High level of resistance to Sclerotinia sclerotiorum in introgression lines derived from hybridization between wild crucifers and the crop Brassica species B. napus and B. juncea Harsh Garg a , Chhaya Atri b , Prabhjodh S. Sandhu b , Balvir Kaur b , Michael Renton a,c , Shashi K. Banga b , Hardeep Singh b , Charandeep Singh b , Martin J. Barbetti a,d, *, Surinder S. Banga b a School of Plant Biology, Faculty of Natural and Agricultural Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia b Department of Plant Breeding and Genetics, Punjab Agricultural University, Ludhiana, 141004 Punjab, India c CSIRO Sustainable Ecosystems, Floreat, WA 6014, Australia d Department of Agriculture and Food Western Australia, Baron-Hay Court, South Perth, WA 6151, Australia 1. Introduction Sclerotinia disease, caused by the fungal pathogen Sclerotinia sclerotiorum, is a serious threat to oilseed rape production with substantial yield losses recorded worldwide including India, Europe, China, North America and Australia (Li et al., 1999; McCartney et al., 1999; Sprague and Stewart-Wade, 2002; Hind et al., 2003; Koch et al., 2007; Malvarez et al., 2007; Singh et al., 2008). Various methods used for managing Sclerotinia disease include cultural control, chemical control and varietal resistance (Bardin and Huang, 2001). The persistent nature of sclerotia and the wide host range of this pathogen from taxonomically diverse hosts (over 408 plant species) generally render cultural practices such as crop rotation to be ineffective (Williams and Stelfox, 1980; Boland and Hall, 1994). Further, disease management through chemical control is also largely ineffective due to difficulty in timing the fungicide application with the release of ascospores (Bolton et al., 2006). Host resistance offers the only economic and sustainable method for effectively managing this disease (Zhao et al., 2004; Li et al., 2006). While partial resistance against this pathogen has been observed in certain genotypes of sunflower (Helianthus annuus) (Godoy et al., 2005), beans (Phaseolus coccineus)(Gilmore et al., 2002), peas (Pisum sativum)(Porter et al., 2009), peanut (Arachis hypogea)(Cruickshank et al., 2002), or soybean (Glycine max) (Hartman et al., 2000), complete resistance has not been reported in the field. Partial resistance was also identified in some of the Brassica napus and, to a lesser extent B. juncea, genotypes from China (Li et al., 1999, 2006, 2008; Zhao et al., 2004), Australia (Li et al., 2006, 2008) and India (Singh et al., 2008). Although, a Field Crops Research 117 (2010) 51–58 ARTICLE INFO Article history: Received 29 September 2009 Received in revised form 11 December 2009 Accepted 29 January 2010 Keywords: Erucastrum cardaminoides Diplotaxis tenuisiliqua E. abyssinicum Brassicas Introgression lines Resistance White mold ABSTRACT Sclerotinia rot caused by the fungus Sclerotinia sclerotiorum is one of the most serious and damaging diseases of oilseed rape and there is keen worldwide interest to identify Brassica genotypes with resistance to this pathogen. Complete resistance against this pathogen has not been reported in the field, with only partial resistance being observed in some Brassica genotypes. Introgression lines were developed following hybridization of three wild crucifers (viz. Erucastrum cardaminoides, Diplotaxis tenuisiliqua and E. abyssinicum) with B. napus or B. juncea. Their resistance responses were characterized by using a stem inoculation test. Seed of 54 lines of B. napus and B. juncea obtained from Australia, India and China through an Australian Centre for International Agricultural Research (ACIAR) collaboration programme were used as susceptible check comparisons. Introgression lines derived from E. cardaminoides, D. tenuisiliqua and E. abyssinicum had much higher levels (P < 0.001) of resistance compared with the ACIAR germplasm. Median values of stem lesion length of introgression lines derived from the wild species were 1.2, 1.7 and 2.0 cm, respectively, as compared with the ACIAR germplasm where the median value for stem lesion length was 8.7 cm. This is the first report of high levels of resistance against S. sclerotiorum in introgression lines derived from E. cardaminoides, D. tenuisiliqua and E. abyssinicum. The novel sources of resistance identified in this study are a highly valuable resource that can be used in oilseed Brassica breeding programmes to enhance resistance in future B. napus and B. juncea cultivars against Sclerotinia stem rot. ß 2010 Elsevier B.V. All rights reserved. * Corresponding author at: School of Plant Biology, Faculty of Natural and Agricultural Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia. Tel.: +61 8 64883924; fax: +61 8 64887077. E-mail address: [email protected] (M.J. Barbetti). Contents lists available at ScienceDirect Field Crops Research journal homepage: www.elsevier.com/locate/fcr 0378-4290/$ – see front matter ß 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.fcr.2010.01.013

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Field Crops Research 117 (2010) 51–58
High level of resistance to Sclerotinia sclerotiorum in introgression lines derivedfrom hybridization between wild crucifers and the crop Brassica species B. napusand B. juncea
Harsh Garg a, Chhaya Atri b, Prabhjodh S. Sandhu b, Balvir Kaur b, Michael Renton a,c, Shashi K. Banga b,Hardeep Singh b, Charandeep Singh b, Martin J. Barbetti a,d,*, Surinder S. Banga b
a School of Plant Biology, Faculty of Natural and Agricultural Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australiab Department of Plant Breeding and Genetics, Punjab Agricultural University, Ludhiana, 141004 Punjab, Indiac CSIRO Sustainable Ecosystems, Floreat, WA 6014, Australiad Department of Agriculture and Food Western Australia, Baron-Hay Court, South Perth, WA 6151, Australia
A R T I C L E I N F O
Article history:
Received 29 September 2009
Received in revised form 11 December 2009
Accepted 29 January 2010
Keywords:
Erucastrum cardaminoides
Diplotaxis tenuisiliqua
E. abyssinicum
Brassicas
Introgression lines
Resistance
White mold
A B S T R A C T
Sclerotinia rot caused by the fungus Sclerotinia sclerotiorum is one of the most serious and damaging
diseases of oilseed rape and there is keen worldwide interest to identify Brassica genotypes with
resistance to this pathogen. Complete resistance against this pathogen has not been reported in the field,
with only partial resistance being observed in some Brassica genotypes. Introgression lines were
developed following hybridization of three wild crucifers (viz. Erucastrum cardaminoides, Diplotaxis
tenuisiliqua and E. abyssinicum) with B. napus or B. juncea. Their resistance responses were characterized
by using a stem inoculation test. Seed of 54 lines of B. napus and B. juncea obtained from Australia, India
and China through an Australian Centre for International Agricultural Research (ACIAR) collaboration
programme were used as susceptible check comparisons. Introgression lines derived from E.
cardaminoides, D. tenuisiliqua and E. abyssinicum had much higher levels (P < 0.001) of resistance
compared with the ACIAR germplasm. Median values of stem lesion length of introgression lines derived
from the wild species were 1.2, 1.7 and 2.0 cm, respectively, as compared with the ACIAR germplasm
where the median value for stem lesion length was 8.7 cm. This is the first report of high levels of
resistance against S. sclerotiorum in introgression lines derived from E. cardaminoides, D. tenuisiliqua and
E. abyssinicum. The novel sources of resistance identified in this study are a highly valuable resource that
can be used in oilseed Brassica breeding programmes to enhance resistance in future B. napus and B.
juncea cultivars against Sclerotinia stem rot.
� 2010 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Field Crops Research
journa l homepage: www.e lsev ier .com/ locate / fc r
1. Introduction
Sclerotinia disease, caused by the fungal pathogen Sclerotinia
sclerotiorum, is a serious threat to oilseed rape production withsubstantial yield losses recorded worldwide including India,Europe, China, North America and Australia (Li et al., 1999;McCartney et al., 1999; Sprague and Stewart-Wade, 2002; Hindet al., 2003; Koch et al., 2007; Malvarez et al., 2007; Singh et al.,2008). Various methods used for managing Sclerotinia diseaseinclude cultural control, chemical control and varietal resistance(Bardin and Huang, 2001). The persistent nature of sclerotia andthe wide host range of this pathogen from taxonomically diverse
* Corresponding author at: School of Plant Biology, Faculty of Natural and
Agricultural Sciences, The University of Western Australia, 35 Stirling Highway,
Crawley, WA 6009, Australia. Tel.: +61 8 64883924; fax: +61 8 64887077.
E-mail address: [email protected] (M.J. Barbetti).
0378-4290/$ – see front matter � 2010 Elsevier B.V. All rights reserved.
doi:10.1016/j.fcr.2010.01.013
hosts (over 408 plant species) generally render cultural practicessuch as crop rotation to be ineffective (Williams and Stelfox, 1980;Boland and Hall, 1994). Further, disease management throughchemical control is also largely ineffective due to difficulty intiming the fungicide application with the release of ascospores(Bolton et al., 2006). Host resistance offers the only economic andsustainable method for effectively managing this disease (Zhaoet al., 2004; Li et al., 2006).
While partial resistance against this pathogen has beenobserved in certain genotypes of sunflower (Helianthus annuus)(Godoy et al., 2005), beans (Phaseolus coccineus) (Gilmore et al.,2002), peas (Pisum sativum) (Porter et al., 2009), peanut (Arachis
hypogea) (Cruickshank et al., 2002), or soybean (Glycine max)(Hartman et al., 2000), complete resistance has not been reportedin the field. Partial resistance was also identified in some of theBrassica napus and, to a lesser extent B. juncea, genotypes fromChina (Li et al., 1999, 2006, 2008; Zhao et al., 2004), Australia (Liet al., 2006, 2008) and India (Singh et al., 2008). Although, a

Fig. 1. The general breeding scheme used to introgress segments of the genome
from wild species Erucastrum cardaminoides into cultivated lines of Brassica juncea.
Fig. 2. The schematic representation for developing introgression lines by crossing
Brassica napus with Erucastrum cardaminoides.
Fig. 3. The schematic representation for the development of introgression lines
from wild crucifers Erucastrum abyssinicum and Diplotaxis tenuisiliqua.
H. Garg et al. / Field Crops Research 117 (2010) 51–5852
significant number of at least partially resistant genotypes havebeen identified, breeding to increase the levels of resistance againstSclerotinia disease, in B. napus and/or B. juncea has been ineffective.This is mainly because resistance to S. sclerotiorum in existingcultivars of Brassica and in other cultivated germplasm appears tobe of a complex nature, i.e., it can either be monogenic and/orpolygenic depending on the different plant species and materialsunder investigation (Abawi et al., 1978; Baswana et al., 1991; Zhaoand Meng, 2003; Zhao et al., 2006). Genotypes with higher levels ofresistance are urgently required for inclusion in oilseed Brassica
breeding programmes to enhance the level of field resistance incultivated species such as B. napus and B. juncea.
Lack of effective resistance to Sclerotinia disease in cultivatedspecies has stimulated the interest of researchers towardsexploitation of wild crucifer species to diversify the existing genepool. Higher levels of resistance against Sclerotinia have alreadybeen reported in the secondary gene pool of bean (Abawi et al.,1978; Gilmore et al., 2002; Schwartz et al., 2006), wild Helianthus
species (Seiler, 1992; Gulya et al., 2009) and in a Pisum corecollection (Porter et al., 2009). Several successful attempts havebeen reported to introgress the resistance from the secondary genepool of bean (Phaseolus vulgaris) into the cultivated bean speciesthrough interspecific hybridization followed by backcrossing (e.g.,Schwartz et al., 2006; Singh et al., 2009). Introgression of genomicsegments responsible for resistance against Sclerotinia from wildto cultivated species of sunflower has been attempted in the past(e.g., Ronicke et al., 2004). Despite the Brassicaceae familycomprising of a wide array of different species, to date, it appearsthat only two wild crucifers, Capsella bursa-pastoris (Chen et al.,2007) and Erucastrum gallicum (Lefol et al., 1997a; Seguin-Swartzand Lefol, 1999), have been previously reported to show high levelsof resistance against Sclerotinia disease. Although introgressivehybrids were successfully obtained between different Brassica (B.
rapa and B. napus) species and Capsella bursa-pastoris (Chen et al.,2007), it remains to be confirmed if the introgression of resistanceagainst S. sclerotiorum from E. gallicum into cultivated species hasin fact been accomplished (Lefol et al., 1997a,b; Seguin-Swartz andLefol, 1999). There remains substantial potential both to identifywild crucifers with high levels of resistance to Sclerotinia diseaseand for its successful introgression to the cultivated species.
Three wild crucifers, viz. Erucastrum cardaminoides, Diplotaxis
tenuisiliqua and E. abyssinicum, have been identified with very highlevels of resistance against S. sclerotiorum (S.S. Banga, unpublisheddata). The aim of this study was to introgress the genomicsegments responsible for resistance against S. sclerotiorum fromthese three wild species into the cultivated germplasm. This paperreports the results of screening highly fertile (S4/S5) introgressionlines of B. juncea and B. napus derived from three different wildBrassica species, viz. E. cardaminoides, D. tenuisiliqua and E.
abyssinicum, against S. sclerotiorum.
2. Materials and methods
2.1. Plant materials
2.1.1. Experiment 1
Introgression lines, carrying alien genomic segments, weredeveloped for B. juncea and B. napus by introgression from threewild crucifers viz. E. cardaminoides (2n = 18), D. tenuisiliqua
(2n = 18) and E. abyssinicum (2n = 32). The general outlines ofbreeding schemes to introgress genomic segments from wildcrucifers into these cultivated species are detailed in Figs. 1–3.These lines were developed by S. S. Banga ([email protected]) and co-workers, Punjab Agricultural University from identi-fied wild crucifers by synthesizing intergeneric hybrids, E.
cardaminoides/B. rapa Chandra et al. (2004), E. cardaminoides/B.
nigra Chandra et al. (2004), B. napus/E. cardaminoides, B. juncea/D.
tenuisiliqua, and B. juncea/E. abyssinicum. Chromosome doublingwas achieved using colchicine in cross combinations involvingmonogenomic Brassica species to restore seed fertility. In crosses ofwild crucifers with digenomics, chromosome doubling was notrequired as these were partially fertile. The synthetic amphiploidsor the trigenomic hybrids were subsequently used as pollen/seedparents to hybridize with cultivated digenomics, B. juncea or B.
napus (Figs. 1–3). This was followed by three to four generations ofselfing using the single pod descent method. Special attempts were
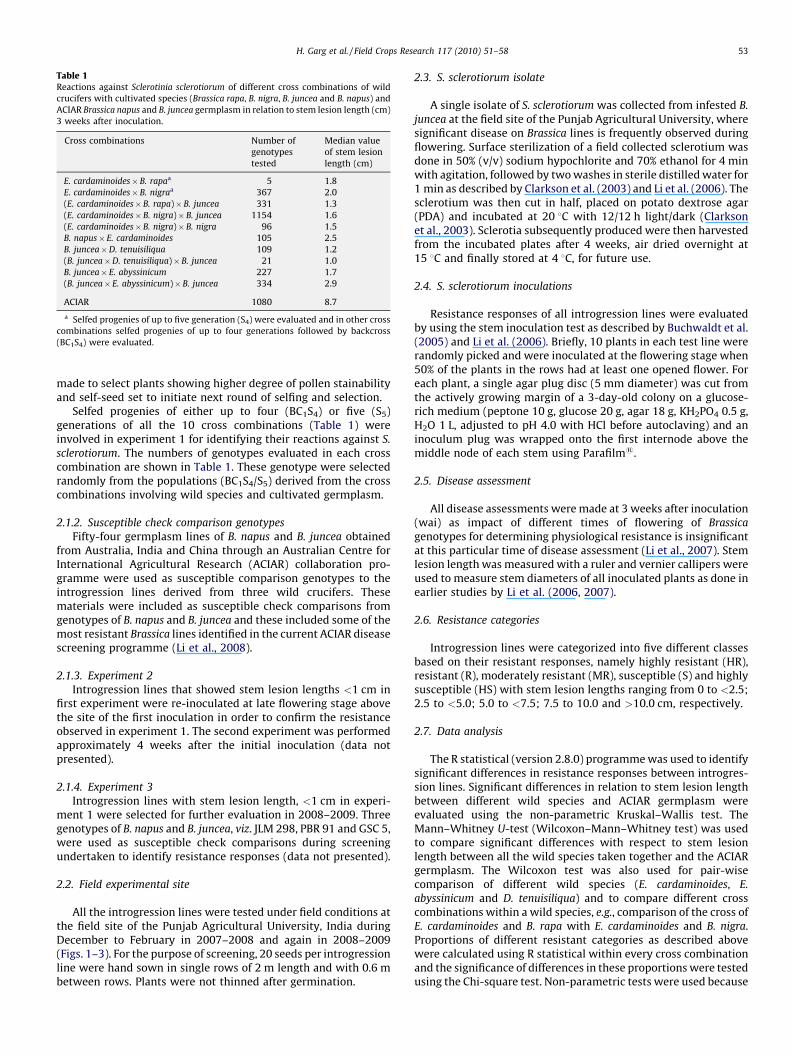
Table 1Reactions against Sclerotinia sclerotiorum of different cross combinations of wild
crucifers with cultivated species (Brassica rapa, B. nigra, B. juncea and B. napus) and
ACIAR Brassica napus and B. juncea germplasm in relation to stem lesion length (cm)
3 weeks after inoculation.
Cross combinations Number of
genotypes
tested
Median value
of stem lesion
length (cm)
E. cardaminoides�B. rapaa 5 1.8
E. cardaminoides�B. nigraa 367 2.0
(E. cardaminoides�B. rapa)�B. juncea 331 1.3
(E. cardaminoides�B. nigra)�B. juncea 1154 1.6
(E. cardaminoides�B. nigra)�B. nigra 96 1.5
B. napus�E. cardaminoides 105 2.5
B. juncea�D. tenuisiliqua 109 1.2
(B. juncea�D. tenuisiliqua)�B. juncea 21 1.0
B. juncea�E. abyssinicum 227 1.7
(B. juncea�E. abyssinicum)�B. juncea 334 2.9
ACIAR 1080 8.7
a Selfed progenies of up to five generation (S4) were evaluated and in other cross
combinations selfed progenies of up to four generations followed by backcross
(BC1S4) were evaluated.
H. Garg et al. / Field Crops Research 117 (2010) 51–58 53
made to select plants showing higher degree of pollen stainabilityand self-seed set to initiate next round of selfing and selection.
Selfed progenies of either up to four (BC1S4) or five (S5)generations of all the 10 cross combinations (Table 1) wereinvolved in experiment 1 for identifying their reactions against S.
sclerotiorum. The numbers of genotypes evaluated in each crosscombination are shown in Table 1. These genotype were selectedrandomly from the populations (BC1S4/S5) derived from the crosscombinations involving wild species and cultivated germplasm.
2.1.2. Susceptible check comparison genotypes
Fifty-four germplasm lines of B. napus and B. juncea obtainedfrom Australia, India and China through an Australian Centre forInternational Agricultural Research (ACIAR) collaboration pro-gramme were used as susceptible comparison genotypes to theintrogression lines derived from three wild crucifers. Thesematerials were included as susceptible check comparisons fromgenotypes of B. napus and B. juncea and these included some of themost resistant Brassica lines identified in the current ACIAR diseasescreening programme (Li et al., 2008).
2.1.3. Experiment 2
Introgression lines that showed stem lesion lengths <1 cm infirst experiment were re-inoculated at late flowering stage abovethe site of the first inoculation in order to confirm the resistanceobserved in experiment 1. The second experiment was performedapproximately 4 weeks after the initial inoculation (data notpresented).
2.1.4. Experiment 3
Introgression lines with stem lesion length, <1 cm in experi-ment 1 were selected for further evaluation in 2008–2009. Threegenotypes of B. napus and B. juncea, viz. JLM 298, PBR 91 and GSC 5,were used as susceptible check comparisons during screeningundertaken to identify resistance responses (data not presented).
2.2. Field experimental site
All the introgression lines were tested under field conditions atthe field site of the Punjab Agricultural University, India duringDecember to February in 2007–2008 and again in 2008–2009(Figs. 1–3). For the purpose of screening, 20 seeds per introgressionline were hand sown in single rows of 2 m length and with 0.6 mbetween rows. Plants were not thinned after germination.
2.3. S. sclerotiorum isolate
A single isolate of S. sclerotiorum was collected from infested B.
juncea at the field site of the Punjab Agricultural University, wheresignificant disease on Brassica lines is frequently observed duringflowering. Surface sterilization of a field collected sclerotium wasdone in 50% (v/v) sodium hypochlorite and 70% ethanol for 4 minwith agitation, followed by two washes in sterile distilled water for1 min as described by Clarkson et al. (2003) and Li et al. (2006). Thesclerotium was then cut in half, placed on potato dextrose agar(PDA) and incubated at 20 8C with 12/12 h light/dark (Clarksonet al., 2003). Sclerotia subsequently produced were then harvestedfrom the incubated plates after 4 weeks, air dried overnight at15 8C and finally stored at 4 8C, for future use.
2.4. S. sclerotiorum inoculations
Resistance responses of all introgression lines were evaluatedby using the stem inoculation test as described by Buchwaldt et al.(2005) and Li et al. (2006). Briefly, 10 plants in each test line wererandomly picked and were inoculated at the flowering stage when50% of the plants in the rows had at least one opened flower. Foreach plant, a single agar plug disc (5 mm diameter) was cut fromthe actively growing margin of a 3-day-old colony on a glucose-rich medium (peptone 10 g, glucose 20 g, agar 18 g, KH2PO4 0.5 g,H2O 1 L, adjusted to pH 4.0 with HCl before autoclaving) and aninoculum plug was wrapped onto the first internode above themiddle node of each stem using Parafilm1.
2.5. Disease assessment
All disease assessments were made at 3 weeks after inoculation(wai) as impact of different times of flowering of Brassica
genotypes for determining physiological resistance is insignificantat this particular time of disease assessment (Li et al., 2007). Stemlesion length was measured with a ruler and vernier callipers wereused to measure stem diameters of all inoculated plants as done inearlier studies by Li et al. (2006, 2007).
2.6. Resistance categories
Introgression lines were categorized into five different classesbased on their resistant responses, namely highly resistant (HR),resistant (R), moderately resistant (MR), susceptible (S) and highlysusceptible (HS) with stem lesion lengths ranging from 0 to <2.5;2.5 to <5.0; 5.0 to <7.5; 7.5 to 10.0 and >10.0 cm, respectively.
2.7. Data analysis
The R statistical (version 2.8.0) programme was used to identifysignificant differences in resistance responses between introgres-sion lines. Significant differences in relation to stem lesion lengthbetween different wild species and ACIAR germplasm wereevaluated using the non-parametric Kruskal–Wallis test. TheMann–Whitney U-test (Wilcoxon–Mann–Whitney test) was usedto compare significant differences with respect to stem lesionlength between all the wild species taken together and the ACIARgermplasm. The Wilcoxon test was also used for pair-wisecomparison of different wild species (E. cardaminoides, E.
abyssinicum and D. tenuisiliqua) and to compare different crosscombinations within a wild species, e.g., comparison of the cross ofE. cardaminoides and B. rapa with E. cardaminoides and B. nigra.Proportions of different resistant categories as described abovewere calculated using R statistical within every cross combinationand the significance of differences in these proportions were testedusing the Chi-square test. Non-parametric tests were used because

Fig. 4. An example of reactions of introgression lines against Sclerotinia sclerotiorum developed from wild crucifers Erucastrum cardaminoides, Diplotaxis tenuisiliqua and
Erucastrum abyssinicum in relation to stem lesion length assessed 3 weeks after inoculation.
Fig. 5. Box plot representation of resistance responses against Sclerotinia
sclerotiorum of ACIAR Brassica napus and B. juncea germplasm and introgression
lines derived from wild crucifers Erucastrum cardaminoides, Diplotaxis tenuisiliqua
and Erucastrum abyssinicum in relation to stem lesion length assessed 3 weeks after
inoculation. The central bar represents the median value and each box corresponds
to the range between 25th and 75th percentiles. Whiskers from the boxes include
the lowest and highest values. A, B and C represent introgression lines developed
from E. cardaminoides, D. tenuisiliqua and E. abyssinicum, respectively.
H. Garg et al. / Field Crops Research 117 (2010) 51–5854
the data was clearly not normally distributed and standardtransformations failed to normalise it.
3. Results
3.1. Resistance responses of introgression lines derived from three
wild species (E. cardaminoides, D. tenuisiliqua and E. abyssinicum)
and of ACIAR germplasm
3.1.1. Experiment 1
The Kruskal–Wallis test indicated significant differences(P < 0.001) between introgression lines derived from three wildspecies and the ACIAR germplasm in relation to the stem lesionlength 3 wai. Introgression lines derived from D. tenuisiliqua werehighly resistant with a median value for stem lesion length of1.2 cm as compared with wild species E. cardaminoides and E.
abyssinicum (median values = 1.7 and 2.0 cm, respectively; Figs. 4–6). The range of stem lesion lengths for introgression lines derivedfrom three wild species viz. E. cardaminoides, D. tenuisiliqua and E.
abyssinicum varied from 0 to 20, 0.2 to 12.0 and 0 to 25 cm,respectively (Figs. 5 and 6). The median value for stem lesionlength of ACIAR germplasm was very high (i.e., 8.7 cm as comparedto introgression lines) with stem lesion lengths ranging up to47.8 cm, which also confirms the high level of pathogenicity of theisolate of S. sclerotiorum utilized for this study.
Significant differences were also observed (P < 0.001) betweendifferent crosses involving the wild species with the ACIARgermplasm. The most resistant cross was (B. juncea � D.
tenuisiliqua) � B. juncea with a median value for stem lesionlength of 1.0 cm (Fig. 7). The maximum median value for stemlesion length in the (B. juncea � E. abyssinicum) � B. juncea crosscombination was 2.9 cm, which is much smaller than in the ACIARgermplasm which had a median value of 8.7 cm (Table 1).
3.1.2. Experiment 2
All introgression lines with a stem lesion length of <1.0 cmwere re-inoculated at the late flowering stage above the site ofinoculation for the first experiment. Overall, genotypes showedreasonably consistent results between experiments 1 and 2(significant linear relationship; P < 0.001), confirming the physio-logical resistance identified in experiment 1.
3.1.3. Experiment 3
As determined by the Kruskal–Wallis test, there were significantoverall differences (P < 0.001) between the introgression linesderived from three different wild crucifers and with the susceptiblecheck comparison genotypes, in relation to stem lesion lengths3 wai. The mean stem lesion length of genotypes used as susceptiblecheck comparisons in experiment 3 was 4.0 cm with a range varyingfrom 0 to 15.5 cm. However, mean values for stem lesion length forintrogression lines developed from the wild crucifers E. cardami-
noides, D. tenuisiliqua and E. abyssinicum were much lower, viz. 0.42,0.36 and 0.36 cm, with ranges for stem lesion length of 0–1.5, 0–1.2and 0–1.2 cm, respectively. The highly reduced stem lesion lengthsrecorded on the introgression lines were in accordance with what
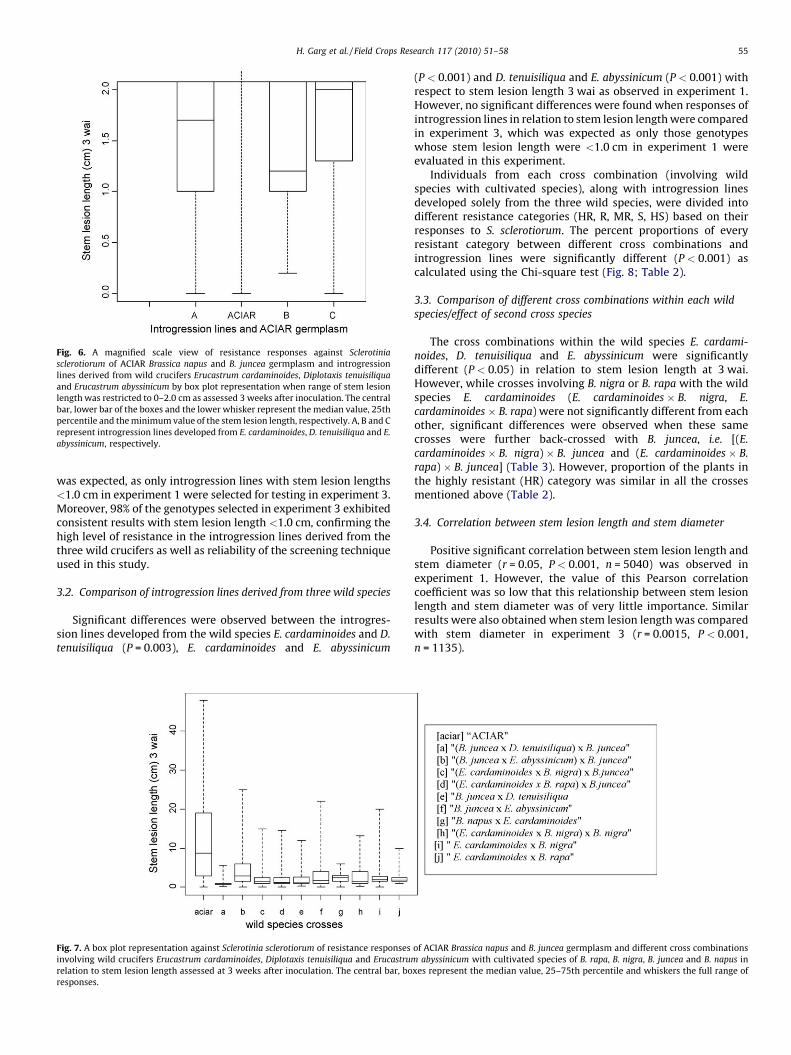
Fig. 6. A magnified scale view of resistance responses against Sclerotinia
sclerotiorum of ACIAR Brassica napus and B. juncea germplasm and introgression
lines derived from wild crucifers Erucastrum cardaminoides, Diplotaxis tenuisiliqua
and Erucastrum abyssinicum by box plot representation when range of stem lesion
length was restricted to 0–2.0 cm as assessed 3 weeks after inoculation. The central
bar, lower bar of the boxes and the lower whisker represent the median value, 25th
percentile and the minimum value of the stem lesion length, respectively. A, B and C
represent introgression lines developed from E. cardaminoides, D. tenuisiliqua and E.
abyssinicum, respectively.
H. Garg et al. / Field Crops Research 117 (2010) 51–58 55
was expected, as only introgression lines with stem lesion lengths<1.0 cm in experiment 1 were selected for testing in experiment 3.Moreover, 98% of the genotypes selected in experiment 3 exhibitedconsistent results with stem lesion length <1.0 cm, confirming thehigh level of resistance in the introgression lines derived from thethree wild crucifers as well as reliability of the screening techniqueused in this study.
3.2. Comparison of introgression lines derived from three wild species
Significant differences were observed between the introgres-sion lines developed from the wild species E. cardaminoides and D.
tenuisiliqua (P = 0.003), E. cardaminoides and E. abyssinicum
Fig. 7. A box plot representation against Sclerotinia sclerotiorum of resistance responses
involving wild crucifers Erucastrum cardaminoides, Diplotaxis tenuisiliqua and Erucastrum
relation to stem lesion length assessed at 3 weeks after inoculation. The central bar, bo
responses.
(P < 0.001) and D. tenuisiliqua and E. abyssinicum (P < 0.001) withrespect to stem lesion length 3 wai as observed in experiment 1.However, no significant differences were found when responses ofintrogression lines in relation to stem lesion length were comparedin experiment 3, which was expected as only those genotypeswhose stem lesion length were <1.0 cm in experiment 1 wereevaluated in this experiment.
Individuals from each cross combination (involving wildspecies with cultivated species), along with introgression linesdeveloped solely from the three wild species, were divided intodifferent resistance categories (HR, R, MR, S, HS) based on theirresponses to S. sclerotiorum. The percent proportions of everyresistant category between different cross combinations andintrogression lines were significantly different (P < 0.001) ascalculated using the Chi-square test (Fig. 8; Table 2).
3.3. Comparison of different cross combinations within each wild
species/effect of second cross species
The cross combinations within the wild species E. cardami-
noides, D. tenuisiliqua and E. abyssinicum were significantlydifferent (P < 0.05) in relation to stem lesion length at 3 wai.However, while crosses involving B. nigra or B. rapa with the wildspecies E. cardaminoides (E. cardaminoides � B. nigra, E.
cardaminoides � B. rapa) were not significantly different from eachother, significant differences were observed when these samecrosses were further back-crossed with B. juncea, i.e. [(E.
cardaminoides � B. nigra) � B. juncea and (E. cardaminoides � B.
rapa) � B. juncea] (Table 3). However, proportion of the plants inthe highly resistant (HR) category was similar in all the crossesmentioned above (Table 2).
3.4. Correlation between stem lesion length and stem diameter
Positive significant correlation between stem lesion length andstem diameter (r = 0.05, P < 0.001, n = 5040) was observed inexperiment 1. However, the value of this Pearson correlationcoefficient was so low that this relationship between stem lesionlength and stem diameter was of very little importance. Similarresults were also obtained when stem lesion length was comparedwith stem diameter in experiment 3 (r = 0.0015, P < 0.001,n = 1135).
of ACIAR Brassica napus and B. juncea germplasm and different cross combinations
abyssinicum with cultivated species of B. rapa, B. nigra, B. juncea and B. napus in
xes represent the median value, 25–75th percentile and whiskers the full range of
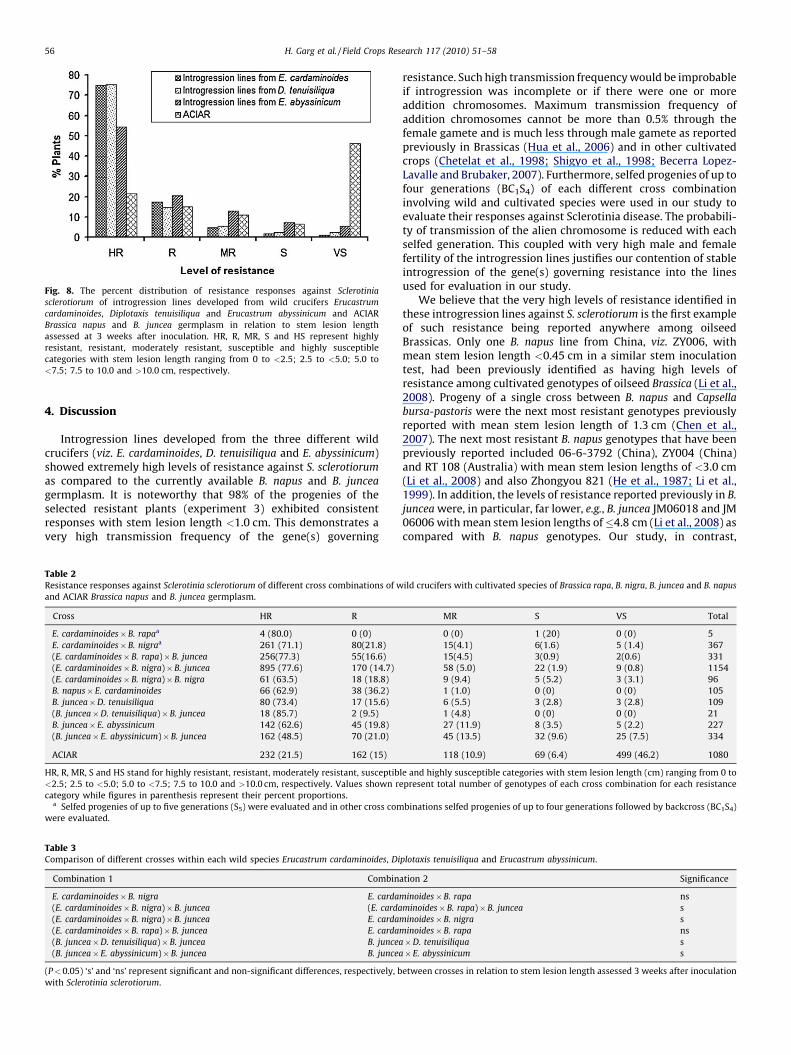
Fig. 8. The percent distribution of resistance responses against Sclerotinia
sclerotiorum of introgression lines developed from wild crucifers Erucastrum
cardaminoides, Diplotaxis tenuisiliqua and Erucastrum abyssinicum and ACIAR
Brassica napus and B. juncea germplasm in relation to stem lesion length
assessed at 3 weeks after inoculation. HR, R, MR, S and HS represent highly
resistant, resistant, moderately resistant, susceptible and highly susceptible
categories with stem lesion length ranging from 0 to <2.5; 2.5 to <5.0; 5.0 to
<7.5; 7.5 to 10.0 and >10.0 cm, respectively.
H. Garg et al. / Field Crops Research 117 (2010) 51–5856
4. Discussion
Introgression lines developed from the three different wildcrucifers (viz. E. cardaminoides, D. tenuisiliqua and E. abyssinicum)showed extremely high levels of resistance against S. sclerotiorum
as compared to the currently available B. napus and B. juncea
germplasm. It is noteworthy that 98% of the progenies of theselected resistant plants (experiment 3) exhibited consistentresponses with stem lesion length <1.0 cm. This demonstrates avery high transmission frequency of the gene(s) governing
Table 2Resistance responses against Sclerotinia sclerotiorum of different cross combinations of w
and ACIAR Brassica napus and B. juncea germplasm.
Cross HR R
E. cardaminoides�B. rapaa 4 (80.0) 0 (0)
E. cardaminoides�B. nigraa 261 (71.1) 80(21.8)
(E. cardaminoides�B. rapa)�B. juncea 256(77.3) 55(16.6)
(E. cardaminoides�B. nigra)�B. juncea 895 (77.6) 170 (14.7)
(E. cardaminoides�B. nigra)�B. nigra 61 (63.5) 18 (18.8)
B. napus�E. cardaminoides 66 (62.9) 38 (36.2)
B. juncea�D. tenuisiliqua 80 (73.4) 17 (15.6)
(B. juncea�D. tenuisiliqua)�B. juncea 18 (85.7) 2 (9.5)
B. juncea�E. abyssinicum 142 (62.6) 45 (19.8)
(B. juncea�E. abyssinicum)�B. juncea 162 (48.5) 70 (21.0)
ACIAR 232 (21.5) 162 (15)
HR, R, MR, S and HS stand for highly resistant, resistant, moderately resistant, susceptib
<2.5; 2.5 to <5.0; 5.0 to <7.5; 7.5 to 10.0 and >10.0 cm, respectively. Values shown re
category while figures in parenthesis represent their percent proportions.a Selfed progenies of up to five generations (S5) were evaluated and in other cross com
were evaluated.
Table 3Comparison of different crosses within each wild species Erucastrum cardaminoides, Di
Combination 1 Combin
E. cardaminoides�B. nigra E. carda
(E. cardaminoides�B. nigra)�B. juncea (E. carda
(E. cardaminoides�B. nigra)�B. juncea E. carda
(E. cardaminoides�B. rapa)�B. juncea E. carda
(B. juncea�D. tenuisiliqua)�B. juncea B. juncea
(B. juncea�E. abyssinicum)�B. juncea B. juncea
(P<0.05) ‘s’ and ‘ns’ represent significant and non-significant differences, respectively, b
with Sclerotinia sclerotiorum.
resistance. Such high transmission frequency would be improbableif introgression was incomplete or if there were one or moreaddition chromosomes. Maximum transmission frequency ofaddition chromosomes cannot be more than 0.5% through thefemale gamete and is much less through male gamete as reportedpreviously in Brassicas (Hua et al., 2006) and in other cultivatedcrops (Chetelat et al., 1998; Shigyo et al., 1998; Becerra Lopez-Lavalle and Brubaker, 2007). Furthermore, selfed progenies of up tofour generations (BC1S4) of each different cross combinationinvolving wild and cultivated species were used in our study toevaluate their responses against Sclerotinia disease. The probabili-ty of transmission of the alien chromosome is reduced with eachselfed generation. This coupled with very high male and femalefertility of the introgression lines justifies our contention of stableintrogression of the gene(s) governing resistance into the linesused for evaluation in our study.
We believe that the very high levels of resistance identified inthese introgression lines against S. sclerotiorum is the first exampleof such resistance being reported anywhere among oilseedBrassicas. Only one B. napus line from China, viz. ZY006, withmean stem lesion length <0.45 cm in a similar stem inoculationtest, had been previously identified as having high levels ofresistance among cultivated genotypes of oilseed Brassica (Li et al.,2008). Progeny of a single cross between B. napus and Capsella
bursa-pastoris were the next most resistant genotypes previouslyreported with mean stem lesion length of 1.3 cm (Chen et al.,2007). The next most resistant B. napus genotypes that have beenpreviously reported included 06-6-3792 (China), ZY004 (China)and RT 108 (Australia) with mean stem lesion lengths of <3.0 cm(Li et al., 2008) and also Zhongyou 821 (He et al., 1987; Li et al.,1999). In addition, the levels of resistance reported previously in B.
juncea were, in particular, far lower, e.g., B. juncea JM06018 and JM06006 with mean stem lesion lengths of�4.8 cm (Li et al., 2008) ascompared with B. napus genotypes. Our study, in contrast,
ild crucifers with cultivated species of Brassica rapa, B. nigra, B. juncea and B. napus
MR S VS Total
0 (0) 1 (20) 0 (0) 5
15(4.1) 6(1.6) 5 (1.4) 367
15(4.5) 3(0.9) 2(0.6) 331
58 (5.0) 22 (1.9) 9 (0.8) 1154
9 (9.4) 5 (5.2) 3 (3.1) 96
1 (1.0) 0 (0) 0 (0) 105
6 (5.5) 3 (2.8) 3 (2.8) 109
1 (4.8) 0 (0) 0 (0) 21
27 (11.9) 8 (3.5) 5 (2.2) 227
45 (13.5) 32 (9.6) 25 (7.5) 334
118 (10.9) 69 (6.4) 499 (46.2) 1080
le and highly susceptible categories with stem lesion length (cm) ranging from 0 to
present total number of genotypes of each cross combination for each resistance
binations selfed progenies of up to four generations followed by backcross (BC1S4)
plotaxis tenuisiliqua and Erucastrum abyssinicum.
ation 2 Significance
minoides�B. rapa ns
minoides�B. rapa)�B. juncea s
minoides�B. nigra s
minoides�B. rapa ns
�D. tenuisiliqua s
�E. abyssinicum s
etween crosses in relation to stem lesion length assessed 3 weeks after inoculation

H. Garg et al. / Field Crops Research 117 (2010) 51–58 57
identified 60% and 3.8% of the genotypes with mean stem lesionlengths <3.0 and <0.45 cm, respectively, among introgressionlines involving the cross combinations of three wild crucifers withB. juncea and B. napus, signifying the outstanding levels ofresistance present in these genotypes.
Median values for stem lesion length for introgression linesderived from D. tenuisiliqua, E. cardaminoides and E. abyssinicum
were 1.2, 1.7 and 2.0 cm, respectively, as compared to 8.7 cm forthe ACIAR genotypes. Clearly, all three wild crucifers used in thisstudy had extremely high levels of resistance against S. scler-
otiorum. Even though the wild crucifer E. gallicum had beenpreviously reported to have better resistance to Sclerotinia diseaseas compared to cultivated lines of Brassica species (Lefol et al.,1997a; Seguin-Swartz and Lefol, 1999), it was only evaluated usinga cut leaf bioassay with ascospores as the inoculation source. Theefficacy of cut leaf bioassays or detached leaf methods remainsdebatable, as this method correlates poorly with the fieldperformance of the Brassica genotypes (Bradley et al., 2006; C-X.Li and M.J. Barbetti, unpublished). However, our study utilized afield stem inoculation technique across two consecutive years, amethodology shown to be reliable and repeatable in differentiatingoilseed B. napus genotypes responses to Sclerotinia disease (Liet al., 2004, 2006; Buchwaldt et al., 2005). Consistent resistantreactions were observed in introgression lines across experiments1 and 2 and also across experiments 1 and 3, confirming the veryhigh level of physiological resistance identified in the developedintrogression lines.
Previous studies have reported that some of the morphologicaltraits such as stem diameter and growth stage of canola can affectthe reaction of Brassica genotypes to Sclerotinia disease (Zhao et al.,2004; Zhao and Wang, 2004; Li et al., 2006). While our studiesshowed some relationship between stem lesion length and stemdiameter in both experiments 1 and 3, this relationship was weakand not considered meaningful. This finding, taken in conjunctionwith the consistent results between experiments 1 and 2, whichwere performed at two different stages of flowering, is indicativethat high levels of resistance identified in these introgression lineswere not related to these morphological traits.
The novel sources of resistance identified in our study arehighly valuable genetic resources that can be utilized in oilseedBrassica breeding programmes to increase resistance in newcultivars against Sclerotinia disease. The present study constitutesthe first report of successful introgression of very high levels ofresistance against S. sclerotiorum from three wild crucifers, namelyE. cardaminoides, D. tenuisiliqua and E. abyssinicum, into cultivatedBrassicas. Future work will focus on the identification and mappingof the genes governing resistance against S. sclerotiorum.
Acknowledgments
Ms Harsh Garg gratefully acknowledges the financial assistanceof the Australian Centre for International Agricultural Research,Canberra, Australia, by way of a John Allwright PhD fellowship. IPRof the introgression lines used in the studies vests exclusively withPunjab Agricultural University, Ludhiana, India.
References
Abawi, G.S., Provvidenti, R., Crosier, D.C., Hunter, J.E., 1978. Inheritance of resistanceto white mold disease in Phaseolus coccineus. J. Hered. 69, 200–202.
Bardin, S.D., Huang, H.C., 2001. Research on biology and control of Sclerotiniadisease in Canada. Can. J. Plant Pathol. 23, 88–98.
Baswana, K.S., Rastogi, K.B., Sharma, P.P., 1991. Inheritance of stalk rot resistance incauliflower (Brassica oleracea var. Botrytis L.). Euphytica 57, 93–96.
Becerra Lopez-Lavalle, L.A., Brubaker, C.L., 2007. Frequency and fidelity of alienchromosome transmission in Gossypium hexaploid bridging populations. Ge-nome 50, 479–491.
Boland, G.J., Hall, R., 1994. Index of plant hosts of Sclerotinia sclerotiorum. Can. J.Plant Pathol. 16, 93–108.
Bolton, M.D., Thomma, B.P.H.J., Nelson, B.D., 2006. Sclerotinia sclerotiorum (Lib.) deBary: biology and molecular traits of a cosmopolitan pathogen. Mol. PlantPathol. 7, 1–16.
Bradley, C.A., Henson, R.A., Porter, P.M., LeGare, D.G., del Rıo, L.E., Khot, S.D., 2006.Response of canola cultivars to Sclerotinia sclerotiorum in controlled and fieldenvironments. Plant Dis. 90, 215–219.
Buchwaldt, L., Li, R., Hegedus, D.D., Rimmer, S., 2005. Pathogenesis of Sclerotiniasclerotiorum in relation to screening for resistance. In: Proceedings of the 13thInternational Sclerotinia Workshop, Monterey, CA, USA, p. 22.
Chandra, A., Gupta, M.L., Ahuja, I., Kaur, G., Banga, S.S., 2004. Intergeneric hybrid-ization between Erucastrum cardaminoides and two diploid crop Brassica spe-cies. Theor. Appl. Genet. 108, 1620–1626.
Chen, H.-F., Wang, H., Li, Z.-Y., 2007. Production and genetic analysis of partialhybrids in intertribal crosses between Brassica species (B. rapa, B. napus) andCapsella bursa-pastoris. Plant Cell Rep. 26, 1791–1800.
Chetelat, R.T., Rick, C.M., Cisneros, P., Alpert, K.B., DeVerna, J.W., 1998. Identification,transmission, and cytological behavior of Solanum lycopersicoides Dun. mono-somic alien addition lines in tomato (Lycopersicon esculentum Mill.). Genome 41,40–50.
Clarkson, J.P., Staveley, J., Phelps, K., Young, C.S., Whipps, J.M., 2003. Ascosporerelease and survival in Sclerotinia sclerotiorum. Mycol. Res. 107, 213–222.
Cruickshank, A.W., Cooper, M., Ryley, M.J., 2002. Peanut resistance to Sclerotiniaminor and S. sclerotiorum. Aust. J. Agric. Res. 53, 1105–1110.
Gilmore, B., Myers, J.R., Kean, D., 2002. Completion of testing of Phaseolus coccineusplant introductions (PIs) for white mold, Sclerotinia sclerotiorum, resistance.Ann. Rept. Bean Improv. Coop. 45 .
Godoy, M., Castano, F., Re, J., Rodriguez, R., 2005. Sclerotinia resistance in sunflower:genotypic variations of hybrids in three environment of Argentina. Euphytica145, 147–154.
Gulya, T., Block, C., Marek, L.F., 2009. Identifying sources of Sclerotinia stalk rotresistance in wild annual Helianthus species for improvement of cultivatedsunflower. In: Proceedings of the 14th International Sclerotinia Workshop,Wilmington, NC, USA.
Hartman, G.L., Gardner, M.E., Hymowitz, T., Naidoo, G.C., 2000. Evaluation ofperennial Glycine species for resistance to soybean fungal pathogens that causeSclerotinia stem rot and sudden death syndrome. Crop Sci. 40, 545–549.
He, Y.H., Yang, R.F., Luo, S.Q., 1987. Development and study of a new rapeseedvariety Zhongyou 821 with high yield and disease resistance (tolerance). OilCrops China 2, 11–15.
Hind, T.L., Ash, G.J., Murray, G.M., 2003. Prevalence of Sclerotinia stem rot of canolain New South Wales. Aust. J. Exp. Agric. 43, 163–168.
Hua, Y.W., Liu, M., Li, Z.Y., 2006. Parental genome separation and elimination of cellsand chromosomes revealed by GISH and AFLP analyses in a Brassica carina-ta � Orychophragmus violaceus cross. Ann. Bot. 97, 993–998.
Koch, S., Dunker, S., Kleinhenz, B., Rohrig, M., von Tiedemann, A., 2007. A crop loss-related forecasting model for Sclerotinia stem rot in winter oilseed rape.Phytopathology 97, 1186–1194.
Lefol, C., Seguin-Swartz, G., Morrall, R.A.A., 1997a. Resistance to Sclerotinia scler-otiorum in a weed related to canola. Can. J. Plant Pathol. 19, 113 (Abstract).
Lefol, E., Seguin-Swartz, G., Downey, R.K., 1997b. Sexual hybridisation in crosses ofcultivated Brassica species with the crucifers Erucastrum gallicum and Raphanusraphanistrum: potential for gene introgression. Euphytica 95, 127–139.
Li, C.X., Li, H., Siddique, A.B., Sivasithamparam, K., Salisbury, P., Banga, S.S., Banga, S.,Chattopadhyay, C., Kumar, A., Singh, R., Singh, D., Agnihotri, A., Liu, S.Y., Li, Y.C.,Tu, J., Fu, T.D., Wang, Y.F., Barbetti, M.J., 2007. The importance of the type andtime of inoculation and assessment in the determination of resistance in Brassianapus and B. juncea to Sclerotinia sclerotiorum. Aust. J. Agric. Res. 58, 1198–1203.
Li, C.X., Li, H., Sivasithamparam, K., Fu, T.D., Li, Y.C., Liu, S.Y., Barbetti, M.J., 2006.Expression of field resistance under Western Australian conditions to Sclerotiniasclerotiorum in Chinese and Australian Brassica napus and Brassica junceagermplasm and its relation with stem diameter. Aust. J. Agric. Res. 57, 1131–1135.
Li, C.X., Liu, S.Y., Sivasithamparam, K., Barbetti, M.J., 2008. New sources of resistanceto Sclerotinia stem rot caused by Sclerotinia sclerotiorum in Chinese and Aus-tralian Brassica napus and Brassica juncea germplasm screened under WesternAustralian conditions. Australas. Plant Pathol. 38, 149–152.
Li, R., Rimmer, R., Buchwaldt, L., Sharpe, A.G., Seguin-Swartz, G., Coutu, C., Hegedus,D.D., 2004. Interaction of Sclerotinia sclerotiorum with a resistant Brassica napuscultivar: expressed sequence tag analysis identifies genes associated withfungal pathogenesis. Fungal Genet. Biol. 41, 735–753.
Li, Y., Chen, J., Bennett, R., Kiddle, G., Wallsgrove, R., Huang, Y., He, Y., 1999. Breeding,inheritance, and biochemical studies on Brassica napus cv. Zhongyou 821:tolerance to Sclerotinia sclerotiorum (stem rot). In: Wratten, N., Salisbury, P.A.(Eds.), Proceedings of the 10th International Rapeseed Congress. Canberra,Australia, September 26-29, 1999, Available at:, p. 61.
Malvarez, M., Carbone, I., Grunwald, N.J., Subbarao, K.V., Schafer, M., Kohn, L.M.,2007. New populations of Sclerotinia sclerotiorum from lettuce in California andpeas and lentils in Washington. Phytopathology 97, 470–483.
McCartney, H.A., Lacey, M.E., Li, Q., Heran, A., 1999. Airborne ascospore concentrationand the infection of oilseed rape and sunflowers by Sclerotinia sclerotiorum. In:Wratten, N., Salisbury, P.A. (Eds.),Proceedings of the 10th International RapeseedCongress. Canberra, Australia, September 26–29, 1999, Available at:, p. 132.
Porter, L.D., Hoheisel, G., Coffman, V.A., 2009. Resistance of peas to Sclerotiniasclerotiorum in the Pisum core collection. Plant Pathol. 58, 52–60.

H. Garg et al. / Field Crops Research 117 (2010) 51–5858
Ronicke, S., Hahn, V., Horn, R., Grone, I., Brahm, L., Schnabl, H., Friedt, W., 2004.Interspecific hybrids of sunflower as a source of Sclerotinia resistance. PlantBreed. 123, 152–157.
Schwartz, H.F., Otto, K., Teran, H., Lema, M., Singh, S.P., 2006. Inheritance of white moldresistance in Phaseolus vulgaris� P. coccineus crosses. Plant Dis. 90, 1167–1170.
Seguin-Swartz, G., Lefol, C., 1999. Sclerotinia stem rot resistance in crucifers. In:Wratten, N.,Salisbury, P.A. (Eds.),Proceedings of the 10th International RapeseedCongress. Canberra, Australia, September 26-29, 1999, Available at:, p. 153.
Seiler, G.J., 1992. Utilization of wild sunflower species for the improvement ofcultivated sunflower. Field Crops Res. 30, 195–230.
Shigyo, M., Iino, M., Ino, H., Tashiro, Y., 1998. Transmission rates of extra chromo-somes in alien monosomic addition lines of Japanese bunching onion with extrachromosomes from shallot. J. Jpn. Soc. Hortic. Sci. 68, 8–22.
Singh, R., Singh, D., Li, H., Sivasithamparam, S., Yadav, N.R., Salisbury, P., Barbetti,M.J., 2008. Management of Sclerotinia rot of oilseed Brassicas—a focus on India.Brassica 10, 1–27.
Singh, S.P., Teran, H., Schwartz, H.F., Otto, K., Lema, M., 2009. White mold-resistantinterspecific common bean germplasm lines VCW 54 and VCW 55. J. PlantRegist. 3, 191–197.
Sprague, S., Stewart-Wade, S., 2002. Sclerotinia in canola—results from petal anddisease surveys across Victoria in 2001. In: Grains Research and DevelopmentCorporation Research Update—Southern Region, Australia, Grains Research andDevelopment Corporation, Victoria, Australia.
Williams, J.R., Stelfox, D., 1980. Influence of farming practices in Alberta ongermination and apothecium production of sclerotia of Sclerotinia sclerotiorum.Can. J. Plant Pathol. 2, 169–172.
Zhao, Y., Wang, M.L., 2004. Inheritance and agronomic performance of an apetalousflower mutant in Brassica napus L. Euphytica 137, 381–386.
Zhao, J., Udall, J.A., Quijada, P.A., Grau, C.R., Meng, J., Osborn, T.C., 2006. Quantitativetrait loci for resistance to Sclerotinia sclerotiorum and its association with ahomeologous non-reciprocal transposition in Brassica napus L. Theor. Appl.Genet. 112, 509–516.
Zhao, J., Meng, J., 2003. Genetic analysis of loci associated with partial resistance toSclerotinia sclerotiorum in rapeseed (Brassica napus L.). Theor. Appl. Genet. 106,759–764.
Zhao, J., Peltier, A.J., Meng, J., Osborn, T.C., Grau, C.R., 2004. Evaluation of Sclerotiniastem rot resistance in oilseed Brassica napus using a petiole inoculation tech-nique under green house conditions. Plant Dis. 88, 1033–1039.
Related Documents











