catches fromshers were government enterprises of the Ministry of the Fish-ries Industry, and most landings were commercial. In the mid990s, subsistence fishing increased due to an economic crisis. Thisntroduced uncertainty regarding the reliability of catch and effortnformation because subsistence catch was sold on the black mar-et or used for family consumption and is typically not reflected inovernment landings. As an indication of the relative importancef subsistence versus commercial landings, anecdotal and sporticense information suggest that sport fishing in the Archipelagoabana-Camagüey is about 1800 tonnes yearly (Claro et al., 2004)nd illegal fishing may reach approximately 500 tonnes. This com-ares with the much higher total commercial finfish catch over theame period in the same zone at about 6000 tonnes. In other fishingones, the subsistence fishery is not as important because fishingrounds are less accessible to subsistence fishers. Hence, the com-ercial fishery is not only the most important by weight, it is also
hat which is most reliably recorded.Examining the coastal finfish fishery as a whole, the total catches
Fig. 2) show the typical phases of a fishery that is moving fromn under- to an over-fished phase, as described by Caddy (1984);nder-utilization before 1960, and fast growth from 1961 to 1975ollowed by marked declines in species of key commercial impor-ance, as determined in the present study (Nassau grouper, lanenapper, grey/cubera snapper, mullets). This situation provoked thentroduction of management measures from the 1980s onwardsTable 1). Simultaneously, the organization and efficiency of theommercial fishery increased during the same decade. From 1994o 2000, fishing effort decreased, evidently favouring an increasen catch per unit effort (CPUE). Overall catches and CPUE declinedfter 2000 (Fig. 2). Review of commercial landings between 1978nd 1995 for species other than those addressed in the currenttudy reveals very different trends, although there is not the sameevel of species-specific detail as for the six focal species of thistudy. Among other reef fishes, notable declines occurred in mul-et (several species), fluctuating landings or less marked declines
ere noted for grunts (Haemulidae) and jacks (Carangidae), whilencreases were recorded for mojarras (Gerreidae) (Claro et al.,001).
.1. E. striatus (Nassau grouper)
Fishing pressure on the Nassau grouper increased notably after959, reaching 1700 tonnes in 1963, after which time landingseclined despite the increase in fishing pressure on the Cuban shelf
(boat-fishing days) and catch per unit effort (CPUE—measured as catch/boat-fishing
(Fig. 3A). Most catches of Nassau grouper (35–50% of the nationalcapture of the species) were historically taken in the ArchipelagoSabana-Camagüey (north-central area, Fig. 1; Claro et al., 2001).Although up until 1969 an important proportion of this catch wasobtained from the Bahamas shelf, the distribution of fishing effort
Fig. 3. Annual catches of Nassau grouper, Epinephelus striatus, in Cuba (A) and aver-age monthly catches by five-year periods (B).
R. Claro et al. / Fisheries Research 99 (2009) 7–16 11
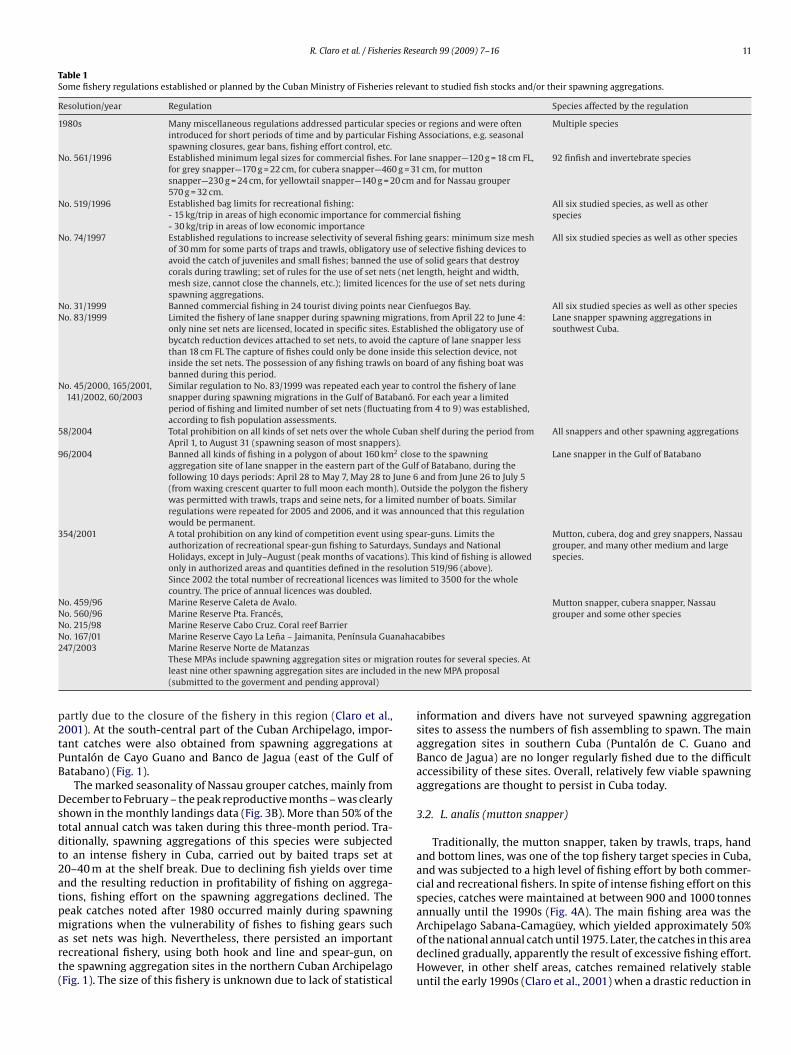
Table 1Some fishery regulations established or planned by the Cuban Ministry of Fisheries relevant to studied fish stocks and/or their spawning aggregations.
Resolution/year Regulation Species affected by the regulation
1980s Many miscellaneous regulations addressed particular species or regions and were oftenintroduced for short periods of time and by particular Fishing Associations, e.g. seasonalspawning closures, gear bans, fishing effort control, etc.
Multiple species
No. 561/1996 Established minimum legal sizes for commercial fishes. For lane snapper—120 g = 18 cm FL,for grey snapper—170 g = 22 cm, for cubera snapper—460 g = 31 cm, for muttonsnapper—230 g = 24 cm, for yellowtail snapper—140 g = 20 cm and for Nassau grouper570 g = 32 cm.
92 finfish and invertebrate species
No. 519/1996 Established bag limits for recreational fishing: All six studied species, as well as otherspecies- 15 kg/trip in areas of high economic importance for commercial fishing
- 30 kg/trip in areas of low economic importanceNo. 74/1997 Established regulations to increase selectivity of several fishing gears: minimum size mesh
of 30 mm for some parts of traps and trawls, obligatory use of selective fishing devices toavoid the catch of juveniles and small fishes; banned the use of solid gears that destroycorals during trawling; set of rules for the use of set nets (net length, height and width,mesh size, cannot close the channels, etc.); limited licences for the use of set nets duringspawning aggregations.
All six studied species as well as other species
No. 31/1999 Banned commercial fishing in 24 tourist diving points near Cienfuegos Bay. All six studied species as well as other speciesNo. 83/1999 Limited the fishery of lane snapper during spawning migrations, from April 22 to June 4:
only nine set nets are licensed, located in specific sites. Established the obligatory use ofbycatch reduction devices attached to set nets, to avoid the capture of lane snapper lessthan 18 cm FL The capture of fishes could only be done inside this selection device, notinside the set nets. The possession of any fishing trawls on board of any fishing boat wasbanned during this period.
Lane snapper spawning aggregations insouthwest Cuba.
No. 45/2000, 165/2001,141/2002, 60/2003
Similar regulation to No. 83/1999 was repeated each year to control the fishery of lanesnapper during spawning migrations in the Gulf of Batabanó. For each year a limitedperiod of fishing and limited number of set nets (fluctuating from 4 to 9) was established,according to fish population assessments.
58/2004 Total prohibition on all kinds of set nets over the whole Cuban shelf during the period fromApril 1, to August 31 (spawning season of most snappers).
All snappers and other spawning aggregations
96/2004 Banned all kinds of fishing in a polygon of about 160 km2 close to the spawningaggregation site of lane snapper in the eastern part of the Gulf of Batabano, during thefollowing 10 days periods: April 28 to May 7, May 28 to June 6 and from June 26 to July 5(from waxing crescent quarter to full moon each month). Outside the polygon the fisherywas permitted with trawls, traps and seine nets, for a limited number of boats. Similarregulations were repeated for 2005 and 2006, and it was announced that this regulationwould be permanent.
Lane snapper in the Gulf of Batabano
354/2001 A total prohibition on any kind of competition event using spear-guns. Limits theauthorization of recreational spear-gun fishing to Saturdays, Sundays and NationalHolidays, except in July–August (peak months of vacations). This kind of fishing is allowedonly in authorized areas and quantities defined in the resolution 519/96 (above).
Mutton, cubera, dog and grey snappers, Nassaugrouper, and many other medium and largespecies.
Since 2002 the total number of recreational licences was limited to 3500 for the wholecountry. The price of annual licences was doubled.
No. 459/96 Marine Reserve Caleta de Avalo. Mutton snapper, cubera snapper, Nassaugrouper and some other speciesNo. 560/96 Marine Reserve Pta. Francés,
No. 215/98 Marine Reserve Cabo Cruz. Coral reef BarrierNo. 167/01 Marine Reserve Cayo La Lena – Jaimanita, Península Guanahacabibes2
ation rin th
p2tPB
Dstdt2atpmart(
47/2003 Marine Reserve Norte de MatanzasThese MPAs include spawning aggregation sites or migrleast nine other spawning aggregation sites are included(submitted to the goverment and pending approval)
artly due to the closure of the fishery in this region (Claro et al.,001). At the south-central part of the Cuban Archipelago, impor-ant catches were also obtained from spawning aggregations atuntalón de Cayo Guano and Banco de Jagua (east of the Gulf ofatabano) (Fig. 1).
The marked seasonality of Nassau grouper catches, mainly fromecember to February – the peak reproductive months – was clearly
hown in the monthly landings data (Fig. 3B). More than 50% of theotal annual catch was taken during this three-month period. Tra-itionally, spawning aggregations of this species were subjectedo an intense fishery in Cuba, carried out by baited traps set at0–40 m at the shelf break. Due to declining fish yields over timend the resulting reduction in profitability of fishing on aggrega-ions, fishing effort on the spawning aggregations declined. Theeak catches noted after 1980 occurred mainly during spawning
igrations when the vulnerability of fishes to fishing gears suchs set nets was high. Nevertheless, there persisted an importantecreational fishery, using both hook and line and spear-gun, onhe spawning aggregation sites in the northern Cuban ArchipelagoFig. 1). The size of this fishery is unknown due to lack of statistical
outes for several species. Ate new MPA proposal
information and divers have not surveyed spawning aggregationsites to assess the numbers of fish assembling to spawn. The mainaggregation sites in southern Cuba (Puntalón de C. Guano andBanco de Jagua) are no longer regularly fished due to the difficultaccessibility of these sites. Overall, relatively few viable spawningaggregations are thought to persist in Cuba today.
3.2. L. analis (mutton snapper)
Traditionally, the mutton snapper, taken by trawls, traps, handand bottom lines, was one of the top fishery target species in Cuba,and was subjected to a high level of fishing effort by both commer-cial and recreational fishers. In spite of intense fishing effort on thisspecies, catches were maintained at between 900 and 1000 tonnesannually until the 1990s (Fig. 4A). The main fishing area was the
Archipelago Sabana-Camagüey, which yielded approximately 50%of the national annual catch until 1975. Later, the catches in this areadeclined gradually, apparently the result of excessive fishing effort.However, in other shelf areas, catches remained relatively stableuntil the early 1990s (Claro et al., 2001) when a drastic reduction in
12 R. Claro et al. / Fisheries Research 99 (2009) 7–16
Fm
ltoaItarf
tatt((2bttdicabf
ig. 4. Annual catches of mutton snapper, Lutjanus analis, in Cuba (A) and averageonthly catches by five-year periods (B).
andings occurred in all regions. Several factors likely contributedo this overall trend: a sharp reduction in commercial fishing effortccurred due to a national economic crisis that resulted in short-ges of fuel, materials and spare parts for boats and fishing gears.n addition, an intense subsistence fishery developed, mainly inhe Archipelago Sabana-Camagüey. Although this fishery affectedlmost all demersal and shore-pelagic species the mutton snappereportedly received greater fishing pressure due to high demandor the species.
The highest catches of mutton snapper were obtained duringhe reproductive period, which extended from April to August,nd at spawning aggregations; between May and June 35–40% ofhe national annual catch was taken (Fig. 4B). During this periodhe highest proportion of adult fish was captured, with juveniles<45 cm FL) prevailing in catches during other months of the yearPozo, 1979; Claro, 1981a; Claro et al., 2001; Claro and Lindeman,008). Aggregations were fished with hook and line at the shelfreak, but also with ‘set’ nets and trawling during spawning migra-ions. Since 2004 a new regulation banned the use of set nets overhe entire Cuban shelf (Table 1). Recreational fishermen also fishedirectly on spawning aggregations but there are no data on the
ntensity of this fishery. In most years, a moderate increment in
atches was observed in October, known by fishermen as “arrib-zones” (fish shoals); these differ from spawning aggregations,ecause the fish are not mature, and seem to be associated witheeding migrations (Claro, 1981a).Fig. 5. Annual catches of lane snapper, Lutjanus synagris, in Cuba (A) and averagemonthly catches by five-year periods (B).
3.3. L. synagris (lane snapper)
Lane snapper was the most important fishery species in Cubaand has been severely over-fished. Catches grew quickly in the1960s due to the widespread introduction of bottom trawls and thelarge numbers of traps, and because of the intense exploitation ofgroups of fish during their migration towards spawning aggregationareas at the shelf break. From 1960 to 1969, lane snapper was fishedmainly using trawl nets, but, starting in 1968, large set nets wereintroduced. Being highly efficient and productive, this gear blockedthe migration of lane snapper schools moving towards the edge ofthe platform. Other measures also led to higher fishing pressure onthis species after 1965, including reception of the catch in fishingareas by large transporter ships. Due to the combined use of trawlsand set nets, catches surpassed 4200 tonnes in 1969 and stayed atmore than 3000 tonnes up until 1977 (Fig. 5A), when catastrophicover-fishing of the species was identified (Claro et al., 2001).
The main catches of lane snapper were obtained in the Gulf ofBatabano, where from 1969 to 1975 more than 60% of the annuallane snapper catch was taken (Claro et al., 2001). The most spec-tacular reproductive aggregations known in Cuba were found inthe eastern region of the Gulf of Batabano. These groupings were
observed only in shallow waters in the north-central half of the Gulfand increased in number of fish as they advanced towards the shelfbreak at the Gulf of Cazones. In shallow waters, close to the shelfbreak, the fish schools remained during the day, and at dusk they
es Research 99 (2009) 7–16 13
mprfelet
fidfiSr
oriGa2gsAo
dsrs1cwmrsbpcs
3
blqe1err
ictatwsrFa(
o
R. Claro et al. / Fisheri
oved towards deeper waters, where mating and spawning of oneortion of the school took place. Immediately after spawning, fisheturned to nearby shallow waters where they remained until theollowing day, repeating the process over 4–5 days (García Cagidet al., 2001; Claro and Lindeman, 2008). The temporary nature ofane snapper schools at the spawning site may be an adaptation toscape predators such as sharks, great barracuda, and groupers athe shelf break (Claro, 1982).
Lane snapper spawning aggregations were also intensivelyshed in the Archipelago Sabana-Camagüey and Archipelago Jar-ines de la Reina (Fig. 1). In all known cases, the schools wereshed in shallow waters mainly with trawl nets. In the Archipelagoabana-Camagüey, catches were highly variable, probably theesult of intensive fishing effort.
Approximately 60–70% of the annual catch of lane snapper wasbtained during 10–12 days of fishing during the month of peakeproductive activity, generally coinciding with the period preced-ng full moon. The reproductive peak usually occurred in May in theulf of Batabano and the northwest shelf, and in June in the southnd northeast zones of the Cuban platform (Claro and Lindeman,003). The concentration of fishing effort on the spawning aggre-ations was likely a major factor in the dramatic collapse of the lanenapper fishery in the Gulf of Batabanó during the 1970s (Fig. 5B).less marked decline was noted in northeast and southeast areas
f Cuba (Claro et al., 2001).Various protective measures were introduced to halt the
eclines in this species but were not successful. After 1978, lanenapper fishing in the Gulf of Batabano was banned during theeproductive period (three months), and for the remaining (non-pawning) months an annual limited quota was introduced. From987, a permanent closure was established in shallow waters adja-ent to the spawning site where the schools generally concentratedhen not spawning. These protective measures were extended forore than 15 years without evident recovery. After 1995, limited
ecovery was observed in the Gulf of Batabanó and fishing of thepawning aggregations was authorized for a limited number ofoats and set nets; this may explain the increase in catches in theeriod 1996–2000. Over the next three years further reductions inatches occurred and a regulation prohibiting the deployment ofet nets over the entire Cuban platform was introduced in 2004.
.4. L. cyanopterus + L. griseus (cubera snapper + grey snapper)
Since fishery statistics in Cuba do not reliably distinguishetween captures of cubera and grey snapper, we combined the
andings data of these two species for analysis. Their fishery grewuickly after 1960, peaking in the late 1970s, with highest fishingffort directed principally towards the grey snapper (Fig. 6A). After982, a marked reduction of this resource took place while fishingffort remained constant. After 1990, social and economic factorsesulted in large shifts up and down in fishing effort and someecovery for the species may have occurred after 2000.
The two snappers are similar in morphology and their spawn-ng aggregations occurred over the same general time period; mostatches came from spawning aggregations. Cubera snappers wereaken predominantly from its spawning aggregations with hooknd line, while grey snappers were taken mainly (85–90%) byrawls, set nets and traps during spawning migrations in shallowaters (Claro, 1983b). The landings of both species showed marked
easonal trends with very high catches during the two-montheproductive period when they aggregated to spawn (July–August;
ig. 6B). Catches for these species also increased in October, prob-bly associated with feeding migrations of non-reproductive fishClaro, 1983b).Although both species reproduced at similar places and monthsf the year, they exhibited different reproductive strategies. Cubera
Fig. 6. Annual catches of cubera and grey snappers, Lutjanus griseus, and L.cyanopterus, in Cuba (A) and average monthly catches by five-year periods (B).
snapper spawned during and after the full moon (Claro andLindeman, 2003), while grey snapper spawned just prior to thenew moon (Domeier et al., 1996). Aggregated grey snapper werecaptured mainly in the channels and straits between the islandsthat skirt the Cuban platform close to the shelf break. Most fishescaptured had ripe gonads, as determined by high gonado-somaticindex (a measure that examines gonad weight relative to bodyweight; gonads increase in size in the spawning season) and thepresence of hydrated oocytes (the oocyte stage that occurs veryshortly before egg release at spawning) (Claro, 1983a), and theirdensities in areas close to the shelf edge increased sharply duringthese months. Aggregations of the grey snapper remained in shal-low waters during the day, moved towards the adjacent shelf breakin the evening to spawn, and returned immediately after spawningto nearby reefs and seagrass beds. This process repeated itself forindividual fish 4–5 times during 8–10 days in one lunar phase and,in the following month other individuals underwent similar cycles(Claro, 1983a; Claro and Lindeman, 2008). Large aggregations arenot known for grey snapper which probably forms smaller groupsfor spawning. Large groups of cubera snapper are also captured dur-ing their spawning migrations in the straits among the keys, close
to spawning sites. Contrary to the grey snapper, however, cuberasnapper remained at the shelf break for several days, where itsaggregations are seen together with those of L. jocu, dog snapper(García Cagide et al., 2001; Heyman et al., 2001, 2005; Claro and
14 R. Claro et al. / Fisheries Res
Fa
Lw
3
dfibaWtnCtlysatspcia
ig. 7. Annual catches of yellowtail snapper, Ocyurus chrysurus, in Cuba (A) andverage monthly catches by five-year periods (B).
indeman, 2003). In these places, the species is fished intensivelyith hook and line by commercial fishermen.
.5. O. chrysurus (yellowtail snapper)
Up until 1955–1958 annual catches of yellowtail snapper catchid not exceed 500 tonnes but subsequently catches increased asshing pressure grew. Initially a large proportion of fish taken wereig specimens because most fishing was carried out with hooknd line at the shelf break, where big fish were more abundant.ith the introduction of bottom trawls (in shallow waters) and
he increase in trap number in the early 1960s, catches increasedotably (Fig. 7A) with small size fish prevailing (Claro, 1983d;arrillo de Albornoz and Ramiro, 1988; Claro et al., 2004). The use ofhese large fishing gears resulted in the abandonment of hook andine fishing and was probably a major factor in the decline in size ofellowtail taken in catches. After 1969, the fishery underwent sub-tantial changes, apparently due to the adoption of managementctions that indirectly affected the yellowtail fishery. Althoughhese actions were not specifically directed towards the yellowtail
napper, its target species, such as lane, mutton and grey snap-ers, partly share the same habitat. After 1987 yellowtail snapperatches declined, partly because of high mortality of juveniles takenn trawl fisheries for this species (Bustamante et al., 1982; Claro etl., 2004). The south-central area of Cuba (Archipelago Jardines deearch 99 (2009) 7–16
la Reina; Fig. 1) contributed 35–40% of the national catch of yel-lowtail snapper, and the Gulf of Batabano 25–35% of the nationalcatch. Other catches came from shelf areas of northern Cuba.Insufficient information precludes a more detailed analysis of thechanges in catch volumes and patterns observed over time for thisspecies.
The reproductive period of yellowtail snapper in Cuba extendedfrom March to October and spawning aggregations were less welldefined, either spatially or temporally, compared to the otherspecies included in the present study. The spawning peak in the Gulfof Batabano was observed during May and June, while in the north-west region of Cuba it occurred in April and May (Claro, 1983c).Although specific spawning aggregation sites were not recorded,there were clear increases in density of yellowtail snapper at mul-tiple locations of the reef close to the edge of the platform duringthe respective reproductive periods, compared to non-reproductiveperiods, and most landings were greatest when fish were ripe(Fig. 7B). During the reproductive season, most individuals hadswollen abdomens, many had hydrated oocytes, and there was anincrease in the gonado-somatic index (Claro, 1983c).
4. Discussion
The fishery database of Cuba is unique among coral reef fish-eries, both in terms of the long-term nature of the dataset as wellas in its relative reliability and monthly detail for several key com-mercially important species. As such, it is valuable for evaluatingdetails of landings trends, hypothesizing on the major causes ofchanges observed over time, and for developing options for man-agement. The data clearly show that, over the last 4–5 decades,there were marked and often striking changes in the fishery of sev-eral important commercial fish species over the coastal platformof Cuba, with overall trends differing between species according totheir biology and fishery vulnerability. While it is not possible toestablish cause and effect for each study species, some clear trendsfrom the data, and through comparisons between species, emergedthat allow for an in-depth, albeit qualitative, evaluation of Cuba’sfisheries. The following discussion considers a range of factors inrelation to the trends in the six study fish species, inclusive of fishingeffort, management interventions and economic shifts, environ-mental and biological factors, and draws several key conclusionsof fishery significance.
In all studied species there were clear signs of over-fishing, espe-cially among those that aggregate to spawn with high predictability,despite a range of management interventions since the 1980s tocombat over-fishing. While some of the management measuresappear to have resulted in positive outcomes for the fishery inthe short-term, enforcement was evidently variable in effective-ness and the fishery overall declined. As a result, there was a steadyincrease in the number and variety of management measures intro-duced and considered to address the problem of over-fishing overseveral decades. Two factors were strongly implicated as contrib-utors to over-fishing; excessive and largely uncontrolled fishingeffort in terms of both number of vessels and scale and types offishing gears, and intense fishing focused on spawning aggrega-tions and accounting for a large proportion of annual catches for allspecies. The study species are the only ones in the fishery exploitedat, or close to, spawning aggregations, although mullets, whichalso have declined markedly, are taken during spawning aggre-gation and migrations in some areas. In addition, environmental
changes may have contributed to the trends noted in these reef fishfisheries. For example, the degradation of coral reefs as result ofcoral bleaching and algal overgrowth during several ENSO events,may have negatively affected aspects of the life cycle of these reef-associated species (Alcolado, 2004; Claro et al., 2006). In the case
es Rese
om
essmtetNslrelIimS
tiedvsmr1isipnytcthha
wmbonlMottas(cp
lbltpsn
R. Claro et al. / Fisheri
f the mullets, fished largely in estuaries and coastal lagoons, riverodifications may also have affected their abundance.The differences in responses to similar overall fishing pressures
xperienced by the six study species, which vary biologically ineveral respects, allow for interesting comparisons. Of all studypecies, the Nassau grouper and lane snapper exhibited the mostarked declines, almost certainly related to the heavy concentra-
ion of fishing effort on their spawning aggregations and lack offfective species-specific regulations. Both fisheries were subjectedo heavy subsistence use in addition to the commercial fishery. Forassau grouper, there was an almost complete absence of species-
pecific protective management, with the exception of a minimumegal size that is too small for the species (32 cm TL). Moreover,elative to other study species the Nassau grouper has the short-st reproductive season, is highly vulnerable to traps and hook andine, and there are few spawning sites (Claro and Lindeman, 2003).n addition, this species is long-lived with late maturation, makingt particularly vulnerable to over-fishing and in need of appropriate
anagement action (Claro et al., 1990; Sadovy and Eklund, 1999;ala et al., 2001; Sadovy de Mitcheson et al., 2008).
For the lane snapper, its collapse in the Gulf of Babatano inhe 1970s was likely the result of several non-sustainable fish-ng actions: intensive fishing throughout the year and high fishingffort with highly effective fishing gear (set nets and bottom trawls)uring spawning migrations. Given the various management inter-entions introduced in the 1980s for this important commercialpecies, its relatively short life cycle, fast growth rate and earlyaturation (Claro, 1981b, 1982; Claro and Lindeman, 2008) some
ecovery was expected. While a slow increase in catches over5 years was noted, and increasingly strict fishing regulationsntroduced (limited fishing effort, a quota and a minimum legalize), reintroduction of intense fishing activity during the spawn-ng season was probably the major factor that again reduced theopulation. Although fishing regulations were enforced, they mayot have provided sufficient protection; for example, in someears catches were permitted during spawning seasons or quo-as were too high. Over the last decade, lane snapper stocksontinued to decline despite additional regulations (Table 1). Habi-at degradation may also have influenced this species throughyper-salinization of inner lagoons, damage to Thalassia beds fromurricanes, and changes in water circulation in coastal areas (Clarond Lindeman, 2008).
Although mutton snapper catches exhibited clear declines, theseere less marked than for the Nassau grouper and lane snapper. Theutton snapper was not found in large concentrated aggregations
ut rather in gatherings close to the shelf break over a short periodf time. It is noteworthy that fishing on spawning aggregations wasot as intense for the mutton snapper as for the Nassau grouper and
ane snapper largely due to differences in reproductive behaviour.utton snapper did not congregate in shallow waters and were
nly fished by hook and line, which does not have the same nega-ive impact as trawls or set nets. Another factor that might influencehe maintenance of high landings in the mutton snapper is the rel-tively high number of spawning sites compared to other speciestudied (Fig. 1). Moreover, the existence of extensive seagrass bedsfeeding areas), close to the reefs and mangroves (refuge areas)reates favourable conditions for the persistence of an abundantopulation for mutton snapper in Cuba (Claro and Lindeman, 2008).
Grey snapper have similar reproductive behaviour to theane snapper, are intermediate in their biological characteristicsetween the lane and mutton snappers and exhibited declines in
andings. They have a longer life span and later sexual maturationhan lane snapper, and the converse compared to the mutton snap-er (Claro, 1981a, 1983b). Management measures affecting the greynapper have largely been driven by changes occurring in the eco-omically more valuable lane snapper stocks. However, due to the
arch 99 (2009) 7–16 15
absence of large and highly concentrated spawning aggregations,grey snappers are less vulnerable to intense fishing during theirspawning season. This species is in need of protective regulations,especially during the reproductive season.
Reproductive behaviours and other biological attributes ofcubera snapper resemble those of the mutton snapper, but theirnatural population levels are relatively smaller, possibly due tomore specialized habitat requirements (Starck, 1970; Lindemanand DeMaria, 2005). For the cubera snapper, there is little focusedfishing effort other than during short periods when spawningmigrations form and at the spawning sites, and the species is notas intensively fished as other snappers. Protective measures clearlyare needed, however, particularly on spawning aggregations.
Of all the species studied yellowtail snapper forms the least con-centrated and predictable spawning aggregations but nonethelesshas undergone declines in catches. The species is intensively fishedduring the spawning season and no protective measures wereestablished except for a minimum legal size that is arguably toosmall (Claro et al., 2004). The yellowtail snapper is fished togetherwith lane and grey snappers, all them coincident in time andspace during their respective spawning periods. Yellowtail snapper,however, seem to be less vulnerable to fishing because their aggre-gations are not highly concentrated, which may explain the lesspronounced decline in catches over the last few decades in com-parison with other snappers. On the other hand, the fishery mainlyextracts small size adults since larger adults prefer deeper waters,where trawling is not possible. This means that an important partof the population may continue to spawn in less accessible deeperwaters. Another negative impact on yellowtail snapper is the highproportion of juvenile fishes taken by trawls and traps. For example,in the Archipelago Sabana-Camagüey 85.8% of all yellowtail snap-per in trawl catches, and 40.5% taken in traps, are juveniles (Claroet al., 2004).
There is evidence that other, anthropogenically driven environ-mental factors may have contributed to declines in snapper andNassau grouper populations in Cuba. These include reduction ofseagrass areas in some regions, such as the inner waters of theArchipelago Sabana-Camagüey (Alcolado et al., 1999), increases incoral bleaching and diseases associated with increases in watertemperatures during ENSO events (Claro et al., 2006). Other fac-tors may include damage to coastal marine habitats from variouscauses, such as pollution and other kinds of habitat degradationthat have increased in recent years (Martínez, Montalvo, Miravet,Lugioyo, Loza and Pérez, unpublished report 2004; Claro, 2007).
Considering all six study species, the greatest overall depletionsoccurred in those species that are most intensively exploited dur-ing their spawning aggregations. In this respect is noteworthy thatmarked declines have also been observed in mullet species, whichare often targeted on or moving to spawning aggregations. This is incontrast to other groups of fishes such as jacks, mojarras and gruntsthat are not knowingly targeted in association with aggregations.In the Nassau grouper and lane snapper, spawning aggregationsappear to be particularly distinctive with animals predictably con-centrated in large numbers and readily accessible to fishermen inwell-known months. Even though the mutton snapper is also ahighly aggregative species and is heavily exploited, the use of selec-tive and small-scale fishing gears at the spawning sites and the largenumber of spawning sites relative to Nassau grouper and lane snap-per, appear to allow for persistence of the fishery even under highfishing pressure. Likewise, species such as the grey and yellowtailsnappers seem to be less affected by heavy exploitation during the
spawning season, although there are clear signs of stock declines inboth species.The overall resilience, or response, of different species to fishingultimately depends on the intensity and nature of fishing activ-ity, vulnerability to fishing and other biological features, such as

1 es Res
re1ttbmtFk
5
meaamanaaielcvimhevi
A
tcSwAtt
R
A
A
A
B
B
B
B
6 R. Claro et al. / Fisheri
eproductive behaviour, longevity, and maturation size, as well asnvironmental factors and management effectiveness (Ault et al.,998). Although the development of a sustainable fishery needs toake such factors into account, it is clear from a comparison acrosshe six species that one measure likely to benefit all species woulde the successful implementation of species-specific conservationanagement. For some species this needs to focus on spawning
imes and/or sites during the spawning season. The Ministry of theisheries Industry is working towards the total elimination of allinds of bottom trawls on the Cuban shelf.
. Conclusion
The long-term species-specific database and related manage-ent and social information available in Cuba provide a rare and
xtremely valuable opportunity for understanding fishing historynd for identifying and evaluating management options. Man-gement is so reactive in many countries that data from pastanagement interventions are often never gathered, much less
nalysed, and, remarkably, the possible effects on the fishery oftenot considered. Many problems with scientific prediction as wells problems among agency–fisher relations arise because of thebsence of follow-up analyses of how management interventionsmpact various processes over time. In many countries, even if fish-ry data are collected and available for analysis they often involveumping of species or give no, or little, indication of effort or socialhanges that could have affected landings. The Cuban dataset pro-ides an important lesson on the value of detailed species-specificnformation and illustrates clearly the importance of sustained
anagement and precautionary approaches. It also demonstratesow the biology of exploited species, particularly species thatxhibit differences in their spawning behaviour in relation to theirulnerability to exploitation, can influence their responses to fish-ng practices.
cknowledgements
We are most grateful to the many fishers who assisted us. Fromhe Ministry of the Fisheries Industry of Cuba, we thank the Provin-ial Fisheries Associations, the Directorate of Coastal Fisheries andervando Valle (Centre of Fisheries Research). Funding for this workas provided by the Society for the Conservation of Reef Fishggregations, which is partly supported by the Packard Founda-
ion. Additional funding was provided by a Doherty Fellowship athe Department of Marine and Environmental Systems, FIT.
eferences
lcolado, P.M., García, E.E., Espinosa, N. (Eds.), 1999. Protecting biodiversity andestablishing sustainable development in the Sabana-Camagüey Ecosystem.Global Environmental Facility (GEF)/United Nations Development Program(UNDP) Project CUB/92/G31 Sabana-Camagüey, Cuba, 145 pp.
lcolado, P.M., 2004. Manual de capacitación para el monitoreo voluntario de alertatemprana en arrecifes coralinos. Proyecto PNUD/GEF, CUB/98/G32, CUB/99/G81– Capacidad 21. “Acciones Prioritarias para consolidar la Protección de la Biodi-versidad en el Ecosistema Sabana-Camagüey”. La Habana, 80 pp. (in Spanish).
ult, J.S., Bohnsack, J.A., Meester, G.A., 1998. A retrospective (1979–1996) multi-species assessment of coral reef fish stocks in the Florida Keys. Fishery Bulletin96 (3), 395–414.
aisre, J.A., 1985. Los recursos pesqueros marinos de Cuba: Fundamentos ecológicosy estrategias para su utilización. Tesis doctoral en Ciencias Biológicas. Univ. dela Habana, 189 pp. (in Spanish).
aisre, J.A., 2000. Chronicle of Cuban marine fisheries (1935–1995). Trend analysisand fisheries potential. FAO Fish. Tech. Pap. 394, 26 pp.
aisre, J.A., 2004. La pesca marítima en Cuba. Editorial Científico-Técnica. La Habana,372 pp. (in Spanish).
aisre, J.A., Páez, J., 1981. Los recursos pesqueros del archipiélago cubano. EstudiosWECAF 8, 1–79 (in Spanish).
earch 99 (2009) 7–16
Bustamante, G., García-Jorge, J.E., García-Arteaga, J.P., 1982. La pesca con chinchorroen la región oriental del Golfo de Batabanó y algunos datos sobre las pesqueríasen la plataforma cubana. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 4, 1–31 (inSpanish).
Caddy, J.F., 1984. An alternative to equilibrium theory for management of fisheries.FAO Fish. Rep. No. 289, Suppl. 2. Rome, Italy, 214 pp.
Carrillo de Albornoz, C., Ramiro, M.E., 1988. Estudio biológico de la rabirrubia (Ocyu-rus chrysurus) en el W de la plataforma SE de Cuba. I. Edad y crecimiento. Rev.Invest. Mar. 9 (1), 9–24 (in Spanish).
Claro, R., 1981a. Ecología y ciclo de vida del pargo criollo, Lutjanus analis (Cuvier), enla plataforma cubana. Inf. Cient. -Téc., Acad. Cienc. Cuba. 186, 1–83 (in Spanish).
Claro, R., 1981b. Ecología y ciclo de vida de la biajaiba, Lutjanus synagris (Linnaeus),en la plataforma cubana. II. Biología pesquera. Inf. Cient. -Téc., Acad. Cienc. Cuba177, 1–53 (in Spanish).
Claro, R., 1982. Ecología y ciclo de vida de la biajaiba, Lutjanus synagris (Linnaeus), enla plataforma cubana. IV. Reproducción. Rep. Invest. Inst. Oceanol. Acad. Cienc.Cuba 5, 1–37 (in Spanish).
Claro, R., 1983a. Ecología y ciclo de vida del caballerote, Lutjanus griseus (Linnaeus), enla plataforma cubana. Identidad, distribución y hábitat, nutrición y reproducción.Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 7, 1–30.
Claro, R., 1983b. Ecología y ciclo de vida del caballerote, Lutjanus griseus (Linnaeus),en la plataforma cubana. II. Edad y crecimiento, estructura de las poblaciones,pesquerías. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 8, 1–28 (in Spanish).
Claro, R., 1983c. Ecología y ciclo de vida de la rabirrubia, Ocyurus chrysurus (Bloch),en la plataforma cubana. I. Identi dad, distribución, hábitat, reproducción y ali-mentación. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 15, 1–34 (in Spanish).
Claro, R., 1983d. Ecología y ciclo de vida de la rabirrubia Ocyurus chrysurus (Bloch),en la plataforma cubana. II. Edad y crecimiento, estructura de poblaciones ypesquerías. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 19, 1–33 (in Spanish).
Claro, R., Lindeman, K.C., 2003. Spawning aggregation sites of snapper and grouperspecies (Lutjanidae and Serranidae) on the Insular Shelf of Cuba. Gulf Carib. Res.14 (2), 91–106.
Claro, R., Lindeman, K.C., 2008. Biología y manejo de los pargos (Lutjanidae)en el Atlántico occidental. Instituto de Oceanología, CITMA, La Habana,Cuba. 472 pp en CD-ROM. ISBN: 878-959-298-011-2. Available on line at:http://www.redciencia.cu (in Spanish).
Claro, R., García-Cagide, A., Sierra, L.M., García-Arteaga, J.P., 1990. Característicasbiológico-pesqueras de la cherna criolla Epinephelus striatus (Bloch) (Pisces:Serranidae) en la plataforma cubana. Cienc. Biol. 23, 23–42 (in Spanish).
Claro, R., Baisre, J.A., Lindeman, K.C., García-Arteaga, J.P., 2001. Cuban fisheries: his-torical trends and current status. In: Claro, R., Lindeman, K.C., Parenti, L.R. (Eds.),Ecology of the Marine Fishes of Cuba. Smithsonian Institution Press, Washingtonand London, pp. 194–218.
Claro, R., Garcia-Arteaga, J.P., Gobert, B., Cantelar,.K., Valle Gomez, S.V., Pina-Amargos,F., 2004. Situacion actual de los recursos pesqueros del archipielago Sabana-Camaguey. Cuba. Bol. Invest. Mar. Cost. 33, 41–57 (in Spanish).
Claro, R., Cantelar, K., Pina Amargós, F., García-Arteaga, J.P., 2006. Cambios enlas comunidades de peces de los arrecifes coralinos del Archipiélago Sabana-Camagüey. Cuba Biología Tropical 55 (2), 537–547 (in Spanish).
Claro R., 2007. Conservación y manejo. En R. Claro (Ed.), La Biodiversidad marina deCuba. (CD-ROM). Instituto de Oceanología, Ministerio de Ciencia, Tecnología yMedio Ambiente, La Habana, Cuba. ISBN: 978-959-298-001-3. Available on lineat: http://www.redciencia.cu (in Spanish).
Domeier, M.L., Koenig, C., Coleman, F., 1996. Reproductive biology of the grey snapper(Lutjanus griseus), with notes on spawning for other western Atlantic snappers(Lutjanidae). In: Arreguin-Sanchez, F., Munro, J.L., Balgos, M.C., Pauly, D. (Eds.),Biology, fisheries and culture of tropical groupers and snappers. Makati CityPhilippines ICLARM 1996 no. 48, pp. 189–201.
García Cagide, A., Claro, R., Koshelev, B.V., 2001. Reproductive patterns of fishes ofthe Cuban Shelf. In: Claro, R., Lindeman, K.C., Parenti, L.R. (Eds.), Ecology of theMarine Fishes of Cuba. Smithsonian Institution Press, Washington and London,pp. 73–114.
Heyman, W.D., Graham, R.T., Kjerfve, B., Johannes, R.E., 2001. Whale sharks Rhincodontypus aggregate to feed on fish spawn in Belize. Mar. Ecol. Prog. Ser. 215, 275–282.
Heyman, W.D., Kjerfve, B., Graham, R.T., Rhodes, K.L., Garbutt, L., 2005. Spawningaggregations of Lutjanus cyanopterus (Cuvier) on the Belize Barrier Reef over a6 year period. J. Fish Biol. 67, 83–101, Available online at http://www.blackwell-synergy.com.
Lindeman, K.C., DeMaria, D., 2005. Juveniles of the Caribbean’s largest coral reefsnapper do not use reefs. Coral Reefs 24, 359.
Pozo, E., 1979. Edad y crecimiento del pargo criollo (Lutjanus analis [Cuvier, 1828])en la plataforma noroccidental de Cuba. Rev. Cub. Invest. Pesq. 4 (2), 1–24.
Sadovy de Mitcheson, Y., Cornish, A., Domeier, M., Colin, P., Russell, M., Lindeman, K.,2008. A global baseline for spawning aggregations of reef fishes. ConservationBiol. 22 (5), 1233–1244.
Sadovy, Y., Eklund, A., 1999. Synopsis of biological data on the Nassau grouper,Epinephelus striatus (Bloch, 1792), and the jewfish, E. itajara (Lichtenstein, 1822).
NOAA Tech. Rep. NMFS 146, 65 pp.Sala, E., Ballesteros, E., Starr, R., 2001. Rapid decline of Nassau grouper spawningaggregations in Belize: fishery management and conservation needs. Fisheries26, 23–30.
Starck, W.A., 1970. Biology of the grey snapper, Lutjanus griseus (Linnaeus), in theFlorida Keys. Stud. Trop. Oceanogr. Univ. Miami 10, 1–150.
Related Documents











