HISTOCHEMICALSTAININGAND CHARACTERIZATIONOFGLYCOPROTEINSIN ACID-SECRETINGCELLSOFFROGSTOMACH TRUDYM .FORTE and JOHNG .FORTE Fromthe Donner LaboratoryandDepartmentofPhysiology-Anatomy,University ofCalifornia, Berkeley,California 94720 ABSTRACT Glycoproteinswerehistochemicallylocalizedinoxynticcellsofthefrog stomachbystain- ingwithperiodicacid-silvermethenamine .Reductionofsilverwas mostintenseon(a) theouteraspectoftheapicalplasmalemma,(b)withinthe tubularsmoothmembrane systemcharacteristicofoxynticcells,and(c)withincisternaeandvesiclesofthe Golgicom- plex .Othermembranecomponentssuchasthosefromthemitochondria,nucleus,junctional complex,lateralandbasalcellmembranesshowedlittleornostainability . Gastricmucosal homogenateswerefractionatedbycentrifugationforfurthermorphologicalandchemical analysis .Thestainingreactionofthemicrosomalfraction(40,000g X60min)wassimilar tothatofthetubularmembranouscomponentsofintactoxynticcells .Carbohydrate analyses showedthatallcellfractionsareextremelylowinacidicsugars, uronicandsialicacids, whileneutralsugarsandhexosaminesarerelativelyabundant . Themicrosomalfraction containsthelargestproportionofcarbohydrates,ca.9%ofthefat-freedryweight . Another distinguishingfeatureisthatglucosamineistheonlydetectablehexosamine inthemicro- somalfraction .Thesehistochemicalandchemicaldataindicatethatneutral glycoproteins areassociatedwithmembranouscomponentswhichhavebeenimplicatedintheprocess of HC1secretionbyoxynticcells .Thestainingpatternwithinthecellssupports thehypothesis ofinterrelationshipsbetweentheGolgimembranes,tubularsmoothmembranes, andapical surfacemembrane . INTRODUCTION Theoxynticoracid-secretingcellsofthefrog stomacharecharacterizedbythepossessionof abundant smooth-surfaced tubular structures withintheapicalcytoplasm(1,2) .Parallelstudies relatingsecretoryactivitytogeneralmorphologi- calappearancehavesupportedtheproposalthat thesetubularmembranousunitsareintimately concernedwiththeH+transportprocessofthe stomach (2-5) . Anotherpointofevidencetofavor thisviewwasderivedfromourexperimentswith metamorphosingbullfrogtadpole stomachsin THEJOURNALOFCELLBIOLOGY • VOLUME47,1970 • pages 437-459 whichitwasfoundthatthedevelopmentalap- pearanceofH+secretorycapacitywastemporally correlatedwiththemorphogenesisofthetubular membraneswithinoxynticcells (6) . Inthissame studyontadpolestomachs,wepresentedevidence whichsuggestedthattheGolgiapparatuswasin factthecellularsiteforsynthesisandelaboration ofthetubularmembranous elementswhich accumulatedatthecommencementofsecretory activity. Onthebasisoftheavailableinformation,we 437 Downloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 March 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

HISTOCHEMICAL STAINING ANDCHARACTERIZATION OF GLYCOPROTEINS IN
ACID-SECRETING CELLS OF FROG STOMACH
TRUDY M. FORTE and JOHN G . FORTE
From the Donner Laboratory and Department of Physiology-Anatomy, University of California,Berkeley, California 94720
ABSTRACT
Glycoproteins were histochemically localized in oxyntic cells of the frog stomach by stain-ing with periodic acid-silver methenamine . Reduction of silver was most intense on (a)the outer aspect of the apical plasmalemma, (b) within the tubular smooth membranesystem characteristic of oxyntic cells, and (c) within cisternae and vesicles of the Golgi com-plex. Other membrane components such as those from the mitochondria, nucleus, junctionalcomplex, lateral and basal cell membranes showed little or no stainability . Gastric mucosalhomogenates were fractionated by centrifugation for further morphological and chemicalanalysis . The staining reaction of the microsomal fraction (40,000 g X 60 min) was similarto that of the tubular membranous components of intact oxyntic cells . Carbohydrate analysesshowed that all cell fractions are extremely low in acidic sugars, uronic and sialic acids,while neutral sugars and hexosamines are relatively abundant . The microsomal fractioncontains the largest proportion of carbohydrates, ca. 9% of the fat-free dry weight . Anotherdistinguishing feature is that glucosamine is the only detectable hexosamine in the micro-somal fraction. These histochemical and chemical data indicate that neutral glycoproteinsare associated with membranous components which have been implicated in the process ofHC1 secretion byoxyntic cells. The staining pattern within the cells supports the hypothesisof interrelationships between the Golgi membranes, tubular smooth membranes, and apicalsurface membrane .
INTRODUCTION
The oxyntic or acid-secreting cells of the frogstomach are characterized by the possession ofabundant smooth-surfaced tubular structureswithin the apical cytoplasm (1, 2) . Parallel studiesrelating secretory activity to general morphologi-cal appearance have supported the proposal thatthese tubular membranous units are intimatelyconcerned with the H+ transport process of thestomach (2-5) . Another point of evidence to favorthis view was derived from our experiments withmetamorphosing bullfrog tadpole stomachs in
THE JOURNAL OF CELL BIOLOGY • VOLUME 47, 1970 • pages 437-459
which it was found that the developmental ap-pearance of H+ secretory capacity was temporallycorrelated with the morphogenesis of the tubularmembranes within oxyntic cells (6) . In this samestudy on tadpole stomachs, we presented evidencewhich suggested that the Golgi apparatus was infact the cellular site for synthesis and elaborationof the tubular membranous elements whichaccumulated at the commencement of secretoryactivity.
On the basis of the available information, we
437
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

developed an hypothesis that the tubular mem-branes of mature oxyntic cells represent an inter-mediate morphological stage between their genesisin the Golgi region and their participation ingastric HCl secretion via intercommunicationwith the apical plasmalemma . One approach toascertaining such interrelationships betweenvarious membranous components would employspecific cytochemical staining techniques on bothintact tissue and cell membrane fractions .
The technique of Rambourg and Leblond (7)for glycoprotein staining was of particular interestsince they found that outer cell membrane sur-faces and inner aspects of the Golgi saccules wereamong the most reactive sites. These authorsstressed the possible interrelationship of Golgistructures with the plasmalemma, especially withrespect to synthesis of constituents of the cell coat .
In the present communication, we employ theperiodic acid-silver methenamine staining reactionof Rambourg and Leblond (7) for investigatingmembranous structures of the bullfrog oxynticcell . The histochemical procedures are coupledwith morphological and chemical analysis of iso-lated cell fractions. We present evidence for a gly-coprotein material which stains with equal in-tensity within the smooth-surfaced tubules, withinGolgi saccules, and on the outer aspect of the apicalcell membrane . The periodic acid-silver stainingreaction of isolated gastric microsomes fuithersupports the notion that this cell fraction is largelyderived from the smooth tubules of oxyntic cells .Furthermore, carbohydrate analysis demonstratesthe unique nature of the glycoprotein moietiesassociated with various gastric cell fractions .
METHODS
Fixation of Stomach Tissue
The frogs (Rana pipiens or Rana catesbeiana) weresacrificed by pithing, and the stomachs were imme-diately removed. The underlying muscle coat wasstripped from the mucosa, and the latter portion wasplaced in a fixative of 3% glutaraldehyde in 0 .1 M
sodium cacodylate buffer, pH 6 .8, and minced intosmall pieces (less than 1 mm3) . The pieces were placedin fresh glutaraldehyde fixative for an additional 60min at room temperature . The tissue was then washedfor 90 min in three changes of 0 .1 M cacodylate buffer.Dehydration was carried out through a graded seriesof alcohol, and the tissue was finally embedded inEpon. Silver sections were cut with glass knives andmounied on uncoated, 200-mesh, stainless steel grids .
438
THE JOURNAL OF CELL BIOLOGY . VOLUME 47, 1970
These grids were stored in a desiccator until subse-quent staining procedures .
Staining Procedures : PeriodicAcid-Silver Methenamine
Sections were stained according to the methoddescribed by Rambourg (8) . All staining was carriedout at 60 °C in a darkened room . The sodium borate,silver nitrate, and silver methenamine solutions werefreshly prepared each time . Staining for 20 min withsilver methenamine gave good positive results . Non-specific staining was assessed by deleting the periodicacid (P.A .) oxidation step in the treatment of somesections .
Periodic-Chromic Acid-Silver Methenamine
The method recently described by Rambourg,Hernandez, and Leblond (9) was followed, exceptthat Epon and not Vestopol was used for embeddingmaterial. The sections were stained in silver methen-amine for 40 min .All sections were examined in the Hitachi HUmicroscope .
Isolation of Cell Fractions fromMueosal Homogenates
11
The gastric mucosa was obtained as describedabove, cut along the lesser curvature to form a sheet,and stretched out on filter paper. The mucosal, orsecretory, surface was gently scraped, then blottedvigorously with filter paper so as to remove adheringmucus and some of the surface epithelial cells (10) .The secretory surface of the fundic mucosa was thenscraped vigorously with a stainless steel spatula. Thisremoved most of the gastric glands from the under-lying muscularis mucosae and connective tissue . Theglandular scrapings from one or more stomachs werethen triturated in a Potter-Elvehjem homogenizerwith approximately 40 volumes of homogenizingmedium. The latter was composed of 110 mm KC1,10 mm Tris HCI, and 2 mm Na ethylenediamine-tetraacetate (EDTA), all adjusted to pH 7 .3 . Frac-tionation by differential centrifugation is given inTable I . The four cell fractions were designated (I)sediment, (II) mitochondria, (III) microsomes, and(IV) supernatant. The cell fractions were subse-quently used for ultrastructural and cytochemicalstudies or for chemical analysis. It is acknowledgedthat these cell fractions are not homogeneous withrespect to cell organelles or their fragments . Themethod of separation is based primarily upon earlierwork on enzymatic functions of gastric cell fractions(11-13) .
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

TABLE I
Procedure for Preparation of Gastric Mucosal Cell Fractions by Differential Centrifugation
Homogenize scraped gastriccells in isolation medium*
15000 g X 5 min
IPellet
resuspended andhomogenized inisolation medium*
5000 g X 5 min
IPellet
I. SEDIMENT
PelletII. MITOCHONDRIA
Pellet
IV. SUPERNATANTIII. MICROSOMES* Isolation medium contained 110 ram KCI, 10 rum Tris-HCI, 2 mm EDTA adjusted to pH 7 .3 .
Fixation
A small aliquot of an isolated cell fraction wasplaced in a conical BEEM' capsule, and an equalquantity of 2 .5 0/c glutaraldehyde in 0 .1 M sodiumcacodylate was added . The membranes were fixed inthis fashion for 60 min and then centrifuged at 10,000g for 10 min in order to pellet the material . The restof the fixation was similar to that described for wholetissue, except that all steps were carried out in theconical BEEM capsules .
Negative Staining
A sample of the microsomal fraction, approxi-mately 1 mg of protein/ml, was fixed in 2% glu-taraldehyde in 0.1 M phosphate buffer, pH 7 .4,overnight at 4 °C. The fixed preparation was dilutedwith an equal volume of 2% sodium phosphotung-state, pH 7.4, and a small drop of this suspension wasplaced on a Formvar-carbon-coated grid . After 1min, excess fluid was withdrawn with filter paper andthe grid was immediately examined in the microscope .
Preparation of Cell Fractions for
Supernate40,000 g X 60 min
twice with PCA and twice with distilled water . Theprecipitates were dried (90 °C overnight), weighed,treated with two washes of CHC13-CH3OH (2 :1),subsequently dried, and reweighed . The fat-freeprecipitates were digested with papain by suspensionin a digestion medium containing 1 M -NaCl, 0 .1 M
Na phosphate, 5 mm cysteine and 5 sum EDTAadjusted to pH 6 .5 . Approximately 85 mg of papainper mg fat-free dry weight was added to each tubeaccording to the procedure of Pamer et al . (14) .Digestion was carried out at 60 ° C for 24-48 hr withperiodic adjustment of the pH of the medium back topH 6.5 with NaOH . After digestion, PCA was addedto a final concentration of 0 .3 M and the tissue digestswere centrifuged at 10,000 g for 10 min . The super-nates were placed in washed dialysis tubing anddialyzed against 600 volumes of 0.02 m NaCl for 48hr. The dialysates were collected and evaporated todryness in vacuo over NaOH and P 2O5. The driedresidue containing carbohydrates and peptides(glycopeptide residue) was taken up in a volume ofH2O and stored at 4 °C for further analysis .
Carbohydrate Analysis
Supernate
I
CombinedSupernate--Supernate
12,000 g X 10 min
Chemical Analysis Sialic acid was measured both on the "purified"glycopeptide residues and on the crude cell fractionsprepared by differential centrifugation . The proce-dure of Warren (15) was used for these assays . Neutralsugars were analyzed according to the methods
t Better Equipment for Electron Microscopy, Bronx, described by Dische (16) . D-Glucose and L-fucose wereN.Y .
used as the respective standards . The neutral sugar
Cell fractions were treated with 0 .3 M perchloricacid (PCA) and the resulting precipitates were washed
T. M. FORTE AND J. G. FORTE Histochemical Staining of Glycoproteins
439
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

analyses were carried out both directly on the glyco-peptide residues and on aliquots hydrolyzed insealed vials containing I M HC1 at 100 °C for 4 hr.The latter procedure resulted in a destruction of some25-50% of hexose, and thus was discontinued .Uronic acid was determined by the method of
Bitter and Muir (17), using n-glucuronolactone(CalBiochem, Los Angeles, Calif.) and chondroitinsulfate "C" (CalBiochem) as standards .
For hexosamine determinations, aliquots of theglycopeptide residues were sealed under vacuum invials containing 6 M HCl. Hydrolysis proceeded at95 ° C for 4 hr, and the resulting hydrolysate wasevaporated to dryness in vacuo over NaOH andH2SO4 at room temperature . The hydrolysates wereagain evaporated to dryness after approximately2 ml of H2O was added to each sample . Hexosaminewas determined either directly on the hydrolysate byElson-Morgan method (18) or after the chromato-graphic purification described by Boas (19) . Theeffectivity of hydrolysis and the ultimate yield of theassay procedures were ascertained by standards ofchondroitin sulfate and internal standard of n-galac-tosamine-1- 14C which were carried on through theentire hydrolysis and purification procedure . Thesetests showed at least 90% recovery for hexosamines .The ratio of glucosamine :galactosamine was deter-mined by the borate depression method of Tracey(20) .
RESULTS
Morphology
The fine structure of oxyntic cells fixed onlywith glutaraldehyde is seen in Fig. 1 . Althoughover-all preservation is excellent, there appears tobe some vacuolization especially within the apicalregion of the cells . The lumen of the gastric glandis partially occluded by cytoplasmic extensionsfrom the apical surface of the cells : these surfaceextensions have often been described as micro-villi (2, 3, 5) . An outstanding feature within theoxyntic cells is the abundance of tubular smoothmembrane profiles seen both in cross-section andin longitudinal section . The tubular elements arepacked extremely closely and in a very regularpattern as is evident from the hexagonal arrayindicated by the arrows in the micrograph . Mito-chondria are well preserved but appear in nega-tive contrast since the cristae have little electronopacity without osmium tetroxide postfixation .The rough endoplasmic reticulum is interspersedwith the mitochondria in the more basal regionsof the cells . Selected areas of the oxyntic cell are
440
THE JOURNAL OF CELL BIOLOGY • VOLUME 47, 1970
shown at higher magnification in Fig . 2 a-c . It isapparent that the staining intensity of specific cellstructures differs considerably with glutaraldehydeas the sole fixative, as compared to previous workin which aldehyde fixation was followed byosmium tetroxide postfixation (2, 5, 21) .
A section through a region dense with tubulesis shown in Fig . 2 a . Inspection of transverse pro-files of tubules reveals that the membrane is atrilaminar structure : an electron-opaque innerleaflet adjacent to the tubular lumen ; an electron-transparent region ; and a dense outer leaflet .The cytoplasmic regions between the tubulesappear quite dense under these conditions offixation, as compared to those which were post-fixed in osmium tetroxide, and the outer leaflet ofthe tubule often blends with the cytoplasmicdensity . The mean over-all diameter of the tubuleis about 360 A, whereas the diameter across theinner dense leaflet of the tubule is 200 A . Thecenter-to-center spacing from one tubule toanother in hexagonally packed regions is approxi-mately 400 A .
A region from the more basal portion of anoxyntic cell is shown in Fig . 2 b . The cisternae ofthe rough endoplasmic reticulum appear to have adense core which may result from the appositionof the inner faces of the cisternal membranes .Dense, round ribosomal particles can be seen liningthe outer surfaces of the membranes .functional complexes between adjacent oxyntic
cells often show interesting detail after only glu-taraldehyde fixation, as seen in Fig . 2 c . A numberof electron-opaque filamentous structures may beseen joining the adjacent faces of the desmosome .These structures, which are ordinarily not seenafter osmium tetroxide postfxation, have ratherregularly spaced intervals of approximately 100 A .
P.A .-Silver Methenamine Staining
Sections oxidized with periodic acid were in-tensely stained after silver reduction .
This phenomenon is seen in Fig. 3 which showsparts of a lumen surrounded by several oxynticcells . It is clearly evident that the luminal surfacemembranes and apical cytoplasmic membranes ofthese cells are heavily stained. By nature of thestaining procedure, i .e. no osmium or counter-staining with uranyl and lead salts, it is not pos-sible to visualize the individual profiles of thecytoplasmic tubules ; however, the strandlikedeposition of reduced silver indicates that this is
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGURE 1 Section through a portion of two oxyntic cells which have been fixed with only glutaraldehyde .Numerous irregular cytoplasmic extensions project into the gland lumen (Lu) . Just beneath the apicalsurface is a cytoplasmic region relatively free of structures, and adjacent to this region are innumerableprofiles of tubular smooth membranes seen both in transverse and in longitudinal aspects . The hexagonalpacking of the tubular structures is indicated by arrows . Mitochondria (M) and the rough-surfacedendoplasmic reticulum (rer) are generally confined to the more basal portion of the cell . ICS refers tothe intercellular space while je indicates the junctional complex. An occasional zymogen granule (Z) canalso be seen. Section was stained with uranyl and lead salts . X 33,600 .
T. M. FORTE AND J. G . FORTE Hiatochemical Staining of Glycoproteins
441
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGURE 2 An enlarged view of selected areas from oxyntic cells fixed with only glutaraldehyde. Fig(2a, Section through smooth tubular elements in the apical region of the cell . The tubule membranes lacetrilaminar structures consisting of two dense layers separated by an electron-transparent middle layer .Lumina of the tubules are electron transparent while the surrounding cytoplasm is relatively dense .The hexagonal arrangement of the tightly packed tubules is also seen with a center-to-center distanceof approximately 400 A . X 79,500. Fig. 2 b, More basal region of oxyntic cell showing rough-surfacedendoplasmic reticulum (rer) and mitochondria (M) . The outer surface of the cisternae of the rer arestudded with dense ribosomal particles, and a dense line, probably representing apposed membranes, canbe seen within the cisternae. The mitochondria, whose cristae are far less dense than the surroundingprotein matrix, appear in negative contrast compared to conditions in which postosmication is employed .X 56,000 . Fig . 2 c, A section through a junctional complex between two oxyntic cells . The tight junc-tion (tj) displays the usual pentalaminar structure . Adjacent faces of the desmosqme (des) are joined byfine, dense, filamentous structures which are spaced at rather regular intervals . X 111,900 . All sectionswere strained with uranyl and lead salts .
indeed the tubular membrane system . The mito-chondria, on the other hand, are almost devoid ofstain . The tight junctions, lateral borders, and thebasal membranes including the basal infoldingsare also only lightly stained . The basement mem-brane of oxyntic glands shows little affinity for thestain ; this is in contrast to the heavily reactivebasement membrane shown by Rambourg andLeblond for several epithelial tissues (7) . Some ofthe stained structures, however, such as the nu-cleus, ribosomes, and extracellular collagen, werenonspecifically stained as judged by silver methe-namine treatment of nonoxidized sections .
The apical portion of an oxyntic cell is seen in
442
THE JOURNAL OF CELL BIOLOGY . VOLUME 47, 1970
greater detail in Fig. 4 . The region just beneath thesurface is almost devoid of structures ; this corre-sponds to the cytoplasmic zone frequently seen inconventionally stained sections (cf. Fig. 1) . Occa-sional heavily stained vesicles are seen within theclear zone, and these profiles probably representoblique sections through surface infoldings . Thebulk of the cell is intensely stained in the apicalregion corresponding to the location of the smooth-surfaced tubular membranes. The reduced silvergrains appear spherical or as beaded strands sug-gesting transverse or longitudinal sections throughthe tubular system .
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGuRE 3 Transverse section through a portion of a gastric gland treated with periodic acid-silvermethenamine . Apical cytoplasmic extensions, whose surfaces are intensely stained, can be seen liningthe gland lumen (Lu) . The apical portion of the cell corresponding to the tubular smooth-surfaced mem-branes (ser) is also heavily stained . Mitochondria (M) are almost devoid of stain while the nucleus (N)and extracellular collagen (col) are nonspecifically stained . The lateral cell borders (ICS region) andbasal membrane infoldings (BF) are only moderately stained. X 15,000 .
The qualitative difference in stainability of the surfaces are much less intensely stained while thevarious regions of the surface membranes is strik- tight junction is almost devoid of stain. Such varia-ingly evident in Fig. 4. The apical portions of the tions in staining of the cell surface suggest differ-plasma membrane avidly reduce silver methena- ences in the chemical nature of various portionsmine; on the other hand, the lateral and basal of the cell membrane .
T. M. FORTE AND J. G. FORTE Histochemical Staining of Glycoproteins
443
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022
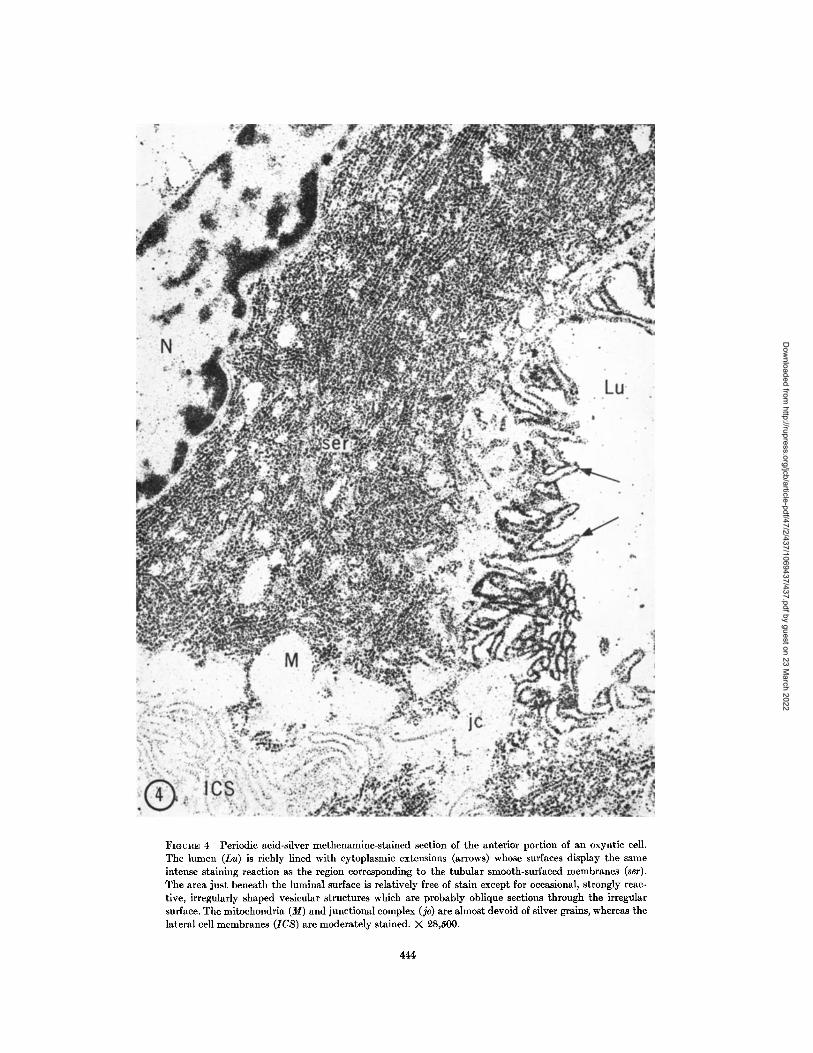
FIGURE 4 Periodic acid-silver methenamine-stained section of the anterior portion of an oxyntic cell .The lumen (Lu) is richly lined with cytoplasmic extensions (arrows) whose surfaces display the sameintense staining reaction as the region corresponding to the tubular smooth-surfaced membranes (ser) .The area just beneath the luminal surface is relatively free of stain except for occasional, strongly reac-tive, irregularly shaped vesicular structures which are probably oblique sections through the irregularsurface . The mitochondria (M) and junctional complex (je) are almost devoid of silver grains, whereas thelateral cell membranes (ICS) are moderately stained . X 28,500 .
444
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGURE 5 Periodic acid-chromic acid-silver methenamine-stained section through several oxyntic cells .The lumen (Lu) is almost completely occluded by the apical cytoplasmic extensions which are heavilystained . Vesicular structures just beneath the surface are also stained . The internal aspects of the tubularsmooth membranes are intensely stained in both their longitudinal and transverse profiles . Regionsshowing hexagonal packing are indicated by arrows . The Golgi apparatus (Go) is highly reactive whereasnuclei (N), mitochondria (M), lateral cell membranes (ICS) and junctional complexes (je) show little or noreactivity. X 9.8,500 . The inset shows in greater detail the structure and arrangement of the hexagonallypacked tubular smooth membranes where a center-to-center spacing of 400 A is found (cf . Fig. 9. a) .X 79,500 .
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGURE 6 Tubular and vesicular structures from the microsomal fraction of the frog stomach seen innegative contrast after sodium phosphotungstate staining . X 58,500 .
P.A.-Chromic Acid-Silver
Methenamine Staining
Rambourg, Hernandez, and Leblond (9) dem-onstrated that when chromic acid oxidation wasused in conjunction with periodic acid the stainingreaction with silver methenamine was far morespecific and sensitive . This was borne out in ourpreparation as typified in Fig. 5. Nuclear andribosomal stainability are very much reduced .For the oxyntic cells shown in Fig . 5, the glandularlumen is almost completely occluded by the apicalprojections which are intensely stained at theirsurfaces. Just beneath the surface, there is a rela-tively clear area with a few vesicular structureswhich are also stained and probably representoblique sections through the folds of the highlyirregular surface . Densely stained tubular ele-ments of the smooth-surfaced membranes in bothlongitudinal and transverse aspects are seenthroughout the apical portions of the cell ; in fact,in some areas the hexagonal packing of the tubulesis clearly evident with a center-to-center spacing ofapproximately 400 A . In cross-section, the tubulesappear to contain a ring of dense silver granules ;in some instances, the tubular lumina are alsofilled with stain (Fig. 5, inset) . Although a finestructural analysis of stain distribution is difficult
446
THE JOURNAL OF CELL BIOLOGY . VOLUME 47, 1970
owing to the large grain siz of reduced silveerparticles, we have measured the outer diameterof the dense outlines in areas of hexagonal packing .The mean diameter was 280 A. Since the outerreaches of the distribution of stain approximatelycorrespond to the position and dimensions of themiddle, electron-transparent lamina of the tubulemembrane, it is likely that the reactive chemicalgroups are predominantly associated with theinner aspects of the tubular membrane .
The Golgi apparatus which is found close to thenucleus is also highly reactive with silver methena-mine. Stain is found associated with both thecisternal and vesicular elements of the Golgisaccules. Mitochondria membranes and lateral andbasal plasma membranes, on the other hand, havelittle affinity for the stain . As was the case in Figs .3 and 4, the junctional complex is barely dis-cernible .
Staining Properties of Isolated Cell Fractions
The microsomal fraction visualized after nega-tive staining (Fig . 6) contains tubular structureswhich often form closed loops. Occasionally,however, vesicular structures can also be seen .Such variously shaped elements are also found insections of the microsomal pellet which has been
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

FIGURE 7 Section though a glutaraldehyde-fixed microsomal pellet . Numerous lamellar and vesicularstructures are present ; however, other structures such as multivesicular bodies (mvb) and clear, ovoidgranules (g) can also be seen . Section was stained with uranyl and lead salts . X 44,400 .
FIGURE 8 Same microsomal preparation as in Fig . 7, except that the section was stained with periodicacid-chromic acid-silver methenamine . There are abundant highly reactive strands and rounded masseswhich correspond to the membranous lamellae and vesicles seen in the previous micrograph . The largeovoid transparent areas probably identify the region of the granules seen in Fig . 7 . X 44,400 .
FIGURE 9 Periodic acid-chromic-silver methenamine-treated section of glutaraldehyde-fixed mito-chondrial fraction . Outlines of mitochondria (M) are barely discernible by this staining technique . Thedense granules (arrows) may represent fragments of adhering mucus . X 42,000 .
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022
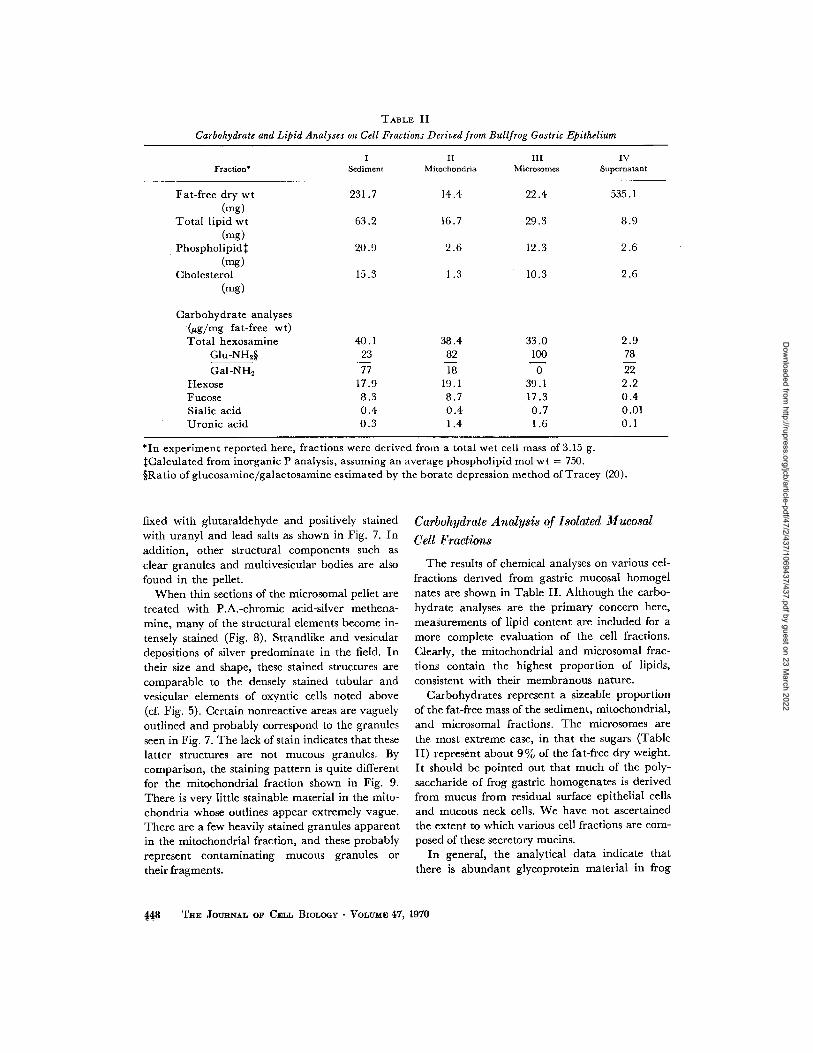
TABLE II
Carbohydrate and Lipid Analyses on Cell Fractions Derived from Bullfrog Gastric Epithelium
*In experiment reported here, fractions were derived from a total wet cell mass of 3 .15 g.$Calculated from inorganic P analysis, assuming an average phospholipid mol wt = 750 .§Ratio of glucosamine/galactosamine estimated by the borate depression method of Tracey (20) .
fixed with glutaraldehyde and positively stainedwith uranyl and lead salts as shown in Fig . 7 . Inaddition, other structural components such asclear granules and multivesicular bodies are alsofound in the pellet .
When thin sections of the microsomal pellet aretreated with P.A.-chromic acid-silver methena-mine, many of the structural elements become in-tensely stained (Fig. 8) . Strandlike and vesiculardepositions of silver predominate in the field . Intheir size and shape, these stained structures arecomparable to the densely stained tubular andvesicular elements of oxyntic cells noted above(cf. Fig . 5) . Certain nonreactive areas are vaguelyoutlined and probably correspond to the granulesseen in Fig . 7 . The lack of stain indicates that theselatter structures are not mucous granules . Bycomparison, the staining pattern is quite differentfor the mitochondrial fraction shown in Fig . 9 .There is very little stainable material in the mito-chondria whose outlines appear extremely vague .There are a few heavily stained granules apparentin the mitochondrial fraction, and these probablyrepresent contaminating mucous granules ortheir fragments .
448 THE JOURNAL OF CELL BIOLOGY - VOLUME 47,1970
Carbohydrate Analysis of Isolated MucosalCell Fractions
The results of chemical analyses on various cel-fractions derived from gastric mucosal homogelnates are shown in Table II. Although the carbo-hydrate analyses are the primary concern here,measurements of lipid content are included for amore complete evaluation of the cell fractions .Clearly, the mitochondrial and microsomal frac-tions contain the highest proportion of lipids,consistent with their membranous nature .
Carbohydrates represent a sizeable proportionof the fat-free mass of the sediment, mitochondrial,and microsomal fractions . The microsomes arethe most extreme case, in that the sugars (TableII) represent about 9% of the fat-free dry weight .It should be pointed out that much of the poly-saccharide of frog gastric homogenates is derivedfrom mucus from residual surface epithelial cellsand mucous neck cells. We have not ascertainedthe extent to which various cell fractions are com-posed of these secretory mucins .
In general, the analytical data indicate thatthere is abundant glycoprotein material in frog
Fraction*I
SedimentII
MitochondriaIII
MicrosomesIV
Supernatant
Fat-free dry wt 231 .7 14 .4 22 .4 535 .1(mg)
Total lipid wt 63.2 16 .7 29.3 8.9(mg)
Phospholipid$ 20.9 2 .6 12 .3 2 .6(-g)
Cholesterol 15 .3 1 .3 10 .3 2 .6(mg)
Carbohydrate analyses(µg/mg fat-free wt)Total hexosamine 40 .1 38 .4 33 .0 2 .9
Glu-NH2§ 23 82 100 78Gal-NH2 77 18 0 22
Hexose 17.9 19 .1 39 .1 2 .2Fucose 8.3 8 .7 17 .3 0 .4Sialic acid 0 .4 0 .4 0 .7 0 .01Uronic acid 0.3 1 .4 1 .6 0 .1
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

gastric tissue. The acidic sugars, uronic, andsialic acids, are found in rather low concentra-tions in all cell fractions. In contrast, the neutralsugars, hexoses, and methylpentoses, are relatively
abundant in all fractions and especially prevalentin the microsomal fraction . Amino sugars alsorepresent a major carbohydrate component .Characteristic distinctions occur in the ratio ofglucosamine : galactosamine for each fraction . Itis of interest that no detectable galactosamine wasobserved in the microsomes .
DISCUSSION
Nature of the P .A.-Silver Methenamine-Staining Reaction
Through the use of various staining techniques,a cell coat which is rich in carbohydrate has beendemonstrated for many vertebrate cells (7) . Thecytochemical silver-staining reaction used in thiswork has been attributed to the reduction of thesilver tetramine by the aldehydic groups producedvia oxidation of 1, 2-glycols and a-amino alcoholsby periodic and chromic acids (9) . Glegg, Cler-mont, and Leblond (22) had shown earlier thatneutral sugars are readily stainable whereas acidicmucopolysaccharides, such as heparin andchondroitin sulfate, are not reactive . In their ap-plication of the silver methenamine-staining tech-
nique to numerous tissues, Rambourg, Hernandez,and Leblond (9) demonstrated that the reactionwas most pronounced at the cell surface, the inneraspects of the Golgi apparatus, and in vesicularstructures having their origin from the Golgiregion. From the results of "in vitro" studies onthe stainability of various carbohydrates, theseauthors felt that glycoproteins were most likely tocontain the reactive groups which take up stainin the cell structures.
Histochemical tests on gastric mucosa withthe light microscope (23, 24) and at the ultrastructural level (25) demonstrate that oxynticcells contain essentially no acid mucosubstance .Although acidic glycoproteins, such as sialo-proteins and sulfated glycoproteins, have beenlocalized in the gastric surface epithelial cells, the
only fundic cell type in which a characteristic
mucopolysaccharide occurs is the mammalianchief cell (23, 24) . The low levels of uronic acidsobserved in the frog fundic mucosal cell fractions
are consistent with the histochemical data,
especially since a typical chief cell is not present inthe main body of amphibian stomachs (26) .
On the other hand, periodic acid-Schiff stainingof acid-secreting cells from mammalian stomachhas been demonstrated by several groups (23, 24) .In these reports, there were no accompanyingfine structural studies, and thus it was not possibleto localize the intracellular site of the stain . Re-cently, Sedar (25) has shown that a polysaccharidesubstance, probably a neutral carbohydrate, linesthe smooth-surfaced membrane system and outer
cell coats of parietal cells of the rat after phospho-tungstic acid staining . The staining pattern whichwe have demonstrated with P .A.-silver methena-mine in oxyntic cells of the frog is similar to that
described by Sedar . The bulk of evidence, bothhistochemical and direct analytical, indicates that
the reactive material is glycoprotein .
Interrelations between Membrane Structures
The fact that both the cytoplasmic tubules andapical surface membranes have similar stainingproperties with the P .A.-silver methenamine reac-tion makes the general hypothesis of membranetransformations between these two componentsmore plausible. We attempted to evaluate thespecific location of the stain on the apical plasma-lemma. In untreated sections, similar to thatshown in Fig . 1, the combined thickness of theinterspace plus the adjacent cell membranes oneither side of closely apposed cytoplasmic exten-sions was found to be 200-250 A. In sectionsstained by the P .A.-chromic acid-silver methena-mine procedure, we found that a single dense lineof silver grains, 70-100 A thick, is seen at theinterface between closely apposed apical cell mem-branes (cf. Fig . 5) . This single line of stain is toonarrow to encompass both cell membranes and theinterspace, and thus it is likely that the reactingcarbohydrate groups are on the outer aspects ofthe apical plasmalemma . This and similar argu-ments have been used by Rambourg and Leblond(7) to localize carbohydrate materials of the cellcoat. Specific deposition of the stain on the outerfaces of the apical plasmalemma and at the inneraspect of the tubular smooth membranes is con-sistent with several schematic interpretations of themorphological events which accompany stimula-tion of HCl secretion (2, 27) .
The positive reaction of Golgi membranes withsilver methenamine is consistent with widespreadreactivity of Golgi structures reported by Ram-
T. M. FORTE AND J. G. FORTE Histochemical Staining of Glycoproteins
449
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

bourg, Hernandez, and Leblond (9) . In a previousstudy on the development of tadpole gastricmucosa, we presented morphological evidence tosuggest that the Golgi apparatus was the source oftubular membranes found in mature oxvntic cells(6) . The similar silver-staining reactivity of therespective membranous components supports thisproposed origin . However, such evidence is notintended to imply identical membrane compositionbut merely similarity of specific chemical group-ings.
Carbohydrate Staining in IsolatedCell Fractions
The microsomal fraction of gastric mucosalhomogenates gives a positive reaction with theP.A.-silver methenamine staining procedure andthe pattern of silver deposition is similar to that ofthe tubules seen in intact oxyntic cells. Thus, it isreasonable to speculate that the microsomal frac-tion is largely derived from these cellular struc-tures. This supports previous evidence based uponmorphological comparison (12), and permits aninterpretation of the chemical and enzymatic com-position of gastric microsomes in terms of intactcell components . However, contamination of themicrosomes with other cell constituents is a recog-nized problem in the evaluation of chemical data .
Nature of the Carbohydrate Components
Although a great deal is known about the carbo-hydrate composition of mucins secreted by thestomach, relatively little information is availableon the sugar content of gastric tissue or cell frac-tions derived therefrom. Pamer, Glass, and Horo-witz (14) have given a detailed analysis of theacidic polysaccharides of rabbit gastric mucosa .However, their results are not comparable tothose reported here because their procedures werenot intended to recover neutral polysaccharides orglycoproteins. In a recent report, Nemoto andYosizawa (28) succeeded in isolating four oligo-saccharide fractions from rabbit gastric mucosaldigests : these consisted of a glycopeptide moietyand three acidic glycosamino-glycan (mucopoly-saccharide)-peptides . Our studies are in generalagreement with the analytical data of Nemoto andYosizawa, although a detailed comparison is notpossible since their analytical sugar data are re-ported as a fractional per cent of each polysaccha-ride subfraction. There is a considerable quantity
4 50 THE JOURNAL OF CELL BIOLOGY . VOLUME 47, 1970
of glycoprotein in frog gastric mucosa, On theother hand, the levels of acidic glycosamino-glycans in various cell fractions are rather low(less than 10% of total gastric tissue carbohy-drates) . Again, it may be pointed out that suchdifferences in distribution of neutral and acidiccarbohydrate units between amphibian and mam-malian (rabbit) gastric mucosa are consistent withhistochemical evidence cited earlier that abundantacidic mucopolysaccharide can be demonstratedin the mammalian chief cell only .
Of the various cell fractions isolated from gastricmucosal homogenates, the microsomes are ofspecial interest since (a) they appear to be derivedfrom the tubular membrane system of oxynticcells ; (b) morphological evidence implicates a rolefor this tubular membranous system in the HClsecretory process (2-5) ; and (c) enzyme systems,indirectly linked to H+ secretion, have beenlocalized in the microsomal fraction (11-13)Thus, an insight into the chemical nature andmacromolecular architecture of the microsomalmembranes may prove useful for interpreting themode of their participation in gastric secretoryactivity . The glycoprotein layer which has beenhistochemically localized in the tubular mem-branous system is one such structural andchemical entity .
The level of the present results does not permitan assessment of the number and composition ofspecific glycopeptides within the gastric cell frac-tions. However, the homogeneity of hexosamine(glucosamine) in the microsomes suggests that asingle major glycoprotein might be present in thisfraction . The high content of fucose, especially inthe microsomal fraction, warrants the generalclassification of frog gastric glycoprotein into thefucopeptide of fucoprotein category. On the otherhand, sialic acid represents a relatively low pro-portion of gastric membrane carbohydrates, asituation which is quite different from that ofseveral other membrane systems (29-31) .
The function of the stainable glycoproteinlayer on the oxyntic cell membranes is as yetunknown. Several possible modes of involvementwith the secretory activity of oxyntic cells may beproposed, among which are (a) the glycoproteinmay facilitate membrane transformations duringactive HCl secretion, perhaps by serving as a"recognition" factor at specific fusion points, (b)the carbohydrate moieties may operate as protec-tive groups to prevent denaturation of membrane
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

proteins during H+ production, (c) the carbo-hydrates may have a direct role in the transloca-tion of H+. It had been proposed earlier that amucosubstance within the tubular membranoussystem might act as an anionic exchange matrix forH+ secretion (2, 32) . The paucity of negativelycharged carbohydrate moieties within the gastricmembrane glycoprotein precludes such a possi-bility .
We would like to express our gratitude to Mrs.Frances Taylor for her technical assistance in pre-paring samples for electron microscopy.
This work was supported in part by United StatesPublic Health Service Grants AM10141 andHE 12710, and by a grant from the American MedicalAssociation Educational and Research Foundation .Received for publication 31 March 1970, and in revised form4 June 1970.
REFERENCES
1 . SEDAR, A. W. 1962 . Electron microscopy of theoxyntic cell in the gastric glands of the bullfrog,Rana catesbeiana . III . Permanganate fixation ofthe endoplasmic reticulum . J . Cell Biol. 14:152 .
2 . SEDAR, A. W. 1965 . Fine structure of the stimu-lated oxyntic cell . Fed. Proc. 24:1360.
3 . VIAL, J. D., and H . ORREGO. 1960. Electronmicroscope observations on the fine structureof parietal cells . J . Biophys. Biochem . Cytol.7 :367 .
4 . SEDAR, A. W. 1961 . Electron microscopy of theoxyntic cell in the gastric glands of the bullfrog(Rana catesbeiana) . II . The acid-secretinggastric mucosa . J. Biophys . Biochem . Cvtol. 10 :47 .
5 . KASBEKAR, D. K., G. M. FORTE, and J. G .FORTE . 1968 . Phospholipid turnover and ultra-structural changes in resting and secretingbullfrog gastric mucosa . Biochim . Biophys.Acta . 163 :1 .
6 . FORTE, G. M., L . LIMLOMWONGSE, and J. G.FORTE . 1969. The development of intracellularmembranes concomitant with the appearanceof HC1 secretion in oxyntic cells of the meta-morphosing bullfrog tadpole . J. Cell Sci . 4:709 .
7 . RAMBOURG, A., and C. P. LEBLOND . 1967 . Elec-tron microscopic observations on the carbo-hydrate-rich cell coat present at the surface ofcells in the rat. J. Cell Biol. 32:27 .
8 . RAMBOURG, A. 1967. An improved silver methen-amine technique for the detection of periodicacid reactive complex carbohydrates with theelectron microscope . J . Histochem . Cytochem .15:409.
9. RAMBOURG, A., W . HERNANDEZ, and C. P.
LEBLOND . 1969. Detection of complex carbo-hydrates in the Golgi apparatus of rat cells .J. Cell Biol. 40:395 .
10 . DAVENPORT, H. W., and R. A. ALLEN . 1969. Aresurface epithelial cells necessary for gastricacid secretion? Gastroenterology . 37:158 .
11 . KASBEKAR, D. K., and R. P. DURBIN . 1965. Anadenosine triphosphatase from frog gastricmucosa . Biochim . Biophys. Acta . 105 :472 .
12. FORTE, J. G., G. M . FORTE, and P. SALTMAN .1967. K+-stimulated phosphatase of micro-somes from gastric mucosa. J . Cell. Physiol . 69 :293 .
13 . FORTE, G. M., J . G . FORTE, and R . F . BILS. 1965 .ATPase activity of subcellular fractions ofrabbit gastric mucosa . Fed. Proc . 24:714 .
14 . PAMER, I ., G . B. J . GLASS, and M. I . HOROWITZ .1969. Purification and characterisation ofsulfated glycoproteins and hyaluronidase-resistant mucopolysaccharides from dog gastricmucosa. Biochemistry . 7 :3821 .
15 . WARREN, L. 1959 . The thiobarbituric acid assayof sialic acids . J. Biol . Chem. 234:1971 .
16 . DISCHE, Z . 1955 . New color reactions for deter-mination of sugars in polysaccharides . InMethods of Biochemical Analysis. D. Glick,editor. 2 :325.
17 . BITTER, T., and H . M. MUIR . 1962. A modifieduronic acid carbazole reaction. Anal . Biochem .4:330 .
18. ELSON, L. A., and W. T . J . MORGAN. 1933. Acolorimetric method for the determination ofglucosamine and chondrosamine . Biochem . J.27 :1824 .
19 . BOAS, N. F. 1953 . Method for the determinationof hexosamines in tissues . J. Biol . Chem.204:553 .
20 . TRACEY, M. V. 1959 . A rapid colorimetric dis-tinction between glucosamine and galactos-amine . Biochim. Biophys. Acta . 17:159.
21 . LILLIBRIDGE, C. B. 1968. Electron microscopicmeasurements of the thickness of various mem-branes in oxyntic cells from frog stomachs . J.Ultrastruct. Res. 23:243 .
22 . GLEGG, R. E., Y . CLERMONT, and C . P . LEBLOND .1952 . The use of lead tetraacetate, benzidine,0-dianisidine and a "film test" in investigatingthe periodic-acid-Schiff technique . StainTechnol. 27:277.
23 . SPICER, S. S., and D . C. H . SUN . 1967. Carbo-hydrate histochemistry of gastric epithelialsecretions in dog . Ann. N.Y. Acad. Sci. 140:762 .
24 . GERARD, A. R . LEV, and G. B . J . GLASS . 1967 .Histochemical study of the mucosubstances inthe canine stomach . I . The resting stomach .Amer . J. Dig . Dis . 12 :891 .
25 . SEDAR, A. W. 1969 . Electron microscopic demon-
T. M. FORTE AND J . G. FORTE Histochemical Staining of Glycoproteins
4 5 1
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022

stration of polysaccharides associated withacid-secreting cells of the stomach after "inertdehydration." J. Ultrastruct. Res. 28:112 .
26 . SEDAR, A. S . 1961 . Electron microscopy of theoxyntic cell in the gastric glands of the bullfrog(Rana catesbeiana) . I . The non-acid-secretinggastric mucosa. J. Biophys. Biochem . Cytol . 9 :1 .
27 . FORTE, J . G. 1970 . Hydrochloric acid secretion bygastric mucosa. In Membranes and Ion Trans-port. E . Bittar, editor . Wiley & Sons, Sussex,England. Chap . 4, 3:165.
28. NEMOTO, T ., and Z . YOSIZAWA . 1969 . A glyco-peptide and glycosamino-glycanpeptides iso-lated from rabbit gastric mucosa . J. Biochem .66 :627 .
29. PATTERSON, M . K., and O. TOUSTER . 1962 .
452
THE JOURNAL OF CELL BIOLOGY • VOLUME 47, 1970
Intracellular distribution of sialic acid and itsrelationship to membranes . Biochim . Biophys .Acta . 56:626 .
30. BAKERMAN, S., and G . WASEMILLER. 1967 . Studieson structural units of human erythrocyte mem-brane. I . separation, isolation, and partialcharacterization. Biochemistry . 6 :1100 .
31 . BOSMANN, H. B., A. HAGOPIAN, and E. H . EYLAR.1968 . Cellular membranes : the isolation andcharacterization of the plasma and smoothmembranes of HeLa cells . Arch. Biochem . Bio-phys. 128 :51 .
32 . FORTE, T., and J . G . FORTE . 1969 . Ultrastruc-tural studies on oxyntic cells of bullfrog stomachusing heavy metal tracer, specific stain andfreeze-etch technique . Physiologist . 12 :228 .
Dow
nloaded from http://rupress.org/jcb/article-pdf/47/2/437/1069437/437.pdf by guest on 23 M
arch 2022
Related Documents











