ORIGINAL PAPER High diversity beetle assemblages attracted to carrion and dung in threatened tropical oak forests in Southern Mexico Ubaldo Caballero • Jorge L. Leo ´n-Corte ´s Received: 23 March 2011 / Accepted: 19 September 2011 / Published online: 30 September 2011 Ó Springer Science+Business Media B.V. 2011 Abstract Despite high diversity levels of beetles inhab- iting dung and carcasses, very few studies have attempted a comparative assessment of copro-necrophile beetle com- munities in relation to spatio-temporal variations, particu- larly in the tropics where the vast majority of beetles occur. We compared beetle assemblages attracted to pads of cattle dung and rat carcasses in four contrasting vegetation types associated with oak forest. In a total of 52 transects including 3,952 dung pad days and 2,600 carcass-trap days we recorded 14,989 beetles representing 406 species and 33 families. Necrophiles (323 species and 33 families) were considerably more diverse than coprophiles (172 species and 16 families). Staphylinidae were taxonomically and numerically dominant, comprising 45% of species and 66% of individuals, respectively. Species estimators (Chao 2) suggested that the observed beetle richness represented 68% of coprophile and 56% of necrophile species richness, with rare species constituting the majority of the species: singletons and doubletons = 89 species (52%) of copro- philes and 166 species (51%) of necrophiles. Beetle assemblages varied in diversity and composition as regards to vegetation type and season: samples from less disturbed vegetation types (continuous oak forest and ravines) had higher beetle diversity, and a strong seasonality effect was recorded for necrophiles, but not for coprophiles. In addi- tion, an indicator value analysis (IndVal), showed that relatively preserved vegetation types recorded more indi- cator species as compared to disturbed sites. Our studies clearly demonstrates that the least fragmented oak forest and ravine are the most valuable areas for necrophile and coprophile beetles in Neotropical Mexico, especially for specialist beetles. Keywords Carrion Chiapas Coleoptera Community ecology Dung Neotropical region Introduction Dung pads and carcasses represent discrete, ephemeral, patchy resources of highly concentrated energy which are widespread throughout a variety of habitats, and to which many insect species are adapted (Hanski 1983; Putman 1983; Doube 1987; Shorrocks 1990). By taking part in the decomposition processes, insects promote persistence of local plants and a variety of trophic interactions (including bacteria, fungi, protozoa, nematodes; Carter et al. 2007; Parmenter and MacMahon 2009). Insects, especially Coleoptera, are one of the most diverse components of dung and carrion substrates, with over 100 and 200 beetle species recorded on single cattle dung pads and carcasses, respectively (Payne 1965; Bernon 1981). An important implication of the spatio-temporal patch- iness of carrion and dung relates to the tight dependency of their insect faunas, rendering them highly sensitive to habitat loss and fragmentation (Klein 1989; Trumbo and Bloch 2000; Halffter and Arellano 2002). This is particu- larly true in tropical regions, where large areas of forest have been converted into complex landscape mosaics dominated by secondary forest fragments and agro-pastoral systems (Dirzo and Raven 2003; Lamb et al. 2005). Many other taxonomic groups have been negatively influenced by these effects (Fahring 2003). U. Caballero J. L. Leo ´n-Corte ´s (&) Departamento de Ecologı ´a y Sistema ´tica Terrestre, El Colegio de la Frontera Sur, Carr. Panamericana y Av. Perife ´rico Sur S/N, San Cristo ´bal de las Casas, Chiapas 29290, Mexico e-mail: [email protected] 123 J Insect Conserv (2012) 16:537–547 DOI 10.1007/s10841-011-9439-y

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
High diversity beetle assemblages attracted to carrion and dungin threatened tropical oak forests in Southern Mexico
Ubaldo Caballero • Jorge L. Leon-Cortes
Received: 23 March 2011 / Accepted: 19 September 2011 / Published online: 30 September 2011
� Springer Science+Business Media B.V. 2011
Abstract Despite high diversity levels of beetles inhab-
iting dung and carcasses, very few studies have attempted a
comparative assessment of copro-necrophile beetle com-
munities in relation to spatio-temporal variations, particu-
larly in the tropics where the vast majority of beetles occur.
We compared beetle assemblages attracted to pads of cattle
dung and rat carcasses in four contrasting vegetation types
associated with oak forest. In a total of 52 transects
including 3,952 dung pad days and 2,600 carcass-trap days
we recorded 14,989 beetles representing 406 species and
33 families. Necrophiles (323 species and 33 families)
were considerably more diverse than coprophiles (172
species and 16 families). Staphylinidae were taxonomically
and numerically dominant, comprising 45% of species and
66% of individuals, respectively. Species estimators (Chao
2) suggested that the observed beetle richness represented
68% of coprophile and 56% of necrophile species richness,
with rare species constituting the majority of the species:
singletons and doubletons = 89 species (52%) of copro-
philes and 166 species (51%) of necrophiles. Beetle
assemblages varied in diversity and composition as regards
to vegetation type and season: samples from less disturbed
vegetation types (continuous oak forest and ravines) had
higher beetle diversity, and a strong seasonality effect was
recorded for necrophiles, but not for coprophiles. In addi-
tion, an indicator value analysis (IndVal), showed that
relatively preserved vegetation types recorded more indi-
cator species as compared to disturbed sites. Our studies
clearly demonstrates that the least fragmented oak forest
and ravine are the most valuable areas for necrophile and
coprophile beetles in Neotropical Mexico, especially for
specialist beetles.
Keywords Carrion � Chiapas � Coleoptera � Community
ecology � Dung � Neotropical region
Introduction
Dung pads and carcasses represent discrete, ephemeral,
patchy resources of highly concentrated energy which are
widespread throughout a variety of habitats, and to which
many insect species are adapted (Hanski 1983; Putman
1983; Doube 1987; Shorrocks 1990). By taking part in the
decomposition processes, insects promote persistence of
local plants and a variety of trophic interactions (including
bacteria, fungi, protozoa, nematodes; Carter et al. 2007;
Parmenter and MacMahon 2009). Insects, especially
Coleoptera, are one of the most diverse components of
dung and carrion substrates, with over 100 and 200 beetle
species recorded on single cattle dung pads and carcasses,
respectively (Payne 1965; Bernon 1981).
An important implication of the spatio-temporal patch-
iness of carrion and dung relates to the tight dependency of
their insect faunas, rendering them highly sensitive to
habitat loss and fragmentation (Klein 1989; Trumbo and
Bloch 2000; Halffter and Arellano 2002). This is particu-
larly true in tropical regions, where large areas of forest
have been converted into complex landscape mosaics
dominated by secondary forest fragments and agro-pastoral
systems (Dirzo and Raven 2003; Lamb et al. 2005). Many
other taxonomic groups have been negatively influenced by
these effects (Fahring 2003).
U. Caballero � J. L. Leon-Cortes (&)
Departamento de Ecologıa y Sistematica Terrestre, El Colegio
de la Frontera Sur, Carr. Panamericana y Av. Periferico Sur S/N,
San Cristobal de las Casas, Chiapas 29290, Mexico
e-mail: [email protected]
123
J Insect Conserv (2012) 16:537–547
DOI 10.1007/s10841-011-9439-y

Previous surveys have assessed the impacts of distur-
bance and habitat fragmentation upon complex and diverse
copro-necrophile beetle communities (Davis 1994; Dunn
and Danoff-Burg 2007; Ulrich et al. 2008). Sometimes
studies narrowly focus on Scarabaeinae dung beetle
assessments (see Nichols et al. 2007), whereas only a
limited number of surveys have considered the assessment
of others key groups such as carrion beetles (Silphidae; e.
g. Gibbs and Stanton 2001; Trumbo and Bloch 2000; Wolf
and Gibbs 2004; Creighton et al. 2009) and rove beetles
(Staphylinidae; e.g. Pohl et al. 2007; Caballero et al. 2009;
Vasquez-Velez et al. 2010). It would therefore be extre-
mely important to quantify comparative responses of the
copro-necrophile beetle communities particularly in Neo-
tropical systems, where, to our knowledge, no attempt has
been made to do so, and where a complete understanding
on the effects of habitat fragmentation and on the relative
importance of habitat type and conditions on complex
guilds are needed for effective management and conser-
vation. Both carrion and dung have been extensively used
to estimate and monitor beetle local populations safely
(Payne 1965; Putman 1983), and hence this procedure
allowed us to maximize insect collections along the whole
decomposition process (Holter 1982; Putman 1983).
We conducted a comparative detailed study of copro-
necrophile beetles in four vegetation types associated with
fragmented oak forests in southern Mexico. Our particular
aims were (i) to evaluate the relative effects of vegetation
type and seasonality on diversity and composition of
these two beetle communities in oak forest remnants, and
(ii) based on species association and diversity, to identify
the relative importance of particular vegetation types for
species conservation. A set of important predictions of
our assessment relates to significant negative changes in
species composition and abundance for sensitive groups
(i.e. Hydrophilidae, Leiodidae, Nitidulidae, Silphidae)
moderate or no changes for relatively generalist guilds
(i.e. Carabidae, Histeridae), and positive changes for
common and widely distributed groups (i.e. Scarabaei-
deae, Staphylinidae), in relation to vegetation small-scale
variations.
Materials and methods
Study area
Four vegetation types associated with oak forests were
studied in southern Mexico. Three of which were located in
the San Fernando Valley (16�4804800N, 93�1004200W): (1) a
highly fragmented oak forest, referred to as ‘‘fragmented
oak forest’’; (2) an area dominated by savannah vegetation
with scattered quarries referred as ‘‘savannah’’; (3) an area
at the bottom of a canyon that has seasonal streams and
interspersed with oak forest patches, referred as ‘‘ravines’’.
Because continuous, little-disturbed oak forests are
practically nonexistent in the San Fernando Valley, we
chose a forest in the highlands, approximately 55 km to the
east, as point of comparison. This non-fragmented tropical
oak forest is located in the Biological Reserve Cerro Hu-
itepec (16�4403800N, 92�4001500W), referred to as ‘‘contin-
uous oak forest’’ (for further details see Caballero et al.
2009). Mean annual temperature and rainfall are 26�C and
883 mm for San Fernando and 15�C and 1,200 mm for
Huitepec (CNA 2006). San Fernando is highly seasonal,
with the wet season concentrated from May to October, and
the dry season from November to April; 81% of the pre-
cipitation falls between June and September. In Huitepec,
rainfall does fluctuate seasonally, but its dry season
encompasses a very short period of the year (from
December to March; Anonymous 2006).
Sampling design
In each vegetation type, at least four 50-m linear transects
were established (one transect corresponding to one sam-
ple, and the number of transects per vegetation type during
both seasons as replicates, N = 52, Table 2). Along each
transect, we exposed fresh baits (rat carcasses and dung
cattle pads) to quantify beetle species and abundance
through time. In order to register the fauna during con-
trasting conditions of the year, we carried out field work
distributed in two sampling periods during dry (March–
early June) and wet seasons (August–October, 2005),
except for ‘‘continuous oak forest’’ sites where only sam-
ples during the rainy season were taken (May–July, 2006).
Coprophilous beetle sampling
Along each transect, a 1 kg pad of fresh cattle dung was
placed every 5 m, making a total of 10 dung pads per
transect. Every day we removed one pad from each transect
for the first 7 days and then after 10, 15 and 23 days
(N = 520 dung pads). Beetles were extracted by putting
each pad in water (ca 20 l); then collecting the beetles as
they floated to the surface (Moore 1954). All insects were
placed in 70% ethanol. Beetles which burrowed beneath
the dung were hand-collected.
Necrophilous beetle sampling
Recent killed laboratory rats (Rattus norvegicus L.) were
used as baits. A fresh rat carcass (weight 80–100 g) was
placed in a pitfall trap as described by Kocarek (2000). The
trap was placed in a wire mesh cylinder with 1.5 cm holes
in order to avoid disturbance by vertebrate scavengers.
538 J Insect Conserv (2012) 16:537–547
123

A fixing liquid of ethylene glycol–water in a 1:1 proportion
was added to a vessel below the carrion, and the cylinder
was buried at the soil surface (Kocarek 2000).
Along each transect, one trap was located (Total N = 52
traps) and inspected for insects daily for the first 7 days,
every other day from day 9 to 20th, and twice a week for
the rest of the sampling period. The total time of bait
exposure was 3,952 dung-pad days and 2,600 carcass-trap
days.
Beetles were identified to the family level using Borror
et al. (1989) and Arnett et al. (2002). All specimens were
sorted to morphospecies using the guidelines of Oliver and
Beattie (1996). In problematic specimens, genitalia were
dissected and sent to specialists for species identification
(see Acknowledgments). A reference series of each mor-
phospecies was deposited in the entomological collection
of El Colegio de la Frontera Sur (ECOSC-E), San Cristobal
de las Casas, Mexico. For simplicity, we hereafter refer to
morphospecies as species.
Data analysis
We compared sample-based rarefaction curves (at compa-
rable levels of sampling effort) by scaling the x-axis to the
number of transects (obtaining species density curves;
Gotelli and Colwell 2001). This method is particularly useful
if assemblages have been assessed under different sampling
intensity or success (Gotelli and Colwell 2001), as in our
case. We contrasted values for: (a) the total number of spe-
cies recorded for coprophiles and necrophiles, (b) the total
number of species recorded for coprophiles and necrophiles
between dry and rainy seasons, (c) the total number of spe-
cies recorded for coprophiles and necrophiles among four
vegetation types, and (d) the number of species recorded in
each vegetation type for coprophiles and necrophiles
between dry and rainy seasons.
To compare the diversity as stated above in (c) and (d),
we computed Fisher’s alpha, a widely-used diversity index
that is relatively unbiased to sampling intensity (Magurran
2004). In addition, species richness for dominant
beetle families was compared using Mann–Whitney U or
Kruskal–Wallis tests among vegetation types and between
dry and rainy seasons. For all tests, statistical significance
was taken after Bonferroni correction for multiple com-
parisons (Sokal and Rohlf 1995).
To estimate total species richness numbers for vegeta-
tion types we used Chao2, a non-parametric statistical
estimator to reduce potential bias due to potential incom-
plete sampling (Chao 2005).
We also calculated the number of ‘‘rare’’ species defined
as singletons and doubletons (i.e. those which recorded:
only one individual and only two individuals, respectively,
see Novotny and Basset 2000).
We examined the similarity for coprophiles and necro-
philes among vegetation types by computing an abun-
dance-based Sørensen index of similarity developed by
Chao et al. (2005), since this approach reduces the potential
bias of undersampling when differences are in sample size
(Chao et al. 2005). In addition, as sampling effort within
vegetation types was equivalent, seasonal similarity within
vegetation types was examined using Sørensen index
(Magurran 2004). All calculations were performed using
EstimateS 8.2.0 (Colwell 2009).
We calculated a measure of dominance using the pro-
portion of individuals for the commonest species in a given
vegetation type (Berger-Parker index) (Magurran 2004).
Dominance values (for coprophiles and necrophiles) were
estimated for dry and rainy seasons and for each vegetation
type in each season. In addition, to elucidate dominance
patterns within local communities, we plotted species-
abundance distributions for coprophiles and necrophiles
(Whittaker plots) recorded in each vegetation type. By
regressing species rank and species abundance (Magurran
2004), we examined potential deviations of the slopes of
the linear equations as regards to vegetation type—the
lower the value of the slope of each regression line the
greater the evenness of the assemblage (Tokeshi 1993). All
statistical analyses were performed using SPSS 15�.
Species-vegetation associations were identified based on
the indicator value method (IndVal, see Dufrene and
Legendre 1997). IndVal analysis allowed us to identify
indicator species within each vegetation type. All species
that recorded at least 10 individuals in samples were con-
sidered for analysis.
Results
Beetle fauna
We collected a total of 14,989 beetles representing 406
species and 33 families, of which 5,255 individuals, 172
species and 16 families were recorded in dung pads, and
9,734 individuals, 323 species and 33 families in rat car-
casses (Table 1). Family richness among vegetation types
ranged from 7 to 9 families for coprophiles and from 14 to
21 for necrophiles (Table 2). Staphylinidae was the most
diverse family, followed by Scarabaeidae and Histeridae
(Table 1). Staphylinids were represented by 83 species of
coprophiles and 142 species of necrophiles, including 73
and 63% of the total abundance, respectively (Table 1).
Rare species constituted the majority of the species: sin-
gletons and doubletons = 89 species (52%) of coprophiles
and 166 species (51%) of necrophiles (Table 2). However,
for coprophiles, the singletons’ curve appeared to reach an
asymptotic behavior (Fig. 1a). The 20 commonest species
J Insect Conserv (2012) 16:537–547 539
123
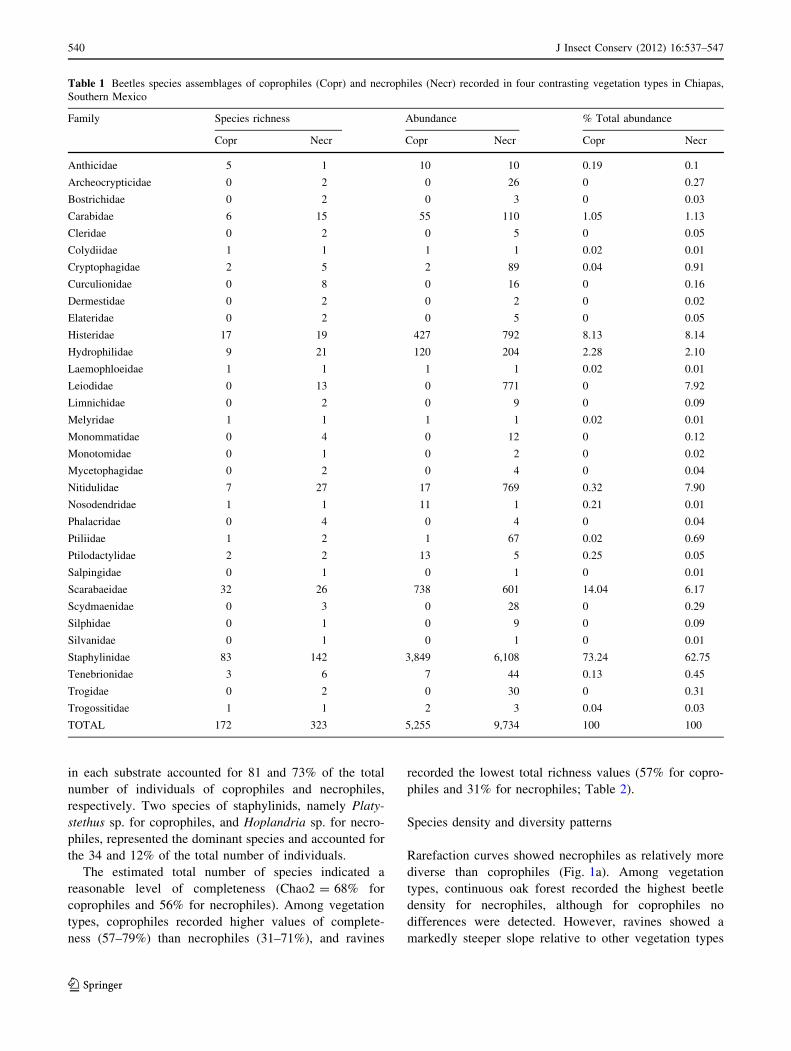
in each substrate accounted for 81 and 73% of the total
number of individuals of coprophiles and necrophiles,
respectively. Two species of staphylinids, namely Platy-
stethus sp. for coprophiles, and Hoplandria sp. for necro-
philes, represented the dominant species and accounted for
the 34 and 12% of the total number of individuals.
The estimated total number of species indicated a
reasonable level of completeness (Chao2 = 68% for
coprophiles and 56% for necrophiles). Among vegetation
types, coprophiles recorded higher values of complete-
ness (57–79%) than necrophiles (31–71%), and ravines
recorded the lowest total richness values (57% for copro-
philes and 31% for necrophiles; Table 2).
Species density and diversity patterns
Rarefaction curves showed necrophiles as relatively more
diverse than coprophiles (Fig. 1a). Among vegetation
types, continuous oak forest recorded the highest beetle
density for necrophiles, although for coprophiles no
differences were detected. However, ravines showed a
markedly steeper slope relative to other vegetation types
Table 1 Beetles species assemblages of coprophiles (Copr) and necrophiles (Necr) recorded in four contrasting vegetation types in Chiapas,
Southern Mexico
Family Species richness Abundance % Total abundance
Copr Necr Copr Necr Copr Necr
Anthicidae 5 1 10 10 0.19 0.1
Archeocrypticidae 0 2 0 26 0 0.27
Bostrichidae 0 2 0 3 0 0.03
Carabidae 6 15 55 110 1.05 1.13
Cleridae 0 2 0 5 0 0.05
Colydiidae 1 1 1 1 0.02 0.01
Cryptophagidae 2 5 2 89 0.04 0.91
Curculionidae 0 8 0 16 0 0.16
Dermestidae 0 2 0 2 0 0.02
Elateridae 0 2 0 5 0 0.05
Histeridae 17 19 427 792 8.13 8.14
Hydrophilidae 9 21 120 204 2.28 2.10
Laemophloeidae 1 1 1 1 0.02 0.01
Leiodidae 0 13 0 771 0 7.92
Limnichidae 0 2 0 9 0 0.09
Melyridae 1 1 1 1 0.02 0.01
Monommatidae 0 4 0 12 0 0.12
Monotomidae 0 1 0 2 0 0.02
Mycetophagidae 0 2 0 4 0 0.04
Nitidulidae 7 27 17 769 0.32 7.90
Nosodendridae 1 1 11 1 0.21 0.01
Phalacridae 0 4 0 4 0 0.04
Ptiliidae 1 2 1 67 0.02 0.69
Ptilodactylidae 2 2 13 5 0.25 0.05
Salpingidae 0 1 0 1 0 0.01
Scarabaeidae 32 26 738 601 14.04 6.17
Scydmaenidae 0 3 0 28 0 0.29
Silphidae 0 1 0 9 0 0.09
Silvanidae 0 1 0 1 0 0.01
Staphylinidae 83 142 3,849 6,108 73.24 62.75
Tenebrionidae 3 6 7 44 0.13 0.45
Trogidae 0 2 0 30 0 0.31
Trogossitidae 1 1 2 3 0.04 0.03
TOTAL 172 323 5,255 9,734 100 100
540 J Insect Conserv (2012) 16:537–547
123
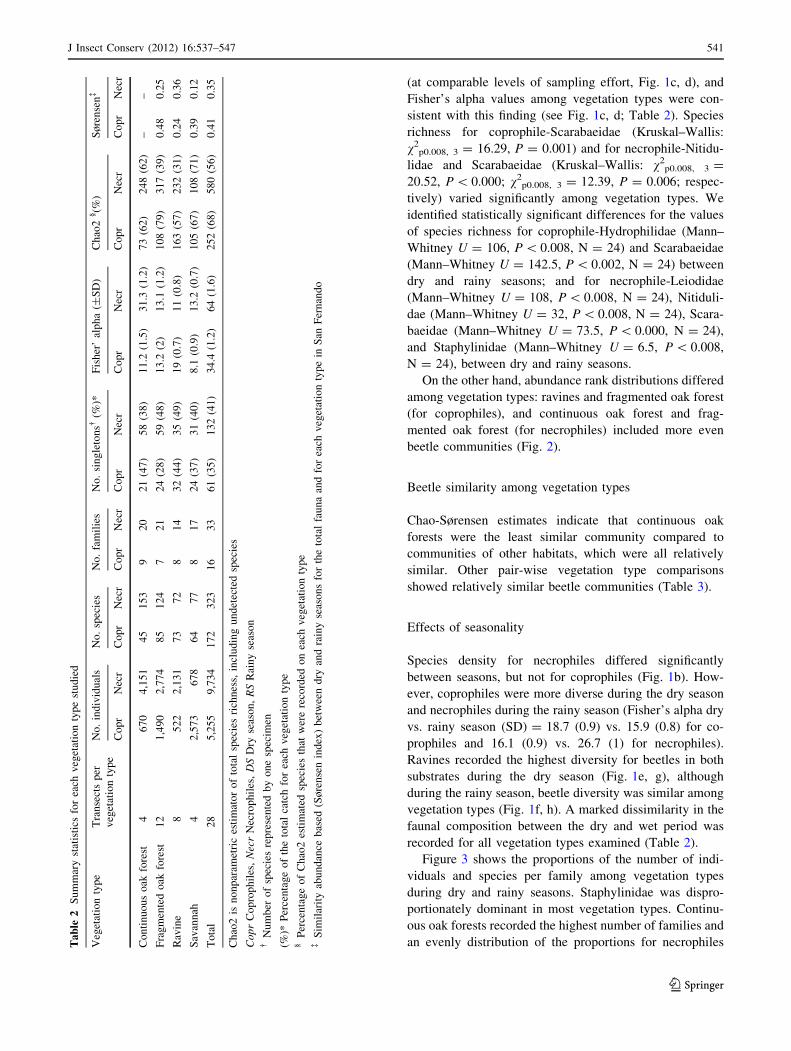
(at comparable levels of sampling effort, Fig. 1c, d), and
Fisher’s alpha values among vegetation types were con-
sistent with this finding (see Fig. 1c, d; Table 2). Species
richness for coprophile-Scarabaeidae (Kruskal–Wallis:
v2p0.008, 3 = 16.29, P = 0.001) and for necrophile-Nitidu-
lidae and Scarabaeidae (Kruskal–Wallis: v2p0.008, 3 =
20.52, P \ 0.000; v2p0.008, 3 = 12.39, P = 0.006; respec-
tively) varied significantly among vegetation types. We
identified statistically significant differences for the values
of species richness for coprophile-Hydrophilidae (Mann–
Whitney U = 106, P \ 0.008, N = 24) and Scarabaeidae
(Mann–Whitney U = 142.5, P \ 0.002, N = 24) between
dry and rainy seasons; and for necrophile-Leiodidae
(Mann–Whitney U = 108, P \ 0.008, N = 24), Nitiduli-
dae (Mann–Whitney U = 32, P \ 0.008, N = 24), Scara-
baeidae (Mann–Whitney U = 73.5, P \ 0.000, N = 24),
and Staphylinidae (Mann–Whitney U = 6.5, P \ 0.008,
N = 24), between dry and rainy seasons.
On the other hand, abundance rank distributions differed
among vegetation types: ravines and fragmented oak forest
(for coprophiles), and continuous oak forest and frag-
mented oak forest (for necrophiles) included more even
beetle communities (Fig. 2).
Beetle similarity among vegetation types
Chao-Sørensen estimates indicate that continuous oak
forests were the least similar community compared to
communities of other habitats, which were all relatively
similar. Other pair-wise vegetation type comparisons
showed relatively similar beetle communities (Table 3).
Effects of seasonality
Species density for necrophiles differed significantly
between seasons, but not for coprophiles (Fig. 1b). How-
ever, coprophiles were more diverse during the dry season
and necrophiles during the rainy season (Fisher’s alpha dry
vs. rainy season (SD) = 18.7 (0.9) vs. 15.9 (0.8) for co-
prophiles and 16.1 (0.9) vs. 26.7 (1) for necrophiles).
Ravines recorded the highest diversity for beetles in both
substrates during the dry season (Fig. 1e, g), although
during the rainy season, beetle diversity was similar among
vegetation types (Fig. 1f, h). A marked dissimilarity in the
faunal composition between the dry and wet period was
recorded for all vegetation types examined (Table 2).
Figure 3 shows the proportions of the number of indi-
viduals and species per family among vegetation types
during dry and rainy seasons. Staphylinidae was dispro-
portionately dominant in most vegetation types. Continu-
ous oak forests recorded the highest number of families and
an evenly distribution of the proportions for necrophilesTa
ble
2S
um
mar
yst
atis
tics
for
each
veg
etat
ion
typ
est
ud
ied
Veg
etat
ion
typ
eT
ran
sect
sp
er
veg
etat
ion
typ
e
No
.in
div
idu
als
No
.sp
ecie
sN
o.
fam
ilie
sN
o.
sin
gle
ton
s�(%
)*F
ish
er’
alp
ha
(±S
D)
Ch
ao2
§(%
)S
øre
nse
n�
Co
pr
Nec
rC
op
rN
ecr
Co
pr
Nec
rC
op
rN
ecr
Co
pr
Nec
rC
op
rN
ecr
Co
pr
Nec
r
Co
nti
nu
ou
so
akfo
rest
46
70
4,1
51
45
15
39
20
21
(47
)5
8(3
8)
11
.2(1
.5)
31
.3(1
.2)
73
(62
)2
48
(62
)–
–
Fra
gm
ente
do
akfo
rest
12
1,4
90
2,7
74
85
12
47
21
24
(28
)5
9(4
8)
13
.2(2
)1
3.1
(1.2
)1
08
(79
)3
17
(39
)0
.48
0.2
5
Rav
ine
85
22
2,1
31
73
72
81
43
2(4
4)
35
(49
)1
9(0
.7)
11
(0.8
)1
63
(57
)2
32
(31
)0
.24
0.3
6
Sav
ann
ah4
2,5
73
67
86
47
78
17
24
(37
)3
1(4
0)
8.1
(0.9
)1
3.2
(0.7
)1
05
(67
)1
08
(71
)0
.39
0.1
2
To
tal
28
5,2
55
9,7
34
17
23
23
16
33
61
(35
)1
32
(41
)3
4.4
(1.2
)6
4(1
.6)
25
2(6
8)
58
0(5
6)
0.4
10
.35
Ch
ao2
isn
on
par
amet
ric
esti
mat
or
of
tota
lsp
ecie
sri
chn
ess,
incl
ud
ing
un
det
ecte
dsp
ecie
s
Co
pr
Co
pro
ph
iles
,N
ecr
Nec
rop
hil
es,
DS
Dry
seas
on
,R
SR
ain
yse
aso
n�
Nu
mb
ero
fsp
ecie
sre
pre
sen
ted
by
on
esp
ecim
en
(%)*
Per
cen
tag
eo
fth
eto
tal
catc
hfo
rea
chv
eget
atio
nty
pe
§P
erce
nta
ge
of
Ch
ao2
esti
mat
edsp
ecie
sth
atw
ere
reco
rded
on
each
veg
etat
ion
typ
e�
Sim
ilar
ity
abu
nd
ance
bas
ed(S
øre
nse
nin
dex
)b
etw
een
dry
and
rain
yse
aso
ns
for
the
tota
lfa
un
aan
dfo
rea
chv
eget
atio
nty
pe
inS
anF
ern
and
o
J Insect Conserv (2012) 16:537–547 541
123
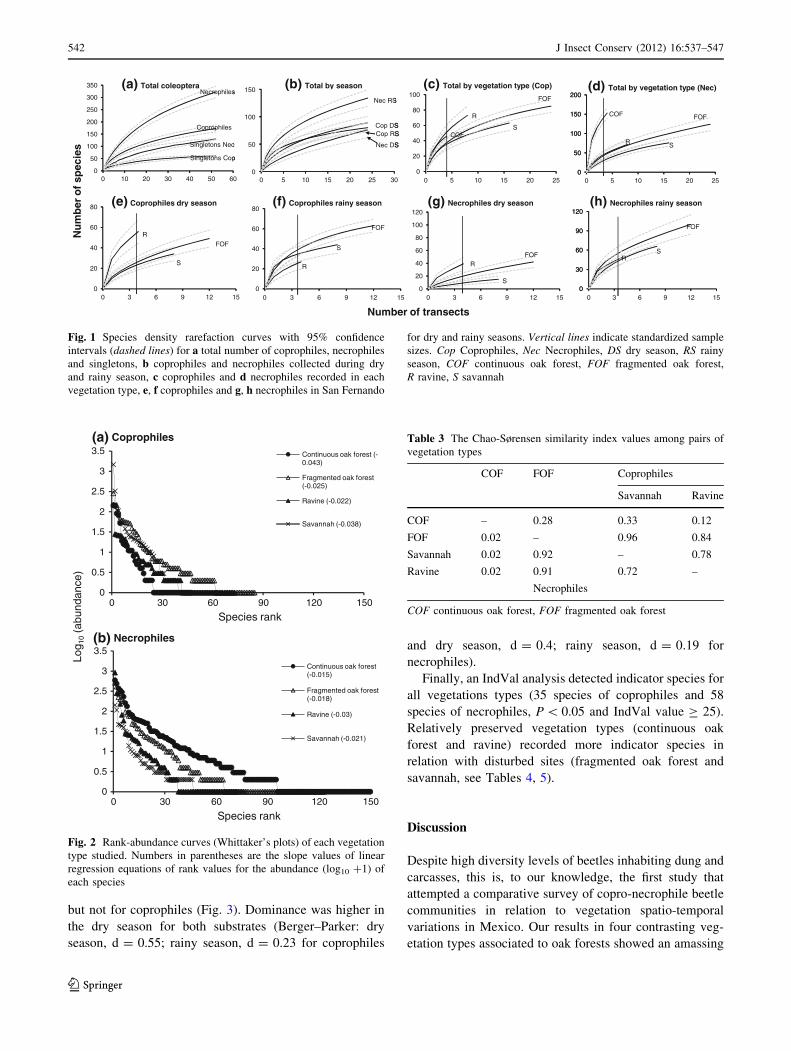
but not for coprophiles (Fig. 3). Dominance was higher in
the dry season for both substrates (Berger–Parker: dry
season, d = 0.55; rainy season, d = 0.23 for coprophiles
and dry season, d = 0.4; rainy season, d = 0.19 for
necrophiles).
Finally, an IndVal analysis detected indicator species for
all vegetations types (35 species of coprophiles and 58
species of necrophiles, P \ 0.05 and IndVal value C 25).
Relatively preserved vegetation types (continuous oak
forest and ravine) recorded more indicator species in
relation with disturbed sites (fragmented oak forest and
savannah, see Tables 4, 5).
Discussion
Despite high diversity levels of beetles inhabiting dung and
carcasses, this is, to our knowledge, the first study that
attempted a comparative survey of copro-necrophile beetle
communities in relation to vegetation spatio-temporal
variations in Mexico. Our results in four contrasting veg-
etation types associated to oak forests showed an amassing
0
20
40
60
80
100
120Necrophiles dry season
S
RFOF
Number of transects
0
20
40
60
80
100Total by vegetation type (Cop)
S
FOF
R
COF
0
20
40
60
80Coprophiles rainy season
R
FOF
S
0
30
60
90
120Necrophiles rainy season
R
FOF
S
0
50
100
150
200Total by vegetation type (Nec)
S
COF FOF
R
0
20
40
60
80 Coprophiles dry season
S
R
FOF
0
50
100
150
200
250
300
350Necrophiles
Coprophiles
Singletons Cop
Singletons Nec
Nu
mb
er o
f sp
ecie
s
0
50
100
150 Total by season
Nec DS
Nec RS
Cop DSCop RS
(g)
S
RFOF
(c)
S
FOF
R
COF
(f)
R
FOF
S
0
30
60
90
120(h)
R
FOF
S
0
50
100
150
200(d)
S
COF FOF
R
(e)
S
R
FOF
(a) Necrophiles
Coprophiles
Singletons Cop
Singletons Nec
0 3 6 9 12 15
0 5 10 15 20 25
0 3 6 9 12 15 0 3 6 9 12 15
0 5 10 15 20 25
0 3 6 9 12 15
0 10 20 30 40 50 60 0 5 10 15 20 25 30
(b)
Nec DS
Nec RS
Cop DSCop RS
Total coleoptera
Fig. 1 Species density rarefaction curves with 95% confidence
intervals (dashed lines) for a total number of coprophiles, necrophiles
and singletons, b coprophiles and necrophiles collected during dry
and rainy season, c coprophiles and d necrophiles recorded in each
vegetation type, e, f coprophiles and g, h necrophiles in San Fernando
for dry and rainy seasons. Vertical lines indicate standardized sample
sizes. Cop Coprophiles, Nec Necrophiles, DS dry season, RS rainy
season, COF continuous oak forest, FOF fragmented oak forest,
R ravine, S savannah
0
0.5
1
1.5
2
2.5
3
3.5
Species rank
Coprophiles
Continuous oak forest (-0.043)
Fragmented oak forest (-0.025)
Ravine (-0.022)
Savannah (-0.038)
(a)
0
0.5
1
1.5
2
2.5
3
3.5
0 30 60 90 120 150
0 30 60 90 120 150
Species rank
Necrophiles
Continuous oak forest (-0.015)
Fragmented oak forest (-0.018)
Ravine (-0.03)
Savannah (-0.021)
(b)
Log 1
0(a
bund
ance
)
Fig. 2 Rank-abundance curves (Whittaker’s plots) of each vegetation
type studied. Numbers in parentheses are the slope values of linear
regression equations of rank values for the abundance (log10 ?1) of
each species
Table 3 The Chao-Sørensen similarity index values among pairs of
vegetation types
COF FOF Coprophiles
Savannah Ravine
COF – 0.28 0.33 0.12
FOF 0.02 – 0.96 0.84
Savannah 0.02 0.92 – 0.78
Ravine 0.02 0.91 0.72 –
Necrophiles
COF continuous oak forest, FOF fragmented oak forest
542 J Insect Conserv (2012) 16:537–547
123

beetle diversity including 14,989 beetles representing 406
species and 33 families. This recorded number of necro-
phile and coprophile beetles represents a 29% of the beetle
families reported for Mexico and 26% for the Neotropics
(Costa 2000; Navarrete–Heredia and Fierros–Lopez 2001).
Nonetheless, in our habitat surveys we detected dissimilar
0
20
40
60
80
100
FOF (DS)FOF (RS) Rav (DS) Rav (RS) Sav (DS) Sav (RS) COFN
o. s
peci
es (
%)
(d) 2011 8 9 101312†
0
20
40
60
80
100
FOF (DS)FOF (RS) Rav (DS) Rav (RS) Sav (DS) Sav (RS) COF
No.
indi
vidu
als
(%)
(a) 0.215* 0.333 0.181 0.817 0.3610.1290.409
0
20
40
60
80
100
FOF (DS)FOF (RS) Rav (DS) Rav (RS) Sav (DS) Sav (RS) COF
No.
spe
cies
(%
)
(b) 1067 9 7 8 6†
Coprophiles
0
20
40
60
80
100
FOF (DS)FOF (RS) Rav (DS) Rav (RS) Sav (DS) Sav (RS) COF
No.
indi
vidu
als
(%)
0.143* 0.491 0.4 0.793 0.2630.1430.778
Necrophiles
Stap Scar Hist Niti Car Leio Other BeetlesHydr Trog
(c)
Fig. 3 Family proportion of coprophiles and necrophiles beetles
collected in four vegetation types in dry and rainy seasons. The
category of ‘‘other beetles’’ comprises 25 families representing 10 and
18% of the species and 1 and 3% of the abundance of coprophiles and
necrophiles, respectively. COF continuous oak forest, FOF frag-
mented oak forest, R ravine, S savannah, DS dry season, RS rainy
season. *Dominance (Berger-Parker index), �No. families
Table 4 Coprophilous beetle indicator species of each vegetation type using the IndVal analysis
COF FOF Ravine Savannah
Hydrophilidae Staphylinidae Histeridae Carabidae
Cercyon sp. 6 (100)** Aleochara oxypodia (51)* Euspilotus sp.2 (50)** Apristus mexicanus (25)*
Scarabaeidae Histeridae sp. 2 (38)* Scarabaeidae
Aphodinae sp. 10 (100)** Scarabaeidae Euoniticellus intermedius (52)**
Onthophagus cyanellus (100)** Uroxys sp. 1 (73)** Aphodinae sp. 5 (45)**
Onthophagus sp. 1 (100)** Aphodinae sp. 3 (44)** Staphylinidae
Aphodinae sp. 2 (99)** Staphylinidae Platystethus sp. (80)**
Staphylinidae Coproporus hepaticus (86)** Aleochara notula (69)**
Anotylus sp. 3 (100)** Anotylus sp. 1 (75)** Tinotus sp. 1 (62)**
Atheta sp. 10 (100)** Lissohypnus sp. 1 (50)** Phylonthus sp.1 (45)*
Myllaena sp. (100)** Belonuchus rufipennis (48)*
Oxytelus sp. (100)** Lithocharis sp. (33)*
Tinotus caviceps (100)** Neohypnus sp.1 (32)*
Atheta sp. 9 (85)**
Indicator value is given in parenthesis. Significance of Monte Carlo test: *P \ 0.05; **P \ 0.01
Names in bold denote beetle families
J Insect Conserv (2012) 16:537–547 543
123

responses between beetle assemblages among vegetation
types that might be related to particular preferences of each
group as well as to inherent effects of seasonally and
vegetation type conditions.
First, an examination of the diversity for coprophile
beetle families revealed that Staphylinidae and Scarabaei-
deae showed no strong differences for vegetation type
preferences (although see below; Davis et al. 1988; Dunn
and Danoff-Burg 2007; Caballero et al. 2009; Hopp et al.
2010). Staphylinidae are generalist predators (Hanski
1987), and most dung beetles—Scarabaeideae- are con-
sidered opportunistic—they tend to use a large variety of
dung types (Hanski and Cambefort 1991). The Histeridae
were relatively common and widely distributed across
vegetation types—except in continuous oak forests where
they were absent, likely because the margins of Histeridae
populations might be affected by physiographic habitat
boundaries—previous findings have shown a negative
effect of elevation on their distributions (Kovarik and
Caterino 2001). The Hydrophilidae, which are consumers
of dung (Didham et al. 1998), were more abundantly
trapped in ravines and continuous oak forests. Intriguingly,
the Nitidulidae, although mostly saprophagous and myce-
tophagous, were observed in the ravine during the rainy
season (in San Fernando), most likely reflecting their
opportunist foraging habits: they have been observed
feeding on a widely decaying substrates (Didham et al.
1998).
On the other hand, necrophile beetles exhibited con-
trasting patterns. The Staphylinidae, Scarabaeideae, and
Histeridae families were evenly distributed across vegeta-
tion types (except by the absence of Histeridae in contin-
uous oak forests), and showing a clear decrease in
abundance in the Savannah. The Hydrophilidae showed
relatively constant abundances across vegetation types,
except in the Savannah where they were absent. The Lei-
odidae are mostly scavengers that feed on various decaying
organic materials in usually moist forest (Peck 2001),
which might explain their presence in ‘‘forested’’ condi-
tions during the rainy season (oak forests and ravines).
Interestingly, the Carabidae were almost exclusively
present during the dry season in fragmented oak forests,
Table 5 Necrophilous beetle indicator species of each vegetation type using the IndVal analysis
COF FOF Ravine Savannah
Carabidae Staphylinidae Scarabaeidae Histeridae Staphylinidae
Dyscolus reflexicollis (100)** Aleocharinae sp. 2 (100)** Uroxys sp. 5 (35)* Euspilotus sp.2 (85)** Aleochara notula (31)*
Dyscolus teter (100)** Anotylus sp. 1 (100)** Hister coenosus (50)** Tenebrionidae
Hydrophilidae Atheta sp. 1 (100)** Xerosaprinus sp.3 (36)** Tenebrionidae sp. 8 (30)*
Cercyon sp. 6 (100)** Atheta sp. 4 (100)** Leiodidae
Cercyon sp. 7 (100)** Belonuchus sp. 1 (100)** Dissochetus sp. 2 (33)**
Cercyon sp. 9 (75)** Diestota sp. 1 (100)** Nitidulidae
Leiodidae Diestota sp. 2 (100)** Stelidota sp. 2 (45)**
Dissochetus sp. 1 (100)** Falagria sp. 1 (100)** Scarabaeidae
Dissochetus sp. 4 (100)** Gnypeta sp. (100)** Uroxys sp. 1 (86)**
Ptomaphagus sp. (100)** Homalotini sp. (100)** Uroxys sp. 4 (75)**
Nitidulidae Hoplandria sp. (100)** Uroxys sp. 2 (41)*
Aethina villosa (100)** Myllaena sp. (100)** Staphylinidae
Epuraea sp. 1 (100)** Parocyusa sp. 1 (100)** Hoplandria peltata (92)**
Stelidota sp. 3 (100)** Parocyusa sp. 2 (100)** Belonuchus rufipennis (76)**
Stelidota sp. 4 (100)** Phloenomus sp. 1 (100)** Coproporus hepaticus (70)**
Stelidota sp. 5 (100)** Phylonthus sp. 2 (100)** Aleocharinae sp. 23 (45)*
Scarabaeidae Platydracus sp. 1 (100)**
Onthophagus cyanellus (100)** Quedius sp. (100)**
Other families Tinotus caviceps (100)**
Cryptophagidae sp. 3 (100)** Ocyota sp. 1 (75)**
Cryptophagidae sp. 4 (100)** Phylonthus sp. 1 (75)**
Ptiliidae sp. 1 (100)** Diestota sp. 3 (75)**
Scydmaenidae sp. 4 (100)** Ocyota sp. 2 (50)**
Tenebrionidae sp. 2 (75)** Phloenomus sp. 2 (50)**
Indicator value is given in parenthesis. Significance of Monte Carlo test: *P \ 0.05; **P \ 0.01
Names in bold denote beetle families
544 J Insect Conserv (2012) 16:537–547
123

ravines, and savannah (Ball and Bousquet 2001), being a
potential explanation an early emergence of Carabidae to
avoid competition with other groups of comparable feeding
preferences (Andresen 2005).
Second, seasonality had a profound effect on the struc-
ture of beetle assemblages: changes in diversity of necro-
philes were particularly dramatic from dry to wet seasons
(see Fig. 1b, g, h). However, while no changes in diversity
levels for coprophiles between seasons were distinguished,
both diversity and community structure differed signifi-
cantly among vegetation types between seasons. In this
sense, most of the information available for dung beetles
(dung and carrion Scarabaeidae beetles) suggests this group
to be particularly sensitive to habitat suitability (Hanski
and Cambefort 1991), as well as to experience significant
decreases in diversity in dry tropical areas with a pro-
nounced seasonality (Janzen 1983; Andresen 2005, 2008;
Mendes and Linhares 2006). Furthermore, we found a high
dominance and a low diversity in most stressful conditions:
during the dry season and in the most disturbed vegetation
type (the case for savannah). These findings are related to
the fact that many coprophilous communities tend to be
numerically dominated by a few species (Doube 1987), that
this pattern is relatively common in open and climatically
unstable habitats (Magurran 2004), and that this phenom-
enon has been recorded for dung beetles in other Neo-
tropical dry forests (Escobar 1997; Andresen 2005, 2008).
Third, our results suggest that vegetation type and
conditions play an important role for population persis-
tence, and particularly for specialist beetles. For instance,
relatively large tracks of forest (i.e. ‘‘Huitepec’’ site) con-
tained the highest diversity of necrophiles, very likely
because an important number of vertebrates occur there
(Naranjo and Espinosa-Medinilla 2003) and hence the
availability of carcasses might increase accordingly. Con-
versely, in fragmented tropical areas (like in San Fernando)
certain vegetation types such as ravines constituted
potential refuges for insect fauna. These results are in line
with previous findings with beetle groups in which certain
pieces of land represent key elements for population per-
sistence (Gibbs and Stanton 2001; Halffter and Arellano
2002; Caballero et al. 2009; Vasquez-Velez et al. 2010). In
these fragmented landscapes, carrion of small mammals,
amphibians, birds and reptilians—which tend to concen-
trate in oak forest remnants—became the primary source of
food for necrophile populations. However, this pattern does
not hold for coprophiles perhaps because a large number of
mammals in the Neotropics tend to be rather small (\2 kg)
and only producing small amounts of dung (Emmons
1997)—the establishment and free movement of cattle in
areas like San Fernando represent a relatively important
supply of dung (Arellano et al. 2008a). We must consider
though that deliberately offering of dung or carrion in the
field could potentially bias beetle attraction, which in turn
might inflate diversity levels for a given vegetation type i.e.
the supply of trap baits could have altered the movement
behavior of beetles as these insects are able to locate baits
from a distance. Nonetheless, we consider the amount of
dung or carrion used in the baits small compared to what is
naturally available in the landscape, and hence movements
and behavior of beetles through the landscape are relatively
unbiased (see Arellano et al. 2008b).
Overall, our data highlight the need to preserve few
large forest tracts and key vegetation types (such as
ravines); these pieces of land not only counteract the
adverse effects of habitat loss and the impacts of stressful
conditions caused by seasonality, but constitute the reser-
voir of key species that occur there: [41% of the necro-
phile indicator species are Staphylinidae associated to
relatively pristine oak forests. Whilst seasonality and
vegetation type are important predictors of beetle diversity,
(measured as density, evenness, and distinctiveness), con-
ditions during an extended dry period coupled with
anthropogenic pressures underline the fact that key vege-
tation types (e.g.[21% of coprophile indicator species are
Staphylinidae restricted to ravines) play important roles as
refugees for many beetle species. These pieces of land
provide a diverse array of habitats, landforms and com-
munities that are naturally rich in biodiversity (Corbacho
et al. 2003). In a wider context, the native oak forests of our
study area have undergone high rates of forest loss during
the last three decades (in the case of the ‘‘highlands of
Chiapas’’ exhibiting a deforestation rate of 3.4% per year;
Echeverria et al. 2007), compared to many other forested
landscapes in the world (Cohen et al. 2002; Staus et al.
2002). These forests have been reduced severely and
degraded over time owing to logging for timber and fuel
wood, clearance for cultivation, and quarry developments.
The worry is that previous surveys (including ours) have
demonstrated that these patchy oak forests still contain
unusual levels of biodiversity and many uniquely Mexican
taxa of plants and animals (Peterson et al. 1993), thus
fragmentation and loss of forests can broadly be expected
to lose substantial amounts of biodiversity over the long
term (e.g. Bawa and Seidler 1998).
Our studies clearly demonstrates that the least frag-
mented oak forest and ravine are the most valuable areas
for necrophile and coprophile beetles in Neotropical
Mexico, especially for specialist beetles. Fragmentation,
via land-use conversion associated with agriculture and
exploitation will have a negative impact on beetle diversity
and overall biodiversity within this Neotropical area.
Acknowledgments We are grateful to San Fernando landowners
and PRONATURA, the administrator of Huitepec Reserve, for
allowing access to field sites. Taxonomical identifications were
J Insect Conserv (2012) 16:537–547 545
123

confirmed by S. J. Ashe� (Staphylinidae), G. E. Ball (Carabidae), M.
S. Caterino (Histeridae), A. R. Cline (Nitidulidae), S. B. Peck (Lei-
odidae). Dolores Vidal (UNICACH) provided laboratory rats. Manuel
Giron, Arcangel Molina and Irma Miss helped in the field. Michael
Caterino, Jack Longino, Robert Anderson and two anonymous
reviewers provided useful comments on early versions of this paper.
Funding was provided by The National Council for Science and
Technology (CONACYT: Grants J35230-V, CHIS-2005-C03-054,
CONACYT-SEMARNAT 108222, PIFOP-ECOSUR, and PATM-
ECOSUR).
References
Andresen E (2005) Effects of season and vegetation type on
community organization of dung beetles in a tropical dry forest.
Biotropica 37:291–300
Andresen E (2008) Dung beetle assemblages in primary forest and
disturbed habitats in a tropical dry forest landscape in Western
Mexico. J Insect Conserv 12:639–650
Anonymous (2006) Temperatura y precipitacion media mensual
1951–2006. Estaciones meteorologicas Tuxtla Gutierrez (DGE)
y San Cristobal de las Casas (La Cabana) Chiapas. Mexico.
Comision Nacional del Agua, Chiapas
Arellano L, Leon-Cortes JL, Halffter G (2008a) Response of dung
beetle assemblages to landscape structure in remnant natural and
modified habitats in southern Mexico. Insect Conserv Divers
1:253–262
Arellano L, Leon-Cortes JL, Ovaskainen O (2008b) Patterns of
abundance and movement in relation to landscape structure: a
study of a common scarab (Canthon cyanellus cyanellus) in
Southern Mexico. Landscape Ecol 23:69–78
Arnett RH Jr, Thomas MC, Skelley PE, Frank JH (2002) American
Beetles, Volume II: Polyphaga: Scarabaeoidea through Curcu-
lionoidea. CRC Press, Boca Raton
Ball GE, Bousquet Y (2001) Carabidae. In: Arnett RH Jr, Thomas
MC (eds) American Beetles. Volume 1. Archostemata, Myx-
ophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press
LLC, Boca Raton, pp 32–131
Bawa KS, Seidler R (1998) Natural forest management and conser-
vation of biodiversity in tropical forests. Conserv Biol 12:46–55
Bernon G (1981) Species abundance and diversity of the Coleoptera
component of a South African cow dung community, and
associated insect predators. Ph D dissertation, University of
Bowling Green, USA
Borror DJ, Triplehorn CA, Johnson NF (1989) An introduction to the
study of insects. Saunders College Publishing, Philadelphia
Caballero U, Leon-Cortes JL, Moron-Rıos A (2009) Response of rove
beetles (Staphylinidae) to various habitat types and change in
Southern Mexico. J Insect Conserv 13:67–75
Carter DO, Yellowlees D, Tibbett M (2007) Cadaver decomposition
in terrestrial ecosystems. Naturwissenschaften 94:12–24
Chao A (2005) Species richness stimation. In: Balakrishnan N, Read
CB, Vidakovic B (eds) Encyclopedia of statistical sciences.
Wiley, New York, pp 7909–7916
Chao A, Chazdon RL, Colwell RK, Shen T-J (2005) A new statistical
approach for assessing similarity of species composition with
incidence and abundance data. Ecol Lett 8:148–159
Cohen WB, Spies TA, Alig RJ, Oetter DR, Maiersperger TK, Fiorella
M (2002) Characterizing 23 years (1972–95) of stand replace-
ment disturbance in western Oregon forests with Landsat
imagery. Ecosystems 5:122–137
Colwell RK (2009) EstimateS: Statistical estimation of species
richness and shared species from samples, (Software and User’s
guide), Version 8.2. Persistent Available from URL: http://purl.
oclc.org/estimates
Corbacho C, Sanchez JM, Costillo E (2003) Patterns of structural
complexity and human disturbance of riparian vegetation in
agricultural landscapes of a Mediterranean area. Agric Ecosyst
Environ 95:495–507
Costa C (2000) Estado de conocimiento de los Coleoptera Neotrop-
icales. In: Martin-Piera F, Morrone J, Melic A (eds) Hacia un
proyecto CYTED para el inventario y estimacion de la divers-
idad entomologica en Iberoamerica. Pri-Bes-2000.m3 m: Mo-
nografıas Tercer Milenio 1:99–114
Creighton JC, Bastarache R, Lomolino MV, Belk MC (2009) Effect
of forest removal on the abundance of the endangered American
burying beetle, Nicrophorus americanus (Coleoptera: Silphidae).
J Insect Conserv 13:37–43
Davis ALV (1994) Habitat fragmentation in southern Africa anddistributional response patterns in five specialist or generalist
dung beetle families (Coleoptera). Afr J Ecol 32:192–207
Davis ALV, Doube BM, Maclennan PD (1988) Habitat associations
and seasonal abundance of dung beetles (Coleoptera: Staphylin-
idae, Hydrophilidae, Histeridae) in the Hluhluwe region of South
Africa. Bull Entomol Res 78:425–434
Didham RK, Lawton JH, Hammond PM, Eggleton P (1998) Trophic
structure stability and extinction dynamics of beetles (Coleop-
tera) in tropical forest fragments. Philos T Roy Soc B 353:437–
451
Dirzo R, Raven PH (2003) Global state of biodiversity and loss. Annu
Rev Environ Resour 28:133–167
Doube BM (1987) Spatial and temporal organization in communities
associated with dung pads and carcasses. In: Gee JHR, Giller PS
(eds) Organization of communities: past and present. British
Ecological Society symposium. Blackwell, Oxford, pp 255–280
Dufrene M, Legendre P (1997) Species assemblages and indicator
species: the need for a flexible assymetrical approach. Ecol
Monogr 67:345–366
Dunn RR, Danoff-Burg JA (2007) Road size and carrion beetle
assemblages in a New York forest. J Insect Conserv 11:325–332
Echeverria C, Cayuela L et al (2007) Spatial and temporal patterns of
forest loss and fragmentation in Mexico and Chile. In: Newton A
(ed) Biodiversity loss and conservation in fragmented forest
landscapes the forest of Montane Mexico and temperate South
America. CABI internacional, Wallingford, pp 14–42
Emmons LH (1997) Neotropical rainforest mammals: a field guide.
University of Chicago Press, Chicago
Escobar F (1997) Estudio de la comunidad de coleopteros coprofagos
(Scarabaeidae) en un remanente de bosque seco al norte del
Tolima, Colombia. Caldasia 19:419–430
Fahring L (2003) Effects of habitat loss and fragmentation on
biodiversity. Annu Rev Ecol Syst 34:487–515
Gibbs JP, Stanton EJ (2001) Habitat fragmentation and arthropod
community change: Burying beetles, phoretic mites and flies.
Ecol Appl 11:79–85
Gotelli N, Colwell RK (2001) Quantifying biodiversity: Procedures
and pitfalls in the measurement and comparison of species
richness. Ecol Lett 4:379–391
Halffter G, Arellano L (2002) Response of dung beetle diversity to
human-induced changes in a tropical landscape. Biotropica
34:144–154
Hanski I (1983) Coexistence of competitors in patchy environment.
Ecology 64:493–500
Hanski I (1987) Colonization of ephemeral habitats. In: Gray AJ,
Crawley MJ, Edwards PJ (eds) Colonization, succession and
stability. Blackwell, Oxford, pp 155–185
Hanski I, Cambefort Y (1991) Resource partitioning. In: Hanski I,
Cambefort Y (eds) Dung beetle ecology. Princeton University
Press, Princeton New Jersey, pp 330–349
546 J Insect Conserv (2012) 16:537–547
123

Holter P (1982) Resource utilization and local coexistence in a guild
of scarabaeid dung beetles (Aphodius spp.). Oikos 39:213–227
Hopp PW, Ottermanns R, Caron E, Meyer S, Roß-Nickoll M (2010)
Recovery of litter inhabiting beetle assemblages during forest
regeneration in the Atlantic forest of Southern Brazil. Insect
Conserv Divers 3:103–113
Janzen DH (1983) Seasonal changes in abundance of large nocturnal
dung beetles (Scarabaeidae) in Costa Rica deciduous forest and
adjacent horse pasture. Oikos 41:274–283
Klein B (1989) Effects of forest fragmentation on dung and carrion
beetle assemblages in central Amazonia. Ecology 70:1715–1725
Kocarek P (2000) A pitfall trap for ecology studies. Biol Bratislava
55:575–577
Kovarik PW, Caterino MS (2001) Histeridae. In: Arnett RH, Thomas
MC (eds) American Beetles, Volume 1. CRC Press, Boca Raton,
pp 212–227
Lamb D, Erskine P, Parrotta JA (2005) Restoration of degraded
tropical forest landscapes. Science 310:1628–1632
Magurran AE (2004) Measuring biological diversity. Blackwell
Publishing, Oxford
Mendes J, Linhares A (2006) Coleoptera associated with undisturbed
cow pats in pastures in Southeastern Brazil. Neotrop Entomol
35:715–723
Moore I (1954) An efficient method of collecting dung beetles. Pan-
Pac Entomol 30:208
Naranjo P, Espinosa-Medinilla E (2003) Los mamıferos de la reserva
ecologica Huitepec, Chiapas, Mexico. Rev Mex Mastozool
5:58–63
Navarrete–Heredia JL, Fierros–Lopez H (2001) Coleoptera de
Mexico: situacion actual y perspectivas de estudio. In: Navarr-
ete–Heredia JL, Fierros–Lopez H, Burgos–Solorio A (eds)
Topicos selectos sobre Coleoptera de Mexico. Universidad de
Guadalajara Jalisco/Universidad Autonoma del Estado de More-
los, Cuernavaca, pp 1–21
Nichols E, Larsen T, Spector S, Davis AL, Escobar F, Favila M,
Vulinec K (2007) Global dung beetle response to tropical forest
modification and fragmentation: a quantitative literature review
and meta-analysis. Biol Conserv 137:1–19
Novotny V, Basset Y (2000) Rare species in communities of tropical
insect herbivores: pondering the mystery of singletons. Oikos
89:564–572
Oliver I, Beattie AJ (1996) Invertebrate morphospecies as surrogates
for species: a case study. Conserv Biol 10:99–109
Parmenter RR, MacMahon JA (2009) Carrion decomposition and
nutrient cycling in a semi-arid shrub-steppe ecosystem. Ecol
Monogr 79:37–661
Payne JA (1965) A summer carrion study of the baby pig Sus scrofa
Linnaeus. Ecology 46:592–602
Peck SB (2001) Leiodidae. In: Arnett RH, Thomas MC (eds)
American Beetles, vol 1. CRC Press, Boca Raton, pp 250–258
Peterson AT, Flores-Villela A et al (1993) Conservation priorities in
Mexico: moving up in the world. Biodivers Lett 1:33–38
Pohl GR, Langor DW, Spence JR (2007) Rove beetles and ground
beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of
harvest and regeneration practices in western Canadian foothills
forests. Biol Conserv 137:294–307
Putman RJ (1983) Carrion and Dung: the decomposition of animal
wastes. The Institute of Biology’s Studies in Biology No. 156,
Edward Arnold, London
Shorrocks B (1990) Coexistence in a patchy environment. In:
Shorrocks B, Swingland R (eds) Living in a patchy environment.
Oxford University Press, Oxford, pp 91–116
Sokal RR, Rohlf FJ (1995) Biometry: the principles and practice of
statistics in Biological Research, 3rd edn. WH Freeman and
Company, New York
Staus NL, Strittholt JR, Della-Sala DA, Robinson R (2002) Rate and
pattern of forest disturbance in the Klamath-Siskiyou ecoregion,
USA between 1972 and 1992. Landscape Ecol 17:455–470
Tokeshi M (1993) Species abundance patterns and community
structure. Adv Ecol Res 24:111–186
Trumbo ST, Bloch PL (2000) Habitat fragmentation and burying
beetle abundance and success. J Insect Conserv 4:245–252
Ulrich W, Komosinski K, Zalewski M (2008) Body size and biomass
distributions of carrion visiting beetles: do cities host smaller
species? Ecol Res 23:241–248
Vasquez-Velez L, Bermudez C, Chacon P, Lozano-Zambrano F
(2010) Analysis of the richness of Staphylinidae (Coleoptera) on
different scales of a sub-Andean rural landscape in Colombia.
Biodivers Conserv 19:1917–1931
Wolf JM, Gibbs JP (2004) Silphids in urban forests: diversity and
function. Urban Ecosyst 7:371–384
J Insect Conserv (2012) 16:537–547 547
123
Related Documents











