Vol. 6, 597-601, August 1997 Cancer Epidem1ologj, Biomarkers & Prevention 597 phy. High Dietary w-6 Polyunsaturated Fatty Acids Drastically Increase the Formation of Etheno-DNA Base Adducts in White Blood Cells of Female Subjects’ Jagadeesan Nair, Carlos E. Vaca, Ivana Velic, Marja Mutanen, Lilsa M. Valsta, and Helmut Bartsch2 Division of Toxicology and Cancer Risk Factors, Deutsches Krebsforschungszentrum (DKFZ), Im Neuenheimer Feld 280, D-69l20 Heidelberg, Germany [J. N., I. V., H. B.]; Molecular Epidemiology Unit, Center for Nutrition and Toxicology, Department of Biosciences at Novum, Karolinska Institute, 5-1 1457 Huddinge, Sweden [C. E. V.]; Department of Applied Chemistry and Microbiology (Nutrition), University of Helsinki, SF-00710 Helsinki, Finland [M. M.]; and Department of Nutrition, National Public Health Institute, SF-00300 Helsinki, Finland [L. M. V.] Abstract Lipid peroxidation generates reactive aldehydes such as trans-4-hydroxy-2-nonenal and malonaldehyde, which form promutagemc exocyclic DNA adducts in human cells and may contribute to diet-related cancers. Using ultrasensitive detection methods, analysis of WBC DNA from volunteers in a dietary study revealed that high intake of w-6 polyunsaturated fatty acids increased malonaldehyde-derived adducts in male and female subjects. In contrast, etheno adducts (1,M-ethenodeoxyadenosine; 3,N’-ethenodeoxycytidine) were not elevated in males but were, on average, 44) times higher in females, displaying a huge intersubject variation in lipid peroxidation-denved DNA damage. Exocycic DNA adducts are promising biomarkers for examining the hypothesis of possible links between increased intake of dietary w-6 polyunsaturated fatty acids, DNA damage, and elevated cancer risk for breast, colon, and prostate. Introduction At least one-third of all human cancers may be associated with diet (1); dietary fat is thought to be one of the main risk factors because positive correlations between dietary fat intake and increased risk of breast, colon, and prostate cancer have been reported (2-4). Although the role of individual fatty acids in human cancer risk is still poorly understood, epidemiological data have linked a high dietary w-6 PUFA3 intake (linoleic acid Received 1/8/97; revised 4/1/97; accepted 5/2/97. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. I This study was partly supported by a contract with the European Union (EV5V- CT94-0409) and was presented in part at an intemational meeting entitled “DNA Adducts and Mutations in Human Biomomtonng,” held in Stockholm, Sweden, June 9-13, 1996. 2 To whom requests for reprints should be addressed. 3 The abbreviations used are: PUFA, polyunsaturated fatty acid; HNE, trans-4- hydroxy-2-nonenal: MUFA, monounsaturated fatty acid; adA, l,M-ethenode- oxyadenosine; #{128}dC, 3,W-ethenodeoxycytidine; FA, fatty acid; MA-dG, malon- Cl8:2), especially in connection with a low w-3 PUFA (doco- sahexaenoic acid C22:6) consumption, to an increased risk of breast and colon cancer (5, 6). This can be explained, on one hand, by the findings that w-6 PUFAs enhance mammary tumorigenesis and tumor cell proliferation in experimentally induced mammary tumors, whereas w-3 PUFAs can act as growth inhibitors of initiated cancer cells (5, 6). On the other hand, oxidative stress and enhanced peroxidation of PUFAs in cell membranes, often involving hydroxyl radicals, are widely hypothesized to stimulate the development of human malignant diseases (7), raising the question of whether the carcinogenic process could be initiated and/or accelerated through tu-6 PUFA lipid peroxidation-derived DNA damage (8, 9). Recent studies reported the presence of such DNA-damaging adducts in hu- man tissues (10-13). As cell replication before repair of the adducts leads to mutations, DNA adduction is commonly re- garded as the initiating step in carcinogenesis. HNE, one of the major products of lipid peroxidation (14) formed after oxidation of linoleic acid and arachidonic acid (15), is readily oxidized by fatty acid hydroperoxides (16) to form 2,3-epoxy-4-hydroxynonanal, a bifunctional alkylating agent. It reacts with DNA bases to yield etheno adducts, such as l,N2-ethenodeoxyguanosine (17). Likewise, oxidation of rat liver microsomal membranes or of arachidonic acid with cu- mene hydroperoxide or Fe2 yielded edA and #{128}dC in the presence of the respective deoxynucleotides (Ref. 1 8; see scheme for formation of etheno adducts from PUFA in Fig. 1). The relevance of the three etheno adducts in carcinogenesis is supported by the following: they are promutagenic DNA le- sions (19); there is strong evidence that they initiate carcino- genesis by vinyl chloride and urethane (19); edA and #{128}dC are elevated in the liver of Long-Evans-Cinnamon rats (animal model for Wilsons’s disease), a rat strain that develops spon- taneous hepatitis and later hepatocellular carcinoma due to aberrant copper accumulation (20); and much higher hepatic levels of etheno adducts were detected in patients with metal storage diseases at elevated risk for liver cancer, e.g. , Wilson’s disease and primary hemochromatosis, as compared to liver DNA from healthy individuals (21). To investigate the impact of a diet high in w-6 PUFAs or MUFAs on DNA damage, samples from a carefully controlled dietary study (22) were analyzed for promutagenic exocyclic DNA adducts, edA, and #{128}dC. aldehyde-deoxyguanosine adduct; ThARS, thiobarbutaric acid reactive substances; 1’WBC, total WBC; HPLC, high-performance liquid chromatogra- on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 6, 597-601, August 1997 Cancer Epidem1ologj�, Biomarkers & Prevention 597
phy.
High Dietary w-6 Polyunsaturated Fatty Acids Drastically Increase
the Formation of Etheno-DNA Base Adducts in White Blood
Cells of Female Subjects’
Jagadeesan Nair, Carlos E. Vaca, Ivana Velic,Marja Mutanen, Lilsa M. Valsta, and Helmut Bartsch2
Division of Toxicology and Cancer Risk Factors, Deutsches
Krebsforschungszentrum (DKFZ), Im Neuenheimer Feld 280, D-69l20
Heidelberg, Germany [J. N., I. V., H. B.]; Molecular Epidemiology Unit,
Center for Nutrition and Toxicology, Department of Biosciences at Novum,
Karolinska Institute, 5-1 1457 Huddinge, Sweden [C. E. V.]; Department of
Applied Chemistry and Microbiology (Nutrition), University of Helsinki,
SF-00710 Helsinki, Finland [M. M.]; and Department of Nutrition, National
Public Health Institute, SF-00300 Helsinki, Finland [L. M. V.]
Abstract
Lipid peroxidation generates reactive aldehydes such astrans-4-hydroxy-2-nonenal and malonaldehyde, whichform promutagemc exocyclic DNA adducts in humancells and may contribute to diet-related cancers. Usingultrasensitive detection methods, analysis of WBC
DNA from volunteers in a dietary study revealedthat high intake of w-6 polyunsaturated fatty acidsincreased malonaldehyde-derived adducts in male and
female subjects. In contrast, etheno adducts(1,M�-ethenodeoxyadenosine; 3,N’�-ethenodeoxycytidine)were not elevated in males but were, on average, 44) timeshigher in females, displaying a huge intersubjectvariation in lipid peroxidation-denved DNA damage.Exocycic DNA adducts are promising biomarkers for
examining the hypothesis of possible links betweenincreased intake of dietary w-6 polyunsaturated fattyacids, DNA damage, and elevated cancer risk for breast,colon, and prostate.
Introduction
At least one-third of all human cancers may be associated with
diet (1); dietary fat is thought to be one of the main risk factorsbecause positive correlations between dietary fat intake andincreased risk of breast, colon, and prostate cancer have beenreported (2-4). Although the role of individual fatty acids in
human cancer risk is still poorly understood, epidemiological
data have linked a high dietary w-6 PUFA3 intake (linoleic acid
Received 1/8/97; revised 4/1/97; accepted 5/2/97.The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked advertisement in
accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
I This study was partly supported by a contract with the European Union (EV5V-
CT94-0409) and was presented in part at an intemational meeting entitled “DNA
Adducts and Mutations in Human Biomomtonng,” held in Stockholm, Sweden,
June 9-13, 1996.
2 To whom requests for reprints should be addressed.
3 The abbreviations used are: PUFA, polyunsaturated fatty acid; HNE, trans-4-
hydroxy-2-nonenal: MUFA, monounsaturated fatty acid; adA, l,M�-ethenode-
oxyadenosine; #{128}dC,3,W�-ethenodeoxycytidine; FA, fatty acid; MA-dG, malon-
Cl8:2), especially in connection with a low w-3 PUFA (doco-
sahexaenoic acid C22:6) consumption, to an increased risk of
breast and colon cancer (5, 6). This can be explained, on one
hand, by the findings that w-6 PUFAs enhance mammary
tumorigenesis and tumor cell proliferation in experimentally
induced mammary tumors, whereas w-3 PUFAs can act as
growth inhibitors of initiated cancer cells (5, 6). On the other
hand, oxidative stress and enhanced peroxidation of PUFAs in
cell membranes, often involving hydroxyl radicals, are widely
hypothesized to stimulate the development of human malignant
diseases (7), raising the question of whether the carcinogenic
process could be initiated and/or accelerated through tu-6 PUFA
lipid peroxidation-derived DNA damage (8, 9). Recent studies
reported the presence of such DNA-damaging adducts in hu-
man tissues (10-13). As cell replication before repair of the
adducts leads to mutations, DNA adduction is commonly re-
garded as the initiating step in carcinogenesis.
HNE, one of the major products of lipid peroxidation (14)
formed after oxidation of linoleic acid and arachidonic acid
(15), is readily oxidized by fatty acid hydroperoxides (16) to
form 2,3-epoxy-4-hydroxynonanal, a bifunctional alkylating
agent. It reacts with DNA bases to yield etheno adducts, such
as l,N2-ethenodeoxyguanosine (17). Likewise, oxidation of rat
liver microsomal membranes or of arachidonic acid with cu-
mene hydroperoxide or Fe2� yielded edA and #{128}dCin the
presence of the respective deoxynucleotides (Ref. 1 8; see
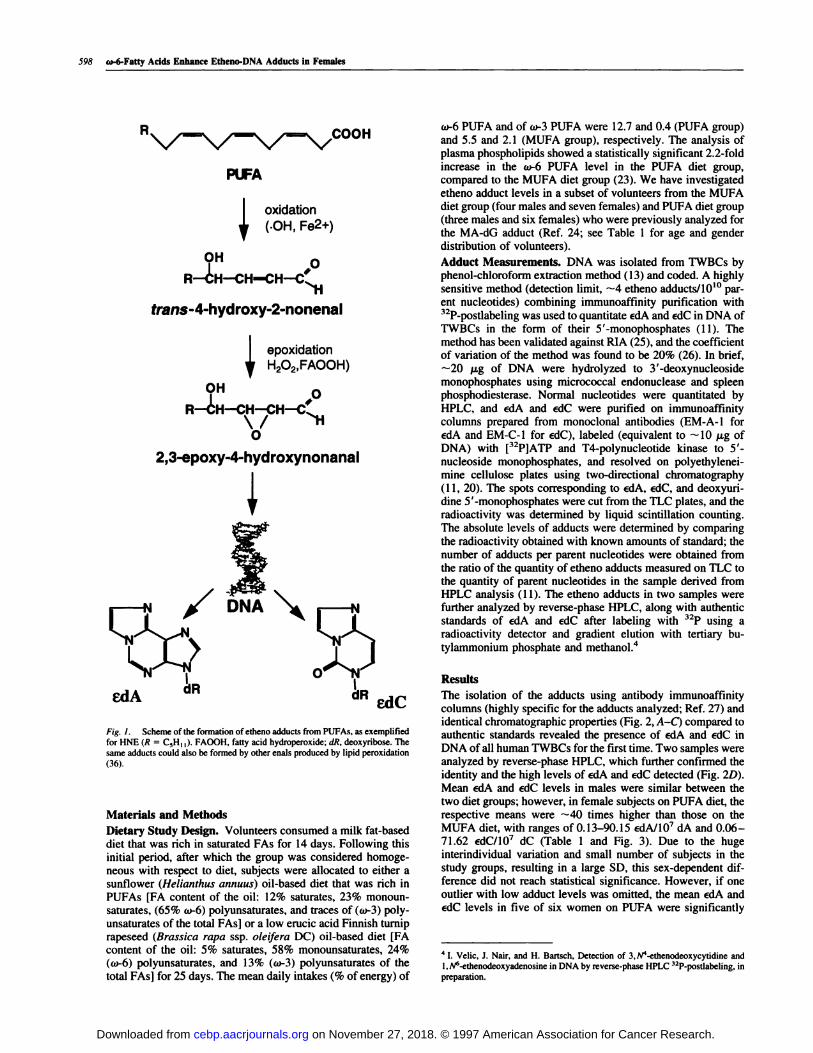
scheme for formation of etheno adducts from PUFA in Fig. 1).
The relevance of the three etheno adducts in carcinogenesis is
supported by the following: they are promutagenic DNA le-
sions (19); there is strong evidence that they initiate carcino-
genesis by vinyl chloride and urethane (19); edA and #{128}dCare
elevated in the liver of Long-Evans-Cinnamon rats (animal
model for Wilsons’s disease), a rat strain that develops spon-
taneous hepatitis and later hepatocellular carcinoma due to
aberrant copper accumulation (20); and much higher hepatic
levels of etheno adducts were detected in patients with metal
storage diseases at elevated risk for liver cancer, e.g. , Wilson’s
disease and primary hemochromatosis, as compared to liver
DNA from healthy individuals (21). To investigate the impact
of a diet high in w-6 PUFAs or MUFAs on DNA damage,
samples from a carefully controlled dietary study (22) were
analyzed for promutagenic exocyclic DNA adducts, edA, and
#{128}dC.
aldehyde-deoxyguanosine adduct; ThARS, thiobarbutaric acid reactivesubstances; 1’WBC, total WBC; HPLC, high-performance liquid chromatogra-
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
PlEA
OH 0
R4H-CH-CH--C’
\f0
2,3-epoxy-4-hydroxynonanal
I
SDNA/
EdA
598 w-6.Fatty Acids Enhance Etheno-DNA Adducts in Females
I oxidation
+ (.OH, Fe2+)
R4H_CH_CH_-C#{231}
trans-4-hydroxy-2-nonenal
I epoxidation
* H202,FAOOH)
#{231}i��JR�JC
Fig. 1. Scheme of the formation of etheno adducts from PUFAs, as exemplified
for HNE (R = C,H1 ). FAOOH, fatty acid hydroperoxide; dR, deoxyribose. Thesame adducts could also be formed by other enals produced by lipid peroxidation
(36).
Materials and Methods
Dietary Study Design. Volunteers consumed a milk fat-baseddiet that was rich in saturated FAs for 14 days. Following this
initial period, after which the group was considered homoge-neous with respect to diet, subjects were allocated to either asunflower (Helianthus annuus) oil-based diet that was rich inPUFAs [FA content of the oil: 12% saturates, 23% monoun-
saturates, (65% w-6) polyunsaturates, and traces of (w-3) poly-unsaturates of the total FAs] or a low erucic acid Finnish turniprapeseed (Brassica rapa ssp. oleifera DC) oil-based diet [FAcontent of the oil: 5% saturates, 58% monounsaturates, 24%(w-6) polyunsaturates, and 13% (w-3) polyunsaturates of thetotal FAs] for 25 days. The mean daily intakes (% of energy) of
w-6 PUFA and of w-3 PUFA were 12.7 and 0.4 (PUFA group)
and 5.5 and 2.1 (MUFA group), respectively. The analysis ofplasma phospholipids showed a statistically significant 2.2-fold
increase in the w-6 PUFA level in the PUFA diet group,compared to the MUFA diet group (23). We have investigatedetheno adduct levels in a subset of volunteers from the MUFA
diet group (four males and seven females) and PUFA diet group(three males and six females) who were previously analyzed forthe MA-dG adduct (Ref. 24; see Table 1 for age and gender
distribution of volunteers).
Adduct Measurements. DNA was isolated from TWBCs byphenol-chloroform extraction method (13) and coded. A highly
sensitive method (detection limit, -4 etheno adducts/101#{176} par-ent nucleotides) combining immunoaffinity purification with
32P-postlabeling was used to quantitate edA and #{128}dCin DNA ofTWBCs in the form of their 5’-monophosphates (11). The
method has been validated against lilA (25), and the coefficientof variation of the method was found to be 20% (26). In brief,-20 ,.�g of DNA were hydrolyzed to 3’-deoxynucleosidemonophosphates using micrococcal endonuclease and spleenphosphodiesterase. Normal nucleotides were quantitated byHPLC, and edA and #{128}dCwere purified on immunoaffinity
columns prepared from monoclonal antibodies (EM-A-i foredA and EM-C-i for #{128}dC),labeled (equivalent to -10 �g of
DNA) with [32P]ATP and T4-polynucleotide kinase to 5’-nucleoside monophosphates, and resolved on polyethylenei-mine cellulose plates using two-directional chromatography
(11, 20). The spots corresponding to edA, #{128}dC,and deoxyuri-dine 5’-monophosphates were cut from the TLC plates, and theradioactivity was determined by liquid scintillation counting.The absolute levels of adducts were determined by comparing
the radioactivity obtained with known amounts of standard; thenumber of adducts per parent nucleotides were obtained from
the ratio of the quantity of etheno adducts measured on TLC tothe quantity of parent nucleotides in the sample derived fromHPLC analysis (1 1). The etheno adducts in two samples were
further analyzed by reverse-phase HPLC, along with authenticstandards of edA and #{128}dCafter labeling with 32P using aradioactivity detector and gradient elution with tertiary bu-tylammonium phosphate and methanol.4
Results
The isolation of the adducts using antibody immunoaffinitycolumns (highly specific for the adducts analyzed; Ref. 27) andidentical chromatographic properties (Fig. 2, A-C) compared to
authentic standards revealed the presence of edA and #{128}dCinDNA of all human TWBCs for the first time. Two samples wereanalyzed by reverse-phase HPLC, which further confirmed theidentity and the high levels of edA and edC detected (Fig. 2D).
Mean edA and #{128}dClevels in males were similar between the
two diet groups; however, in female subjects on PUFA diet, therespective means were -40 times higher than those on theMUFA diet, with ranges of 0.13-90.15 #{128}dA/l07dA and 0.06-71.62 #{128}dC/i07 dC (Table 1 and Fig. 3). Due to the hugeinterindividual variation and small number of subjects in the
study groups, resulting in a large SD, this sex-dependent dif-ference did not reach statistical significance. However, if oneoutlier with low adduct levels was omitted, the mean edA andedC levels in five of six women on PUFA were significantly
4 I. Velic, J. Nair, and H. Bartsch, Detection of 3,W’-ethenodeoxycytidine and
1, IV”-ethenodeoxyadenosine in DNA by reverse-phase HPLC 32P-postlabeling, in
preparation.
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 599
Table 1 Description of study subjects and param eters analyzed
Group Subject no. Sex edA/iO7dA edC/lO7dC MA-dG/l07#{176} MAb (pJ�4) ��4,p�5b � Conjugated dienes (,.�M)
MUFA
PUFA
MUFA
PUFA
1
2
3
4
S
6
7
1
2
3
4
S
6
7
8
9
10
1 1
12
13
MC
M
M
M
M
M
M
F
F
F
F
F
F
F
F
F
F
F
F
F
34
24
60
31
24
23
26
23
23
23
23
43
32
24
24
24
24
65
21
46
0.96
0.06
0.53
1.20
0.43
0.15
0.33
0.44
0.03
0.05
2.52
0.1 1
0.23
1.16
0.13
20.86
8.51
12.35
90.15
10.94
0.04
0.05
0.21
1.10
0.13
0.09
0.26
0.22
0.06
0.07
0.79
0.15
0.08
0.86
0.06
24.51
12.71
3.04
71.62
8.95
0.2
5.4
0.2
2.3
1.7
4.4
2.1
2.5
1.5
1 . 1
0.2
1.2
0.2
1.2
1.2
28.7
2.2
3.6
12.0
28.0
1 .0
0.66
0.74
1.74
0.97
0.86
1.17
1.26
1.53
1 .09
1.22
0.85
0.80
0.96
0.70
0.64
0.70
1.45
0.75
1.51
5.56
3.39
6.1 1
4.32
8.79
10.68
n/a
7.74
5.67
7.44
5.49
4.59
3.55
3.61
7.33
11.19
5.23
5.94
5.61
7.82
4.63
4.63
1.85
5.00
n/a
n/a
5.56
n/a
n/a
n/a
n/a
3.33
4.44
n/a
5.37
3.70
3.70
10.93
5.37
12.22
a From Ref. 24.b From Ref. 23.
C M, male; F, female; n/a, not analyzed.
(45.4-fold and 75.7-fold, respectively) higher than those onMUFA diet (P < 0.06 and P < 0.05, respectively). Linearregression analysis of all samples showed a significant cone-
lation between edA and #{128}dClevels (r = 0.98, P = 1.79 X10� 15) suggesting that, in the DNA of human TWBCs, bothadducts were formed by a similar pathway. There was a positivecorrelation trend for the levels of etheno adducts and MA-dG
adducts in the same sample (edC versus MA-dG: r = 0.48, P =
0.01; edA versus MA-dG: r = 0.40, P = 0.08; Table 1).Surprisingly, no correlations were found between etheno ad-
ducts and other indices of lipid peroxidation in serum, such asThARS, MA, and conjugated dienes, that were measured pre-viously (Ref. 23; Table 1). Also, no significant differences were
noted for lipid peroxidation indices between the MUFA and
PUFA groups (22).
Discussion
The high mean levels (26.48/i0� edA and 22.48/i0� #{128}dC)and
the huge variation of etheno adducts in the TWBCs of femalesubjects on PUFA diet were unexpected. In human liver DNA
from healthy subjects (the only other human tissue analyzedthus far), levels were found to be in the range of 0.02-0.28
#{128}dA/lO7dA and 0.06-0.38 #{128}dC/lO7dC (1 1, 21), whereas inliver cancer-prone patients with metal storage diseases, such asWilson’s disease and primary hemochromatosis, the hepatic
levels were increased 2-3-fold (21). edA and edC adduct levelsin TWBCs of males on either MUFA or PUFA diet and in
female subjects on MUFA diet were of similar magnitude tothose seen in the liver of adult patients with metal storagediseases. The available information by questionnaire on age andlifestyle, such as smoking (none of the participants were regularsmokers), alcohol consumption, physical exercise, and the useof oral contraceptives, did not explain the -40 times increasedmean levels of etheno adducts in females on a high PUFA diet.All study subjects were kept on a similar diet (22), except for
the type of oil used, whereby PUFA diet led to a -4 times
higher mean daily intake of w-6 PUFA than the baseline diet.
Therefore, our results clearly indicate that the DNA adduction
was due to consumption of a high co-6 PUFA diet, which
resulted in elevated levels of polyunsaturated membrane lipids,
implying increased lipid peroxidation (23, 28), although com-monly used markers for lipid peroxidation status in serum, such
as MA, TBARS, and conjugated dienes, failed to reveal differ-ences between the MUFA and PUPA diet groups (23). How-
ever, significantly higher levels of MA-dG adducts were de-
tected previously in the TWBCs of subjects on PUFA diet
versus MUFA diet, and the highest mean levels of MA-dG
adducts were formed in the DNA of TWBCs of female volun-tears in the PUFA dietary group (24).
The marked diet-related difference of etheno adducts inDNA of TWBC in some females on PUFA diet may be due to
increased dietary w-6 PUFA intake and hormonal metabolism
acting synergistically, which is supported by experimental data:
treatment of male Syrian hamsters with the steroidal hormonesestradiol and diethylstilbesterol increased both lipid hydroper-
oxides and MA-DNA adducts in liver and kidney (29). Studiesin vitro suggested generation of hydroxyl radicals from estro-
gens by redox cycling, involving the 4-hydroxylated metabo-lites and their quinones (30). High intake of w-6 PUFAs,
linoleic acid and arachidonic acid, has been shown to inhibit the
detoxification of estrogens by 2-hydroxylation (31) and toelevate 16-a-hydroxylation, leading to metabolites with the
potential of redox cycling, thereby possibly enhancing the gen-
eration of hydroxyl radicals. Whether these fatty acids also
enhance the formation of 4-hydroxylated estrogens is notknown.
There was a - 1000-fold variation in etheno adduct levelsamong female individuals on w-6 PUFA diet; only one of the
female subjects had similarly low levels as males or females onMUFA diet. This interindividual variation may be due to the
following host factors: different detoxification rate of HNE,mostly by glutathione S-transferases (32); differences in repair
of etheno adducts by specific glycosylases that have been
characterized in human tissues, each capable of removing edA
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

6(X) w-6-Fatty Acids Enhance Etheno-DNA Adducts in Females
100’ , 100
.� Mean±SDn80 80
2 EJedA �cdCU
� 60 � MUFA(M) 0.7±0.6 0.4±0.5 .
� PUFA(M) 0.3±0.1 0.2±0.1
!. �#{176}. MUFA(F) 0.6±0.9 0.6±0.3 �� PUFA(F) 26.5±36.6 22.5±29.0�20 ii 20
I � IIrIIl1�t�2 [1 2
0 LiEI � EL�J�.J -
MUFA(M) PUFA(M) MUFA(F) PUFA(F)
Fig. 3. Columns, individual and mean levels of etheno adducts/l07 parent
nucleotides in human TWBCs (for sequence see Table 1). Individual volunteers
on diets: MUFA, rapeseed oil; PUFA, sunflower oil, M. male; F, female.
and #{128}dCfrom DNA (33); differential induction of specificcytochrome P-450s that catalyze 4-hydroxylation or 16 a-hy-droxylation of estradiol; indeed, a cytochrome P-450 isozymerelated to CYP1B1 that increases the 4-hydroxylation of estra-diol 5-fold has been identified in human uterine myometrium
(34).On the basis of our results, two important questions can be
addressed: Does a high dietary o-6 PUFA intake also result inincreased levels of edA and #{128}dCin tissues, such as breast,
colon, and prostate, in which fat intake or hormones are sus-pected carcinogenic risk factors? Is there a synergism betweenhormonal metabolism and exocyclic DNA adduct formation inhumans? Should such adducts be formed via hydroxyl radicals
by the w-6 PUFA pathway in these target organs, they mayoffer new etiological and mechanistic clues for tumor inductionrelated to dietary fatty acids. Etheno adducts have been proven
to be highly miscoding lesions in mammalian cells (35) and are
thought to initiate the carcinogenic process through specificpoint mutations by the known or suspected human carcinogensvinyl chloride or urethane, respectively (19).
Acknowledgments
We thank Dr. A. Barbin, IARC, Lyon, France, for preparing the standards and
antigens; Dr. M. Rajewsky and collaborators, University of Essen, Essen. Ger-many. for providing the antibodies EM-A-I and EM-C-I used in immunoaffinity
columns; M. Turpeinen, University of Helsinki, for analyzing the serum MA,
TBARS. and conjugated dienes; J-L. Fang. Karolinska Institute, Huddinge, for
DNA extraction of WBCs; C. Ditrich and I. Kling, Deutsches Krebsforschungs-zentrum (DKFZ), for excellent technical assistance; and G. Bielefeldt for secre-
tarial help.
Fig. 2. Autoradiograms of #{128}dA.#{128}dC.and deoxyuridine (dU; internal standard)
as 5-monophosphates. A, standard; B, DNA sample from volunteer on rapeseed
oil diet; C. DNA sample from volunteer on sunflower oil diet. DI, acetic acid (IM.pH 3.5); D2, saturated ammonium sulfate at -24”C (pH 3.5); X. Y. and Z,
unknown adducts retained by immunoaffinity columns. D, comparison of reverse-phase HPLC profile between standard 5’-monophosphates of edA and #{128}dC(UV)
and 32P-postlabeled adA and #{128}dC(radioactive) of one DNA sample from the
PUFA diet group. Undesignated spots or peaks are unremoved normal nucleotides
from samples and impurities from commercial [32P]ATP.
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 601
References
1. Doll, R., and Peto, R. The causes ofcancer: quantitative estimates of avoidable
risks of cancer in the United States today. J. Nail. Cancer Inst. (Bethesda), 66:
1191-1308, 1981.
2. Lipworth, L. Epidemiology of breast cancer. Eur. J. Cancer Prey., 4: 7-30,
1995.
3. Potter, J. D. Risk factors for colon neoplasia: epidemiology and biology. Eur.
J. Cancer, 31A: 1033-1038, 1995.
4. Key, T. Risk factors for prostate cancer. Cancer Sun’., 23: 63-77, 1995.
5. Bueno-de-Mesquita, H. B. Highlights of the International conference on food
factors (ICoFF): chemistry and prevention, Hamamatsu, Japan. Eur. J. Cancer
Prey., 5: 209-215, 1995.
6. Noguchi, M., Rose, D. P., Earashi, M., and Miyazaki, I. The role offatty acidsand eicosanoid synthesis inhibitors in breast carcinoma. Oncology (Basel), 52:
265-271, 1995.
7. Dreher, D., and Junod, A. F. Role of oxygen free radicals in cancer develop-
ment. Eur. J. Cancer, 31A: 30-38, 1996.
8. Vaca, C. E., Wilhelm, J., and Harms Ringdahl, M. Interaction of lipid per-
oxidation products with DNA. A review. Mutat. Res., 195: 137-149, 1988.
9. HeM, P. M., Ortner, A., and Esterbauer, H. Genotoxic properties of 4-hydroxy-
alkenals and analogous aldehydes. Mutat. Res., 290: 183-192, 1993.
10. Chaudhary, A. K., Nokubo, M., Reddy, G. R., Yeola, S. N., Morrow, J. D.,
Blair, I. A., and Mameu, L. J. Detection of endogenous malondialdehyde-
deoxyguanosine adducts in human liver. Science (Washington DC), 265: 1580-
1582, 1994.
11. Nair, J., Barbin, A., Guichard, Y., and Bartsch, H. l,N”-ethenodeoxyade-
nosine and 3,JV”-ethenodeoxycytidine in liver DNA from humans and untreated
rodents detected by immunoaffinity/32P-postlabeling. Carcinogenesis (Lond.),
16: 613-617, 1995.
12. Nath, R. G., Ocando, J. E., and Chung, F. L. Detection of l,N2-propanode-
oxyguanosine adducts as potential endogenous DNA lesions in rodent and human
tissues. Cancer Res., 56: 452-456, 1996.
13. Vaca, C. E., Fang, J. L., Mutanen, M., and Valsta, L. 32P-postlabelling
determination of DNA adducts of malonaldehyde in humans: total white blood
cells and breast tissue. Carcinogenesis (Lond.), 16: 1847-1851, 1995.
14. Poli, G., Dianzani, M. U., Cheeseman, K. H., Slater, T. F., Lang, J., and
Esterbauer, H. Separation and characterization of the aldehydic products of lipid
peroxidation stimulated by carbon tetrachloride or ADP-iron in isolated rat
hepatocytes and rat liver microsomal suspensions. Biochem. J., 227: 629-638,
1985.
15. Wu, H. Y., and Lin, J. K. Determination of aldehydic lipid peroxidation
products with dabsylhydrazine by high-performance liquid chromatography.
Anal. Chem., 67: 1603-1612, 1996.
16. Chen, H-J. C., and Chung, F-L, Epoxidation of trans-4-hydroxy-2-nonenal
by fatty acid hydroperoxides and hydrogen peroxide. (1cm. Res. Toxicol., 9:
306-312, 1996.
17. Sodum, R. S., and Chung, F. L. 1, N�-ethenodeoxyguanosine as a potential
marker for DNA adduct formation by trans-4-hydroxy-2-nonenal. Cancer Res.,
48: 320-323, 1988.
18. El Ghissassi, F., Barbin, A., Nair, J., and Bartsch, H. Formation of lJV”-
ethenoadenine and 3,N”-ethenocytosine by lipid peroxidation products and flu-
cleic acid bases. Chem. Rca. Toxicol., 8: 278-283, 1995.
19. Bartsch, H., Barbin, A., Marion, M. J., Nair, J., and Guichard, Y. Formation,
detection, and role in carcinogenesis of ethenobases in DNA. Drug Metab. Rev.,
26: 349-371, 1994.
20. Nair, J., Sone, H., Nagao, M., Barbin, A., and Bartsch, H. Copper-dependent
formation of miscoding etheno-DNA adducts in the liver of Long Evans cinna-
mon (LEC) rats developing hereditary hepatitis and hepatocellular carcinoma.
CancerRes., 56: 1267-1271, 1996.
21. Nair, J. Carmichael, P. L., Fernando, R. C., Phillips, D. H., and Bartsch, H.
Elevated I, IV”-ethenodeoxyadenosine (adA) and 3-N”-ethenodeoxycytidine(#{128}dC)in liver with Wilson’s disease and primary hemochromatosis. Proc. Am.
Assoc. Cancer Res., 37: 813, 1996.
22. Valsta, L M., Jauhiainen, M., Am, A., Katan, M. B., and Mutanen, M.
Effects of a monounsaturated rapeseed oil and a polyunsaturated sunflower oil
diet on lipoprotein levels in humans. Arterioscler. Thromb., 12: 50-57, 1992.
23. Turpeinen. A. M., Alfthan, G., Valsta, L, Hietanen, E., Salonen, J. T.,
Schunk, H., Nyyssonen, K., and Mutanen, M. Plasma and lipoprotein lipid
peroxidation in humans on sunflower and rapeseed oil diets. Lipids, 30: 485-492,
1995.
24. Fang, J. L, Vaca, C. E., Valsta, L. M., and Mutanen, M. Determination ofDNA adducts of malonaldehyde in humans: effects of dietary fatty acid compo-
sition. Carcinogenesis (Land.), 17: 1035-1040, 1996.
25. Guichard, Y., El Ghissassi, F., Nair, J., Bartsch, H., and Barbin, A. Formationand accumulation of DNA ethenobases in adult Sprague-Dawley rats exposed to
vinyl chloride. Carcinogenesis (Lond.), 17: 1553-1559, 1996.
26. Fernando, R. C., Nair, J., Barbin, A., Miller, J. A., and Bartsch, H. Detection
of 1, N”- ethenodeoxyadenosine and 3,N’-ethenodeoxycytidine by immunoaffin-
ity/32P- postlabelling in liver and lung DNA of mice treated with ethyl carbamate
(urethane) or its metabolites. Carcinogenesis (Lond.), 17: 171 1-1559, 1996.
27. Eberle, G., Barbin, A., Laib, R. J., Ciroussel, F., Thomale, J., Bartsch, H., and
Rajewsky, M. F. I,JV”-etheno-2’-deoxyadenosine and 3,W’-etheno-2’-deoxycy-
tidine detected by monoclonal antibodies in lung and liver DNA of rats exposed
to vinyl chloride. Carcinogenesis (Lond.), 10: 209-212, 1989.
28. Hammer, C. T., and Wills, E. D. The effect of dietary fats on the composition
of the liver endoplasmic reticulum and oxidative drug metabolism. Br. J. Nutr.,
41: 465-475, 1979.
29. Wang, M. Y., and Liehr, J. G. Induction by estrogens of lipid peroxidationand lipid peroxide-derived malonaldehyde-DNA adducts in male Syrian ham-
sters: role of lipid peroxidation in estrogen-induced kidney carcinogenesis. Car-
cinogenesis (Land.), 16: 1941-1945, 1995.
30. Han, X., and Liehr, J. G. Microsome-mediated 8-hydroxylation of guanine
bases of DNA by steroid estrogens: correlation of DNA damage by free radicalswith metabolic activation to quinones. Carcinogenesis (Lond.), 16: 257 1-2574,
1995.
31. Davis, D. L., Bradlow, H. L., Wolff, M., Woodruff, T., Hod, D. G., andAnton Culver, H. Medical hypothesis: xenoestrogens as preventable causes of
breast cancer. Environ. Health Perspect., 101: 372-377, 1993.
32. Hartley, D. P., Ruth, J. A., and Petersen, D. R. The hepatocellular metabolism
of 4-hydroxynonenal by alcohol dehydrogenase, aldehyde dehydrogenase. and
glutathione S-transferase. Arch. Biochem. Biophys., 316: 197-205, 1995.
33. Hang, B., Chenna, A., Rao, S., and Singer, B. I, N”-ethei.uadenine and
3, N�-ethenocytosine are excised by separate human DNA glycosylases. Carci-
nogenesis (Lond.), 17: 155-157, 1996.
34. Liehr, J. G., Ricci, M. J., Jefcoate, C. R., Hannigan, E. V., Hokanson, J. A.,
and Thu. B. T. 4-Hydroxylation of estradiol by human uterine myometrium and
myoma microsomes: implications for the mechanism of uterine tumorigenesis.
Proc. NatI. Acad. Sci. USA, 92: 9220-9224, 1995.
35. Moriya, M., Zhang, W., Johnson, F., and Grollman, A. P. Mutagenic potency
of exocyclic DNA adducts: marked differences between Escherichia coli and
simian kidney cells. Proc. Nail. Acad. Sci. USA, 91: 1 1899-1 1903, 1994.
36. Chung. F-L., Chen, H-J. C., and Nath, R. G. Lipid peroxidation as a potential
endogenous source for the formation of exocyclic DNA adducts. Carcinogenesis
(Land.), 17: 2105-2111, 1996.
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

1997;6:597-601. Cancer Epidemiol Biomarkers Prev J Nair, C E Vaca, I Velic, et al. blood cells of female subjects.increase the formation of etheno-DNA base adducts in white High dietary omega-6 polyunsaturated fatty acids drastically
Updated version
http://cebp.aacrjournals.org/content/6/8/597
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cebp.aacrjournals.org/content/6/8/597To request permission to re-use all or part of this article, use this link
on November 27, 2018. © 1997 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
Related Documents











