Marine Biology 97, 533-540 (1988) Marine ,=,=.Biology :L~,,oTgL~ 9 Springer-Verlag 1988 Histochemical demonstration of mercury in the intestine, nephridia and epidermis of the marine polychaete Nereis virens exposed to inorganic mercury P.K. Jensen 1 and E. Baatrup 2 1 Institute of Zoology and Zoophysiology, University of Aarhus, DK-8000, Aarhus C, Denmark 2 Institute of Anatomy B, Neurobiology, University of Aarhus, DK-8000, Aarhus C, Denmark Abstract Marine polychaetes Nereis virens (Sars) were exposed to 9 ppb Hg as mercuric chloride in the aquarium water. Con- centrations of mercury in the water and in the worms were monitored using radiolabelled mercury. After 11 d, the average mercury concentration in the nereids was 8.41 ppm Hg and the bioconcentration factor was calculat- ed to be 930. Accumulations of mercury in the tissues were made visible for light and electron microscopy by autometallographic silver enhancement. Mercury was demonstrated in the intestine, nephridia, epidermis and cuticula. In the intestine, mercury was found to be located predominantly in the apical part of the epithelial cells. In the nephridia, mercury deposits were apparent in the peri- toneal cells of the nephridial tubules. Silver-enhanced mer- cury was also observed at the epicuticula, in the collagen fibres of the endocuticula and in the epidermal basal cells. Intracellularly, the observed mercury accumulations were localised predominantly in lysosomes. Extracellularly, mer- cury was observed in the basal laminae of the intestine and the epidermis, and also in the intestinal peritrophic mem- brane. How N. virens copes with mercury toxicity is dis- cussed. Introduction It is known that marine polychaetes accumulate mercury from the environment (Luoma 1977a, b, Kendall 1978, Medeiros etal. 1981) and they have therefore been em- ployed in marine pollution monitoring programs to detect the degree of mercury pollution (Bryan et al. 1980, Kiorboe et al. 1983, Langston 1986). Mercury is highly toxic to adult (Brown and Ah- sanullah 1971, Reish et al. 1976, Reish 1978) and juvenile (Reish et al. 1976) marine polychaetes. Sublethal concen- trations of mercury cause suppression of reproduction (Reish 1978) and have been reported to inhibit the ab- sorption of glycine across the body surface of the poly- chaete Nereis diversicolor (Siebers and Ehlers 1979). On the other hand, only little information regarding the sites of accumulation within polychaetes and the handling of mercury by these animals is available. Luoma (1977b) used autoradiography to show the distribution of 2~ in whole specimens of Neanthes succinea. However, autoradi- ography is unacceptable at higher resolution because of the relatively energetic beta-radiation from the 2~ isotope. Autometallography is a sensitive method for light and electron microscopic demonstration of mercury deposits within organs and cells. According to Danscher and MM- ler-Madsen (1985), accumulations of possibly less than ten mercury atoms bound to sulphide or selenide can be made visible. In the present study, autometallography has been ap- plied to demonstrate mercury deposits in tissues of the polychaete Nereis virens (Sars) exposed to sublethal con- centrations of water-borne mercury chloride. Materials and methods Animals Specimens of the polychaete Nereis virens (Sars) were col- lected from Kysing Fjord, a shallow, Inesohaline estuary near Aarhus in Denmark. Water temperature and salinity at the sampling site was 9 ~ and 19%0 S, respectively. Only intact worms were used for the experiments. In order to obtain homogeneous material, 30 worms with average length of 5-+0.7cm and weighing 340-+ 10 mg (wet wt) were used. According to the length/ age relationship reported by Kristensen (1984), the worms in the present study were estimated to be approximately 1.5 yr old. Prior to experimentation, the nereids were placed in a darkened aquarium for 24 h in order to void their guts. The worms were not fed during the experimen- tal period.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Marine Biology 97, 533-540 (1988) Marine ,=,=.Biology :L ~,,oTgL~
�9 Springer-Verlag 1988
Histochemical demonstration of mercury in the intestine, nephridia and epidermis of the marine polychaete Nereis virens exposed to inorganic mercury
P . K . J e n s e n 1 and E. Baatrup 2
1 Institute of Zoology and Zoophysiology, University of Aarhus, DK-8000, Aarhus C, Denmark 2 Institute of Anatomy B, Neurobiology, University of Aarhus, DK-8000, Aarhus C, Denmark
Abstract
Marine polychaetes Nereis virens (Sars) were exposed to 9 ppb Hg as mercuric chloride in the aquarium water. Con- centrations of mercury in the water and in the worms were monitored using radiolabelled mercury. After 11 d, the average mercury concentration in the nereids was 8.41 ppm Hg and the bioconcentration factor was calculat- ed to be 930. Accumulations of mercury in the tissues were made visible for light and electron microscopy by autometallographic silver enhancement. Mercury was demonstrated in the intestine, nephridia, epidermis and cuticula. In the intestine, mercury was found to be located predominantly in the apical part of the epithelial cells. In the nephridia, mercury deposits were apparent in the peri- toneal cells of the nephridial tubules. Silver-enhanced mer- cury was also observed at the epicuticula, in the collagen fibres of the endocuticula and in the epidermal basal cells. Intracellularly, the observed mercury accumulations were localised predominantly in lysosomes. Extracellularly, mer- cury was observed in the basal laminae of the intestine and the epidermis, and also in the intestinal peritrophic mem- brane. How N. virens copes with mercury toxicity is dis- cussed.
Introduction
It is known that marine polychaetes accumulate mercury from the environment (Luoma 1977a, b, Kendall 1978, Medeiros etal. 1981) and they have therefore been em- ployed in marine pollution monitoring programs to detect the degree of mercury pollution (Bryan et al. 1980, Kiorboe et al. 1983, Langston 1986).
Mercury is highly toxic to adult (Brown and Ah- sanullah 1971, Reish et al. 1976, Reish 1978) and juvenile (Reish et al. 1976) marine polychaetes. Sublethal concen- trations of mercury cause suppression of reproduction (Reish 1978) and have been reported to inhibit the ab-
sorption of glycine across the body surface of the poly- chaete Nereis diversicolor (Siebers and Ehlers 1979).
On the other hand, only little information regarding the sites of accumulation within polychaetes and the handling of mercury by these animals is available. Luoma (1977b) used autoradiography to show the distribution of 2~ in whole specimens of Neanthes succinea. However, autoradi- ography is unacceptable at higher resolution because of the relatively energetic beta-radiation from the 2~ isotope.
Autometallography is a sensitive method for light and electron microscopic demonstration of mercury deposits within organs and cells. According to Danscher and MM- ler-Madsen (1985), accumulations of possibly less than ten mercury atoms bound to sulphide or selenide can be made visible.
In the present study, autometallography has been ap- plied to demonstrate mercury deposits in tissues of the polychaete Nereis virens (Sars) exposed to sublethal con- centrations of water-borne mercury chloride.
Materials and methods
Animals
Specimens of the polychaete Nereis virens (Sars) were col- lected from Kysing Fjord, a shallow, Inesohaline estuary near Aarhus in Denmark. Water temperature and salinity at the sampling site was 9 ~ and 19%0 S, respectively. Only intact worms were used for the experiments.
In order to obtain homogeneous material, 30 worms with average length of 5-+0.7cm and weighing 340-+ 10 mg (wet wt) were used. According to the length/ age relationship reported by Kristensen (1984), the worms in the present study were estimated to be approximately 1.5 yr old. Prior to experimentation, the nereids were placed in a darkened aquarium for 24 h in order to void their guts. The worms were not fed during the experimen- tal period.
534 P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues
Quantitative measurements
The amounts of mercury in the water and within the worms were determined using radiolabelled mercury. The experiment was performed in a 20-1itre darkened aquari- um filled with 14 litre of pre-filtered seawater, adjusted to a salinity of 20%o S and a temperature of 12 ~ The water was aerated to oxygen saturation (9.5 mg 02 1-1).
The nereids were placed in 10 to 12 cm long trans- parent polyethylene tubes with an inner diameter of 4.5 ram, simulating the worms' natural burrow within the sediment. A 1 mm mesh, placed at either end of the tube, prevented the worm from leaving the tube. The nereids were allowed to acclimatize to the experimental conditions for 36 h. During this period the worms made periodic un- dulating movements.
Following acclimatization, 14 ml of a mercury stock so- lution was added to the water. This solution contained 10 big Hg ml-1 (as HgC12) including 0.37 MBq 2~ (as 2~ Thus, a final concentration of 0.37 MBq and 10/~gHg -1 1-1 of water (10ppb) was obtained in the aquarium. The 2~ isotope was purchased from Research Establishment Riso, Denmark (as 2~ and unla- belled HgC12 of analytical grade from Merck, Darmstadt, FRG. The mercury concentration in the water was assessed twice daily by measuring the radioactivity emitted from a 5 ml water sample. The radioactivity was measured in a 2 x l~r (5.0 x 4.45 cm) NaI (T1) well-crystal connected to a one-channel analyser. During the course of the measurements, the mercury concentration in the water decreased by between 5 and 25%. Sufficient stock solution was then added in order to reach the initial concentration. Thus, an average mercury concentration of 9 ppb was ob- tained.
The uptake of mercury by the worms was determined by measuring the radioactivity emitted from the absorbed 2~ at various time intervals. Prior to the measurements, the worms were carefully removed from their tubes and then rinsed in uncontaminated seawater from the sampling site. The amount of mercury in each worm was determined by comparing the counting rate with that of an 'artificial worm' in the form of a plastic tube containing a known quantity of mercury.
Preparation for microscopy
At the end of the experimental period, three worms treated with mercury and two untreated (controls) were prepared for microscopy. Each worm was dissected in a fixative con- taining 1% paraformaldehyde and 2% glutaraldehyde in 0.15 M phosphate buffer (pH 7.2). The following parts of the worms were sampled: (1) the head region, including the prostomium and the peristomium; (2) the oesophagus and the oesophageal diverticulum; (3) Segments Nos. 25 and 26 counted from the peristomium; and (4) the hind- most three segments, together with the pygidium. The tis- sue samples were immersed in the fixative as described
above for 3 h at room temperature. The tissues were then rinsed in the buffer, dehydrated and subsequently embed- ded in Epon 812. Sections 3/~m thick were cut and fas- tened to glass slides on a heating plate at 40 ~
The mercury accumulations in the tissues were made visible for microscopy according to the autometallographic method of Danscher and Moller-Madsen (1985) (see also Danscher et al. 1987). Briefly, this method involves silver enhancement of mercury accumulations associated with sulphide or selenide. The mercury sulphide or mercury selenide complexes act as catalysts (electron carriers). When the mercury-containing tissue is treated with silver ions (electron acceptors) and a reducing agent (electron doner), silver ions adhering to the surface of the complex of mercury sulphide or mercury selenide will be trans- formed into metallic silver. Elemental silver formed in this way is itself catalytic and the process thus continues; conse- quently, the mercury becomes encapsulated in layers of sil- ver. The location of mercury can subsequently be demon- strated by light and electron microscopy. Prior to devel- opment the slides were coated with 0.5% gelatin. The de- veloper was made up immediately before use by mixing (1) 60 ml gum arabic; (2) 15 ml reducing agent (0.51 M hydro- quinone); (3) 10ml buffer (0.80M sodium citrate in 1.27 M citric acid); (4) 15 ml silver ion supply (0.03 M sil- ver lactate). The slides were placed in jars filled with the developer and kept in a dark box in a 26 ~ water bath for 70 min. After development the slides were washed in 40 ~ hot water for 20 rain in order to remove the gelatin coat. The slides were then dipped in Farmer's solution (9 parts of 10% sodium thiosulfate plus 1 part of 10% potassium fer- ricyanid) for 10 s and rinsed in distilled water. Hereafter the slides were dipped in a film fixative for 30 s and rinsed in distilled water.
ppm Hg
lO
9
8
7
6
5
4
3
2
1
D a y s i i i i i i r i i I J
1 2 3 4 5 6 7 8 9 10 11
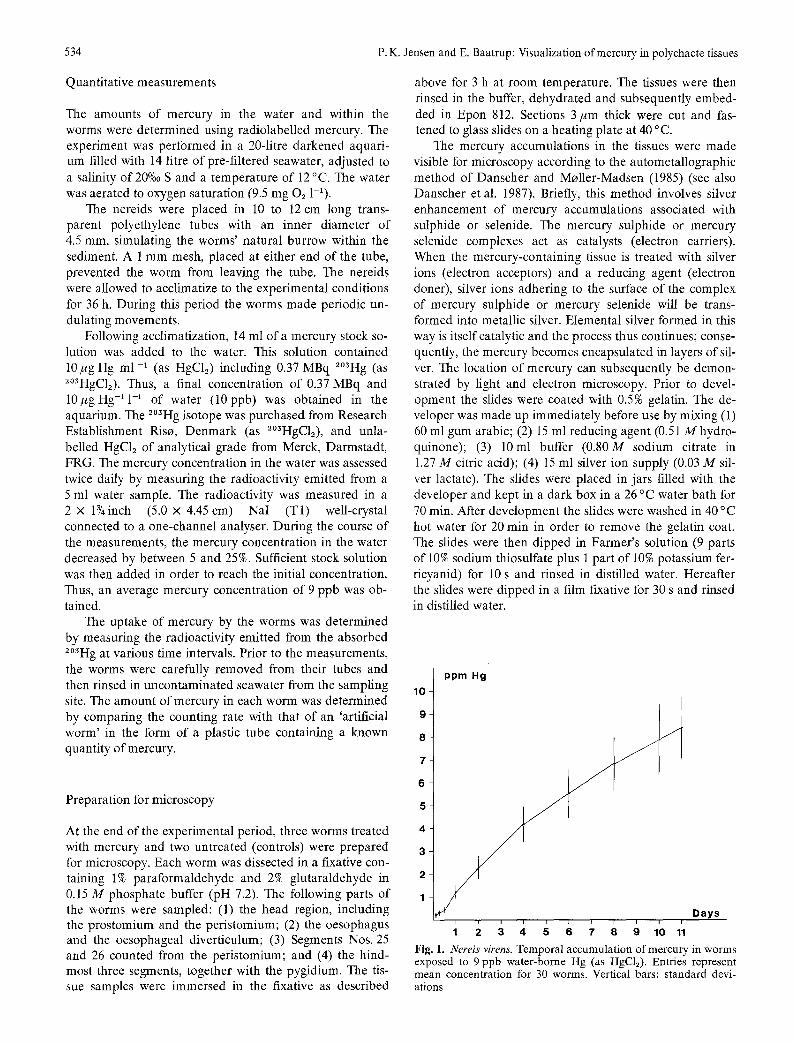
Fig. l. Nereis virens. Temporal accumulation of mercury in worms exposed to 9 ppb water-borne Hg (as HgC12). Entries represent mean concentration for 30 worms. Vertical bars: standard devi- ations

P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues 535
Fig. 2. Nereis virens. Light micrograph showing the intestinal epithelium. Following autometallographic silver enhancement, numerous silver grains are visible in sections from worms treated with Hg, predominantly in the apical part of the epithelium (A), whereas sections from worms not treated with Hg (controls; B) are void of silver grains. Scale bars = 20/~m
Fig. 3. Nereis virens. Apical portion of an epithelial cell from the intestine of a worm exposed to HgC12 for 11 d. The deposits of Hg are almost exclusively located within lysosome-like organelles. Scale bar = 1 pm
Following light microscopy, selected sections were covered with a drop of liquid Epon, and an Epon block was then placed on top. After polymerization, the block plus attached section were removed from the slide by briefly heating on a 90 ~ heating plate. Thin sections were cut for electron microscopy and then counterstained with lead citrate and uranyl acetate.
Results
Uptake of mercury
Nereis virens absorbed mercury from the water. The con- centration of mercury in N. virens increased with increasing exposure time, reaching an average concentration of

536 P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues
Fig. 4. Nereis virens. In worms exposed to HgC12 an extensive deposition of mercury appears in the tubular cells of the convoluted neph- ridial ducts. (A) and (B). Appearance of silver-enhanced Hg under normal light microscopy and dark field microscopy, respectively. Scale bars = 30 #m
Fig. 5. Nereis virens. Ultrastructurally, silver-enhanced Hg is located within lysosomes (ly) of the nephridial tubular cells, tl: tubular lu- men. Scale bar = 1 #m
8.41 + 1.4 ppm Hg after 11 d of treatment. Compared with the average concentration o f 9 ppb Hg in the water, the bioconcentration factor (i.e., the concentration of mercury in the worm divided by the mercury concentration in the water) was determined to be 930 after 11 d. The uptake of mercury by N. virens from the water as a function of time is given in Fig. 1.
Locations of mercury accumulations
As the nereids used in this work were rather small, 3/~m thick cross-sections of whole individuals could be obtained. Following autometallographic staining, deposits of silver- enhanced mercury were observed in the intestine, nephri- dia, epidermis, and in the cuticula of mercury-treated indi-
P. K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues 537
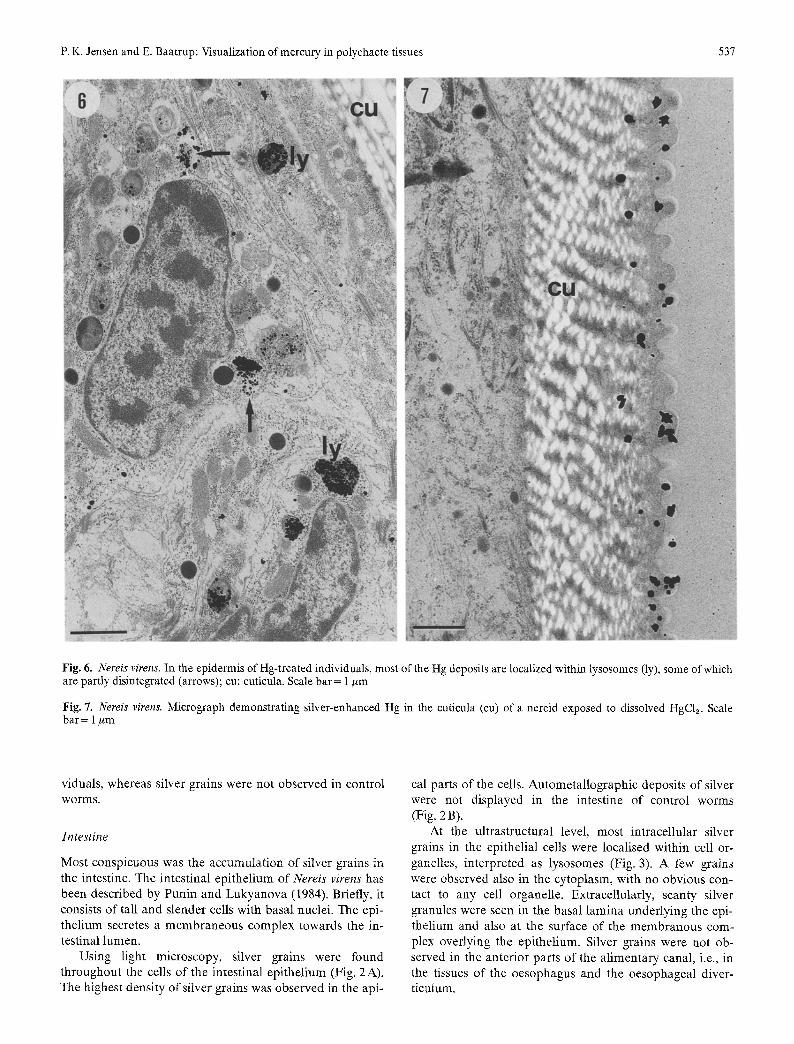
Fig. 6. Nereis virens. In the epidermis of Hg-treated individuals, most of the Hg deposits are localized within lysosomes (ly), some of which are partly disintegrated (arrows); cu: cuticula. Scale bar= 1 ~m
Fig. 7. Nereis virens. Micrograph demonstrating silver-enhanced Hg in the cuticula (cu) of a nereid exposed to dissolved HgCla. Scale bar = 1 #rn
viduals, whereas silver grains were not observed in control worms.
In tes t ine
Most conspicuous was the accumulation of silver grains in the intestine. The intestinal epithelium of Nereis virens has been described by Punin and Lukyanova (1984). Briefly, it consists of tall and slender cells with basal nuclei. The epi- thelium secretes a membraneous complex towards the in- testinal lumen.
Using light microscopy, silver grains were found throughout the cells of the intestinal epithelium (Fig. 2A). The highest density of silver grains was observed in the api-
cal parts of the cells. Autometallographic deposits of silver were not displayed in the intestine of control worms (Fig. 2 B).
At the ultrastructural level, most intracellular silver grains in the epithelial cells were localised within cell or- ganelles, interpreted as lysosomes (Fig. 3). A few grains were observed also in the cytoplasm, with no obvious con- tact to any cell organelle. Extracellularly, scanty silver granules were seen in the basal lamina underlying the epi- thelium and also at the surface of the membranous com- plex overlying the epithelium. Silver grains were not ob- served in the anterior parts of the alimentary canal, i.e., in the tissues of the oesophagus and the oesophageal diver- ticulum.

538 P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues
Nephridia
The nephridia of nereids are metanephridia. Each neph- rostome opens into the coelomic cavity and the convoluted nephridial duct opens on the exterior surface. The tubular cells lining the inner surface of the nephridial duct are ciliated. A more detailed description of nephridial mor- phology in nereids has been given by Jones (1957).
Under the light microscope, silver grains were observed in the tubular cells of mercury-treated worms (Fig. 4). At the ultrastructural level, the silver deposits were localized within lysosomes of the tubular cells (Fig. 5). A few silver grains were also found on the luminal surface of blood vessels passing by the nephridia. The nephridia of worms not treated with mercury were void of silver deposits.
Epidermis
In the epidermis, silver grains were difficult to distinguish from pigment granules using light microscopy, but the dif- ference became evident with electron microscopy (Fig. 6). In the cells of the epidermis, the highest density of silver deposits was observed within lysosomes, but a few grains were seen also in the cytoplasmic matrix outside the ly- sosomes. Extracellularly, single grains were observed in the basal lamina subjacent to the epithelium.
Cuticula
In the cuticula, silver-enhanced mercury was observed on the epicuticular expansions and in the collagenous fibres of the endocuticula (Fig. 7). Silver grains were not observed in the epidermis and the cuticle of controls.
Discussion
The marine polychaete Nereis virens rapidly retained dis- solved mercury from the water. This is in accordance with observations on other marine invertebrates (Fowler et al. 1978, Jensen et al. 1981, Riisggtrd et al. 1985, Riisggrd and Famine 1986), including marine polychaetes (Luoma 1977a, b, Kendall 1978). The uptake of mercury was rela- tively constant during the 11 d of mercury treatment, which is in accordance with Kendall (1978), who exposed N. suceinea to low concentrations of HgC12 over a 72 d period without observing any plateau effect.
The bioconcentration factor found for Nereis virens is comparable to those found for other marine invertebrates (Luoma 1977a, Fowler etal. 1978, Riisg~rd and Famme 1986). It is, however, difficult to compare the bio- concentration factors obtained in the various studies be- cause of different experimental conditions. For example, even within the same species the accumulation of mercury depends on the body size (Fowler et al. 1978), the ambient temperature of the water (Kendall 1978, Denton and Bur- don-Jones 1981), its salinity (Luomo 1977a, Axiak and Schembri 1982), and the physiochemical form of mercury (Fowler et al. 1978, Nagashima et al. 1983, Riisggtrd and
Famine 1986). Between species, additional factors such as anatomical and physiological differences influence the de- gree of mercury accumulation (see Windom and Kendall 1979).
The autometallographic method has been worked out for microscopical demonstration of metallic gold and sil- ver, and sulphides and selenides of zinc and mercury (Danscher 1984, Danscher and Moller-Madsen 1985, Baatrup et al. 1986, Danscher et al. 1987). Previously, the technique had not been applied to invertebrates, but the fact that silver grains appeared only in those worms treated with mercury, and not in the controls, suggests strongly that mercury was the catalytic substrate for silver enhance- ment in Nereis virens. Autometallography allows determi- nation of the distribution of mercury deposits within or- ganisms and organs of sizes beyond the range of other methods. Such a localization can indicate the target organs of mercury accumulation. Although quantitative measure- ments have not yet been applied to the autometallographic method, the density of silver grains in the tissues provides information regarding the relative distribution of mercury among organs and tissues and thereby suggests how nereids cope with mercury toxicity.
The presence of silver-enhanced mercury in cells of the intestine, nephridia, epidermis, and also in the two cuticu- lar layers, demonstrates that mercury was absorbed by the nereids, which is in accordance with Luoma (1977 b).
In the intestine, heavy accumulations of silver-en- hanced mercury were found in the apical part of the epi- thelium, and also in the membranous complex overlying the epithelium. This complex forms the peritrophic mem- brane which has been reported to surround the faeces of nereids (see Michel and De Villez 1978). It is not known whether mercury was absorbed or excreted across the epi- thelium of the digestive tract. However, as the worms were not fed during the experimental period, and because Fletcher (1974) showed that nereids do not pass water through their digestive tract, our results favour the hypoth- esis that when the worm is exposed to water-borne mer- cury, mercury is excreted into the intestinal lumen rather than absorbed from it. Accumulations of heavy metals have been found in the intestinal wall of other armelids (Ireland and Richards 1977, Andersen and Laursen 1982), and there is evidence that the nereid intestine is involved in the handling of exogenous substances (Zeeck 1980).
In the nephridia, mercury was accumulated in the fin- ing of the nephridial tubules. When compared with the vertebrate kidney, this finding parallels the high density of mercury deposits in the proximal tubular epithelium offish exposed to water-borne mercury (Baatrup et al. 1986). It is not known at present whether the visualized mercury de- posits in the nereid tubular epithelium originated from the blood or whether they had been absorbed from the tubular lumen. The nereid metanephridium is, however, known to be an excretory organ (Fretter and Graham 1976, Barnes 1980) which is involved in the turnover of exogenous ma- terials (see Oglesby 1978). It is possible, therefore, that nephridial excretion of mercury in nereids does occur.

P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues 539
Accumulations of mercury were also found in the cuticula and in the cells of the epidermis. An epidermal absorption of mercury cannot be concluded from the present study, but the mercury chloride was administered in the aquari- um water and nereids are believed to use their surface for respiration (Fretter and Graham 1976). Since the mercury was probably not taken up from the gut, as discussed above, an epidermal absorption of water-borne mercury is therefore likely to occur in nereids. It has previously been shown that mercury is rapidly accumulated in the respi- ratory organs of marine molluscs (Denton and Burdon- Jones 1981, Nagashima et al. 1983) and crustaceans (Vern- berg and Vernberg 1972, Ray and Yripp 1976).
The finding that the mercury present in all three tissues was localized predominantly within lysosome-like bodies accords with the lysosomal accumulation of mercury in fish (Baatrup et al. 1986) and mammal ian (Norseth and Bren- deford 1971, Danscher and Rungby 1986, Thorlacius-Uss- ing and Moller Graab~ek 1986) tissues. Fowler et al. (1975) observed a dose-related increase of lysosome-like bodies in the mantle tentacle epithelial cells of mercury-treated clams and suggested that these ceils deposit mercury within lysosomes in order to avoid more harmful effects. Likewise, but using the rubeanic acid method, Bryan and Hum- merstone (1971) showed that copper appears as granular deposits in the epidermis and the nephridia of Nereis diver- sicolor collected from copper-contaminated estuarine sedi- ments. Granular accumulations of copper were found also by X-ray microanalysis within marine Crustacea (Walker 1977) and molluscs (George et al. 1978). Thus, it appears that granular deposition is a common response o f inverte- brates to heavy metal intoxication.
Studies on Nereis Wrens sampled at a mercury-polluted area are in progress. Preliminary results suggest that the distribution of Hg in nereids exposed under natural con- ditions parallel the present experimental findings.
Acknowledgements. This work was supported by Aarhus University Research Foundation and The Danish Research Council's Pro- gram for Technical Development, assigned to Dr. G. Danscher. We thank Dr. G. Danscher for the subsidy. The authors are also grateful to Dr. J. G. Mather for linguistic corrections and com- ments on the manuscript. Further, we acknowledge the technical assistance of Mrs. H. B. Andersen, Mrs. Pia Kj~ergaard Moller, Mrs. E. Frost, Mr. Bj. Krunderup, Mr. A. Meier, and Mr. A. Kjaer.
Literature cited
Andersen, C., Laursen, J. (1982). Distribution of heavy metals in Lumbrieus terrestris, Aporreetodea longa and A. rosea mea- sured by atomic absorption and X-ray fluorescence spec- trometry. Pedobiologia 24:347-356
Axiak, V., Schembri, J. L. (1982). Effects of temperature on the toxicity of mercury and cadmium to the littoral gastropod Monodonta turbinata. Mar. Pollut. Bull. 13:383-386
Baatrup, E., Nielsen, M. G., Danscher, G. (1986). Histochemical demonstration of two mercury pools in trout tissues: mercury in kidney and liver after mercuric chloride exposure. Eeotoxi- col. Envir. Saf. 12:267-282
Barnes, R. D. (1980). Invertebrate zoology. Holt-Saunders, Tokyo
Brown, B., Ahsanullah, M. (1971). Effect of heavy metals on mor- tality and growth. Mar. Pollut. Bull. 2:182-187
Bryan, G. W., Hummerstone, L. G. (1971). Adaptation of the poly- chaete Nereis diversicolor to estuarine sediments containing high concentrations of heavy metals I. General observation and adaptation to copper. J. mar. biol. Ass. U.K. 51:845-863
Bryan, G. W., Langston, W. J., Hummerstone, L. G. (1980). The use of biological indicators of heavy metal contamination in es- tuaries; with special reference to an assessment of the biologi- cal availability of metals in estuarine sediment from South- West Britain. Occ. Publs. mar. biol. Ass. U.K. 1:1-73
Danscher, G. (1984). Autometallography. A new technique for light and electron microscopic visualization of metals in bio- logical tissues (gold, silver, metal sulphides and metal selenides). Histochemistry 81: 331-335
Danscher, G., Moller-Madsen, B. (1985). Silver amplification of mercury sulfide and selenide: a histochemical method for light and electron microscopic localization of mercury in tissue. J. Histochem. Cytochem. 33:219-228
Danscher, G., Norgaard, J. O. R., Baatrup, E. (1987). Autometal- lography: tissue metals demonstrated by a silver enhancement kit. Histochemistry 86:465-469
Danscher, G., Rungby, J. (1986). Differentiation of histochemi- cally visualized mercury and silver. Histochem. J. 18:109-114
Denton, G. R. W., Burdon-Jones, C. (1981). Influence of tempera- ture and salinity on the uptake, distribution and deputation of mercury, cadmium and lead by the black-lip oyster Saccostrea echinata. Mar. Biol. 64:317-326
Fletcher, C. R. (1974). Volume regulation in Nereis diversicolor I. The steady state. Comp. Biochem. Physiol. 47 A: 1199-1214
Fowler, S. W., Heyraud, M., La Rosa, J. (1978). Factors affecting methyl and inorganic mercury dynamics in mussels and shrimp. Mar. Biol. 46:267-276
Fowler, B. A., Wolfe, D. A., Hettler, W. F. (1975). Mercury and iron uptake by cytosomes in mantle epithelial cells of quahog clams (Mercenaria mercenaria) exposed to mercury. J. Fish. Res. Bd. Can. 32:1767-1775
Fretter, V., Graham, A. (1976). A functional anatomy of inverte- brates. Academic Press, London
George, S. G., Pirie, B. J. S., Cheyne, A. R., Coombs, L., Grant, P. T. (1978). Detoxication of metals by marine bivalves: An ul- trastructural study of the compartmentation of copper and zinc in the oyster Ostrea edulis. Mar. Biol. 45:147-156
Ireland, M. P., Richards, K. S. (1977). The occurrence and local- isation of heavy metals and glycogen in the earthworms Lum- bricus rubellus and Dendrobaena rubida from a heavy metal site. Histoehemistry 51: 153-166
Jensen, K., Randlov, A., Riisggtrd, H. U. (1981). Heavy metal pol- lution from a point source demonstrated by means of mussels, Mytilus edulis. Chemosphere (U.K.) 10:761-765
Jones, M. L. (1957). On the morphology of the nephridium of Nereis vexillosa Grube. Biol. Bull. mar. biol. Lab., Woods Hole 113:407-413
Kendall, D. R. (1978). The role of macrobenthic organisms in mer- cury, cadmium, copper and zinc transfers in Georgia salt marsh ecosystems. Ph.D. thesis, Emory University
Kiorboe, T., Mohlenberg, F., Riisg~rd, H. U. (1983). Mercury levels in fish, invertebrates and sediment in a recently recorded polluted area (Nissum Broad, Western Limt]ord, Denmark) Mar. Pollut. Bull. 14:21-24
Kristensen, E. (1984). Life cycle, growth and production in estuar- ine populations of the polychaetes Nereis Wrens and N. diver- sieolor. Holarct. Ecol., Copenhagen 7:249-256
Langston, W. J. (1986). Metals in sediments and benthic organisms in the Mersey estuary. Estuar. coast. Shelf Sci. 23:239-261
Luoma, S. N. (1977 a). The dynamics of biologically available mer- cury in a small estuary. Estuar. coast, mar. Sci. 5:643-652
Luoma, S. N. (1977 b). Physiological characteristics of mercury up- take by two estuarine species. Mar. Biol. 41 : 269-273
Medeiros, D. M., Cadwell, L. L., Preston, R. L. (1981). A possible physiological uptake mechanism of methylmercury by the ma-

540 P.K. Jensen and E. Baatrup: Visualization of mercury in polychaete tissues
rine bloodworm (Glycera dibranchiata). Bull. envir. Contam. Toxicol. 24:97-101
Michel, C., De Villez, J., (1978). Digestion. In: Mill, P. J. (ed.) Physiology of annelids. Academic Press, London, p. 509-554
Nagashima, Y., Shiomi, K., Yamanaka, H., Kikuchi, T. (1983). Ac- cumulation of mercury by tissues in the short-necked clam Tapes japonica. Bull. Jap. Soc. scient. Fish. 49:801-804
Norseth, T., Brendeford, M. (1971). Intracellular distribution of inorganic and organic mercury in rat liver after exposure to methylmercury salts. Biochem. Pharm. 20:1101-1107
Oglesby, L. C. (1978). Salt and waterbalance. In: Mill, P. J. (ed.) Physiology of annelids. Academic Press, London, p. 555-658
Punin, M. Y., Lukyanova, E. L. (1984). Midgut epithelium organi- zation of the errant polychaete Nereis Wrens. I. Absorptive and secretory ceils. Tsitologiya 26:60-68
Ray, G. L., Tripp, M. R. (1976). The uptake of mercury from water by the grass shrimp, Palaemonetes vulgaris (Say). J. envirl. Qual. 5:193-197
Reish, D. J. (1978). The effects of heavy metals on polychaetous annelids. Revue int. Oc6anogr. retd. 49:99-104
Reish, D. J., Martin, J. M., Piltz, F. M., Word, J. Q. (1976). The ef- fect of heavy metals on laboratory populations of two poly- chaetes with comparisons to the water quality conditions and standards in Southern California marine waters. Wat. Res. 10: 299-3O2
Riisg~rd, H. U., Famme, P. (1986). Accumulation of inorganic and organic mercury in shrimp, Crangon crangon. Mar. Pollut. Bull. 17:255-257
Riisggtrd, H. U., Kiorboe, T., Mohlenberg, F., Drabeek, I., Pheiffer Madsen, P. (1985). Accumulation, elimination and chemical speciation of mercury in the bivalves Mytilus edulis and Ma- coma balthica. Mar. Biol. 86:55-62
Siebers, D., Ehlers, U. (1979). Heavy metal action on trans- integumentary absorption of glycine in two annelid species. Mar. Biol. 50:175-179
Thorlacius-Ussing, O., Moller Graab~ek, P. (1986). Simultaneous ultrastructural demonstration of heavy metals (silver, mercury) and acid phosphatase. Histochem. J. 18:639-646
Vernberg, W. B., Vernberg, J. (1972). The synergistic effects of temperature, salinity, and mercury on survival and metabolism of the adult fiddler crab, Uca pugilator. Fish. Bull. U.S. 70: 415-420
Walker, G. (1977). Copper granules in the barnacle Balanus balanoides. Mar. Biol. 39:343-349
Windom, H. L., Kendall, D. R. (1979). Accumulation and bio- transformation of mercury in coastal and marine biota. In: Nri- agua, J. O. (ed.) The biogeochemistry of mercury in the en- vironment. Biomedical Press, Elsevier/North-Holland, p. 303-323
Zeeck, E. (1980). Determination and localization of oil com- ponents in living benthic organisms by fluorescence mi- croscopy. Helgol~inder wiss. Meeresunters. 33:272-277
Date of final manuscript acceptance: October 26, 1987. Communicated by T. Fenchel, Helsingsr
Related Documents











