Hidden diversity in bent-winged bats (Chiroptera: Miniopteridae) of the Western Palaearctic and adjacent regions: implications for taxonomy JAN ŠRÁMEK 1 *, VÁCLAV GVOŽDÍK 2,3 and PETR BENDA 1,2 1 Department of Zoology, Faculty of Science, Charles University, Vinic ˇná 7, CZ–128 44 Prague, Czech Republic 2 Department of Zoology, National Museum (Natural History), Václavské nám. 68, CZ–115 79 Prague, Czech Republic 3 Laboratory of Molecular Ecology, Institute of Animal Physiology and Genetics, Academy of Sciences of the Czech Republic, Rumburská 89, CZ–277 21 Libe ˇchov, Czech Republic Received 6 December 2011; revised 28 August 2012; accepted for publication 29 August 2012 The taxonomic status of bent-winged bats (Miniopterus) in the Western Palaearctic and adjacent regions is unclear, particularly in some areas of the eastern Mediterranean, Middle East and Arabia. To address this, we analysed an extensive collection of museum materials from all principal parts of this distribution range, i.e. North Africa, Europe and southwest Asia, using morphological (skull) and genetic approaches (mitochondrial DNA). Linear and geometric morphometric analysis of cranial and dental characteristics, together with molecular phylogeny, suggested that Miniopterus populations comprise four separate species: (1) M. schreibersii sensu strictissimo (s.str.) – occurring in Europe, coastal Anatolia, Levant, Cyprus, western Transcaucasia, and North Africa; (2) M. pallidus – occurring in inland Anatolia, Jordan, eastern Transcaucasia, Turkmenistan, Iran and southern Afghanistan (Kandahar); (3) a Miniopterus sp. – recorded from Nangarhar province in eastern Afghanistan, which we tentatively assign to M. cf. fuliginosus; and (4) a Miniopterus sp. with Afro-tropic affinities confirmed from south-western Arabia and Ethiopia, which we tentatively name M. cf. arenarius. The latter two species are well differentiated by skull morphology, while M. pallidus possesses very similar skull morphology to M. schreibersii. The results also suggest the existence of a possible new taxon (subspecies) within M. schreibersii s.str. inhabiting the Atlas Mountains of Morocco. © 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190. doi: 10.1111/j.1096-3642.2012.00870.x ADDITIONAL KEYWORDS: Arabia – bent-winged bats – cryptic species – Europe – Middle East – mitochondrial DNA – morphology – North Africa – phylogeography – systematics. INTRODUCTION Bent-winged bats, family Miniopteridae, are repre- sented by a single genus, Miniopterus Bonaparte, 1837. The genus includes up to 19 species occurring mostly in the tropics and subtropics of the Old World, viz. Africa (except the Sahara), southern and central Europe, southern Asia from Anatolia, across the Middle East and Transcaucasia to China and Japan, the Sunda archipelago, the Philippines, and the Australasian region (Simmons, 2005). Morphological analysis suggests that the named forms (species/ subspecies) of this genus are very similar in their cranial and external characteristics (e.g. Tate, 1941; Maeda, 1982; Benda et al., 2006), meaning that *Corresponding author. Current address: Department of Cell and Molecular Biology, Third Faculty of Medicine, Charles University, Ruská 87, CZ–100 00 Prague, Czech Republic. E-mail: [email protected] Zoological Journal of the Linnean Society, 2013, 167, 165–190. With 4 figures © 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190 165

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hidden diversity in bent-winged bats (Chiroptera:Miniopteridae) of the Western Palaearctic and adjacentregions: implications for taxonomy
JAN ŠRÁMEK1*, VÁCLAV GVOŽDÍK2,3 and PETR BENDA1,2
1Department of Zoology, Faculty of Science, Charles University, Vinicná 7, CZ–128 44 Prague, CzechRepublic2Department of Zoology, National Museum (Natural History), Václavské nám. 68, CZ–115 79 Prague,Czech Republic3Laboratory of Molecular Ecology, Institute of Animal Physiology and Genetics, Academy of Sciencesof the Czech Republic, Rumburská 89, CZ–277 21 Libechov, Czech Republic
Received 6 December 2011; revised 28 August 2012; accepted for publication 29 August 2012
The taxonomic status of bent-winged bats (Miniopterus) in the Western Palaearctic and adjacent regions is unclear,particularly in some areas of the eastern Mediterranean, Middle East and Arabia. To address this, we analysed anextensive collection of museum materials from all principal parts of this distribution range, i.e. North Africa,Europe and southwest Asia, using morphological (skull) and genetic approaches (mitochondrial DNA). Linearand geometric morphometric analysis of cranial and dental characteristics, together with molecular phylogeny,suggested that Miniopterus populations comprise four separate species: (1) M. schreibersii sensu strictissimo (s.str.)– occurring in Europe, coastal Anatolia, Levant, Cyprus, western Transcaucasia, and North Africa; (2) M. pallidus– occurring in inland Anatolia, Jordan, eastern Transcaucasia, Turkmenistan, Iran and southern Afghanistan(Kandahar); (3) a Miniopterus sp. – recorded from Nangarhar province in eastern Afghanistan, which wetentatively assign to M. cf. fuliginosus; and (4) a Miniopterus sp. with Afro-tropic affinities confirmed fromsouth-western Arabia and Ethiopia, which we tentatively name M. cf. arenarius. The latter two species are welldifferentiated by skull morphology, while M. pallidus possesses very similar skull morphology to M. schreibersii.The results also suggest the existence of a possible new taxon (subspecies) within M. schreibersii s.str. inhabitingthe Atlas Mountains of Morocco.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190.doi: 10.1111/j.1096-3642.2012.00870.x
ADDITIONAL KEYWORDS: Arabia – bent-winged bats – cryptic species – Europe – Middle East –mitochondrial DNA – morphology – North Africa – phylogeography – systematics.
INTRODUCTION
Bent-winged bats, family Miniopteridae, are repre-sented by a single genus, Miniopterus Bonaparte,1837. The genus includes up to 19 species occurring
mostly in the tropics and subtropics of the Old World,viz. Africa (except the Sahara), southern and centralEurope, southern Asia from Anatolia, across theMiddle East and Transcaucasia to China and Japan,the Sunda archipelago, the Philippines, and theAustralasian region (Simmons, 2005). Morphologicalanalysis suggests that the named forms (species/subspecies) of this genus are very similar in theircranial and external characteristics (e.g. Tate, 1941;Maeda, 1982; Benda et al., 2006), meaning that
*Corresponding author. Current address: Department ofCell and Molecular Biology, Third Faculty of Medicine,Charles University, Ruská 87, CZ–100 00 Prague, CzechRepublic. E-mail: [email protected]
bs_bs_banner
Zoological Journal of the Linnean Society, 2013, 167, 165–190. With 4 figures
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190 165

identification of many taxa is difficult and the classi-fication of many populations of this genus remainsunclear. Further, a number of recent molecular phy-logenetic studies have indicated that the taxonomy ofthis genus is in urgent need of revision (Appleton,McKenzie & Christidis, 2004; Tian et al., 2004; Miller-Butterworth et al., 2005; Furman et al., 2009, 2010c;Furman, Öztunç & Çoraman, 2010b). This is particu-larly true for Miniopterus schreibersii (Kuhl, 1817)sensu lato (s.l.), the only species considered as inhab-iting the whole south-western portion of the Palae-arctic region (Koopman, 1993, 1994; Simmons, 2005).
For a long time, this species was considered apolytypic and widespread bat, with up to 25 subspe-cies recognised within its distribution range, whichis almost identical to that of the genus and com-prises most of the Old World region (e.g. Tate, 1941;Hayman & Hill, 1971; Corbet, 1978; Harrison &Bates, 1991; Corbet & Hill, 1992; Koopman, 1994).Nowadays, M. schreibersii sensu stricto (s.s.) is mostoften accepted as a south-western Palaearctic faunalelement occurring in southern and central Europe,supra-Saharan Africa, south-west Asia, and easternAfghanistan (cf. Appleton et al., 2004; Tian et al.,2004; Miller-Butterworth et al., 2005; Benda et al.,2006; Bilgin et al., 2006, 2008; Furman et al., 2009,2010b). It is interesting to note, however, that thesegeographical limits for M. schreibersii had alreadybeen proposed by Maeda (1982) in his precise mor-phometric analysis of the Palaearctic and Orientalpopulations of the genus.
The newly delimited species rank of M. schreibersii(as reviewed by Simmons, 2005) consists of twosubspecies, M. s. schreibersii [type locality (t.l.):Kolumbács Cave, left bank of the River Danube,near Coronini, Romania; sensu Ansell & Topál, 1976]and M. s. pallidus Thomas, 1907 [t.l.: vicinity ofBandar-i-Gaz (Golestan Province), Iran; sensu Lay,1967]. These subspecies are very similar in bothexternal and cranial characteristics (e.g. Ognev,1928; Albayrak & Coskun, 2000; Benda et al., 2006;Furman et al., 2009) and differ mainly in pelage col-oration. Evidence of seasonal and geographic changesin this trait, however, has shown it to be unsuit-able for taxonomic identification (Kuzâkin, 1950;Lay, 1967; Karatas & Sözen, 2004; Gazaryan, 2005).Furman et al. (2010c) studied differentiation betweenthese taxa in populations inhabiting Asia Minor andfound statistically significant genetic, morphological(body size and wing shape) and echolocation differ-ences. Following these findings, they suggested thatthe two taxa were reproductively isolated and consid-ered them to be two separately evolving units repre-senting distinct cryptic species, M. schreibersii sensustrictissimo (s.str.) and M. pallidus. This taxonomicproposal, however, was based almost solely on the
populations from Turkey, while most of the Palaearc-tic range of the bent-winged bats still remains ques-tionable from a taxonomic and phylogeographic pointof view (cf. Bilgin, 2011, 2012).
The subspecific taxonomic rank of the taxonpallidus has been applied, particularly by Russianauthors (e.g. Ellerman & Morrison-Scott, 1951;Kuzâkin, 1965; Strelkov, Sosnovcena & Babaev, 1978;Rahmatulina, 2005), for populations occurring insome areas of the former Soviet Union (currently theCaucasus region and southern Turkmenistan; Ognev,1927, 1928; Gazaryan, 2005). Distribution of thisform has also been reported from the Levant (Syria,Lebanon, Israel, Jordan), Iraq, Iran, Afghanistan andfrom the inland/highland areas of central and easternTurkey (e.g. Lay, 1967; Gaisler, 1970; Maeda, 1982;Horácek, Hanák & Gaisler, 2000; Boye, 2004; Bendaet al., 2006; Furman et al., 2010c); Ferguson (2002),however, reported the occurrence of a subspecies,schreibersii, for Israel. Populations from the Euro-pean distribution range of M. schreibersii, s.s. as wellas populations from North Africa and the larger Medi-terranean islands, have traditionally been attributedto the nominotypical subspecies (e.g. Aellen & Stri-nati, 1970; Spitzenberger, 1981; Gaisler, 1983; Cru-citti, 1989; Kowalski & Rzebik-Kowalska, 1991); thesituation in Cyprus, however, remains unclear. Boye(2004), for example, mentions the occurrence of sub-species pallidus, while others assume the island to beinhabited by the nominotypical form (e.g. Horáceket al., 2000).
In the Middle East, a morphologically distinctpopulation of M. schreibersii s.l. has been suggestedas present in the Nangarhar province of easternAfghanistan, on the border of the Palaearctic andOriental regions (Gaisler, 1970), and was thought torepresent M. s. fuliginosus Hodgson, 1835 (t.l.: Nepal)(Gaisler, 1970; Hill, 1983; Koopman, 1994; Bates &Harrison, 1997). A further population traditionallyassigned to M. schreibersii s.l. occurs at the border ofthe Palaearctic in south-western Arabia (Harrison &Bates, 1991). These populations were originally clas-sified as M. s. arenarius Heller, 1912 (t.l.: Nanyuki,Kenya) (Nader & Kock, 1987; Harrison & Bates, 1991;Koopman, 1994). This taxon, however, is currentlyconsidered as part of M. natalensis Smith, 1834 (t.l.:Durban, South Africa), which was recently revali-dated to species level within M. schreibersii s.l and isreported to occur in sub-Saharan Africa and south-western Arabia (Koopman, 1994; Simmons, 2005).
To summarise, the taxonomic status and dis-tribution ranges of particular taxa/populations of Pal-aearctic Miniopterus bats have tended to be reportedmore-or-less intuitively (mainly on a geographicbasis) and its status remains unclear in most of therespective areas. Classification of the Levantine,
166 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Middle Eastern, North African and, especially,European populations remains in need of revision.Delimitation of the geographical margins and contactzones between respective taxa, especially in the lightof new findings (cf. Furman et al., 2010c), is alsonecessary.
Here, we present a revision of the taxonomy ofMiniopterus populations of Western Palaearctic andsome adjacent regions, based on a synthesis of resultsfrom morphological and molecular analysis of a richmuseum-material collection from all principal parts ofthe Miniopterus distribution range (i.e. southernEurope, south-west Asia and North Africa). In doingso, we aim to answer two main questions arisingfrom several recent studies (e.g. Appleton et al., 2004;Miller-Butterworth et al., 2005; Bilgin et al., 2008;Furman et al., 2010b, c): (1) what are the phylogeneticand phenotypic relationships between particularWestern Palaearctic Miniopterus populations (aswell as their taxonomic status), and (2) is M. pallidus[demonstrated as representing a separate species in arecent study (Furman et al., 2010c)] morphologicallywell differentiated and what is its present distribu-tion range?
MATERIAL AND METHODS
In order to assess the taxonomic status of Miniopteruspopulations from the Western Palaearctic and adja-cent regions (i.e. Afghanistan, Yemen and Ethiopia),we examined 352 skulls morphologically (Appendix 1)and 52 samples genetically. Fifty-seven additionalsequences of Miniopterus spp. from around the OldWorld were retrieved from GenBank (Table 1). Areview of the geographic origin of all the materialinvestigated is displayed in Figure 1A.
MOLECULAR ANALYSIS
Total genomic DNA was extracted from tissuesamples (c. 1 mm2 of wing membrane) using theGenomed JetQuick Tissue DNA Spin Kit (Löhne,Germany), following the manufacturer’s protocol. Asegment of extracted mitochondrial DNA (mtDNA),the complete gene for the second subunit of NADHdehydrogenase (ND2 – 1044 bp), was amplified byPolymerase Chain Reaction (PCR) using the primersND2-1 and ND2-2 (Kirchman et al., 2001) underthe following thermal profile: initial denaturation of93 °C for 3 min, 35 subsequent cycles of 93 °C for 30 s,52 °C for 40 s and 72 °C for 1 min, and a final exten-sion of 72 °C for 10 min. Sequencing was carriedout by Macrogen Inc. (Seoul, South Korea, http://www.macrogen.com) using a combination of theabove mentioned PCR primers, one formerly pub-lished internal forward primer (mmND2.1; Osborne
& Christidis, 2001), and one newly developedMiniopterus-specific internal reverse primer(mND2inR: 5′-TGAATRACYGCCGTACTA-3′). Newsequences of different haplotypes were depositedin GenBank (see Table 1 for Accession Nos.). Fifty-seven additional sequences from various Miniopterusspecies were added to our dataset from GenBank(AY169435–46, AY169448–71, Appleton et al., 2004;GU290290–310, Furman et al., 2010b), as well asfour outgroup taxa: Myotis muricola (AY504566;J. M. Worthington Wilmer, C. J. Schneider & M. D.Sorenson, unpubl. data), Chalinolobus tuberculatus(AF321051; Lin & Penny, 2001), Chalinolobus nigrog-riseus (AY504561; J. M. Worthington Wilmer, C. J.Schneider & M. D. Sorenson, unpubl. data), andChalinolobus morio (AY169472; Appleton et al., 2004).For phylogenetic analysis, we shortened the newsequences from this study to 1034 bp in orderto match the additional GenBank sequences. Allsequences were aligned in BioEdit 7.0 (Hall, 1999)and examined by translation into amino acids withthe vertebrate mitochondrial genetic code usingDnaSP 5.10 (Librado & Rozas, 2009); no stop codonswere detected.
Phylogenetic trees were constructed using a datasetof 89 sequences that comprised only unique haplo-types (haplotype dataset). The trees were constructedusing the maximum likelihood (ML), Bayesianapproach (BA), and neighbour-joining (NJ) methods.For ML and BA, the jModelTest 0.1.1 softwarepackage (Posada, 2008) was employed prior to analy-sis to calculate the best-fit model of nucleotide evolu-tion (selected according to the Akaike informationcriterion for the whole sequence length in ML, andeach codon position separately in BA). ML analysiswas performed using PhyML 3.0 (Guindon et al.,2010). The best-fit substitution model correspondedwith the transitional model with a proportion ofinvariant positions and gamma distribution ofrate heterogeneity (TIM2 + I + G). The best branch-swapping approach was applied, which combinesnearest neighbour interchanges with subtree pruningand regrafting, and optimisation of topology andbranch length settings. Bootstrap branch support wascalculated based on 1000 resampled datasets. The BAwas carried out using MrBayes 3.2 (Huelsenbeck& Ronquist, 2001; Ronquist & Huelsenbeck, 2003),with partitions for codon positions and parametersoptimised during runs. The likelihood settings corre-sponded with the general time-reversible model,which was the closest approximation of the best-fitsubstitution model for each partition available inMrBayes (we applied GTR + G/GTR + G/GTR + I + Gfor codon position 1/2/3). BA analysis was performedfor six million generations with two runs (to checkconvergence) and four coupled chains for each run,
TAXONOMY OF MINIOPTERUS 167
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Tab
le1.
Spe
cim
ens
exam
ined
gen
etic
ally
Spe
cies
Hap
loty
peL
inea
ge/
subl
inea
geC
oun
try
Loc
alit
yC
oord
inat
esG
enB
ank
Acc
.N
o.V
ouch
er/R
efer
ence
M.s
chre
iber
sii
MS
C1
WM
Ital
yE
tna,
Sic
ily
37.7
2N
,14
.92
EJX
0121
35bi
opsy
M.s
chre
iber
sii
MS
C1
WM
Ital
yE
tna,
Sic
ily
37.7
2N
,14
.92
EJX
0121
35bi
opsy
M.s
chre
iber
sii
MS
C1
WM
Ital
yE
tna,
Sic
ily
37.7
2N
,14
.92
EJX
0121
35bi
opsy
M.s
chre
iber
sii
MS
C1
WM
Rom
ania
Bet
fia
46.9
8N
,22
.02
EJX
0121
35bi
opsy
M.s
chre
iber
sii
MS
C1
WM
Rom
ania
Bet
fia
46.9
8N
,22
.02
EJX
0121
35bi
opsy
M.s
chre
iber
sii
MS
C1
WM
Slo
vaki
aD
rien
ovec
48.6
2N
,20
.95
EJX
0121
35N
MP
pb42
61M
.sch
reib
ersi
iM
SC
1W
MS
lova
kia
Dri
enov
ec48
.62
N,
20.9
5E
JX01
2135
NM
Ppb
4260
M.s
chre
iber
sii
MS
C2
WM
Rom
ania
Du
bova
47.6
2N
,22
.25
EJX
0121
36N
MP
pb44
19M
.sch
reib
ersi
iM
SC
2W
MR
oman
iaB
etfi
a46
.98
N,
22.0
2E
JX01
2136
NM
Ppb
4256
M.s
chre
iber
sii
MS
C3
WM
Rom
ania
Bet
fia
46.9
8N
,22
.02
EJX
0121
37N
MP
pb42
58M
.sch
reib
ersi
iM
SC
4W
MG
reec
eM
ilat
os,
Cre
te35
.30
N,
25.5
8E
JX01
2138
NM
P91
116
M.s
chre
iber
sii
MS
C4
WM
Gre
ece
Mil
atos
,C
rete
35.3
0N
,25
.58
EJX
0121
38N
MP
9111
3M
.sch
reib
ersi
iM
SC
4W
MG
reec
eO
mal
os,
Cre
te35
.35
N,
23.9
0E
JX01
2138
NM
P91
166
M.s
chre
iber
sii
MS
C4
WM
Gre
ece
Vre
ikos
Cav
e,C
rete
35.0
8N
,26
.00
EJX
0121
38N
MP
9231
6M
.sch
reib
ersi
iM
SC
5W
MG
reec
eL
efko
gia,
Cre
te35
.18
N,
24.4
7E
JX01
2139
NM
P92
311
M.s
chre
iber
sii
MS
C5
WM
Gre
ece
Om
alos
,C
rete
35.3
5N
,23
.90
EJX
0121
39N
MP
9117
2M
.sch
reib
ersi
iM
SC
6E
MC
ypru
sK
akop
etri
a34
.97
N,
32.8
7E
JX01
2140
NM
PC
H10
8M
.sch
reib
ersi
iM
SC
6E
MC
ypru
sK
akop
etri
a34
.97
N,
32.8
7E
JX01
2140
NM
PC
H46
M.s
chre
iber
sii
MS
C7
EM
Cyp
rus
Aka
mas
Pen
insu
la35
.05
N,
32.3
3E
JX01
2141
NM
PC
H12
3M
.sch
reib
ersi
iM
SC
7E
MC
ypru
sK
akop
etri
a34
.97
N,
32.8
7E
JX01
2141
NM
P90
406
M.s
chre
iber
sii
MS
C7
EM
Cyp
rus
Kak
opet
ria
34.9
7N
,32
.87
EJX
0121
41N
MP
9040
5M
.sch
reib
ersi
iM
SC
7E
MC
ypru
sK
alav
asos
34.8
0N
,33
.27
EJX
0121
41N
MP
9043
4M
.sch
reib
ersi
iM
SC
8E
MS
yria
Qal
a’at
al-H
osn
35.6
5N
,40
.73
EJX
0121
42N
MP
pb49
989
M.s
chre
iber
sii
MS
C8
EM
Syr
iaTa
lsh
’hab
32.7
0N
,35
.96
EJX
0121
42N
MP
4886
1M
.sch
reib
ersi
iM
SC
9E
MTu
rkey
Akb
ez36
.51
N,
36.3
0E
JX01
2143
NM
Ptr
099
M.s
chre
iber
sii
MS
C10
EM
Leb
anon
Aaq
ura
34.1
2N
,35
.92
EJX
0121
44N
MP
9177
8M
.sch
reib
ersi
iM
SC
10E
MS
yria
Safi
ta34
.83
N,
36.1
2E
JX01
2144
NM
P48
883
M.s
chre
iber
sii
MS
C11
EM
Syr
iaQ
ala’
atal
-Hos
n35
.65
N,
40.7
3E
JX01
2145
NM
P48
873
M.s
chre
iber
sii
MS
C11
EM
Syr
iaS
afita
34.8
3N
,36
.12
EJX
0121
45N
MP
4888
1M
.sch
reib
ersi
iM
SC
12E
MC
ypru
sK
akop
etri
a34
.97
N,
32.8
7E
JX01
2146
NM
PC
H45
M.s
chre
iber
sii
MS
C12
EM
Leb
anon
Aam
chit
e34
.15
N,
35.6
7E
JX01
2146
NM
P91
808
M.s
chre
iber
sii
MS
C13
EM
Leb
anon
Aaq
ura
34.1
2N
,35
.92
EJX
0121
47N
MP
9177
7M
.sch
reib
ersi
iM
SC
14M
OM
oroc
coTa
zou
guer
te32
.02
N,
03.7
8W
JX01
2148
NM
Ppb
3906
M.s
chre
iber
sii
MS
C14
MO
Mor
occo
Tazo
ugu
erte
32.0
2N
,03
.78
WJX
0121
48N
MP
pb39
08M
.sch
reib
ersi
iM
SC
15M
OM
oroc
coS
ebt-
des-
Ait
-S
erh
rou
chen
34.0
3N
,04
.57
WJX
0121
49N
MP
9010
3
M.s
chre
iber
sii
MS
C15
MO
Mor
occo
Talk
out
31.6
8N
,07
.28
WJX
0121
49N
MP
9004
7M
.sch
reib
ersi
iM
SC
16M
OM
oroc
coTa
lkou
t31
.68
N,
07.2
8W
JX01
2150
NM
P90
051
168 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

M.s
chre
iber
sii
MS
C17
WM
Bu
lgar
iaP
azar
dzik
AY
1694
46A
pple
ton
etal
.(2
004)
M.s
chre
iber
sii
MS
C18
WM
Bu
lgar
iaS
ofia
AY
1694
45A
pple
ton
etal
.(2
004)
M.s
chre
iber
sii
MS
C19
WM
Geo
rgia
Gh
lian
aG
U29
0307
Fu
rman
etal
.(2
010b
)M
.sch
reib
ersi
iM
SC
20W
MG
eorg
iaG
hli
ana
GU
2903
08F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C21
WM
Mor
occo
Aga
dir
AY
1694
50A
pple
ton
etal
.(2
004)
M.s
chre
iber
sii
MS
C21
WM
Mor
occo
Aga
dir
AY
1694
49A
pple
ton
etal
.(2
004)
M.s
chre
iber
sii
MS
C22
WM
Spa
inC
adiz
AY
1694
48A
pple
ton
etal
.(2
004)
M.s
chre
iber
sii
MS
C23
WM
Turk
eyH
ızar
GU
2903
01F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C24
WM
Turk
eyH
orat
ası
GU
2903
02F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C25
EM
Turk
eyK
aran
lık
GU
2903
04F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C26
EM
Turk
eyK
aran
lık
GU
2903
05F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C27
EM
Turk
eyO
bru
kG
U29
0309
Fu
rman
etal
.(2
010b
)M
.sch
reib
ersi
iM
SC
28E
MTu
rkey
Obr
uk
GU
2903
10F
urm
anet
al.
(201
0b)
M.s
chre
iber
sii
MS
C29
EM
Turk
eyZ
inda
nG
U29
0303
Fu
rman
etal
.(2
010b
)M
.sch
reib
ersi
iM
SC
30E
MTu
rkey
Zin
dan
GU
2903
06F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA1
ME
Iran
Dor
ud
33.4
5N
,49
.02
EJX
0121
51N
MP
4815
4M
.pal
lid
us
MPA
2M
EIr
anB
isot
un
34.3
8N
,47
.43
EJX
0121
52N
MP
4815
1M
.pal
lid
us
MPA
3M
EIr
anM
ina
37.3
0N
,58
.97
EJX
0121
53N
MP
9082
5M
.pal
lid
us
MPA
3M
EIr
anM
ina
37.3
0N
,58
.97
EJX
0121
53N
MP
9082
6M
.pal
lid
us
MPA
4M
EIr
anB
isot
un
34.3
8N
,47
.43
EJX
0121
54N
MP
4814
9M
.pal
lid
us
MPA
5M
EJo
rdan
Kh
ash
ibah
32.2
2N
,35
.72
EJX
0121
55N
MP
9253
2M
.pal
lid
us
MPA
6M
EA
zerb
aija
nA
zıx
GU
2902
93F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA7
ME
Aze
rbai
jan
Azı
xG
U29
0290
Fu
rman
etal
.(2
010b
)M
.pal
lid
us
MPA
8M
EA
zerb
aija
nA
zıx
GU
2902
95F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA8
ME
Turk
eyE
pçik
GU
2902
96F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA9
ME
Iran
Kar
aftu
GU
2902
92F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA10
ME
Iran
Kar
aftu
GU
2902
94F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA11
ME
Turk
eyD
elik
liG
U29
0299
Fu
rman
etal
.(2
010b
)M
.pal
lid
us
MPA
12M
EIr
anS
arin
Ab-
Gar
ma
GU
2902
91F
urm
anet
al.
(201
0b)
M.p
alli
du
sM
PA13
ME
Turk
eyD
elik
liG
U29
0300
Fu
rman
etal
.(2
010b
)M
.pal
lid
us
MPA
14M
ETu
rkey
Epç
ikG
U29
0297
Fu
rman
etal
.(2
010b
)M
.pal
lid
us
MPA
14M
ETu
rkey
Epç
ikG
U29
0298
Fu
rman
etal
.(2
010b
)M
.afr
ican
us
MA
F1
Eth
iopi
aS
ofO
mar
06.0
9N
,40
.85
EJX
0121
61N
MP
9212
9M
.afr
ican
us
MA
F2
Eth
iopi
aS
ofO
mar
06.0
9N
,40
.85
EJX
0121
62N
MP
9212
7M
.‘a
ust
rali
s’M
AU
1A
ust
rali
aC
ape
York
AY
1694
53A
pple
ton
etal
.(2
004)
M.
‘au
stra
lis’
MA
U2
Au
stra
lia
Sh
oalw
ater
Bay
AY
1694
52A
pple
ton
etal
.(2
004)
M.
‘au
stra
lis’
MA
U3
Indo
nes
iaJa
vaA
Y16
9444
App
leto
net
al.
(200
4)M
.‘a
ust
rali
s’M
AU
4P
hil
ippi
nes
Ley
teIs
lan
dA
Y16
9458
App
leto
net
al.
(200
4)M
.‘a
ust
rali
s’M
AU
5P
hil
ippi
nes
Neg
ros
Isla
nd
AY
1694
57A
pple
ton
etal
.(2
004)
TAXONOMY OF MINIOPTERUS 169
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Tab
le1.
Con
tin
ued
Spe
cies
Hap
loty
peL
inea
ge/
subl
inea
geC
oun
try
Loc
alit
yC
oord
inat
esG
enB
ank
Acc
.N
o.V
ouch
er/R
efer
ence
M.
‘au
stra
lis’
MA
U6
Van
uat
uA
ore
Isla
nd
AY
1694
54A
pple
ton
etal
.(2
004)
M.
‘au
stra
lis’
MA
U7
Van
uat
uTe
gua
Isla
nd
AY
1694
55A
pple
ton
etal
.(2
004)
M.
‘au
stra
lis’
MA
U8
Van
uat
uTo
gaIs
lan
dA
Y16
9456
App
leto
net
al.
(200
4)M
.bas
san
iiM
BA
Au
stra
lia
Nar
acoo
rte
AY
1694
35A
pple
ton
etal
.(2
004)
M.f
uli
gin
osu
sM
FU
1C
hin
aY
un
anA
Y16
9468
App
leto
net
al.
(200
4)M
.fu
ligi
nos
us
MF
U2
Japa
nW
akay
ama
AY
1694
69A
pple
ton
etal
.(2
004)
M.i
nfl
atu
sM
INU
gan
daR
wen
zori
Mou
nta
ins
AY
1694
65A
pple
ton
etal
.(2
004)
M.m
agn
ater
MM
AP
apu
aN
ewG
uin
eaN
ong
Riv
erA
Y16
9443
App
leto
net
al.
(200
4)M
.man
avi
MM
NM
adag
asca
rA
ndr
ingi
tra
Res
erve
AY
1694
64A
pple
ton
etal
.(2
004)
M.
‘med
ius’
MM
E1
Pap
ua
New
Gu
inea
Mag
idob
oA
Y16
9441
App
leto
net
al.
(200
4)M
.‘m
ediu
s’M
ME
2P
apu
aN
ewG
uin
eaS
olR
iver
AY
1694
42A
pple
ton
etal
.(2
004)
M.
cf.
aren
ariu
sM
AR
1Y
EYe
men
Hal
hal
15.7
3N
,43
.62
EJX
0121
56N
MP
pb37
47M
.cf
.ar
enar
ius
MA
R2
YE
Yem
enR
iqab
14.8
7N
,43
.42
EJX
0121
57N
MP
pb31
27M
.cf
.ar
enar
ius
MA
R3
YE
Eth
iopi
aM
ash
a07
.87
N,
35.4
8E
JX01
2158
NM
P92
178
M.
cf.
aren
ariu
sM
AR
4Y
EE
thio
pia
Mas
ha
07.8
7N
,35
.48
EJX
0121
59N
MP
9217
7M
.cf
.ar
enar
ius
MA
R5
YE
Yem
enR
iqab
14.8
7N
,43
.42
EJX
0121
60N
MP
pb31
28M
.nat
alen
sis
MN
A1
Sou
thA
fric
aS
teen
kam
pskr
aal
AY
1694
67A
pple
ton
etal
.(2
004)
M.n
atal
ensi
sM
NA
2S
outh
Afr
ica
Su
dwal
aA
Y16
9466
App
leto
net
al.
(200
4)M
.oce
anen
sis
MO
CA
ust
rali
aN
owa
Now
aA
Y16
9436
App
leto
net
al.
(200
4)M
.ori
anae
MO
R1
Au
stra
lia
Dar
win
AY
1694
37A
pple
ton
etal
.(2
004)
M.o
rian
aeM
OR
2A
ust
rali
aK
imbe
rley
Ran
ges
AY
1694
38A
pple
ton
etal
.(2
004)
M.p
ropr
itri
stis
MP
RP
apu
aN
ewG
uin
eaW
aro
AY
1694
40A
pple
ton
etal
.(2
004)
M.
sp.
MS
P1
Pap
ua
New
Gu
inea
War
oA
Y16
9439
App
leto
net
al.
(200
4)M
.sp
.M
SP
2P
hil
ippi
nes
Neg
ros
Isla
nd
AY
1694
51A
pple
ton
etal
.(2
004)
M.
sp.
MS
P3
Sol
omon
Isla
nds
San
taIs
abel
AY
1694
59A
pple
ton
etal
.(2
004)
M.
sp.
MS
P4
Sol
omon
Isla
nds
San
taIs
abel
AY
1694
60A
pple
ton
etal
.(2
004)
M.
sp.
MS
P5
Sol
omon
Isla
nds
San
taIs
abel
AY
1694
61A
pple
ton
etal
.(2
004)
M.
sp.
MS
P6
Tan
zan
iaG
onja
For
est
Res
erve
AY
1694
62A
pple
ton
etal
.(2
004)
M.
sp.
MS
P7
Tan
zan
iaU
sam
bara
Mou
nta
ins
AY
1694
63A
pple
ton
etal
.(2
004)
M.t
rist
isM
TR
1P
hil
ippi
nes
Ley
teIs
lan
dA
Y16
9471
App
leto
net
al.
(200
4)M
.tri
stis
MT
R2
Ph
ilip
pin
esN
egro
sIs
lan
dA
Y16
9470
App
leto
net
al.
(200
4)M
yoti
sm
uri
cola
AY
5045
66W
orth
ingt
onW
ilm
eret
al.
(un
publ
.)C
hal
inol
obu
sm
orio
AY
1694
72A
pple
ton
etal
.(2
004)
Ch
alin
olob
us
nig
rogr
iseu
sA
Y50
4561
Wor
thin
gton
Wil
mer
etal
.(u
npu
bl.)
Ch
alin
olob
us
tube
rcu
latu
sA
F32
1051
Lin
&P
enn
y(2
001)
NM
P=
Nat
ion
alM
use
um
inP
ragu
e,C
zech
Rep
ubl
ic.
170 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190
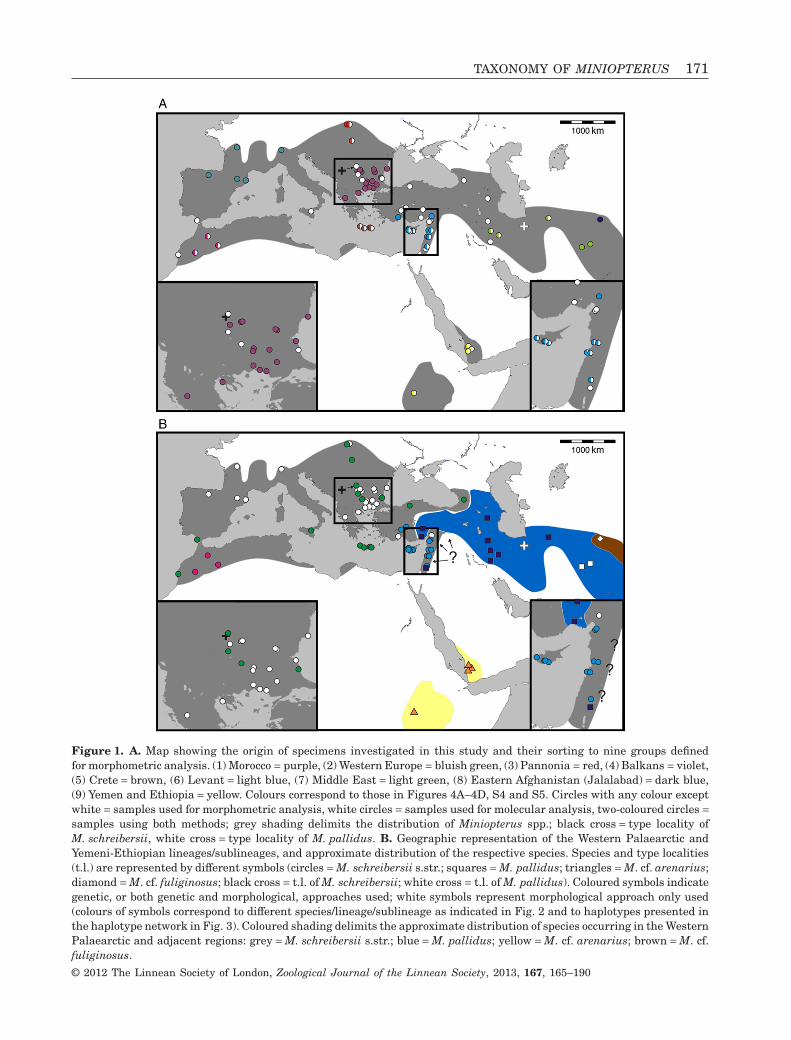
Figure 1. A. Map showing the origin of specimens investigated in this study and their sorting to nine groups definedfor morphometric analysis. (1) Morocco = purple, (2) Western Europe = bluish green, (3) Pannonia = red, (4) Balkans = violet,(5) Crete = brown, (6) Levant = light blue, (7) Middle East = light green, (8) Eastern Afghanistan (Jalalabad) = dark blue,(9) Yemen and Ethiopia = yellow. Colours correspond to those in Figures 4A–4D, S4 and S5. Circles with any colour exceptwhite = samples used for morphometric analysis, white circles = samples used for molecular analysis, two-coloured circles =samples using both methods; grey shading delimits the distribution of Miniopterus spp.; black cross = type locality ofM. schreibersii, white cross = type locality of M. pallidus. B. Geographic representation of the Western Palaearctic andYemeni-Ethiopian lineages/sublineages, and approximate distribution of the respective species. Species and type localities(t.l.) are represented by different symbols (circles = M. schreibersii s.str.; squares = M. pallidus; triangles = M. cf. arenarius;diamond = M. cf. fuliginosus; black cross = t.l. of M. schreibersii; white cross = t.l. of M. pallidus). Coloured symbols indicategenetic, or both genetic and morphological, approaches used; white symbols represent morphological approach only used(colours of symbols correspond to different species/lineage/sublineage as indicated in Fig. 2 and to haplotypes presented inthe haplotype network in Fig. 3). Coloured shading delimits the approximate distribution of species occurring in the WesternPalaearctic and adjacent regions: grey = M. schreibersii s.str.; blue = M. pallidus; yellow = M. cf. arenarius; brown = M. cf.fuliginosus.
TAXONOMY OF MINIOPTERUS 171
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

and parameter and tree samples saved every 100generations. A 50% majority-rule consensus tree wasconstructed from the sampled trees after discardingthe first 20 000 (two million generations) as burn-in,which included samples before attainment of thestationarity plateau on the log-likelihood score plotaccording to Tracer 1.4 (Drummond & Rambaut,2007). Posterior probabilities were calculated as thefrequency of samples recovering any particular clade(Huelsenbeck & Ronquist, 2001). The NJ tree wasinferred using PAUP* 4.0b10 (Swofford, 2003), basedon uncorrected p-distances and calculating the boot-strap branch support based on 1000 resampled data-sets. Haplotype networks were prepared using thestatistical parsimony algorithm implemented in TCS1.21 (Clement, Posada & Crandall, 2000) under a 95%limit of parsimony. Based on the results obtainedthrough phylogenetic analysis, ingroup haplotypes forbent-winged bat populations were divided into sevensets and average uncorrected p-distances betweenthem calculated using MEGA 4.0 (Tamura et al.,2007). In the text, we always refer to uncorrectedp-distances as they are easily comparable with mostprevious molecular-taxonomic studies.
MORPHOLOGICAL ANALYSIS
Morphometric analysis was based on skull traits. Thecomplete set of specimens was divided into ninegroups (Fig. 1A) based on the results of both pub-lished (Appleton et al., 2004; Bilgin et al., 2006, 2008;Furman et al., 2009, 2010b, c) and our own preli-minary genetic analysis, geographic origin of thesamples, and obvious differences observed in biomet-ric data (cranial and dental metrics and non-metrictraits): viz. (1) Morocco – specimens from the AtlasMountains (N = 18); (2) Western Europe – specimensfrom Spain, France, Italy, and Austria (N = 37); (3)Pannonia – specimens from Slovakia and Romania(N = 49); (4) Balkans – specimens from Bulgariaand continental Greece (N = 84); (5) Crete (N = 19); (6)Levant – specimens from southern Turkey, westernSyria, Cyprus, and Lebanon (N = 93); (7) Middle East– specimens from Iran and southern Afghanistan(N = 14); (8) Eastern Afghanistan (Jalalabad area)(N = 28); and (9) Yemen and Ethiopia – includingone specimen from Sudan (N = 11). Only individualsmorphologically assignable to M. schreibersii s.l. wereincluded. Group 6 (Levant) contained some samplesfrom the area of the zone of sympatry of M. schreib-ersii and M. pallidus. These samples were classifiedaccording to the prevailing genotypes present in theplace of origin. Explanation of some geographic termsused in this study (considering the grouping of theexamined material): Levant = western Syria andLebanon, but including southern Turkey and Cyprus;
Middle East = central and eastern Turkey, southernAzerbaijan, Iran, southern Afghanistan and north-western Jordan. The Statistica 6.0 software package(StatSoft) was used for all morphological statisticalanalyses.
Linear morphometricsWe recorded 24 cranio-dental measurements (11 skullor mandible measurements and 13 upper or lowertooth-row dimensions) taken using a digital calliper(by JŠ) to the nearest 0.01 mm (Fig. S1). Further, werecorded 57 dental measurements (width, length andhigh dimensions of respective teeth; Fig. S2) to thenearest 0.0125 mm using an optical calliper (for acomplete list of all measurements, see Appendix S1).
Basic descriptive statistical parameters (mean,minimum, maximum, and standard deviation) werecalculated for each measurement and for each group(1–9). We further used the morphometric data toperform factor (FA) and discriminant function (DFA)analyses to test: (1) grouping and/or separation of theabove groups (1–9); (2) similarity/dissimilarity of par-ticular populations/taxa; and (3) the importance ofparticular dimensions for intrageneric, inter-specificand intraspecific variation and differentiation. FA andDFA were first performed on samples from all groups(1–9), and subsequently on groups 1–7, in order tobetter demonstrate differences between geographi-cally and morphologically close populations. Cranialand dental characteristics were divided into six data-sets (maxillary, mandibular, cranial, cranio-dental,all dental, and all cranial characteristics) and thesetested separately in consecutive FAs and DFAs toassess the influence of different character sets on thegrouping/separation of individuals in morphospace.The FA and DFA canonical scores were plotted toshow relationships among the examined groups ofsamples. Morphological data of the Balkan (withaddition of genotyped samples of the Levantinegroup) and Middle Eastern sample sets were analysedby analysis of variance (ANOVA).
Geometric morphometrics and non-metric traitsGeometric morphometrics were used to analysegeographic variation in Miniopterus skulls and man-dibles. This technique has been demonstrated to beboth objective and efficient compared to traditionalmethods (e.g. Zelditch, Fink & Swiderski, 1995; Rohlf,Loy & Corti, 1996; Rohlf, 1998), including in studieson bats (e.g. Velazco, Gardner & Patterson, 2010). Thesame material was used as for linear morphometrics(Appendix 1).
Images of skulls (lateral, ventral and dorsal view),mandibles (lateral and occlusal view) and dentition(details of the upper and lower tooth-row) weretaken with a digital camera, archived (jpeg format;
172 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

1360 ¥ 1200 pixels resolution), and processed usingQuickPhoto 4.1 software (Promicra, Prague). Imagesof mandibles were taken separately. All images weretaken at an identical angle. Images of skulls andmandibles were converted to thin-plate spline format(tps) using tpsUtil 1.46 software (Rohlf, 2010).Homologous and topologically equivalent landmarkswere plotted on the skull (lateral, dorsal, and ventralviews) and mandible (lateral view) images using thetpsDig 1.40 program (Rohlf, 2004) in order to describesize and shape variation (for landmark definitions seeAppendix S1).
Landmark coordinates were converted into millime-tres using an established conversion factor (pixel/mm)and the original scale. The centroid size (CS) scoresof all view types for each specimen (CS1 – lateralview of mandible, CS2 – lateral view of skull, CS3 –ventral view of skull, and CS4 – dorsal view of skull)were calculated using the tpsRegr 1.36 program(Rohlf, 2009), and subsequently plotted to show sizedifferences between the groups examined. In orderto compare the shape of specimens from differentgroups, the coordinates for each specimen werescaled, aligned and transformed by general procrustesalignment (which generates a consensus configura-tion based on the landmark coordinates of all speci-mens) using the tpsRelw 1.46 software package(Rohlf, 2008) with a = 0, and orthogonal projectionand uniform component included. Shape differencesbetween the consensus landmark configuration andeach individual specimen were obtained and used tocompute a matrix of partial warp (PW) scores. Rela-tive warp (RW) scores were computed over the cov-ariance matrix of the PW scores; these are, therefore,analogous to a principal components analysis (PCA)in the sense that they describe the axes of greatestvariation in shape for all specimens investigated. ThePW matrix was used in a DFA to describe differencesbetween the studied groups and to confirm patternspreviously suggested by the RW scores. The scoresfrom canonical variant 1 of the DFA (of partial matrixdata) and the CS of skull and mandible were plottedin order to visualise and evaluate how size and shapecontributed to the arrangement of these groups. Dataobtained by geometric morphometrics (RW scores) ofthe Balkan (with addition the genotyped Levantinesamples) and Middle Eastern sample sets were ana-lysed by ANOVA.
The status of 49 non-metric cranial and dentalcharacteristics (44 dental and five skull or mandible;see Table S1) were investigated based on images ofskulls, mandibles and dentition. Each characteristicwas evaluated based on a pre-defined scale system1–5 (see Fig. S3). Non-metric data were analysed inthe same way as the linear metric data (basic descrip-tive statistics, FA and DFA computed).
RESULTS
MOLECULAR ANALYSIS
Eighty-nine haplotypes were registered from 111shortened (1034 bp) sequences (including GenBankand outgroup sequences) of the mitochondrial ND2gene. Within this haplotype dataset, 591 characterswere variable and 527 parsimony-informative. Topolo-gies from all analyses performed (ML, BA, NJ),as well as the log likelihood values (lnL), weresimilar (Fig. 2; ML lnL = -10614.95; BA meanlnL = -10316.71). Three well supported Miniopterusbat clades were identified within the completedata set: (A) an Australian-Oriental clade (Australa-sian, Oriental and Eastern-Palaearctic regions) inthe basal position; (B) an Afro-Arabian clade (sub-Saharan Africa and south-western Arabia); and (C) aWest Palaearctic clade (Europe, North Africa, AsiaMinor and the Middle East). The Western Palaearcticclade could be further divided into three well sepa-rated lineages: a Middle Eastern lineage (ME) (Iran,inland Turkey, Azerbaijan, Jordan) in the basal posi-tion, differing by 4.3 and 5.4% from the remaining twolineages; a Moroccan lineage (MO) from the AtlasMts.; and a Mediterranean lineage (M) identified fromSpain, Sicily, Slovakia, Romania, Bulgaria, Crete, theAtlantic coast of Morocco, the eastern Mediterraneanregion (southern Turkey, Cyprus, Syria, Lebanon),and the Black Sea region (northern Turkey, Georgia).The latter two lineages differed from each other by2.5%. Within the third lineage, we detected a furthersubdivision into two seemingly parapatric subline-ages with 1.2% divergence: a West Mediterraneansublineage (WM) from Europe, the Atlantic coastof Morocco, and the Black Sea region; and an EastMediterranean sublineage (EM) from southernTurkey, Cyprus, Syria and Lebanon. The mutual rela-tionships between the West Palaearctic clade samplesare also demonstrated through the parsimony haplo-type network (Fig. 3). All the above-mentionedMiniopterus clades/lineages/sublineages were highlysupported by ML bootstrap values (� 80%), NJ boot-strap values (� 85%), and BA posterior probabilities(� 0.98), except for clade A and lineage MO by MLbootstrap (62% and 71%), and sublineage WM by BAposterior probabilities (0.81).
All samples from south-western Arabia and Ethio-pia, border areas of the Palaearctic and Afro-tropicregions, were embedded within clade B, where theyformed two lineages (Fig. 2), one represented by amorphologically distinct Afro-tropic species, M. afri-canus Sanborn, 1936, collected from Ethiopia (andinitially used as an outgroup species); and the otherformed by individuals from Yemen and westernEthiopia [hereafter known as the Yemeni-Ethiopianlineage/group (YE)]. Representatives of the YE
TAXONOMY OF MINIOPTERUS 173
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190
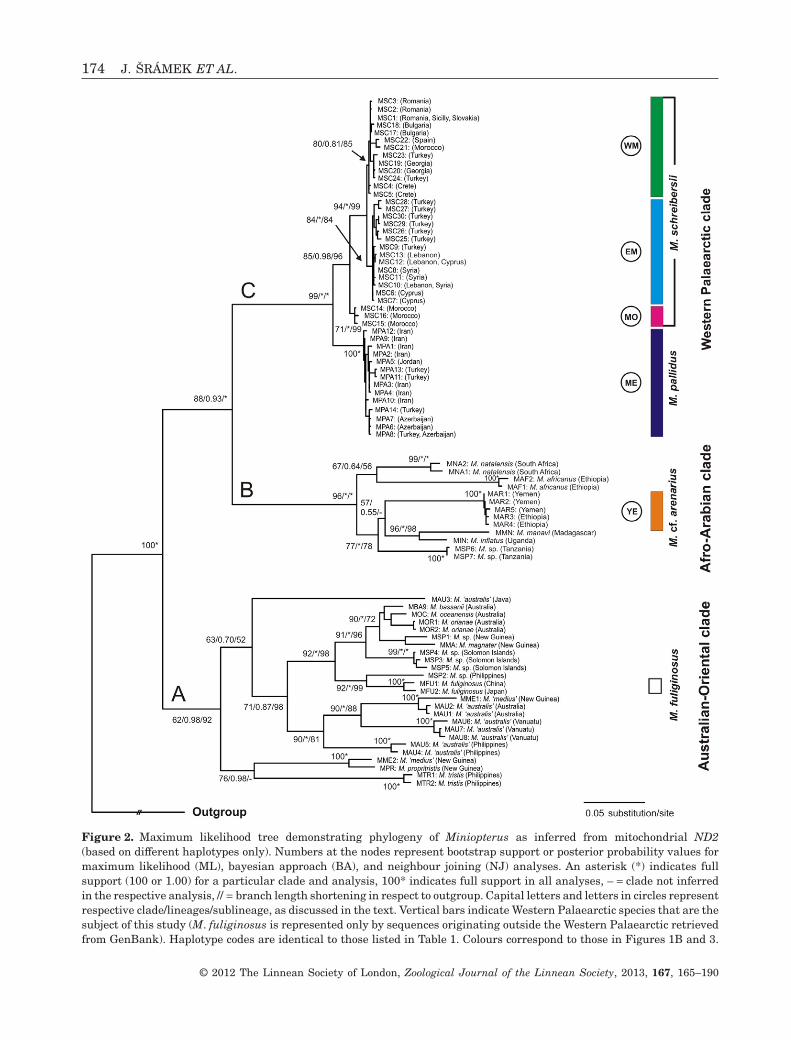
Figure 2. Maximum likelihood tree demonstrating phylogeny of Miniopterus as inferred from mitochondrial ND2(based on different haplotypes only). Numbers at the nodes represent bootstrap support or posterior probability values formaximum likelihood (ML), bayesian approach (BA), and neighbour joining (NJ) analyses. An asterisk (*) indicates fullsupport (100 or 1.00) for a particular clade and analysis, 100* indicates full support in all analyses, – = clade not inferredin the respective analysis, // = branch length shortening in respect to outgroup. Capital letters and letters in circles representrespective clade/lineages/sublineage, as discussed in the text. Vertical bars indicate Western Palaearctic species that are thesubject of this study (M. fuliginosus is represented only by sequences originating outside the Western Palaearctic retrievedfrom GenBank). Haplotype codes are identical to those listed in Table 1. Colours correspond to those in Figures 1B and 3.
174 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

lineage morphologically resemble M. cf. schreibersiibut have been recently assigned to M. natalensis(e.g. Simmons, 2005). The latter lineage, however,was differentiated by 11.5% from populations inSouth Africa, where the type locality of M. natalensisis registered. Moreover, the South African haplotypeswere not in sister position to the YE lineage, withother species, such as M. manavi Thomas, 1906; M.inflatus Thomas, 1903; and Miniopterus sp. from Tan-zania, being interspersed.
Genetic distances within and between selectedpopulations/taxa are presented in Table 2; while ageographic representation of the Western Palaearcticand YE lineages/sublineages, and the approximatedistribution of the respective species, is presented inFigure 1B.
MORPHOLOGICAL ANALYSIS
Linear morphometricsAll CS values, and all cranial and dental measure-ments for the nine different groups and their simple
comparisons (Table 3; Tables S2 and S3), indicatedthe same size patterns. Bats from eastern Afghani-stan (Jalalabad area) were markedly larger in com-parison to European and Levantine samples (sizedifferences between the latter two bat groups werevery small). Specimens from Crete, Yemen and Ethio-pia were clearly the smallest; while samples from theMiddle East were slightly smaller than those fromeastern Afghanistan, but markedly larger than batsfrom Europe and the Levant, and similar in size toMoroccan bats. Both skull and dentition shape differ-ences (expressed by ratios of cranial or dental dimen-sions) were much less expressive than differences ingeneral size. This pattern was more pronounced incranial than dental characteristics.
The results of FA and DFA analysis of skull anddental dimensions generally showed similar follow-ing patterns (results of FA not shown; for DFA seeFig. 4A, B; and Fig. S4), as did the comparison of rawskull and tooth dimensions and their ratios: (1)samples from eastern Afghanistan, the YE group and
Figure 3. Graphic illustration of relationships between ND2 haplotypes of M. schreibersii s.str. (green, light blue, purple)and M. pallidus (dark blue), as inferred through the maximum parsimony network approach. Size of circles correspondsto the number of samples within a particular haplotype (1, 2, 4 or 7 samples). Small dots between haplotypes indicatehypothetical haplotypes (or number of substitutions between them). Geographic abbreviations: MO – Morocco, Agadir(coast); MA – Morocco, Atlas Mts.; ES – Spain; SI – Sicily (Italy); SK – Slovakia; RO – Romania; BG – Bulgaria;CR – Crete (Greece); CY – Cyprus; NT – northern Turkey; ST – southern Turkey; GE – Georgia; AZ – Azerbaijan;LB – Lebanon; SY – Syria; JO – Jordan; IR – Iran. Colours correspond to those in Figures 1B and 2. Haplotype codesare identical to those listed in Table 1.
TAXONOMY OF MINIOPTERUS 175
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Morocco clustered separately from the European,Levantine and Middle Eastern groups, however, incase of Morocco markedly less distinct; (2) samplesfrom the Balkans and Pannonia formed a commoncluster, as did samples from Crete and the Levant; (3)Middle Eastern samples overlapped substantiallywith the West European and Levantine samples, andwere positioned close to the Pannonian and Balkansamples; (4) samples from western Europe weregrouped together with other groups from Europe, theMiddle East and the Levant (based on cranial dimen-sions), and very closely with samples from the Levant(based solely on dental traits). Using factor loadingvalues, we were able to identify the 10 cranial and 13dental dimensions that affected observed variationmost significantly (DFA, P < 0.0001), i.e. LaZ, LaInf,LaM, ACr, ACo, CC, M3M3, I1M3, CM3, M1M3; and LI2,WCsup, WP2, WP4, LiM1, W2M1, LiM2, W3M2, LI2, LI3,LDinf, WDinf and LP2, respectively. For a description ofmorphometric differentiation between the Miniopt-erus groups examined, see Appendix S2.
Results of ANOVA (Table S4) showed significantdifferences in 34 of 85 of the examined characteristics(mainly cranial) between the Balkan (with additionof the genotyped Levantine samples) and MiddleEastern samples.
Geometric morphometrics and non-metric traitsTwenty-two RWs were generated for the lateral skullview, 18 for the ventral view, 14 for the dorsal view,and 14 for the lateral view of the mandible. The firstfour RWs, which together represented more than 50%of total variation for each view, were used in allsubsequent analyses (Table S5).
Results of PCA and DFA demonstrated a number ofdifferences between the sample sets examined, andparticularly in the lateral view of the skull; however,neither PCA nor DFA were able to demonstrate anyclear separation between most of the groups exam-ined (results of PCA not shown; for DFA see Fig. 4C),with the European and Middle Eastern samples inparticular frequently showing a substantial overlap.Nevertheless, distinctive separations were observedin the samples from eastern Afghanistan when plot-ting the first two DFA canonical variables consideringall views of skull (Fig. 4C); the YE group consideringthe dorsal and ventral views of skull; the Moroccansamples in the skull ventral view; and in the Panno-nian samples for the skull lateral view. While therelationships between the groups differed for indi-vidual views, some general patterns were observable:(1) Pannonian and Levantine samples were distinctfrom each other; (2) samples from Crete were mostlysimilar to those from the Levant; and (3) MiddleEastern samples were mostly grouped together withsamples from Western Europe. In general, all analy-T
able
2.U
nco
rrec
ted
p-di
stan
ces
betw
een
and
wit
hin
diff
eren
tsp
ecie
s/(s
ub)
lin
eage
sof
Min
iopt
eru
sba
sed
onth
eh
aplo
type
data
set
(in
perc
enta
ge;
mea
nin
pare
nth
eses
)
M.s
chre
iber
sii,
Wes
tern
-M
edit
erra
nea
nsu
blin
eage
(WM
)
M.s
chre
iber
sii,
Eas
tern
-M
edit
erra
nea
nsu
blin
eage
(EM
)
M.s
chre
iber
sii,
Mor
occa
nli
nea
ge(M
O)
M.p
alli
du
s,M
iddl
eE
aste
rnli
nea
ge(M
E)
M.c
f.ar
enar
ius,
Yem
eni-
Eth
iopi
anli
nea
ge(Y
E)
M.a
fric
anu
s,E
thio
pia
M.n
atal
ensi
s,S
outh
Afr
ica
M.s
chre
iber
sii,
Wes
tern
-Med
iter
ran
ean
subl
inea
ge(W
M)
0.1–
1.4
(0.6
)
M.s
chre
iber
sii,
Eas
tern
-Med
iter
ran
ean
subl
inea
ge(E
M)
0.8–
2.2
(1.2
)0.
1–1.
1(0
.5)
M.s
chre
iber
sii,
Mor
occa
nli
nea
ge(M
O)
2.0–
3.0
(2.4
)2.
2–3.
1(2
.5)
0.3–
0.5
(0.4
)M
.pal
lid
us,
Mid
dle
Eas
tern
lin
eage
(ME
)4.
9–6.
3(5
.4)
4.7–
5.9
(5.3
)3.
9–4–
7(4
.3)
0.1–
1.1
(0.6
)
M.
cf.
aren
ariu
s,Ye
men
i-E
thio
pian
lin
eage
(YE
)14
.1–1
5.1
(14.
6)13
.9–1
5.0
(14.
4)14
.2–1
4.9
(14.
5)14
.8–1
5.8
(15.
2)0.
1–0.
5(0
.4)
M.a
fric
anu
s,E
thio
pia
15.5
–16.
1(1
5.7)
15.5
–16.
0(1
5.8)
15.0
–15.
7(1
5.3)
15.1
–16.
2(1
5.6)
12.8
–13.
4(1
3.1)
1.0
M.n
atal
ensi
s,S
outh
Afr
ica
12.8
–13.
6(1
3.2)
12.8
–13.
9(1
3.2)
12.9
–13.
3(1
3.2)
14.0
–15.
0(1
4.5)
11.2
–11.
8(1
1.5)
10.6
–10.
9(1
0.8)
1.5
176 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Tab
le3.
Sel
ecte
dcr
ania
lan
dde
nta
ldi
men
sion
s(i
nm
m)
ofM
inio
pter
us
exam
ined
inth
isst
udy
Ch
arac
ter
Mor
occo
(MO
)W
este
rnE
uro
pe(W
M)
Pan
non
ia(W
M)
Bal
kan
s(W
M)
Cre
te(W
M)
NM
min
max
SD
NM
min
max
SD
NM
min
max
SD
NM
min
max
SD
NM
min
max
SD
LC
r18
15.3
6515
.12
15.6
50.
164
3615
.229
14.8
015
.49
0.17
345
15.3
9214
.94
15.8
80.
206
7615
.229
14.5
415
.83
0.21
619
14.9
1614
.48
15.1
90.
185
LaI
183.
777
3.63
3.98
0.08
936
3.71
13.
573.
880.
071
473.
761
3.53
4.02
0.10
882
3.71
03.
523.
930.
087
193.
621
3.48
3.77
0.08
4L
aIn
f18
4.20
24.
024.
320.
086
364.
023
3.74
4.29
0.10
445
4.02
53.
854.
240.
088
803.
974
3.45
4.18
0.13
619
3.89
43.
784.
050.
091
LaM
188.
788
8.64
8.97
0.10
535
8.67
58.
108.
990.
190
468.
757
8.10
9.21
0.18
076
8.74
68.
129.
040.
155
198.
524
8.40
8.67
0.09
1A
Cr
187.
973
7.73
8.34
0.18
530
7.49
76.
668.
010.
420
437.
672
6.79
8.04
0.32
577
7.74
36.
838.
210.
324
197.
765
7.36
7.99
0.15
4L
Md
1810
.977
10.8
011
.28
0.14
436
10.8
5210
.56
11.1
50.
143
4810
.923
10.2
811
.30
0.17
575
10.8
4710
.15
11.0
70.
154
1910
.622
10.3
210
.88
0.11
8A
Co
182.
598
2.48
2.81
0.07
736
2.59
42.
402.
930.
109
482.
533
2.17
2.93
0.12
075
2.50
82.
042.
930.
109
192.
538
2.41
2.69
0.07
6C
S1
1811
.168
10.8
411
.52
0.18
934
11.1
3010
.67
11.9
70.
298
4411
.431
10.9
412
.03
0.27
866
11.1
0310
.64
11.9
00.
261
1910
.730
10.1
511
.02
0.21
1C
S2
1820
.813
20.3
921
.21
0.22
832
20.4
7820
.15
20.8
90.
199
4220
.659
20.0
421
.19
0.27
668
20.5
0219
.87
21.1
10.
261
1920
.163
19.7
220
.59
0.20
5C
S3
1816
.935
16.7
317
.33
0.15
932
16.7
6016
.37
17.0
90.
173
4016
.738
16.3
017
.43
0.25
073
16.7
5616
.23
17.1
90.
186
1916
.394
16.0
516
.75
0.17
3C
S4
1817
.716
17.4
418
.03
0.19
334
17.5
1517
.18
17.9
30.
194
3717
.762
17.1
818
.44
0.26
474
17.5
8017
.14
18.1
00.
204
1917
.179
16.6
417
.51
0.22
1L
Csu
p18
1.05
51.
001.
100.
031
281.
084
1.03
1.13
0.02
644
1.06
20.
981.
150.
036
811.
071
1.00
1.15
0.03
719
1.05
21.
001.
130.
031
WC
sup
180.
883
0.81
0.93
0.03
528
0.89
60.
810.
980.
032
440.
840
0.78
0.90
0.03
081
0.83
30.
760.
930.
033
190.
825
0.78
0.85
0.02
4H
Csu
p4
1.65
31.
631.
710.
041
211.
573
1.45
1.93
0.10
629
1.54
71.
331.
690.
074
561.
550
1.15
1.70
0.09
215
1.52
01.
301.
630.
097
LP
218
0.84
10.
750.
900.
046
360.
816
0.78
0.88
0.02
748
0.82
90.
730.
930.
047
820.
829
0.78
0.94
0.03
619
0.81
80.
780.
880.
025
WP
218
1.13
11.
031.
200.
048
361.
138
1.08
1.19
0.03
148
1.08
70.
901.
160.
051
821.
094
1.00
1.20
0.04
019
1.08
31.
041.
130.
027
HP
24
0.58
40.
550.
630.
031
250.
568
0.48
0.65
0.04
733
0.53
50.
430.
630.
049
580.
561
0.43
0.65
0.05
315
0.52
10.
450.
600.
045
WP
418
1.39
01.
311.
460.
041
361.
444
1.38
1.51
0.03
248
1.32
81.
151.
480.
083
821.
377
1.23
1.50
0.05
319
1.38
91.
261.
450.
046
LP
418
1.22
01.
101.
380.
068
361.
283
1.18
1.35
0.04
248
1.23
11.
131.
380.
066
821.
231
1.13
1.38
0.05
719
1.18
11.
151.
230.
023
HP
44
1.56
91.
481.
650.
072
251.
586
1.44
1.71
0.06
533
1.49
71.
351.
600.
071
581.
543
1.28
1.68
0.07
115
1.40
70.
831.
650.
279
LoM
118
1.43
81.
351.
500.
039
351.
473
1.43
1.55
0.03
448
1.44
71.
351.
530.
039
831.
452
1.24
1.55
0.04
319
1.43
91.
401.
480.
021
LiM
118
0.92
40.
881.
030.
035
350.
983
0.93
1.05
0.03
148
0.94
70.
831.
040.
049
830.
984
0.85
1.09
0.04
119
0.97
00.
901.
030.
036
LC
inf
180.
726
0.70
0.76
0.02
031
0.75
40.
700.
840.
031
450.
736
0.63
0.85
0.04
775
0.72
30.
640.
790.
029
190.
717
0.68
0.78
0.02
1W
Cin
f18
0.82
90.
780.
880.
032
310.
821
0.79
0.86
0.02
245
0.79
90.
740.
860.
024
750.
804
0.68
0.85
0.02
719
0.78
90.
780.
810.
013
HC
inf
41.
503
1.49
1.53
0.01
621
1.39
41.
031.
500.
108
301.
435
1.30
1.56
0.06
351
1.43
51.
251.
630.
064
151.
424
1.30
1.50
0.05
8L
P2
180.
594
0.55
0.64
0.02
328
0.58
70.
550.
630.
021
460.
567
0.53
0.60
0.02
275
0.56
60.
530.
630.
022
190.
551
0.53
0.58
0.02
0W
P2
180.
651
0.63
0.68
0.01
728
0.62
60.
590.
650.
017
460.
645
0.60
0.71
0.02
875
0.64
30.
530.
690.
027
190.
626
0.59
0.68
0.02
0H
P2
40.
575
0.53
0.63
0.04
620
0.51
90.
450.
630.
045
310.
509
0.40
0.61
0.05
151
0.50
40.
450.
600.
028
150.
481
0.43
0.52
0.02
8W
P4
180.
791
0.73
0.85
0.02
934
0.76
20.
700.
850.
030
480.
781
0.68
0.85
0.03
577
0.76
70.
630.
880.
040
190.
759
0.63
0.79
0.03
6L
P4
180.
607
0.55
0.68
0.03
734
0.61
30.
510.
750.
055
480.
639
0.54
0.71
0.04
176
0.60
30.
500.
780.
050
190.
605
0.54
0.68
0.03
9H
P4
40.
925
0.85
0.98
0.06
122
0.87
40.
580.
980.
076
330.
836
0.74
0.98
0.06
252
0.87
20.
731.
000.
050
150.
845
0.75
0.95
0.04
7L
M3
181.
219
1.15
1.29
0.03
636
1.27
31.
191.
350.
032
481.
253
1.19
1.33
0.03
278
1.25
41.
181.
330.
027
191.
235
1.15
1.31
0.03
5W
M3
180.
666
0.60
0.73
0.02
936
0.63
40.
600.
680.
019
480.
659
0.59
0.78
0.03
678
0.64
20.
600.
730.
024
190.
641
0.59
0.70
0.02
4
TAXONOMY OF MINIOPTERUS 177
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190
Tab
le3.
Con
tin
ued
Ch
arac
ter
Lev
ant
(EM
)M
iddl
eE
ast
(ME
)E
aste
rnA
fgh
anis
tan
Yem
en&
Eth
iopi
a(Y
E)
NM
min
max
SD
NM
min
max
SD
NM
min
max
SD
NM
min
max
SD
LC
r93
15.1
4814
.71
15.7
40.
217
1415
.497
15.1
115
.84
0.22
228
15.6
3615
.03
16.1
30.
254
1015
.058
14.7
115
.50
0.22
9L
aI93
3.67
53.
413.
970.
084
143.
686
3.55
3.88
0.09
628
3.92
83.
764.
150.
083
113.
711
3.58
3.96
0.10
8L
aIn
f93
3.98
73.
744.
180.
086
144.
039
3.94
4.18
0.08
028
4.12
43.
824.
430.
141
123.
712
3.46
3.87
0.11
5L
aM93
8.69
78.
279.
050.
142
148.
914
8.74
9.09
0.09
228
8.84
08.
519.
210.
168
108.
299
7.97
8.68
0.22
4A
Cr
907.
819
7.14
8.11
0.18
814
8.00
57.
768.
340.
162
287.
779
6.99
8.36
0.43
411
7.50
47.
137.
870.
215
LM
d91
10.7
6310
.32
11.1
70.
155
1411
.072
10.7
411
.23
0.14
928
11.3
0411
.00
11.6
00.
166
1110
.645
10.2
911
.04
0.20
8A
Co
912.
568
2.35
2.93
0.11
314
2.62
12.
482.
730.
074
282.
647
2.44
3.04
0.11
711
2.41
22.
242.
650.
115
CS
185
11.0
0510
.45
11.8
90.
312
1311
.153
10.9
011
.60
0.21
023
11.5
5111
.03
12.2
90.
291
1110
.677
10.3
910
.87
0.15
7C
S2
9120
.437
19.8
521
.37
0.29
114
20.8
3020
.17
21.3
90.
342
2821
.076
20.4
221
.74
0.31
110
20.0
7319
.52
20.7
30.
327
CS
392
16.6
2116
.13
17.3
90.
245
1417
.045
16.6
317
.42
0.24
328
17.2
2716
.80
17.7
00.
243
1016
.548
16.0
217
.06
0.28
5C
S4
9217
.321
16.5
617
.99
0.26
014
17.8
3817
.42
18.3
30.
306
2817
.902
17.2
518
.39
0.29
311
17.2
7816
.79
17.8
90.
302
LC
sup
921.
072
1.00
1.18
0.03
414
1.08
81.
051.
140.
030
261.
114
1.03
1.20
0.04
812
1.05
60.
961.
130.
042
WC
sup
920.
865
0.80
1.15
0.04
114
0.85
80.
810.
900.
025
260.
933
0.85
1.00
0.04
212
0.86
00.
800.
950.
036
HC
sup
571.
579
1.25
1.73
0.08
814
1.56
91.
301.
730.
130
261.
645
1.28
2.00
0.17
83
1.62
91.
601.
660.
031
LP
293
0.83
20.
780.
930.
034
140.
835
0.80
0.89
0.02
727
0.89
50.
791.
000.
055
120.
850
0.75
0.90
0.04
1W
P2
931.
124
0.91
1.25
0.05
114
1.11
81.
061.
160.
031
271.
081
0.90
1.19
0.06
112
1.02
80.
801.
150.
096
HP
258
0.58
00.
450.
700.
052
140.
542
0.25
0.68
0.10
727
0.55
60.
380.
700.
077
30.
596
0.58
0.64
0.03
6W
P4
931.
403
1.30
1.53
0.03
914
1.36
81.
291.
430.
046
271.
413
1.33
1.58
0.05
712
1.35
01.
261.
430.
060
LP
493
1.26
61.
081.
400.
064
141.
196
1.09
1.28
0.05
627
1.28
21.
231.
450.
057
121.
200
1.03
1.28
0.06
9H
P4
581.
514
0.88
1.70
0.15
314
1.41
10.
791.
710.
311
271.
531
1.35
1.75
0.10
43
1.53
31.
501.
600.
058
LoM
193
1.45
51.
311.
560.
035
141.
482
1.43
1.55
0.03
227
1.49
11.
381.
550.
050
121.
447
1.36
1.50
0.04
7L
iM1
930.
957
0.85
1.05
0.04
214
0.99
40.
951.
050.
034
270.
991
0.90
1.10
0.04
412
0.95
10.
881.
030.
040
LC
inf
890.
736
0.68
0.80
0.02
614
0.74
10.
700.
780.
027
260.
787
0.71
0.86
0.04
012
0.74
50.
680.
800.
033
WC
inf
890.
807
0.75
0.86
0.02
514
0.82
10.
780.
850.
027
260.
853
0.75
0.93
0.04
512
0.78
00.
730.
850.
033
HC
inf
551.
452
1.15
1.55
0.08
814
1.43
61.
251.
550.
084
251.
557
1.29
1.80
0.14
83
1.42
51.
381.
530.
087
LP
289
0.56
90.
530.
630.
022
140.
549
0.51
0.58
0.01
726
0.57
50.
530.
630.
032
120.
567
0.53
0.60
0.02
2W
P2
890.
627
0.58
0.68
0.02
114
0.63
40.
600.
660.
018
260.
651
0.61
0.68
0.01
912
0.61
00.
580.
650.
024
HP
255
0.49
00.
430.
550.
025
130.
468
0.25
0.55
0.07
926
0.47
20.
360.
550.
056
30.
463
0.45
0.48
0.01
3W
P4
900.
761
0.68
0.85
0.03
014
0.78
10.
700.
830.
035
270.
767
0.73
0.83
0.02
712
0.75
10.
710.
800.
022
LP
490
0.60
10.
500.
680.
037
140.
634
0.55
0.70
0.03
727
0.66
80.
580.
760.
058
120.
633
0.53
0.70
0.04
9H
P4
550.
883
0.75
0.95
0.05
014
0.91
80.
781.
000.
056
270.
879
0.75
1.03
0.07
63
0.90
00.
850.
980.
066
LM
390
1.24
91.
151.
330.
031
141.
257
1.20
1.31
0.03
327
1.26
81.
201.
310.
029
121.
221
1.15
1.28
0.03
4W
M3
900.
638
0.58
0.88
0.03
414
0.66
50.
630.
700.
023
270.
688
0.64
0.75
0.03
212
0.66
50.
600.
730.
034
Cod
esin
brac
kets
stan
dfo
rth
ere
spec
tive
gen
etic
lin
eage
orsu
blin
eage
(see
text
).S
eeA
ppen
dix
S1
for
expl
anat
ion
ofdi
men
sion
abbr
evia
tion
s.D
ata
for
all
dim
ensi
ons
are
pres
ente
din
Tabl
esS
2an
dS
3.M
=m
ean
,m
in=
min
imu
mva
lue,
max
=m
axim
um
valu
e,an
dS
D=
stan
dard
devi
atio
n.
178 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190
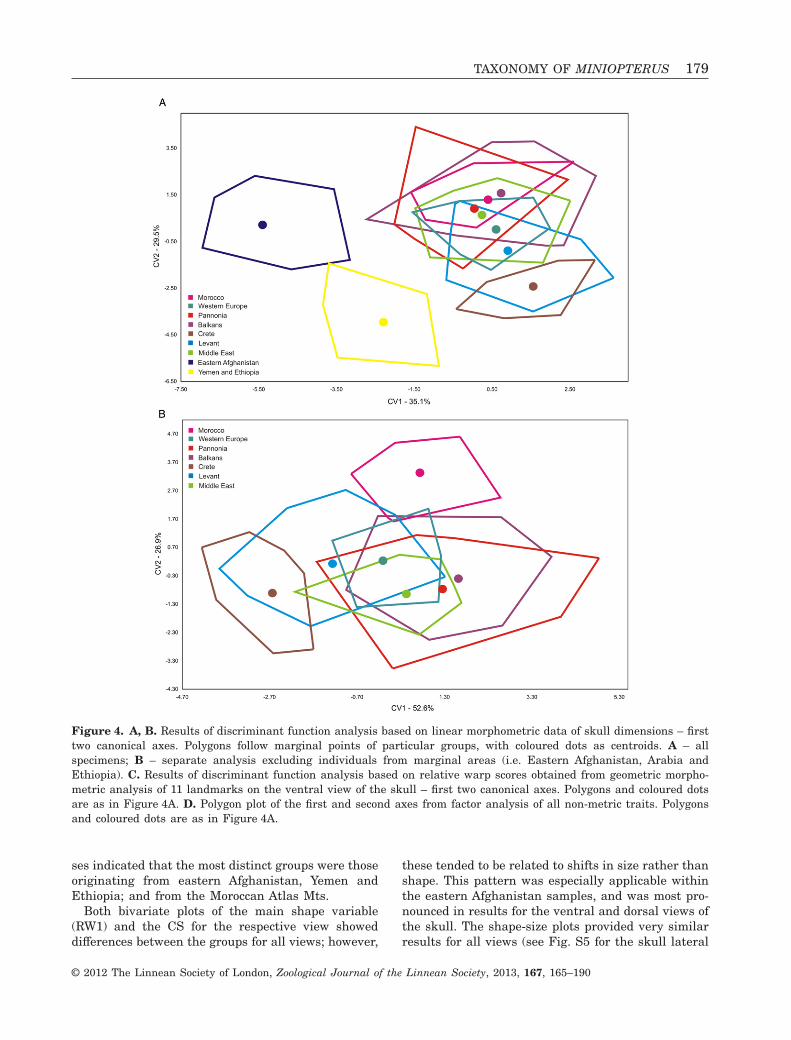
ses indicated that the most distinct groups were thoseoriginating from eastern Afghanistan, Yemen andEthiopia; and from the Moroccan Atlas Mts.
Both bivariate plots of the main shape variable(RW1) and the CS for the respective view showeddifferences between the groups for all views; however,
these tended to be related to shifts in size rather thanshape. This pattern was especially applicable withinthe eastern Afghanistan samples, and was most pro-nounced in results for the ventral and dorsal views ofthe skull. The shape-size plots provided very similarresults for all views (see Fig. S5 for the skull lateral
Figure 4. A, B. Results of discriminant function analysis based on linear morphometric data of skull dimensions – firsttwo canonical axes. Polygons follow marginal points of particular groups, with coloured dots as centroids. A – allspecimens; B – separate analysis excluding individuals from marginal areas (i.e. Eastern Afghanistan, Arabia andEthiopia). C. Results of discriminant function analysis based on relative warp scores obtained from geometric morpho-metric analysis of 11 landmarks on the ventral view of the skull – first two canonical axes. Polygons and coloured dotsare as in Figure 4A. D. Polygon plot of the first and second axes from factor analysis of all non-metric traits. Polygonsand coloured dots are as in Figure 4A.
TAXONOMY OF MINIOPTERUS 179
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190
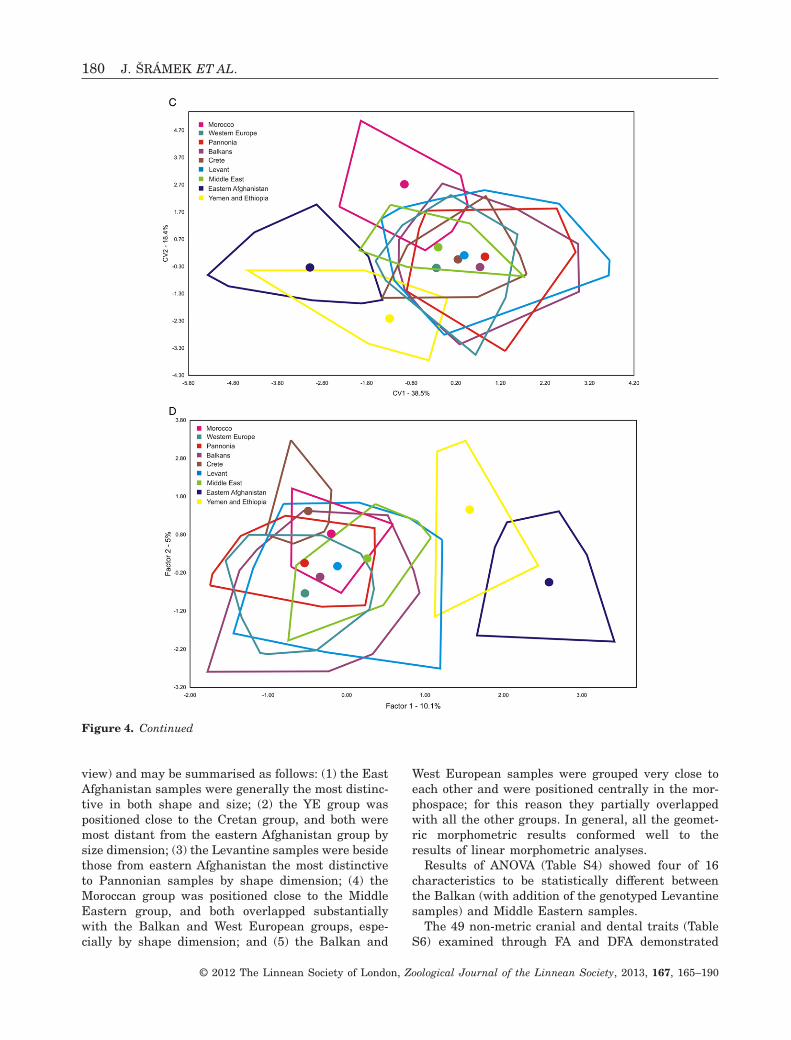
view) and may be summarised as follows: (1) the EastAfghanistan samples were generally the most distinc-tive in both shape and size; (2) the YE group waspositioned close to the Cretan group, and both weremost distant from the eastern Afghanistan group bysize dimension; (3) the Levantine samples were besidethose from eastern Afghanistan the most distinctiveto Pannonian samples by shape dimension; (4) theMoroccan group was positioned close to the MiddleEastern group, and both overlapped substantiallywith the Balkan and West European groups, espe-cially by shape dimension; and (5) the Balkan and
West European samples were grouped very close toeach other and were positioned centrally in the mor-phospace; for this reason they partially overlappedwith all the other groups. In general, all the geomet-ric morphometric results conformed well to theresults of linear morphometric analyses.
Results of ANOVA (Table S4) showed four of 16characteristics to be statistically different betweenthe Balkan (with addition of the genotyped Levantinesamples) and Middle Eastern samples.
The 49 non-metric cranial and dental traits (TableS6) examined through FA and DFA demonstrated
Figure 4. Continued
180 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

pronounced differentiation of the eastern Afghanistangroup from the other groups. Similar differentiationwas noted for the YE group, while all other groupsformed a cluster of broadly overlapping samples(for FA see Fig. 4D; results of DFA not shown).These analyses also enabled selection of maxilla char-acteristics that most affected observed variationin the non-metric traits (P < 0.0001 in DFAs). For adescription of non-metric differentiation between theMiniopterus groups see Appendix S2.
DISCUSSION
Revision of particularly Western Palaearctic bent-winged bat populations over their whole range (i.e.from the Maghreb to Afghanistan, and from CentralEurope to Arabia) revealed unexpected hidden diver-sity, even in the light of recent discoveries by Furmanet al. (2009, 2010c). Synthesis of the results from twodifferent analytical approaches suggests that M. sch-reibersii s.l. (sensu, e.g. Corbet, 1978), a traditionallypolytypic species, should be split into several allopat-ric or parapatric population groups, differing fromeach other in genetic and morphological traits. Thesegroups can be delimited geographically as follows: (1)Europe, northern Turkey and Georgia; (2) the Levant,including southern Turkey and Cyprus; (3) the moun-tains of Morocco; (4) the Middle East (except for theLevant and Turkish coastal areas); (5) south-westernArabia and Ethiopia; and (6) eastern Afghanistan(Jalalabad area). This order mirrors the degree ofrelatedness of the respective populations to those ofEurope (i.e. group two is closer related to the Euro-pean population than group three). Although the geo-graphical groupings may, at first, appear surprising,the findings are in general accordance with the opin-ions of earlier authors (namely Tate, 1941 and Maeda,1982) who stressed morphological similarities amongindividual species of the genus Miniopterus and pre-sumed the existence of more species, rather than asingle universal morphotype. These conclusions havealso recently gained support through several molecu-lar studies (Appleton et al., 2004; Tian et al., 2004;Miller-Butterworth et al., 2005; Furman et al., 2009,2010b, c), and we supplement these findings withadditional molecular phylogeny and new morphologi-cal evidence.
Our results indicate that, though the populationsdiffer only slightly in skull size, these differences weremore pronounced than differences in skull shape.These findings are in accordance with those of previ-ous authors (Tate, 1941; Maeda, 1982). Differences incranial measurements were also more expressivethan differences in dental measurements. Levels ofsignificance for differentiation between populationswere then followed by the results of non-metric
cranial and dental characteristics. Genetic differencesbetween populations were markedly more expressivefor some groups (i.e. the Middle East, Morocco, Yemenand Ethiopia) than differences observed for morpho-logical traits. No clinal shift in size or other morpho-logical data was found between populations, contraryto morphometric analysis results for other bat speciesoccurring in the Western Palaearctic (Hanák &Horácek, 1984; Bogdanowicz, 1990; Benda & Horácek,1995; Benda et al., 2006).
EUROPE AND EASTERN MEDITERRANEAN
European, Black Sea region and eastern Mediterra-nean Miniopterus populations evidently representidentical taxa that can be co-identified with thespecies M. schreibersii s.str. (sensu Furman et al.,2010c). Our results support the opinions of mostprevious authors (e.g. Spitzenberger, 1981; Crucitti,1989; Fernandez & Ibañez, 1989; Appleton et al.,2004; Boye, 2004; Gazaryan, 2005; Furman et al.,2009, 2010b, c) who suggest that all European popu-lations of M. schreibersii (in its traditional concept)belong to the nominotypical form.
Samples of the WM sublineage (that includessamples from Europe, coastal Morocco, coastal areasof northern Turkey and Georgia) showed relativelylow genetic variation (0.1–1.4%). Only one samplefrom Spain and two from the Atlantic coast of Moroccoshowed more differentiation from the cluster ofother European haplotypes (0.8–1.3%). This diver-gence could correspond to the ‘isolation by distance’model suggested for other bat species in the region(e.g. Pipistrellus – Hulva et al., 2004, 2007a, 2010). Inorder to confirm this genetic pattern, however, addi-tional samples from less distant localities of WesternEurope (e.g. France, the Italian peninsular, otherIberian samples) need to be studied. Morphometricdata did not show any clinal pattern, and onlyshallow morphological variation. Bent-winged batsfrom Crete represented the only exception, thesebeing significantly smaller. Ondrias (1978) andIliopoulou-Georgudaki (1986), who studied bats fromthe Greek islands, including Crete, found similarmorphological evidence and suggested that smallersize in island Miniopterus populations could haveresulted from climatic influence, i.e. strong winds.These authors, however, did not take account ofthe general ecological factors associated with islandbiogeography (MacArthur & Wilson, 1967), whichwe consider a more likely explanation for this mor-phological effect. Interestingly, genetic divergencebetween Cretan and mainland populations is minutecompared to morphometric divergence. Morphometricdifferentiation, however, appears to relate to skullsize rather than skull shape. This suggests that
TAXONOMY OF MINIOPTERUS 181
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

size-based morphological evidence does not correlatewith genetic evidence in Miniopterus bats, as hasbeen demonstrated for other bat groups (e.g. Hulva,Horácek & Benda, 2007b; Benda, Vallo & Reiter,2011).
Levantine populations (including Cyprus), forwhich taxonomic position has hitherto been unclear,belong to the same taxon as European populations,i.e. M. schreibersii s.str. However, they formed adistinct clade under mtDNA genealogy, the EM sub-lineage, which diverged by 0.8–2.2% from the Euro-pean and Black Sea region samples that formedtheir sister clade. Our genetic results thus supportthe opinions of Horácek et al. (2000) and Karatas &Sözen (2004), i.e. that the Mediterranean parts ofthe Levant are inhabited by the European form.When comparing Levantine bent-winged bats tothe population more to the east, i.e. M. pallidus (seebelow), there was a substantial divergence in genetictraits (5.3%) but, interestingly, almost no distinctionin cranial or dental morphology. It appears, there-fore, that both M. schreibersii and M. pallidus areconservative in their morphology, and especiallyin skull shape. A further interesting point is thatMiller-Butterworth et al. (2005) uncovered, andFurman et al. (2010b) consequently re-analysed,another genetic lineage from northern Israel(Alma Cave), based on the mitochondrial cytochromeb gene of the only sample. This was, however, com-pletely outside the species ranks of both M. schreib-ersii s.str. and M. pallidus (c. 6–8% genetic distance;cf. Miller-Butterworth et al., 2005; Furman et al.,2010b; see also the phylogenetic tree topology in thelatter paper). This lineage was most closely relatedto an Afro-tropical species, M. natalensis, and thussuggests the possible presence of the Arabian speciesM. cf. arenarius (see below) in southern Levant [ifthe sequence (AY614736) is correct – several ambi-guity codes are present]. A similar disjunct distri-bution can be seen in the Arabian tree frog, Hylafelixarabica (Gvoždík et al., 2010). Considering thehigh systematic and biogeographic importance ofthis possible Israeli lineage, it is necessary toconfirm the finding with more numerous samples infuture analyses. [Considering the apparent absenceof this lineage in our rich dataset, which coverssurrounding areas of the Levant (Lebanon, south-western Syria, north-western Jordan), a mislabellingof the sample would appear to be a more probableexplanation of this curiosity.]
MOROCCO
Our results show that a unique evolutionary lineageof M. schreibersii s.str. inhabits the Atlas Mountainsof Morocco. The inland Moroccan samples formed
a clade that diverged by 2.4–2.5% from the M lineage.Similar evidence was also provided by the morpho-metric analysis results. On the other hand, a pub-lished haplotype from Agadir (Atlantic coast ofsouth-west Morocco; Appleton et al., 2004) repre-sented part of the M lineage (WM sublineage). Thehaplotype most similar to this was one detected fromSpain and published by the same author (Appletonet al., 2004). Unfortunately, we were unable to obtainsamples from the Atlantic coast in order to investi-gate their morphological characteristics. Moroccansamples that separated into two genealogical lineageswere also found by Furman et al. (2010b), based onmitochondrial cytochrome b sequences taken fromGarcía-Mudarra, Ibáñez & Juste (2009), althoughexact locations of the samples were, unfortunately,not published. The available results, however, clearlysuggest that there are two distinct lineages presentin Morocco, the West Mediterranean M. schreibersii,occurring along the Atlantic coast, and an unnamedMoroccan form of M. schreibersii s.str., occurring ininland areas of the Atlas Mts. A similar geographicpattern of haplotype distribution was also docu-mented in Morocco for the vespertilionid bats Myotismystacinus (García-Mudarra et al., 2009) and Pipist-rellus pipistrellus (Hulva et al., 2010), as well as forthe freshwater terrapin Mauremys leprosa (Fritzet al., 2005). Miniopterus populations from the AtlasMts. may thus represent a separate taxon. This sug-gestion contradicts the traditional view on taxonomicaffiliation of Maghrebian populations, which are con-sidered to represent the nominotypical form by mostauthors (e.g. Ellerman & Morrison-Scott, 1951; Aellen& Strinati, 1970; Qumsiyeh & Schlitter, 1982; Gaisler,1983; Kowalski & Rzebik-Kowalska, 1991; Boye,2004). An in-depth study of the North African Mini-opterus population is necessary in order to reveal thephylogenetic and taxonomic position of the respectivesub-populations.
MIDDLE EAST
Miniopterus populations of the Middle East, includingthose of southern Afghanistan, Iran, Azerbaijan,inland plateau areas of central and eastern Turkey,and north-western Jordan, represent a further dis-tinct evolutionary lineage. This ME lineage, tradition-ally considered a subspecies M. schreibersii pallidus(e.g. Corbet, 1978; Koopman, 1994), displayed markedgenetic divergence (5.4% distance to the M lineageand 4.3% distance to the MO lineage). The 5% valueset as a species level indicator according to thegenetic species concept in mammals, and particularlyin bats, was, therefore, exceeded [the 5% value wasoriginally suggested for mitochondrial markerswith similar mutation rates, i.e. genes for cytochrome
182 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

b (Baker & Bradley, 2006) and ND1 (Mayer, Dietz &Kiefer, 2007)]. Cranial and dental morphologicaltraits investigated in this study as well as performedanalyses (FA, DFA, ANOVA), however, indicated thatindividuals of this group were in shape almost iden-tical to those from Europe and the Levant, whereas insize were slightly bigger. ANOVA results otherwiseshoved many significant mainly cranial size charac-teristics (in shape minimum) between the Balkan(with addition of genotyped samples of the Levant;M lineage; representing M. schreibersii species) andMiddle Eastern samples (ME lineage; representingpossible M. pallidus species), nevertheless, these dif-ferences were at the same level or even smaller thandifferences between populations of M. schreibersii (seeTable S4). Found significant morphological differencesbetween representative samples of M. schreibersiiand possible M. pallidus species in a way correspondto those found by Furman et al. (2010c) (and partlyby Bilgin et al., [2012]) between the Turkish inland(ME lineage) and Turkish coastal (M lineage) popu-lations, based on forearm length, body mass and wingshape. Any of these three characteristics (particularlyforearm length data – a possible important diagno-stic character [see Furman et al., 2010c]) were notanalysed in our study thus comparison with dataobtained by Furman et al. (2010c) and Bilgin et al.(2012) was not possible. Following a series of molecu-lar studies (Bilgin et al., 2006, 2008; Furman et al.,2009, 2010b), Furman et al. (2010c) suggested theraising of Middle Eastern bent-winged bats to specieslevel. Support for this came from Maraci et al. (2010)and Bilgin et al. (2012), who found the two taxa insympatry, and even in syntopy, in the same roosts.Considering all these and ours findings, we agree thatthe ME lineage represents a separate species, M. pal-lidus, a sister species to M. schreibersii s.str., thoughthis remains rather cryptic morphologically (i.e. noteasily distinguishable in the field).
Genetic comparison of the Al Wardeh Cave popula-tion from north-western Jordan indicates thatthis population belongs to M. pallidus. Up to now,however, representatives of this taxon are known onlyfrom the belt of mountainous habitats that stretchfrom central Turkey to Afghanistan. The record fromJordan, therefore, represents a significant extensionof this taxon’s range southward to the Levant. Unfor-tunately, we had an insufficient number of specimensto provide a well-founded morphological analysis ofthe Jordanian population. The Jordanian site is geo-graphically very close (c. 55 km) to Talsh’hab, south-west Syria, where an individual of M. schreibersii wasfound. A transition zone between M. schreibersiiand M. pallidus may run along the Great Rift in thenorth-south transect of the Levant, therefore, andboth taxa may be present there in sympatry [simi-
larly as in Turkey (Maraci et al., 2010; Bilgin et al.,2012)] or in close parapatry, as in the case of treefrogs or geckos (Gvoždík et al., 2010; Moravec et al.,2011).
EASTERN AFGHANISTAN
The Jalalabad (Nangarhar Province of Afghanistan)population, usually considered as representing M. s.fuliginosus (e.g. Ellerman & Morrison-Scott, 1951;Gaisler, 1970; Hill, 1983; Yoshiyuki, 1989; Corbet &Hill, 1992; Koopman, 1994; Bates & Harrison, 1997;Simmons, 2005), differed strongly from all otherWestern Palaearctic populations in both linear andgeometric morphometrics, as well as in non-metrictraits. These findings, therefore, support a hypothesispreviously put forward by Maeda (1982), i.e. that thissubspecies should be regarded as a separate species,M. fuliginosus. Regrettably, there were no geneticsamples available to us to back-up the morphologicalfindings through molecular analysis. Nevertheless,according to the published phylogenetic analyses ofChinese and Japanese populations affiliated to M.fuliginosus (Appleton et al., 2004; Furman et al.,2010b), the species status of this form appears to havebeen demonstrated sufficiently as it has been shownto be genetically very distant from species of the C(West Palaearctic) clade (Fig. 2). A complex morpho-logical and molecular genetic analysis of Indiansubcontinent Miniopterus populations is needed,however, to confirm taxonomic assignation of EastAfghanistan and other Oriental region populationsformerly co-identified with M. schreibersii s.l. (cf.Gaisler, 1970; Bates & Harrison, 1997). Here, wetentatively suggest using the name M. cf. fuliginosusfor the Jalalabad populations, in accordance withprevious authors, but as a full species.
SOUTH-WESTERN ARABIA AND ETHIOPIA
Of the Western Palaearctic Miniopterus populationsexamined, that of south-western Arabia (Yemen) wasone of the most distinct. These bats demonstratedsubstantial similarities to African populations, andyet genetically were very close to the samples exam-ined from Ethiopia. Yemeni and Ethiopian samplesappear to represent an identical taxon with regard toboth genetic data (a low distance of 0.1–0.5%) andmorphology. Previously assigned to M. schreibersii s.l.(Nader & Kock, 1987; Harrison & Bates, 1991), thesepopulations have more recently been regarded as partof M. natalensis (Simmons, 2005). The separation ofAfrican populations from M. schreibersii, suggestedpreviously by Koopman (1994), has been confirmedthrough molecular analysis (Appleton et al., 2004;Miller-Butterworth et al., 2005), and our results
TAXONOMY OF MINIOPTERUS 183
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

further support this conclusion. M. natalensis,however, is a species described as from South Africa,and bats of that origin represent a genetic lineagesubstantially distant from the YE lineage (11.5%). Asthe level of genetic differentiation clearly exceeds the5% level recommended for species recognition accord-ing to the genetic species concept (Baker & Bradley,2006; Mayer et al., 2007), and the two lineages(natalensis s.str. and YE) are clearly not in a sisterphylogenetic relationship, it may be appropriate toconsider the YE lineage as a species distinct from theSouth African M. natalensis. Nader & Kock (1987),the first to attempt taxonomic determination of south-west Arabian Miniopterus populations, identifiedthese bats as M. schreibersii arenarius, based on mor-phological and parasitological evidence. Accordingto earlier classification, and in the light of our newresults, we regard south-west Arabian and EthiopianMiniopterus bats, formerly assigned to M. schreibersiiarenarius or M. natalensis arenarius (see Harrison& Bates, 1991; Koopman, 1994), as representing aseparate species tentatively named M. cf. arenariusHeller, 1912. As the name originates from Kenya, agenetic and/or morphologic comparison with type/topotypic material is needed to confirm this.
HISTORICAL BIOGEOGRAPHY OF
MINIOPTERUS SCHREIBERSII
Observed genetic variation in M. schreibersii s.str.also brings new insights into the species’ phylogeog-raphy. Furman et al. (2010a) suggested that shallowgenetic differentiation between the western andeastern European colonies, and the relatively highgenetic diversity observed in the eastern colonies,may indicate a re-colonisation of Europe from a singleglacial refugium located in north-western Anatolia.Alternatively, Bilgin et al. (2008) localised such apossible refugium in Turkish Thrace, while Pereiraet al. (2009) suggested either southern Iberia or NorthAfrica. Furman et al. (2010a) further speculated onthe existence of another glacial refugium in Italy. Ourresults, however, do not support such a hypothesis asa widely distributed haplotype was detected in south-ern Italy (Sicily; WM sublineage). To confirm such ahypothesis, an in-depth analysis of both Italian andsurrounding populations is needed. Moreover, accord-ing to the available evidence, M. schreibersii fossilsare absent in Pleistocene-Holocene transition cave-deposits in Italy (Tata & Kotsakis, 2005). Taking allthe intraspecific genetic data available (Bilgin et al.,2008; Pereira et al., 2009; Furman et al., 2010a; thisstudy) into consideration, it would appear that, inaddition to the apparent existence of a refugium inthe Atlas Mts. of Morocco (see above) dating from theMiddle Pleistocene (cf. molecular clock by Furman
et al., 2010b), the presence of three or four additionalglacial refugia can also be detected within the Mlineage. Based on the genetic structure observed inthis study (e.g. Fig. 3), we hypothesise, in accordancewith Furman et al. (2010a), that the main refugium ofthe WM sublineage was in the east in the Black Searegion. However, as the Spanish and coastal Moro-ccan samples form a distinct clade within the WMsublineage, we cannot exclude the possibility of afurther, western refugium in south-western Europeor lowland North Africa (cf. Pereira et al., 2009). Twodistinct haplotype clusters observed within the EMsublineage, comprising southern Turkish and Levan-tine samples (including Cyprus), appear to correspondwith the locations of glacial refugia in southernTurkey and the Levant (western Syria, Lebanon). Thehaplotypes of the Cypriot population do not form amonophyletic lineage, which suggests that colonisa-tion of Cyprus from the adjacent mainland probablyoccurred recently and through repeated episodes (asalso suggested for some other Cypriot bats; see Bendaet al., 2007). Considering the supposed migratorynature of M. schreibersii (cf. Rodrigues & Palmeirim,2008; Pereira et al., 2009), and the fact that geo-graphic barriers do not appear to have a substantialeffect on the evolutionary history of the species(Dobson, 1998; Appleton et al., 2004; Ibáñez et al.,2006; Bilgin et al., 2008; García-Mudarra et al., 2009;Furman et al., 2010c), one could also speculate on theexistence of additional Miniopterus refugia. It wouldappear, therefore, that the genetic structure of M. sch-reibersii is a result of complex ecological-evolutionarycausalities that may be diverse in different regions ofthe Western Palaearctic.
ACKNOWLEDGEMENTS
We thank Rainer Hutterer (Bonn) and Riyad Sadek(Beirut) for access to the museum specimens undertheir care, and Ivan Horácek and Pavel Hulva (Prague)for providing tissue samples. We are obliged to IvanHorácek for valuable comments regarding the researchtopic and previous versions of the manuscript. Thisstudy was supported by the Czech Science Foundation(# 206/09/0888) and the Ministry of Culture of theCzech Republic (#DKRVO 00023272).
REFERENCES
Aellen V, Strinati P. 1970. Chauves-souris cavernicoles deTunisie [Cave bats of Tunisia]. Mammalia 34: 228–236.
Albayrak I, Coskun S. 2000. Geographic variations andtaxonomic status of Miniopterus schreibersii (Kuhl, 1819)in Turkey (Chiroptera: Vespertilionidae). Turkish Journal ofZoology 24: 125–133.
184 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Ansell WFH, Topál G. 1976. The type locality of Miniopterusschreibersii (KUHL) (Mammalia: Chiroptera). VertebrataHungarica 17: 15–17.
Appleton BR, McKenzie JA, Christidis L. 2004. Molecularsystematics and biogeography of the bent-wing bat complexMiniopterus schreibersii (Kuhl, 1817) (Chiroptera: Vesper-tilionidae). Molecular Phylogenetics and Evolution 31: 431–439.
Baker RJ, Bradley RD. 2006. Speciation in mammals andthe genetic species concept. Journal of Mammaliology 87:643–662.
Bates PJJ, Harrison DL. 1997. Bats of the Indian Subcon-tinent. Sevenoaks: Harrison Zoological Museum.
Benda P, Andreas M, Kock D, Lucan RK, Munclinger P,Nová P, Obuch J, Ochman K, Reiter A, Uhrin M,Weinfurtová D. 2006. Bats (Mammalia: Chiroptera) ofthe Eastern Mediterranean. Part 4. Bat fauna of Syria:distribution, systematics, ecology. Acta Societatis ZoologicaeBohemicae 70: 1–329.
Benda P, Hanák V, Horácek I, Hulva P, Lucan R,Ruedi M. 2007. Bats (Mammalia: Chiroptera) of theEastern Mediterranean. Part 5. Bat fauna of Cyprus: reviewof records with confirmation of six species new for the islandand description of a new subspecies. Acta Societatis Zoo-logicae Bohemicae 71: 71–130.
Benda P, Horácek I. 1995. Geographic variation in threespecies of Myotis (Mammalia: Chiroptera) in South of theWestern Palaearctics. Acta Societatis Zoologicae Bohemicae59: 17–39.
Benda P, Vallo P, Reiter A. 2011. Taxonomic revision of thegenus Asellia (Chiroptera: Hipposideridae) with a descrip-tion of a new species from southern Arabia. Acta Chirop-terologica 13: 245–270.
Bilgin IR. 2012. The conservation genetics of three cave-dwelling bat species in southeastern Europe and Anatolia.Turkish Journal of Zoology 36: 275–282.
Bilgin R. 2011. Back to the suture: the distribution ofintraspecific genetic diversity in and around Anatolia.International Journal of Molecular Sciences 12: 4080–4103.
Bilgin R, Gürün K, Maracı Ö, Furman A, Hulva P,Çoraman E, Lucan RK, Bartonicka T, Horácek I. 2012.Syntopic occurrence in Turkey supports separate speciesstatus for Miniopterus schreibersii schreibersii and Miniopt-erus schreibersii pallidus (Mammalia: Chiroptera). ActaChiropterologica 14. In press.
Bilgin R, Karatas A, Çoraman E, Disotell T, Morales JC.2008. Regionally and climatically restricted patterns of dis-tribution and genetic diversity in a migratory bat species,Miniopterus schreibersii (Chiroptera: Vespertilionidae).BMC Evolutionary Biology 8: 209.
Bilgin R, Karatas A, Çoraman E, Pandurski I,Papadatou E, Morales JC. 2006. Molecular taxonomyand phylogeography of Miniopterus schreibersii (Kuhl, 1817)(Chiroptera: Vespertilionidae), in the Eurasian transi-tion. Biological Journal of the Linnean Society 87: 577–582.
Bogdanowicz W. 1990. Geographic variation of the taxonomy
of Daubenton’s bat, Myotis daubentoni, in Europe. Journal ofMammalogy 71: 205–218.
Boye P. 2004. Miniopterus schreibersii Natterer in Kuhl, 1819– Langflügelfledermaus. In: Krapp F, ed. Handbuch derSäugetiere Europas. Band 4: Fledertiere. Teil II: ChiropteraII. Vespertilionidae 2. Molossidae, Nycteridae [Handbook ofMammals of Europe. Volume 4: Bats. No. II: Chiroptera II.Vespertilionidae 2. Molossidae, Nycteridae]. Wiebelsheim:Aula-Verlag, 1093–1122.
Clement M, Posada D, Crandall KA. 2000. TCS: a computerprogram to estimate gene genealogies. Molecular Ecology 9:1657–1659.
Corbet GB. 1978. The mammals of the Palearctic region: ataxonomic review. London and Ithaca: British Museum(Natural History) and Cornell University Press.
Corbet GB, Hill JE. 1992. The mammals of the Indomalayanregion: a systematic review. London: Oxford UniversityPress.
Crucitti P. 1989. Distribution, diversity and abundance ofcave bats in Latium (Central Italy). In: Hanák V, HorácekI, Gaisler J, eds. European bat research 1987. Prague:Charles University Press, 381–388.
Dobson M. 1998. Mammal distributions in the western Medi-terranean: the role of human intervention. Mammal Review28: 77–88.
Drummond AJ, Rambaut A. 2007. BEAST: Bayesian evo-lutionary analysis by sampling trees. BMC EvolutionaryBiology 7: 214.
Ellerman JR, Morrison-Scott TCS. 1951. Checklist ofPalearctic and Indian mammals 1758 to 1946. London:British Museum (Natural History).
Ferguson WW. 2002. The mammals of Israel. Jerusalem:Gefen Publishing House.
Fernandez R, Ibañez C. 1989. Patterns of distribution ofbats in the Iberian Peninsula. In: Hanák V, Horácek I,Gaisler J, eds. European bat research 1987. Prague: CharlesUniversity Press, 357–361.
Fritz U, Fritzsch G, Lehr E, Ducotterd J-M, Müller A.2005. The Atlas Mountains, not the Straits of Gibraltar, asa biogeographic barrier for Mauremys leprosa. Salamandra41: 97–106.
Furman A, Çoraman E, Bilgin R, Karatas A. 2009.Molecular ecology and phylogeography of the bent-wing batcomplex (Miniopterus schreibersii) (Chiroptera: Vespertilio-nidae) in Asia Minor and adjacent regions. Zoologica Scripta38: 129–141.
Furman A, Öztunç T, Postawa T, Çoraman E. 2010a.Shallow genetic differentiation in Miniopterus schreibersii(Chiroptera: Vespertilionidae) indicates a relatively recentre-colonization of Europe from a single glacial refugium.Acta Chiroperologica 12: 51–59.
Furman A, Öztunç T, Çoraman E. 2010b. On the phylog-eny of Miniopterus schreibersii schreibersii and Miniopterusschreibersii pallidus from Asia Minor in reference to otherMiniopterus taxa (Chiroptera: Vespertilionidae). Acta Chi-ropterologica 12: 61–72.
Furman A, Postawa T, Öztunç T, Çoraman E. 2010c.Cryptic diversity of the bent-wing bat, Miniopterus schreib-
TAXONOMY OF MINIOPTERUS 185
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

ersii (Chiroptera: Vespertilionidae), in Asia Minor. BMCEvolutionary Biology 10: 121–133.
Gaisler J. 1970. The bats (Chiroptera) collected in Afghanistanby the Czechoslovak Expedition of 1965–67. Acta Scien-tiarum Naturalium Academiae Scientiarum BohemoslovacaeBrno, Series Nova 4: 1–56.
Gaisler J. 1983. Nouvelles donées sur les chiroptères du nordalgérien. Mammalia 47: 359–369.
García-Mudarra JL, Ibáñez C, Juste J. 2009. The Straitsof Gibraltar: barrier or bridge to Ibero-Moroccan bat diver-sity? Biological Journal of the Linnean Society 96: 434–450.
Gazaryan SV. 2005. Geograficeskaâ izmencivost dlinnokry-lov Miniopterus schreibersii (Chiroptera) na territorii vos-tocnoj Evropy i severo-zapadnoj Azii [Geographical variationof Schreibers’ bat Miniopterus schreibersii (Chiroptera)in the territory of eastern Europe and northwestern Asia].Zoologicheskiy Zhurnal 84: 1136–1143. (in Russian, with anabstract in English).
Guindon S, Dufayard JF, Lefort V, Anisimova M, HordijkW, Gascuel O. 2010. New algorithms and methods toestimate maximum-likelihood phylogenies: assessing theperformance of PhyML 3.0. Systematic Biology 59: 307–321.
Gvoždík V, Moravec J, Klütsch C, Kotlík P. 2010. Phylo-geography of the Middle Eastern tree frogs (Hyla, Hylidae,Amphibia) as inferred from nuclear and mitochondrial DNAvariation, with a description of a new species. MolecularPhylogenetics and Evolution 55: 1146–1166.
Hall TA. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95–98.
Hanák V, Horácek I. 1984. Some comments of the taxonomyof Myotis daubentoni (Kuhl, 1819) (Chiroptera, Mammalia).Myotis 21–22: 7–19.
Harrison DL, Bates PJJ. 1991. The mammals of Arabia.Second Edition. Sevenoaks: Harrison Zoological Museum.
Hayman RW, Hill JE. 1971. Part 2. Order Chiroptera. In:Meester J, Setzer HW, eds. The mammals of Africa: anidentification manual. Washington: Smithsonian InstitutionPress, 1–73.
Heller E. 1912. New races of insectivores, bats, and lemursfrom British East Africa. Smithsonian Miscellaneous Col-lections 60: 1–13.
Hill JE. 1983. Bats from Indo-Australia (Mammalia: Chirop-tera). Bulletin of the British Museum of Natural History 45:103–208.
Horácek I, Hanák V, Gaisler J. 2000. Bats of the Palearcticregion: a taxonomic and biogeographic review. In: WoloszynBW, ed. Proceedings of the VIIIth European Bat ResearchSymposium. Vol. 1. Approaches to Biogeography and Ecologyof Bats. Kraków: Chiropterological Information Center, Insti-tute of Systematics and Evolution of Animals PAS, 11–157.
Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesianinference of phylogeny. Bioinformatics 17: 754–755.
Hulva P, Benda P, Hanák V, Evin A, Horácek I.2007a. New mitochondrial lineages within the Pipistrelluspipistrellus complex from Mediterranean Europe. Folia Zoo-logica 56: 378–388.
Hulva P, Horácek I, Benda P. 2007b. Molecules,
morphometrics and new fossils provide an integrated viewof the evolutionary history of Rhinopomatidae (Mammalia:Chiroptera). BMC Evolutionary Biology 7: 165–180.
Hulva P, Fornusková A, Chudárková A, Evin A,Allegrini B, Benda P, Bryja J. 2010. Mechanisms ofradiation in a bat group from the genus Pipistrellus inferredby phylogeography, demography and population genetics.Molecular Ecology 19: 5417–5431.
Hulva P, Horácek I, Strelkov PP, Benda P. 2004. Molecu-lar architecture of Pipistrellus pipistrellus / P. pygmaeuscomplex (Chiroptera: Vespertilionidae): further crypticspecies and Mediterranean origin of the divergence. Molecu-lar Phylogenetics and Evolution 32: 1023–1035.
Ibanez C, García-Mudarra JL, Ruedi M, Stadelmann B,Juste J. 2006. The Iberian contribution to cryptic diversityin European bats. Acta Chiropterologica 8: 277–297.
Iliopoulou-Georgudaki J. 1986. The relationship betweenclimatic factors and forearm length of bats: evidence fromthe chiropterofauna of Lesbos island (Greece – EastAegean). Mammalia 50: 475–482.
Karatas A, Sözen M. 2004. Contribution to karyology, dis-tribution and taxonomic status of the Long-winged Bat,Miniopterus schreibersii (Chiroptera: Vespertilionidae), inTurkey. Zoology in the Middle East 33: 51–64.
Kirchman JJ, Hackett SH, Goodman SM, Bates JM.2001. Phylogeny and systematics of ground rollers (Brach-ypteraciidae) of Madagaskar. The Auk 118: 849–863.
Koopman KF. 1993. Order Chiroptera. In: Wilson DE,Reeder DM, eds. Mammal species of the world. A taxonomicand geographic reference. Washington and London: Smith-sonian Institution Press, 137–241.
Koopman KF. 1994. Chiroptera: systematics. In: Nietham-mer J, Schliemann H, Starck D, eds. Handbuch der Zoolo-gie. Band VIII. Mammalia. Teilband 60. Berlin and NewYork: Walter de Gruyter, 1–217.
Kowalski K, Rzebik-Kowalska B. 1991. Mammals ofAlgeria. Kraków: Zakład Narodowy im. Ossolinskich –Wydawnictwo.
Kuzâkin AP. 1950. Letucie myši (Sistematika, obraz žizni ipol’za dlâ sel’skogo i lesnogo hozâjstva) [Bats (Systematic,Life History and Utility in Agriculture and Forestry)].Moskva: Sovetskaja Nauka, (in Russian).
Kuzâkin AP. 1965. Otrjad Rukokrylye. Ordo Chiroptera[Order Bats. Ordo Chiroptera]. In: Bobrinskij NA, KuznecovBA, Kuzâkin AP, eds. Opredelitel’ mlekopitaûsih SSSR [Keyto identification of Mammals of the USSR]. Moskva: Pros-vešcenije, 79–116. (in Russian).
Lay DM. 1967. A study of the mammals of Iran. Resultingfrom the Street Expedition of 1962–63. Fieldiana: Zoology54: 1–282.
Librado P, Rozas J. 2009. DnaSP v5: a software for com-prehensive analysis of DNA polymorphism data. Bioinfor-matics 25: 1451–1452.
Lin Y-H, Penny D. 2001. Implications for bat evolution fromtwo new complete mitochondrial genomes. MolecularBiology and Evolution 18: 684–688.
MacArthur RH, Wilson EO. 1967. The theory of islandbiogeography. Princeton, NJ: Princeton University Press.
186 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Maeda K. 1982. Studies on the classification of Miniopterusin Eurasia, Australia and Melanesia. Honyurui Kagaku(Mammalian Science), Supplement 1: 1–176.
Maraci Ö, Bilgin R, Lucan RK, Bartonicka P, Hulva P,Horácek I. 2010. The sympatry of Miniopterus schreibersiischreibersii and Miniopterus s. pallidus in three caves: thesmoking gun for their elevation to full species status. In:Horácek I, Benda P, eds. 15th IBRC – the ConferenceManual: Programme, Abstracts, List of Participants. Volumeof Abstracts of the 15th International Bat Research Confer-ence, held in Prague, 23–27 August 2010. Kostelec nadCernými lesy: Lesnická práce s.r.o., 220–221.
Mayer F, Dietz C, Kiefer A. 2007. Molecular species iden-tification boosts bat diversity. Frontiers in Zoology 4: 1–5.
Miller-Butterworth CM, Eick G, Jacobs DS, SchoemanMC, Halley EH. 2005. Genetic and phenotypic differencesbetween South African long-fingered bats, with a globalminiopterinae phylogeny. Journal of Mammalogy 86: 1121–1135.
Moravec J, Kratochvíl L, Amr ZS, Jandzik D, Šmíd J,Gvoždík V. 2011. High genetic differentiation within theHemidactylus turcicus complex (Reptilia: Gekkonidae) inthe Levant, with comments on the phylogeny and system-atics of the genus. Zootaxa 2894: 21–38.
Nader IA, Kock D. 1987. First record of Miniopterus sch-reibersii (Kuhl 1819) (Mammalia: Chiroptera) from NorthYemen with zoogeographical relationship evidenced by wingmites (Acarina: Spinturnicidae). Senckenbergiana Biologica67: 225–229.
Ognev SI. 1927. A synopsis of the Russian bats. Journal ofMammalogy 8: 140–157.
Ognev SI. 1928. Zveri vostocnoj Evropy i severnoj Azii.Tom I. [The Mammals of the Eastern Europe and of theNorthern Asia. Vol.I]. Moskva & Leningrad: Gosudarstven-noe Izdatel’stvo, (in Russian).
Ondrias JC. 1978. Population variation in Miniopterusschreibersii from Greece. Biologia Gallo-Hellenica 7: 223–232.
Osborne MJ, Christidis L. 2001. Molecular Phylogenetics ofAustralo-Papuan Possums and Gliders (Family Petauridae).Molecular Phylogenetics and Evolution 20: 211–224.
Pereira MJR, Salgueiro P, Rodrigues L, Coelho MM,Palmeirim JM. 2009. Population structure of a cave dwell-ing bat, Miniopterus schreibersii: does it reflect history andsocial organization? Journal of Heredity 100: 533–544.
Posada D. 2008. JModelTest: phylogenetic model averaging.Molecular Biology and Evolution 25: 1253–1256.
Qumsiyeh MB, Schlitter DA. 1982. The bat fauna of JabalAl Akhdar, northeast Libya. Annals of Carnegie Museum 51:377–389.
Rahmatulina IK. 2005. Rukokrylye Azerbajdžana (fauna,ekologiâ, zoogeografiâ) [Bats of Azerbaijan (Fauna, Ecology,Zoogeography)]. Baku: Nacional’naâ Akademiâ NaukAzerbajdžana, (in Russian).
Rodrigues L, Palmeirim JM. 2008. Migratory behaviour ofthe Schreiber’s bat: when, where and why do cave batsmigrate in a Mediterranean region? Journal of Zoology 264:116–125.
Rohlf FJ. 1998. On applications of geometrics to studiesof ontogeny and phylogeny. Systematic Biology 47: 147–158.
Rohlf FJ. 2004. Tpsdig. Version 1.40. Stony Brook: StateUniversity of New York at Stony Brook, Department ofEcology and Evolution.
Rohlf FJ. 2008. Relative warps. Version 1.46. Stony Brook:State University of New York at Stony Brook, Departmentof Ecology and Evolution.
Rohlf FJ. 2009. Tpsregr. Version 1.36. Stony Brook: StateUniversity of New York at Stony Brook, Department ofEcology and Evolution.
Rohlf FJ. 2010. Tps Utility program. Version 1.46. StonyBrook: State University of New York at Stony Brook,Department of Ecology and Evolution.
Rohlf FJ, Loy A, Corti M. 1996. Morphometric analysis ofOld World Talpidae (Mammalia, Insectivora) using partial-warps scores. Systematic Biology 45: 344–362.
Ronquist F, Huelsenbeck JP. 2003. MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19: 1572–1574.
Simmons NB. 2005. Order Chiroptera. In: Wilson DE,Reeder DM, eds. Mammal species of the world. A taxonomicand geographic reference. Third Edition. Volume 1. Balti-more: The John Hopkins University Press, 312–529.
Spitzenberger F. 1981. Die Langflügelfledermaus (Miniopt-erus schreibersii Kuhl, 1819) in Österreich – Mammaliaaustriaca 5. Mitteilungen der Abteilung für Zoologie amLandesmuseum Joanneum 2: 139–156.
Strelkov PP, Sosnovcena VP, Babaev NV. 1978. Letuciemyši (Chiroptera) Turkmenii [The bats of Turkmenia].Trudy Zoologiceskogo Instituta Akademii Nauk SSSR 79:3–71. (in Russian, with a subtitle in English).
Swofford DL. 2003. PAUP*. Phylogenetic Analysis UsingParsimony (* and other methods). Version 4. Sinauer Asso-ciates, Sunderland.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4:Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Molecular Biology and Evolution 24: 1596–1599.
Tata C, Kotsakis T. 2005. Italian fossil chiropteran assem-blages: a preliminary report. Geologia Delle Alpi 2: 53–60.
Tate GHH. 1941. Results of the Archbold Expeditions.No. 40. Notes on vespertilionid bats of the subfamiliesMiniopterinae, Murininae, Kerivoulinae, and Nyctophilinae.Bulletin of the American Museum of Natural History 78:567–597.
Tian L, Liang B, Maeda K, Metzner W, Zhang S. 2004.Molecular studies on the classification of Miniopterusschreibersii (Chiroptera: Vespertilionidae) inferred frommitochondrial cytochrome b sequences. Folia Zoologica 53:303–311.
Velazco PM, Gardner AL, Patterson BD. 2010. Systemat-ics of the Platyrrhinus helleri species complex (Chiroptera:Phyllostomidae), with descriptions of two new species. Zoo-logical Journal of the Linnean Society 159: 785–812.
Yoshiyuki M. 1989. A systematic study of the Japanese Chi-roptera. Tokyo: The National Science Museum.
TAXONOMY OF MINIOPTERUS 187
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Zelditch ML, Fink WL, Swiderski DL. 1995. Morphomet-rics, homology, and phylogenetics: quantified characters assynapomorphies. Systematic Biology 44: 179–189.
APPENDIX 1
Specimens examined morphologically. Abbreviations:NMP = National Museum in Prague, Czech Republic;ZFMK = Zoological Research Museum AlexanderKoenig in Bonn, Germany; AUB = American Univer-sity of Beirut, Lebanon. Mare = Miniopterus cf. are-narius; Mful = M. cf. fuliginous; Msch = M. schreibersiis.str.; Mpal = M. pallidus; Msp = unidentified taxonof M. cf. schreibersii; f = female; m = male; x =unidentified sex.
GROUP 1: MOROCCO
Morocco: Azigza Cave (Tazouguerte) 6f (NMP:pb3910, pb3912–pb3914, pb3916, pb3917 – 26. 4.2008), 4m (pb3907–pb3909, pb3911 – 26. 4. 2008),Msch, leg.: P. Benda; Oued Tessaout Valley (Talkout)3f (NMP: 90046, 90049, 90054 – 30. 8. 2003), 4m(NMP: 90050–90052, 90055 – 30. 8. 2003), Msch, leg.:P. Benda; Oued El-Ammar River (Sebt-des-Ait-Serhrouchen) 1f (NMP: 90103 – 9. 9. 2003), Msch,leg.: P. Benda.
GROUP 2: WESTERN EUROPE
Spain: Bei Tremp (Pyrenees) 3f (ZFMK: 56.735,56.737, 56.738 – 28. 5. 1955), 4 m (ZFMK: 56.1068,56.733, 56.734, 56.736 – 28. 5. 1955), Msch, leg.: J.Niethammer; Ramales de la Victoria 1m (ZFMK:97.246 – 19. 4. 1963), Msch, leg.: J. Niethammer.France: Grotte de Povade (Banyuls) 9f (ZFMK:59.120, 59.124, 59.127-59.131, 59.133, 59.134 – 8. 4.1959), 7m (NMP: 59.121-59.123, 59.125, 59.126,59.132 – 8. 4. 1959, 59.350 – 23. 5. 1959), Msch, leg.:A. Heymer; Chateau de Collioure (Banuyls) 2f(ZFMK: 59.348, 59.349 – 11. 5. 1959), 1m (ZFMK:59.347 – 11. 5. 1959), Msch, leg.: A. Heymer; St. Remy3f (ZFMK: 59.531b, 59.531c, 59.531d – 5. 11. 1959),1m (ZFMK: 59.531a – 5. 11. 1959), Msch, leg.: H.Roer. Italy: Gargano 2f (ZFMK: 66.338 – 2. 8. 1961,66.360 – 4. 8. 1961), 2x (ZFMK: 66.359 – 2. 8. 1961,66.357 – date unspecified), Msch, leg.: G’. Witte.Country not stated: Kaiserstuhl 1x (ZFMK: 84.529– 27. 3. 1952), Msch, leg.: Eisentraut.
GROUP 3: PANNONIA
Slovakia: Certova diera Cave (Dornica) 16f (NMP:70/58, 76/58, 85/58, 89/58, 92/58, 94/58, 96/58, 100/58– 3. 2. 1958; J – 114, J – 117, J – 118, J – 123, J – 176,J – 177, J – 178, J – 180 – 10. 12. 1956), 4m (NMP:93/58, 99/58 – 3. 2. 1958; J – 174, J – 181 – 10. 12.
1956), 9x (NMP: 80/66 – date unspecified; CD, CD 1,CD 2, CD 3, CD 4, CD 5, CD 6, CD-NX – 5. 11. 1958),Msch, leg.: V. Hanák; Drienovecká vyvieracka Cave(Drienovec) 6f (NMP: 150/58, 155/58 – 6. 2. 1958;246/61 – 17. 2. 1961; 613/59 – 1. 6. 1959; pb4260,pb4261 – 17. 7. 2009), 2m (NMP: 156/58 – 6. 2. 1958;570/59 – 31. 5. 1959), Msch, leg.: V. Hanák; P. Benda.Romania: Betfia Cave (Betfia) 11f (NMP: pb4247–pb4254, pb4256–pb4258 – 13. 7. 2009), Msch, leg.:P. Benda.
GROUP 4: BALKANS
Bulgaria: Maslen nos (Primorsko) 21f (NMP: 49191,49198, 49205, 49206, 49214, 49220, 49222, 49227,49228, 49341 – 5. 6. 1957; 49686 – 27. 8. 1961; 49688– 7. 8. 1961; 49690, 49692–49698, 49700 – 27. 8.1961), 12m (NMP: 49186, 49192, 49197, 49207,49226, 49229, 49231, 49232 – 5. 6. 1957; 49691,49699, 49703 – 27. 8. 1961), Msch, leg.: V. Hanák;Zmejovi Dupki Cave (Sliven) 7f (NMP: 49148, 49151,49152, 49165 – 25. 5. 1957; 49177, 49178, 49180 – 27.5. 1957), 4m (NMP: 49150, 49166 – 25. 5. 1957;49179, 49181 – 27. 5. 1957), Msch, leg.: V. Hanák;Karlukovo 5f (NMP: 49351 – 3. 7. 1976; 49356, 49361,49362 – 5. 7. 1976; 49367 – 6. 7. 1976), 1m (NMP:49357 – 5. 7. 1976), Msch, leg.: M. Braniš et al.;Gardina Dupka Cave (Mostovo) 3f (NMP: 50059,50061, 50062 – 22. 8. 1987), 3m (NMP: 50040 – 22. 6.1984; 50058, 50060 – 22. 8. 1987), Msch, leg.: P.Musil; Hajduška Peštera Cave (Devenci) 3f (NMP:49647–49649 – 14. 6. 1977), Msch, leg.: V. Bejceket al.; Nirica Peštera Cave (Kotel) 1f (NMP: 49799 –15. 7. 1979), 1m (NMP: 49798 – 15. 7. 1979), Msch,leg.: P.Donát et al.; Kamen Brjag 2f (NMP: 50049,50050 – 12. 7. 1986), Msch, leg.: V. Hanzal et al.;Ivanova voda Cave (Dobrostan) 1m (NMP: 49806 –23. 7. 1979), Msch, leg.: P. Donát et al.; RažiškataCave (Lakatnik) 1m (NMP: 50143 – 21.12. 1956),Msch, leg.: J. Figala et al. Greece: Didimotichon(Thrakia) 1x (ZFMK: 97.247 – 3. 8. 1971), Msch, leg.:J. Niethammer; Xánthi (Kimmeria) 3f (NMP: 48622–48624 – 16. 6. 1989), 1m (NMP: 48625 – 16. 6. 1989),Msch, leg.: V. Hanák & V. Vohrálík; Evros Cave (Didi-motiho) 2f (NMP: 48665, 48667 – 22. 6. 1989), 1m(NMP: 48666 – 22. 6. 1989), Msch, leg.: V. Hanák & V.Vohrálík; Polyphemos (Maronia) 2f (NMP: 48632,48633 – 18. 6. 1989), 1m (NMP: 48642 – 19. 6. 1989),Msch, leg.: V. Hanák & V. Vohrálík; Petralona 1f(NMP: 48611 – 28. 9. 1988), 1m (NMP: 48610 – 28. 9.1988), leg.: V. Hanák & V. Vohrálík et al., 1x (ZFMK:77.51 – 25. 5. 1962), leg.: Wolf, Msch; Ioánnina Cave(Papigo) 2f (NMP: 48578, 48579 – 26. 9. 1988), Msch,leg.: V. Hanák & V. Vohrálík et al.; Avas 1m (NMP:48657 – 20. 6. 1989), Msch, leg.: V. Hanák & V.Vohrálík.
188 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

GROUP 5: CRETE
Greece – Crete: Spilion Tsanis Cave (Omalos) 1f(NMP: 91055 – 1. 10. 2006), 11m (NMP: 91054,91056–91064, 91069 – 1. 10. 2006), Msch, leg.: P.Benda; Spilia Milatou Cave (Milatos) 2f (NMP: 91115,91118 – 7. 10.), 3m (NMP: 91112–91114 – 7. 10. 2006),Msch, leg.: P. Benda; Vreikos Cave (Crete) 1f (NMP:92316 – 12. 10. 2007), Msch, leg.: unspecified; MoniKato Preveli (Lefkogia) 1m (NMP: 92311 – 11. 10.2007), Msch, leg.: unspecified.
GROUP 6: LEVANT
Syria: Safita (Hama) 4f (NMP: 48880–48883 – 29. 5.2001), Msp, leg.: P. Benda; Qala’ et al. Hosn (Hama) 2f(NMP: 49989 – 10. 5. 2001, pb1904 – 29. 5. 2001),Msp, leg.: R. Lucan, P. Benda; Talsh’hab (Der’a) 1m(NMP: 48861 – 25. 5. 2001), Msp, leg.: P. Benda.Lebanon: Er Rouais Cave (Aaqura) 3f (NMP: 91778,91779 – 22. 1. 2007; LE 86 – 26. 6. 2006), 7f (NMP:91776, 91777 – 22. 1. 2007; LE 87–LE 91 – 26. 6.2006), Msp, leg.: P. Benda et al.; I. Horácek et al.;Saleh Cave (Amchite) 22f (NMP: LE 77 – 25. 6. 2006,91808 – 28. 1. 2007; AUB: M – 085–M – 089 – 13. 10.1960; M – 091 – 14. 8. 1960; M – 108–M – 111 – 13.10. 1960; M – 113 – 13. 10. 1960; M – 1162, M – 1165– 17. 4. 1960; M – 119, M – 124, M – 127, M – 129,M – 133, M – 139, M – 140 – 18. 3. 1961), 31m (NMP:LE 78 – 25. 6. 2006; AUB: M – 084 – 13. 10. 1960; M– 092–M – 094 – 14. 8. 1960; M – 097 – 13. 10. 1960;M – 101–M – 105, M – 112 – 13. 10. 1960; M – 115 –18. 3. 1961; M – 1163, M – 1164 – 17. 4. 1965; M –120–M – 123, M – 125, M – 126, M – 128, M – 130–M– 132, M – 134–M – 138, M – 142 – 18. 3. 1961), Msp,leg.: I. Horácek et al.; P. Benda, R. E. Lewis. Turkey:Indigu Majarasi Cave (Antalya) 1f (ZFMK: 66.626 –11. 4. 1966), 6m (ZFMK: 66.619 – 20. 4. 1966, 66.625,66.627–630 – 11. 4. 1966), Msch, leg.: K. Dobat; Haru-niye 2x (ZFMK: 58.282, 58.283 – 1953 (unspecified),Msch, leg.: unspecified. Cyprus: Smigies Trail(Akamas Peninsula) 5f (NMP: CH 32, CH 33, CH 35,CH 38 – 27. 3. 2005; CH 129 – 12. 10. 2005), 3m(NMP: CH 34, CH 36, CH 39 – 27. 3. 2005), Msp, leg.:I. Horácek et al.; Troodos forest – valley N of Kako-
petria (Kakopetria) 5m (NMP: pb2805–pb2807 – 11.4. 2005), Msp, leg.: P. Benda; Troodos Forest – valley4 km SW of Kakopetria (Kakopetria) 2m (NMP: CH45, CH 46 – 29. 3. 2005), Msp, leg.: I. Horácek et al.;Kalavasos 1m (pb2836 – 19. 4. 2005), Msp, leg.: P.Benda.
GROUP 7: MIDDLE EAST
Afghanistan: Samphshir Ghor (Kala bust) 1f(ZFMK: 97.237 – 29. 3. 1972), 2m (ZFMK: 97.235,97.236 – 29. 3. 1972), Mpal, leg.: J. Niethammer;Kandahar 1f (ZFMK: 97.245 – 28. 2. 1965), Mpal, leg.:J. Niethammer. Iran: Mina 7m (NMP: 90825 – 90830– 22. 5. 2006), Mpal, leg.: P. Benda; Bisotun (Kerman-shah) 1f (NMP: 48150 – 10. 8. 1998), 2m (NMP:48149, 48151 – 10. 8. 1998), Mpal, leg.: P. Benda;Dorud (Lorestan) 1m (48154 – 10. 8. 1998), Mpal, leg.:P. Benda.
GROUP 8: EASTERN AFGHANISTAN
(JALALABAD AREA)
Afghanistan: Jalalabad 9f (ZFMK: 97.226, 97.227,97.228, 97.229, 97.232, 97.234 – 1. 3. 1966; 97.238 –14. 5. 1965; 97.243, 97.243 – 4. 3. 1966), 16m (ZFMK:97.215, 97.216, 97.218, 97.220, 97.221, 97.222, 97.224,97.225, two samples without number – 14. 5. 1965;97.231, 97.233 – 1. 3. 1966; 97.239, 97.240, 97.241 –14. 5. 1965; 97.244 – 4. 3. 1966), 1x (ZFMK: 97.230 –1. 3. 1966), Mful, leg.: J. Niethammer.
GROUP 9: YEMEN AND ETHIOPIA
Yemen: Jebel Bura (Riqab) 1f (NMP: pb3129 – 30. 10.2005), 5m (NMP: pb3126–pb3128, pb3130, pb3131 –30. 10. 2005), Mare, leg.: P. Benda; At Tur (Hajjah) 1m(ZFMK: 85.64 – 1. 3. 1985), 1x (ZFMK: 85.63 – 1. 3.1985), Mare, leg.: F. Schutte, H.P. Fritéz; Halhal(Haja) 1m (NMP: pb3747 – 2. 11. 2007), Mare, leg.: P.Benda. Ethiopia: Baro River (Masha) 2f (NMP:92177, 92178 – 5. 9. 2003), Mare, leg.: P. Benda.Sudan: (unspecified) 1x (ZFMK: 212 – date unspeci-fied), Mare, leg.: unspecified.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Figure S1. Cranio-dental measurements and landmarks used in the linear and geometric morphometricanalyses.Figure S2. Dental measurements used in the linear morphometric analyses.Figure S3. Non-metric dental and cranial characters.Figure S4. Results of the discriminant function analyses based on the linear morphometric data of dentaldimensions.
TAXONOMY OF MINIOPTERUS 189
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

Figure S5. The main shape variable (RW1) plotted against the centroid size (CS2) of the lateral view of skull.Table S1. Non-metric dental and cranial characters. Letter codes are associated to those in Fig. S3.Table S2. Skull dimensions of the examined Miniopterus.Table S3. Dental dimensions of the examined Miniopterus.Table S4. Results of ANOVA analyses of Middle Eastern (representing M. pallidus) and Balkan (containingsequenced samples from the Levant) (representing M. schreibersii) sample sets.Table S5. Percentage share-values of the total variation of the first four relative warps of the examined samplesets for the respective view of skull and mandible.Table S6. Non-metric dental and cranial characters of the examined Miniopterus.Appendix S1. Supporting information to the methods. List of cranio-dental measurements. List of dentalmeasurements. Landmark definitions for respective views of skull and mandible.Appendix S2. Supporting information to the results. Description of morphometric cranial and dental differ-entiation among the examined groups of Miniopterus. Description of non-metric dental and cranial differen-tiation among the examined groups of Miniopterus.
190 J. ŠRÁMEK ET AL.
© 2012 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 165–190

1
SUPPORTING INFORMATION
Appendix S1: Supporting information to the methods
- List of cranio-dental measurements.
- List of dental measurements.
- Landmark definitions for respective views of skull and mandible.
- Table S1. Non-metric dental and cranial characters.
- Figure S1. Cranio-dental measurements and landmarks used in the linear and
geometric morphometric analyses.
- Figure S2. Dental measurements used in the linear morphometric analyses.
- Figure S3. Non-metric dental and cranial characters.
Appendix S2: Supporting information to the results
- Description of morphometric cranial and dental differentiation among the examined
groups of Miniopterus.
- Description of non-metric dental and cranial differentiation among the examined
groups of Miniopterus.
- Table S2. Skull dimensions of the examined Miniopterus.
- Table S3. Dental dimensions of the examined Miniopterus.
- Table S4. Results of ANOVA analyses of Middle Eastern (representing M. pallidus)
and Balkan (containing sequenced samples from the Levant) (representing M.
schreibersii) sample sets.
- Table S5. Percentage share-values of the total variation of the first four relative warps
of the examined sample sets for the respective view of skull and mandible.
- Table S6. Non-metric dental and cranial characters of the examined Miniopterus.
- Figure S4. Results of the discriminant function analyses based on the linear
morphometric data of dental dimensions.
- Figure S5. The main shape variable (RW1) plotted against the centroid size (CS2) of
the lateral view of skull.

2
Appendix S1
SUPPORTING INFORMATION TO THE METHODS
List of cranio-dental measurements:
Skull length (LCr); condylobasal length (LCb); zygomatic width (LaZ); interorbital
constriction width (LaI); rostral width – between infraorbital foramens (LaInf); neurocranium
width (LaN); mastoidal width (LaM); neurocranium height (ANc); skull height (incl.
tympanic bullae) (ACr); mandible length – condylar (LMd); coronoid process height (ACo);
rostral width across the upper canines (CC); rostral width across the upper premolars (P4P4);
rostral width across the third upper molars (M3M3); upper tooth-row length – between the first
incisor and third molar (I1M3), – between the canine and third molar (CM3), – between the
second (larger) premolar and third molar (P4M3); upper molar-row length (M1M3); upper
tooth-row length – between the canine and second premolar (CP4). The last five
measurements were taken also on mandible (I1M3, CM3, P4M3, M1M3, CP4). See Figure S1
below.
List of dental measurements:
First and second incisor length (LI1, LI2), canine length (LCsup), first and second premolar
length (LP2, LP4), third molar length (LM3) – in the upper tooth-row; first, second and third
incisor length (LI1, LI2, LI3), canine length (LCinf), first, second and third premolar length
(LP2, LP3, LP4), first, second and third molar length (LM1, LM2, LM3) – in the lower tooth-
row; first and second incisor width (WI1, WI2), canine width (WCsup), first and second
premolar width (WP2, WP4), third molar width (WM3) – in the upper tooth-row; first, second
and third incisor width (WI1, WI2, WI3), canine width (WCinf), first, second and third
premolar width (WP2, WP3, WP4) – in the lower tooth-row; first and second molar diagonal
width (W1M1, W1M2); first and second molar largest (external) length (LoM1, LoM2); first
and second molar smallest (internal) length (LiM1, LiM2); first and second molar width (in
central part) (W2M1, W2M2); first and second molar largest width (in the distal half) (W3M1,
W3M2); first and second molar width (in central part of the mesial half) (W1M1, W1M2); first
and second molar width (in central part of the distal half) (W2M1, W2M2); first molar
diagonal width (W3M1); third molar smallest width (WM3); coronal height of all teeth except
molars (HI1, HI2, Csup, HP2, HP4, HI1, HI2, HI3, Cinf, HP2, HP3, HP4). See Figure S2 below.

3
Landmark definitions for respective views of skull and mandible:
Lateral view of mandible (Fig. S1A) – (1) mandible anterior extremity, on the level of the first
incisor; (2) third molar posterior extremity; (3) lateral extremity of the mesial margin of
ramus mandibulae; (4) top point of the processus coronoideus; (5) top point of the processus
condylaris; (6) convex extremity on the level of incisura mandibulae inferior; (7) convex
extremity at the base of processus angularius ; (8) convex extremity of the ventral margin of
corpus mandibulae, in its distal half; (9) ventral extremity on the level of the contact of left
and right corpora mandibulae.
Lateral view of skull (Fig. S1B) – (1) skull anterior extremity, on the level of the first
incisor; (2) nasal fossa dorsal extremity; (3) rostrum/skull congruent point; (4) skull anterior
lateral extremity; (5) concave extremity of dorsal neurocranium; (6) intersection between
interparietal and supraoccipital bone; (7) supraoccipital basis; (8) convex extremity of the
occipital condyle; (9) top point of the processus mastoideus; (10) latero-anterior border of
ocular orbit; (11) third molar posterior extremity; (12) alveolar limit between second premolar
and first molar; (13) alveolar limit between canine and first premolar.
Ventral view of skull (Fig. S1C) – (1) premaxillar incisura; (2) skull anterior
extremity, on the level of the first incisor; (3) alveolar limit between canine and first
premolar; (4) anterior extremity of the first molar; (5) lateral extremity of the third molar; (6)
mandibular arcade incisura; (7) right lateral extremity of lacerated foramen; (8) left lateral
extremity of articular fossa; (9) lateral extremity of processus mastoideus; (10) occipital
condyle posterior extremity; (11) foramen magnum incisura.
Dorsal view of skull (Fig. S1D) – (1) premaxillar incisura; (2) skull anterior extremity,
on the level of the first incisor; (3) rostrum lateral extremity, on the level of canine; (4) largest
rostrum lateral extremity; (5) interorbital most convex extremity; (6) neurocranial lateral
extremity; (7) lateral mastoidal extremity; (8) lateral margin of squama occipitalis; (9) skull
occipital posterior extremity.

4
Table S1. Non-metric dental and cranial characters. Letter codes are associated to those in
Fig. S3.
Variable Sign Description
P4inf A The slant of the third lower premolar (P4)
P4inf2 B The shape of the third lower premolar cingulum (P4)
P3inf C The shape of the second lower premolar cingulum (P3)
P3inf2 D The rate of extension of latero-distal arch of the second lower premolar (P3)
P3inf3 E The slant of the second lower premolar (P3)
P2P4inf F Relative distance between tips of the first (P2) and third (P4) lower premolars
P2P4inf2 G Relative high of the first lower premolar (P2) to the third lower premolar (P4)
P3P4inf H Relative high of the second lower premolar (P3) to the third lower premolar (P4)
FmenP2inf I Position of foramen mentale to the first lower premolar (P2)
Fmen J Absolute size of foramen mentale
Cinf K The shape of the lower canine cingulum on the occlusal side
CingM1inf L The rate of the first lower molar (M1) protocingulum convexity (in ventral direction)
CingM1inf2 M The rate of the first lower molar (M1) protocingulum progression to postcingulum progresion (in ventral direction)
CingM1inf3 O Position of the top point of the first lower molar (M1) cingulum concavity
CingM1inf4 P The rate of deflection of the first lower molar (M1) postcingulum
CingM2inf Q The rate of the second lower molar (M2) protocingulum convexity (in ventral direction)
CingM2inf2 R The rate of the second lower molar (M2) protocingulum progression to postcingulum progresion (in ventral direction)
CingM2inf3 S Position of the top point of the second lower molar (M2) cingulum concavity
M3sup T The rate of the third lower molar (M3) metacone progression (in ventral direction)
M3sup2 U The rate of the third upper molar (M3) palatal side progression
M2sup V The slant of the second upper molar (M2) occlusal cant
M2sup2 W The rate of the second upper molar (M2) deflection on mesial side
M2sup3 X The greatest rate of the second upper molar (M2) depression on distal side
M2Sup4 Y The rate of the second upper molar (M2) distal side progression (in lateral half)
CingM2sup Z The rate of the second upper molar (M2) cingulum extremity progression on occluso-distal arch
M1sup AA The slant of the first upper molar (M1) occlusal cant
M1sup2 AB The greatest rate of the first upper molar (M1) depression on distal side
M1sup3 AC The rate of the first upper molar (M1) distal side progression (in lateral half)
M1sup4 AD The rate of the first upper molar (M1) occlusal side depression
M1sup5 AE Position of the first upper molar (M1) palatal side depression
P4sup AF The rate of the second upper premolar (P4) mesio-occlusal arch progression
P4sup2 AG The rate of the second upper premolar (P4) depression on mesial side
P4sup3 AH The rate of the second upper premolar (P4) depression on distal side
P4sup4 AI The rate of the extremity progression on mesio-occlusal side of the second upper premolar (P4)
P4sup5 AJ The rate of the second upper premolar (P4) mesio-lateral arch progression
P4sup6 AK The rate of the second upper premolar (P4) deflection on lateral side (in mesial half of cingulum)
P4sup7 AL The rate of the second upper premolar (P4) deflection on lateral side (in distal half of cingulum)
P4sup8 AM Central position of the second upper premolar (P4) lateral side depression (in distal half of cingulum)
P2sup AN Relative width of mesial arch of the first upper premolar (P2)
P2sup2 AO The rate of depression on the first upper premolar (P2) distal side
P2sup3 AP The rate of depression on the first upper premolar (P2) occlusal side
P2sup4 AQ The rate of depression on the first upper premolar (P2) mesio-lateral side
CingCsup AR The rate of upper canine cingular extremity progression
ZygW AS The rate of zygomatic arch intension
InfO AT Position of infraorbital foramen to first upper premolar (P2)
I2supCsup AU Depth of depression between second upper incisor (I2) and upper canine
ProcCW AV Relative width of procesus coronoideus
RmanW AW Relative size of ramus mandibulae extremity before processus coronoideus
Isup AX The slant of upper incisives
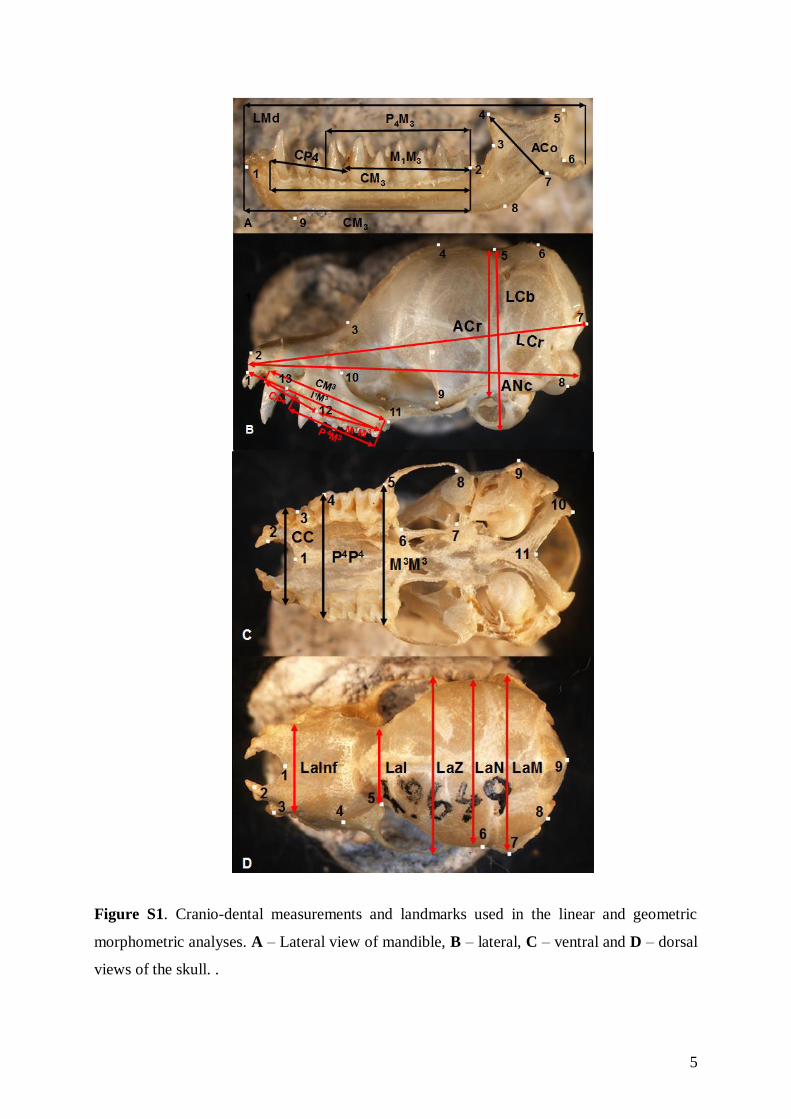
5
Figure S1. Cranio-dental measurements and landmarks used in the linear and geometric
morphometric analyses. A – Lateral view of mandible, B – lateral, C – ventral and D – dorsal
views of the skull. .

6
Figure S2. Dental measurements used in the linear morphometric analyses. View of A – right
upper, B – left lower tooth-row, C – lower incisives, D – upper incisives, E – lower canine, F
– from left to right: upper canine, 2nd premolar, 4th premolar; G – lower premolar-row, from
left to right: 2nd, 3rd and 4th premolar.
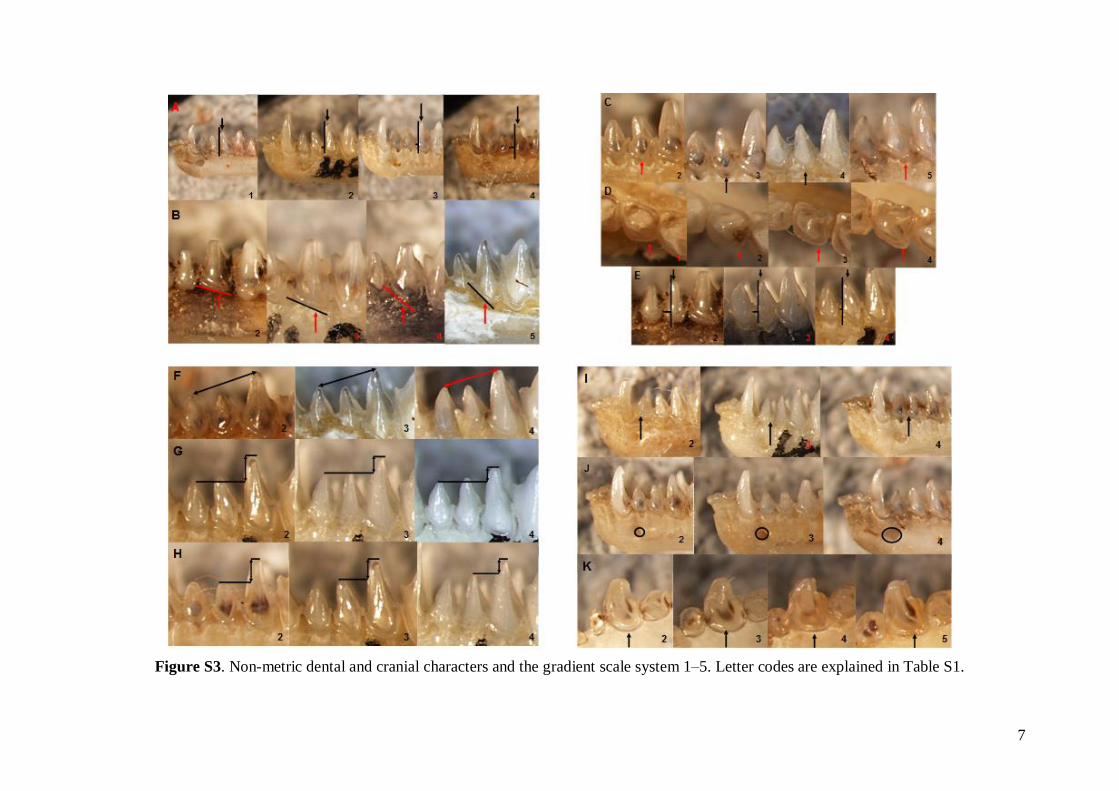
7
Figure S3. Non-metric dental and cranial characters and the gradient scale system 1–5. Letter codes are explained in Table S1.
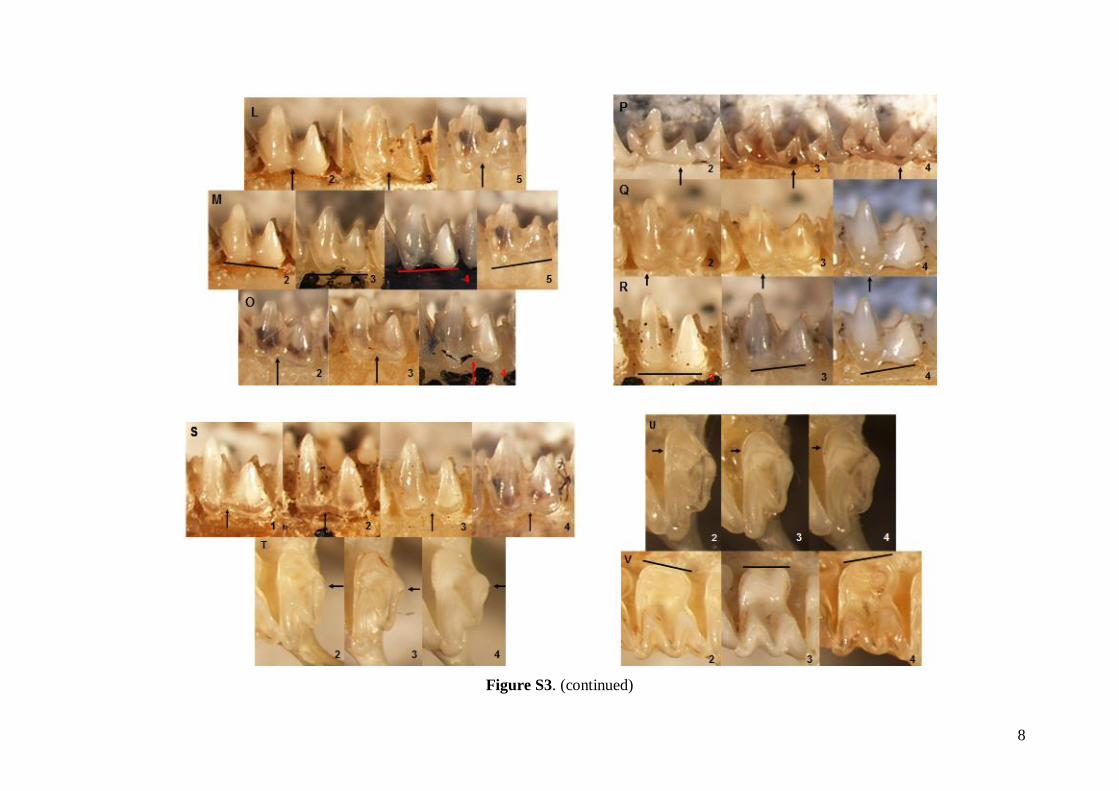
8
Figure S3. (continued)

9
Figure S3. (continued)
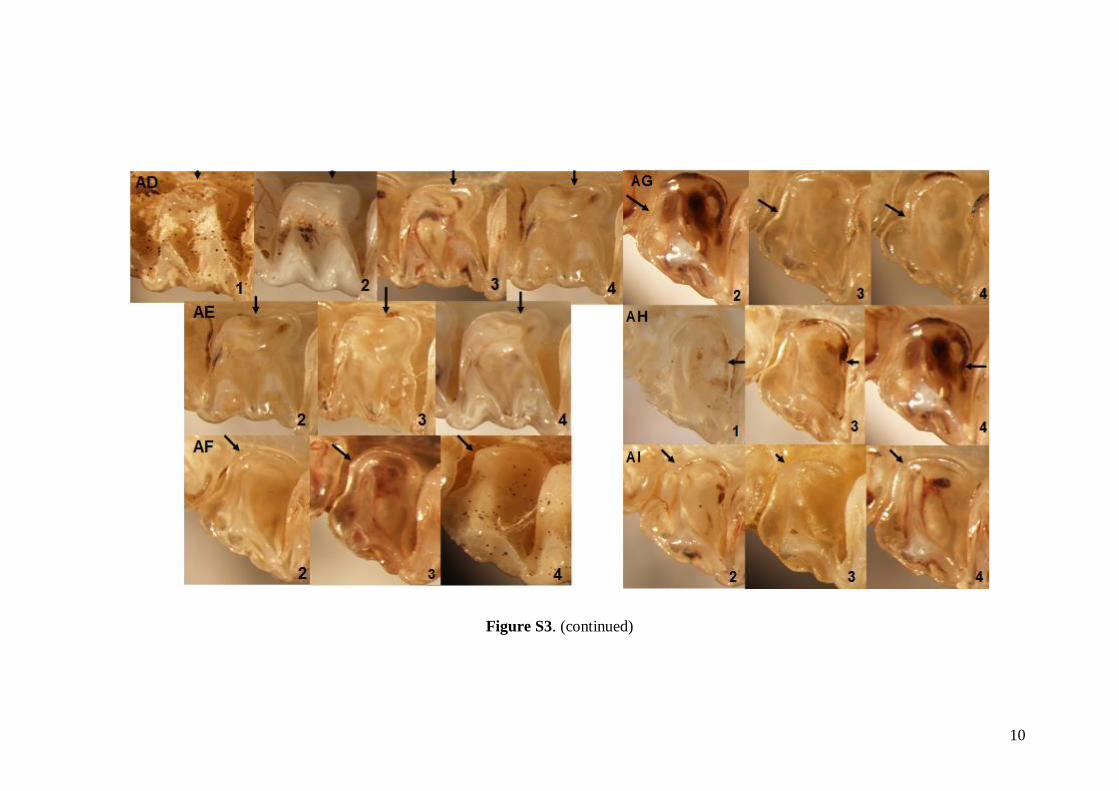
10
Figure S3. (continued)

11
Figure S3. (continued)
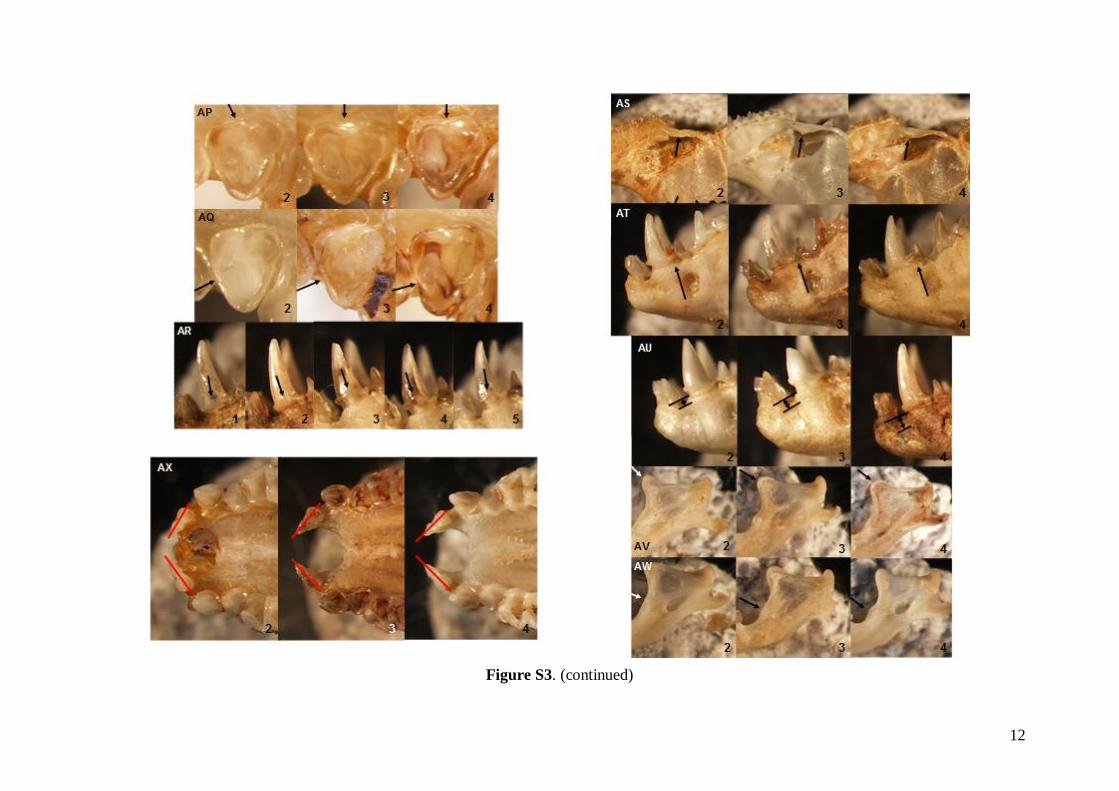
12
Figure S3. (continued)

13
Appendix S2
SUPPORTING INFORMATION TO THE RESULTS
Description of morphometric cranial and dental differentiation among the examined
groups of Miniopterus:
Morphometric differences among the examined groups as described below represent
distinctions related to the material from Pannonia, which contains samples originating from
the region of type locality of M. schreibersii in Romania (unless otherwise stated).
The bats from the Moroccan inland were characterized by relatively wider rostrum in
comparison to other examined samples, particularly at the level of infraorbital foramens (see
LaInf, LCr/LaInf in Table S2), their skulls were relatively very high and short. Mandibles of
the Moroccan bats were, among the all examined groups, the most distinct – the coronoid
processes were relatively very high, molar-rows quite short (M1M3; LMd/M1M3 in table S2).
Lower canines were relatively high, second upper premolars (P4) wide. The lower premolars
(P2 and P3) were markedly high and generally large, lower molariform teeth (P4 and M3) wide,
third lower premolar (P4) generally small.
The bats from Western Europe possessed skulls with relatively wide rostrum and
markedly long upper molar-row (M1M3; M3M3/M1M3 in table S2). Their coronoid processes
were very high (ACo; LMd/ACo in table S2), second upper premolars (P4) were markedly
high and generally large, first upper molars (M1) were large. First lower incisors (I1) were
relatively wide; lower canines low but large; lower molariforms P2, P3, M1 and M3 long (in
the mesiodistal direction); lower premolars (P3 and P4) were high; all three lower premolars
were generally small, canines large.
The bent-winged bats from the Balkans showed their skulls to be relatively high
(ACr) and their upper molar-rows (M1M3) relatively long. The second upper incisors (I2) were
relatively long, second upper premolars (P4) low and wide, first upper molars (M1) were –
relatively to the largest length of the tooth – large in the smallest length of the tooth. First
lower incisors (I1) were relatively wide; first lower molars (M1) were generally large.
The samples from Crete were characterized by their relatively high skulls and very
high coronoid processes and long lower molariform tooth-rows (P4M3). These bats showed
relatively long second lower incisors (I2), while their second upper premolars (P4) were low
and wide, first upper molars (M1) – in comparison to the largest length of the tooth –

14
relatively large in the smallest length of the tooth. The first lower incisors (I1) were relatively
wide, the first lower molars (M1) large.
The bats from the Levant showed high skulls, similarly as the Cretan bats and very
high coronoid processes. The Levantine bats possessed generally large second upper
premolars (P4), relatively wide first lower incisors (I1), markedly low first lower premolars
(P2), long lower molars (M1 and M3), small lower premolars (especially P4) and large lower
canines.
The bats from the Middle East were characterized by relatively markedly narrow
width of interorbital constriction (LaI, LCr/LaI in Table S2) and high skulls, mandibles with
relatively high coronoid processes, in upper tooth-row relatively wide and low second (large)
premolars (P4) and generally smaller second incisors (I2), premolars (P2, P3) and third molars
(M3); in lower tooth-row relatively wide third incisors (I3), long first molars (M1), markedly
low second premolars (P2), but very high third and fourth premolars (P3, P4), very small
canines and all premolars.
The samples from Eastern Afghanistan (Jalalabad) showed relatively high coronoid
processes, short mandibular molar-rows, relatively small and long second upper incisors (I2),
long first upper premolars (P2), high second upper premolars (P4), large and markedly high
upper canines, but the first upper incisors (I1) markedly small, relatively long second upper
molars (M2) in their smallest length of the tooth. In the mandibles, the first incisors (I1) were
relatively wide, second incisors (I2), third and fourth premolars (P3, P4) long, second and third
premolars (P2, P3) low, canines markedly large (especially in their high), but premolars rather
small, as well as the third molars (M3) that were moreover relatively wide.
The material of the M. schreibersii-like form from Yemen and Ethiopia were
particularly distinct from other examined samples, particularly in their width dimensions –
skulls were relatively narrow (LaM; LCr/LaM in Table S2), rostra generally very narrow
(LaInf); mandibles had the coronoid processes relatively low; lower canines were relatively
high, first upper premolars (P2) high and long, but generally small. Upper molars (M2 and M3)
were generally small, second upper incisors (I2) markedly. In the lower dentition, incisors (I1,
I3) and the last molars (M3) were relatively wide, first premolars (P2) low and markedly small,
second premolars (P3) very high – as well as the third premolars (P4) – and very markedly
small. For details concerning the skull sizes and other cranial and dental dimensions see
Tables S2 and S3.

15
Description of non-metric dental and cranial differentiation among the examined groups
of Miniopterus:
Observed values of non-metric characters showed that the most distinct samples, in
comparison to those from Pannonia, were samples from Eastern Afghanistan (Jalalabad), such
a trend was less apparent concerning the group of the Afro-Arabian samples, Moroccan and
the Middle Eastern samples. Differences among all other examined groups were rather
minute. Distinctions of the above mentioned mostly differentiated groups were as follows:
Moroccan samples had cingulum of canine palatal side without any extremity (Cinf),
first upper molar occlusal side deflection quite expressive (M1sup4) and relatively expressive
extremity on ramus mandibulae before processus coronoideus (RmanW). Samples from the
Middle East possesed with first upper molar occlusal side deflection relatively expressive
(M1sup4) and strong zygomatic arch intension (ZygW).
Samples from Eastern Afghanistan (Jalalabad) were characterized by relatively
small slant of the third lower premolar (P4inf), markedly relatively low first and second lower
premolars in comparison to third lower premolar (P2P4inf2, P3P4inf), less expressive
protocingulum in comparison to postcingulum of the first and second lower molars
(CingM1inf2, CingM2inf2), mesial position of the top point of the second lower molar
cingulum concavity (CingM2inf3), small rate of the third upper molar palatal side progression
(M3sup2), small depression on the mesial side and small progression on the distal side of the
second upper molar (M2sup2, M2sup4), small rate of the second upper premolar deflection on
lateral side (in mesial half of cingulum) (P4sup6), small rate of deflection of the first upper
molar mesio-lateral side (P2sup4) and small depth of depression between second upper
incisor and upper canine (I2supCsup).
Samples of the M. schreibersii-like form from Yemen and Ethiopia had mesial
position of the top point of the first lower molar cingulum concavity (CingM1inf3), small rate
of the third upper molar palatal side progression (M3sup2), distal slant of the second upper
molar occlusal cant (M2sup) and its cingulum extremity of distal-occlusal arch markedly
relatively small (CingM2sup), distal slant of the first upper molar occlusal cant (M1sup),
strong rate of the second upper premolar mesio-occlusal arch extremity progression (P4sup4),
relatively small rate of depression of the first upper premolar distal side (P2sup2), distal
position of infraorbital foramen in comparison to the first upper premolar (InfO). For details
concerning the non-metric traits see Table S5.
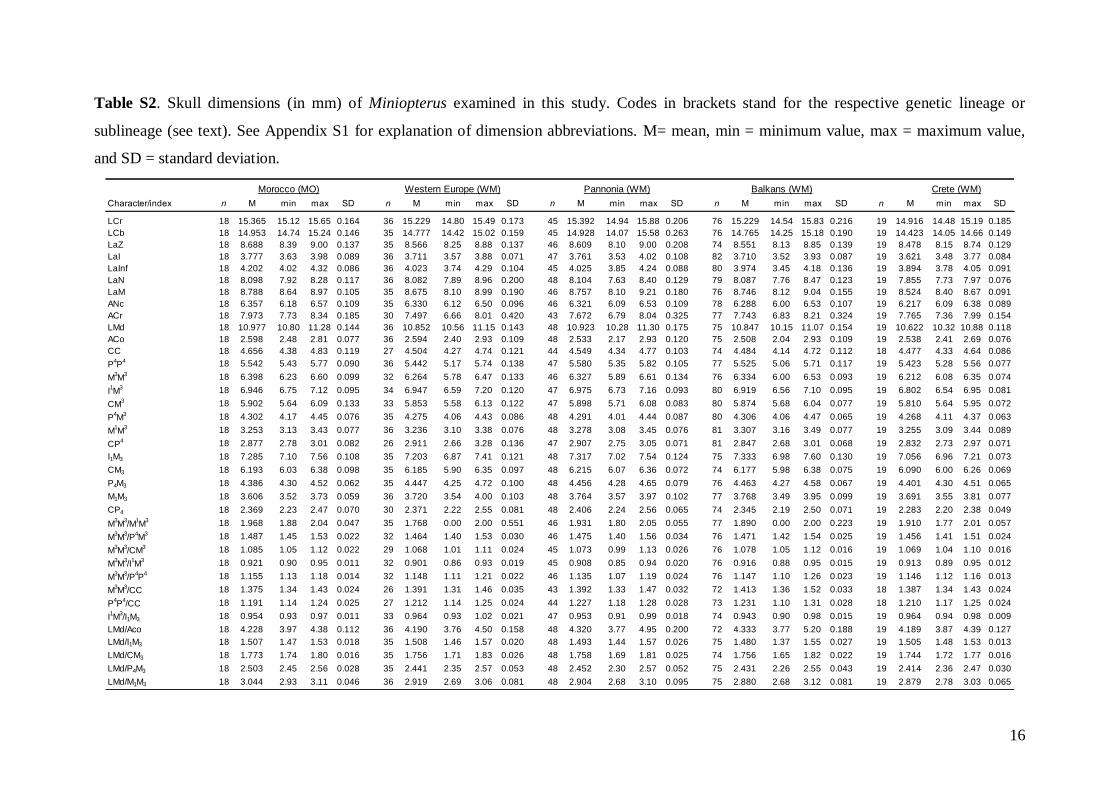
16
Table S2. Skull dimensions (in mm) of Miniopterus examined in this study. Codes in brackets stand for the respective genetic lineage or
sublineage (see text). See Appendix S1 for explanation of dimension abbreviations. M= mean, min = minimum value, max = maximum value,
and SD = standard deviation.
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
LCr 18 15.365 15.12 15.65 0.164 36 15.229 14.80 15.49 0.173 45 15.392 14.94 15.88 0.206 76 15.229 14.54 15.83 0.216 19 14.916 14.48 15.19 0.185
LCb 18 14.953 14.74 15.24 0.146 35 14.777 14.42 15.02 0.159 45 14.928 14.07 15.58 0.263 76 14.765 14.25 15.18 0.190 19 14.423 14.05 14.66 0.149
LaZ 18 8.688 8.39 9.00 0.137 35 8.566 8.25 8.88 0.137 46 8.609 8.10 9.00 0.208 74 8.551 8.13 8.85 0.139 19 8.478 8.15 8.74 0.129
LaI 18 3.777 3.63 3.98 0.089 36 3.711 3.57 3.88 0.071 47 3.761 3.53 4.02 0.108 82 3.710 3.52 3.93 0.087 19 3.621 3.48 3.77 0.084
LaInf 18 4.202 4.02 4.32 0.086 36 4.023 3.74 4.29 0.104 45 4.025 3.85 4.24 0.088 80 3.974 3.45 4.18 0.136 19 3.894 3.78 4.05 0.091
LaN 18 8.098 7.92 8.28 0.117 36 8.082 7.89 8.96 0.200 48 8.104 7.63 8.40 0.129 79 8.087 7.76 8.47 0.123 19 7.855 7.73 7.97 0.076
LaM 18 8.788 8.64 8.97 0.105 35 8.675 8.10 8.99 0.190 46 8.757 8.10 9.21 0.180 76 8.746 8.12 9.04 0.155 19 8.524 8.40 8.67 0.091
ANc 18 6.357 6.18 6.57 0.109 35 6.330 6.12 6.50 0.096 46 6.321 6.09 6.53 0.109 78 6.288 6.00 6.53 0.107 19 6.217 6.09 6.38 0.089
ACr 18 7.973 7.73 8.34 0.185 30 7.497 6.66 8.01 0.420 43 7.672 6.79 8.04 0.325 77 7.743 6.83 8.21 0.324 19 7.765 7.36 7.99 0.154
LMd 18 10.977 10.80 11.28 0.144 36 10.852 10.56 11.15 0.143 48 10.923 10.28 11.30 0.175 75 10.847 10.15 11.07 0.154 19 10.622 10.32 10.88 0.118
ACo 18 2.598 2.48 2.81 0.077 36 2.594 2.40 2.93 0.109 48 2.533 2.17 2.93 0.120 75 2.508 2.04 2.93 0.109 19 2.538 2.41 2.69 0.076
CC 18 4.656 4.38 4.83 0.119 27 4.504 4.27 4.74 0.121 44 4.549 4.34 4.77 0.103 74 4.484 4.14 4.72 0.112 18 4.477 4.33 4.64 0.086
P4P
4 18 5.542 5.43 5.77 0.090 36 5.442 5.17 5.74 0.138 47 5.580 5.35 5.82 0.105 77 5.525 5.06 5.71 0.117 19 5.423 5.28 5.56 0.077
M3M
3 18 6.398 6.23 6.60 0.099 32 6.264 5.78 6.47 0.133 46 6.327 5.89 6.61 0.134 76 6.334 6.00 6.53 0.093 19 6.212 6.08 6.35 0.074
I1M
3 18 6.946 6.75 7.12 0.095 34 6.947 6.59 7.20 0.120 47 6.975 6.73 7.16 0.093 80 6.919 6.56 7.10 0.095 19 6.802 6.54 6.95 0.081
CM3 18 5.902 5.64 6.09 0.133 33 5.853 5.58 6.13 0.122 47 5.898 5.71 6.08 0.083 80 5.874 5.68 6.04 0.077 19 5.810 5.64 5.95 0.072
P4M
3 18 4.302 4.17 4.45 0.076 35 4.275 4.06 4.43 0.086 48 4.291 4.01 4.44 0.087 80 4.306 4.06 4.47 0.065 19 4.268 4.11 4.37 0.063
M1M
3 18 3.253 3.13 3.43 0.077 36 3.236 3.10 3.38 0.076 48 3.278 3.08 3.45 0.076 81 3.307 3.16 3.49 0.077 19 3.255 3.09 3.44 0.089
CP4 18 2.877 2.78 3.01 0.082 26 2.911 2.66 3.28 0.136 47 2.907 2.75 3.05 0.071 81 2.847 2.68 3.01 0.068 19 2.832 2.73 2.97 0.071
I1M3 18 7.285 7.10 7.56 0.108 35 7.203 6.87 7.41 0.121 48 7.317 7.02 7.54 0.124 75 7.333 6.98 7.60 0.130 19 7.056 6.96 7.21 0.073
CM3 18 6.193 6.03 6.38 0.098 35 6.185 5.90 6.35 0.097 48 6.215 6.07 6.36 0.072 74 6.177 5.98 6.38 0.075 19 6.090 6.00 6.26 0.069
P4M3 18 4.386 4.30 4.52 0.062 35 4.447 4.25 4.72 0.100 48 4.456 4.28 4.65 0.079 76 4.463 4.27 4.58 0.067 19 4.401 4.30 4.51 0.065
M1M3 18 3.606 3.52 3.73 0.059 36 3.720 3.54 4.00 0.103 48 3.764 3.57 3.97 0.102 77 3.768 3.49 3.95 0.099 19 3.691 3.55 3.81 0.077
CP4 18 2.369 2.23 2.47 0.070 30 2.371 2.22 2.55 0.081 48 2.406 2.24 2.56 0.065 74 2.345 2.19 2.50 0.071 19 2.283 2.20 2.38 0.049
M3M
3/M
1M
3 18 1.968 1.88 2.04 0.047 35 1.768 0.00 2.00 0.551 46 1.931 1.80 2.05 0.055 77 1.890 0.00 2.00 0.223 19 1.910 1.77 2.01 0.057
M3M
3/P
4M
3 18 1.487 1.45 1.53 0.022 32 1.464 1.40 1.53 0.030 46 1.475 1.40 1.56 0.034 76 1.471 1.42 1.54 0.025 19 1.456 1.41 1.51 0.024
M3M
3/CM
3 18 1.085 1.05 1.12 0.022 29 1.068 1.01 1.11 0.024 45 1.073 0.99 1.13 0.026 76 1.078 1.05 1.12 0.016 19 1.069 1.04 1.10 0.016
M3M
3/I
1M
3 18 0.921 0.90 0.95 0.011 32 0.901 0.86 0.93 0.019 45 0.908 0.85 0.94 0.020 76 0.916 0.88 0.95 0.015 19 0.913 0.89 0.95 0.012
M3M
3/P
4P
4 18 1.155 1.13 1.18 0.014 32 1.148 1.11 1.21 0.022 46 1.135 1.07 1.19 0.024 76 1.147 1.10 1.26 0.023 19 1.146 1.12 1.16 0.013
M3M
3/CC 18 1.375 1.34 1.43 0.024 26 1.391 1.31 1.46 0.035 43 1.392 1.33 1.47 0.032 72 1.413 1.36 1.52 0.033 18 1.387 1.34 1.43 0.024
P4P
4/CC 18 1.191 1.14 1.24 0.025 27 1.212 1.14 1.25 0.024 44 1.227 1.18 1.28 0.028 73 1.231 1.10 1.31 0.028 18 1.210 1.17 1.25 0.024
I1M
3/I1M3 18 0.954 0.93 0.97 0.011 33 0.964 0.93 1.02 0.021 47 0.953 0.91 0.99 0.018 74 0.943 0.90 0.98 0.015 19 0.964 0.94 0.98 0.009
LMd/Aco 18 4.228 3.97 4.38 0.112 36 4.190 3.76 4.50 0.158 48 4.320 3.77 4.95 0.200 72 4.333 3.77 5.20 0.188 19 4.189 3.87 4.39 0.127
LMd/I1M3 18 1.507 1.47 1.53 0.018 35 1.508 1.46 1.57 0.020 48 1.493 1.44 1.57 0.026 75 1.480 1.37 1.55 0.027 19 1.505 1.48 1.53 0.013
LMd/CM3 18 1.773 1.74 1.80 0.016 35 1.756 1.71 1.83 0.026 48 1.758 1.69 1.81 0.025 74 1.756 1.65 1.82 0.022 19 1.744 1.72 1.77 0.016
LMd/P4M3 18 2.503 2.45 2.56 0.028 35 2.441 2.35 2.57 0.053 48 2.452 2.30 2.57 0.052 75 2.431 2.26 2.55 0.043 19 2.414 2.36 2.47 0.030
LMd/M1M3 18 3.044 2.93 3.11 0.046 36 2.919 2.69 3.06 0.081 48 2.904 2.68 3.10 0.095 75 2.880 2.68 3.12 0.081 19 2.879 2.78 3.03 0.065
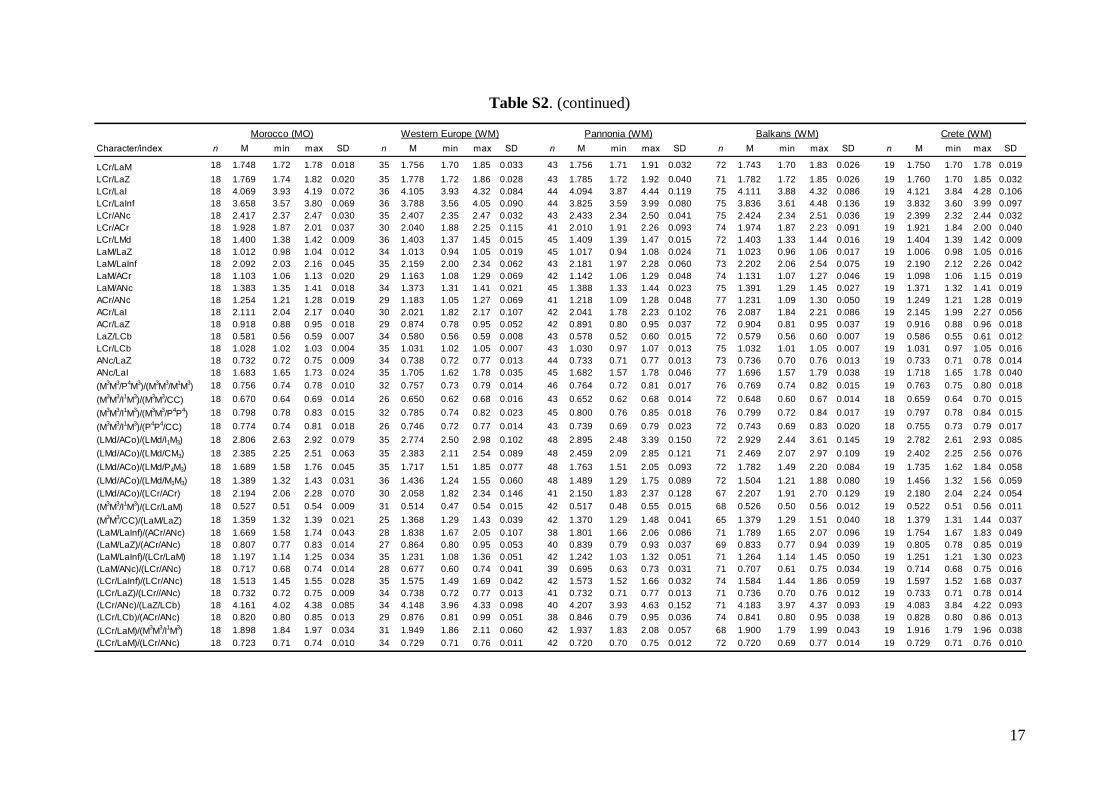
17
Table S2. (continued)
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
LCr/LaM 18 1.748 1.72 1.78 0.018 35 1.756 1.70 1.85 0.033 43 1.756 1.71 1.91 0.032 72 1.743 1.70 1.83 0.026 19 1.750 1.70 1.78 0.019
LCr/LaZ 18 1.769 1.74 1.82 0.020 35 1.778 1.72 1.86 0.028 43 1.785 1.72 1.92 0.040 71 1.782 1.72 1.85 0.026 19 1.760 1.70 1.85 0.032
LCr/LaI 18 4.069 3.93 4.19 0.072 36 4.105 3.93 4.32 0.084 44 4.094 3.87 4.44 0.119 75 4.111 3.88 4.32 0.086 19 4.121 3.84 4.28 0.106
LCr/LaInf 18 3.658 3.57 3.80 0.069 36 3.788 3.56 4.05 0.090 44 3.825 3.59 3.99 0.080 75 3.836 3.61 4.48 0.136 19 3.832 3.60 3.99 0.097
LCr/ANc 18 2.417 2.37 2.47 0.030 35 2.407 2.35 2.47 0.032 43 2.433 2.34 2.50 0.041 75 2.424 2.34 2.51 0.036 19 2.399 2.32 2.44 0.032
LCr/ACr 18 1.928 1.87 2.01 0.037 30 2.040 1.88 2.25 0.115 41 2.010 1.91 2.26 0.093 74 1.974 1.87 2.23 0.091 19 1.921 1.84 2.00 0.040
LCr/LMd 18 1.400 1.38 1.42 0.009 36 1.403 1.37 1.45 0.015 45 1.409 1.39 1.47 0.015 72 1.403 1.33 1.44 0.016 19 1.404 1.39 1.42 0.009
LaM/LaZ 18 1.012 0.98 1.04 0.012 34 1.013 0.94 1.05 0.019 45 1.017 0.94 1.08 0.024 71 1.023 0.96 1.06 0.017 19 1.006 0.98 1.05 0.016
LaM/LaInf 18 2.092 2.03 2.16 0.045 35 2.159 2.00 2.34 0.062 43 2.181 1.97 2.28 0.060 73 2.202 2.06 2.54 0.075 19 2.190 2.12 2.26 0.042
LaM/ACr 18 1.103 1.06 1.13 0.020 29 1.163 1.08 1.29 0.069 42 1.142 1.06 1.29 0.048 74 1.131 1.07 1.27 0.046 19 1.098 1.06 1.15 0.019
LaM/ANc 18 1.383 1.35 1.41 0.018 34 1.373 1.31 1.41 0.021 45 1.388 1.33 1.44 0.023 75 1.391 1.29 1.45 0.027 19 1.371 1.32 1.41 0.019
ACr/ANc 18 1.254 1.21 1.28 0.019 29 1.183 1.05 1.27 0.069 41 1.218 1.09 1.28 0.048 77 1.231 1.09 1.30 0.050 19 1.249 1.21 1.28 0.019
ACr/LaI 18 2.111 2.04 2.17 0.040 30 2.021 1.82 2.17 0.107 42 2.041 1.78 2.23 0.102 76 2.087 1.84 2.21 0.086 19 2.145 1.99 2.27 0.056
ACr/LaZ 18 0.918 0.88 0.95 0.018 29 0.874 0.78 0.95 0.052 42 0.891 0.80 0.95 0.037 72 0.904 0.81 0.95 0.037 19 0.916 0.88 0.96 0.018
LaZ/LCb 18 0.581 0.56 0.59 0.007 34 0.580 0.56 0.59 0.008 43 0.578 0.52 0.60 0.015 72 0.579 0.56 0.60 0.007 19 0.586 0.55 0.61 0.012
LCr/LCb 18 1.028 1.02 1.03 0.004 35 1.031 1.02 1.05 0.007 43 1.030 0.97 1.07 0.013 75 1.032 1.01 1.05 0.007 19 1.031 0.97 1.05 0.016
ANc/LaZ 18 0.732 0.72 0.75 0.009 34 0.738 0.72 0.77 0.013 44 0.733 0.71 0.77 0.013 73 0.736 0.70 0.76 0.013 19 0.733 0.71 0.78 0.014
ANc/LaI 18 1.683 1.65 1.73 0.024 35 1.705 1.62 1.78 0.035 45 1.682 1.57 1.78 0.046 77 1.696 1.57 1.79 0.038 19 1.718 1.65 1.78 0.040
(M3M
3/P
4M
3)/(M
3M
3/M
1M
3) 18 0.756 0.74 0.78 0.010 32 0.757 0.73 0.79 0.014 46 0.764 0.72 0.81 0.017 76 0.769 0.74 0.82 0.015 19 0.763 0.75 0.80 0.018
(M3M
3/I
1M
3)/(M
3M
3/CC) 18 0.670 0.64 0.69 0.014 26 0.650 0.62 0.68 0.016 43 0.652 0.62 0.68 0.014 72 0.648 0.60 0.67 0.014 18 0.659 0.64 0.70 0.015
(M3M
3/I
1M
3)/(M
3M
3/P
4P
4) 18 0.798 0.78 0.83 0.015 32 0.785 0.74 0.82 0.023 45 0.800 0.76 0.85 0.018 76 0.799 0.72 0.84 0.017 19 0.797 0.78 0.84 0.015
(M3M
3/I
1M
3)/(P
4P
4/CC) 18 0.774 0.74 0.81 0.018 26 0.746 0.72 0.77 0.014 43 0.739 0.69 0.79 0.023 72 0.743 0.69 0.83 0.020 18 0.755 0.73 0.79 0.017
(LMd/ACo)/(LMd/I1M3) 18 2.806 2.63 2.92 0.079 35 2.774 2.50 2.98 0.102 48 2.895 2.48 3.39 0.150 72 2.929 2.44 3.61 0.145 19 2.782 2.61 2.93 0.085
(LMd/ACo)/(LMd/CM3) 18 2.385 2.25 2.51 0.063 35 2.383 2.11 2.54 0.089 48 2.459 2.09 2.85 0.121 71 2.469 2.07 2.97 0.109 19 2.402 2.25 2.56 0.076
(LMd/ACo)/(LMd/P4M3) 18 1.689 1.58 1.76 0.045 35 1.717 1.51 1.85 0.077 48 1.763 1.51 2.05 0.093 72 1.782 1.49 2.20 0.084 19 1.735 1.62 1.84 0.058
(LMd/ACo)/(LMd/M1M3) 18 1.389 1.32 1.43 0.031 36 1.436 1.24 1.55 0.060 48 1.489 1.29 1.75 0.089 72 1.504 1.21 1.88 0.080 19 1.456 1.32 1.56 0.059
(LMd/ACo)/(LCr/ACr) 18 2.194 2.06 2.28 0.070 30 2.058 1.82 2.34 0.146 41 2.150 1.83 2.37 0.128 67 2.207 1.91 2.70 0.129 19 2.180 2.04 2.24 0.054
(M3M
3/I
1M
3)/(LCr/LaM) 18 0.527 0.51 0.54 0.009 31 0.514 0.47 0.54 0.015 42 0.517 0.48 0.55 0.015 68 0.526 0.50 0.56 0.012 19 0.522 0.51 0.56 0.011
(M3M
3/CC)/(LaM/LaZ) 18 1.359 1.32 1.39 0.021 25 1.368 1.29 1.43 0.039 42 1.370 1.29 1.48 0.041 65 1.379 1.29 1.51 0.040 18 1.379 1.31 1.44 0.037
(LaM/LaInf)/(ACr/ANc) 18 1.669 1.58 1.74 0.043 28 1.838 1.67 2.05 0.107 38 1.801 1.66 2.06 0.086 71 1.789 1.65 2.07 0.096 19 1.754 1.67 1.83 0.049
(LaM/LaZ)/(ACr/ANc) 18 0.807 0.77 0.83 0.014 27 0.864 0.80 0.95 0.053 40 0.839 0.79 0.93 0.037 69 0.833 0.77 0.94 0.039 19 0.805 0.78 0.85 0.019
(LaM/LaInf)/(LCr/LaM) 18 1.197 1.14 1.25 0.034 35 1.231 1.08 1.36 0.051 42 1.242 1.03 1.32 0.051 71 1.264 1.14 1.45 0.050 19 1.251 1.21 1.30 0.023
(LaM/ANc)/(LCr/ANc) 18 0.717 0.68 0.74 0.014 28 0.677 0.60 0.74 0.041 39 0.695 0.63 0.73 0.031 71 0.707 0.61 0.75 0.034 19 0.714 0.68 0.75 0.016
(LCr/LaInf)/(LCr/ANc) 18 1.513 1.45 1.55 0.028 35 1.575 1.49 1.69 0.042 42 1.573 1.52 1.66 0.032 74 1.584 1.44 1.86 0.059 19 1.597 1.52 1.68 0.037
(LCr/LaZ)/(LCr//ANc) 18 0.732 0.72 0.75 0.009 34 0.738 0.72 0.77 0.013 41 0.732 0.71 0.77 0.013 71 0.736 0.70 0.76 0.012 19 0.733 0.71 0.78 0.014
(LCr/ANc)/(LaZ/LCb) 18 4.161 4.02 4.38 0.085 34 4.148 3.96 4.33 0.098 40 4.207 3.93 4.63 0.152 71 4.183 3.97 4.37 0.093 19 4.083 3.84 4.22 0.093
(LCr/LCb)/(ACr/ANc) 18 0.820 0.80 0.85 0.013 29 0.876 0.81 0.99 0.051 38 0.846 0.79 0.95 0.036 74 0.841 0.80 0.95 0.038 19 0.828 0.80 0.86 0.013
(LCr/LaM)/(M3M
3/I
1M
3) 18 1.898 1.84 1.97 0.034 31 1.949 1.86 2.11 0.060 42 1.937 1.83 2.08 0.057 68 1.900 1.79 1.99 0.043 19 1.916 1.79 1.96 0.038
(LCr/LaM)/(LCr/ANc) 18 0.723 0.71 0.74 0.010 34 0.729 0.71 0.76 0.011 42 0.720 0.70 0.75 0.012 72 0.720 0.69 0.77 0.014 19 0.729 0.71 0.76 0.010
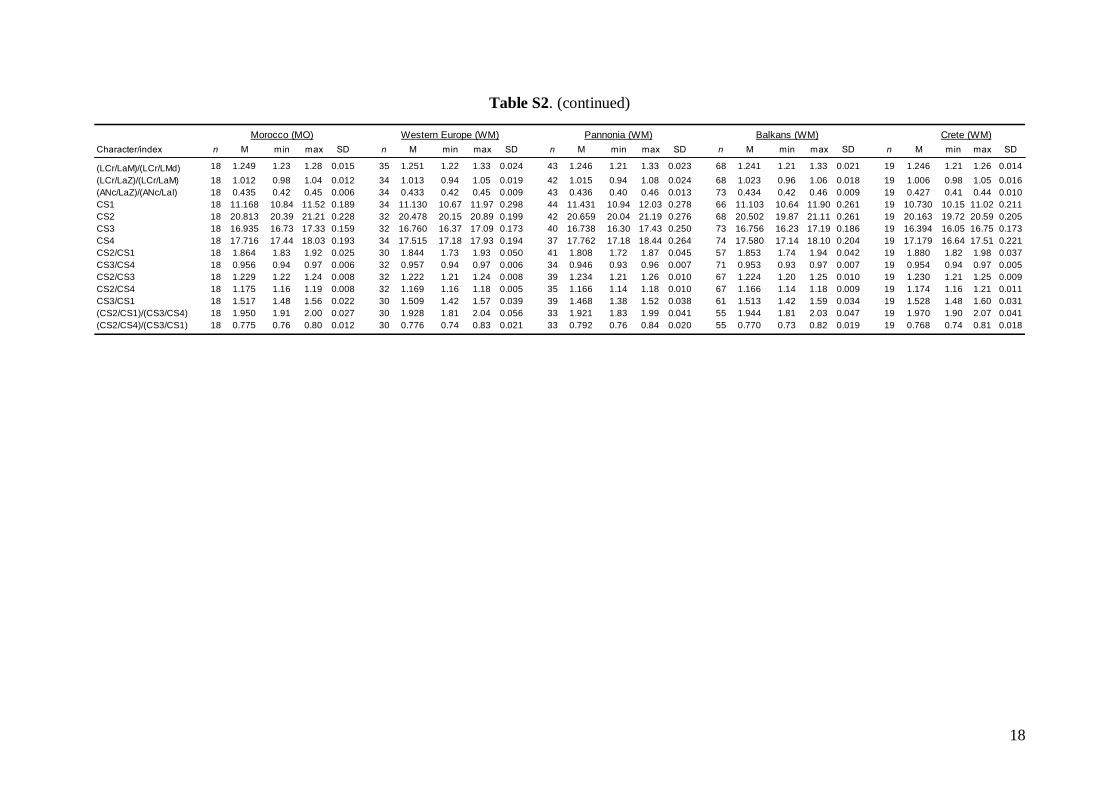
18
Table S2. (continued)
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
(LCr/LaM)/(LCr/LMd) 18 1.249 1.23 1.28 0.015 35 1.251 1.22 1.33 0.024 43 1.246 1.21 1.33 0.023 68 1.241 1.21 1.33 0.021 19 1.246 1.21 1.26 0.014
(LCr/LaZ)/(LCr/LaM) 18 1.012 0.98 1.04 0.012 34 1.013 0.94 1.05 0.019 42 1.015 0.94 1.08 0.024 68 1.023 0.96 1.06 0.018 19 1.006 0.98 1.05 0.016
(ANc/LaZ)/(ANc/LaI) 18 0.435 0.42 0.45 0.006 34 0.433 0.42 0.45 0.009 43 0.436 0.40 0.46 0.013 73 0.434 0.42 0.46 0.009 19 0.427 0.41 0.44 0.010
CS1 18 11.168 10.84 11.52 0.189 34 11.130 10.67 11.97 0.298 44 11.431 10.94 12.03 0.278 66 11.103 10.64 11.90 0.261 19 10.730 10.15 11.02 0.211
CS2 18 20.813 20.39 21.21 0.228 32 20.478 20.15 20.89 0.199 42 20.659 20.04 21.19 0.276 68 20.502 19.87 21.11 0.261 19 20.163 19.72 20.59 0.205
CS3 18 16.935 16.73 17.33 0.159 32 16.760 16.37 17.09 0.173 40 16.738 16.30 17.43 0.250 73 16.756 16.23 17.19 0.186 19 16.394 16.05 16.75 0.173
CS4 18 17.716 17.44 18.03 0.193 34 17.515 17.18 17.93 0.194 37 17.762 17.18 18.44 0.264 74 17.580 17.14 18.10 0.204 19 17.179 16.64 17.51 0.221
CS2/CS1 18 1.864 1.83 1.92 0.025 30 1.844 1.73 1.93 0.050 41 1.808 1.72 1.87 0.045 57 1.853 1.74 1.94 0.042 19 1.880 1.82 1.98 0.037
CS3/CS4 18 0.956 0.94 0.97 0.006 32 0.957 0.94 0.97 0.006 34 0.946 0.93 0.96 0.007 71 0.953 0.93 0.97 0.007 19 0.954 0.94 0.97 0.005
CS2/CS3 18 1.229 1.22 1.24 0.008 32 1.222 1.21 1.24 0.008 39 1.234 1.21 1.26 0.010 67 1.224 1.20 1.25 0.010 19 1.230 1.21 1.25 0.009
CS2/CS4 18 1.175 1.16 1.19 0.008 32 1.169 1.16 1.18 0.005 35 1.166 1.14 1.18 0.010 67 1.166 1.14 1.18 0.009 19 1.174 1.16 1.21 0.011
CS3/CS1 18 1.517 1.48 1.56 0.022 30 1.509 1.42 1.57 0.039 39 1.468 1.38 1.52 0.038 61 1.513 1.42 1.59 0.034 19 1.528 1.48 1.60 0.031
(CS2/CS1)/(CS3/CS4) 18 1.950 1.91 2.00 0.027 30 1.928 1.81 2.04 0.056 33 1.921 1.83 1.99 0.041 55 1.944 1.81 2.03 0.047 19 1.970 1.90 2.07 0.041
(CS2/CS4)/(CS3/CS1) 18 0.775 0.76 0.80 0.012 30 0.776 0.74 0.83 0.021 33 0.792 0.76 0.84 0.020 55 0.770 0.73 0.82 0.019 19 0.768 0.74 0.81 0.018
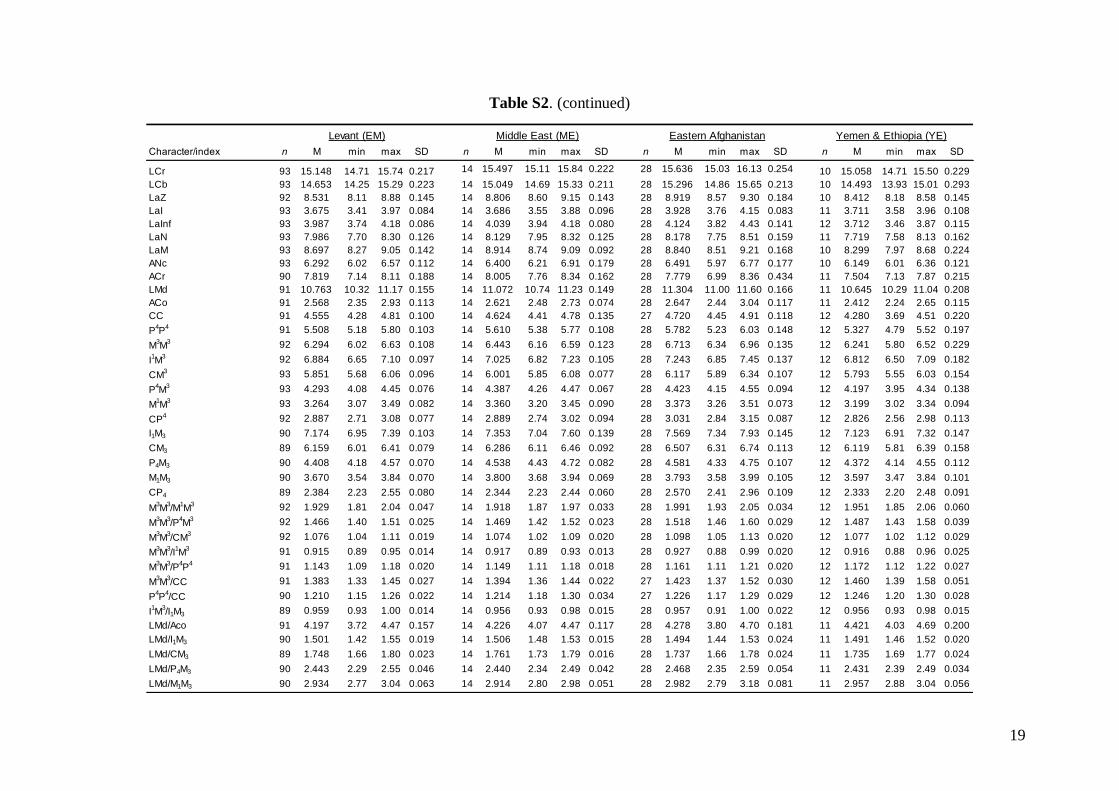
19
Table S2. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
LCr 93 15.148 14.71 15.74 0.217 14 15.497 15.11 15.84 0.222 28 15.636 15.03 16.13 0.254 10 15.058 14.71 15.50 0.229
LCb 93 14.653 14.25 15.29 0.223 14 15.049 14.69 15.33 0.211 28 15.296 14.86 15.65 0.213 10 14.493 13.93 15.01 0.293
LaZ 92 8.531 8.11 8.88 0.145 14 8.806 8.60 9.15 0.143 28 8.919 8.57 9.30 0.184 10 8.412 8.18 8.58 0.145
LaI 93 3.675 3.41 3.97 0.084 14 3.686 3.55 3.88 0.096 28 3.928 3.76 4.15 0.083 11 3.711 3.58 3.96 0.108
LaInf 93 3.987 3.74 4.18 0.086 14 4.039 3.94 4.18 0.080 28 4.124 3.82 4.43 0.141 12 3.712 3.46 3.87 0.115
LaN 93 7.986 7.70 8.30 0.126 14 8.129 7.95 8.32 0.125 28 8.178 7.75 8.51 0.159 11 7.719 7.58 8.13 0.162
LaM 93 8.697 8.27 9.05 0.142 14 8.914 8.74 9.09 0.092 28 8.840 8.51 9.21 0.168 10 8.299 7.97 8.68 0.224
ANc 93 6.292 6.02 6.57 0.112 14 6.400 6.21 6.91 0.179 28 6.491 5.97 6.77 0.177 10 6.149 6.01 6.36 0.121
ACr 90 7.819 7.14 8.11 0.188 14 8.005 7.76 8.34 0.162 28 7.779 6.99 8.36 0.434 11 7.504 7.13 7.87 0.215
LMd 91 10.763 10.32 11.17 0.155 14 11.072 10.74 11.23 0.149 28 11.304 11.00 11.60 0.166 11 10.645 10.29 11.04 0.208
ACo 91 2.568 2.35 2.93 0.113 14 2.621 2.48 2.73 0.074 28 2.647 2.44 3.04 0.117 11 2.412 2.24 2.65 0.115
CC 91 4.555 4.28 4.81 0.100 14 4.624 4.41 4.78 0.135 27 4.720 4.45 4.91 0.118 12 4.280 3.69 4.51 0.220
P4P
4 91 5.508 5.18 5.80 0.103 14 5.610 5.38 5.77 0.108 28 5.782 5.23 6.03 0.148 12 5.327 4.79 5.52 0.197
M3M
3 92 6.294 6.02 6.63 0.108 14 6.443 6.16 6.59 0.123 28 6.713 6.34 6.96 0.135 12 6.241 5.80 6.52 0.229
I1M
3 92 6.884 6.65 7.10 0.097 14 7.025 6.82 7.23 0.105 28 7.243 6.85 7.45 0.137 12 6.812 6.50 7.09 0.182
CM3 93 5.851 5.68 6.06 0.096 14 6.001 5.85 6.08 0.077 28 6.117 5.89 6.34 0.107 12 5.793 5.55 6.03 0.154
P4M
3 93 4.293 4.08 4.45 0.076 14 4.387 4.26 4.47 0.067 28 4.423 4.15 4.55 0.094 12 4.197 3.95 4.34 0.138
M1M
3 93 3.264 3.07 3.49 0.082 14 3.360 3.20 3.45 0.090 28 3.373 3.26 3.51 0.073 12 3.199 3.02 3.34 0.094
CP4 92 2.887 2.71 3.08 0.077 14 2.889 2.74 3.02 0.094 28 3.031 2.84 3.15 0.087 12 2.826 2.56 2.98 0.113
I1M3 90 7.174 6.95 7.39 0.103 14 7.353 7.04 7.60 0.139 28 7.569 7.34 7.93 0.145 12 7.123 6.91 7.32 0.147
CM3 89 6.159 6.01 6.41 0.079 14 6.286 6.11 6.46 0.092 28 6.507 6.31 6.74 0.113 12 6.119 5.81 6.39 0.158
P4M3 90 4.408 4.18 4.57 0.070 14 4.538 4.43 4.72 0.082 28 4.581 4.33 4.75 0.107 12 4.372 4.14 4.55 0.112
M1M3 90 3.670 3.54 3.84 0.070 14 3.800 3.68 3.94 0.069 28 3.793 3.58 3.99 0.105 12 3.597 3.47 3.84 0.101
CP4 89 2.384 2.23 2.55 0.080 14 2.344 2.23 2.44 0.060 28 2.570 2.41 2.96 0.109 12 2.333 2.20 2.48 0.091
M3M
3/M
1M
3 92 1.929 1.81 2.04 0.047 14 1.918 1.87 1.97 0.033 28 1.991 1.93 2.05 0.034 12 1.951 1.85 2.06 0.060
M3M
3/P
4M
3 92 1.466 1.40 1.51 0.025 14 1.469 1.42 1.52 0.023 28 1.518 1.46 1.60 0.029 12 1.487 1.43 1.58 0.039
M3M
3/CM
3 92 1.076 1.04 1.11 0.019 14 1.074 1.02 1.09 0.020 28 1.098 1.05 1.13 0.020 12 1.077 1.02 1.12 0.029
M3M
3/I
1M
3 91 0.915 0.89 0.95 0.014 14 0.917 0.89 0.93 0.013 28 0.927 0.88 0.99 0.020 12 0.916 0.88 0.96 0.025
M3M
3/P
4P
4 91 1.143 1.09 1.18 0.020 14 1.149 1.11 1.18 0.018 28 1.161 1.11 1.21 0.020 12 1.172 1.12 1.22 0.027
M3M
3/CC 91 1.383 1.33 1.45 0.027 14 1.394 1.36 1.44 0.022 27 1.423 1.37 1.52 0.030 12 1.460 1.39 1.58 0.051
P4P
4/CC 90 1.210 1.15 1.26 0.022 14 1.214 1.18 1.30 0.034 27 1.226 1.17 1.29 0.029 12 1.246 1.20 1.30 0.028
I1M
3/I1M3 89 0.959 0.93 1.00 0.014 14 0.956 0.93 0.98 0.015 28 0.957 0.91 1.00 0.022 12 0.956 0.93 0.98 0.015
LMd/Aco 91 4.197 3.72 4.47 0.157 14 4.226 4.07 4.47 0.117 28 4.278 3.80 4.70 0.181 11 4.421 4.03 4.69 0.200
LMd/I1M3 90 1.501 1.42 1.55 0.019 14 1.506 1.48 1.53 0.015 28 1.494 1.44 1.53 0.024 11 1.491 1.46 1.52 0.020
LMd/CM3 89 1.748 1.66 1.80 0.023 14 1.761 1.73 1.79 0.016 28 1.737 1.66 1.78 0.024 11 1.735 1.69 1.77 0.024
LMd/P4M3 90 2.443 2.29 2.55 0.046 14 2.440 2.34 2.49 0.042 28 2.468 2.35 2.59 0.054 11 2.431 2.39 2.49 0.034
LMd/M1M3 90 2.934 2.77 3.04 0.063 14 2.914 2.80 2.98 0.051 28 2.982 2.79 3.18 0.081 11 2.957 2.88 3.04 0.056
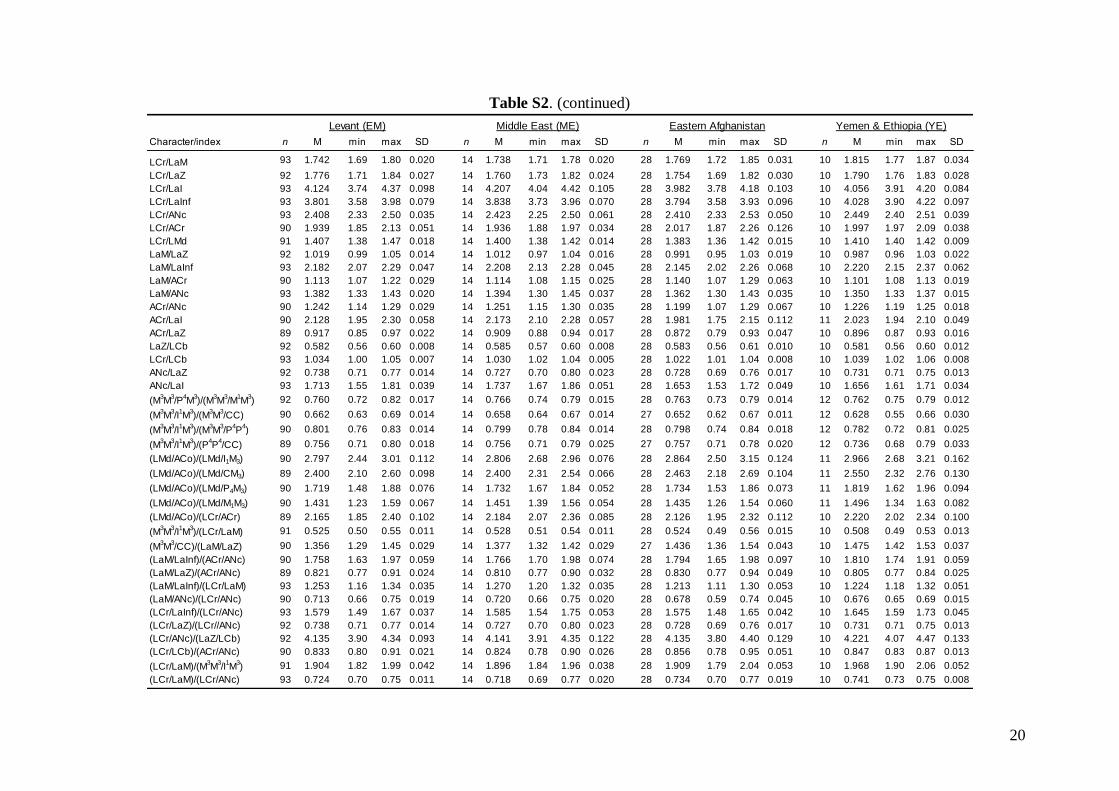
20
Table S2. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
LCr/LaM 93 1.742 1.69 1.80 0.020 14 1.738 1.71 1.78 0.020 28 1.769 1.72 1.85 0.031 10 1.815 1.77 1.87 0.034
LCr/LaZ 92 1.776 1.71 1.84 0.027 14 1.760 1.73 1.82 0.024 28 1.754 1.69 1.82 0.030 10 1.790 1.76 1.83 0.028
LCr/LaI 93 4.124 3.74 4.37 0.098 14 4.207 4.04 4.42 0.105 28 3.982 3.78 4.18 0.103 10 4.056 3.91 4.20 0.084
LCr/LaInf 93 3.801 3.58 3.98 0.079 14 3.838 3.73 3.96 0.070 28 3.794 3.58 3.93 0.096 10 4.028 3.90 4.22 0.097
LCr/ANc 93 2.408 2.33 2.50 0.035 14 2.423 2.25 2.50 0.061 28 2.410 2.33 2.53 0.050 10 2.449 2.40 2.51 0.039
LCr/ACr 90 1.939 1.85 2.13 0.051 14 1.936 1.88 1.97 0.034 28 2.017 1.87 2.26 0.126 10 1.997 1.97 2.09 0.038
LCr/LMd 91 1.407 1.38 1.47 0.018 14 1.400 1.38 1.42 0.014 28 1.383 1.36 1.42 0.015 10 1.410 1.40 1.42 0.009
LaM/LaZ 92 1.019 0.99 1.05 0.014 14 1.012 0.97 1.04 0.016 28 0.991 0.95 1.03 0.019 10 0.987 0.96 1.03 0.022
LaM/LaInf 93 2.182 2.07 2.29 0.047 14 2.208 2.13 2.28 0.045 28 2.145 2.02 2.26 0.068 10 2.220 2.15 2.37 0.062
LaM/ACr 90 1.113 1.07 1.22 0.029 14 1.114 1.08 1.15 0.025 28 1.140 1.07 1.29 0.063 10 1.101 1.08 1.13 0.019
LaM/ANc 93 1.382 1.33 1.43 0.020 14 1.394 1.30 1.45 0.037 28 1.362 1.30 1.43 0.035 10 1.350 1.33 1.37 0.015
ACr/ANc 90 1.242 1.14 1.29 0.029 14 1.251 1.15 1.30 0.035 28 1.199 1.07 1.29 0.067 10 1.226 1.19 1.25 0.018
ACr/LaI 90 2.128 1.95 2.30 0.058 14 2.173 2.10 2.28 0.057 28 1.981 1.75 2.15 0.112 11 2.023 1.94 2.10 0.049
ACr/LaZ 89 0.917 0.85 0.97 0.022 14 0.909 0.88 0.94 0.017 28 0.872 0.79 0.93 0.047 10 0.896 0.87 0.93 0.016
LaZ/LCb 92 0.582 0.56 0.60 0.008 14 0.585 0.57 0.60 0.008 28 0.583 0.56 0.61 0.010 10 0.581 0.56 0.60 0.012
LCr/LCb 93 1.034 1.00 1.05 0.007 14 1.030 1.02 1.04 0.005 28 1.022 1.01 1.04 0.008 10 1.039 1.02 1.06 0.008
ANc/LaZ 92 0.738 0.71 0.77 0.014 14 0.727 0.70 0.80 0.023 28 0.728 0.69 0.76 0.017 10 0.731 0.71 0.75 0.013
ANc/LaI 93 1.713 1.55 1.81 0.039 14 1.737 1.67 1.86 0.051 28 1.653 1.53 1.72 0.049 10 1.656 1.61 1.71 0.034
(M3M
3/P
4M
3)/(M
3M
3/M
1M
3) 92 0.760 0.72 0.82 0.017 14 0.766 0.74 0.79 0.015 28 0.763 0.73 0.79 0.014 12 0.762 0.75 0.79 0.012
(M3M
3/I
1M
3)/(M
3M
3/CC) 90 0.662 0.63 0.69 0.014 14 0.658 0.64 0.67 0.014 27 0.652 0.62 0.67 0.011 12 0.628 0.55 0.66 0.030
(M3M
3/I
1M
3)/(M
3M
3/P
4P
4) 90 0.801 0.76 0.83 0.014 14 0.799 0.78 0.84 0.014 28 0.798 0.74 0.84 0.018 12 0.782 0.72 0.81 0.025
(M3M
3/I
1M
3)/(P
4P
4/CC) 89 0.756 0.71 0.80 0.018 14 0.756 0.71 0.79 0.025 27 0.757 0.71 0.78 0.020 12 0.736 0.68 0.79 0.033
(LMd/ACo)/(LMd/I1M3) 90 2.797 2.44 3.01 0.112 14 2.806 2.68 2.96 0.076 28 2.864 2.50 3.15 0.124 11 2.966 2.68 3.21 0.162
(LMd/ACo)/(LMd/CM3) 89 2.400 2.10 2.60 0.098 14 2.400 2.31 2.54 0.066 28 2.463 2.18 2.69 0.104 11 2.550 2.32 2.76 0.130
(LMd/ACo)/(LMd/P4M3) 90 1.719 1.48 1.88 0.076 14 1.732 1.67 1.84 0.052 28 1.734 1.53 1.86 0.073 11 1.819 1.62 1.96 0.094
(LMd/ACo)/(LMd/M1M3) 90 1.431 1.23 1.59 0.067 14 1.451 1.39 1.56 0.054 28 1.435 1.26 1.54 0.060 11 1.496 1.34 1.63 0.082
(LMd/ACo)/(LCr/ACr) 89 2.165 1.85 2.40 0.102 14 2.184 2.07 2.36 0.085 28 2.126 1.95 2.32 0.112 10 2.220 2.02 2.34 0.100
(M3M
3/I
1M
3)/(LCr/LaM) 91 0.525 0.50 0.55 0.011 14 0.528 0.51 0.54 0.011 28 0.524 0.49 0.56 0.015 10 0.508 0.49 0.53 0.013
(M3M
3/CC)/(LaM/LaZ) 90 1.356 1.29 1.45 0.029 14 1.377 1.32 1.42 0.029 27 1.436 1.36 1.54 0.043 10 1.475 1.42 1.53 0.037
(LaM/LaInf)/(ACr/ANc) 90 1.758 1.63 1.97 0.059 14 1.766 1.70 1.98 0.074 28 1.794 1.65 1.98 0.097 10 1.810 1.74 1.91 0.059
(LaM/LaZ)/(ACr/ANc) 89 0.821 0.77 0.91 0.024 14 0.810 0.77 0.90 0.032 28 0.830 0.77 0.94 0.049 10 0.805 0.77 0.84 0.025
(LaM/LaInf)/(LCr/LaM) 93 1.253 1.16 1.34 0.035 14 1.270 1.20 1.32 0.035 28 1.213 1.11 1.30 0.053 10 1.224 1.18 1.32 0.051
(LaM/ANc)/(LCr/ANc) 90 0.713 0.66 0.75 0.019 14 0.720 0.66 0.75 0.020 28 0.678 0.59 0.74 0.045 10 0.676 0.65 0.69 0.015
(LCr/LaInf)/(LCr/ANc) 93 1.579 1.49 1.67 0.037 14 1.585 1.54 1.75 0.053 28 1.575 1.48 1.65 0.042 10 1.645 1.59 1.73 0.045
(LCr/LaZ)/(LCr//ANc) 92 0.738 0.71 0.77 0.014 14 0.727 0.70 0.80 0.023 28 0.728 0.69 0.76 0.017 10 0.731 0.71 0.75 0.013
(LCr/ANc)/(LaZ/LCb) 92 4.135 3.90 4.34 0.093 14 4.141 3.91 4.35 0.122 28 4.135 3.80 4.40 0.129 10 4.221 4.07 4.47 0.133
(LCr/LCb)/(ACr/ANc) 90 0.833 0.80 0.91 0.021 14 0.824 0.78 0.90 0.026 28 0.856 0.78 0.95 0.051 10 0.847 0.83 0.87 0.013
(LCr/LaM)/(M3M
3/I
1M
3) 91 1.904 1.82 1.99 0.042 14 1.896 1.84 1.96 0.038 28 1.909 1.79 2.04 0.053 10 1.968 1.90 2.06 0.052
(LCr/LaM)/(LCr/ANc) 93 0.724 0.70 0.75 0.011 14 0.718 0.69 0.77 0.020 28 0.734 0.70 0.77 0.019 10 0.741 0.73 0.75 0.008
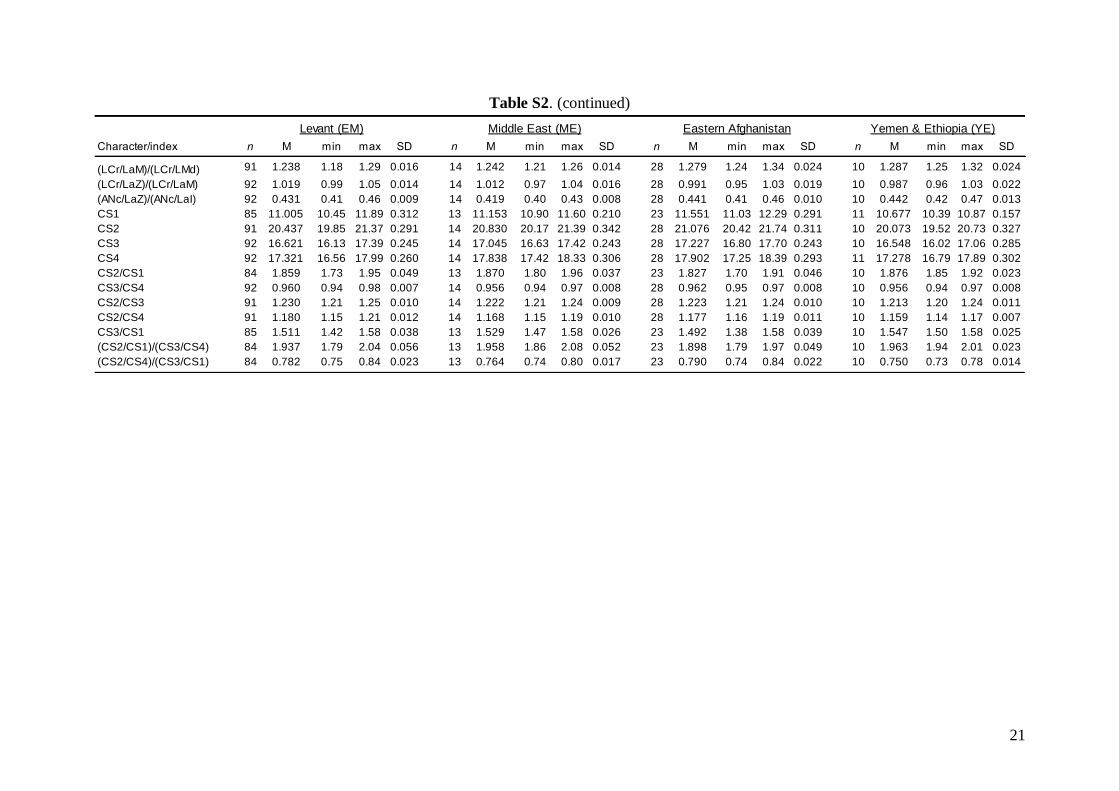
21
Table S2. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
(LCr/LaM)/(LCr/LMd) 91 1.238 1.18 1.29 0.016 14 1.242 1.21 1.26 0.014 28 1.279 1.24 1.34 0.024 10 1.287 1.25 1.32 0.024
(LCr/LaZ)/(LCr/LaM) 92 1.019 0.99 1.05 0.014 14 1.012 0.97 1.04 0.016 28 0.991 0.95 1.03 0.019 10 0.987 0.96 1.03 0.022
(ANc/LaZ)/(ANc/LaI) 92 0.431 0.41 0.46 0.009 14 0.419 0.40 0.43 0.008 28 0.441 0.41 0.46 0.010 10 0.442 0.42 0.47 0.013
CS1 85 11.005 10.45 11.89 0.312 13 11.153 10.90 11.60 0.210 23 11.551 11.03 12.29 0.291 11 10.677 10.39 10.87 0.157
CS2 91 20.437 19.85 21.37 0.291 14 20.830 20.17 21.39 0.342 28 21.076 20.42 21.74 0.311 10 20.073 19.52 20.73 0.327
CS3 92 16.621 16.13 17.39 0.245 14 17.045 16.63 17.42 0.243 28 17.227 16.80 17.70 0.243 10 16.548 16.02 17.06 0.285
CS4 92 17.321 16.56 17.99 0.260 14 17.838 17.42 18.33 0.306 28 17.902 17.25 18.39 0.293 11 17.278 16.79 17.89 0.302
CS2/CS1 84 1.859 1.73 1.95 0.049 13 1.870 1.80 1.96 0.037 23 1.827 1.70 1.91 0.046 10 1.876 1.85 1.92 0.023
CS3/CS4 92 0.960 0.94 0.98 0.007 14 0.956 0.94 0.97 0.008 28 0.962 0.95 0.97 0.008 10 0.956 0.94 0.97 0.008
CS2/CS3 91 1.230 1.21 1.25 0.010 14 1.222 1.21 1.24 0.009 28 1.223 1.21 1.24 0.010 10 1.213 1.20 1.24 0.011
CS2/CS4 91 1.180 1.15 1.21 0.012 14 1.168 1.15 1.19 0.010 28 1.177 1.16 1.19 0.011 10 1.159 1.14 1.17 0.007
CS3/CS1 85 1.511 1.42 1.58 0.038 13 1.529 1.47 1.58 0.026 23 1.492 1.38 1.58 0.039 10 1.547 1.50 1.58 0.025
(CS2/CS1)/(CS3/CS4) 84 1.937 1.79 2.04 0.056 13 1.958 1.86 2.08 0.052 23 1.898 1.79 1.97 0.049 10 1.963 1.94 2.01 0.023
(CS2/CS4)/(CS3/CS1) 84 0.782 0.75 0.84 0.023 13 0.764 0.74 0.80 0.017 23 0.790 0.74 0.84 0.022 10 0.750 0.73 0.78 0.014
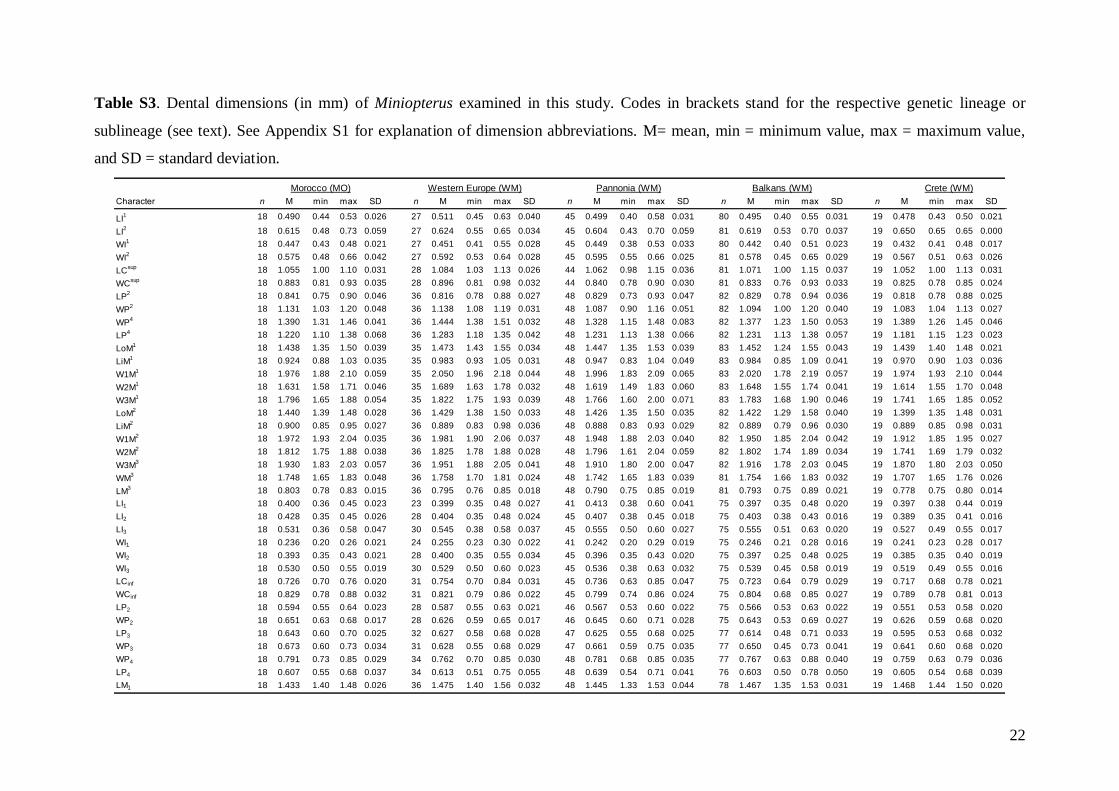
22
Table S3. Dental dimensions (in mm) of Miniopterus examined in this study. Codes in brackets stand for the respective genetic lineage or
sublineage (see text). See Appendix S1 for explanation of dimension abbreviations. M= mean, min = minimum value, max = maximum value,
and SD = standard deviation.
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
LI1 18 0.490 0.44 0.53 0.026 27 0.511 0.45 0.63 0.040 45 0.499 0.40 0.58 0.031 80 0.495 0.40 0.55 0.031 19 0.478 0.43 0.50 0.021
LI2 18 0.615 0.48 0.73 0.059 27 0.624 0.55 0.65 0.034 45 0.604 0.43 0.70 0.059 81 0.619 0.53 0.70 0.037 19 0.650 0.65 0.65 0.000
WI1 18 0.447 0.43 0.48 0.021 27 0.451 0.41 0.55 0.028 45 0.449 0.38 0.53 0.033 80 0.442 0.40 0.51 0.023 19 0.432 0.41 0.48 0.017
WI2 18 0.575 0.48 0.66 0.042 27 0.592 0.53 0.64 0.028 45 0.595 0.55 0.66 0.025 81 0.578 0.45 0.65 0.029 19 0.567 0.51 0.63 0.026
LCsup 18 1.055 1.00 1.10 0.031 28 1.084 1.03 1.13 0.026 44 1.062 0.98 1.15 0.036 81 1.071 1.00 1.15 0.037 19 1.052 1.00 1.13 0.031
WCsup 18 0.883 0.81 0.93 0.035 28 0.896 0.81 0.98 0.032 44 0.840 0.78 0.90 0.030 81 0.833 0.76 0.93 0.033 19 0.825 0.78 0.85 0.024
LP2 18 0.841 0.75 0.90 0.046 36 0.816 0.78 0.88 0.027 48 0.829 0.73 0.93 0.047 82 0.829 0.78 0.94 0.036 19 0.818 0.78 0.88 0.025
WP2 18 1.131 1.03 1.20 0.048 36 1.138 1.08 1.19 0.031 48 1.087 0.90 1.16 0.051 82 1.094 1.00 1.20 0.040 19 1.083 1.04 1.13 0.027
WP4 18 1.390 1.31 1.46 0.041 36 1.444 1.38 1.51 0.032 48 1.328 1.15 1.48 0.083 82 1.377 1.23 1.50 0.053 19 1.389 1.26 1.45 0.046
LP4 18 1.220 1.10 1.38 0.068 36 1.283 1.18 1.35 0.042 48 1.231 1.13 1.38 0.066 82 1.231 1.13 1.38 0.057 19 1.181 1.15 1.23 0.023
LoM1 18 1.438 1.35 1.50 0.039 35 1.473 1.43 1.55 0.034 48 1.447 1.35 1.53 0.039 83 1.452 1.24 1.55 0.043 19 1.439 1.40 1.48 0.021
LiM1 18 0.924 0.88 1.03 0.035 35 0.983 0.93 1.05 0.031 48 0.947 0.83 1.04 0.049 83 0.984 0.85 1.09 0.041 19 0.970 0.90 1.03 0.036
W1M1 18 1.976 1.88 2.10 0.059 35 2.050 1.96 2.18 0.044 48 1.996 1.83 2.09 0.065 83 2.020 1.78 2.19 0.057 19 1.974 1.93 2.10 0.044
W2M1 18 1.631 1.58 1.71 0.046 35 1.689 1.63 1.78 0.032 48 1.619 1.49 1.83 0.060 83 1.648 1.55 1.74 0.041 19 1.614 1.55 1.70 0.048
W3M1 18 1.796 1.65 1.88 0.054 35 1.822 1.75 1.93 0.039 48 1.766 1.60 2.00 0.071 83 1.783 1.68 1.90 0.046 19 1.741 1.65 1.85 0.052
LoM2 18 1.440 1.39 1.48 0.028 36 1.429 1.38 1.50 0.033 48 1.426 1.35 1.50 0.035 82 1.422 1.29 1.58 0.040 19 1.399 1.35 1.48 0.031
LiM2 18 0.900 0.85 0.95 0.027 36 0.889 0.83 0.98 0.036 48 0.888 0.83 0.93 0.029 82 0.889 0.79 0.96 0.030 19 0.889 0.85 0.98 0.031
W1M2 18 1.972 1.93 2.04 0.035 36 1.981 1.90 2.06 0.037 48 1.948 1.88 2.03 0.040 82 1.950 1.85 2.04 0.042 19 1.912 1.85 1.95 0.027
W2M2 18 1.812 1.75 1.88 0.038 36 1.825 1.78 1.88 0.028 48 1.796 1.61 2.04 0.059 82 1.802 1.74 1.89 0.034 19 1.741 1.69 1.79 0.032
W3M3 18 1.930 1.83 2.03 0.057 36 1.951 1.88 2.05 0.041 48 1.910 1.80 2.00 0.047 82 1.916 1.78 2.03 0.045 19 1.870 1.80 2.03 0.050
WM3 18 1.748 1.65 1.83 0.048 36 1.758 1.70 1.81 0.024 48 1.742 1.65 1.83 0.039 81 1.754 1.66 1.83 0.032 19 1.707 1.65 1.76 0.026
LM3 18 0.803 0.78 0.83 0.015 36 0.795 0.76 0.85 0.018 48 0.790 0.75 0.85 0.019 81 0.793 0.75 0.89 0.021 19 0.778 0.75 0.80 0.014
LI1 18 0.400 0.36 0.45 0.023 23 0.399 0.35 0.48 0.027 41 0.413 0.38 0.60 0.041 75 0.397 0.35 0.48 0.020 19 0.397 0.38 0.44 0.019
LI2 18 0.428 0.35 0.45 0.026 28 0.404 0.35 0.48 0.024 45 0.407 0.38 0.45 0.018 75 0.403 0.38 0.43 0.016 19 0.389 0.35 0.41 0.016
LI3 18 0.531 0.36 0.58 0.047 30 0.545 0.38 0.58 0.037 45 0.555 0.50 0.60 0.027 75 0.555 0.51 0.63 0.020 19 0.527 0.49 0.55 0.017
WI1 18 0.236 0.20 0.26 0.021 24 0.255 0.23 0.30 0.022 41 0.242 0.20 0.29 0.019 75 0.246 0.21 0.28 0.016 19 0.241 0.23 0.28 0.017
WI2 18 0.393 0.35 0.43 0.021 28 0.400 0.35 0.55 0.034 45 0.396 0.35 0.43 0.020 75 0.397 0.25 0.48 0.025 19 0.385 0.35 0.40 0.019
WI3 18 0.530 0.50 0.55 0.019 30 0.529 0.50 0.60 0.023 45 0.536 0.38 0.63 0.032 75 0.539 0.45 0.58 0.019 19 0.519 0.49 0.55 0.016
LCinf 18 0.726 0.70 0.76 0.020 31 0.754 0.70 0.84 0.031 45 0.736 0.63 0.85 0.047 75 0.723 0.64 0.79 0.029 19 0.717 0.68 0.78 0.021
WCinf 18 0.829 0.78 0.88 0.032 31 0.821 0.79 0.86 0.022 45 0.799 0.74 0.86 0.024 75 0.804 0.68 0.85 0.027 19 0.789 0.78 0.81 0.013
LP2 18 0.594 0.55 0.64 0.023 28 0.587 0.55 0.63 0.021 46 0.567 0.53 0.60 0.022 75 0.566 0.53 0.63 0.022 19 0.551 0.53 0.58 0.020
WP2 18 0.651 0.63 0.68 0.017 28 0.626 0.59 0.65 0.017 46 0.645 0.60 0.71 0.028 75 0.643 0.53 0.69 0.027 19 0.626 0.59 0.68 0.020
LP3 18 0.643 0.60 0.70 0.025 32 0.627 0.58 0.68 0.028 47 0.625 0.55 0.68 0.025 77 0.614 0.48 0.71 0.033 19 0.595 0.53 0.68 0.032
WP3 18 0.673 0.60 0.73 0.034 31 0.628 0.55 0.68 0.029 47 0.661 0.59 0.75 0.035 77 0.650 0.45 0.73 0.041 19 0.641 0.60 0.68 0.020
WP4 18 0.791 0.73 0.85 0.029 34 0.762 0.70 0.85 0.030 48 0.781 0.68 0.85 0.035 77 0.767 0.63 0.88 0.040 19 0.759 0.63 0.79 0.036
LP4 18 0.607 0.55 0.68 0.037 34 0.613 0.51 0.75 0.055 48 0.639 0.54 0.71 0.041 76 0.603 0.50 0.78 0.050 19 0.605 0.54 0.68 0.039
LM1 18 1.433 1.40 1.48 0.026 36 1.475 1.40 1.56 0.032 48 1.445 1.33 1.53 0.044 78 1.467 1.35 1.53 0.031 19 1.468 1.44 1.50 0.020
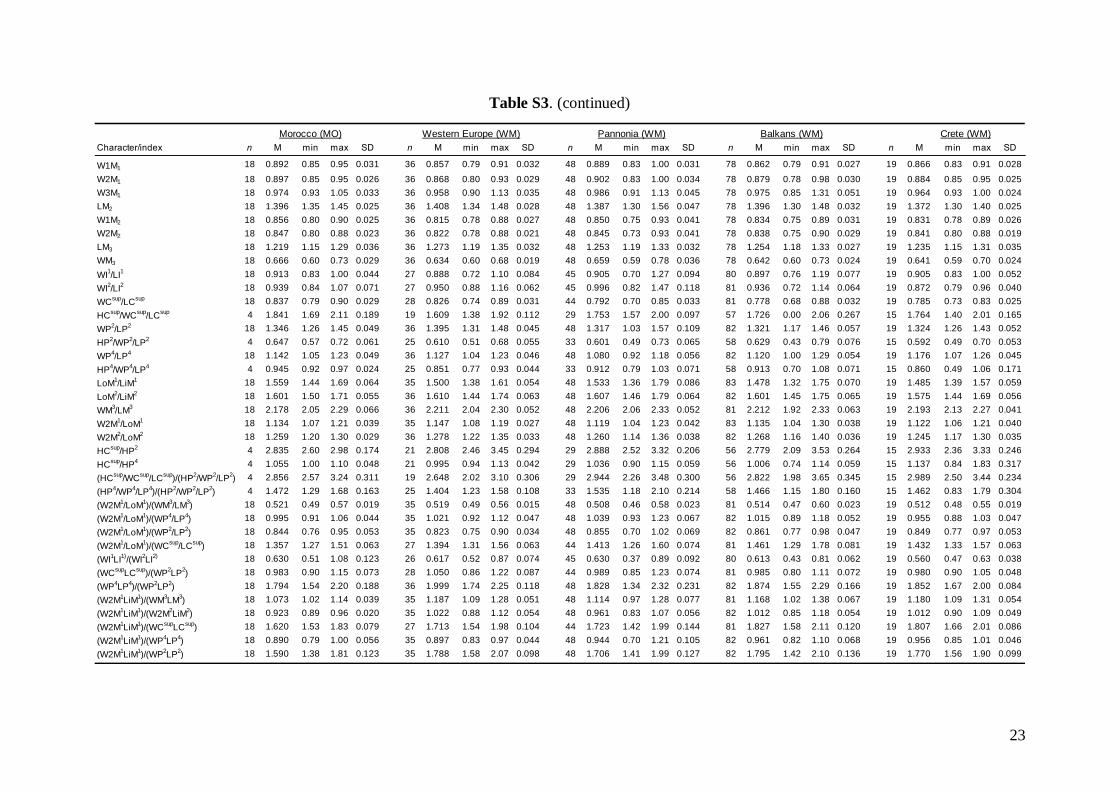
23
Table S3. (continued)
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
W1M118 0.892 0.85 0.95 0.031 36 0.857 0.79 0.91 0.032 48 0.889 0.83 1.00 0.031 78 0.862 0.79 0.91 0.027 19 0.866 0.83 0.91 0.028
W2M1 18 0.897 0.85 0.95 0.026 36 0.868 0.80 0.93 0.029 48 0.902 0.83 1.00 0.034 78 0.879 0.78 0.98 0.030 19 0.884 0.85 0.95 0.025
W3M1 18 0.974 0.93 1.05 0.033 36 0.958 0.90 1.13 0.035 48 0.986 0.91 1.13 0.045 78 0.975 0.85 1.31 0.051 19 0.964 0.93 1.00 0.024
LM2 18 1.396 1.35 1.45 0.025 36 1.408 1.34 1.48 0.028 48 1.387 1.30 1.56 0.047 78 1.396 1.30 1.48 0.032 19 1.372 1.30 1.40 0.025
W1M2 18 0.856 0.80 0.90 0.025 36 0.815 0.78 0.88 0.027 48 0.850 0.75 0.93 0.041 78 0.834 0.75 0.89 0.031 19 0.831 0.78 0.89 0.026
W2M2 18 0.847 0.80 0.88 0.023 36 0.822 0.78 0.88 0.021 48 0.845 0.73 0.93 0.041 78 0.838 0.75 0.90 0.029 19 0.841 0.80 0.88 0.019
LM3 18 1.219 1.15 1.29 0.036 36 1.273 1.19 1.35 0.032 48 1.253 1.19 1.33 0.032 78 1.254 1.18 1.33 0.027 19 1.235 1.15 1.31 0.035
WM3 18 0.666 0.60 0.73 0.029 36 0.634 0.60 0.68 0.019 48 0.659 0.59 0.78 0.036 78 0.642 0.60 0.73 0.024 19 0.641 0.59 0.70 0.024
WI1/LI
1 18 0.913 0.83 1.00 0.044 27 0.888 0.72 1.10 0.084 45 0.905 0.70 1.27 0.094 80 0.897 0.76 1.19 0.077 19 0.905 0.83 1.00 0.052
WI2/LI
2 18 0.939 0.84 1.07 0.071 27 0.950 0.88 1.16 0.062 45 0.996 0.82 1.47 0.118 81 0.936 0.72 1.14 0.064 19 0.872 0.79 0.96 0.040
WCsup
/LCsup 18 0.837 0.79 0.90 0.029 28 0.826 0.74 0.89 0.031 44 0.792 0.70 0.85 0.033 81 0.778 0.68 0.88 0.032 19 0.785 0.73 0.83 0.025
HCsup
/WCsup
/LCsup 4 1.841 1.69 2.11 0.189 19 1.609 1.38 1.92 0.112 29 1.753 1.57 2.00 0.097 57 1.726 0.00 2.06 0.267 15 1.764 1.40 2.01 0.165
WP2/LP
2 18 1.346 1.26 1.45 0.049 36 1.395 1.31 1.48 0.045 48 1.317 1.03 1.57 0.109 82 1.321 1.17 1.46 0.057 19 1.324 1.26 1.43 0.052
HP2/WP
2/LP
2 4 0.647 0.57 0.72 0.061 25 0.610 0.51 0.68 0.055 33 0.601 0.49 0.73 0.065 58 0.629 0.43 0.79 0.076 15 0.592 0.49 0.70 0.053
WP4/LP
4 18 1.142 1.05 1.23 0.049 36 1.127 1.04 1.23 0.046 48 1.080 0.92 1.18 0.056 82 1.120 1.00 1.29 0.054 19 1.176 1.07 1.26 0.045
HP4/WP
4/LP
4 4 0.945 0.92 0.97 0.024 25 0.851 0.77 0.93 0.044 33 0.912 0.79 1.03 0.071 58 0.913 0.70 1.08 0.071 15 0.860 0.49 1.06 0.171
LoM1/LiM
1 18 1.559 1.44 1.69 0.064 35 1.500 1.38 1.61 0.054 48 1.533 1.36 1.79 0.086 83 1.478 1.32 1.75 0.070 19 1.485 1.39 1.57 0.059
LoM2/LiM
2 18 1.601 1.50 1.71 0.055 36 1.610 1.44 1.74 0.063 48 1.607 1.46 1.79 0.064 82 1.601 1.45 1.75 0.065 19 1.575 1.44 1.69 0.056
WM3/LM
3 18 2.178 2.05 2.29 0.066 36 2.211 2.04 2.30 0.052 48 2.206 2.06 2.33 0.052 81 2.212 1.92 2.33 0.063 19 2.193 2.13 2.27 0.041
W2M1/LoM
1 18 1.134 1.07 1.21 0.039 35 1.147 1.08 1.19 0.027 48 1.119 1.04 1.23 0.042 83 1.135 1.04 1.30 0.038 19 1.122 1.06 1.21 0.040
W2M2/LoM
2 18 1.259 1.20 1.30 0.029 36 1.278 1.22 1.35 0.033 48 1.260 1.14 1.36 0.038 82 1.268 1.16 1.40 0.036 19 1.245 1.17 1.30 0.035
HCsup
/HP2 4 2.835 2.60 2.98 0.174 21 2.808 2.46 3.45 0.294 29 2.888 2.52 3.32 0.206 56 2.779 2.09 3.53 0.264 15 2.933 2.36 3.33 0.246
HCsup
/HP4 4 1.055 1.00 1.10 0.048 21 0.995 0.94 1.13 0.042 29 1.036 0.90 1.15 0.059 56 1.006 0.74 1.14 0.059 15 1.137 0.84 1.83 0.317
(HCsup
/WCsup
/LCsup
)/(HP2/WP
2/LP
2) 4 2.856 2.57 3.24 0.311 19 2.648 2.02 3.10 0.306 29 2.944 2.26 3.48 0.300 56 2.822 1.98 3.65 0.345 15 2.989 2.50 3.44 0.234
(HP4/WP
4/LP
4)/(HP
2/WP
2/LP
2) 4 1.472 1.29 1.68 0.163 25 1.404 1.23 1.58 0.108 33 1.535 1.18 2.10 0.214 58 1.466 1.15 1.80 0.160 15 1.462 0.83 1.79 0.304
(W2M1/LoM
1)/(WM
3/LM
3) 18 0.521 0.49 0.57 0.019 35 0.519 0.49 0.56 0.015 48 0.508 0.46 0.58 0.023 81 0.514 0.47 0.60 0.023 19 0.512 0.48 0.55 0.019
(W2M1/LoM
1)/(WP
4/LP
4) 18 0.995 0.91 1.06 0.044 35 1.021 0.92 1.12 0.047 48 1.039 0.93 1.23 0.067 82 1.015 0.89 1.18 0.052 19 0.955 0.88 1.03 0.047
(W2M1/LoM
1)/(WP
2/LP
2) 18 0.844 0.76 0.95 0.053 35 0.823 0.75 0.90 0.034 48 0.855 0.70 1.02 0.069 82 0.861 0.77 0.98 0.047 19 0.849 0.77 0.97 0.053
(W2M1/LoM
1)/(WC
sup/LC
sup) 18 1.357 1.27 1.51 0.063 27 1.394 1.31 1.56 0.063 44 1.413 1.26 1.60 0.074 81 1.461 1.29 1.78 0.081 19 1.432 1.33 1.57 0.063
(WI1LI
1)/(WI
2LI
2) 18 0.630 0.51 1.08 0.123 26 0.617 0.52 0.87 0.074 45 0.630 0.37 0.89 0.092 80 0.613 0.43 0.81 0.062 19 0.560 0.47 0.63 0.038
(WCsup
LCsup
)/(WP2LP
2) 18 0.983 0.90 1.15 0.073 28 1.050 0.86 1.22 0.087 44 0.989 0.85 1.23 0.074 81 0.985 0.80 1.11 0.072 19 0.980 0.90 1.05 0.048
(WP4LP
4)/(WP
2LP
2) 18 1.794 1.54 2.20 0.188 36 1.999 1.74 2.25 0.118 48 1.828 1.34 2.32 0.231 82 1.874 1.55 2.29 0.166 19 1.852 1.67 2.00 0.084
(W2M1LiM
1)/(WM
3LM
3) 18 1.073 1.02 1.14 0.039 35 1.187 1.09 1.28 0.051 48 1.114 0.97 1.28 0.077 81 1.168 1.02 1.38 0.067 19 1.180 1.09 1.31 0.054
(W2M1LiM
1)/(W2M
2LiM
2) 18 0.923 0.89 0.96 0.020 35 1.022 0.88 1.12 0.054 48 0.961 0.83 1.07 0.056 82 1.012 0.85 1.18 0.054 19 1.012 0.90 1.09 0.049
(W2M1LiM
1)/(WC
supLC
sup) 18 1.620 1.53 1.83 0.079 27 1.713 1.54 1.98 0.104 44 1.723 1.42 1.99 0.144 81 1.827 1.58 2.11 0.120 19 1.807 1.66 2.01 0.086
(W2M1LiM
1)/(WP
4LP
4) 18 0.890 0.79 1.00 0.056 35 0.897 0.83 0.97 0.044 48 0.944 0.70 1.21 0.105 82 0.961 0.82 1.10 0.068 19 0.956 0.85 1.01 0.046
(W2M1LiM
1)/(WP
2LP
2) 18 1.590 1.38 1.81 0.123 35 1.788 1.58 2.07 0.098 48 1.706 1.41 1.99 0.127 82 1.795 1.42 2.10 0.136 19 1.770 1.56 1.90 0.099
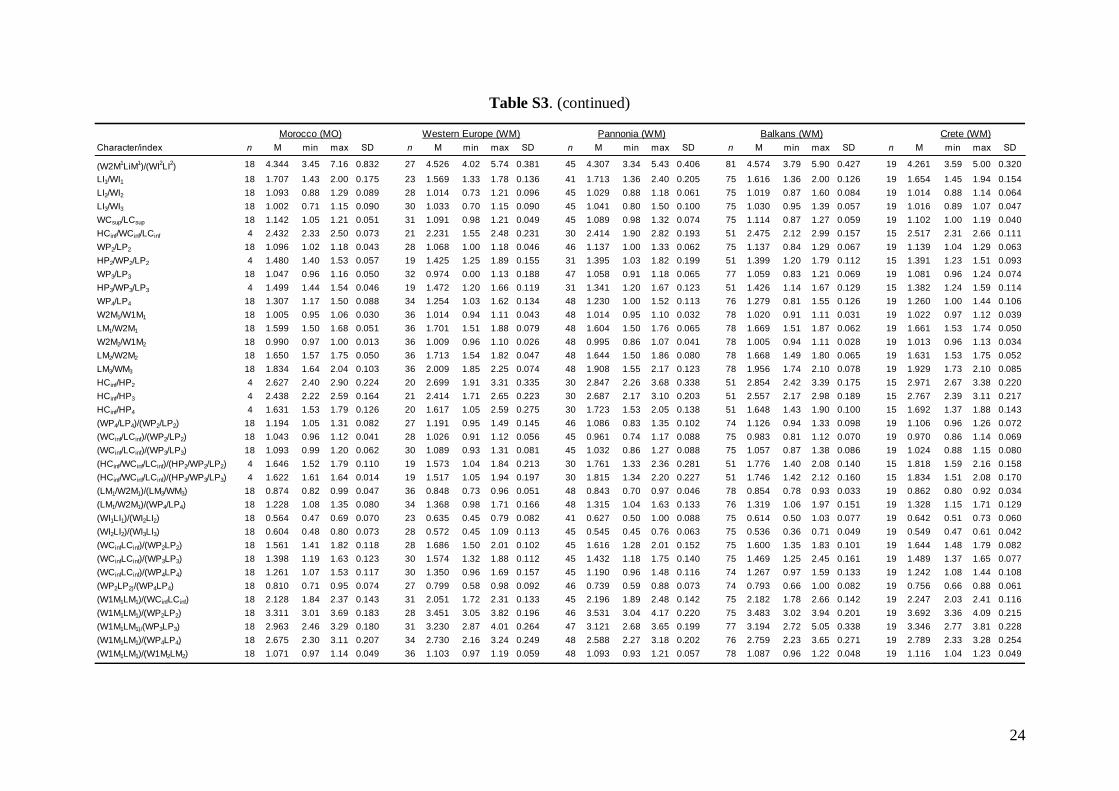
24
Table S3. (continued)
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
(W2M1LiM
1)/(WI
2LI
2) 18 4.344 3.45 7.16 0.832 27 4.526 4.02 5.74 0.381 45 4.307 3.34 5.43 0.406 81 4.574 3.79 5.90 0.427 19 4.261 3.59 5.00 0.320
LI1/WI1 18 1.707 1.43 2.00 0.175 23 1.569 1.33 1.78 0.136 41 1.713 1.36 2.40 0.205 75 1.616 1.36 2.00 0.126 19 1.654 1.45 1.94 0.154
LI2/WI2 18 1.093 0.88 1.29 0.089 28 1.014 0.73 1.21 0.096 45 1.029 0.88 1.18 0.061 75 1.019 0.87 1.60 0.084 19 1.014 0.88 1.14 0.064
LI3/WI3 18 1.002 0.71 1.15 0.090 30 1.033 0.70 1.15 0.090 45 1.041 0.80 1.50 0.100 75 1.030 0.95 1.39 0.057 19 1.016 0.89 1.07 0.047
WCsup/LCsup 18 1.142 1.05 1.21 0.051 31 1.091 0.98 1.21 0.049 45 1.089 0.98 1.32 0.074 75 1.114 0.87 1.27 0.059 19 1.102 1.00 1.19 0.040
HCinf/WCinf/LCinf 4 2.432 2.33 2.50 0.073 21 2.231 1.55 2.48 0.231 30 2.414 1.90 2.82 0.193 51 2.475 2.12 2.99 0.157 15 2.517 2.31 2.66 0.111
WP2/LP2 18 1.096 1.02 1.18 0.043 28 1.068 1.00 1.18 0.046 46 1.137 1.00 1.33 0.062 75 1.137 0.84 1.29 0.067 19 1.139 1.04 1.29 0.063
HP2/WP2/LP2 4 1.480 1.40 1.53 0.057 19 1.425 1.25 1.89 0.155 31 1.395 1.03 1.82 0.199 51 1.399 1.20 1.79 0.112 15 1.391 1.23 1.51 0.093
WP3/LP3 18 1.047 0.96 1.16 0.050 32 0.974 0.00 1.13 0.188 47 1.058 0.91 1.18 0.065 77 1.059 0.83 1.21 0.069 19 1.081 0.96 1.24 0.074
HP3/WP3/LP3 4 1.499 1.44 1.54 0.046 19 1.472 1.20 1.66 0.119 31 1.341 1.20 1.67 0.123 51 1.426 1.14 1.67 0.129 15 1.382 1.24 1.59 0.114
WP4/LP4 18 1.307 1.17 1.50 0.088 34 1.254 1.03 1.62 0.134 48 1.230 1.00 1.52 0.113 76 1.279 0.81 1.55 0.126 19 1.260 1.00 1.44 0.106
W2M1/W1M1 18 1.005 0.95 1.06 0.030 36 1.014 0.94 1.11 0.043 48 1.014 0.95 1.10 0.032 78 1.020 0.91 1.11 0.031 19 1.022 0.97 1.12 0.039
LM1/W2M1 18 1.599 1.50 1.68 0.051 36 1.701 1.51 1.88 0.079 48 1.604 1.50 1.76 0.065 78 1.669 1.51 1.87 0.062 19 1.661 1.53 1.74 0.050
W2M2/W1M2 18 0.990 0.97 1.00 0.013 36 1.009 0.96 1.10 0.026 48 0.995 0.86 1.07 0.041 78 1.005 0.94 1.11 0.028 19 1.013 0.96 1.13 0.034
LM2/W2M2 18 1.650 1.57 1.75 0.050 36 1.713 1.54 1.82 0.047 48 1.644 1.50 1.86 0.080 78 1.668 1.49 1.80 0.065 19 1.631 1.53 1.75 0.052
LM3/WM3 18 1.834 1.64 2.04 0.103 36 2.009 1.85 2.25 0.074 48 1.908 1.55 2.17 0.123 78 1.956 1.74 2.10 0.078 19 1.929 1.73 2.10 0.085
HCinf/HP2 4 2.627 2.40 2.90 0.224 20 2.699 1.91 3.31 0.335 30 2.847 2.26 3.68 0.338 51 2.854 2.42 3.39 0.175 15 2.971 2.67 3.38 0.220
HCinf/HP3 4 2.438 2.22 2.59 0.164 21 2.414 1.71 2.65 0.223 30 2.687 2.17 3.10 0.203 51 2.557 2.17 2.98 0.189 15 2.767 2.39 3.11 0.217
HCinf/HP4 4 1.631 1.53 1.79 0.126 20 1.617 1.05 2.59 0.275 30 1.723 1.53 2.05 0.138 51 1.648 1.43 1.90 0.100 15 1.692 1.37 1.88 0.143
(WP4/LP4)/(WP2/LP2) 18 1.194 1.05 1.31 0.082 27 1.191 0.95 1.49 0.145 46 1.086 0.83 1.35 0.102 74 1.126 0.94 1.33 0.098 19 1.106 0.96 1.26 0.072
(WCinf/LCinf)/(WP2/LP2) 18 1.043 0.96 1.12 0.041 28 1.026 0.91 1.12 0.056 45 0.961 0.74 1.17 0.088 75 0.983 0.81 1.12 0.070 19 0.970 0.86 1.14 0.069
(WCinf/LCinf)/(WP3/LP3) 18 1.093 0.99 1.20 0.062 30 1.089 0.93 1.31 0.081 45 1.032 0.86 1.27 0.088 75 1.057 0.87 1.38 0.086 19 1.024 0.88 1.15 0.080
(HCinf/WCinf/LCinf)/(HP2/WP2/LP2) 4 1.646 1.52 1.79 0.110 19 1.573 1.04 1.84 0.213 30 1.761 1.33 2.36 0.281 51 1.776 1.40 2.08 0.140 15 1.818 1.59 2.16 0.158
(HCinf/WCinf/LCinf)/(HP3/WP3/LP3) 4 1.622 1.61 1.64 0.014 19 1.517 1.05 1.94 0.197 30 1.815 1.34 2.20 0.227 51 1.746 1.42 2.12 0.160 15 1.834 1.51 2.08 0.170
(LM1/W2M1)/(LM3/WM3) 18 0.874 0.82 0.99 0.047 36 0.848 0.73 0.96 0.051 48 0.843 0.70 0.97 0.046 78 0.854 0.78 0.93 0.033 19 0.862 0.80 0.92 0.034
(LM1/W2M1)/(WP4/LP4) 18 1.228 1.08 1.35 0.080 34 1.368 0.98 1.71 0.166 48 1.315 1.04 1.63 0.133 76 1.319 1.06 1.97 0.151 19 1.328 1.15 1.71 0.129
(WI1LI1)/(WI2LI2) 18 0.564 0.47 0.69 0.070 23 0.635 0.45 0.79 0.082 41 0.627 0.50 1.00 0.088 75 0.614 0.50 1.03 0.077 19 0.642 0.51 0.73 0.060
(WI2LI2)/(WI3LI3) 18 0.604 0.48 0.80 0.073 28 0.572 0.45 1.09 0.113 45 0.545 0.45 0.76 0.063 75 0.536 0.36 0.71 0.049 19 0.549 0.47 0.61 0.042
(WCinfLCinf)/(WP2LP2) 18 1.561 1.41 1.82 0.118 28 1.686 1.50 2.01 0.102 45 1.616 1.28 2.01 0.152 75 1.600 1.35 1.83 0.101 19 1.644 1.48 1.79 0.082
(WCinfLCinf)/(WP3LP3) 18 1.398 1.19 1.63 0.123 30 1.574 1.32 1.88 0.112 45 1.432 1.18 1.75 0.140 75 1.469 1.25 2.45 0.161 19 1.489 1.37 1.65 0.077
(WCinfLCinf)/(WP4LP4) 18 1.261 1.07 1.53 0.117 30 1.350 0.96 1.69 0.157 45 1.190 0.96 1.48 0.116 74 1.267 0.97 1.59 0.133 19 1.242 1.08 1.44 0.108
(WP2LP2)/(WP4LP4) 18 0.810 0.71 0.95 0.074 27 0.799 0.58 0.98 0.092 46 0.739 0.59 0.88 0.073 74 0.793 0.66 1.00 0.082 19 0.756 0.66 0.88 0.061
(W1M1LM1)/(WCinfLCinf) 18 2.128 1.84 2.37 0.143 31 2.051 1.72 2.31 0.133 45 2.196 1.89 2.48 0.142 75 2.182 1.78 2.66 0.142 19 2.247 2.03 2.41 0.116
(W1M1LM1)/(WP2LP2) 18 3.311 3.01 3.69 0.183 28 3.451 3.05 3.82 0.196 46 3.531 3.04 4.17 0.220 75 3.483 3.02 3.94 0.201 19 3.692 3.36 4.09 0.215
(W1M1LM1)/(WP3LP3) 18 2.963 2.46 3.29 0.180 31 3.230 2.87 4.01 0.264 47 3.121 2.68 3.65 0.199 77 3.194 2.72 5.05 0.338 19 3.346 2.77 3.81 0.228
(W1M1LM1)/(WP4LP4) 18 2.675 2.30 3.11 0.207 34 2.730 2.16 3.24 0.249 48 2.588 2.27 3.18 0.202 76 2.759 2.23 3.65 0.271 19 2.789 2.33 3.28 0.254
(W1M1LM1)/(W1M2LM2) 18 1.071 0.97 1.14 0.049 36 1.103 0.97 1.19 0.059 48 1.093 0.93 1.21 0.057 78 1.087 0.96 1.22 0.048 19 1.116 1.04 1.23 0.049
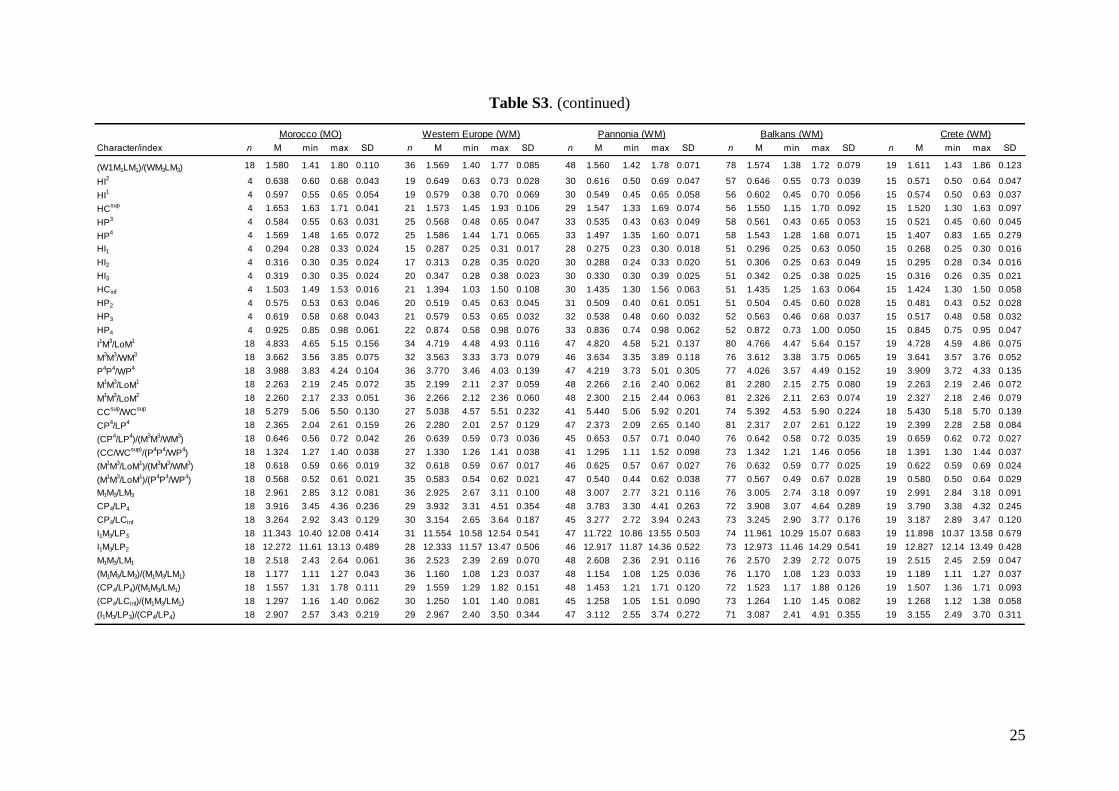
25
Table S3. (continued)
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
(W1M1LM1)/(WM3LM3) 18 1.580 1.41 1.80 0.110 36 1.569 1.40 1.77 0.085 48 1.560 1.42 1.78 0.071 78 1.574 1.38 1.72 0.079 19 1.611 1.43 1.86 0.123
HI2 4 0.638 0.60 0.68 0.043 19 0.649 0.63 0.73 0.028 30 0.616 0.50 0.69 0.047 57 0.646 0.55 0.73 0.039 15 0.571 0.50 0.64 0.047
HI1 4 0.597 0.55 0.65 0.054 19 0.579 0.38 0.70 0.069 30 0.549 0.45 0.65 0.058 56 0.602 0.45 0.70 0.056 15 0.574 0.50 0.63 0.037
HCsup 4 1.653 1.63 1.71 0.041 21 1.573 1.45 1.93 0.106 29 1.547 1.33 1.69 0.074 56 1.550 1.15 1.70 0.092 15 1.520 1.30 1.63 0.097
HP3 4 0.584 0.55 0.63 0.031 25 0.568 0.48 0.65 0.047 33 0.535 0.43 0.63 0.049 58 0.561 0.43 0.65 0.053 15 0.521 0.45 0.60 0.045
HP4 4 1.569 1.48 1.65 0.072 25 1.586 1.44 1.71 0.065 33 1.497 1.35 1.60 0.071 58 1.543 1.28 1.68 0.071 15 1.407 0.83 1.65 0.279
HI1 4 0.294 0.28 0.33 0.024 15 0.287 0.25 0.31 0.017 28 0.275 0.23 0.30 0.018 51 0.296 0.25 0.63 0.050 15 0.268 0.25 0.30 0.016
HI2 4 0.316 0.30 0.35 0.024 17 0.313 0.28 0.35 0.020 30 0.288 0.24 0.33 0.020 51 0.306 0.25 0.63 0.049 15 0.295 0.28 0.34 0.016
HI3 4 0.319 0.30 0.35 0.024 20 0.347 0.28 0.38 0.023 30 0.330 0.30 0.39 0.025 51 0.342 0.25 0.38 0.025 15 0.316 0.26 0.35 0.021
HCinf 4 1.503 1.49 1.53 0.016 21 1.394 1.03 1.50 0.108 30 1.435 1.30 1.56 0.063 51 1.435 1.25 1.63 0.064 15 1.424 1.30 1.50 0.058
HP2 4 0.575 0.53 0.63 0.046 20 0.519 0.45 0.63 0.045 31 0.509 0.40 0.61 0.051 51 0.504 0.45 0.60 0.028 15 0.481 0.43 0.52 0.028
HP3 4 0.619 0.58 0.68 0.043 21 0.579 0.53 0.65 0.032 32 0.538 0.48 0.60 0.032 52 0.563 0.46 0.68 0.037 15 0.517 0.48 0.58 0.032
HP4 4 0.925 0.85 0.98 0.061 22 0.874 0.58 0.98 0.076 33 0.836 0.74 0.98 0.062 52 0.872 0.73 1.00 0.050 15 0.845 0.75 0.95 0.047
I1M
3/LoM
1 18 4.833 4.65 5.15 0.156 34 4.719 4.48 4.93 0.116 47 4.820 4.58 5.21 0.137 80 4.766 4.47 5.64 0.157 19 4.728 4.59 4.86 0.075
M3M
3/WM
3 18 3.662 3.56 3.85 0.075 32 3.563 3.33 3.73 0.079 46 3.634 3.35 3.89 0.118 76 3.612 3.38 3.75 0.065 19 3.641 3.57 3.76 0.052
P4P
4/WP
4 18 3.988 3.83 4.24 0.104 36 3.770 3.46 4.03 0.139 47 4.219 3.73 5.01 0.305 77 4.026 3.57 4.49 0.152 19 3.909 3.72 4.33 0.135
M1M
3/LoM
1 18 2.263 2.19 2.45 0.072 35 2.199 2.11 2.37 0.059 48 2.266 2.16 2.40 0.062 81 2.280 2.15 2.75 0.080 19 2.263 2.19 2.46 0.072
M1M
3/LoM
2 18 2.260 2.17 2.33 0.051 36 2.266 2.12 2.36 0.060 48 2.300 2.15 2.44 0.063 81 2.326 2.11 2.63 0.074 19 2.327 2.18 2.46 0.079
CCsup
/WCsup 18 5.279 5.06 5.50 0.130 27 5.038 4.57 5.51 0.232 41 5.440 5.06 5.92 0.201 74 5.392 4.53 5.90 0.224 18 5.430 5.18 5.70 0.139
CP4/LP
4 18 2.365 2.04 2.61 0.159 26 2.280 2.01 2.57 0.129 47 2.373 2.09 2.65 0.140 81 2.317 2.07 2.61 0.122 19 2.399 2.28 2.58 0.084
(CP4/LP
4)/(M
3M
3/WM
3) 18 0.646 0.56 0.72 0.042 26 0.639 0.59 0.73 0.036 45 0.653 0.57 0.71 0.040 76 0.642 0.58 0.72 0.035 19 0.659 0.62 0.72 0.027
(CC/WCsup)
/(P4P
4/WP
4) 18 1.324 1.27 1.40 0.038 27 1.330 1.26 1.41 0.038 41 1.295 1.11 1.52 0.098 73 1.342 1.21 1.46 0.056 18 1.391 1.30 1.44 0.037
(M1M
3/LoM
1)/(M
3M
3/WM
3) 18 0.618 0.59 0.66 0.019 32 0.618 0.59 0.67 0.017 46 0.625 0.57 0.67 0.027 76 0.632 0.59 0.77 0.025 19 0.622 0.59 0.69 0.024
(M1M
3/LoM
1)/(P
4P
4/WP
4) 18 0.568 0.52 0.61 0.021 35 0.583 0.54 0.62 0.021 47 0.540 0.44 0.62 0.038 77 0.567 0.49 0.67 0.028 19 0.580 0.50 0.64 0.029
M1M3/LM3 18 2.961 2.85 3.12 0.081 36 2.925 2.67 3.11 0.100 48 3.007 2.77 3.21 0.116 76 3.005 2.74 3.18 0.097 19 2.991 2.84 3.18 0.091
CP4/LP4 18 3.916 3.45 4.36 0.236 29 3.932 3.31 4.51 0.354 48 3.783 3.30 4.41 0.263 72 3.908 3.07 4.64 0.289 19 3.790 3.38 4.32 0.245
CP4/LCinf 18 3.264 2.92 3.43 0.129 30 3.154 2.65 3.64 0.187 45 3.277 2.72 3.94 0.243 73 3.245 2.90 3.77 0.176 19 3.187 2.89 3.47 0.120
I1M3/LP3 18 11.343 10.40 12.08 0.414 31 11.554 10.58 12.54 0.541 47 11.722 10.86 13.55 0.503 74 11.961 10.29 15.07 0.683 19 11.898 10.37 13.58 0.679
I1M3/LP2 18 12.272 11.61 13.13 0.489 28 12.333 11.57 13.47 0.506 46 12.917 11.87 14.36 0.522 73 12.973 11.46 14.29 0.541 19 12.827 12.14 13.49 0.428
M1M3/LM1 18 2.518 2.43 2.64 0.061 36 2.523 2.39 2.69 0.070 48 2.608 2.36 2.91 0.116 76 2.570 2.39 2.72 0.075 19 2.515 2.45 2.59 0.047
(M1M3/LM3)/(M1M3/LM1) 18 1.177 1.11 1.27 0.043 36 1.160 1.08 1.23 0.037 48 1.154 1.08 1.25 0.036 76 1.170 1.08 1.23 0.033 19 1.189 1.11 1.27 0.037
(CP4/LP4)/(M1M3/LM1) 18 1.557 1.31 1.78 0.111 29 1.559 1.29 1.82 0.151 48 1.453 1.21 1.71 0.120 72 1.523 1.17 1.88 0.126 19 1.507 1.36 1.71 0.093
(CP4/LCinf)/(M1M3/LM1) 18 1.297 1.16 1.40 0.062 30 1.250 1.01 1.40 0.081 45 1.258 1.05 1.51 0.090 73 1.264 1.10 1.45 0.082 19 1.268 1.12 1.38 0.058
(I1M3/LP3)/(CP4/LP4) 18 2.907 2.57 3.43 0.219 29 2.967 2.40 3.50 0.344 47 3.112 2.55 3.74 0.272 71 3.087 2.41 4.91 0.355 19 3.155 2.49 3.70 0.311
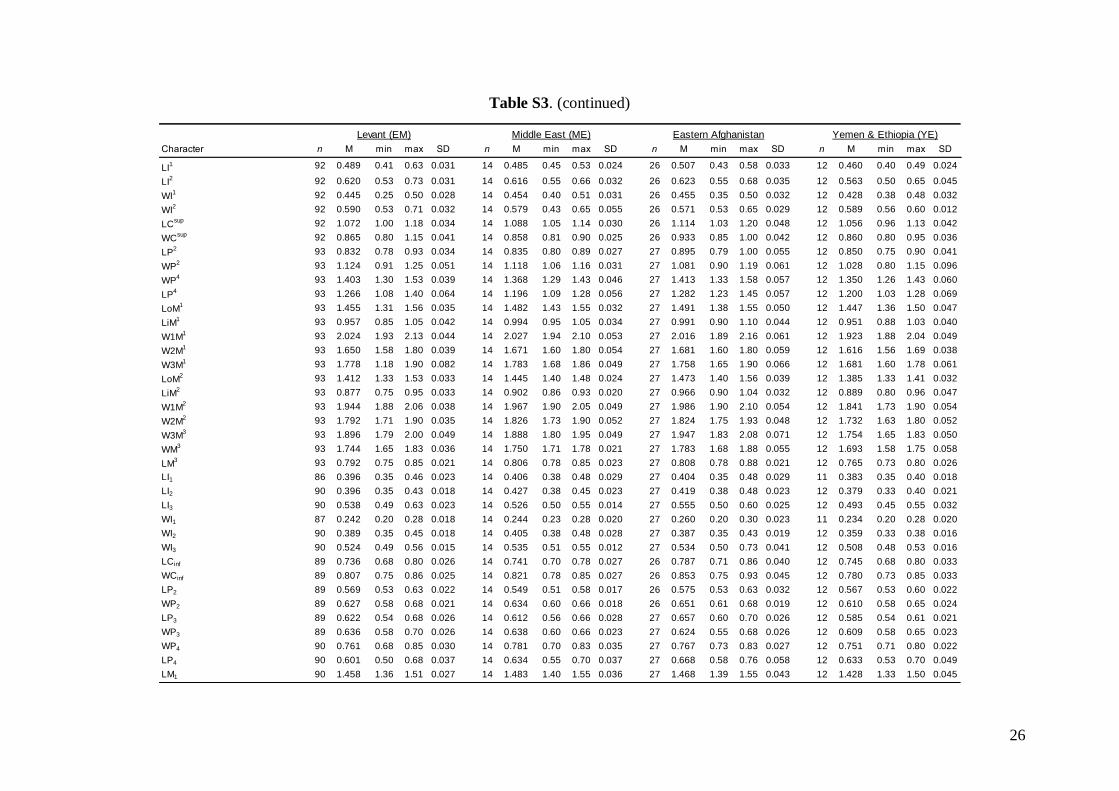
26
Table S3. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character n M min max SD n M min max SD n M min max SD n M min max SD
LI1 92 0.489 0.41 0.63 0.031 14 0.485 0.45 0.53 0.024 26 0.507 0.43 0.58 0.033 12 0.460 0.40 0.49 0.024
LI2 92 0.620 0.53 0.73 0.031 14 0.616 0.55 0.66 0.032 26 0.623 0.55 0.68 0.035 12 0.563 0.50 0.65 0.045
WI1 92 0.445 0.25 0.50 0.028 14 0.454 0.40 0.51 0.031 26 0.455 0.35 0.50 0.032 12 0.428 0.38 0.48 0.032
WI2 92 0.590 0.53 0.71 0.032 14 0.579 0.43 0.65 0.055 26 0.571 0.53 0.65 0.029 12 0.589 0.56 0.60 0.012
LCsup 92 1.072 1.00 1.18 0.034 14 1.088 1.05 1.14 0.030 26 1.114 1.03 1.20 0.048 12 1.056 0.96 1.13 0.042
WCsup 92 0.865 0.80 1.15 0.041 14 0.858 0.81 0.90 0.025 26 0.933 0.85 1.00 0.042 12 0.860 0.80 0.95 0.036
LP2 93 0.832 0.78 0.93 0.034 14 0.835 0.80 0.89 0.027 27 0.895 0.79 1.00 0.055 12 0.850 0.75 0.90 0.041
WP2 93 1.124 0.91 1.25 0.051 14 1.118 1.06 1.16 0.031 27 1.081 0.90 1.19 0.061 12 1.028 0.80 1.15 0.096
WP4 93 1.403 1.30 1.53 0.039 14 1.368 1.29 1.43 0.046 27 1.413 1.33 1.58 0.057 12 1.350 1.26 1.43 0.060
LP4 93 1.266 1.08 1.40 0.064 14 1.196 1.09 1.28 0.056 27 1.282 1.23 1.45 0.057 12 1.200 1.03 1.28 0.069
LoM1 93 1.455 1.31 1.56 0.035 14 1.482 1.43 1.55 0.032 27 1.491 1.38 1.55 0.050 12 1.447 1.36 1.50 0.047
LiM1 93 0.957 0.85 1.05 0.042 14 0.994 0.95 1.05 0.034 27 0.991 0.90 1.10 0.044 12 0.951 0.88 1.03 0.040
W1M1 93 2.024 1.93 2.13 0.044 14 2.027 1.94 2.10 0.053 27 2.016 1.89 2.16 0.061 12 1.923 1.88 2.04 0.049
W2M1 93 1.650 1.58 1.80 0.039 14 1.671 1.60 1.80 0.054 27 1.681 1.60 1.80 0.059 12 1.616 1.56 1.69 0.038
W3M1 93 1.778 1.18 1.90 0.082 14 1.783 1.68 1.86 0.049 27 1.758 1.65 1.90 0.066 12 1.681 1.60 1.78 0.061
LoM2 93 1.412 1.33 1.53 0.033 14 1.445 1.40 1.48 0.024 27 1.473 1.40 1.56 0.039 12 1.385 1.33 1.41 0.032
LiM2 93 0.877 0.75 0.95 0.033 14 0.902 0.86 0.93 0.020 27 0.966 0.90 1.04 0.032 12 0.889 0.80 0.96 0.047
W1M2 93 1.944 1.88 2.06 0.038 14 1.967 1.90 2.05 0.049 27 1.986 1.90 2.10 0.054 12 1.841 1.73 1.90 0.054
W2M2 93 1.792 1.71 1.90 0.035 14 1.826 1.73 1.90 0.052 27 1.824 1.75 1.93 0.048 12 1.732 1.63 1.80 0.052
W3M3 93 1.896 1.79 2.00 0.049 14 1.888 1.80 1.95 0.049 27 1.947 1.83 2.08 0.071 12 1.754 1.65 1.83 0.050
WM3 93 1.744 1.65 1.83 0.036 14 1.750 1.71 1.78 0.021 27 1.783 1.68 1.88 0.055 12 1.693 1.58 1.75 0.058
LM3 93 0.792 0.75 0.85 0.021 14 0.806 0.78 0.85 0.023 27 0.808 0.78 0.88 0.021 12 0.765 0.73 0.80 0.026
LI1 86 0.396 0.35 0.46 0.023 14 0.406 0.38 0.48 0.029 27 0.404 0.35 0.48 0.029 11 0.383 0.35 0.40 0.018
LI2 90 0.396 0.35 0.43 0.018 14 0.427 0.38 0.45 0.023 27 0.419 0.38 0.48 0.023 12 0.379 0.33 0.40 0.021
LI3 90 0.538 0.49 0.63 0.023 14 0.526 0.50 0.55 0.014 27 0.555 0.50 0.60 0.025 12 0.493 0.45 0.55 0.032
WI1 87 0.242 0.20 0.28 0.018 14 0.244 0.23 0.28 0.020 27 0.260 0.20 0.30 0.023 11 0.234 0.20 0.28 0.020
WI2 90 0.389 0.35 0.45 0.018 14 0.405 0.38 0.48 0.028 27 0.387 0.35 0.43 0.019 12 0.359 0.33 0.38 0.016
WI3 90 0.524 0.49 0.56 0.015 14 0.535 0.51 0.55 0.012 27 0.534 0.50 0.73 0.041 12 0.508 0.48 0.53 0.016
LCinf 89 0.736 0.68 0.80 0.026 14 0.741 0.70 0.78 0.027 26 0.787 0.71 0.86 0.040 12 0.745 0.68 0.80 0.033
WCinf 89 0.807 0.75 0.86 0.025 14 0.821 0.78 0.85 0.027 26 0.853 0.75 0.93 0.045 12 0.780 0.73 0.85 0.033
LP2 89 0.569 0.53 0.63 0.022 14 0.549 0.51 0.58 0.017 26 0.575 0.53 0.63 0.032 12 0.567 0.53 0.60 0.022
WP2 89 0.627 0.58 0.68 0.021 14 0.634 0.60 0.66 0.018 26 0.651 0.61 0.68 0.019 12 0.610 0.58 0.65 0.024
LP3 89 0.622 0.54 0.68 0.026 14 0.612 0.56 0.66 0.028 27 0.657 0.60 0.70 0.026 12 0.585 0.54 0.61 0.021
WP3 89 0.636 0.58 0.70 0.026 14 0.638 0.60 0.66 0.023 27 0.624 0.55 0.68 0.026 12 0.609 0.58 0.65 0.023
WP4 90 0.761 0.68 0.85 0.030 14 0.781 0.70 0.83 0.035 27 0.767 0.73 0.83 0.027 12 0.751 0.71 0.80 0.022
LP4 90 0.601 0.50 0.68 0.037 14 0.634 0.55 0.70 0.037 27 0.668 0.58 0.76 0.058 12 0.633 0.53 0.70 0.049
LM1 90 1.458 1.36 1.51 0.027 14 1.483 1.40 1.55 0.036 27 1.468 1.39 1.55 0.043 12 1.428 1.33 1.50 0.045
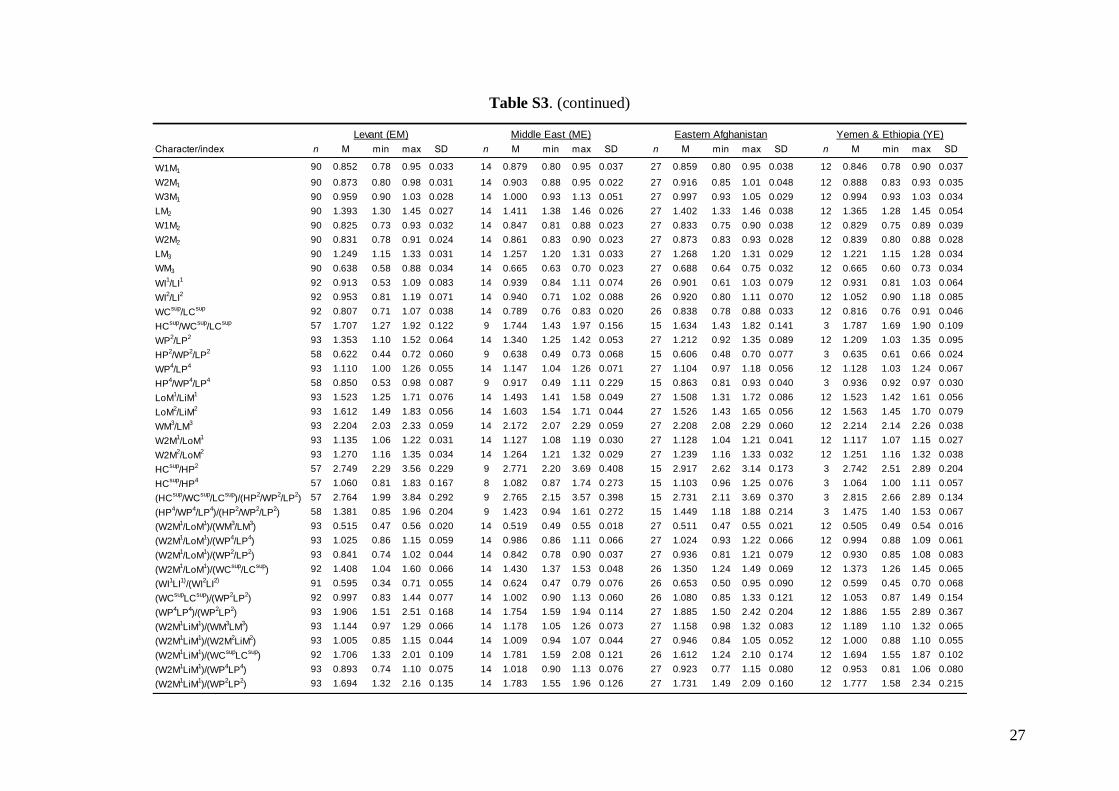
27
Table S3. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
W1M190 0.852 0.78 0.95 0.033 14 0.879 0.80 0.95 0.037 27 0.859 0.80 0.95 0.038 12 0.846 0.78 0.90 0.037
W2M1 90 0.873 0.80 0.98 0.031 14 0.903 0.88 0.95 0.022 27 0.916 0.85 1.01 0.048 12 0.888 0.83 0.93 0.035
W3M1 90 0.959 0.90 1.03 0.028 14 1.000 0.93 1.13 0.051 27 0.997 0.93 1.05 0.029 12 0.994 0.93 1.03 0.034
LM2 90 1.393 1.30 1.45 0.027 14 1.411 1.38 1.46 0.026 27 1.402 1.33 1.46 0.038 12 1.365 1.28 1.45 0.054
W1M2 90 0.825 0.73 0.93 0.032 14 0.847 0.81 0.88 0.023 27 0.833 0.75 0.90 0.038 12 0.829 0.75 0.89 0.039
W2M2 90 0.831 0.78 0.91 0.024 14 0.861 0.83 0.90 0.023 27 0.873 0.83 0.93 0.028 12 0.839 0.80 0.88 0.028
LM3 90 1.249 1.15 1.33 0.031 14 1.257 1.20 1.31 0.033 27 1.268 1.20 1.31 0.029 12 1.221 1.15 1.28 0.034
WM3 90 0.638 0.58 0.88 0.034 14 0.665 0.63 0.70 0.023 27 0.688 0.64 0.75 0.032 12 0.665 0.60 0.73 0.034
WI1/LI
1 92 0.913 0.53 1.09 0.083 14 0.939 0.84 1.11 0.074 26 0.901 0.61 1.03 0.079 12 0.931 0.81 1.03 0.064
WI2/LI
2 92 0.953 0.81 1.19 0.071 14 0.940 0.71 1.02 0.088 26 0.920 0.80 1.11 0.070 12 1.052 0.90 1.18 0.085
WCsup
/LCsup 92 0.807 0.71 1.07 0.038 14 0.789 0.76 0.83 0.020 26 0.838 0.78 0.88 0.033 12 0.816 0.76 0.91 0.046
HCsup
/WCsup
/LCsup 57 1.707 1.27 1.92 0.122 9 1.744 1.43 1.97 0.156 15 1.634 1.43 1.82 0.141 3 1.787 1.69 1.90 0.109
WP2/LP
2 93 1.353 1.10 1.52 0.064 14 1.340 1.25 1.42 0.053 27 1.212 0.92 1.35 0.089 12 1.209 1.03 1.35 0.095
HP2/WP
2/LP
2 58 0.622 0.44 0.72 0.060 9 0.638 0.49 0.73 0.068 15 0.606 0.48 0.70 0.077 3 0.635 0.61 0.66 0.024
WP4/LP
4 93 1.110 1.00 1.26 0.055 14 1.147 1.04 1.26 0.071 27 1.104 0.97 1.18 0.056 12 1.128 1.03 1.24 0.067
HP4/WP
4/LP
4 58 0.850 0.53 0.98 0.087 9 0.917 0.49 1.11 0.229 15 0.863 0.81 0.93 0.040 3 0.936 0.92 0.97 0.030
LoM1/LiM
1 93 1.523 1.25 1.71 0.076 14 1.493 1.41 1.58 0.049 27 1.508 1.31 1.72 0.086 12 1.523 1.42 1.61 0.056
LoM2/LiM
2 93 1.612 1.49 1.83 0.056 14 1.603 1.54 1.71 0.044 27 1.526 1.43 1.65 0.056 12 1.563 1.45 1.70 0.079
WM3/LM
3 93 2.204 2.03 2.33 0.059 14 2.172 2.07 2.29 0.059 27 2.208 2.08 2.29 0.060 12 2.214 2.14 2.26 0.038
W2M1/LoM
1 93 1.135 1.06 1.22 0.031 14 1.127 1.08 1.19 0.030 27 1.128 1.04 1.21 0.041 12 1.117 1.07 1.15 0.027
W2M2/LoM
2 93 1.270 1.16 1.35 0.034 14 1.264 1.21 1.32 0.029 27 1.239 1.16 1.33 0.032 12 1.251 1.16 1.32 0.038
HCsup
/HP2 57 2.749 2.29 3.56 0.229 9 2.771 2.20 3.69 0.408 15 2.917 2.62 3.14 0.173 3 2.742 2.51 2.89 0.204
HCsup
/HP4 57 1.060 0.81 1.83 0.167 8 1.082 0.87 1.74 0.273 15 1.103 0.96 1.25 0.076 3 1.064 1.00 1.11 0.057
(HCsup
/WCsup
/LCsup
)/(HP2/WP
2/LP
2) 57 2.764 1.99 3.84 0.292 9 2.765 2.15 3.57 0.398 15 2.731 2.11 3.69 0.370 3 2.815 2.66 2.89 0.134
(HP4/WP
4/LP
4)/(HP
2/WP
2/LP
2) 58 1.381 0.85 1.96 0.204 9 1.423 0.94 1.61 0.272 15 1.449 1.18 1.88 0.214 3 1.475 1.40 1.53 0.067
(W2M1/LoM
1)/(WM
3/LM
3) 93 0.515 0.47 0.56 0.020 14 0.519 0.49 0.55 0.018 27 0.511 0.47 0.55 0.021 12 0.505 0.49 0.54 0.016
(W2M1/LoM
1)/(WP
4/LP
4) 93 1.025 0.86 1.15 0.059 14 0.986 0.86 1.11 0.066 27 1.024 0.93 1.22 0.066 12 0.994 0.88 1.09 0.061
(W2M1/LoM
1)/(WP
2/LP
2) 93 0.841 0.74 1.02 0.044 14 0.842 0.78 0.90 0.037 27 0.936 0.81 1.21 0.079 12 0.930 0.85 1.08 0.083
(W2M1/LoM
1)/(WC
sup/LC
sup) 92 1.408 1.04 1.60 0.066 14 1.430 1.37 1.53 0.048 26 1.350 1.24 1.49 0.069 12 1.373 1.26 1.45 0.065
(WI1LI
1)/(WI
2LI
2) 91 0.595 0.34 0.71 0.055 14 0.624 0.47 0.79 0.076 26 0.653 0.50 0.95 0.090 12 0.599 0.45 0.70 0.068
(WCsup
LCsup
)/(WP2LP
2) 92 0.997 0.83 1.44 0.077 14 1.002 0.90 1.13 0.060 26 1.080 0.85 1.33 0.121 12 1.053 0.87 1.49 0.154
(WP4LP
4)/(WP
2LP
2) 93 1.906 1.51 2.51 0.168 14 1.754 1.59 1.94 0.114 27 1.885 1.50 2.42 0.204 12 1.886 1.55 2.89 0.367
(W2M1LiM
1)/(WM
3LM
3) 93 1.144 0.97 1.29 0.066 14 1.178 1.05 1.26 0.073 27 1.158 0.98 1.32 0.083 12 1.189 1.10 1.32 0.065
(W2M1LiM
1)/(W2M
2LiM
2) 93 1.005 0.85 1.15 0.044 14 1.009 0.94 1.07 0.044 27 0.946 0.84 1.05 0.052 12 1.000 0.88 1.10 0.055
(W2M1LiM
1)/(WC
supLC
sup) 92 1.706 1.33 2.01 0.109 14 1.781 1.59 2.08 0.121 26 1.612 1.24 2.10 0.174 12 1.694 1.55 1.87 0.102
(W2M1LiM
1)/(WP
4LP
4) 93 0.893 0.74 1.10 0.075 14 1.018 0.90 1.13 0.076 27 0.923 0.77 1.15 0.080 12 0.953 0.81 1.06 0.080
(W2M1LiM
1)/(WP
2LP
2) 93 1.694 1.32 2.16 0.135 14 1.783 1.55 1.96 0.126 27 1.731 1.49 2.09 0.160 12 1.777 1.58 2.34 0.215
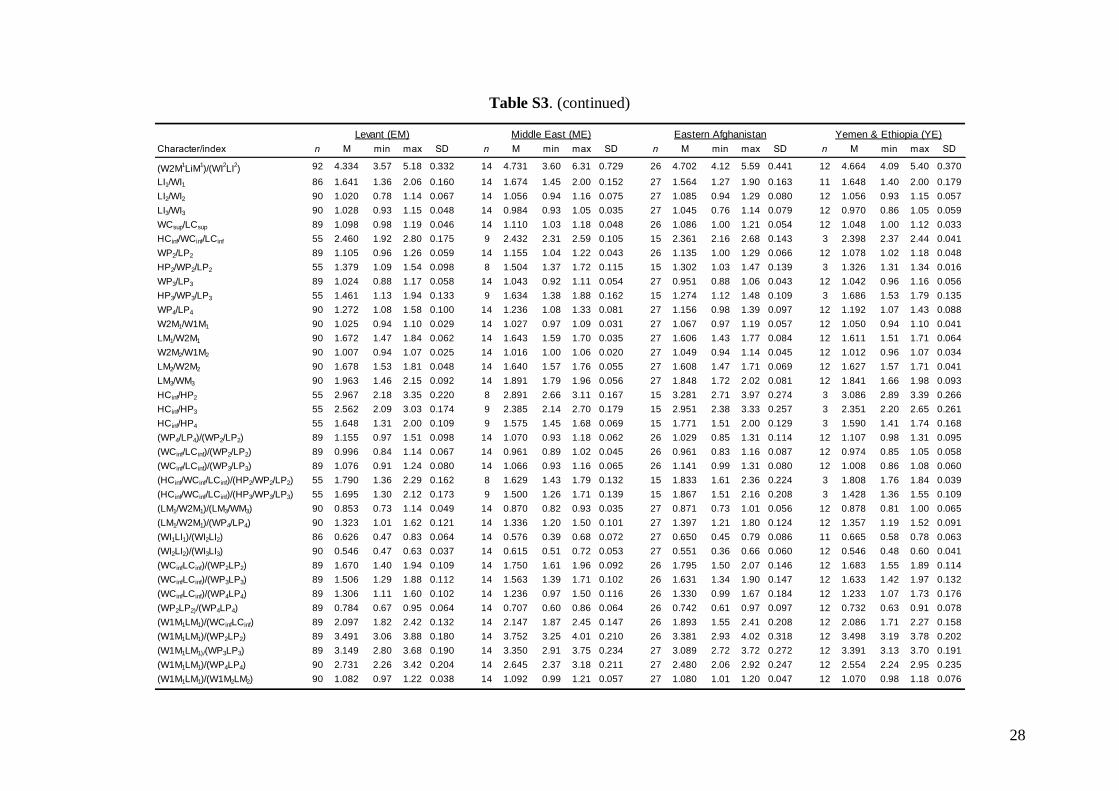
28
Table S3. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
(W2M1LiM
1)/(WI
2LI
2) 92 4.334 3.57 5.18 0.332 14 4.731 3.60 6.31 0.729 26 4.702 4.12 5.59 0.441 12 4.664 4.09 5.40 0.370
LI1/WI1 86 1.641 1.36 2.06 0.160 14 1.674 1.45 2.00 0.152 27 1.564 1.27 1.90 0.163 11 1.648 1.40 2.00 0.179
LI2/WI2 90 1.020 0.78 1.14 0.067 14 1.056 0.94 1.16 0.075 27 1.085 0.94 1.29 0.080 12 1.056 0.93 1.15 0.057
LI3/WI3 90 1.028 0.93 1.15 0.048 14 0.984 0.93 1.05 0.035 27 1.045 0.76 1.14 0.079 12 0.970 0.86 1.05 0.059
WCsup/LCsup 89 1.098 0.98 1.19 0.046 14 1.110 1.03 1.18 0.048 26 1.086 1.00 1.21 0.054 12 1.048 1.00 1.12 0.033
HCinf/WCinf/LCinf 55 2.460 1.92 2.80 0.175 9 2.432 2.31 2.59 0.105 15 2.361 2.16 2.68 0.143 3 2.398 2.37 2.44 0.041
WP2/LP2 89 1.105 0.96 1.26 0.059 14 1.155 1.04 1.22 0.043 26 1.135 1.00 1.29 0.066 12 1.078 1.02 1.18 0.048
HP2/WP2/LP2 55 1.379 1.09 1.54 0.098 8 1.504 1.37 1.72 0.115 15 1.302 1.03 1.47 0.139 3 1.326 1.31 1.34 0.016
WP3/LP3 89 1.024 0.88 1.17 0.058 14 1.043 0.92 1.11 0.054 27 0.951 0.88 1.06 0.043 12 1.042 0.96 1.16 0.056
HP3/WP3/LP3 55 1.461 1.13 1.94 0.133 9 1.634 1.38 1.88 0.162 15 1.274 1.12 1.48 0.109 3 1.686 1.53 1.79 0.135
WP4/LP4 90 1.272 1.08 1.58 0.100 14 1.236 1.08 1.33 0.081 27 1.156 0.98 1.39 0.097 12 1.192 1.07 1.43 0.088
W2M1/W1M1 90 1.025 0.94 1.10 0.029 14 1.027 0.97 1.09 0.031 27 1.067 0.97 1.19 0.057 12 1.050 0.94 1.10 0.041
LM1/W2M1 90 1.672 1.47 1.84 0.062 14 1.643 1.59 1.70 0.035 27 1.606 1.43 1.77 0.084 12 1.611 1.51 1.71 0.064
W2M2/W1M2 90 1.007 0.94 1.07 0.025 14 1.016 1.00 1.06 0.020 27 1.049 0.94 1.14 0.045 12 1.012 0.96 1.07 0.034
LM2/W2M2 90 1.678 1.53 1.81 0.048 14 1.640 1.57 1.76 0.055 27 1.608 1.47 1.71 0.069 12 1.627 1.57 1.71 0.041
LM3/WM3 90 1.963 1.46 2.15 0.092 14 1.891 1.79 1.96 0.056 27 1.848 1.72 2.02 0.081 12 1.841 1.66 1.98 0.093
HCinf/HP2 55 2.967 2.18 3.35 0.220 8 2.891 2.66 3.11 0.167 15 3.281 2.71 3.97 0.274 3 3.086 2.89 3.39 0.266
HCinf/HP3 55 2.562 2.09 3.03 0.174 9 2.385 2.14 2.70 0.179 15 2.951 2.38 3.33 0.257 3 2.351 2.20 2.65 0.261
HCinf/HP4 55 1.648 1.31 2.00 0.109 9 1.575 1.45 1.68 0.069 15 1.771 1.51 2.00 0.129 3 1.590 1.41 1.74 0.168
(WP4/LP4)/(WP2/LP2) 89 1.155 0.97 1.51 0.098 14 1.070 0.93 1.18 0.062 26 1.029 0.85 1.31 0.114 12 1.107 0.98 1.31 0.095
(WCinf/LCinf)/(WP2/LP2) 89 0.996 0.84 1.14 0.067 14 0.961 0.89 1.02 0.045 26 0.961 0.83 1.16 0.087 12 0.974 0.85 1.05 0.058
(WCinf/LCinf)/(WP3/LP3) 89 1.076 0.91 1.24 0.080 14 1.066 0.93 1.16 0.065 26 1.141 0.99 1.31 0.080 12 1.008 0.86 1.08 0.060
(HCinf/WCinf/LCinf)/(HP2/WP2/LP2) 55 1.790 1.36 2.29 0.162 8 1.629 1.43 1.79 0.132 15 1.833 1.61 2.36 0.224 3 1.808 1.76 1.84 0.039
(HCinf/WCinf/LCinf)/(HP3/WP3/LP3) 55 1.695 1.30 2.12 0.173 9 1.500 1.26 1.71 0.139 15 1.867 1.51 2.16 0.208 3 1.428 1.36 1.55 0.109
(LM1/W2M1)/(LM3/WM3) 90 0.853 0.73 1.14 0.049 14 0.870 0.82 0.93 0.035 27 0.871 0.73 1.01 0.056 12 0.878 0.81 1.00 0.065
(LM1/W2M1)/(WP4/LP4) 90 1.323 1.01 1.62 0.121 14 1.336 1.20 1.50 0.101 27 1.397 1.21 1.80 0.124 12 1.357 1.19 1.52 0.091
(WI1LI1)/(WI2LI2) 86 0.626 0.47 0.83 0.064 14 0.576 0.39 0.68 0.072 27 0.650 0.45 0.79 0.086 11 0.665 0.58 0.78 0.063
(WI2LI2)/(WI3LI3) 90 0.546 0.47 0.63 0.037 14 0.615 0.51 0.72 0.053 27 0.551 0.36 0.66 0.060 12 0.546 0.48 0.60 0.041
(WCinfLCinf)/(WP2LP2) 89 1.670 1.40 1.94 0.109 14 1.750 1.61 1.96 0.092 26 1.795 1.50 2.07 0.146 12 1.683 1.55 1.89 0.114
(WCinfLCinf)/(WP3LP3) 89 1.506 1.29 1.88 0.112 14 1.563 1.39 1.71 0.102 26 1.631 1.34 1.90 0.147 12 1.633 1.42 1.97 0.132
(WCinfLCinf)/(WP4LP4) 89 1.306 1.11 1.60 0.102 14 1.236 0.97 1.50 0.116 26 1.330 0.99 1.67 0.184 12 1.233 1.07 1.73 0.176
(WP2LP2)/(WP4LP4) 89 0.784 0.67 0.95 0.064 14 0.707 0.60 0.86 0.064 26 0.742 0.61 0.97 0.097 12 0.732 0.63 0.91 0.078
(W1M1LM1)/(WCinfLCinf) 89 2.097 1.82 2.42 0.132 14 2.147 1.87 2.45 0.147 26 1.893 1.55 2.41 0.208 12 2.086 1.71 2.27 0.158
(W1M1LM1)/(WP2LP2) 89 3.491 3.06 3.88 0.180 14 3.752 3.25 4.01 0.210 26 3.381 2.93 4.02 0.318 12 3.498 3.19 3.78 0.202
(W1M1LM1)/(WP3LP3) 89 3.149 2.80 3.68 0.190 14 3.350 2.91 3.75 0.234 27 3.089 2.72 3.72 0.272 12 3.391 3.13 3.70 0.191
(W1M1LM1)/(WP4LP4) 90 2.731 2.26 3.42 0.204 14 2.645 2.37 3.18 0.211 27 2.480 2.06 2.92 0.247 12 2.554 2.24 2.95 0.235
(W1M1LM1)/(W1M2LM2) 90 1.082 0.97 1.22 0.038 14 1.092 0.99 1.21 0.057 27 1.080 1.01 1.20 0.047 12 1.070 0.98 1.18 0.076
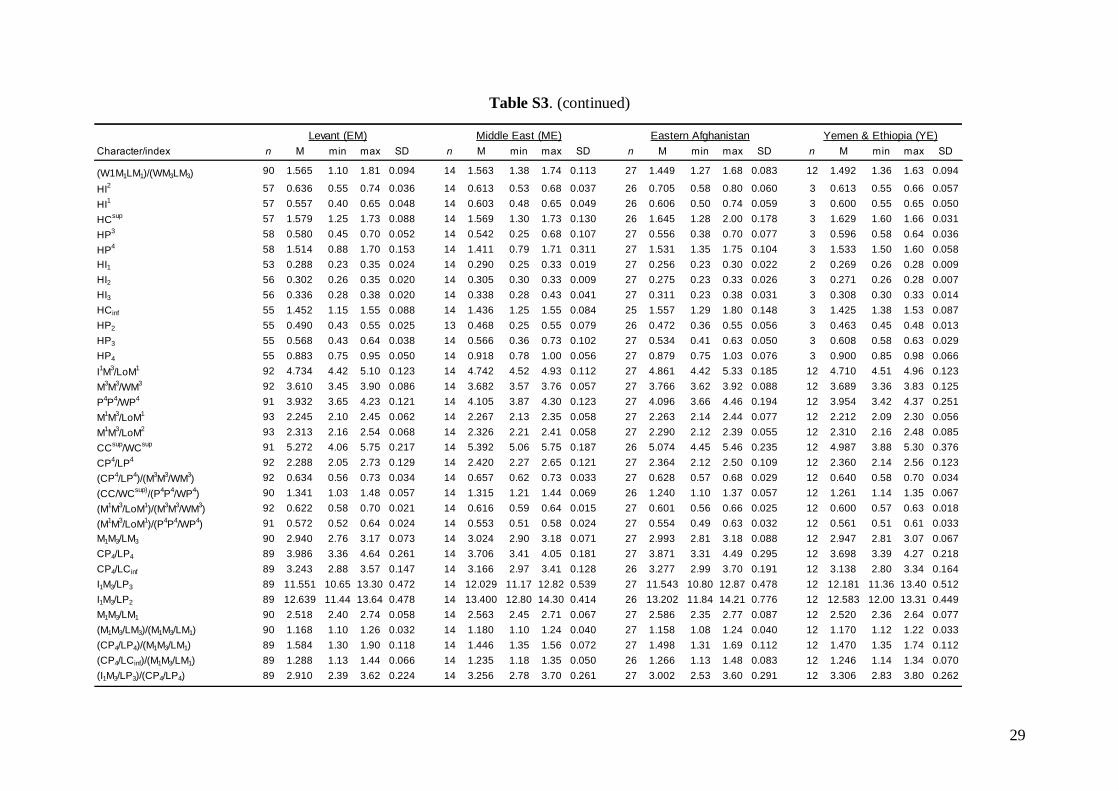
29
Table S3. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character/index n M min max SD n M min max SD n M min max SD n M min max SD
(W1M1LM1)/(WM3LM3) 90 1.565 1.10 1.81 0.094 14 1.563 1.38 1.74 0.113 27 1.449 1.27 1.68 0.083 12 1.492 1.36 1.63 0.094
HI2 57 0.636 0.55 0.74 0.036 14 0.613 0.53 0.68 0.037 26 0.705 0.58 0.80 0.060 3 0.613 0.55 0.66 0.057
HI1 57 0.557 0.40 0.65 0.048 14 0.603 0.48 0.65 0.049 26 0.606 0.50 0.74 0.059 3 0.600 0.55 0.65 0.050
HCsup 57 1.579 1.25 1.73 0.088 14 1.569 1.30 1.73 0.130 26 1.645 1.28 2.00 0.178 3 1.629 1.60 1.66 0.031
HP3 58 0.580 0.45 0.70 0.052 14 0.542 0.25 0.68 0.107 27 0.556 0.38 0.70 0.077 3 0.596 0.58 0.64 0.036
HP4 58 1.514 0.88 1.70 0.153 14 1.411 0.79 1.71 0.311 27 1.531 1.35 1.75 0.104 3 1.533 1.50 1.60 0.058
HI1 53 0.288 0.23 0.35 0.024 14 0.290 0.25 0.33 0.019 27 0.256 0.23 0.30 0.022 2 0.269 0.26 0.28 0.009
HI2 56 0.302 0.26 0.35 0.020 14 0.305 0.30 0.33 0.009 27 0.275 0.23 0.33 0.026 3 0.271 0.26 0.28 0.007
HI3 56 0.336 0.28 0.38 0.020 14 0.338 0.28 0.43 0.041 27 0.311 0.23 0.38 0.031 3 0.308 0.30 0.33 0.014
HCinf 55 1.452 1.15 1.55 0.088 14 1.436 1.25 1.55 0.084 25 1.557 1.29 1.80 0.148 3 1.425 1.38 1.53 0.087
HP2 55 0.490 0.43 0.55 0.025 13 0.468 0.25 0.55 0.079 26 0.472 0.36 0.55 0.056 3 0.463 0.45 0.48 0.013
HP3 55 0.568 0.43 0.64 0.038 14 0.566 0.36 0.73 0.102 27 0.534 0.41 0.63 0.050 3 0.608 0.58 0.63 0.029
HP4 55 0.883 0.75 0.95 0.050 14 0.918 0.78 1.00 0.056 27 0.879 0.75 1.03 0.076 3 0.900 0.85 0.98 0.066
I1M
3/LoM
1 92 4.734 4.42 5.10 0.123 14 4.742 4.52 4.93 0.112 27 4.861 4.42 5.33 0.185 12 4.710 4.51 4.96 0.123
M3M
3/WM
3 92 3.610 3.45 3.90 0.086 14 3.682 3.57 3.76 0.057 27 3.766 3.62 3.92 0.088 12 3.689 3.36 3.83 0.125
P4P
4/WP
4 91 3.932 3.65 4.23 0.121 14 4.105 3.87 4.30 0.123 27 4.096 3.66 4.46 0.194 12 3.954 3.42 4.37 0.251
M1M
3/LoM
1 93 2.245 2.10 2.45 0.062 14 2.267 2.13 2.35 0.058 27 2.263 2.14 2.44 0.077 12 2.212 2.09 2.30 0.056
M1M
3/LoM
2 93 2.313 2.16 2.54 0.068 14 2.326 2.21 2.41 0.058 27 2.290 2.12 2.39 0.055 12 2.310 2.16 2.48 0.085
CCsup
/WCsup 91 5.272 4.06 5.75 0.217 14 5.392 5.06 5.75 0.187 26 5.074 4.45 5.46 0.235 12 4.987 3.88 5.30 0.376
CP4/LP
4 92 2.288 2.05 2.73 0.129 14 2.420 2.27 2.65 0.121 27 2.364 2.12 2.50 0.109 12 2.360 2.14 2.56 0.123
(CP4/LP
4)/(M
3M
3/WM
3) 92 0.634 0.56 0.73 0.034 14 0.657 0.62 0.73 0.033 27 0.628 0.57 0.68 0.029 12 0.640 0.58 0.70 0.034
(CC/WCsup)
/(P4P
4/WP
4) 90 1.341 1.03 1.48 0.057 14 1.315 1.21 1.44 0.069 26 1.240 1.10 1.37 0.057 12 1.261 1.14 1.35 0.067
(M1M
3/LoM
1)/(M
3M
3/WM
3) 92 0.622 0.58 0.70 0.021 14 0.616 0.59 0.64 0.015 27 0.601 0.56 0.66 0.025 12 0.600 0.57 0.63 0.018
(M1M
3/LoM
1)/(P
4P
4/WP
4) 91 0.572 0.52 0.64 0.024 14 0.553 0.51 0.58 0.024 27 0.554 0.49 0.63 0.032 12 0.561 0.51 0.61 0.033
M1M3/LM3 90 2.940 2.76 3.17 0.073 14 3.024 2.90 3.18 0.071 27 2.993 2.81 3.18 0.088 12 2.947 2.81 3.07 0.067
CP4/LP4 89 3.986 3.36 4.64 0.261 14 3.706 3.41 4.05 0.181 27 3.871 3.31 4.49 0.295 12 3.698 3.39 4.27 0.218
CP4/LCinf 89 3.243 2.88 3.57 0.147 14 3.166 2.97 3.41 0.128 26 3.277 2.99 3.70 0.191 12 3.138 2.80 3.34 0.164
I1M3/LP3 89 11.551 10.65 13.30 0.472 14 12.029 11.17 12.82 0.539 27 11.543 10.80 12.87 0.478 12 12.181 11.36 13.40 0.512
I1M3/LP2 89 12.639 11.44 13.64 0.478 14 13.400 12.80 14.30 0.414 26 13.202 11.84 14.21 0.776 12 12.583 12.00 13.31 0.449
M1M3/LM1 90 2.518 2.40 2.74 0.058 14 2.563 2.45 2.71 0.067 27 2.586 2.35 2.77 0.087 12 2.520 2.36 2.64 0.077
(M1M3/LM3)/(M1M3/LM1) 90 1.168 1.10 1.26 0.032 14 1.180 1.10 1.24 0.040 27 1.158 1.08 1.24 0.040 12 1.170 1.12 1.22 0.033
(CP4/LP4)/(M1M3/LM1) 89 1.584 1.30 1.90 0.118 14 1.446 1.35 1.56 0.072 27 1.498 1.31 1.69 0.112 12 1.470 1.35 1.74 0.112
(CP4/LCinf)/(M1M3/LM1) 89 1.288 1.13 1.44 0.066 14 1.235 1.18 1.35 0.050 26 1.266 1.13 1.48 0.083 12 1.246 1.14 1.34 0.070
(I1M3/LP3)/(CP4/LP4) 89 2.910 2.39 3.62 0.224 14 3.256 2.78 3.70 0.261 27 3.002 2.53 3.60 0.291 12 3.306 2.83 3.80 0.262
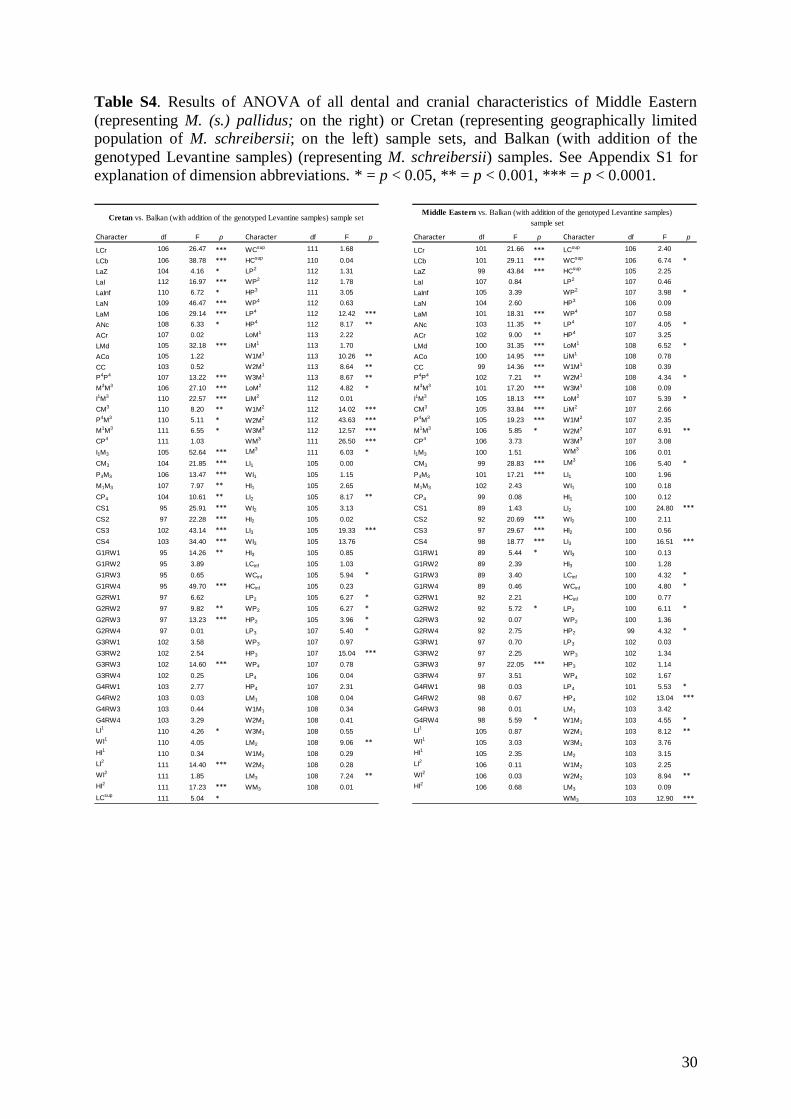
30
Table S4. Results of ANOVA of all dental and cranial characteristics of Middle Eastern
(representing M. (s.) pallidus; on the right) or Cretan (representing geographically limited
population of M. schreibersii; on the left) sample sets, and Balkan (with addition of the
genotyped Levantine samples) (representing M. schreibersii) samples. See Appendix S1 for
explanation of dimension abbreviations. * = p < 0.05, ** = p < 0.001, *** = p < 0.0001.
Character df F p Character df F p Character df F p Character df F p
LCr 106 26.47 *** WCsup 111 1.68 LCr 101 21.66 *** LCsup 106 2.40
LCb 106 38.78 *** HCsup 110 0.04 LCb 101 29.11 *** WCsup 106 6.74 *
LaZ 104 4.16 * LP2 112 1.31 LaZ 99 43.84 *** HCsup 105 2.25
LaI 112 16.97 *** WP2 112 1.78 LaI 107 0.84 LP2 107 0.46
LaInf 110 6.72 * HP3 111 3.05 LaInf 105 3.39 WP2 107 3.98 *
LaN 109 46.47 *** WP4 112 0.63 LaN 104 2.60 HP3 106 0.09
LaM 106 29.14 *** LP4 112 12.42 *** LaM 101 18.31 *** WP4 107 0.58
ANc 108 6.33 * HP4 112 8.17 ** ANc 103 11.35 ** LP4 107 4.05 *
ACr 107 0.02 LoM1 113 2.22 ACr 102 9.00 ** HP4 107 3.25
LMd 105 32.18 *** LiM1 113 1.70 LMd 100 31.35 *** LoM1 108 6.52 *
ACo 105 1.22 W1M1 113 10.26 ** ACo 100 14.95 *** LiM1 108 0.78
CC 103 0.52 W2M1 113 8.64 ** CC 99 14.36 *** W1M1 108 0.39
P4P4 107 13.22 *** W3M1 113 8.67 ** P4P4 102 7.21 ** W2M1 108 4.34 *
M3M3 106 27.10 *** LoM2 112 4.82 * M3M3 101 17.20 *** W3M1 108 0.09
I1M3 110 22.57 *** LiM2 112 0.01 I1M3 105 18.13 *** LoM2 107 5.39 *
CM3 110 8.20 ** W1M2 112 14.02 *** CM3 105 33.84 *** LiM2 107 2.66
P4M3 110 5.11 * W2M2 112 43.63 *** P4M3 105 19.23 *** W1M2 107 2.35
M1M3 111 6.55 * W3M3 112 12.57 *** M1M3 106 5.85 * W2M2 107 6.91 **
CP4 111 1.03 WM3 111 26.50 *** CP4 106 3.73 W3M3 107 3.08
I1M3 105 52.64 *** LM3111 6.03 * I1M3 100 1.51 WM3
106 0.01
CM3 104 21.85 *** LI1 105 0.00 CM3 99 28.83 *** LM3106 5.40 *
P4M3 106 13.47 *** WI1 105 1.15 P4M3 101 17.21 *** LI1 100 1.96
M1M3 107 7.97 ** HI1 105 2.65 M1M3 102 2.43 WI1 100 0.18
CP4 104 10.61 ** LI2 105 8.17 ** CP4 99 0.08 HI1 100 0.12
CS1 95 25.91 *** WI2 105 3.13 CS1 89 1.43 LI2 100 24.80 ***
CS2 97 22.28 *** HI2 105 0.02 CS2 92 20.69 *** WI2 100 2.11
CS3 102 43.14 *** LI3 105 19.33 *** CS3 97 29.67 *** HI2 100 0.56
CS4 103 34.40 *** WI3 105 13.76 CS4 98 18.77 *** LI3 100 16.51 ***
G1RW1 95 14.26 ** HI3 105 0.85 G1RW1 89 5.44 * WI3 100 0.13
G1RW2 95 3.89 LCinf 105 1.03 G1RW2 89 2.39 HI3 100 1.28
G1RW3 95 0.65 WCinf 105 5.94 * G1RW3 89 3.40 LCinf 100 4.32 *
G1RW4 95 49.70 *** HCinf 105 0.23 G1RW4 89 0.46 WCinf 100 4.80 *
G2RW1 97 6.62 LP2 105 6.27 * G2RW1 92 2.21 HCinf 100 0.77
G2RW2 97 9.82 ** WP2 105 6.27 * G2RW2 92 5.72 * LP2 100 6.11 *
G2RW3 97 13.23 *** HP2 105 3.96 * G2RW3 92 0.07 WP2 100 1.36
G2RW4 97 0.01 LP3 107 5.40 * G2RW4 92 2.75 HP2 99 4.32 *
G3RW1 102 3.58 WP3 107 0.97 G3RW1 97 0.70 LP3 102 0.03
G3RW2 102 2.54 HP3 107 15.04 *** G3RW2 97 2.25 WP3 102 1.34
G3RW3 102 14.60 *** WP4 107 0.78 G3RW3 97 22.05 *** HP3 102 1.14
G3RW4 102 0.25 LP4 106 0.04 G3RW4 97 3.51 WP4 102 1.67
G4RW1 103 2.77 HP4 107 2.31 G4RW1 98 0.03 LP4 101 5.53 *
G4RW2 103 0.03 LM1 108 0.04 G4RW2 98 0.67 HP4 102 13.04 ***
G4RW3 103 0.44 W1M1 108 0.34 G4RW3 98 0.01 LM1 103 3.42
G4RW4 103 3.29 W2M1 108 0.41 G4RW4 98 5.59 * W1M1 103 4.55 *
LI1 110 4.26 * W3M1 108 0.55 LI1 105 0.87 W2M1 103 8.12 **
WI1 110 4.05 LM2 108 9.06 ** WI1 105 3.03 W3M1 103 3.76
HI1 110 0.34 W1M2 108 0.29 HI1 105 2.35 LM2 103 3.15
LI2 111 14.40 *** W2M2 108 0.28 LI2 106 0.11 W1M2 103 2.25
WI2 111 1.85 LM3 108 7.24 ** WI2 106 0.03 W2M2 103 8.94 **
HI2 111 17.23 *** WM3 108 0.01 HI2 106 0.68 LM3 103 0.09
LCsup111 5.04 * WM3 103 12.90 ***
Cretan vs. Balkan (with addition of the genotyped Levantine samples) sample setMiddle Eastern vs. Balkan (with addition of the genotyped Levantine samples)
sample set
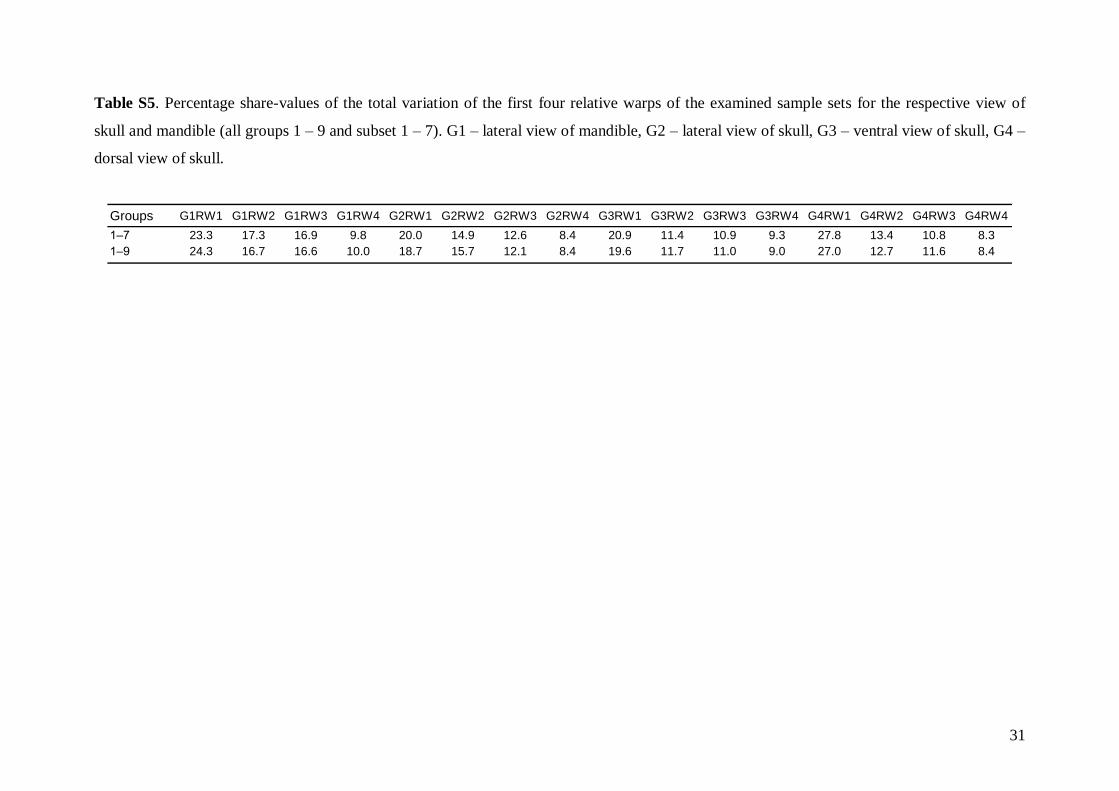
31
Table S5. Percentage share-values of the total variation of the first four relative warps of the examined sample sets for the respective view of
skull and mandible (all groups 1 – 9 and subset 1 – 7). G1 – lateral view of mandible, G2 – lateral view of skull, G3 – ventral view of skull, G4 –
dorsal view of skull.
Groups G1RW1 G1RW2 G1RW3 G1RW4 G2RW1 G2RW2 G2RW3 G2RW4 G3RW1 G3RW2 G3RW3 G3RW4 G4RW1 G4RW2 G4RW3 G4RW4
1–7 23.3 17.3 16.9 9.8 20.0 14.9 12.6 8.4 20.9 11.4 10.9 9.3 27.8 13.4 10.8 8.3
1–9 24.3 16.7 16.6 10.0 18.7 15.7 12.1 8.4 19.6 11.7 11.0 9.0 27.0 12.7 11.6 8.4
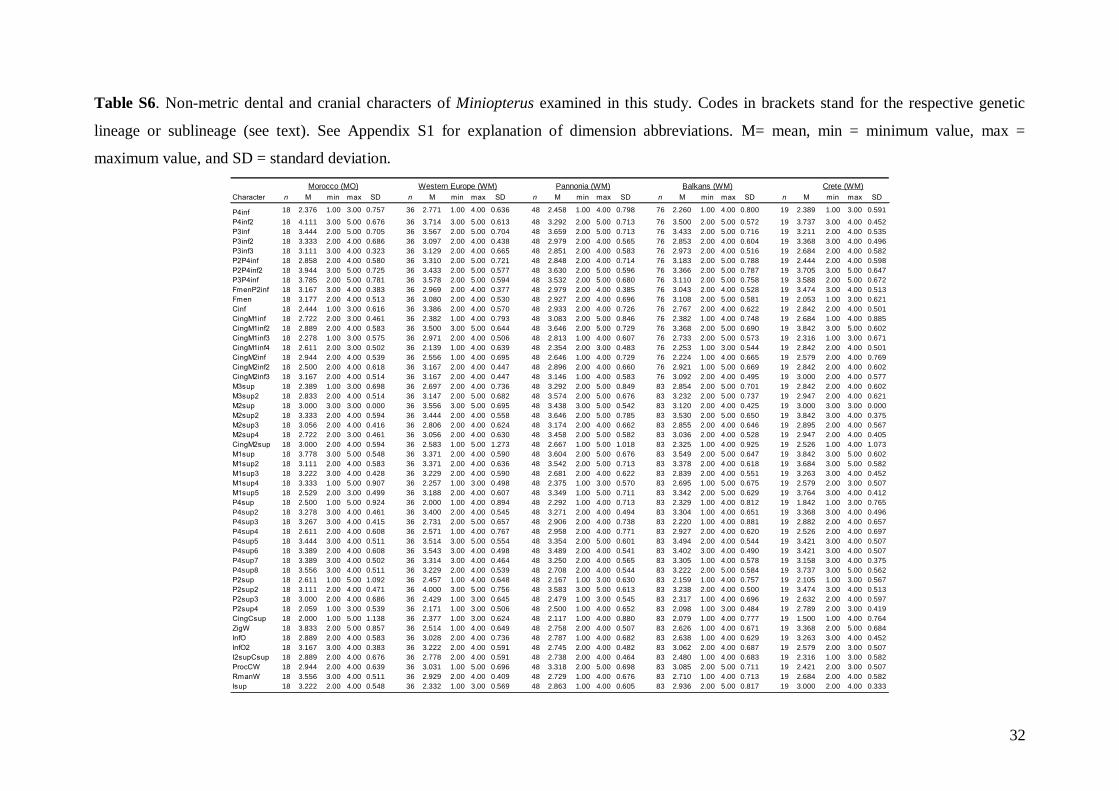
32
Table S6. Non-metric dental and cranial characters of Miniopterus examined in this study. Codes in brackets stand for the respective genetic
lineage or sublineage (see text). See Appendix S1 for explanation of dimension abbreviations. M= mean, min = minimum value, max =
maximum value, and SD = standard deviation.
Morocco (MO) Western Europe (WM) Pannonia (WM) Balkans (WM) Crete (WM)
Character n M min max SD n M min max SD n M min max SD n M min max SD n M min max SD
P4inf 18 2.376 1.00 3.00 0.757 36 2.771 1.00 4.00 0.636 48 2.458 1.00 4.00 0.798 76 2.260 1.00 4.00 0.800 19 2.389 1.00 3.00 0.591
P4inf2 18 4.111 3.00 5.00 0.676 36 3.714 3.00 5.00 0.613 48 3.292 2.00 5.00 0.713 76 3.500 2.00 5.00 0.572 19 3.737 3.00 4.00 0.452
P3inf 18 3.444 2.00 5.00 0.705 36 3.567 2.00 5.00 0.704 48 3.659 2.00 5.00 0.713 76 3.433 2.00 5.00 0.716 19 3.211 2.00 4.00 0.535
P3inf2 18 3.333 2.00 4.00 0.686 36 3.097 2.00 4.00 0.438 48 2.979 2.00 4.00 0.565 76 2.853 2.00 4.00 0.604 19 3.368 3.00 4.00 0.496
P3inf3 18 3.111 3.00 4.00 0.323 36 3.129 2.00 4.00 0.665 48 2.851 2.00 4.00 0.583 76 2.973 2.00 4.00 0.516 19 2.684 2.00 4.00 0.582
P2P4inf 18 2.858 2.00 4.00 0.580 36 3.310 2.00 5.00 0.721 48 2.848 2.00 4.00 0.714 76 3.183 2.00 5.00 0.788 19 2.444 2.00 4.00 0.598
P2P4inf2 18 3.944 3.00 5.00 0.725 36 3.433 2.00 5.00 0.577 48 3.630 2.00 5.00 0.596 76 3.366 2.00 5.00 0.787 19 3.705 3.00 5.00 0.647
P3P4inf 18 3.785 2.00 5.00 0.781 36 3.578 2.00 5.00 0.594 48 3.532 2.00 5.00 0.680 76 3.110 2.00 5.00 0.758 19 3.588 2.00 5.00 0.672
FmenP2inf 18 3.167 3.00 4.00 0.383 36 2.969 2.00 4.00 0.377 48 2.979 2.00 4.00 0.385 76 3.043 2.00 4.00 0.528 19 3.474 3.00 4.00 0.513
Fmen 18 3.177 2.00 4.00 0.513 36 3.080 2.00 4.00 0.530 48 2.927 2.00 4.00 0.696 76 3.108 2.00 5.00 0.581 19 2.053 1.00 3.00 0.621
Cinf 18 2.444 1.00 3.00 0.616 36 3.386 2.00 4.00 0.570 48 2.933 2.00 4.00 0.726 76 2.767 2.00 4.00 0.622 19 2.842 2.00 4.00 0.501
CingM1inf 18 2.722 2.00 3.00 0.461 36 2.382 1.00 4.00 0.793 48 3.083 2.00 5.00 0.846 76 2.382 1.00 4.00 0.748 19 2.684 1.00 4.00 0.885
CingM1inf2 18 2.889 2.00 4.00 0.583 36 3.500 3.00 5.00 0.644 48 3.646 2.00 5.00 0.729 76 3.368 2.00 5.00 0.690 19 3.842 3.00 5.00 0.602
CingM1inf3 18 2.278 1.00 3.00 0.575 36 2.971 2.00 4.00 0.506 48 2.813 1.00 4.00 0.607 76 2.733 2.00 5.00 0.573 19 2.316 1.00 3.00 0.671
CingM1inf4 18 2.611 2.00 3.00 0.502 36 2.139 1.00 4.00 0.639 48 2.354 2.00 3.00 0.483 76 2.253 1.00 3.00 0.544 19 2.842 2.00 4.00 0.501
CingM2inf 18 2.944 2.00 4.00 0.539 36 2.556 1.00 4.00 0.695 48 2.646 1.00 4.00 0.729 76 2.224 1.00 4.00 0.665 19 2.579 2.00 4.00 0.769
CingM2inf2 18 2.500 2.00 4.00 0.618 36 3.167 2.00 4.00 0.447 48 2.896 2.00 4.00 0.660 76 2.921 1.00 5.00 0.669 19 2.842 2.00 4.00 0.602
CingM2inf3 18 3.167 2.00 4.00 0.514 36 3.167 2.00 4.00 0.447 48 3.146 1.00 4.00 0.583 76 3.092 2.00 4.00 0.495 19 3.000 2.00 4.00 0.577
M3sup 18 2.389 1.00 3.00 0.698 36 2.697 2.00 4.00 0.736 48 3.292 2.00 5.00 0.849 83 2.854 2.00 5.00 0.701 19 2.842 2.00 4.00 0.602
M3sup2 18 2.833 2.00 4.00 0.514 36 3.147 2.00 5.00 0.682 48 3.574 2.00 5.00 0.676 83 3.232 2.00 5.00 0.737 19 2.947 2.00 4.00 0.621
M2sup 18 3.000 3.00 3.00 0.000 36 3.556 3.00 5.00 0.695 48 3.438 3.00 5.00 0.542 83 3.120 2.00 4.00 0.425 19 3.000 3.00 3.00 0.000
M2sup2 18 3.333 2.00 4.00 0.594 36 3.444 2.00 4.00 0.558 48 3.646 2.00 5.00 0.785 83 3.530 2.00 5.00 0.650 19 3.842 3.00 4.00 0.375
M2sup3 18 3.056 2.00 4.00 0.416 36 2.806 2.00 4.00 0.624 48 3.174 2.00 4.00 0.662 83 2.855 2.00 4.00 0.646 19 2.895 2.00 4.00 0.567
M2sup4 18 2.722 2.00 3.00 0.461 36 3.056 2.00 4.00 0.630 48 3.458 2.00 5.00 0.582 83 3.036 2.00 4.00 0.528 19 2.947 2.00 4.00 0.405
CingM2sup 18 3.000 2.00 4.00 0.594 36 2.583 1.00 5.00 1.273 48 2.667 1.00 5.00 1.018 83 2.325 1.00 4.00 0.925 19 2.526 1.00 4.00 1.073
M1sup 18 3.778 3.00 5.00 0.548 36 3.371 2.00 4.00 0.590 48 3.604 2.00 5.00 0.676 83 3.549 2.00 5.00 0.647 19 3.842 3.00 5.00 0.602
M1sup2 18 3.111 2.00 4.00 0.583 36 3.371 2.00 4.00 0.636 48 3.542 2.00 5.00 0.713 83 3.378 2.00 4.00 0.618 19 3.684 3.00 5.00 0.582
M1sup3 18 3.222 3.00 4.00 0.428 36 3.229 2.00 4.00 0.590 48 2.681 2.00 4.00 0.622 83 2.839 2.00 4.00 0.551 19 3.263 3.00 4.00 0.452
M1sup4 18 3.333 1.00 5.00 0.907 36 2.257 1.00 3.00 0.498 48 2.375 1.00 3.00 0.570 83 2.695 1.00 5.00 0.675 19 2.579 2.00 3.00 0.507
M1sup5 18 2.529 2.00 3.00 0.499 36 3.188 2.00 4.00 0.607 48 3.349 1.00 5.00 0.711 83 3.342 2.00 5.00 0.629 19 3.764 3.00 4.00 0.412
P4sup 18 2.500 1.00 5.00 0.924 36 2.000 1.00 4.00 0.894 48 2.292 1.00 4.00 0.713 83 2.329 1.00 4.00 0.812 19 1.842 1.00 3.00 0.765
P4sup2 18 3.278 3.00 4.00 0.461 36 3.400 2.00 4.00 0.545 48 3.271 2.00 4.00 0.494 83 3.304 1.00 4.00 0.651 19 3.368 3.00 4.00 0.496
P4sup3 18 3.267 3.00 4.00 0.415 36 2.731 2.00 5.00 0.657 48 2.906 2.00 4.00 0.738 83 2.220 1.00 4.00 0.881 19 2.882 2.00 4.00 0.657
P4sup4 18 2.611 2.00 4.00 0.608 36 2.571 1.00 4.00 0.767 48 2.958 2.00 4.00 0.771 83 2.927 2.00 4.00 0.620 19 2.526 2.00 4.00 0.697
P4sup5 18 3.444 3.00 4.00 0.511 36 3.514 3.00 5.00 0.554 48 3.354 2.00 5.00 0.601 83 3.494 2.00 4.00 0.544 19 3.421 3.00 4.00 0.507
P4sup6 18 3.389 2.00 4.00 0.608 36 3.543 3.00 4.00 0.498 48 3.489 2.00 4.00 0.541 83 3.402 3.00 4.00 0.490 19 3.421 3.00 4.00 0.507
P4sup7 18 3.389 3.00 4.00 0.502 36 3.314 3.00 4.00 0.464 48 3.250 2.00 4.00 0.565 83 3.305 1.00 4.00 0.578 19 3.158 3.00 4.00 0.375
P4sup8 18 3.556 3.00 4.00 0.511 36 3.229 2.00 4.00 0.539 48 2.708 2.00 4.00 0.544 83 3.222 2.00 5.00 0.584 19 3.737 3.00 5.00 0.562
P2sup 18 2.611 1.00 5.00 1.092 36 2.457 1.00 4.00 0.648 48 2.167 1.00 3.00 0.630 83 2.159 1.00 4.00 0.757 19 2.105 1.00 3.00 0.567
P2sup2 18 3.111 2.00 4.00 0.471 36 4.000 3.00 5.00 0.756 48 3.583 3.00 5.00 0.613 83 3.238 2.00 4.00 0.500 19 3.474 3.00 4.00 0.513
P2sup3 18 3.000 2.00 4.00 0.686 36 2.429 1.00 3.00 0.645 48 2.479 1.00 3.00 0.545 83 2.317 1.00 4.00 0.696 19 2.632 2.00 4.00 0.597
P2sup4 18 2.059 1.00 3.00 0.539 36 2.171 1.00 3.00 0.506 48 2.500 1.00 4.00 0.652 83 2.098 1.00 3.00 0.484 19 2.789 2.00 3.00 0.419
CingCsup 18 2.000 1.00 5.00 1.138 36 2.377 1.00 3.00 0.624 48 2.117 1.00 4.00 0.880 83 2.079 1.00 4.00 0.777 19 1.500 1.00 4.00 0.764
ZigW 18 3.833 2.00 5.00 0.857 36 2.514 1.00 4.00 0.649 48 2.758 2.00 4.00 0.507 83 2.626 1.00 4.00 0.671 19 3.368 2.00 5.00 0.684
InfO 18 2.889 2.00 4.00 0.583 36 3.028 2.00 4.00 0.736 48 2.787 1.00 4.00 0.682 83 2.638 1.00 4.00 0.629 19 3.263 3.00 4.00 0.452
InfO2 18 3.167 3.00 4.00 0.383 36 3.222 2.00 4.00 0.591 48 2.745 2.00 4.00 0.482 83 3.062 2.00 4.00 0.687 19 2.579 2.00 3.00 0.507
I2supCsup 18 2.889 2.00 4.00 0.676 36 2.778 2.00 4.00 0.591 48 2.738 2.00 4.00 0.464 83 2.480 1.00 4.00 0.683 19 2.316 1.00 3.00 0.582
ProcCW 18 2.944 2.00 4.00 0.639 36 3.031 1.00 5.00 0.696 48 3.318 2.00 5.00 0.698 83 3.085 2.00 5.00 0.711 19 2.421 2.00 3.00 0.507
RmanW 18 3.556 3.00 4.00 0.511 36 2.929 2.00 4.00 0.409 48 2.729 1.00 4.00 0.676 83 2.710 1.00 4.00 0.713 19 2.684 2.00 4.00 0.582
Isup 18 3.222 2.00 4.00 0.548 36 2.332 1.00 3.00 0.569 48 2.863 1.00 4.00 0.605 83 2.936 2.00 5.00 0.817 19 3.000 2.00 4.00 0.333
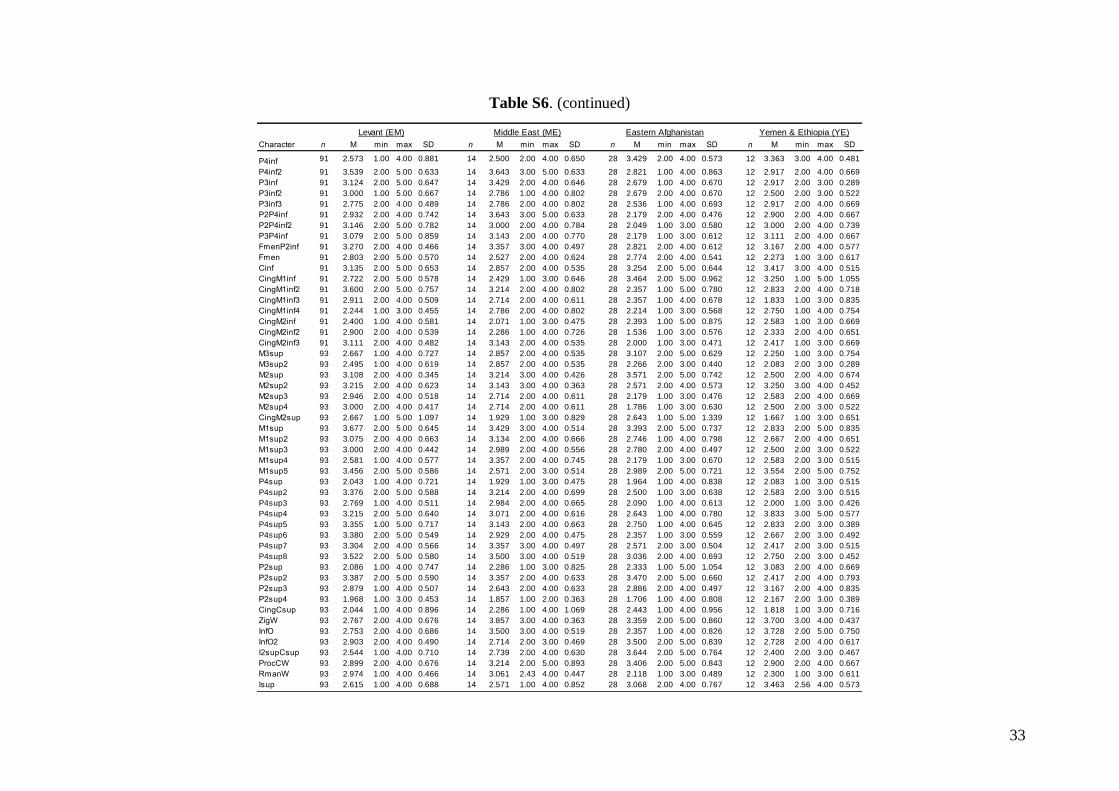
33
Table S6. (continued)
Levant (EM) Middle East (ME) Eastern Afghanistan Yemen & Ethiopia (YE)
Character n M min max SD n M min max SD n M min max SD n M min max SD
P4inf 91 2.573 1.00 4.00 0.881 14 2.500 2.00 4.00 0.650 28 3.429 2.00 4.00 0.573 12 3.363 3.00 4.00 0.481
P4inf2 91 3.539 2.00 5.00 0.633 14 3.643 3.00 5.00 0.633 28 2.821 1.00 4.00 0.863 12 2.917 2.00 4.00 0.669
P3inf 91 3.124 2.00 5.00 0.647 14 3.429 2.00 4.00 0.646 28 2.679 1.00 4.00 0.670 12 2.917 2.00 3.00 0.289
P3inf2 91 3.000 1.00 5.00 0.667 14 2.786 1.00 4.00 0.802 28 2.679 2.00 4.00 0.670 12 2.500 2.00 3.00 0.522
P3inf3 91 2.775 2.00 4.00 0.489 14 2.786 2.00 4.00 0.802 28 2.536 1.00 4.00 0.693 12 2.917 2.00 4.00 0.669
P2P4inf 91 2.932 2.00 4.00 0.742 14 3.643 3.00 5.00 0.633 28 2.179 2.00 4.00 0.476 12 2.900 2.00 4.00 0.667
P2P4inf2 91 3.146 2.00 5.00 0.782 14 3.000 2.00 4.00 0.784 28 2.049 1.00 3.00 0.580 12 3.000 2.00 4.00 0.739
P3P4inf 91 3.079 2.00 5.00 0.859 14 3.143 2.00 4.00 0.770 28 2.179 1.00 3.00 0.612 12 3.111 2.00 4.00 0.667
FmenP2inf 91 3.270 2.00 4.00 0.466 14 3.357 3.00 4.00 0.497 28 2.821 2.00 4.00 0.612 12 3.167 2.00 4.00 0.577
Fmen 91 2.803 2.00 5.00 0.570 14 2.527 2.00 4.00 0.624 28 2.774 2.00 4.00 0.541 12 2.273 1.00 3.00 0.617
Cinf 91 3.135 2.00 5.00 0.653 14 2.857 2.00 4.00 0.535 28 3.254 2.00 5.00 0.644 12 3.417 3.00 4.00 0.515
CingM1inf 91 2.722 2.00 5.00 0.578 14 2.429 1.00 3.00 0.646 28 3.464 2.00 5.00 0.962 12 3.250 1.00 5.00 1.055
CingM1inf2 91 3.600 2.00 5.00 0.757 14 3.214 2.00 4.00 0.802 28 2.357 1.00 5.00 0.780 12 2.833 2.00 4.00 0.718
CingM1inf3 91 2.911 2.00 4.00 0.509 14 2.714 2.00 4.00 0.611 28 2.357 1.00 4.00 0.678 12 1.833 1.00 3.00 0.835
CingM1inf4 91 2.244 1.00 3.00 0.455 14 2.786 2.00 4.00 0.802 28 2.214 1.00 3.00 0.568 12 2.750 1.00 4.00 0.754
CingM2inf 91 2.400 1.00 4.00 0.581 14 2.071 1.00 3.00 0.475 28 2.393 1.00 5.00 0.875 12 2.583 1.00 3.00 0.669
CingM2inf2 91 2.900 2.00 4.00 0.539 14 2.286 1.00 4.00 0.726 28 1.536 1.00 3.00 0.576 12 2.333 2.00 4.00 0.651
CingM2inf3 91 3.111 2.00 4.00 0.482 14 3.143 2.00 4.00 0.535 28 2.000 1.00 3.00 0.471 12 2.417 1.00 3.00 0.669
M3sup 93 2.667 1.00 4.00 0.727 14 2.857 2.00 4.00 0.535 28 3.107 2.00 5.00 0.629 12 2.250 1.00 3.00 0.754
M3sup2 93 2.495 1.00 4.00 0.619 14 2.857 2.00 4.00 0.535 28 2.266 2.00 3.00 0.440 12 2.083 2.00 3.00 0.289
M2sup 93 3.108 2.00 4.00 0.345 14 3.214 3.00 4.00 0.426 28 3.571 2.00 5.00 0.742 12 2.500 2.00 4.00 0.674
M2sup2 93 3.215 2.00 4.00 0.623 14 3.143 3.00 4.00 0.363 28 2.571 2.00 4.00 0.573 12 3.250 3.00 4.00 0.452
M2sup3 93 2.946 2.00 4.00 0.518 14 2.714 2.00 4.00 0.611 28 2.179 1.00 3.00 0.476 12 2.583 2.00 4.00 0.669
M2sup4 93 3.000 2.00 4.00 0.417 14 2.714 2.00 4.00 0.611 28 1.786 1.00 3.00 0.630 12 2.500 2.00 3.00 0.522
CingM2sup 93 2.667 1.00 5.00 1.097 14 1.929 1.00 3.00 0.829 28 2.643 1.00 5.00 1.339 12 1.667 1.00 3.00 0.651
M1sup 93 3.677 2.00 5.00 0.645 14 3.429 3.00 4.00 0.514 28 3.393 2.00 5.00 0.737 12 2.833 2.00 5.00 0.835
M1sup2 93 3.075 2.00 4.00 0.663 14 3.134 2.00 4.00 0.666 28 2.746 1.00 4.00 0.798 12 2.667 2.00 4.00 0.651
M1sup3 93 3.000 2.00 4.00 0.442 14 2.989 2.00 4.00 0.556 28 2.780 2.00 4.00 0.497 12 2.500 2.00 3.00 0.522
M1sup4 93 2.581 1.00 4.00 0.577 14 3.357 2.00 4.00 0.745 28 2.179 1.00 3.00 0.670 12 2.583 2.00 3.00 0.515
M1sup5 93 3.456 2.00 5.00 0.586 14 2.571 2.00 3.00 0.514 28 2.989 2.00 5.00 0.721 12 3.554 2.00 5.00 0.752
P4sup 93 2.043 1.00 4.00 0.721 14 1.929 1.00 3.00 0.475 28 1.964 1.00 4.00 0.838 12 2.083 1.00 3.00 0.515
P4sup2 93 3.376 2.00 5.00 0.588 14 3.214 2.00 4.00 0.699 28 2.500 1.00 3.00 0.638 12 2.583 2.00 3.00 0.515
P4sup3 93 2.769 1.00 4.00 0.511 14 2.984 2.00 4.00 0.665 28 2.090 1.00 4.00 0.613 12 2.000 1.00 3.00 0.426
P4sup4 93 3.215 2.00 5.00 0.640 14 3.071 2.00 4.00 0.616 28 2.643 1.00 4.00 0.780 12 3.833 3.00 5.00 0.577
P4sup5 93 3.355 1.00 5.00 0.717 14 3.143 2.00 4.00 0.663 28 2.750 1.00 4.00 0.645 12 2.833 2.00 3.00 0.389
P4sup6 93 3.380 2.00 5.00 0.549 14 2.929 2.00 4.00 0.475 28 2.357 1.00 3.00 0.559 12 2.667 2.00 3.00 0.492
P4sup7 93 3.304 2.00 4.00 0.566 14 3.357 3.00 4.00 0.497 28 2.571 2.00 3.00 0.504 12 2.417 2.00 3.00 0.515
P4sup8 93 3.522 2.00 5.00 0.580 14 3.500 3.00 4.00 0.519 28 3.036 2.00 4.00 0.693 12 2.750 2.00 3.00 0.452
P2sup 93 2.086 1.00 4.00 0.747 14 2.286 1.00 3.00 0.825 28 2.333 1.00 5.00 1.054 12 3.083 2.00 4.00 0.669
P2sup2 93 3.387 2.00 5.00 0.590 14 3.357 2.00 4.00 0.633 28 3.470 2.00 5.00 0.660 12 2.417 2.00 4.00 0.793
P2sup3 93 2.879 1.00 4.00 0.507 14 2.643 2.00 4.00 0.633 28 2.886 2.00 4.00 0.497 12 3.167 2.00 4.00 0.835
P2sup4 93 1.968 1.00 3.00 0.453 14 1.857 1.00 2.00 0.363 28 1.706 1.00 4.00 0.808 12 2.167 2.00 3.00 0.389
CingCsup 93 2.044 1.00 4.00 0.896 14 2.286 1.00 4.00 1.069 28 2.443 1.00 4.00 0.956 12 1.818 1.00 3.00 0.716
ZigW 93 2.767 2.00 4.00 0.676 14 3.857 3.00 4.00 0.363 28 3.359 2.00 5.00 0.860 12 3.700 3.00 4.00 0.437
InfO 93 2.753 2.00 4.00 0.686 14 3.500 3.00 4.00 0.519 28 2.357 1.00 4.00 0.826 12 3.728 2.00 5.00 0.750
InfO2 93 2.903 2.00 4.00 0.490 14 2.714 2.00 3.00 0.469 28 3.500 2.00 5.00 0.839 12 2.728 2.00 4.00 0.617
I2supCsup 93 2.544 1.00 4.00 0.710 14 2.739 2.00 4.00 0.630 28 3.644 2.00 5.00 0.764 12 2.400 2.00 3.00 0.467
ProcCW 93 2.899 2.00 4.00 0.676 14 3.214 2.00 5.00 0.893 28 3.406 2.00 5.00 0.843 12 2.900 2.00 4.00 0.667
RmanW 93 2.974 1.00 4.00 0.466 14 3.061 2.43 4.00 0.447 28 2.118 1.00 3.00 0.489 12 2.300 1.00 3.00 0.611
Isup 93 2.615 1.00 4.00 0.688 14 2.571 1.00 4.00 0.852 28 3.068 2.00 4.00 0.767 12 3.463 2.56 4.00 0.573
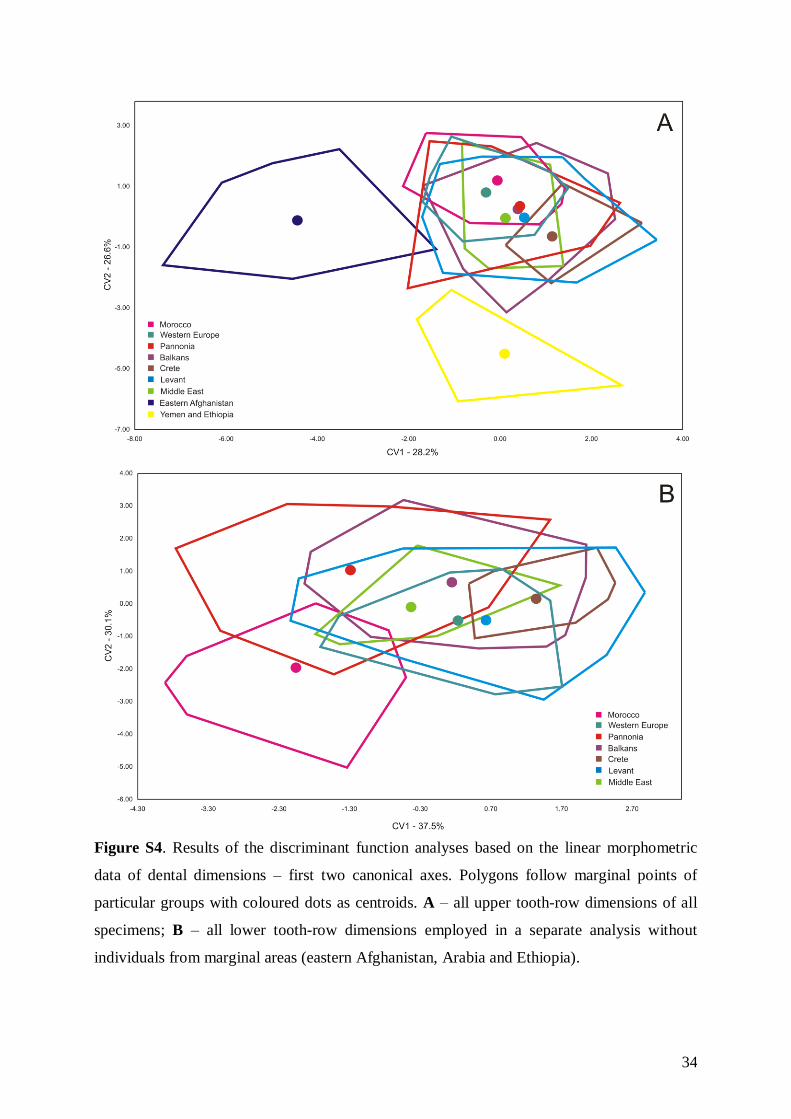
34
Figure S4. Results of the discriminant function analyses based on the linear morphometric
data of dental dimensions – first two canonical axes. Polygons follow marginal points of
particular groups with coloured dots as centroids. A – all upper tooth-row dimensions of all
specimens; B – all lower tooth-row dimensions employed in a separate analysis without
individuals from marginal areas (eastern Afghanistan, Arabia and Ethiopia).

35
Figure S5. The main shape variable (RW1) plotted against the centroid size (CS2) of the
lateral view of skull. Polygons and coloured dots as in Fig. S4.
Related Documents











