RESEARCH ARTICLE High diversity and abundance of putative polyphosphate- accumulating Tetrasphaera -related bacteria in activated sludge systems Hien Thi Thu Nguyen, Vang Quy Le, Aviaja Anna Hansen, Jeppe Lund Nielsen & Per Halkjær Nielsen Department of Biotechnology, Chemistry, and Environmental Engineering, Aalborg University, Aalborg, Denmark Correspondence: Per Halkjær Nielsen, Department of Biotechnology, Chemistry, and Environmental Engineering, Aalborg University, Sohngaardsholmsvej 49, DK-9000 Aalborg, Denmark. Tel.: 145 9940 8503; fax: 145 9814 1808; e-mail: [email protected] Received 18 July 2010; revised 22 December 2010; accepted 29 December 2010. DOI:10.1111/j.1574-6941.2011.01049.x Editor: Alfons Stams Keywords EBPR; PAO; P-removal; microautoradiography; wastewater. Abstract The diversity of the putative polyphosphate-accumulating genus Tetrasphaera in wastewater treatment systems with enhanced biological phosphorus removal (EBPR) was investigated using the full-cycle rRNA approach combined with microautoradiography and histochemical staining. 16S rRNA actinobacterial gene sequences were retrieved from different full-scale EBPR plants, and the sequences belonging to the genus Tetrasphaera (family Intrasporangiaceae) were found to form three clades. Quantitative FISH analyses of the communities in five full-scale EBPR plants using 10 new oligonucleotide probes were carried out. The results showed that the probe-defined Tetrasphaera displayed different morphologies and constituted up to 30% of the total biomass. It was shown that active uptake of orthophosphate and formation of polyphosphate took place in most of the probe- defined Tetrasphaera populations. However, aerobic uptake of orthophosphate only took place after uptake of certain carbon sources under anaerobic conditions and these were more diverse than hitherto assumed: amino acids, glucose, and for some also acetate. Tetrasphaera seemed to occupy a slightly different ecological niche compared with ‘Candidatus Accumulibacter’ contributing to a functional redundancy and stability of the EBPR process. Introduction The enhanced biological phosphorus removal (EBPR) pro- cess has been implemented in many wastewater treatment plants (WWTPs) worldwide. When operated successfully, the EBPR process is a relatively inexpensive and environ- mentally sustainable option for phosphorus (P) removal; however, the stability and reliability of the EBPR process can be a problem (Seviour et al., 2003). EBPR is based on the ability of polyphosphate-accumulating organisms (PAOs) to take up P and accumulate it intracellularly as polyphosphate when exposed to alternating anaerobic [O 2 and nitrite/ nitrate absent] and aerobic conditions (Comeau et al., 1986; Wentzel et al., 1986). Many studies in both lab-scale and full-scale EBPR plants have shown that ‘Candidatus Accumulibacter phosphatis’ (referred to as Accumulibacter hereafter) are important PAOs and their enriched cultures generally behave as the biochemical models predict (Hesselmann et al., 1999; Cro- cetti et al., 2000; Liu et al., 2001; Zilles et al., 2002; Beer et al., 2006; Garcia Martin et al., 2006). Under anaerobic condi- tions, they take up low-molecular-weight organic acids (e.g. acetate, propionate, and pyruvate) using polyphosphate as an energy source. These substrates are stored as intracellular polyhydroxyalkanoates via energy from hydrolysis of intra- cellular polyphosphate and reducing power from glycolysis of intracellular glycogen (Mino et al., 1998), the tricar- boxylic acid cycle (Pereira et al., 1996; Louie et al., 2000; Lemos et al., 2003), or both (Wexler et al., 2009; Zhou et al., 2009). In the subsequent aerobic or denitrifying phase, the PAO can use polyhydroxyalkanoates for growth and for replenishing their polyphosphate and glycogen pools (van Loosdrecht et al., 1997; Garcia Martin et al., 2006; Zhou et al., 2009). Besides Accumulibacter , Gram-positive Tetrasphaera-re- lated organisms (Actinobacteria) are also putative PAOs that are abundant in many full-scale EBPR plants (Eschenhagen et al., 2003; Kong et al., 2005; Nielsen et al., 2010), often in higher numbers than Accumulibacter . Studies of their eco- physiology have, however, shown that they do not share all typical PAO characteristics known from Accumulibacter . They are able to take up phosphate and form polyphosphate FEMS Microbiol Ecol ]] (2011) 1–12 c 2011 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY ECOLOGY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R E S E A R C H A R T I C L E
High diversityandabundanceof putativepolyphosphate-accumulatingTetrasphaera-related bacteria inactivatedsludge systemsHien Thi Thu Nguyen, Vang Quy Le, Aviaja Anna Hansen, Jeppe Lund Nielsen & Per Halkjær Nielsen
Department of Biotechnology, Chemistry, and Environmental Engineering, Aalborg University, Aalborg, Denmark
Correspondence: Per Halkjær Nielsen,
Department of Biotechnology, Chemistry, and
Environmental Engineering, Aalborg
University, Sohngaardsholmsvej 49, DK-9000
Aalborg, Denmark. Tel.: 145 9940 8503; fax:
145 9814 1808; e-mail: [email protected]
Received 18 July 2010; revised 22 December
2010; accepted 29 December 2010.
DOI:10.1111/j.1574-6941.2011.01049.x
Editor: Alfons Stams
Keywords
EBPR; PAO; P-removal; microautoradiography;
wastewater.
Abstract
The diversity of the putative polyphosphate-accumulating genus Tetrasphaera in
wastewater treatment systems with enhanced biological phosphorus removal
(EBPR) was investigated using the full-cycle rRNA approach combined with
microautoradiography and histochemical staining. 16S rRNA actinobacterial gene
sequences were retrieved from different full-scale EBPR plants, and the sequences
belonging to the genus Tetrasphaera (family Intrasporangiaceae) were found to
form three clades. Quantitative FISH analyses of the communities in five full-scale
EBPR plants using 10 new oligonucleotide probes were carried out. The results
showed that the probe-defined Tetrasphaera displayed different morphologies and
constituted up to 30% of the total biomass. It was shown that active uptake of
orthophosphate and formation of polyphosphate took place in most of the probe-
defined Tetrasphaera populations. However, aerobic uptake of orthophosphate
only took place after uptake of certain carbon sources under anaerobic conditions
and these were more diverse than hitherto assumed: amino acids, glucose, and for
some also acetate. Tetrasphaera seemed to occupy a slightly different ecological
niche compared with ‘Candidatus Accumulibacter’ contributing to a functional
redundancy and stability of the EBPR process.
Introduction
The enhanced biological phosphorus removal (EBPR) pro-
cess has been implemented in many wastewater treatment
plants (WWTPs) worldwide. When operated successfully,
the EBPR process is a relatively inexpensive and environ-
mentally sustainable option for phosphorus (P) removal;
however, the stability and reliability of the EBPR process can
be a problem (Seviour et al., 2003). EBPR is based on the
ability of polyphosphate-accumulating organisms (PAOs) to
take up P and accumulate it intracellularly as polyphosphate
when exposed to alternating anaerobic [O2 and nitrite/
nitrate absent] and aerobic conditions (Comeau et al.,
1986; Wentzel et al., 1986).
Many studies in both lab-scale and full-scale EBPR plants
have shown that ‘Candidatus Accumulibacter phosphatis’
(referred to as Accumulibacter hereafter) are important
PAOs and their enriched cultures generally behave as the
biochemical models predict (Hesselmann et al., 1999; Cro-
cetti et al., 2000; Liu et al., 2001; Zilles et al., 2002; Beer et al.,
2006; Garcia Martin et al., 2006). Under anaerobic condi-
tions, they take up low-molecular-weight organic acids (e.g.
acetate, propionate, and pyruvate) using polyphosphate as
an energy source. These substrates are stored as intracellular
polyhydroxyalkanoates via energy from hydrolysis of intra-
cellular polyphosphate and reducing power from glycolysis
of intracellular glycogen (Mino et al., 1998), the tricar-
boxylic acid cycle (Pereira et al., 1996; Louie et al., 2000;
Lemos et al., 2003), or both (Wexler et al., 2009; Zhou et al.,
2009). In the subsequent aerobic or denitrifying phase, the
PAO can use polyhydroxyalkanoates for growth and for
replenishing their polyphosphate and glycogen pools (van
Loosdrecht et al., 1997; Garcia Martin et al., 2006; Zhou
et al., 2009).
Besides Accumulibacter, Gram-positive Tetrasphaera-re-
lated organisms (Actinobacteria) are also putative PAOs that
are abundant in many full-scale EBPR plants (Eschenhagen
et al., 2003; Kong et al., 2005; Nielsen et al., 2010), often in
higher numbers than Accumulibacter. Studies of their eco-
physiology have, however, shown that they do not share all
typical PAO characteristics known from Accumulibacter.
They are able to take up phosphate and form polyphosphate
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY

under aerobic conditions, but cannot take up short-chain
fatty acids under anaerobic conditions, and do not store
polyhydroxyalkanoates (Kong et al., 2005). They can take up
some amino acids, and some Tetrasphaera seem capable of
fermenting glucose (Kong et al., 2008). Furthermore, they
express extracellular surface-associated amylases for degrada-
tion of starch (Xia et al., 2008). These results suggest that the
ecophysiology of PAOs – and particularly Tetrasphaera-
related PAOs – is much more diverse than hitherto anticipated.
A few isolates of the genus Tetrasphaera are available.
These include Tetrasphaera australiensis, Tetrasphaera japo-
nica (Maszenan et al., 2000), Tetrasphaera elongata (strain
LP2) (Hanada et al., 2002), T. elongata (strain ASP12) (Onda
& Takii, 2002), and the filamentous Tetrasphaera jenkinsii,
Tetrasphaera vanveenii, and Tetrasphaera veronensis (McKen-
zie et al., 2006). Storage of intracellular polyphosphate has
been found in all isolates, but only the filamentous species
are able to synthesize polyhydroxyalkanoates (McKenzie
et al., 2006). Interestingly, some of these cultured Tetra-
sphaera have some ecophysiological traits rather similar to
those of the uncultured Tetrasphaera-related PAO from full-
scale EBPR plants. For T. elongata (strain LP2), their aerobic
P uptake depends on an anaerobic uptake of glucose, and for
T. elongata strain ASP12, T. australiensis, and T. japonica, an
amino acid mixture (casamino acids) is required. However,
the strains are difficult to grow in pure culture, and more
detailed studies of their physiology are lacking.
The diversity of the genus Tetrasphaera is not well
described in full-scale EBPR plants. Surveys using FISH
applying probes targeting species from the Tetrasphaera
genus, Actino-221 and Actino-658 (Kong et al., 2005), have
shown that they usually grow as cocci in clusters of tetrads
and short rods in clumps. We have, however, occasionally
observed other morphologies of probe-defined Tetra-
sphaera, and so the entire diversity seems to be not yet well
described among these putative PAOs. Furthermore, it is
uncertain whether they all are PAOs capable of accumulating
intracellular polyphosphate in full-scale plants.
The aim of this study was to investigate the diversity, the
function, and the abundance of the genus Tetrasphaera in
full-scale EBPR WWTPs. A better understanding of their
ecophysiology, ecology, and abundances is needed to under-
stand the role of the Tetrasphaera-related PAOs in full-scale
EBPR plants and be able to manipulate the communities for
optimal P removal and troubleshooting.
Materials and methods
Activated sludge sampling
Activated sludge samples were collected from six Danish
full-scale WWTPs (Ejby Mølle, Hjørring, Abenra, Skive,
Aalborg East, and Odense), all of which were well-working
EBPR plants with nitrogen removal (nitrification and deni-
trification) and stable over several years. Effluent concentra-
tions are always below 1.0 mg total P L�1 and 8 mg total
N L�1. The C/P ratio in incoming wastewater was between
60 and 90 g total chemical oxygen demand g�1 total P for all
plants and 4 90% of the P was removed during treatment.
More information about the plants can be found elsewhere
(Nielsen et al., 2010). Samples from three plants (Ejby
Mølle, Hjørring, and Aalborg East) were used for studies of
the ecophysiology of Tetrasphaera. Two plants (Ejby Mølle
and Aalborg East) have a BioDeniphos configuration (Se-
viour & Blackall, 1999), and Hjørring WWTP has recirculat-
ing operation. Fresh samples from aeration tanks were stored
at 4 1C for o 24 h to carry out all the experiments. The
samples were diluted with filtered nitrate and nitrite-free
effluent to a final concentration of 1 g suspended solids L�1
before the experiments on ecophysiology were performed.
Clone library construction, clone screening,and sequencing
Community DNA was extracted from five activated sludge
samples using the PowerSoilTM DNA Isolation Kit (Mo Bio).
All extracted DNA were pooled and used for construction of
four clone libraries based on PCR amplification of 16S
rRNA genes with four different primer sets to get as many
different sequences as possible. Set A consisted of primers
27F and 1492R targeting Universal bacteria (Lane, 1991); set
B consisted of 27F and actino-1011R targeting T. japonica
(Liu et al., 2001); set C consisted of Actino-221F targeting
many Tetrasphaera (Kong et al., 2005) and 1492R, and set D
consisting of HGC236F targeting most Actinobacteria (Er-
hart et al., 1997) and 1492R. A touchdown PCR program
was applied: 5 min denaturing at 94 1C, followed by 10 cycles
of: (1) 94 1C for 30 s, (2) 1 min annealing at 60, 62, and 68 1C
(decreasing 1 1C per cycle) for primer sets A, B, and C,
respectively, and (3) 72 1C for 45 s, followed by 20 cycles
with the same denaturing and extension conditions, but
with 1 min annealing at 10 1C below the initial annealing
temperature, finalized with a 7-min extension at 72 1C. The
optimized template amount was 20–200 ng DNA per PCR
reaction. The PCR products were ligated into the pCRII-
TOPO vector (Invitrogen, Groningen, the Netherlands), and
clones with the correct 16S rRNA gene inserts (confirmed by
fragment size after PCR amplification) were sequenced with
the PCR primers by Macrogen (Seoul, Korea).
Phylogenetic analysis
The 16S rRNA gene sequences obtained were checked for
their chimeric properties using the MALLARD software (Ashel-
ford et al., 2006). Nearly full-length 16S rRNA gene
sequences were imported into the ARB software package
(Ludwig et al., 2004), aligned, and a phylogenetic tree was
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
2 H.T.T. Nguyen et al.

constructed based on comparative analysis of aligned 16S
rRNA gene nucleotide sequences. Phylogenetic trees were
calculated using the neighbor-joining method with a Pois-
son correction model and a 100-replicate bootstrap analysis.
The branching pattern was compared with trees generated
using the neighbor-joining algorithm.
FISH, polyhydroxyalkanoate and 4,6-diamino-2-phenylindole (DAPI) staining
FISH was performed as detailed in Nielsen (2009). Lysozyme
[0.5 g L�1 in 100 mM Tris (pH 7.5) and 5 mM EDTA] was
used to increase the permeability of the cells. The oligonu-
cleotide probes EUBmix (equimolar concentration of
EUB338, EUB338II, and EUB338III) targeting most bacteria
(Amann et al., 1990; Daims et al., 1999) were labeled by
FLUOS [5(6)-carboxyfluorescein-N-hydroxy-succinimide
ester], whereas PAOmix (equimolar concentration of
PAO462, PAO651, and PAO846) targeting most Accumuli-
bacter (Crocetti et al., 2000), HGC69a targeting actinobacter-
ial high-G1C Gram-positive bacteria (Roller et al., 1994),
HGC236 targeting Actinobacteria (Erhart et al., 1997), acti-
no-1011 targeting T. japonica (Liu et al., 2001), Actino-221
and Actino-658 targeting potential Tetrasphaera-PAOs (Kong
et al., 2005) were labeled with either FLUOS or Cy3 (sulfoin-
docyanine dyes). Their specificities, hybridization require-
ments, and reference information are described in probeBase
(Loy et al., 2003). Twenty separate microscopy images were
taken for quantification FISH analysis, with the final results
reflecting the mean percentages of Tetrasphaera of the entire
bacterial community detected by EUBmix (Nielsen, 2009).
The sample was stained with DAPI (0.2 mM final con-
centration) for 5 min after FISH, rinsed thoroughly with
sterile distilled water, and air-dried. The DAPI signal of
stained cells was examined using a Zeiss Axiovision epi-
fluorescence microscope (Carl Zeiss) equipped with a CCD
camera (Quantex, Photometrics). With this DAPI concen-
tration, the polyphosphate granules appear bright yellow,
while the cells remain pale blue (Kawaharasaki et al., 1999).
For FISH analysis, the same field examined for DAPI was
examined under the corresponding filter sets for fluores-
cence labels Cy3 and FLUOS.
Presence of polyhydroxyalkanoates as intracellular sto-
rage polymers was tested by Nile Blue staining largely as
described elsewhere (Kragelund et al., 2005). FISH was,
however, performed first followed by chemical staining and
relocation of the same microscopic field.
Design and testing of oligonucleotide probes
Tetrasphaera-specific probes were designed using the probe
design tool in the ARB software package (Ludwig et al., 2004).
The specificities of these probes were confirmed using the
CHECK PROBE program in the Ribosomal Database Project
(Maidak et al., 2001). The formamide concentration for
optimum probe stringency was determined by performing a
series of FISH experiments at 5% formamide increments
from 0% to 60% formamide on isolates when possible or on
activated sludge. The fluorescence intensities of single cells
from at least 20 images for each probe were evaluated by
image analysis using IMAGEJ software (Collins, 2007). The
specificities of new Tetrasphaera probes and the formamide
concentrations used are described in Table 1. Three reference
strains, T. elongata, T. jenkinsii, and T. australiensis (pro-
vided by R. Seviour, La Trobe University, Australia), were
used in this study. All strains were grown on GS or R2A
media (Maszenan et al., 2000).
Microautoradiography--FISH
A microautoradiography–FISH method was slightly mod-
ified from that described by Nielsen & Nielsen (2005) and
Kong et al. (2005). Briefly, 2 mL activated sludge samples
were incubated in 9 mL serum bottles with labeled and
unlabeled substrates under aerobic or anaerobic conditions.
All anaerobic preparations were carefully flushed with O2-
free N2. After incubation they were fixed by addition of 96%
ethanol and allowed to stand for 3 h at 4 1C, and they were
subsequently washed in a phosphate-buffered saline (PBS)
at 4 1C and finally resuspended in ice-cold PBS and ice-cold
96% ethanol (1 : 1). The samples were gently homogenized
between two gelatin-coated cover glasses (24� 60 mm).
FISH probing of microautoradiography-incubated bio-
mass, coating with emulsion, exposure, and development of
the hybridized FISH slides before being examined by micro-
scopy were carried out as described previously (Nielsen &
Nielsen, 2005). Each microautoradiography experiment was
repeated at least three times in different WWTPs. Controls
for nonradioactively induced silver grain formation (che-
mography) were always included, and no microautoradio-
graphy-positive cells were ever observed with any of the
nonradioactive chemicals used in this study.
The ability to take up labeled orthophosphate (33P) by
Tetrasphaera was investigated as follows. Before the incuba-
tions with 33P, sludge samples were incubated anaerobically
with different carbon sources (1 mM). Any unconsumed
carbon source was removed by washing the samples three
times with the filtered effluent water before labeled 33P and
nonlabeled P were added to a final concentration of 0.3 mM
at the very beginning of a 4-h incubation period with
oxygen. Samples for microautoradiography analyses were
taken at the end of the incubation.
In order to investigate the storage capacity of Tetra-
sphaera and Accumulibacter, it was investigated whether they
could continuously take up labeled acetate or glucose
(1 mM, 2 h incubation) after prolonged anaerobic conditions
(0, 3, 5, and 9 h) with unlabeled acetate present (2 mM).
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
3Tetrasphaera-related bacteria in activated sludge systems

The sources and specifications of all the radioactive
chemicals in this study have been described previously
(Kong et al., 2005).
Nucleotide sequence accession numbers
The 16S rRNA gene sequences obtained in this study have
been deposited in the GenBank database under accession
numbers GU552242–GU552268.
Results
Clone library analysis
Four clone libraries were constructed based on the commu-
nity 16S rRNA genes amplified by four primer sets. A total of
189 16S rRNA gene sequences were retrieved, and 93
sequences in 10 different actinobacterial families were
identified. Nearly one-third of these sequences belonged to
the family Intrasporangiaceae in the Actinobacteria. Their
phylogenetic relationship with other closely related bacteria
is shown in the phylogenetic tree (Fig. 1). The clone
sequences grouped into three separate clades. Clade 1
includes clones related to sequences of T. elongata, which
was isolated from activated sludge (98.1–99.8% identity),
and Tetrasphaera duodecadis (97.6–98.9% identity). Clade 2
is related to T. jenkinsii (96.5–96.9% identity), T. australien-
sis (97.1–97.4% identity), ‘Candidatus Nostocoida limicola’
(96.3–96.7% identity), and T. veronensis (96.7–97% iden-
tity), all isolated from activated sludge. Clones in clade 3
shared 98.2–99.9% identity and were not related to any
cultured species. Besides these sequences, some clones were
related to T. japonica (97.1–97.5% identity) isolated from
activated sludge and some to Tetrasphaera sp. Ellin176
(96.1–97.0% identity) isolated from a soil sample. These
clones did not form other distinct clades or clusters.
Design, testing, and optimization ofoligonucleotide probes
A number of oligonucleotide probes with different specifi-
cities were designed to investigate morphologies and abun-
dances and for the determination of important
ecophysiological traits (Table 1). Ten probes were designed
to target most, but not all, of the clones belonging to the
family Intrasporangiaceae retrieved in this study and 24
other partial and full sequences (Crocetti et al., 2000; Kong
et al., 2005). Attempts to design probes covering entire
clades (Fig. 1) or all Tetrasphaera sequences were not
successful.
The 10 probes were tested and optimized on samples
from five different EBPR WWTPs (Ejby Mølle, Hjørring,
Abenra, Skive, and Odense) and on pure Tetrasphaera
cultures. The sequences related to T. elongata (clade 1) were
targeted by probes Elo1-1250, Tet1-823, and Tet1-266. The
morphology of these bacteria was very diverse (Table 2).
Bacteria related to T. jenkinsii, T. australiensis, and T.
veronensis (clade 2) were targeted by five probes, and they
also showed large differences in morphology (Table 2 and
Fig. 2). Two morphotypes have not been described before:
the thin filaments (approximately 0.45mm in width and
25–60 mm in length) and the small branched rods. Clade 3
without cultured representatives was targeted by probes
Tet3-654 and Tet3-19. Probe Tet3-19-targeted bacteria were
cocci in clusters of tetrad, whereas Tet3-654-targeted bacter-
ia had four different morphologies (cluster of tetrads, small
cocci, branched rods, and short rods in clumps) (Fig. 2).
Probes targeting bacteria related to T. japonica and Tetra-
sphaera sp. Ellin176 were also designed, but the targeted cells
were present in only one of the treatment plants investigated
and only in very low numbers (o 1% of the EUB-targeted
cells, data not shown). Bacteria targeted by these probes
were not further investigated.
Table 1. Oligonucleotide probes designed for detection of Tetrasphaera in EBPR plants
Name Abbreviation Clade Target group Sequence (50–30) % Formamide
S-S-Elo1-1250-a-A-18 Elo1-1250 1 T. elongata and related clones CGCGATTTCGCAGCCCTT 20
S-S-Tet1-823-a-A-18 Tet1-823 1 T. elongata-related clones TGAGACCCGCACCTAGTT 30
S-S-Tet1-266-a-A-18 Tet1-266 1 Clone ASM31 CCCGTCGTCGCCTGTAGC 25
S-S-Tet2-842-a-A-18 Tet2-842 2 Clones related to T. jenkinsii, T. australiensis,
and T. veronensis
GCGGCACAGAACTCGTGA 30
S-S-Tet2-831-a-A-18 Tet2-831 2 T. australiensis, T. veronensis,
and uncultured Tetrasphaera
TCGTGAAATGAGTCCCAC 10
S-S-Tet2-892-a-A-18 Tet2-892 2 Clone ASM47 TAGTTAGCCTTGCGGCCG 5
S-S-Tet2-87-a-A-18 Tet2-87 2 Uncultured Tetrasphaera TCGCCACTGATCAGGAGA 10
S-S-Tet2-174-a-A-18 Tet2-174 2 T. jenkinsii, T. australiensis, T. veronensis,
and Candidatus N. limicola
GCTCCGTCTCGTATCCGG 20
S-S-Tet3-654-a-A-18 Tet3-654 3 Uncultured Tetrasphaera GGTCTCCCCTACCATACT 35
S-S-Tet3-19-a-A-18 Tet3-19 3 Clone ASM57 CAGCGTTCGTCCTACACA 0
Coverage is also shown in Fig. 1.
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
4 H.T.T. Nguyen et al.

Ecophysiology determined bymicroautoradiography-FISH andchemical staining
All probe-defined Tetrasphaera showed intracellular DAPI-
positive granules indicating the presence of polyphosphate.
In order to investigate whether they had an active P uptake
when changing from anaerobic to aerobic conditions, a
number of experiments using 33P-labeled phosphate were
conducted. The five most abundant probe-defined popula-
tions were tested, and four of them could take up 33P under
aerobic conditions after a 2-h anaerobic preincubation
period with organic substrates (Table 3): clade 1 (targeted
by probe Tet1-266), clade 2 (Tet2-892 and Tet2-174), clade 3
(Tet3-654), but not those targeted by probe Tet3-19. Two
important observations were made: (1) the uptake
Uncultured bacteria, AF387312Uncultured bacteria, AF387313Uncultured bacteria, AF387307Tetrasphaera elongata, AB030911
Uncultured bacteria, AF387308Uncultured bacteria, AF387311
Clone ASM28, GU552252Clone ASM29, GU552253
Clone ASM31, GU552254 Clone ASM26, GU552251
Tetrasphaera duodecadis, AB072496Clone ASM14, GU552247
Clone ASM3, GU552243Clone ASM47, GU552259
Clone ASM12, GU552246Clone ska14, AY710276Clone ska15, AY710277
Clone ska20, AY710275Clone ska2, AY710287Clone ska22, AY710283Candidatus Nostocoida limicola, Y14596Tetrasphaera jenkinsii, DQ007321Tetrasphaera australiensis, AF125091
Tetrasphaera veronensis, Y14595Uncultured bacteria, AF387314
Clone ska9, AY710273Clone ASM24, GU552249
Clone ASM57, GU552268Clone ASM62, GU552263Clone ska3, AY710288Clone ASM59, GU552260
Clone ska6, AY710272Clone ska16, AY710278
Clone ska5, AY710289Clone ska21, AY710284
Clone ska25, AY710284Clone ASM64, GU552265Clone Ska12, AY710274Clone ASM63, GU552264Clone ASM66, GU552267Clone ASM65, GU552266
Clone ska26, AY710285Clone ASM25, GU552250Clone ASM33, GU552255
Clone ASM8, GU552245 Clone ASM44, GU552257Clone ska7, AY710271Clone ASM61, GU552262
Clone ska28, AY710286Clone ASM6, GU552244
Clone ASM60, GU552261Tetrasphaera japonica, AF125092
0.01
Tet
1-82
3T
et1-
266
Tet
2-84
2
Elo
1-12
50T
et3-
654
Tet
3-19
Tet
2-83
1
Tet
2-87
Tet
2-17
4T
et2-
892
Cla
de 1
Cla
de 2
Cla
de 3
Clone ASM1, GU552242Clone ASM39, GU552256
Clone ASM45, GU552251Clone ASM16, GU552248
Clone ska19, AF710281Tetrasphaera sp. Ellin176, AF409018
85
60
87
95
54
57
8758
6856
88
7651
55
66
97
97
9867
8096
9669
82
82
4866
60
86
72
81
Out
grou
p
Ornithinicoccus hortensis, Y17869
Fig. 1. Distance matrix tree of the genus Tetrasphaera in the Actinobacteria showing high diversity of Tetrasphaera-related bacteria in an EBPR-
activated sludge system. Ten betaproteobacterial sequences were used as the outgroup. Boldface names indicate the sequences obtained in this study.
Each sequence represents one clone. The dotted parts of the bracket lines indicate that the sequences are not covered by probes. The bar indicates 1%
sequence divergence.
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
5Tetrasphaera-related bacteria in activated sludge systems
depended on the type of organic substrate present during
the 2-h anaerobic preincubation and (2) only certain
morphotypes from each probe-defined population could
take up P.
When raw wastewater was added as substrate under
anaerobic conditions, all probe-defined morphotypes took
up 33P in the subsequent aerobic stage except some bacteria
from clade 3 targeted by probe Tet3-19, and the filamentous
morphotype of clade 2 (hybridizing with probe Tet2-174).
In control experiments without any added external organic
substrate in the anaerobic preincubation, none of the probe-
defined populations were able to take up 33P in the aerobic
period. This indicated that they were all able to accumulate
energy reserves during the anaerobic period to take up 33P
under aerobic conditions, presumably by taking up organic
substrates. The effect of specific organic substrates during
anaerobic preincubation on the subsequent 33P uptake by
the Tetrasphaera-PAO is shown in Table 4. The four sub-
strates tested (casamino acids, glutamic acid, glucose, and
acetate) were consumed by most Tetrasphaera. Thus, the
physiological traits shown here meet the criteria for PAOs,
and so the four probe-defined populations are called Tet1-
266-PAO, Tet2-892-PAO, Tet2-174-PAO, and Tet3-654-PAO
here.
The ability of Tetrasphaera-PAOs to take up acetate,
glucose, casamino acid, and glutamic acid was investigated
under both aerobic and anaerobic conditions (Table 4). The
uptake pattern was very similar for three of the probe-
defined populations (Tet2-892-PAO, Tet2-174-PAO, and
Tet3-654-PAO) as they could take up all four substrates
under both conditions. Only Tet1-266-PAOs were unable to
take up acetate and glutamic acid. Interestingly, Tet3-19-
defined populations could take up glucose and glutamic
acid under both anaerobic and aerobic conditions (Table 4),
but were not able to take up 33P (Table 3) despite the
presence of intracellular DAPI-positive granules.
None of the probe-defined nonfilamentous Tetrasphaera
bacteria had any intracellular polyhydroxyalkanoates as
tested by Nile Blue staining. Because most Tetrasphaera were
able to take up acetate and other organic compounds under
anaerobic conditions, it was investigated whether they could
continuously take up acetate under prolonged anaerobic
conditions, or whether they reached a saturation level
indicative of storage capacity. No Tetrasphaera-PAOs could
assimilate acetate after 3, 5, and 9 h of preincubation with
unlabeled acetate (Table 5). Therefore, under anaerobic
conditions, the storage capacity of Tetrasphaera-PAO was
saturated after 3 h. This indicated that no growth took place
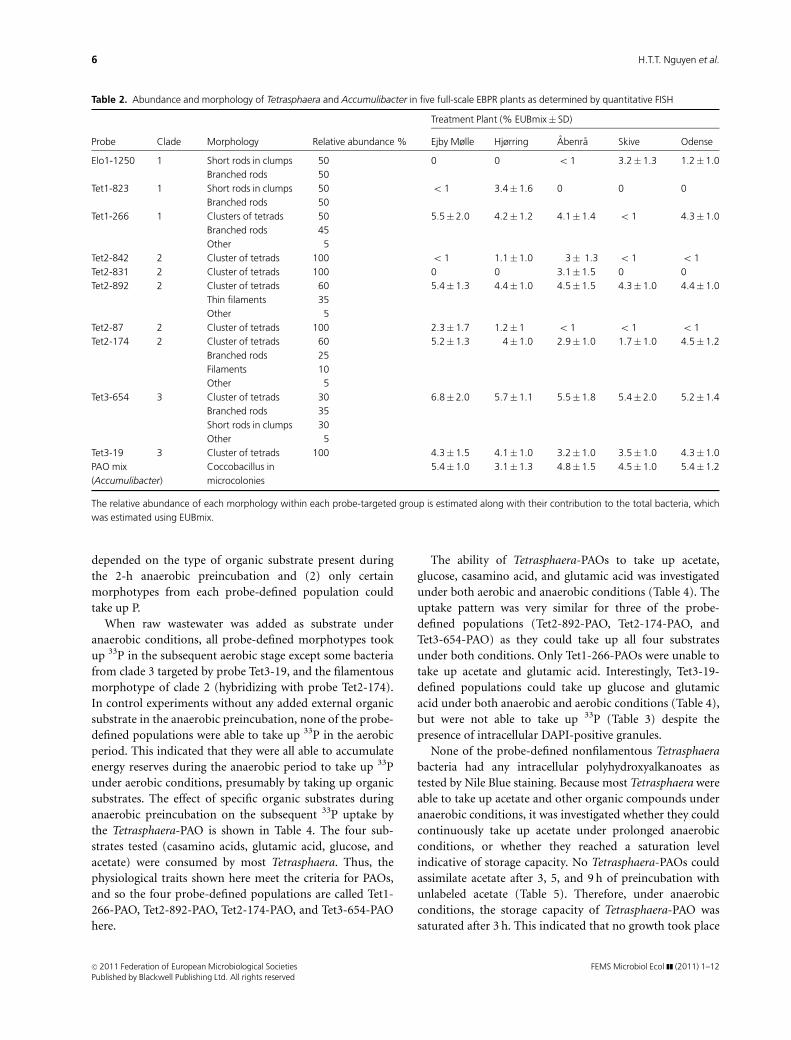
Table 2. Abundance and morphology of Tetrasphaera and Accumulibacter in five full-scale EBPR plants as determined by quantitative FISH
Treatment Plant (% EUBmix� SD)
Probe Clade Morphology Relative abundance % Ejby Mølle Hjørring Abenra Skive Odense
Elo1-1250 1 Short rods in clumps 50 0 0 o 1 3.2� 1.3 1.2�1.0
Branched rods 50
Tet1-823 1 Short rods in clumps 50 o 1 3.4�1.6 0 0 0
Branched rods 50
Tet1-266 1 Clusters of tetrads 50 5.5� 2.0 4.2�1.2 4.1� 1.4 o 1 4.3�1.0
Branched rods 45
Other 5
Tet2-842 2 Cluster of tetrads 100 o 1 1.1�1.0 3� 1.3 o 1 o 1
Tet2-831 2 Cluster of tetrads 100 0 0 3.1� 1.5 0 0
Tet2-892 2 Cluster of tetrads 60 5.4� 1.3 4.4�1.0 4.5� 1.5 4.3� 1.0 4.4�1.0
Thin filaments 35
Other 5
Tet2-87 2 Cluster of tetrads 100 2.3� 1.7 1.2�1 o 1 o 1 o 1
Tet2-174 2 Cluster of tetrads 60 5.2� 1.3 4�1.0 2.9� 1.0 1.7� 1.0 4.5�1.2
Branched rods 25
Filaments 10
Other 5
Tet3-654 3 Cluster of tetrads 30 6.8� 2.0 5.7�1.1 5.5� 1.8 5.4� 2.0 5.2�1.4
Branched rods 35
Short rods in clumps 30
Other 5
Tet3-19 3 Cluster of tetrads 100 4.3� 1.5 4.1�1.0 3.2� 1.0 3.5� 1.0 4.3�1.0
PAO mix
(Accumulibacter)
Coccobacillus in
microcolonies
5.4� 1.0 3.1�1.3 4.8� 1.5 4.5� 1.0 5.4�1.2
The relative abundance of each morphology within each probe-targeted group is estimated along with their contribution to the total bacteria, which
was estimated using EUBmix.
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
6 H.T.T. Nguyen et al.
under anaerobic conditions and that they only had a limited
uptake/storage capacity. However, all Tetrasphaera-PAOs
could take up glucose after 9 h of incubation with unlabeled
acetate, indicating that fermentation took place.
Abundance of Tetrasphaera in full-scaleEBPR plants
The distribution of Tetrasphaera was investigated in five full-
scale, well-functioning EBPR plants with nitrogen removal
(nitrification and denitrification). According to the micro-
autoradiography–FISH results, most of the probe-defined
Tetrasphaera were also PAOs. Only species from clade 3
targeted by probe Tet3-19 were either not active or not
PAOs. The probe-defined Tetrasphaera-PAOs were present
in high abundance (18–30% of all Bacteria targeted by
EUBmix) in all treatment plants investigated (Table 2). This
high abundance was supported by Gram staining of the
samples, showing that 20–30% of the biomasses were indeed
Gram positive. This was much higher than the abundance of
Accumulibacter (3.1–5.4%; Table 2). The abundance of each
probe-defined population varied in the different treatment
plants. Approximately half of the probe-defined populations
[targeted by the specific probes Tet1-266 (clade 1), Tet2-892,
and Tet2-174 (both clade 2), Tet3-19 and the broader Tet3-
654 (clade 3)] were present in all WWTP investigated with
relatively high abundances (4 5%). The others were domi-
nant only in one or a few plants (e.g. Tet1-823-PAOs in
Hjørring WWTP). When probes Actino-221 and Actino-
658 were applied, only approximately 50% of the Tetra-
sphaera were detected because these probes do not target
clade 1 and a large part of clade 2. The new probes cover
most Tetrasphaera in the five plants investigated, and nearly
all cells positive with broad actinobacterial probes (HGC69a
and HGC236) were targeted by one of the new probes. On
the other hand, the broad actinobacterial probes did not
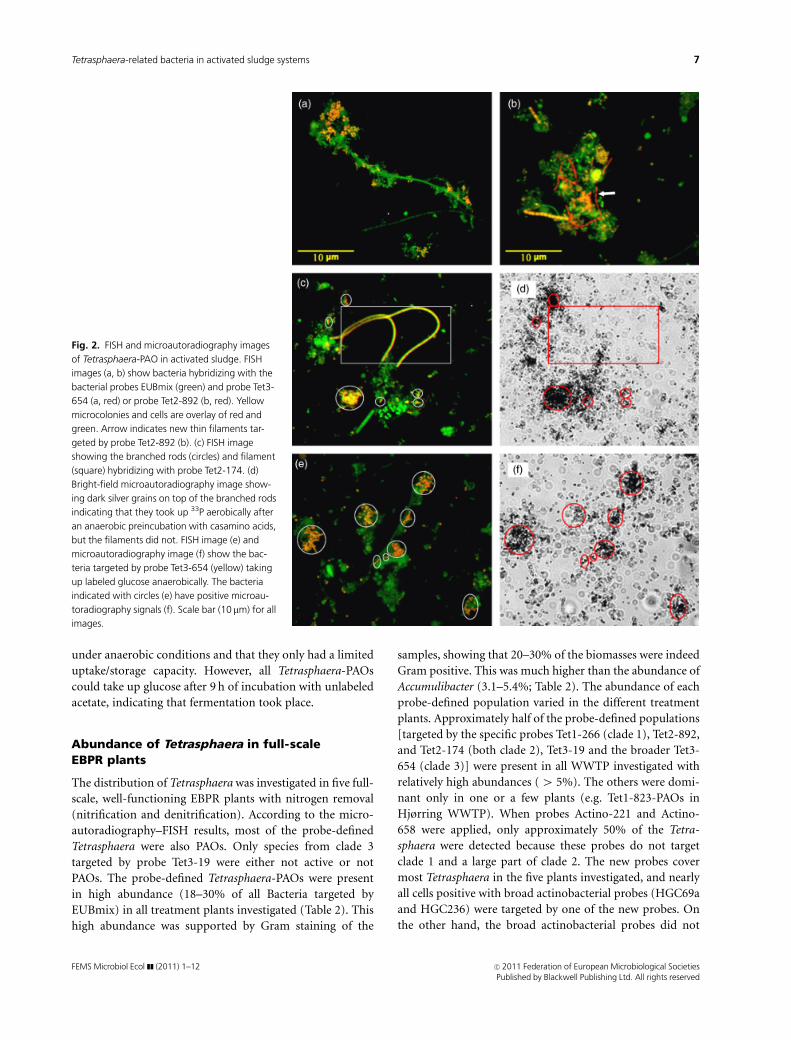
Fig. 2. FISH and microautoradiography images
of Tetrasphaera-PAO in activated sludge. FISH
images (a, b) show bacteria hybridizing with the
bacterial probes EUBmix (green) and probe Tet3-
654 (a, red) or probe Tet2-892 (b, red). Yellow
microcolonies and cells are overlay of red and
green. Arrow indicates new thin filaments tar-
geted by probe Tet2-892 (b). (c) FISH image
showing the branched rods (circles) and filament
(square) hybridizing with probe Tet2-174. (d)
Bright-field microautoradiography image show-
ing dark silver grains on top of the branched rods
indicating that they took up 33P aerobically after
an anaerobic preincubation with casamino acids,
but the filaments did not. FISH image (e) and
microautoradiography image (f) show the bac-
teria targeted by probe Tet3-654 (yellow) taking
up labeled glucose anaerobically. The bacteria
indicated with circles (e) have positive microau-
toradiography signals (f). Scale bar (10 mm) for all
images.
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
7Tetrasphaera-related bacteria in activated sludge systems
target all Tetrasphaera (e.g. the branched rods and the thin
filaments) targeted by the new probes and thus under-
estimated the total abundance of Tetrasphaera (and Actino-
bacteria). Interestingly, the thin filaments and the branched
rods were also not targeted by the EUBmix probes.
Discussion
No further studies of Tetrasphaera-related bacteria have
been made since they were first shown to have PAO behavior
and found in high numbers in full-scale EBPR systems
(Kong et al., 2005, 2008). In this study, we have conducted
a more detailed investigation and demonstrated a high
phylogenetic and physiological diversity. Furthermore, their
high abundance indicates that they play an important role in
full-scale EBPR plants, are perhaps more important than
Accumulibacter, and have a different metabolism.
Phylogeny and identification of Tetrasphaera byoligonucleotide probes
The phylogenetic analysis of clone sequences obtained from
full-scale EBPR plants showed the presence of three phylo-
genetically distinct clades in the genus Tetrasphaera in the
family Intrasporangiaceae. Two of these clades are repre-
sented by described Tetrasphaera isolates, although only
relatively distantly for most (96–98% identity), and they
are all showing a significantly higher diversity of Tetra-
sphaera-related bacteria in full-scale EBPR plants than
described previously (Kong et al., 2005).
Clade 1 (T. elongata-related sequences) was covered by
probes Elo1-1250, Tet1-823, and Tet1-266. Bacteria targeted
by probe Tet1-266 were most abundant. Interestingly, the
probe was designed to target only clone ASM31 (with two
mismatches to any other sequence), but it still showed two
phenotypes (branched rods and cocci in tetrads). Both
phenotypes were PAOs based on uptake of 33P so the probe
may target ecotypes with different morphologies or there is
still an undescribed diversity. This sequence is not targeted
by probe Actino-221 or Actino-658 designed by Kong et al.
(2005), and so these bacteria have not been detected
previously.
Clade 2 contains four isolated species: T. jenkinsii, T.
australiensis, T. veronensis, and the filamentous ‘Candidatus
Nostocoida limicola’. Most abundant were bacteria targeted
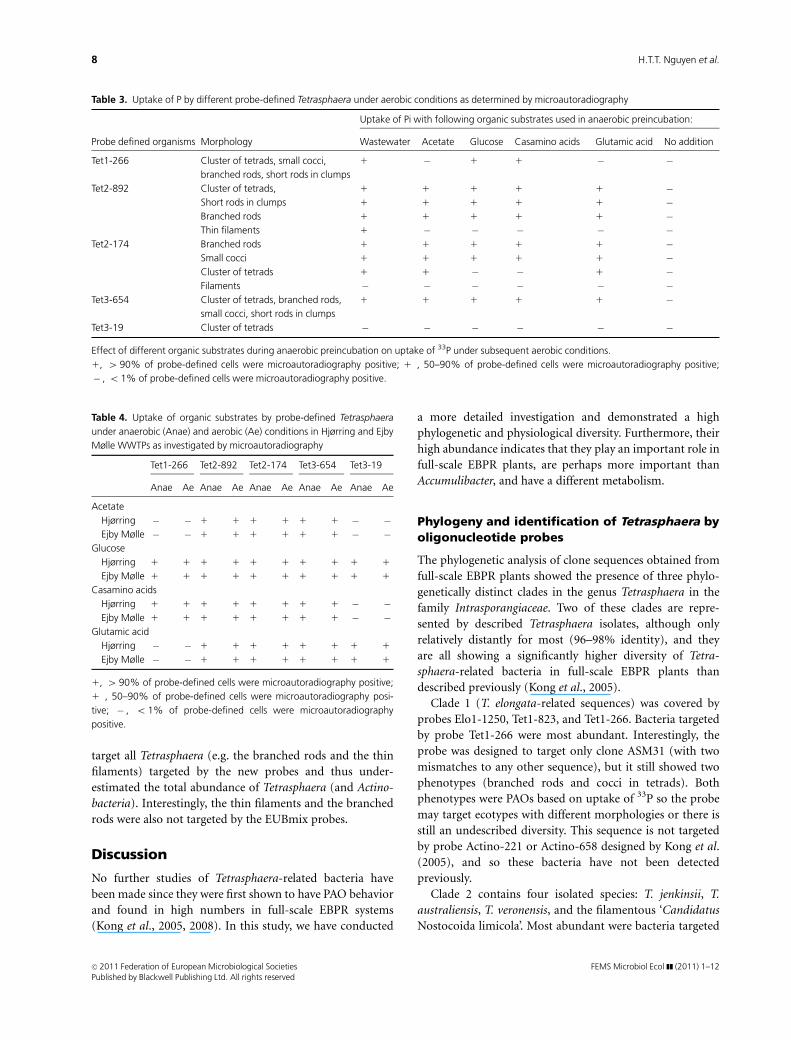
Table 3. Uptake of P by different probe-defined Tetrasphaera under aerobic conditions as determined by microautoradiography
Uptake of Pi with following organic substrates used in anaerobic preincubation:
Probe defined organisms Morphology Wastewater Acetate Glucose Casamino acids Glutamic acid No addition
Tet1-266 Cluster of tetrads, small cocci,
branched rods, short rods in clumps
1 � 1 1 � �
Tet2-892 Cluster of tetrads, 1 1 1 1 1 �Short rods in clumps 1 1 1 1 1 �Branched rods 1 1 1 1 1 �Thin filaments 1 � � � � �
Tet2-174 Branched rods 1 1 1 1 1 �Small cocci 1 1 1 1 1 �Cluster of tetrads 1 1 � � 1 �Filaments � � � � � �
Tet3-654 Cluster of tetrads, branched rods,
small cocci, short rods in clumps
1 1� 1 1 1 �
Tet3-19 Cluster of tetrads � � � � � �
Effect of different organic substrates during anaerobic preincubation on uptake of 33P under subsequent aerobic conditions.
1, 4 90% of probe-defined cells were microautoradiography positive; 1�, 50–90% of probe-defined cells were microautoradiography positive;
� , o 1% of probe-defined cells were microautoradiography positive.
Table 4. Uptake of organic substrates by probe-defined Tetrasphaera
under anaerobic (Anae) and aerobic (Ae) conditions in Hjørring and Ejby
Mølle WWTPs as investigated by microautoradiography
Tet1-266 Tet2-892 Tet2-174 Tet3-654 Tet3-19
Anae Ae Anae Ae Anae Ae Anae Ae Anae Ae
Acetate
Hjørring � � 1 1 1 1 1� 1� � �Ejby Mølle � � 1 1 1 1 1� 1� � �
Glucose
Hjørring 1 1 1 1 1 1 1 1 1 1
Ejby Mølle 1 1 1 1 1 1 1 1 1 1
Casamino acids
Hjørring 1 1 1 1 1 1 1 1 � �Ejby Mølle 1 1 1 1 1 1 1 1 � �
Glutamic acid
Hjørring � � 1 1 1 1 1 1 1 1
Ejby Mølle � � 1 1 1 1 1 1 1 1
1, 4 90% of probe-defined cells were microautoradiography positive;
1�, 50–90% of probe-defined cells were microautoradiography posi-
tive; � , o 1% of probe-defined cells were microautoradiography
positive.
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
8 H.T.T. Nguyen et al.
by Tet2-892 (targeting only one clone) and the probe
targeting the isolates (Tet2-174). Probe Actino-221 covers
largely the same as Tet2-87 and Tet2-892, but Tet2-174 does
not, and so these have been overlooked in previous studies.
Tet2-174 hybridized with several bacteria exhibiting mor-
photypes also described by the four isolates in clade 2.
Clade 3 contains only sequences from uncultured clones,
and probes Tet3-654 and Tet3-19 were designed to cover all
sequences as it was not possible to design a single probe
targeting all. Probe Tet3-19 (targeting clone ASM57) hybri-
dized only with cocci in tetrads and these are likely not PAOs
(Table 3). Probe Tet3-654 targets almost the same sequences
as Actino-658 but Tet3-654 covered more morphotypes than
Actino-658 and a higher number of Tetrasphaera. When the
two probes were used simultaneously with different fluor-
ochromes, only Tet3-654 targeted certain morphotypes,
such as the branched rods. These results show that Tet3-654
is more comprehensive and should be used to target Tetra-
sphaera-PAOs in clade 3.
At least six different morphotypes were found in the
Tetrasphaera clades 1–3 (short rods, branched rods, small
cocci, cocci in tetrads, filaments, and thin filaments). It
shows that the diversity is much greater than previously
known. The morphology of the rod-shaped cells seems very
similar to that of T. elongata, strains ASP12 (Onda & Takii,
2002) and LP12 (Hanada et al., 2002). The typical clusters of
tetrads are very similar to T. australiensis and T. japonica
(Maszenan et al., 2000). The filamentous morphology of
Tet2-174-targeted bacteria was similar in both size and
morphology to ‘Candidatus Nostocoida limicola’ targeted
by probe NLIMII175 (Liu & Seviour, 2001). However, the
thin filaments, branched rods, and small cocci have not been
described previously for Tetrasphaera. Interestingly, the thin
filaments observed in clade 2 (putative PAOs based on
carbon and 33P-uptake) did not hybridize with the EUBmix
or the broad actinobacterial probes (HGC69a and
HGC236), and so they have so far been overlooked. They
are usually hidden within the activated sludge floc, where
they can be quite abundant.
In order to detect the most important Tetrasphaera-PAOs
in full-scale EBPR plants, we propose to apply probe Tet1-
266 for clade 1, Tet2-892 and Tet2-174 for clade 2, and Tet3-
654 for clade 3.
Ecophysiology of Tetrasphaera
Most probe-defined Tetrasphaera were shown to be putative
PAOs as they contained polyphosphate and could actively
take up labeled P under aerobic conditions, provided they
had an organic substrate they could take up in the previous
anaerobic phase. Only bacteria targeted by probe Tet3-19 in
clade 3 and most filamentous morphotypes did not demon-
strate a PAO phenotype.
So far, most probe-defined Tetrasphaera in full-scale
EBPR plants can take up different amino acids and glucose
(Kong et al., 2005, 2008) under both anaerobic and aerobic
conditions, and this was confirmed in this study. This is very
similar to most isolated T. elongata strains in clade 1.
However, the ecophysiology of Tetrasphaera from clades 2
and 3 (probe Tet2-892, Tet2-174, and Tet3-654) seems more
diverse and more similar to that of T. australiensis and T.
japonica (Maszenan et al., 2000): all grow on complex media
and utilize all four substrates tested in this study (glucose,
acetate, casamino acids, and glutamic acid). The pattern of
acetate uptake of Tet3-654-defined PAOs was rather com-
plex. The microautoradiography-positive fraction varied
from plant to plant and even from year to year in the same
plant. Therefore, we have tested several old fixed microau-
toradiography samples from earlier studies with the same
varying results (data not shown). The varying uptake of
substrates may be explained by the broad coverage of probe
Tet3-654 allowing significant undetected temporal changes
of subpopulations within the targeted populations. This
complexity of Tet3-654-PAOs in acetate uptake can also
explain why bacteria targeted by probe Actino-658 (which
does not cover all Tet3-654-PAOs) were not observed to take
up short-chain fatty acids.
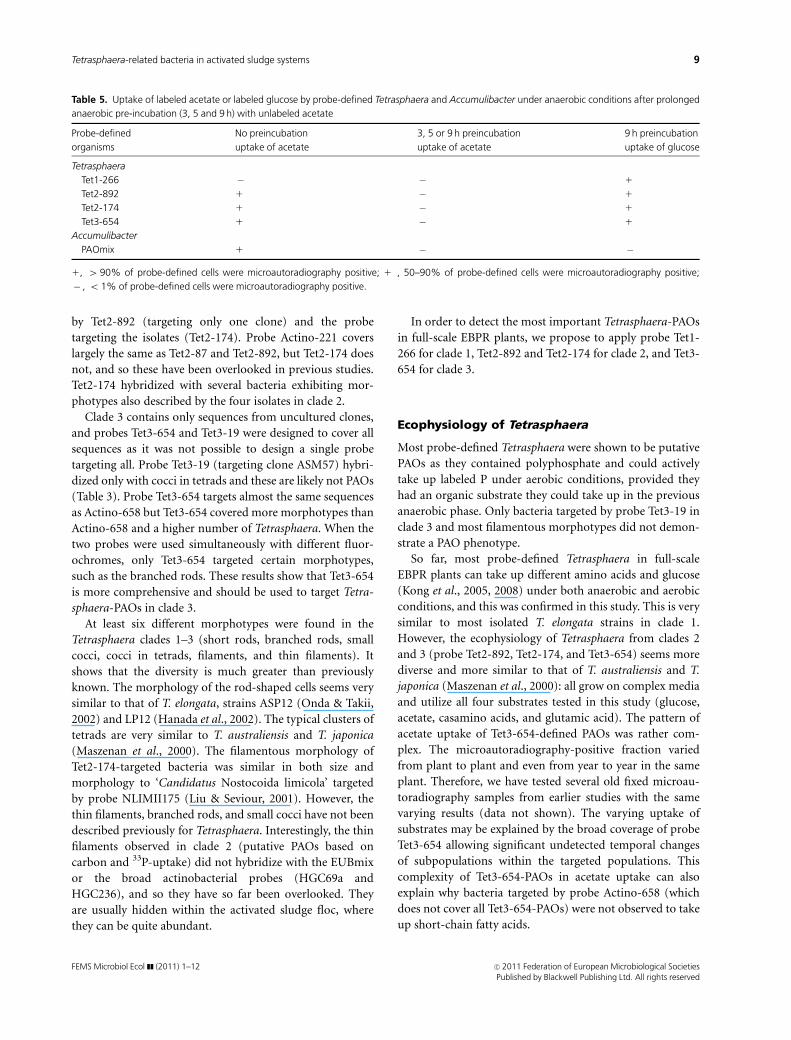
Table 5. Uptake of labeled acetate or labeled glucose by probe-defined Tetrasphaera and Accumulibacter under anaerobic conditions after prolonged
anaerobic pre-incubation (3, 5 and 9 h) with unlabeled acetate
Probe-defined
organisms
No preincubation
uptake of acetate
3, 5 or 9 h preincubation
uptake of acetate
9 h preincubation
uptake of glucose
Tetrasphaera
Tet1-266 � � 1
Tet2-892 1 � 1
Tet2-174 1 � 1
Tet3-654 1� � 1
Accumulibacter
PAOmix 1 � �
1, 4 90% of probe-defined cells were microautoradiography positive; 1�, 50–90% of probe-defined cells were microautoradiography positive;
� , o 1% of probe-defined cells were microautoradiography positive.
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
9Tetrasphaera-related bacteria in activated sludge systems

Also, as observed in the present study, all Tetrasphaera
were able to consume glucose anaerobically, but they did not
all take up P in the following aerobic period. In clade 2, the
filamentous morphotype (very similar to ‘Candidatus Nos-
tocoida limicola’) hybridizing with probe Tet2-174 did not
actively take up P aerobically and may not be active PAOs
despite the presence of intracellular P.
Physiology of Tetrasphaera-PAOs compared withEBPR biochemical models
The physiology of Tetrasphaera-PAOs is more complex and
in several aspects very different from Accumulibacter and the
general biochemical models used to explain the behavior of
PAOs (Garcia Martin et al., 2006; Oehmen et al., 2007; Zhou
et al., 2009).
Most surprising is the ability of Tetrasphaera-PAOs to
take up acetate and other substrates in the anaerobic period,
but their lack of ability to form polyhydroxyalkanoates as a
storage compound. Staining with Nile Blue or Sudan Black
did not detect polyhydroxyalkanoates, and so the presence
and identity of any intracellular compound is still unknown.
Interestingly, the anaerobic uptake capacity of acetate was
exhausted after 3 h, very similar to that of Accumulibacter
(Kong et al., 2005), strongly indicating the presence of an
unknown storage compound. Glucose, on the other hand,
was taken up after 9 h preincubation with acetate and
presumably fermented. Tetrasphaera can take up glucose
even after several days under anaerobic conditions in the
presence of glucose (Kong et al., 2008). In any case, the
anaerobic uptake of substrate, either as acetate, amino acids,
or glucose, was decisive for the subsequent uptake of P in the
aerobic phase. However, for the thin filamentous bacteria
hybridizing with Tet2-892, only wastewater could provide
the right, so far unknown, organic substance during the
anaerobic period.
Role of Tetrasphaera-PAO in full-scaleEBPR systems
Four probe-defined groups in the three clades of Tetra-
sphaera-PAO constituted 18–30% of the total bacterial
biomass present in five well-working EBPR plants with
nitrogen removal. A high number of Tetrasphaera has also
been reported in German EBPR plants (Eschenhagen et al.,
2003), and many Gram-positive bacteria (up to 35% of the
biomass) have been reported in Australian EBPR plants
(Beer et al., 2006), although their identities have not been
investigated in detail. Their abundance is often substantially
greater than that of Accumulibacter, indicating that Tetra-
sphaera-PAO may play an important role in full-scale EBPR
plants.
Tetrasphaera-PAOs seem to occupy a slightly different
ecological niche than Accumulibacter. They are more diverse
in substrate uptake, can take up glucose (but not mannose
or galactose, Kong et al., 2008), and probably also ferment
under anaerobic conditions. Whether they can grow under
anaerobic conditions is, however, uncertain (Kong et al.,
2008). As they excrete extracellular amylases (Xia et al.,
2008), they appear to be specialized degraders and consu-
mers of starch and other polysaccharides in the wastewater.
A certain functional redundancy among the bacteria in
WWTPs is seen for most important processes, for example
among denitrifiers (Thomsen et al., 2007; Hesselsoe et al.,
2009), ensuring high resilience and a stable function. The
same seems to be the case for PAOs with several ecotypes of
Accumulibacter (He et al., 2007), and, as shown in this study,
considerable diversity of Tetrasphaera. The most important
factors determining the presence of either Tetrasphaera-
PAOs or Accumulibacter can be the availability of glucose
and amino acids as well as the anaerobic residence time in
the anaerobic reactor, where the ability to ferment will be
beneficial the longer the residence time.
Acknowledgements
This study was funded by Viborg Energy, Danish Council for
Independent Research, and Aalborg University. We thank
Prof. Robert Seviour for providing Tetrasphaera cultures
and Marianne Stevenson and Artur Tomasz Mielczarek for
technical help.
References
Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R &
Stahl DA (1990) Combination of 16S rRNA-targeted
oligonucleotide probes with flow cytometry for analyzing
mixed microbial populations. Appl Environ Microb 56:
1919–1925.
Ashelford KE, Chuzhanova NA, Fry JC, Jones AJ & Weightman AJ
(2006) New screening software shows that most recent large
16S rRNA gene clone libraries contain chimeras. Appl Environ
Microb 72: 5734–5741.
Beer M, Stratton HM, Griffiths PC & Seviour RJ (2006) Which
are the polyphosphate accumulating organisms in full-scale
activated sludge enhanced biological phosphate removal
systems in Australia? J Appl Microbiol 100: 233–243.
Collins TJ (2007) ImageJ for microscopy. Biotechniques 43:
25–30.
Comeau Y, Hall KJ, Hancock REW & Oldham WK (1986)
Biochemical model for enhanced biological phosphorus
removal. Water Res 20: 1511–1521.
Crocetti GR, Hugenholtz P, Bond PL, Schuler A, Keller J, Jenkins
D & Blackall LL (2000) Identification of polyphosphate-
accumulating organisms and design of 16S rRNA-directed
probes for their detection and quantitation. Appl Environ
Microb 66: 1175–1182.
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
10 H.T.T. Nguyen et al.

Daims H, Bruhl A, Amann R, Schleifer K-H & Wagner M (1999)
The domain-specific probe EUB338 is insufficient for the
detection of all Bacteria: development and evaluation of a
more comprehensive probe set. Syst Appl Microbiol 22:
434–444.
Erhart R, Bradford D, Seviour RJ, Amann R & Blackall LL (1997)
Development and use of fluorescent in situ hybridization
probes for the detection and identification of Microthrix
parvicella in activated sludge. Syst Appl Microbiol 20: 310–318.
Eschenhagen M, Schuppler M & Roske I (2003) Molecular
characterization of the microbial community structure in two
activated sludge systems for the advanced treatment of
domestic effluents. Water Res 37: 3224–3232.
Garcia Martin H, Ivanova N, Kunin V et al. (2006) Metagenomic
analysis of two enhanced biological phosphorus removal
(EBPR) sludge communities. Nat Biotechnol 24: 1263–1269.
Hanada S, Liu WT, Shintani T, Kamagata Y & Nakamura K
(2002) Tetrasphaera elongata sp. nov., a polyphosphate-
accumulating bacterium isolated from activated sludge. Int J
Syst Evol Micr 52: 883–887.
He S, Gall DL & McMahon KD (2007) ‘Candidatus
Accumulibacter’ population structure in enhanced biological
phosphorus removal sludges as revealed by polyphosphate
kinase genes. Appl Environ Microb 73: 5865–5874.
Hesselmann RP, Werlen C, Hahn D, van der Meer JR & Zehnder
AJ (1999) Enrichment, phylogenetic analysis and detection of
a bacterium that performs enhanced biological phosphate
removal in activated sludge. Syst Appl Microbiol 22: 454–465.
Hesselsoe M, Fureder S, Schloter M, Bodrossy L, Iversen N,
Roslev P, Nielsen PH, Wagner M & Loy A (2009) Isotope array
analysis of Rhodocyclales uncovers functional redundancy and
versatility in an activated sludge. ISME J 3: 1349–1364.
Kawaharasaki M, Tanaka H, Kanagawa T & Nakamura K (1999)
In situ identification of polyphosphate-accumulating bacteria
in activated sludge by dual staining with rRNA-targeted
oligonucleotide probes and 40, 60-diamidino-2-phenylindole
(DAPI) at a polyphosphate-probing concentration. Water Res
33: 257–265.
Kong Y, Nielsen JL & Nielsen PH (2004) Microautoradiographic
study of Rhodocyclus-related polyphosphate-accumulating
bacteria in full-scale enhanced biological phosphorus removal
plants. Appl Environ Microbiol 70: 5383–5390.
Kong Y, Nielsen JL & Nielsen PH (2005) Identity and
ecophysiology of uncultured actinobacterial polyphosphate-
accumulating organisms in full-scale enhanced biological
phosphorus removal plants. Appl Environ Microb 71:
4076–4085.
Kong Y, Xia Y & Nielsen PH (2008) Activity and identity of
fermenting microorganisms in full-scale biological nutrient
removing wastewater treatment plants. Environ Microbiol 10:
2008–2019.
Kragelund C, Nielsen JL, Thomsen TR & Nielsen PH (2005)
Ecophysiology of the filamentous Alphaproteobacterium
Meganema perideroedes in activated sludge. FEMS Microbiol
Ecol 54: 111–122.
Lane DJ (1991) 16S/23S rRNA sequencing. Nucleic Acid
Techniques in Bacterial Systematics (Stackebrandt E &
Goodfellow M, eds), pp. 113–175. Wiley Publications,
Chichester, UK.
Lemos PC, Serafim LS, Santos MM, Reis MA & Santos H (2003)
Metabolic pathway for propionate utilization by phosphorus-
accumulating organisms in activated sludge: 13C labeling and
in vivo nuclear magnetic resonance. Appl Environ Microb 69:
241–251.
Liu JR & Seviour RJ (2001) Design and application of
oligonucleotide probes for fluorescent in situ identification of
the filamentous bacterial morphotype Nostocoida limicola in
activated sludge. Environ Microbiol 3: 551–560.
Liu WT, Nielsen AT, Wu JH, Tsai CS, Matsuo Y & Molin S (2001)
In situ identification of polyphosphate- and
polyhydroxyalkanoate-accumulating traits for microbial
populations in a biological phosphorus removal process.
Environ Microbiol 3: 110–122.
Louie TM, Mah TJ, Oldham WK & Ramey WD (2000) Use of
metabolic inhibitors and gas chromatography/mass
spectrometry to study poly-b-hydroxyalkanoates metabolism
involving cryptic nutrients in enhanced biological phosphorus
removal systems. Water Res 34: 1507–1514.
Loy A, Horn M & Wagner M (2003) probeBase: an online
resource for rRNA-targeted oligonucleotide probes. Nucleic
Acids Res 31: 514–516.
Ludwig W, Strunk O, Westram R et al. (2004) ARB: a software
environment for sequence data. Nucleic Acids Res 32:
1363–1371.
Maidak BL, Cole JR, Lilburn TG et al. (2001) The RDP-II
(Ribosomal Database Project). Nucleic Acids Res 29: 173–174.
Maszenan A, Seviour R, Patel B, Schumann P, Burghardt J,
Tokiwa Y & Stratton H (2000) Three isolates of novel
polyphosphate-accumulating Gram-positive cocci, obtained
from activated sludge, belong to a new genus, Tetrasphaera
gen. nov., and description of two new species, Tetrasphaera
japonica sp. nov. and Tetrasphaera australiensis sp. nov. Int J
Syst Evol Micr 50: 593–603.
McKenzie CM, Seviour EM, Schumann P, Maszenan AM, Liu JR,
Webb RI, Monis P, Saint CP, Steiner U & Seviour RJ (2006)
Isolates of ‘Candidatus Nostocoida limicola’ Blackall et al. 2000
should be described as three novel species of the genus
Tetrasphaera, as Tetrasphaera jenkinsii sp. nov., Tetrasphaera
vanveenii sp. nov. and Tetrasphaera veronensis sp. nov. Int J Syst
Evol Micr 56: 2279–2290.
Mino T, van Loosdrecht MCM & Heijnen HJ (1998)
Microbiology and biochemistry of the enhanced biological
phosphate removal process. Water Res 32: 3193–3207.
Nielsen JL (2009) Protocol for Fluorescence in situ hybridization
(FISH) with rRNA-targeted oligonucleotides. FISH Handbook
of Wastewater Treatment (Nielsen PH, Lemmer H & Daims H,
eds), pp. 73–84. IWA publishing, London, UK.
Nielsen JL & Nielsen PH (2005) Advances in microscopy:
microautoradiography of single cells. Method Enzymol 397:
237–256.
FEMS Microbiol Ecol ]] (2011) 1–12 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
11Tetrasphaera-related bacteria in activated sludge systems

Nielsen PH, Mielczarek AT, Kragelund C, Nielsen JL, Saunders
AM, Kong Y, Hansen AA & Vollertsen J (2010) A conceptual
ecosystem model of microbial communities in enhanced
biological phosphorus removal plants. Water Res 44:
5070–5088.
Oehmen A, Lemos PC, Carvalho G, Yuan Z, Keller J, Blackall LL
& Reis MA (2007) Advances in enhanced biological
phosphorus removal: from micro to macro scale. Water Res 41:
2271–2300.
Onda S & Takii S (2002) Isolation and characterization of a
Gram-positive polyphosphate-accumulating bacterium. J Gen
Appl Microbiol 48: 125–133.
Pereira H, Lemos PC, Reis MAM, Crespo JPSG, Carrondo MJT &
Santos H (1996) Model for carbon metabolism in biological
phosphorus removal processes based on in vivo 13C-NMR
labeling experiments. Water Res 30: 2128–2138.
Roller C, Wagner M, Amann R, Ludwig W & Schleifer K-H
(1994) In situ probing of Gram-positive bacteria with high
DNA G1C content using 23S rRNA- targeted
oligonucleotides. Microbiology 140: 2849–2858.
Seviour RJ & Blackall LL (1999) The Microbiology of Activated
Sludge. Kluwer Academic Publishers, Dordrecht.
Seviour RJ, Mino T & Onuki M (2003) The microbiology of
biological phosphorus removal in activated sludge systems.
FEMS Microbiol Rev 27: 99–127.
Thomsen TR, Kong Y & Nielsen PH (2007) Ecophysiology of
abundant denitrifying bacteria in activated sludge. FEMS
Microbiol Ecol 60: 370–382.
van Loosdrecht MCM, Smolders GJ, Kuba T & Heijnen JJ (1997)
Metabolism of micro-organisms responsible for enhanced
biological phosphorus removal from wastewater, Use of
dynamic enrichment cultures. Antonie van Leeuwenhoek 71:
109–116.
Wentzel MC, Lotter LH, Loewenthal RE & Marais GR (1986)
Metabolic behaviour of Acinetobacter spp. in enhanced
biological phosphorus removal - a biochemical model. Water
SA 12: 209–224.
Wexler M, Richardson DJ & Bond PL (2009) Radiolabelled
proteomics to determine differential functioning of
Accumulibacter during the anaerobic and aerobic phases of a
bioreactor operating for enhanced biological phosphorus
removal. Environ Microbiol 11: 3029–3044.
Xia Y, Kong Y, Thomsen TR & Nielsen PH (2008) Identification
and ecophysiological characterization of epiphytic
protein-hydrolyzing Saprospiraceae (‘Candidatus Epiflobacter’
spp.) in activated sludge. Appl Environ Microb 74:
2229–2238.
Zhou Y, Pijuan M, Zeng RJ & Yuan Z (2009) Involvement
of the TCA cycle in the anaerobic metabolism of
polyphosphate-accumulating organisms (PAOs). Water Res
43: 1330–1340.
Zilles JL, Hung CH & Noguera DR (2002) Presence of
Rhodocyclus in a full-scale wastewater treatment plant and
their participation in enhanced biological phosphorus
removal. Water Sci Technol 46: 123–128.
FEMS Microbiol Ecol ]] (2011) 1–12c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
12 H.T.T. Nguyen et al.
Related Documents











