JOURNAL OF MORPHOLOGY 216:179-195 (1993) Histology of the Late-Stage Placentae in the Matrotrophic Skink Chalcides chalcides (Lacerti I i a; Sci n ci d ae) DANIEL G. BLACKBURN Department of Biology, Trinity College, Hartford, Connectscut 06106 ABSTRACT Examination of late-stage placental material of the lizard Chal- cides chalcides from the Hubrecht Laboratorium (Utrecht, The Netherlands) reveals several cytological and histological specializations that appear to have been superimposed over a morphological pattern that is typical for squamates. The chorioallantoic placenta is highly vascularized and consists of a single mesometrial placentome and a generalized paraplacentomal region, both of which are epitheliochorial. The placentome is deciduate, and contains deeply interdigitating folds of hypertrophied uterine and chorioallantoic tissue. Chori- onic epithelium lining the placentome comprises enlarged, microvilliated cells, a small proportion of which are diplokaryocytes. The placentomal uterine epithelium is not syncytial and consists of enlarged cells bearing microvilli. The yolk sac placenta is a true omphaloplacenta (sensu stricto), being formed by juxtaposition of uterine tissues to an avascular, bilaminar omphalopleure. Epithelium of the omphalopleure is stratified and is hypertrophied into papillae that project into detritus of the uterine lumen. The omphalopleure is separated from the yolk sac proper by a yolk cleft that is not confluent with the exocoelom and is not invaded by the allantois. Neither an omphalallantoic placenta nor a true choriovitelline placenta is present in late gestation. Morphologically, the mature placentae of C. chalcides are among the most specialized to have been described in reptiles, reflecting the substantial maternal-fetal nutrient transfer that occurs in this species. Q 1993 Wiley-Lisa, Inc, The viviparous scincid lizard Chalcides chalcides is one of a very few reptiles known to exhibit a substantial degree of maternal- fetal nutrient transfer during gestation (Yaron, '85; Ghiara et al., ,871, a fetal nu- tritional pattern known as matrotrophy (Wourms, '81; Blackburn, '92). Information on the morphological basis for this nutrient transfer derives almost exclusively from an influential paper published over one-half a century ago by Nicolaas Jurjan ten Cate- Hoedemaker ('331, a work that lacks photo- micrographs. The only other works on placen- tation in C. chalcides deal either mainly with macroscopic observations (Studiati, 1853; Giacomini, 1891) or briefly with early devel- opmental stages (Luckett, '77a,b) or chori- onic cytology (Ghiara et al., '87). In the absence of photomicrographic evi- dence and a detailed, contemporary morpho- logical description, several important issues remain unresolved. For example, interpreta- tions of the yolk sac placenta in Chalcides chalcides (Giacomini, 1891; ten Cate-Hoede- maker, '33) are difficult to reconcile with observations on the early-stage yolk sac (Luckett, '77a,b) as well as with information on other squamates (lizards, snakes, and am- phisbaenians). In addition, placental charac- teristics have been inferred for C. chalcides that have not been demonstrated in other squamates, such as the absence of an isolated yolk mass and intravitelline mesoderm (Weekes, '27) and the presence of a deciduate chorioallantoic placenta of the endothelial- endothelial type (ten Cate-Hoedemaker, '33). The possible uniqueness of placentation in C. chalcides cannot be assessed from the infor- mation now available, with the result that data on this species and other squamate rep- tiles have never been fully integrated. The scarcity of information on C. chalcides is especially problematical in that, anatomi- cally and physiologically, placentation in this species is one of the most complex to have been described in reptiles (Yaron, '851, and, evolutionarily, placentation appears to have converged on that of certain eutherian mam- o 1993 WILEY-LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JOURNAL OF MORPHOLOGY 216:179-195 (1993)
Histology of the Late-Stage Placentae in the Matrotrophic Skink Chalcides chalcides (Lacerti I i a; Sci n ci d ae)
DANIEL G. BLACKBURN Department of Biology, Trinity College, Hartford, Connectscut 06106
ABSTRACT Examination of late-stage placental material of the lizard Chal- cides chalcides from the Hubrecht Laboratorium (Utrecht, The Netherlands) reveals several cytological and histological specializations that appear to have been superimposed over a morphological pattern that is typical for squamates. The chorioallantoic placenta is highly vascularized and consists of a single mesometrial placentome and a generalized paraplacentomal region, both of which are epitheliochorial. The placentome is deciduate, and contains deeply interdigitating folds of hypertrophied uterine and chorioallantoic tissue. Chori- onic epithelium lining the placentome comprises enlarged, microvilliated cells, a small proportion of which are diplokaryocytes. The placentomal uterine epithelium is not syncytial and consists of enlarged cells bearing microvilli. The yolk sac placenta is a true omphaloplacenta (sensu stricto), being formed by juxtaposition of uterine tissues to an avascular, bilaminar omphalopleure. Epithelium of the omphalopleure is stratified and is hypertrophied into papillae that project into detritus of the uterine lumen. The omphalopleure is separated from the yolk sac proper by a yolk cleft that is not confluent with the exocoelom and is not invaded by the allantois. Neither an omphalallantoic placenta nor a true choriovitelline placenta is present in late gestation. Morphologically, the mature placentae of C. chalcides are among the most specialized to have been described in reptiles, reflecting the substantial maternal-fetal nutrient transfer that occurs in this species. Q 1993 Wiley-Lisa, Inc,
The viviparous scincid lizard Chalcides chalcides is one of a very few reptiles known to exhibit a substantial degree of maternal- fetal nutrient transfer during gestation (Yaron, '85; Ghiara et al., ,871, a fetal nu- tritional pattern known as matrotrophy (Wourms, '81; Blackburn, '92). Information on the morphological basis for this nutrient transfer derives almost exclusively from an influential paper published over one-half a century ago by Nicolaas Jurjan ten Cate- Hoedemaker ('331, a work that lacks photo- micrographs. The only other works on placen- tation in C. chalcides deal either mainly with macroscopic observations (Studiati, 1853; Giacomini, 1891) or briefly with early devel- opmental stages (Luckett, '77a,b) or chori- onic cytology (Ghiara et al., '87).
In the absence of photomicrographic evi- dence and a detailed, contemporary morpho- logical description, several important issues remain unresolved. For example, interpreta- tions of the yolk sac placenta in Chalcides chalcides (Giacomini, 1891; ten Cate-Hoede-
maker, '33) are difficult to reconcile with observations on the early-stage yolk sac (Luckett, '77a,b) as well as with information on other squamates (lizards, snakes, and am- phisbaenians). In addition, placental charac- teristics have been inferred for C. chalcides that have not been demonstrated in other squamates, such as the absence of an isolated yolk mass and intravitelline mesoderm (Weekes, '27) and the presence of a deciduate chorioallantoic placenta of the endothelial- endothelial type (ten Cate-Hoedemaker, '33). The possible uniqueness of placentation in C. chalcides cannot be assessed from the infor- mation now available, with the result that data on this species and other squamate rep- tiles have never been fully integrated. The scarcity of information on C. chalcides is especially problematical in that, anatomi- cally and physiologically, placentation in this species is one of the most complex to have been described in reptiles (Yaron, '851, and, evolutionarily, placentation appears to have converged on that of certain eutherian mam-
o 1993 WILEY-LISS, INC.

180 D.G. BLACKBURN
mals (ten Cate-Hoedemaker, ’33; Blackburn, ’92).
Through the generosity of the curators of the Embryological Collection at the Hu- brecht Laboratorium in Utrecht, I have had the opportunity to study in detail the histolog- ical specimens that ten Cate-Hoedemaker used in his 1933 investigation. The present paper draws on this invaluable material to provide a new description and interpretation of the histology of the late-stage placentae of Chalcides chalcides, corroborated by means of the first photomicrographs to be published of this material, Particular attention is fo- cused herein on previously unresolved issues, incongruent interpretations, and features of functional or evolutionary significance.
MATERIALS AND METHODS
The histological material of Chalcides chal- cides used in this study is the property of the International Embryological Institute of the Hubrecht Laboratorium in Utrecht, The Netherlands. The material includes serial sec- tions of excised placentomes, excised yolk sac placentae, and oviductal conceptuses with intact placental membranes. The material was found to be in good condition, with some of the series being incomplete. Sections had been mounted consecutively but not always serially. Fixation and sectioning artifacts are sometimes apparent, and several of the slides show evidence of diminished stain intensity, though not more than one would expect from their age.
Fixation reportedly was accomplished by introducing Bouin’s fixative directly into the peritoneal cavity (ten Cate-Hoedemaker, ’33, p. 318). Many details of the embedding and sectioning procedures are no longer avail- able. Records at the Institute indicate that the material was sectioned at 10 p,m (G. Boterenbrood, personal communication) in transverse, sagittal, and frontal planes. Among the histological stains employed were hematoxylin, iron hematoxylin, and hem- alum, with Congo red and van Gieson’s stain applied as counterstains (ten Cate-Hoede- maker, ’33). Consecutive slides often had been treated with different stains, showing alterna- tive features of the same structures.
In the present study, the histological mate- rial was examined and photographed by means of Olympus BH-2 microscopes fitted with S Plan Apochromat objectives with cor- rection collars and equipped with a Nikon camera. Photographs were taken on Kodak T Max-100 professional film, processed at 120%
of the recommended development time. Mag- nification values reported in this paper are of the published micrographs, as calculated from parallel photomicrographs of a stage mi- crometer.
RESULTS Fetal morphology
The fetuses in the material studied all ap- pear to be at equivalent stages of develop- ment. In each fetus, scales have differenti- ated on the head, eyelids, body, limbs, and digits. The tridactyl manus and pes bear claws. The integument is pigmented, as is the peritoneum, choroid, and iris. Neural arches and centra have begun to ossify throughout the vertebral column, including the caudal region. Ossification also is apparent in the skull, mandible, hyoid, ribs, limb girdles, and diaphyses of the stylopodial, zeugopodial, and autopodial elements. Anatomy of the fetuses indicates that they are in a very advanced stage of development.
Uterine oviduct The oviduct is greatly distended by the
conceptuses, with segments between adja- cent fetuses being relatively constricted (Fig. 1). In mounted sections, the gravid segments or “incubation chambers” (“camera incuba- trice” of Giacomini, 1891) averaged at maxi- mum - 5 mm in height (i.e., mesometrial to antimesometrial pole) x 9 mm in transverse width.
The structural organization of the uterine portion of the oviduct is best appreciated through examination of the interembryonic segments (Fig. 2). The epithelium lining the oviductal lumen consists of a single layer of cells, which range from cuboidal, to colum- nar, to pseudostratified columnar. Cell apices are gently rounded, and at least some bear cilia. Cell boundaries are indistinct but gener- ally visible, and terminal bars are apparent. Epithelial cell nuclei are round to vertically elongate, are approximately centrally lo- cated, and contain nucleoli and heterochro- matin. At the base of the epithelium lie capil- laries and pre- and postcapillary vessels. These small vessels protrude upward, displac- ing portions of the overlying epithelial cells; thus basal portions of the epithelial cells ex- hibit a scalloped appearance. Deep to the epithelium and associated vessels lies a lam- ina propria of areolar to mucoid connective tissue, often greatly thinned and containing arterioles and venules. Glands are not evi-

181 PLACENTATION IN CHALCIDES CHALCIDES
dent in the lamina propria. A thin, bilaminar sheet of smooth muscle, consisting of an in- ner circular layer and an outer longitudinal layer, forms the remainder of the oviduct wall. Arterioles, venules, and larger blood vessels are embedded within and between the oviductal muscle layers. Externally, the ovi- duct is lined by the serosa.
Adjacent to the incubation chambers, the internal aspect of the oviduct projects lume- nally into ridges lined by simple and pseu- dostratified columnar epithelium (Fig. 1). Muscle tissue does not protrude into these ridges. The lamina propria core of the oviduc- tal ridges is composed of mucoid and sparse areolar connective tissue and contains prom- inent arterioles and venules that often fill the folds. Tangential sections through the lam- ina propria in this region show an extensive network of small vessels, which intercommu- nicates freely with vessels embedded in the underlying oviductal musculature.
Along the length of the uterine oviduct is attached an elongate bilaminar sheet of mes- entery that is continuous with the serosa (Fig. 1). Based on orientation of the fetuses, and by reference to other squamates, this sheet of tissue presumably represents the mesometrium (dorsal mesentery). The meso- metrium contains arteries, veins, and some of their tributaries, each of which communi- cates with vessels of the oviduct wall.
Topographical relationships Each fetus, surrounded by an amnion, lies
on its left side on the yolk sac. In the speci- mens examined, the yolk sac is small and occupies but a small area at the abembryonic pole. The chorion forms the outer boundary of the egg, except at the abembryonic pole, where it is continued by the omphalopleure. The allantois fills the exocoelom, and its outer (parietal) leaf lines the inner surface of the chorion, forming the chorioallantoic membrane. Apposition of the chorioallantoic membrane to the inner lining of the uterus constitutes the chorioallantoic placenta (al- lantoplacenta), Inner (visceral) and outer al- lantoic membranes typically lie in close con- tact at the circumferential periphery of the egg, with the allantoic cavity being corre- spondingly compressed or obliterated (Figs. 1,3). The amniotic cavity is greatly expanded in many areas, such that the amnion lines the inner surface of the visceral allantoic membrane, forming an amnioallantoic mem- brane (Figs. 1,3) . A single placentome occu- pies the mesometrial pole of the egg (Fig. 4).
The placentome is a specialized allantoplacen- tal region of thickened, interdigitating uter- ine and chorioallantoic tissues.
The omphalopleure lies a t the antimesome- trial pole, ventral to the yolk sac. I t is sepa- rated from the yolk sac by an extraembryonic coelomic cleft that is not continuous with the exocoelom proper (Fig. 5). As is discussed below, this space represents the “yolk cleft” (sensu Stewart and Blackburn, ’88). The width of this cleft varies; typically, it ranges between 39 and 55 pm, but in two specimens it is represented by a narrow gap of 2-4 pm. These differences may reflect preparation ar- tifacts. Only at occasional points are the yolk sac and omphalopleure in direct contact. The exocoelom and the yolk cleft lack continuity due to a thin septum of tissue passing radi- ally from the periphery of the yolk sac to the omphalopleure (Fig. 6). This septum of tis- sue restricts the allantois to the exocoelom proper and thus excludes the allantois from the abembryonic pole. Consequently, the al- lantois lies entirely dorsal and peripheral to the yolk sac in the abembryonic region and does not line the inner surface of the ompha- lopleure (Fig. 5).
Adnexal vascular connections The umbilical cord arises from the abdomi-
nal region of the fetus and splits into a dorsal branch and a ventral branch. The dorsal branch conveys the left umbilical artery and vein dorsally towards the placentome. These two blood vessels give rise to numerous smaller vessels that communicate with the placentome and the outer leaf of the allan- tois, thereby vascularizing the dorsal (embry- onic) pole of the chorionic vesicle. The ven- tral branch of the umbilical cord (Fig. 5) carries the right umbilical artery and vein, the vitelline vessels, the vitellointestinal duct, and in the specimens examined the allantoic stalk (cf. ten Cate-Hoedemaker, ’33). The right umbilical artery and vein give rise to large branches that accompany the parietal leaf of the allantois where it lies atop the yolk sac. These vascular branches pass radially to give rise to the vessels of the parietal allan- tois where it contributes to the chorion, pe- ripheral to the omphalopleure.
The vitelline artery and vein serve the yolk sac via an extensive capillary network. The vitelline vessels lie adjacent to the right um- bilical vessels within the umbilical cord, as well as distally, where the allantois and um- bilical vessels lie atop the yolk sac. No vascu- lar connections between the vitelline and al-
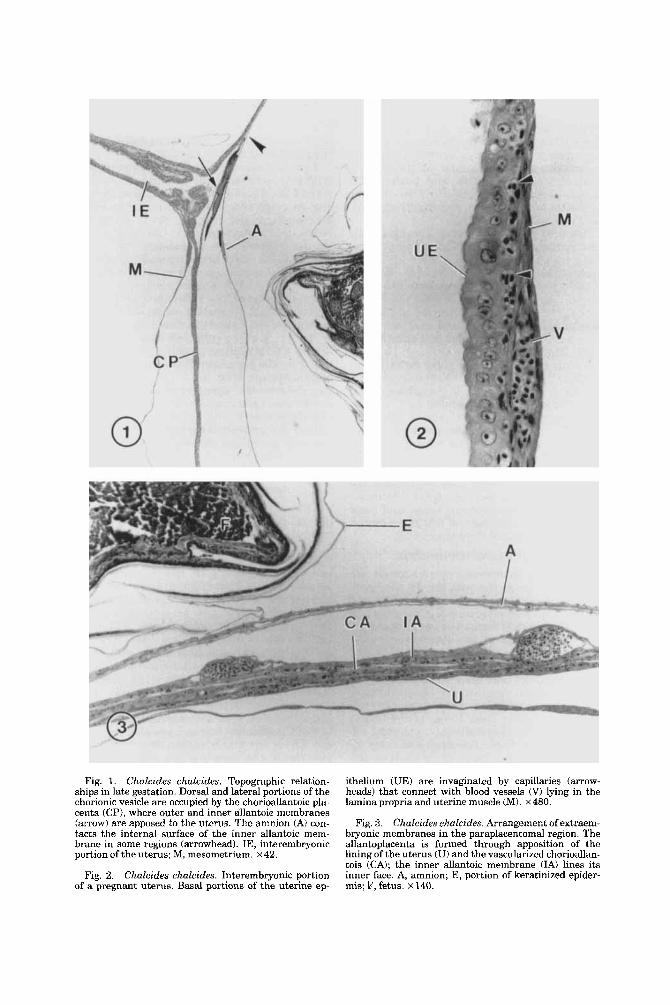
Fig. 1. Chalcides chulcides. Topographic relation- ships in late gestation. Dorsal and lateral portions of the chorionic vesicle are occupied by the chorioallantoic pla- centa (CP), where outer and inner allantoic membranes (arrow) are apposed to the uterus. The amnion (A) con- tacts the internal surface of the inner allantoic mem- brane in some regions (arrowhead). IE, interembryonic portion of the uterus; M, mesometrium. ~ 4 2 .
Fig. 2. Chalcides chalcides. Interembryonic portion of a pregnant uterus. Basal portions of the uterine ep-
ithelium WE) are invaginated by capillaries (arrow- heads) that connect with blood vessels (V) lying in the lamina propria and uterine muscle (M). ~480.
Fig. 3. Chuleides ehalcides. Arrangement of extraem- bryonic membranes in the paraplacentomal region. The allantoplacenta is formed through apposition of the lining of the uterus (U) and the vavcularized chorioallan- tois (CAI; the inner allantoic membrane (IA) lines its inner face. A, amnion; E, portion of keratinized epider- mis; F, fetus. ~140.
PLACENTATION IN CHALCIDES CHALCIDES
Fig. 4. Chakides ch.aZcides. Placentome at low magni- fication. The placentome (P) lies dorsal to the fetus, where chorioallantoic tissue (CA) interdigitates exten- sively with tissue of the uterus (U). IA, inner allantoic membrane; M, mesomctrium. x41.
lantoic circulations are present in these regions. However, the allantoic and vitelline circulations appear to comrnunicate at the periphery of the yolk sac, by means of vessels of the septum of tissue that passes between
183
Fig. 5. Chclcides cftalcides. Topographic relation- ships at the ahemhryonic pole in late gestation. The yolk sac (YS) is separated from the omphalopleure (0) by a yolkcleft (YC). UV, right umbilical vessels. x42.
the yolk sac and the omphalopleure (Fig. 6) . The vessels of this septum connect proxi- mally with capillaries on the external surface of the yolk sac and are identical in size and appearance to the yolk sac capillaries. Dis-

184 D.G. BLACKBURN
tally, the septa1 vessels appear to connect with adjacent allantoic vessels, although the presence of such a connection could not be established with certainty.
Yolk sac In transverse sections, the yolk sac appears
roughly triangular, with its longest axis lying along the abembryonic pole of the egg (Fig. 5). The yolk sac is composed chiefly of large, irregular cells (“yolk endoderm”) organized as a highly infolded sheet around an irregu- lar central lumen (Fig. 7). Examination of serial sections reveals this lumen to be contin- uous with the vitellointestinal duct but not with the yolk cleft or the exocoelom proper. Small amounts of yolk in the form of rounded particles or granules are occasionally evident in the yolk sac lumen as well as in the vitel- lointestinal duct. The yolk sac is bordered dorsolaterally by squamous mesothelial cells that line the exocoelom and ventrally by sim- ilar cells that line the yolk cleft.
The large yolk endoderm cells, which make up the bulk of the yolk sac, are polymorphic and range from rectangular to irregularly polygonal cells of -25-50 pm in maximum diameter (Fig. 7). These cells are intimately associated with basally situated capillaries and other small blood vessels of the yolk sac. The cells closely conform to the shape and tortuous course of the blood vessels; most of the cells are invaginated basally by the ves- sels, and many encircle the vessels com- pletely. The cells are polarized; their rounded apices project into the yolk sac lumen, and some cells appear to bear short, apical projec- tions. Adjacent yolk endoderm cells are linked laterally along their shared borders through thin intercellular bridges and, near their api- ces, via zonulae adherens. Cell nuclei are large and basophilic, with prominent nucle- oli. These nuclei frequently are lobed or irreg- ularly shaped, and some are pyknotic or show other evidence of degeneration. The cyto- plasm contains yolk material in the form of rounded inclusions that show affinity for both hematoxylin and iron hematoxylin. Another cytoplasmic component, probably represent- ing mitochondria, is apparent only in tissue stained with iron hematoxylin; these rod- shaped structures are abundant and appear to be of equivalent size.
Yolk sac placenta The yolk sac placenta of the late-stage Chal-
cides chalcides is formed by juxtaposition of the omphalopleure and uterine tissue at the
antimesometrial pole (Figs 5 , 8 ) . The ompha- lopleure is avascular; its nearest blood supply is the vitelline circulation of the yolk sac proper, from which the omphalopleure i s sep- arated by width of the yolk cleft.
The omphalopleure appears to be bilami- nar, being composed chiefly of two tissue layers: an external (stratified) epithelium and an underlying cellular layer (Fig. 9). A basal lamina, which stains strongly with iron hema- toxylin, separates the epithelium from the underlying layer. Although morphological de- tails were difficult t o discern, the deeper layer appears to consist largely of dorsoventrally flattened cells with elongate nuclei. In occa- sional regions, low cuboidal cells similar to the yolk endoderm cells were observed.
The stratified epithelium of the omphalo- pleure consists of an outer sublayer of small cuboidal to low columnar cells and an inner sublayer of squamous cells. Epithelial cell boundaries are indistinct, and the cuboidal and columnar cells are heavily vacuolated. The epithelium is thrown up into papillae that project into the uterine lumen (Figs. 8, 9). These papillae do not branch or intercon- nect, and range up to 53 pm in length. The papillae are smallest near the junction of the omphalopleure with the chorion, where the omphalopleuric epithelium is continuous with the chorionic epithelium.
Avery thin but distinct monolayer of squa- mow cells separates the omphalopleure from the yolk cleft (Figs. 8, 9). These flattened cells line the yolk cleft and thus lie on the inner (dorsal) surface of the omphalopleure, the ventral surface of the yolk sac (uide su- pra), and the ventral face of the connective tissue septum connecting the yolk sac to the omphalopleure. These cells are termed “intra- vitelline cells” based on their similarities to those of other squamates (see below). The intravitelline cells of the yolk cleft are histo- logically similar to the mesothelium lining the dorsolateral surfaces of the yolk sac.
Maternal tissues of the omphaloplacenta are unspecialized anatomically. The uterine epithelium consists of a monolayer of cuboi- dal cells and overlies a moderately vascular- ized lamina propria. Glands are not apparent in this region. Along the ventral border of the oviduct in the region of the omphaloplacenta, the bilaminar musculature forms a thick layer that is four to five times its thickness else- where in the uterus (Figs. 6,8).
Amass of detritus lies between the ompha- lopleure and the uterine epithelium at the antimesometrial pole (Fig. 8). The amount of
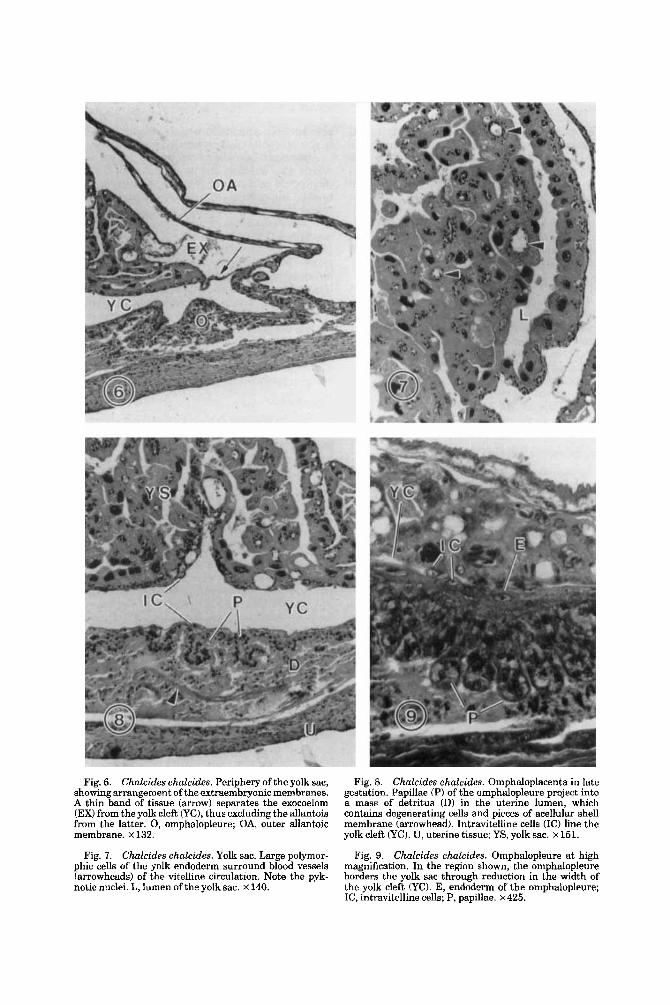
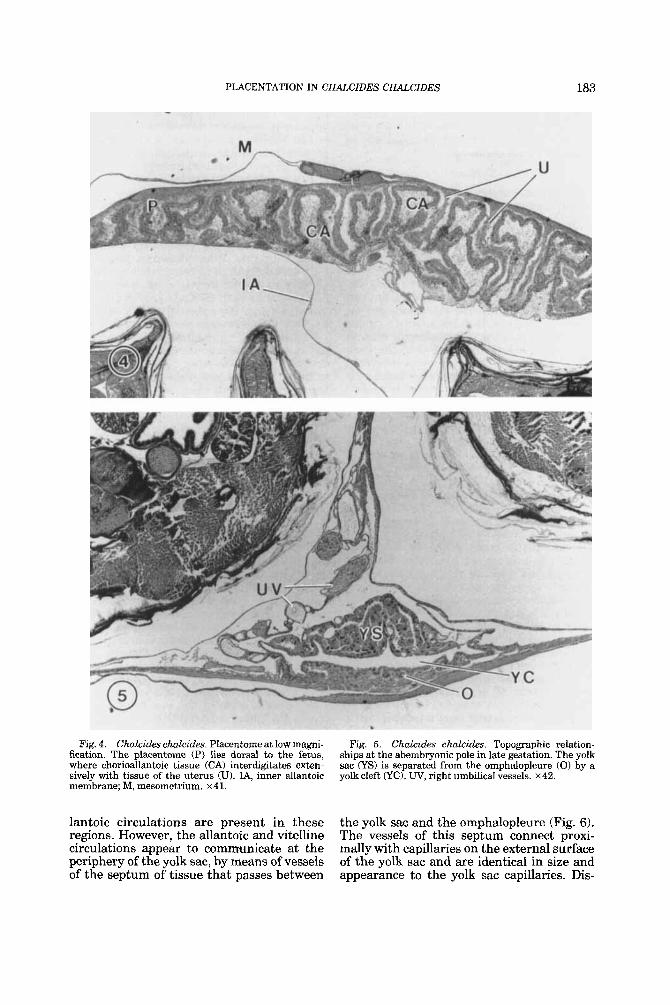
Fig. 6. Chalcides chalcides. Periphery of the yolk sac, showing arrangement of the extraembryonic membranes. A thin band of tissue (arrow) separates the exocoelom (EX) from the yolk cleft CyC), thus excluding the allantois from the latter. 0, omphalopleure; OA, outer allantoic membrane. x 132.
Fig. I. Chalcides chalcides. Yolk sac. Large polymor- phic cells of the yolk endoderm surround blood vessels (arrowheads) of the vitelline circulation. Note the pyk- notic nuclei. L, lumen of the yolk sac. x 140.
Fig. 8. Chalcides chalcides. Omphaloplacenta in late gestation. Papillae (P) of the omphalopleure project into a mass of detritus (D) in the uterine lumen, which contains degenerating cells and pieces of acellular shell membrane (arrowhead). Intravitelline cells (IC) line the yolk cleft (YC). U, uterine tissue; YS, yolk sac. x 151.
Chalcides chalcides. Omphalopleure at high magnification. In the region shown, the omphalopleure borders the yolk sac through reduction in the width of the yolk cleft (YO. E, endodcrm of the omphalopleure; IC, intravitelline cells; P, papillae. x 425.
Fig. 9.

186 D.G. BLACKBURN
this detritus varies between specimens. It consists of degenerating cells intermixed with pieces of acellular membrane. The cells of the detritus are similar in morphology to cells of the uterine epithelium, except that they are shrunken, contain pyknotic nuclei, and ap- pear to be in various stages of degeneration. The pieces of acellular membrane are ran- domly pleated and overlapped and stain mod- erately with hematoxylin and intensely with iron hematoxylin.
Chorioallantoic placenta The chorioallantoic placenta (allantopla-
centa) consists of the chorioallantoic mem- brane in apposition to the uterine epithe- lium. Two major regions can be distinguished: the placentome, which lies subadjacent to the attachment of the mesometrium, and the “paraplacentomal” (nonplacentomal) region, which forms the remainder of the allantopla- centa (Figs. 3, 4). In neither region is any trace of a shell membrane apparent between the uterine and chorionic epithelia. Paraplacentomal region
In the paraplacentomal region of the cho- rioallantoic placenta (Fig. lo), the inner sur- face of the chorioallantois typically is lined by both a vascularized inner (visceral) leaf of the allantoic membrane and the amnion (Figs. 3, 10). The outer surface of the chorioallantois and the apposed uterine tissue do not appear to be fused or interlocked, as indicated by the separation between them that is sometimes evident in the histological preparations. The maternal-fetal interface is smooth, lacking in ridges or folds. However, general contours of the uterus are followed by the chorioallan- tois, and the external surface of the chorion often conforms to slight protrusions or de- pressions in the uterine lining.
A major distinguishing characteristic of the paraplacentomal region of the allantopla- centa is the attenuated condition of the ma- ternal and fetal tissue layers (Figs. 10, ll). The uterine epithelium forms a thin but con- tinuous layer of simple squamous cells over- lying the abundant uterine capillaries. The lamina propria and uterine muscle are also very thin and, consequently, are seldom visi- ble as discrete layers. Nonetheless, presence of both components is readily apparent in regions where tissues have become separated during sectioning. As with the uterine epithe- lium, the chorionic epithelium is represented by a monolayer of squamous cells, although occasional regions of low cuboidal cells are
present. No apical (lumenal) specializations of the epithelial cell membranes are visible.
Abundant allantoic capillaries bulge to- wards the surface of the chorion, typically displacing portions of the epithelial cells lat- eraIIy. Superficial to the capillaries of the outer allantoic membrane, chorionic epithe- lial cells remain in contact via extremely thin processes (Fig. 11)) which can be difficult to discern except under optimal viewing condi- tions. Nevertheless, a continuous layer of chorionic epithelial cells appears to separate the underlying allantoic tissues and capillary endothelium from the uterine lumen and maternal tissue. A layer of allantoic mesen- chyme lies beneath the chorionic epithelium. In most areas, this connective tissue layer is very thin; however, surroundmg the larger allantoic blood vessels, the mesenchyme forms a moderately thick layer. Placentome
The placentome is a specialized allantopla- cental region of hypertrophy, hyperplasia, and interdigitation. Uterine and chorionic tissues are closely associated and often inter- locked, though not fused. The endometrium forms branching folds or outgrowths that protrude into deep invaginations into the chorioallantoic tissue of the placentome (Fig. 4). In dorsal (frontal) sections, the endome- trial folds appear as thin, interconnecting ridges that enclose, and occasionally are sur- rounded by, chorioallantoic tissue (Fig. 12).
The endometrial folds contain uterine epi- thelium and small blood vessels but not uter- ine muscle (Fig. 13, 14). The uterine epithe- lium is formed by columnar, cuboidal, and polymorphic cells that range from 15 to 40 p,m in height and 17-30 pm in width. The cells appear to bear short, apical microvilli. Adjacent epithelial cells are linked laterally at their apices via tight junctions but are often separated by intercellular spaces basal to these junctional complexes. Cell bound- aries are distinct, and no syncytial areas are apparent. Epithelial cell nuclei are large, round or oval structures and are located cen- trally in the cells. The nuclei are pale stain- ing, with little condensed chromatin, and each contains a prominent nucleolus. The uterine epithelium of the placentome rests on a thin lamina propria, which contains abundant blood vessels.
Epithelial cells of the chorion are large and columnar, and occasionally polymorphic, and appear to bear microvilli (Figs. 14, 15). Cyto- plasm of the cells is eosinophilic and faintly
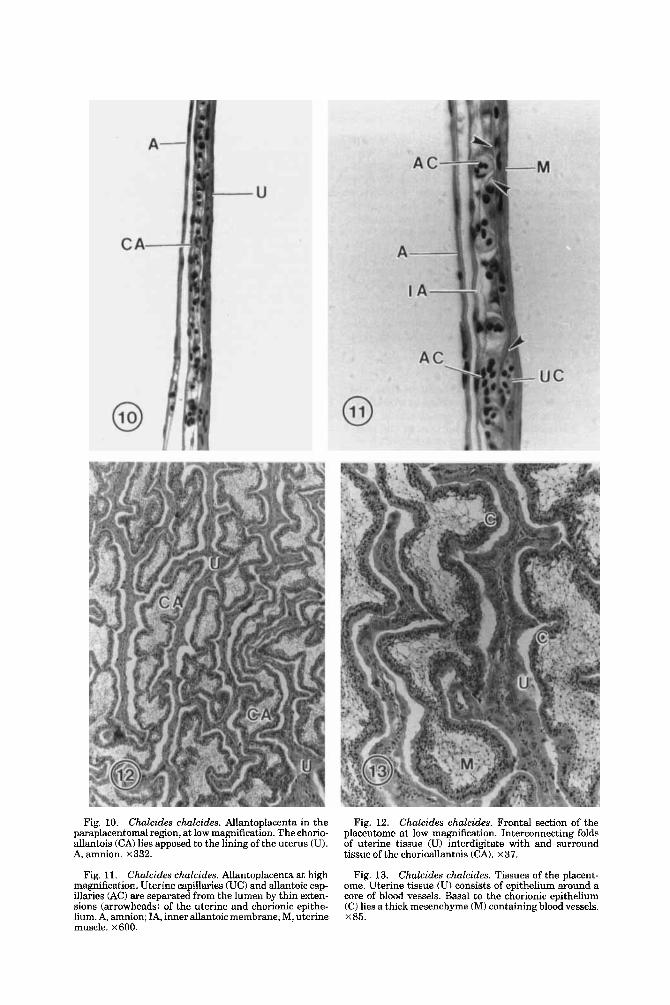
Fig. 10. Chalcides chalcides. Allantoplacenta in the paraplacentomal region, at low magnification. The chorio- allantois (CA) lies apposed to the lining of the uterus (U). A, amnion. X332.
Fig. 11. Chalcides chalcides. Allantoplacenta at high magnification. Uterine capillaries (UC) and allantoic cap- illaries (Act are separated from the lumen by thin exten- sions (arrowheads) of the uterine and chorionic epithe- lium. A, amnion; IA, inner allantoic membrane; M, uterinc muscle. ~ 6 0 0 .
Fig. 12. Chalcides chalcides. Frontal section of the placentome at low magnification. Interconnecting folds of uterine tissue (U) interdigitate with and surround tissue of the chorioallantois (CAI. x37.
Chalcides chalcides. Tissues of the placent- ome. Uterine tissue iU) consists of epitheIium around a core of blood vessels. Basal to the chorionic epithelium (C) lies a thick mesenchyme (MI containing blood vessels. x85.
Fig. 13.

188 D.G. BLACKBURN
granular. Each cell contains a round or oval nucleus with a prominent nucleolus. Occa- sional cells contain two such nuclei. Deep to the chorionic epithelium lies a mass of allan- toic connective tissue (Fig. 13,141. The tissue is mesenchymal to mucoid and contains an abundance of blood vessels. In the interstices of the placentome are individual uterine epi- thelial cells that lie separated from the endo- metrium (Fig. 15). Such cells typically are not in contact with the lamina propria and very occasionally lie adjacent to regions where the lamina propria is exposed to the uterine lumen through epithelial loss. Such regions are small, and in no area were uterine capil- laries observed to be exposed to the lumen or to lie apposed to chorionic tissues.
Postpartum placentome The one postpartum placentome that was
examined histologically contains extraembry- onic tissue as well as maternal tissue (Fig. 16). The extraembryonic portion of the plac- entome is largely intact and consists of chori- onic epithelium, allantoic connective tissue, and allantoic vessels filled with erythrocytes. The chorionic tissue is well preserved and shows no indication of shrinkage or cell de- generation. The nuclei of the chorionic epithe- lium are rounded and contain both euchroma- tin and heterochromatin, and the cytoplasm of most cells exhibits normal staining proper- ties.
Remnants of uterine tissue and cell debris lie trapped in the interstices of the folds of chorionic tissue (Fig. 17). The uterine tissue includes single epithelial cells, clumps of epi- thelial cells attached along their lateral bor- ders, and portions of uterine folds consisting of epithelium and basal connective tissue. Few maternal erythrocytes and no uterine blood vessels are evident. Although occa- sional chunks of uterine tissue present a healthy appearance, most of the uterine tis- sue appears necrotic or otherwise abnormal. Uterine epithelial cells typically are shrunken and misshapen, with pyknotic nuclei and densely staining cytoplasm. Many of these cells are broken and appear to be the source of the abundant cellular debris that is pre- sent. Although the pieces of uterine tissue invariably lie between folds of chorionic tis- sue, they are not directly attached to the adjacent chorionic epithelial cells (Fig. 17).
DISCUSSION General observations
The Chalcides chalcides conceptuses in the study series appear to be at equivalent states
of development. Based on integumentary and osteological data on other lizards, (e.g., Du- faure and Hubert, '61; Hubert, '85; Peter, '04; Raynaud, ,851, I judge the fetuses to be in an advanced developmental stage, repre- senting Dufaure and Hubert ('61) ("D&H") stage 40, or possibly stage 39. Given that D&H stage 40 is the stage at parturition, this assessment is consistent with ten Cate- Hoedemaker's ('33, p. 318) report that some of the lizards gave birth before they could be sacrificed. The fact that the yolk sacs have not been withdrawn into the fetal abdominal cavities (Fig. 5) does not conflict with the inference that the fetuses are immediately prepartum. Unlike the situation in many live- bearing lizards (see Weekes, '351, the yolk sac in C. chalcides reportedly remains external until after birth, and in fact is said to provide the neonate with its first meal (ten Cate- Hoedemaker, '33, pp. 318, 336). In this re- spect, C. chalcides is similar to its congener C. ocellatus (Badir, '55; personal observa- tion). Unrelated viviparous lizards that are born with an external yolk sac include the lacertid Lacerta vivipara (Hrabowski, '271, the gekkonid Hoplodactylus maculatus (Boyd, '421, and the anguid Gerrhonotus coeruleus (Stewart and Castillo, '84).
Histology of the uterus in C. chalcides conforms to the general squamate pattern (see Giacomini, 1893; Fox, '77). The overall topographical relationships between the con- ceptus and the uterus are typical of vivipa- rous squamates, with the omphaloplacenta lying at the antimesometrial pole and with the chorioallantoic placenta contributing to the remainder of the maternal-fetal inter- face. In addition, the placentome of C. chal- ctdes lies subadjacent to the dorsal attach- ment of the mesometrium, as in those few other squamates in which placentomes have been observed (see Weekes, '30; Blackburn et al., '84).
Chalcides chalcides is similar to other live- bearing reptiles in that the portions of the oviduct that contain conceptuses are greatly distended relative to the intervening seg- ments. Weekes ('35, p. 627) raised the ques- tion of whether an active constriction of the uterus between conceptuses was involved. If the interembryonic portions of uterus ex- hibit their normal (i.e., nonpregnant) thick- ness, as ten Cate-Hoedemaker ('33) indicated was true of early development, the difference in oviductal diameters between incubation chambers and intervening segments would represent distension of the former rather than tonic constriction of the latter. This
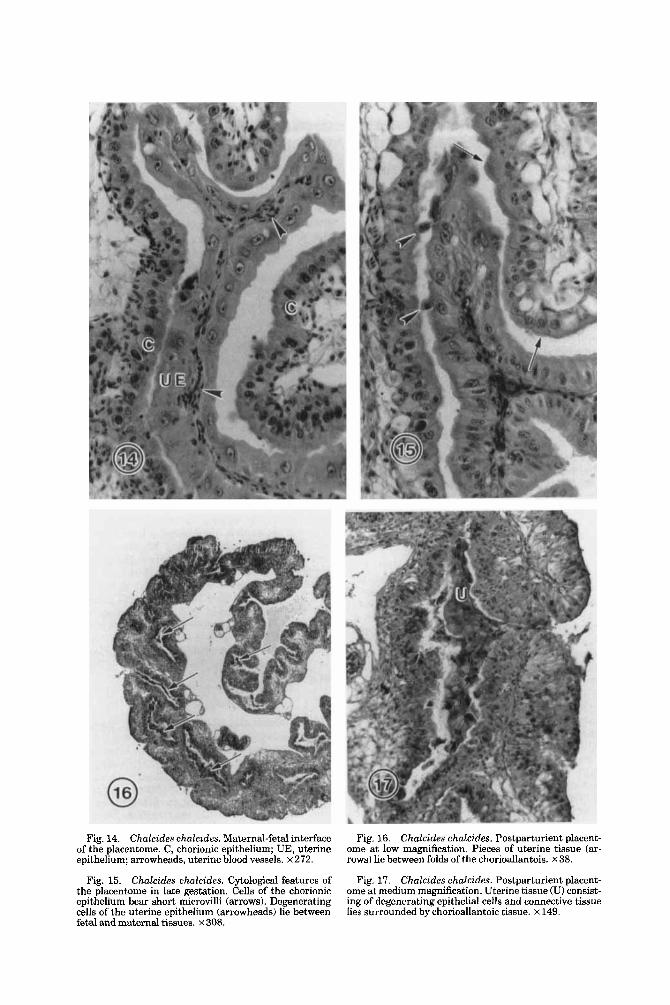
Fig. 14. Chalcides chalcides. Maternal-fetal interface Fig. 16. Chalcides chalcides. Postparturient placent- ome at low magnification. Pieces of uterine tissue (ar- rows) lie between folds of the chorioallantois. x 38.
of the placentome. C, chorionic epithelium; UE, uterine epithelium; arrowheads, uterine blood vessels. x 272.
Fig. 15. Chalcides ehalcides. Cytological features of Fig. 17. Chalcides chalcides. Postparturient placent- ome at medium magnification. Uterine tissue (U) consist- ing of degenerating epithelial cells and connective tissue lies surrounded by chorioallantoic tissue. x 149.
the placentome in late gestation. Cells of the chorionic epithelium bear short microvilli (arrows). Degenerating cells of the uterine epithelium (arrowheads) lie between fetal and maternal tissues. x 308.

190 D.G. BLACKBURN
inference is supported by Giacomini’s (1891) observation that former incubation charn- bers are still apparent in this species 3 months after parturition.
Shell membrane Unlike the condition in many viviparous
squamates (Jacobi, ’36; Panigel, ’51; Hoff- man, ’70; Stewart, ’851, in late-stage C. chal- cides no shell membrane is visible histologi- cally around the conceptus. Absence of intact remnants of shell membrane may require confirmation via electron microscopy; shell membranes become exceedingly thin during gestation in some viviparous squamates, and persistent traces can be difficult to discern histologically in suboptimally fixed tissues (Hoffman, ’70; Blackburn, ’93).
However, the shell membrane of late-stage C. chalcides appears to be represented by the pieces of acellular membrane lying in the oviductal lumen, ventral to the omphalo- pleure (Fig. 8). These remnants are ran- domly intermixed with other detritus along the ventral aspect of the incubation chamber, where they presumably have accumulated after being shed from the egg. Such an onto- genetic loss of the shell membrane commonly has been reported in other viviparous squa- mates (e.g., see Weekes, ’27, ’35; Boyd, ’42; Kasturirangan, ’51; Guillette and Jones, ’85). The identity of the membrane fragments in C. chalcides may be open to question, for Giacomini (1891) opined that tertiary egg investments were not deposited in this spe- cies. He interpreted the detrital membrane as the remains of the vitelline membrane, a fibrous covering of the ovum that he pre- sumed to be secreted by the ovarian follicular epithelium. In contrast, Weekes (’27, p. 507) inferred from Giacomini’s account that the membrane of the detritus was in fact the shell membrane. Ten Cate-Hoedemaker (’33) neither identified nor described the detrital membrane, simply referring to it in figure captions (pp. 330,332) as the “Eimembran,” a term he used indiscriminantly for both secondary and tertiary investments of the ovum (ten Cate-Hoedemaker, ’33, p. 317).
I consider the membrane fragments of the detritus to be remnants of the shell mem- brane rather than the vitelline membrane for several reasons. 1) The membrane is similar in appearance to the shell membrane of other viviparous squamates; 2) egg investments of ovarian origin are far thinner than the frag- ments in question and are not known to be shed in such a fashion; 3) shell membranes
invariably are deposited in squamates, even in viviparous species with much smaller ova and a degree of matrotrophy that exceeds that of C. chalcides (Blackburn et al., ’84; Blackburn, ’93); and 4) shell membranes in viviparous squamates frequently are shed and come to occupy the abembryonic region of the oviductal lumen, where they can contrib- ute to a thick pad of detritus (Weekes, ’29, ’30, ’35; Yaron, ’85). Ongoing work on C. chakides confirms that a shell membrane is in fact deposited early in development (Black- burn, ’88).
Adnexal uasculature In C. chalcides, as in other squamates, the
umbilical and vitelline circulatory systems serve the allantois and yolk sac, respectively. This species may be unusual, however, in that the two circulatory systems apparently communicate through capillaries of the con- nective tissue septum that passes from the yolk sac to the omphalopleure. Direct commu- nication between vitelline and allantoic circu- lations has seldom been described in other squamates (Giacomini, 1892; Hrabowski, ’26). The septal capillaries in C. chalcides connect proximally with the vitelline vascu- lar network and appear to communicate dis- tally with branches of adjacent allantoic ves- sels. Ten Cate-Hoedemaker (’33) suggested that the capillaries carry blood from the allan- toic vessels to the vitelline circulation. This hypothesis requires experimental verifica- tion, in that blood flow in the opposite direc- tion cannot be ruled out on histological grounds. The developmental origin of the septal capillaries also warrants study, al- though, from their morphology and position, the vessels appear to be derived from the vitelline system.
Chorioallantoic placenta In the chorioallantoic placenta of C. chal-
cides, two regions can be distinguished: the placentome and the paraplacentomal region. In response to Giacornini’s (1891) designa- tion of only the former as placental, ten Cate- Hoedemaker (’33) pointed out that the entire allantochorion participates in placentation. Deferring to usage in the mammalian litera- ture, ten Cate-Hoedemaker (’33) reluctantly denoted the nonplacentomal region as “Para- plazentargebiet.” I use the term “paraplacen- tomal region,” which avoids unfortunate functional connotations of the adjective “paraplacental. ”

PLACENTATION IN CIIALCIDES CHALCIRES 191
Paraplacentomal region The paraplacentomal portion of the allan-
toplacenta consists of a relatively unspecial- ized chorioallantoic membrane in apposition to uterine tissue (Fig. 10). The apposition of outer and inner allantoic membranes offers a double set of vascularized tissues on the fetal side of the placenta. Neither active nor quies- cent uterine glands are apparent, nor are elaborate specializations for histotrophic transfer such as are found in some other matrotrophic skinks (Blackburn and Vitt, ’92 j . Rather, the paraplacentomal allantopla- centa presents the appearance of highly vas- cularized membranes in close proximity. Be- cause no shell membrane is visible between maternal and extraembryonic tissues, uter- ine and chorionic epithelia appear to be in direct contact. However, the uterine and ex- traembryonic tissues probably are not fused or interlocked, as indicated by regions of arti- factual separation. Furthermore, the lu- menal surfaces of the chorion and uterus arc nearly smooth and, histologically at least, show no “protoplasmic processes” such as have occasionally been described (Weekes, ’35) in other squamates.
Typically, the epithelial cells are greatly thinned over the abundant allantoic and uter- ine blood vessels (Fig. 11); in fact, the nuclei and most of the cytoplasm of these epithelial cells lie adjacent to the capillaries. Although occasional regions of cuboidal chorionic and uterine epithelium are evident, no pattern was apparent in the position of such regions. A lamina propria is seldom visible histologi- cally between uterine epithelium and the un- derlying capillary endothelium, and allantoic connective tissue is not apparent between the allantoic capillaries and the chorionic epithelium. Thus, at a minimum, the follow- ing tissue layers separate fetal and maternal blood streams: fetal capillary endothelium, a thin chorionic epithelium, a thin uterine epi- thelium, and maternal capillary endothe- lium.
Applying the Grosser (’27) system to C. chalcides, the paraplacentomal allantopla- centa would be classified as “epitheliochor- id.” I have been unable to locate any areas of true (‘chorionic erosion” or “endothelial- endothelial contact” as described by ten Cate- Hoedemaker (’33). Although the epithelium is highly attenuated on maternal and fetal sides of the paraplacentomal region, careful examination at high magnification reveals thin extensions of epithelial cells extending
over the allantoic and uterine capillaries. My observations also do not support previous suggestions (ten Cate-Hoedemaker (’33, pp. 326,328) that uterine and chorionic epithelia are syncytial or bear a striated border. Placentome
Both grossly and microscopically, the plac- entome is a highly specialized region of the chorioallantoic placenta. At the gross level, the placentome appears as an opaque region at the dorsal pole (Ghiara et al., ’871, al- though in preserved specimens this region may be difficult to distinguish (personal obser- vation). As frontal sections reveal, the uter- ine endometrium is hypertrophied into elon- gate, interconnecting folds that interdigitate extensively with chorionic tissue (Fig. 12). The epithelial cells of the chorion and the uterus are enormously enlarged compared to their counterparts in the paraplacentomal region. The chorionic diplokaryocytes noted by ten Cate-Hoedemaker (’33) are randomly distributed and are too rare to be regarded as a major structural component of the placent- ome, unlike the case in certain Brazilian Mubuyu (Blackburn and Vitt, ‘92; Blackburn, ’93). Ten Cate-Hodemaker (’33) described and illustrated an apical 44Stabchensaum” (literally, a “border of little rods”) on the apical surfaces of the uterine and chorionic epithelial cells, which Kasturirangan (’51) tentatively interpreted 8s a mammillate (mi- crovillous) border. This study supports the presence of microvilli on the uterine epithe- lium and is consistent with the report (Ghi- ara et al., ’87) of microvilli on the chorionic epithelium based on ultrastructural observa- tion.
Ten Cate-Hoedemaker (’33, p. 327) de- scribed occasional sites where maternal capil- laries become exposed near the end of preg- nancy through degeneration of areas of uterine epithelium. On this basis, he classi- fied the placentome as transitional between an epitheliochorial and a syndesmochorial condition. My observations agree with Luck- ett’s (’77a) characterization of the placen- tome as “epitheliochorial.” Although areas where individual uterine epithelial cells have been shed into the lumen are evident (see, e.g., Fig. 151, I have been unable to locate sites where capillaries or more than a small portion of the lamina propria are exposed through epithelial degeneration. Interpreta- tion may be complicated by the fact that sectioning and fixation artifacts are abun- dant in the available placentorne specimens.

192 D.G. BLACKBURN
Nevertheless, I find it difficult to account for ten Cate-Hoedemaker’s description of large sites of epithelial degeneration other than to speculate that sections he described were not included in the study series and that such degeneration occurs just prior to birth (see below).
The interdigitation of fetal and maternal tissues a t the placentome is sufficiently exten- sive that a ready separation at parturition would seem difficult to achieve. The fact that the one postpartum placentome examined includes substantial chunks of endometrium suggests that the maternal tissue is decidu- ate, a t least to some extent. Further evidence for a deciduate placentome is provided by the observation that the postpartum uterus com- pletely lacks endometrial folds (i.e., “mutter- licher Septa”; ten Cate-Hoedemaker, ’33, p. 338). Giacomini’s (1891) contrary report, that fetal and maternal tissues do not closely inter- lock and are easily separable, possibly charac- terizes developmental stages preceding those examined herein.
Whether the endometrium is physically torn from the uterus a t birth or undergoes a prepartum degeneration that expedites sepa- ration from the uterus is not known. Ten Cate-Hoedemaker (’33) inferred the latter, based on the apparent degeneration of por- tions of maternal endometrial villi in prepar- turient specimens. Although I have been un- able to corroborate a prepartum degeneration of endometrium that extends beyond loss of individual uterine cells, the fact that the en- dometrium of the postpartum placentome tends to be deteriorated compared to chori- onic tissues (Fig. 17) could reflect a degenera- tion that began prior to parturition. Adegree of prepartum degeneration of decidual tissue might also account for Giacomini’s (1891) description of a cellular coagulum in the inter- stices of the placentome.
Yolk sac placeiita The yolk sac placenta of C. chalcides is best
understood in the context of omphalopleure morphology in other squamate reptiles, and the complexity and unique nature of the squa- mate yolk sac requires detailed consider- ation. The yolk sac of squamates differs strik- ingly from that of all other vertebrates, and a similar developmental pattern has been dis- covered in all lizards and snakes that have been described in detail (Yaron, ’85; Stewart and Blackburn, ’88). Detailed diagrams have been presented recently by Stewart (’85, ’92) and Stewart and Blackburn (’88).
The squamate omphalopleure is a special- ized, avascular membrane that forms at the abembryonic (ventral) pole of the egg. The omphalopleure consists of two tissue layers: 1) an external (ventral) layer of epithelium, which can be stratified, and 2) an internal cellular layer that is associated developmen- tally with the “isolated yolk mass.” The lat- ter becomes separated from the yolk sac proper through invasion of intravitelline cells into the yolk and the associated formation of a yolk cleft, a cavity that often becomes con- tinuous with the exocoelom proper. The intra- vitelline cells usually surround the yolk cleft (cf. Boyd, ’42; Stewart, ’851, and thus line the ornphalopleure on its inner (dorsal) face. De- velopmental origins of the cellular layers of the ornphalopleure have not been resolved definitively (Yaron, ’85; Stewart, ’85). How- ever, most recent researchers have concluded that the omphalopleure is developmentally bilaminar, the epithelium being ectodermal and the isolated yolk mass layer being endo- dermal (Hoffman, ’70; Ibrahim, ’77; Baxter, ’87; Stewart, ’90). The intravitelline cells are widely regarded as mesodermal in origin (Bax- ter, ’87; Stewart and Blackburn, ’88; cf. Moss- man, ’87).
The isolated yolk mass of squamates can regress during development, in a pattern that varies taxonomically (Hoffman, ’70; Yaron, ’85). The fate of the yolk cleft also varies taxonomically and occasionally even intraspe- cifically. In some squamates, the yolk cleft becomes continuous with the exocoelom proper and is completely invaded by the allan- tois (Weekes, ’35; Hoffman, ’70; Yaron, ’85; Stewart, ’901, forming an omphalallantoic membrane (terminology of Stewart and Blackburn, ’88). However, in other squa- mates, the yolk cleft does not become conflu- ent with the exocoelom proper and hence is never penetrated by the allantois (Weekes, ’29, ’35; Boyd, ’42; Stewart, ’85).
Terminology applied to the various types of yolk sac placentae has now been standard- ized (Stewart and Blackburn, ’88; Stewart, ’90). In viviparous squamates, the omphalo- pleure and isolated yolk mass, in apposition to uterine tissue, constitute the “omphalopla- centa” (sensu stricto), whereas the “choriovi- telline placenta” is formed by apposition of uterine tissue to the trilaminar choriovitel- line membrane. Yet another placental type forms in those squamates in which the allan- tois invades the yolk cleft, for which the term “omphalallantoic placenta” is employed. Un-

PLACENTATION IN CHALCIDES CHALCZDES 193
like the choriovitelline placenta, which devel- ops in many therian mammals (Luckett, '77b; Mossman, ,871, the omphaloplacenta and om- phalallantoic placenta have no dose counter- part among mammals (Stewart and Black- burn, '88).
Some authors have expressed uncertainty or doubt about whether yolk sac develop- ment in C. chalcides conforms to the squa- mate pattern. For example, Weekes ('27) in- ferred from Giacomini's account t ha t mesoderm never invades the yolk, implying that intravitelline cells, an isolated yolk mass, and a bilaminar omphalopleure do not form. Similarly, ten Cate-Hoedemaker ('33) raised the question of whether the exocoelom ever penetrates the yolk or, alternatively, whether a "Dottersacknabelblaschen" develops, such as Hrabowski ('26) described for Lacerta uiuipara. In contrast, Boyd ('42) surmised from ten Cate-Hoedemaker's ('33) paper that a yolk cleft forms and then regresses periph- erally, as in the gekkonid Hoplodactylus mac- ulatus.
My observations suggest that C. chalcides is fundamentally similar to most other squa- mates with regard to morphology of the om- phalopleure. The exocoelomic cavity separat- ing the omphalopleure from the yolk sac clearly represents the yolk cleft (Figs. 5 , 8). Accordingly, exclusion of the allantois from this cleft is similar to the condition described and illustrated in Gerrhonotus coeruleus (Stewart, '85) and some specimens of Eger- nia cunninghami 7Weekes, '35). As in other squamates, the attenuated cells lining the yolk cleft would constitute intravitelline cells (Fig. 8). The identity and derivation of these cells from mesoderm are supported by the strong resemblance that the cells bear to intravitelline cells of other squamates as well as to the mesothelium lining the yolk sac.
The mature omphalopleure of C. chalcides consists of a stratified layer of epithelium overlying a layer of flattened, elongate cells. The conspicuous basal lamina separating these two cell populations identifies the strat- ified layer as epithelial, as in Tharnnophis sirtalis (Hoffman, '70; personal observation). By reference to other squamates, the under- lying cells are almost certainly endodermal cells that became separated from the yolk sac proper by the yolk cleft and intravitelline mesoderm. Although the germ layer deriva- tion of this omphalopleuric component war- rants confirmation, as in other squamates (see Yaron, '85), the fact that the deep layer
of the omphalopleure contains cells that are very similar to the distinctive yolk endoder- ma1 cells (ten Cate-Hoedemaker, '33) sup- ports an endodermal origin for this layer. The isolated yolk mass clearly has regressed in these late-stage C. chalcides, as is common among viviparous lizards (Weekes, '29; Hoff- man, '70).
Contrary to the inferences of Weekes ('271, Chalcides chalcides appears to be like certain other viviparous squamates in possessing a true omphaloplacenta in late development. As in some (if not all) other squamates, the omphalopleure appears to be developmen- tally bilaminar, consisting of a stratified layer of epithelial (ectodermal) cells and an under- lying layer derived from endoderm. Due to the absence of a choriovitelline membrane, a true choriovitelline placenta is not present in late gestation. Likewise, because the allan- tois is excluded from the yolk cleft through late development, an omphalallantoic pla- centa does not form.
These interpretations are consistent with recent, brief accounts of early-stage C. chal- cides. From his observations on early develop- mental stages, Luckett ('77b) diagrammed a yolk cleft, which he tentatively regarded as being discontinuous with the exocoelom proper, an interpretation supported with my own observations on midstage embryos (Blackburn, '88). My observations on the ma- ture yolk sac indicate that the relationship Luckett described persists until late gesta- tion. Luckett ('77a) also noted that a true choriovitelline placenta is present in C. chal- cides between the 15 somite and the 50 so- mite stages. Although a choriovitelline pla- centa was not observed in the present study, its absence most probably reflects the devel- opmental loss of this placenta in association with the expansion of the exocoelom (see Stewart and Blackburn, '88).
The ontogeny of the yolk sac and omphalo- pleure in C. chalcides deserves detailed exam- ination, .and, a t present, morphology of the late-stage omphalopleure ought not to be as- sumed to typify earlier development. Giaco- mini (1891) described an interlacing of om- phalopleuric and uterine folds that was not observed in the present study, and ten Cate- Hoedernaker ('33) inferred that the peripar- turn yolk sac placenta is actually in a stage of regression. Other features that deserve atten- tion include the ultrastructure and germ layer composition of the omphalopleure, the fate of the isolated yolk mass, the ontogeny of the

194 D.G. BLACKBURN
yolk cleft, and the developmental origin of the septum that separates the yolk cleft from the exocoelom.
Comparisons to other squamates Placentation in C. chalcides has long been
regarded as the most complex discovered among reptiles (Weekes, '30). This study con- firms the presence of several highly unusual features, some of which may be unique to this skink genus. For example, the placent- ome is a t least partially deciduate; it exhibits an extensive interdigitation of villous endo- metrial and chorionic tissues; and its chori- onic and uterine epithelia consist entirely of enlarged, microvilliated cells, among which only occasional diplokaryocytes occur. In these respects, C. chalcides differs from all other squamates described, including those with complex placentomes (see Weekes, '30; Blackburn and Vitt, '92). Although the cho- rioallantoic placenta of C. chalcides bears a superficial resemblance to that of certain ma- trotrophic lizards of the genus Mabuya, ma- jor differences are apparent at gross, histolog- ical, and cytological levels (Blackburn, '92, '93). C. chalcides differs from these Mdbuya in lacking such specializations as paraplacen- tomal chorionic areolae and abundant pla- centomal diplokaryocytes, and through the presence of a villous deciduate placenta corre- sponding to Weekes' ('35) "type iii" category. The presence of omphalopleuric papillae in C. chalcides may also unique; these papillae do not correspond to the descriptions of un- usual abembryonic features in the lizard Lac- erta uiuipara (Hrabowski, '26).
Other features of C. chakides are unusual but by no means unique to this species. For example, the exclusion of the allantois from the yolk cleft, with the consequent absence of an omphalallantoic membrane, is atypical but has been documented in least three other lizard genera (see Weekes, '35; Boyd, '42; Stewart, '85). The presence of a placentome is also rare, having been demonstrated only in a few distantly related skinks of Australia (Weekes, '30, '35) and South America (Black- burn et al., '84; Blackburn and Vitt, '92). Whether a choriovitelline placenta is rare in viviparous reptiles is not known; although seldom described in squamates (Stewart, '85, 'go), this membrane may simply have been overlooked due to its ephemeral existence (Stewart and Blackburn, '88).
Despite the presence of impressive cytolog- ical specializations, C . chalcides appears to be fundamentally similar to other squamates
in terms of other aspects of placental mor- phology and, presumably, the associated mor- phogenetic patterns. Topographic relation- ships within the egg are essentially like those of other squamates. Likewise, as in other viviparous squamates for which definitive in- formation is available (Blackburn et al., '92; Blackburn, '931, the (nonplacentomal) allan- toplacenta is epitheliochorial, lacks exposed capillaries, and lacks syncytial and microvilli- ated epithelia, all contrary to previous sugges- tions (ten Cate-Hoedemaker, '33). The mor- phology of the yolk sac and omphalopleure in C. chalcides also appears to agree in impor- tant respects with that of typical squamates (e.g., see Stewart and Blackburn, '881, ovipa- rous and viviparous alike. The presence of an avascular omphalopleure, as well as the ab- sence of a choriovitelline membrane in late- stage gestation, is common to all squamates described in detail. The absence of an omphal- allantoic membrane, although not commonly described in squamates, reflects a fairly mi- nor specialization, i.e., a lack of confluence of the exocoelom and the yolk cleft.
Consequently, the placental membranes of C. chalcides &Rer from those of other vivipa- rous reptiles primarily through modification of a condition that is typical for the suborder Squamata. Fetal nutrition in this species is highly matrotrophic (see Ghiara et al., ,871, and, given that matrotrophy in viviparous amniotes entails placentotrophy (Blackburn, '92), morphological specializations of the pla- cental membranes most likely reflect their role in maternal-fetal nutrient transfer. The functional meaning, adaptive significance, and evolutionary history of the placental spe- cializations of this matrotrophic skink and its relatives are promising subjects for future investigations.
ACKNOWLEDGMENTS I gratefully acknowledge the curators and
staff at the Hubrecht Laboratorium in Utrecht for making available histological ma- terial under their care. I thank Ari van Tien- hoven for logistical support, Donald B. Gal- braith for technical assistance, and James R. Stewart for reading the manuscript. Final stages of this work were supported by faculty research grants, equipment grants, and a research Ieave from Trinity College.
LITERATURE CITED Badir, N. (1955) Beitrag zur Fortpflanzungsbiologie der
lebendgebarenden Eidechse Chalczdes ocellatus. Ge- burtshilfe und Frauenheilkunde. Ergebn. Forseh. 15. 746-749

PLACENTATION IN CHAtCILlES CHALCIDES 195
Baxter, D.C. (1987) Placentation in the Viviparous Lined Snake, Tropidoclonion lineatum: Ontogeny of the Ex- traembryonic Membranes and Histochemistry of Pla- cental Tissues. MSc Thesis, University of Tulsa, Tulsa, Oklahoma.
Blackburn, D.G. (1988) Late-stage placentation in the matrotrophic viviparous lizard Chalcides chalcides. Am. Zool. 28r196A (abstract).
Blackburn, D.G. (1992) Convergent evolution of vivipar- ity, matrotrophy, and specializations for fetal nutrition in reptiles and other vertebrates. Am. Zool. 3 2 3 1 3 3 2 1 .
Blackburn, D.G. (1993) Chorioallantoic placentation in squamate reptiles: Structure, function, development, and evolution. J . Exp. Zool. (in press).
Blackburn, D.G., and L.J. Vitt (1992) Reproduction in South American lizards of the genus Mabuya. In W. Hamlett (ed.): Reproductive Biology of South Ameri- can Vertebrates: Aquatic and Terrestrial. New York: Springer-Vcrlag, pp. 150-164.
Blackburn, D.G., J.R. Stewart, D.C. Baxter, and L.H. Hoffman (1992) Scanning electron microscopy of the placcntal membranes in viviparous garter snakes of the genus Thamnophis. Am. Zool. 32:139A (abstract).
Blackburn, D.G., L.J. Vitt, and C.A. Beuchat (1984) Eutherian-like reproductive specializations in a vivipa- rous reptiles. Proc. Natl. Acad. Sci. USA 81:4860- 4863.
Boyd, M.M.M. (1942) The oviduct, foetal membranes, and placentation in Hoplodactylus maculatus Gray. Proc. Zool. SOC. London Series A 112:65-104.
Dufaure, J.P., and J. Hubert (1961) Table de d6veloppe- ment du lezard vivipare Lacerta (Zootoca) uiuipara Jacquin. Arch. Anat. Micr. Morphol. Exp. 50:309-327.
Fox, H. (1977) The urogenital system of reptiles. In C. Gans and T.S. Parsons (eds): Biology of the Reptilia. Vol. 6. London: Academic Press, pp. 1-157.
Ghiara, G., F. Angelini, M. Zerani, A. Gobbetti, G. Cafi- ero, and V. Caputo (1987) Evolution of viviparity in Scincidae (Reptilia, Lacertilia). Acta Embryol. Mor- phol. Exp. 8t187-201.
Giacomini, E. (1891) Materiali per la storia dello svil- IDD DO del seps chalcides. (Cuv.) Bonao. Monitore Zool.
2:i79-i92,198-211. Giacomini, E. (1892) Contrihuto all migliore conoscenza
degli annessi fetali nei Rettili. Monitore Zool. Ital. 3:126-128,157-163.185-196.
Giacomini, E. (1893) Sull’ovidutto dei Sauropsidi. Moni- tor 2001. Ital. 4~202-265.
Grosser, 0. (1927) Fruhentwicklung und Eihautbildung und Plazentation des Menschen und der Saugctiere. Miinchen: Bergmann.
Guillette, L.J., Jr., and R.E. Jones (1985) Ovarian, oviduc- tal, and placental morphology of the reproductively bimodal lizard, Sceloporus mneus. J. Morphol. 184t85- 98.
Hoffman, L.H. (1970) Placentation in the garter snake, Thamnophis sirtalis. J. Morphol. 131:57-88.
Hrabowski, H. (1926) Das Dotterorgan der Eidechsen. Z. Wiss. Zool. 128t305-382.
Hubert, J. (1985) Embryology of the Squamata. In C. Gans and F. Billet (eds): Biology of the Reptilia. Vol. 15. Dcvclopment, B. New York: John Wiley & Sons, pp. 1-34.
Ibrahim, M.M. (1977) Studies on Viviparity in Chalcides ocellatus Forsk. PhD Dissertation, University College of North Wales, Bangor.
Jacobi, L. (1936) Ovoviviparie bei einheimischen Eidech- sen. Z. Wiss. Zool. 148t401-464.
Kasturirangan, L.R. (1951) Placentation in the sea- snake, Enhydrina schistosa (Daudin). Proc. Indian Acad. Sci., B 34:l-32.
Luckett, W.P. (1977a) Placentation in the viviparous reptile, Cha.lcides chalcides. Anat. Rec. 187:642 (ab- stract).
Luckett, W.P. (197713) Ontogeny of amniote fetal mem- branes and their application to phylogeny. In M.K. Hocht, P.C. Goody, and B.M. Hecht (eds): Major Pat- terns in Vertebrate Evolution. New York: Plenum Press, pp. 439-516.
Mossman, H.W. (1987) Vertobrate Fetal Membranes. New Brunswick, NJ: Rutgers University Press.
Panigel, M. (1951) Rapports anatomo-histologiques etab- lis au cows de la gestation entre l’oeuf et l’oviducte maternal chez le lezard ovovivipare, Zootoca uiuzparo W. (Lacerta uiuipora J.) Bull. SOC. Zool. Fr. 76:163- 170.
Peter, K. (1904) Normentafel zur Entwicklunsgeschichte der Zauneidechse (Laceria ogilis). In F. Keibel (ed): Normentafcln zur Entwicklungsgevchichte der Wirbel- tiere. Part 4. Jena: G. Fischer, pp. 1-165.
Raynaud, (1985) Development of limbs and embryonic limb reduction. In C. Gans and F. Billet (eds): Biology of the Reptilia. Vol. 15. Development, B. New York: John Wiley & Sons, pp. 59-148.
Stewart, J.R. (1985) Placentation in the lizard Gerrhono- tus coeruleus with a comparison to the extraembryonic membranes of the oviparous Gerrhonotus multicarina- tus (Sauria, Anguidae). J. Morphol. 185:lOl-114.
Stewart, J.R. (1990) Development of the extraembryonic membranes and histology of the placentae in Virginia striatula (Squamata: Serpentes). J. Morphol. 205:33- 43.
Stewart, J.R. (1992) Placental structure and nutritional pruvision to embryos in predominantly lecithotrophic viviparous reptiles. Am. Zool. 32.303-312.
Stewart, J.R., and D.G. Blackburn (1988) Reptile placen- tation: structural diversity and terminology. Copcia 1988:839-852.
Studiati, C. (1853) Miscellanea di osservazioni zoo- tomiche. I l l . Intorno alle connessioni dell’uovo coll’ovidutto nel Seps tridmtylus. Mem. Reale Accad. Sci. Torino Ser. Sec. 15~101-113.
Ten Cate-Hoedemaker, N.J. (1933) Bcitrage zur Kennt- nis der Plazentation bei Haien und Reptilien. Z. Zellfor- sch. Mikrosk. Anat. 18:299-345.
Weekes, C.H. (1927) Placentation and other phenomena in the scincid lizard Lygosoma (Hinulia) quoui. Proc. Zool. Soc.New South Wales 52:499-554.
Weekes, C.H. (1929) On placentation in reptiles. I. Proc. Linn SOC. New South Wales 54:34-60.
Weekes, C.H. (1930) On placentation in reptiles. 11. Proc. Linn SOC. New South Wales 55:550-576.
Weekes, C.H. (1935) A review of placentation among reptiles, with particular regard to tho function and evolution of the placenta. Proc. Zool. SOC. London 2: 625-645.
Wourms, J.P. (1981) Viviparity: the maternal-fetal rela- tionship in fishes. Am. Zool. 21:473-515.
Yaron, Z. (1985) Reptile placentation and gestation: struc- ture, function, and endocrine control. In C. Gans and F. Billet (eds): Biology of the Reptilia. Vnl. 15. Develop- ment, B. New York: John Wiley & Sons, pp. 527-603.
Related Documents











