CANCER LETTERS Cancer Letters 90(1995) 123-131 Heparin receptors in two murine mammary adenocarcinomas with different metastatic ability: relationship with growth inhibition Gabriel Esteban Bertolesi*a7b, Lilia Lauria de Cidreb, Eugenia Sam-dote de Lustiga, Ana Maria Eijhna aArea Investigacidn, Institute de Oncologia “A.H. Roffo”, Universidad de Buenos Aires, Av. San Martin 5481. Buenos Aires 1417, Argentina bLuboratorio de Histologia Animal, Departamento de Biologia, Facultad de Ciencias Exactas y Noturales, Universidad de Buenos Aires, Ciudad Universitaria, Pobellon II, Buenos Aires 1428, Argentina Received 9 November 1994; revision received IO January 1995; accepted 10 January 1995 Abstract Binding of heparin to primary cultured cellsof two murine mammary adenocarcinomas with low (M3) and high (MM3) lung, metastatic capacitywas determined. Heparinbindingwas rapid, specific and saturable. MM3 cells grown for 24 h in fetal calf serum (FCS)-free.medium exhibited a higher number of binding sitesfor ‘H-heparin [(ll A 1) x 10’ sites per cell] than M3 cells [(6.9 f 0.6) x lo5 sites per cell]. However,when M3 cells weregrown in the presence of 2% FCS, they showed less heparin binding sites [(3.5 f 0.4) x 10’ sites per cell]. In contrast,dissocia- tion constants werevery similarfor MM3 and M3 cells grown with or without FCS (& = 2-4 x 10m9 M). Further- more, heparin inhibited MM3 and M3 cell growth both in the absence or presence of FCS.Competition studies showed that chemically modified heparins lacking antiproliferative effect (0-desulfated; O/N-desulfated N-acetylated and N- desulfated heparins) were not able to inhibit ‘H-heparin binding. N-desulfated N-acetylated heparin,which had par- tial antiproliferativeeffect, partially inhibited ‘H-heparin binding, while heparin with a high antiproliferative activity inhibited more than 90% 3H-heparinbinding. The antiproliferative effect of heparin and chemically modified heparins seems to be related to their binding ability to the cell membrane. Keywork Receptor; Heparin; Growth inhibition; Tumor; Metastasis 1. Introduction Proteoglycans and their polysaccharide chains, glycosaminoglycans (GAGS), are components of the extracellular matrix and cell membrane. GAGS *Corresponding author, Fax: +54 1 7909591, +54 I 5034370. from the extracellular matrix can regulate tumor cell growth by their interaction with cell mem2brane proteins [ 121. Heparin, a molecule closely related to the heparan sulfate component of proteoglycans, strongly inhibits vascular smooth muscle cell growth following arterial injury [8]. Furthermore, it inhibits cell proliferation of several cultured 0304-3835/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0304-3835(95)03693-Q

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CANCER LETTERS
Cancer Letters 90 (1995) 123-131
Heparin receptors in two murine mammary adenocarcinomas with different metastatic ability: relationship with
growth inhibition
Gabriel Esteban Bertolesi*a7b, Lilia Lauria de Cidreb, Eugenia Sam-dote de Lustiga, Ana Maria Eijhna
aArea Investigacidn, Institute de Oncologia “A.H. Roffo”, Universidad de Buenos Aires, Av. San Martin 5481. Buenos Aires 1417, Argentina
bLuboratorio de Histologia Animal, Departamento de Biologia, Facultad de Ciencias Exactas y Noturales, Universidad de Buenos Aires, Ciudad Universitaria, Pobellon II, Buenos Aires 1428, Argentina
Received 9 November 1994; revision received IO January 1995; accepted 10 January 1995
Abstract
Binding of heparin to primary cultured cells of two murine mammary adenocarcinomas with low (M3) and high (MM3) lung, metastatic capacity was determined. Heparin binding was rapid, specific and saturable. MM3 cells grown for 24 h in fetal calf serum (FCS)-free. medium exhibited a higher number of binding sites for ‘H-heparin [(ll A 1) x 10’ sites per cell] than M3 cells [(6.9 f 0.6) x lo5 sites per cell]. However, when M3 cells were grown in the presence of 2% FCS, they showed less heparin binding sites [(3.5 f 0.4) x 10’ sites per cell]. In contrast, dissocia- tion constants were very similar for MM3 and M3 cells grown with or without FCS (& = 2-4 x 10m9 M). Further- more, heparin inhibited MM3 and M3 cell growth both in the absence or presence of FCS. Competition studies showed that chemically modified heparins lacking antiproliferative effect (0-desulfated; O/N-desulfated N-acetylated and N- desulfated heparins) were not able to inhibit ‘H-heparin binding. N-desulfated N-acetylated heparin, which had par- tial antiproliferative effect, partially inhibited ‘H-heparin binding, while heparin with a high antiproliferative activity inhibited more than 90% 3H-heparin binding. The antiproliferative effect of heparin and chemically modified heparins seems to be related to their binding ability to the cell membrane.
Keywork Receptor; Heparin; Growth inhibition; Tumor; Metastasis
1. Introduction
Proteoglycans and their polysaccharide chains, glycosaminoglycans (GAGS), are components of the extracellular matrix and cell membrane. GAGS
*Corresponding author, Fax: +54 1 7909591, +54 I 5034370.
from the extracellular matrix can regulate tumor cell growth by their interaction with cell mem2brane proteins [ 121.
Heparin, a molecule closely related to the heparan sulfate component of proteoglycans, strongly inhibits vascular smooth muscle cell growth following arterial injury [8]. Furthermore, it inhibits cell proliferation of several cultured
0304-3835/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0304-3835(95)03693-Q

tumor cells such as the human sdrcom;t SW i : [ll]. two rhabdomiosarcomas 124.1 md !wo
adenocarcinomas with different metastatic capa- city [3].
Previous studies have shown that ihe <II:- ticoagulant property of heparin is not necessar\- for its antimitogenic actions [3]. Moreover. chemically modified heparins without anticoagu- lant activity. such as O-desulfated and N-desul- fated heparins lost their antiproliferative effect. while N-desulfated N-acetylated heparin partially retained its growth inhibitory activity 1.7.51.
Both normal [6] and malignant celis [4,24f possess high-affinity cell surface binding sites for heparin. Furthermore, heparin does not block the binding of mitogens. such as PDGF or insulin-like growth factor-l, to smooth muscle cell [25]. How- ever. it suppresses the induction of protoon- cogenes C-$X and c-my [2X]. The mechanism by which heparin inhibits cell proliferation and lhc relationship between this suppression of proWon’- cogenes and the antiproliferative activity 01‘ heparin are nor well understood.
We studied whether the antiproliferative effeci of heparin and chemically modified heparins without anticoagulant activity is associated with their cell surface binding capacity.
2. Materials and methods
2.1. Reagents Heparin (average molecular weight = 18 800;
APTI’ = 210 UI/mg) and chemically modified heparins - partially O-desulfated (O-desul), par- tially OIN desulfated N-acetylated (OI!V-desul .V- Ac), partially N-desulfated (X-desul) and partiallq :V-desulfated N-acetylated (N-desui %-AC i (APT’1 < I(! IWmgj, heparin-FIX and X-desul N-Ac heparin-FITC were generously gifted by Syntex S.A. Buenos Aires, Argentina. “H-Heparin (0.57 mCi/mg) was from New England Nuclear (Boston. MA). Pronase was purchased from Serva (Heidelberg,. Germany);. minimum- essential Eagle-5 medium (MEM; 410-1500), glutamine and trypan blue from Gibco (Grand Island, NY); fetal calf serum (FCS) from GEN (Buenos Aires. Argen- tina); 16-mm (24-well) plastic dishes from Falcon (Lincoln Park. NJ): and DNase from Sigma (SI Louis. MO).
Two BALB/c transplantable mammary adem-. carcmomas with low (M3) and high (-MM31 metastatic ability. previously described. were used 111 this work [1.2.21]. Briefly. the M3 tumor has .: 40’%, incidence of spontaneous lung metastasrh,. with an average of six nodules per mouse. and MM3 has 90% incidence and an average of 30 nodules per mouse. M3 and MM3 tumors have ;I tumor latency period of 6 + 3 and 12 + 2 days. respectively.
Female BAL.B!c inbred mice aged II) HW~:, received subcutaneous transplants of M.! or MM.1 tumor fragments by trocar (2 mm in diameter) Tumor-bearing mice were used 30 (M3) or .55 (MM3‘) days after transplantation. ‘This period represents approximately two- thirds ot’ the sur\jI. val time.
To prepare tumor cell suspenslonsl pooled M3 or MM3 tumors were removed, minced into line i‘ragments and enzymatically d&aggregated ab previously described 121. To obtain primary cul- tures, cells were seeded in complete medium (MEM plus 2 mM glutamine and 80 pg/rnl gen- tamicin) supplemented with 10% FCS and plated on 24-well plastic trays at 5 X IO4 cells/wel!.in 0.5 ml. The culture dishes were kept in a humidified 5’!/1 CO,-air atmosphere at 37°C. Cell monolayers were m&ly composed of tumor epithelioid cells w;lth less than 2% contaminating macrophages and fibroblast-like cells [ 1$2].
*‘. 4 C Ai prol~ftvutiork ussu !‘ After 24 h attached cells were washed three
times with phosphate buffered saline (PBS). with lo-min intervals between washes. then the com- plete medium (supplemented or not with 2% FCS) to which heparin or chemically modified heparins (200 @ml) was added. Control wells did not con- tain either heparin or modified heparins. The me- dium was replaced every 2 days.
Cell growth was eValudted by measurement oi protein content [20]. Protein content was directly proportional to the DNA content as measured with Hoescht 33258 reagent [15], and to cell num her, showing that 1 pg of cellular protein conten:

G. Estehan Bertolesi et al. /Cancer Letters 90 ( 199s) 123-131 125
corresponded to 35 ng of DNA and to 1300 M3 cells or to 1500 MM3 cells.
2.5. Measurement of binding of “H-labeled heparin Time-course and Scatchard analyses of binding
of radiolabeled heparin were used to determine presence, affinity and number of heparin binding sites on M3 or MM3 cells.
Up to 70-80% subconfluent cell monolayers were used for 3H-heparin binding assay. Twenty- four hours before performing the assay, cells were washed three times with PBS and complete medi- um with or without 2% FCS was added.
Heparin binding was determined as previously described [6]. Briefly, cells were washed three times with cold PBS and 3H-heparin was added to the cells (120 &well), which were kept at 4°C dur- ing the assay. To eliminate free 3H-heparin the cells were washed with PBS at 4’C and then solubilized in 0.2 N NaOH (100 ~1) and neutraliz- ed with 2 N HCl. Radioactivity was measured using a liquid scintillation counter to determine the amount of bound heparin. Nonspecific binding was measured with lOO-fold excess of unlabeled heparin and was less than 10% of total. Wells without cells were also exposed to 3H-heparin and binding to the plastic dish (< 5%) was considered background and subtracted from the appropriate experimental points.
2.6, Internalization of fluorescein isothiocyanate (FITC)-heparin or FITC N-desulfated N-acetyl- ated heparin
Internalization assay was carried out by the slightly modified method performed by Castellot and co-workers [6]. Briefly, coverslips with 70-80% confluent M3 or MM3 cells grown 24 h without FCS, were incubated 30 min at 4°C with FITC-heparin or FITC-N-desulfated N-acetylated heparin (25 pg/ml). Cells were washed four times with cold PBS and were maintained in complete medium without FCS at 37°C for 6 h. They were then washed with PBS and fixed in 3.7% formalin for 15 min at room temperature and then washed again in PBS. The coverslips were inverted on 90% glycerol-PBS (pH = 7.4) and observed within 2 h by a fluorescence microscope.
2.7. Statistical analyses All the experiments were repeated at least three
times with duplicates for each point in the ‘H- heparin binding assays and quadruplicates in the cell proliferation assay. Significance between groups was analyzed by Student’s t-test and ANOVA using the STAT PRIMER program and was assumed at P < 0.05.
3. Results
3.1. Binding of ‘H-heparin to MS and MM3 ceils The presence of specific heparin receptors was
tested on M3 and MM3 cells. A time course of 3H-heparin binding assay was
performed (Fig. 1). The specific binding reached a plateau after 30 min with both tumor cells. How- ever, MM3 cells bound twofold more 3H-heparin than the M3 type.
Specific binding of 3H-heparin to M3 cells was directly proportional to the cell number, reaching a plateau at a concentration of 2 x lo5 cells per well (under conditions of the assay) (Fig. 2). Maxi- mum binding capacity (MBC) of 3H-heparin was calculated by l/Bound vs. l/cell number plots (Fig. 2, insert). MBC was only 1% of the total 3H- heparin used in the assay. However, more than
/ d/’
4 ,b /
2/
0 lb 2b -- z-- 40 50 so ;o TIME (Min.)
Fig. 1. Time course on )H-heparin binding to M3 (p and MM3 (A) ceils. Subconfluent cultures (about 2.5 x IO5 cell/weli) grown 24 h without FCS, were incubated in 3H- heparin (2 x IO6 counts/min/well, 4.4 &well) at 4OC at the in- dicated time periods. The cells were washed four times with ice- cold PBS and processed for liquid scintillation counting. Data represent the mean of duplicate wells. Standard deviation was less than 10%. Background counts, obtained when empty wells were exposed to ‘H-heparin and nonspecific binding, have been substracted.

126
2 _-- -1 I-.--- .---- 7 r--..--
50 100 150 200 250 390
NUM6ER OF CELLS ( x lo- 3,
Fig. 2. Effect of M3 cell density on ‘H-heparin binding. Different number of M3 cells were incubated with jH-heparin (3. x 10’ counts/min/well) for 30 min at 4’C. Nonspecific binding with 100-fold excess unlabeled heparin were substracted. Insert: l/Bound vs. f/number of cells on M3 cells was analyzed. The line represents linear regression by kast squares analysis of data points. Y = 0.044321 X + 0.353862, r = 0.886.
.-.._- 6.- . ..~ -..-..- ;Tol------ ;; . ..-.--.. T--‘-. Is
3 H-HEPARIN (n&l)
Fig. 3. Heparin binding to MM3 (A) and M3 cells grown 24 h without (m or with 2% of KS (Cl). Cells were incubated with increasing concentrations of 3H-heparin for 30 min at 4’C. The cells were washed four times with ice-cofd PBS and processed for liquid scin- tillation countkg. Cells incubated with IWfold excessof unlabeled iieparin (nonspecific binding) have been substracted from the respective data. Insert: Scatchard plot analysis of heparin binding to MM3 (A) and M3 cells grown without (r, M with 2% of FCS (@ ‘H-heparin free was corrected by MBC. Lines represent linear regressions by least squares analysis of data points.
G. Esreban Bertolesi et al. /Cancer Letters 90 (1995) 123-131
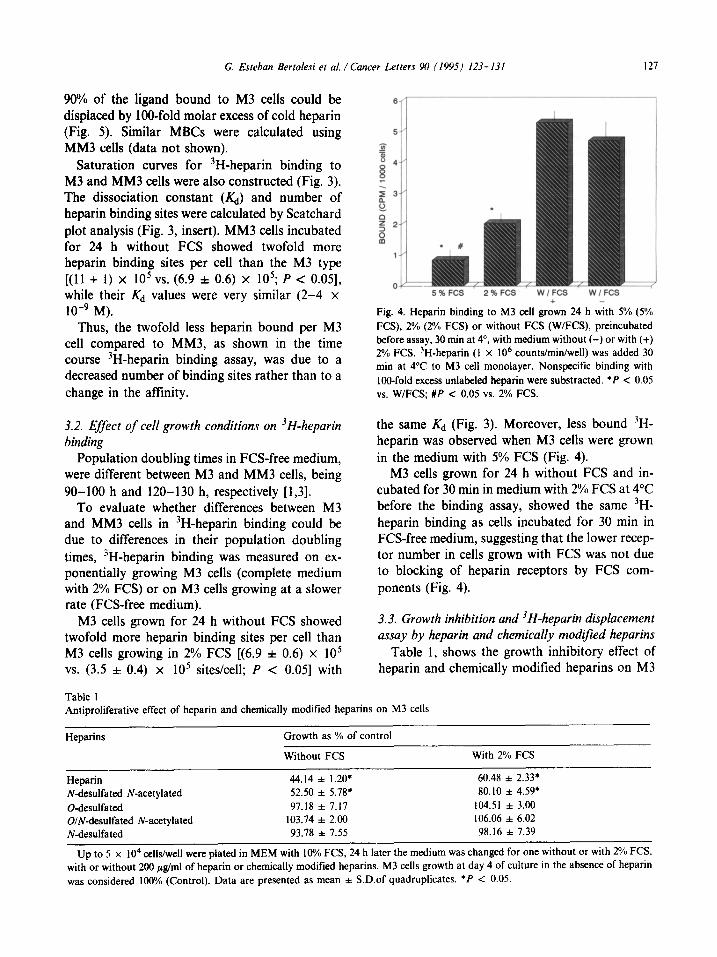
90% of the ligand bound to M3 cells could be displaced by IOO-fold molar excess of cold heparin (Fig. 5). Similar MBCs were calculated using MM3 cells (data not shown).
Saturation curves for 3H-heparin binding to M3 and MM3 cells were also constructed (Fig. 3). The dissociation constant (&) and number of heparin binding sites were calculated by Scatchard plot analysis (Fig. 3, insert). MM3 cells incubated for 24 h without FCS showed twofold more heparin binding sites per cell than the M3 type [(ll + 1) x IO5 vs. (6.9 f 0.6) x 105; P < 0.051, while their Kd values were very similar (2-4 x 1O-9 M).
Thus, the twofold less heparin bound per M3 cell compared to MM3, as shown in the time course 3H-heparin binding assay, was due to a decreased number of binding sites rather than to a change in the affinity.
Fig. 4. Heparin binding to M3 cell griwn 24 h with 5% (SO/; FCS), 2% (2% FCS) or without FCS (W/FCS), preincubatec 1 before assay, 30 mm at 4’. with medium without (-) or with (+) 2% FCS. 3H-heparin (1 x IO6 counts/min/well) was added 30 min at 4°C to M3 cell monolayer. Nonspecific binding with lOO-fold excess unlabeled heparin were substracted. *P < 0.05 vs. WIFCS; #P < 0.05 vs. 2% FCS.
3.2. Efect of cell growth conditions on 3H-heparin binding
Population doubling times in FCS-free medium, were different between M3 and MM3 cells, being 90-100 h and 120-130 h, respectively [1,3].
the same Kd (Fig. 3). Moreover, less bound “H- heparin was observed when M3 cells were grown in the medium with 5% FCS (Fig. 4).
To evaluate whether differences between M3 and MM3 cells in 3H-heparin binding could be due to differences in their population doubling times, “H-heparin binding was measured on ex- ponentially growing M3 cells (complete medium with 2% FCS) or on M3 cells growing at a slower rate (FCS-free medium).
M3 cells grown for 24 h without FCS and in- cubated for 30 min in medium with 2% FCS at 4°C before the binding assay, showed the same 3H- heparin binding as cells incubated for 30 min in FCS-free medium, suggesting that the lower recep- tor number in cells grown with FCS was not due to blocking of heparin receptors by FCS com- ponents (Fig. 4).
M3 cells grown for 24 h without FCS showed twofold more heparin binding sites per cell than M3 cells growing in 2% FCS [(6.9 + 0.6) x lo5 VS. (3.5 f 0.4) x lo5 sites/cell; P < 0.051 with
3.3. Growth inhibition and 3H-heparin displacement assay by heparin and chemically modified heparins
Table 1, shows the growth inhibitory effect of heparin and chemically modified heparins on M3
Table 1 Antiproliferative effect of heparin and chemically modified heparins on M3 cells
! 5%FCS 2%FCS WIFCS WlFCS
Heparins Growth as % of control
Without FCS With 2% FCS
Heparin 44.14 f 1.20* 60.48 f 2.33’ Ndesulfated N-acetylated 52.50 f 5.78* 80.10 f 4.59* O-desulfated 97.18 f 7.17 104.51 & 3.00 O/N-desulfated N-acetylated 103.74 f 2.00 106.06 f 6.02 N-desulfated 93.78 f 7.55 98.16 f 7.39
Up to 5 x IO4 cells/well were plated in MEM with 10% FCS, 24 h later the medium was changed for one without or with 2% FCS, with or without 200 &ml of heparin or chemically modified heparins. M3 cells growth at day 4 of culture in the absence of heparin was consldered 100% (Control). Data are presented as mean f S.D.of quadruplicates. *P < 0.05.

cells, grown with or without 2% FCS, after 4 days treatment. The antiproliferative effect of heparin was more evident in serum-deprived growing cells. Chemical modifications of the heparin molecule, might be translated into a change in its biological activity. O-desul and N-desul heparins (0-desul, O/N-desul N-AC and N-desul) lost their antipro- liferative activity. However, when the positive charge generated by desulfatation was neutralized by acetylation, the N-desul N-Ac heparins partial- ly retained their &proliferative effect 131.
To study whether the antiproliferative effect of chemically modified heparins was related to their capacity to bind heparin cell receptors, we per- formed a 3H-heparin bin ding assay in the presence of increasing amounts of these heparins (Fig. 5).
Unlabeled heparin competitively decreased the binding of ‘H-heparin and at a molar ratio of unlabeled:labeled heparin of 2.2 a 50% decrease was achieved, indicating that the binding charact- eristics of 3H-heparin were similar to that of unlabeled heparin (Fig. 5). A lOO-fold molar ex- cess of heparin decreased the binding of labeled
120 ,L;
, c
0 0 20 40 60 a0 loo 120
MOLAR EXCESS
Fig. 5. Inhibition of ‘H-heparin binding to M3 cells by unlabeled heparin or chemically modified heparins M3 ceils (80% confluent, 2.25 x IO5 cell/well) were incubated with ‘H- heparin (5 x IO5 counts/min/well) in the presence of various molar excess of unlabeled heparin (a, Wesulfated heparin ( 0). N-desulfated heparin (a) O/N-destdfated N-acetylated heparin (Cr, or N-desulfated Iv’-acetylated heparin (A) for 30 min at 4°C. Cell bound radioactivity was then determined. ‘H- heparin bound in the absence of unlabeled heparins was con- sidered 100%.
heparin by more than 90%. In contrast, there was no binding inhibition by heparins-without antipro- liferative activity (O-‘desul, OIN-desul, N-AC and N-desul). On the other hand, a 50% decrease in 3H-heparin binding occurred at 17-fold molar ex- cess of N-desul N-AC heparin, suggesting that this modified heparin is a partial agonist with eightfold less afftnity for cell surface receptors than un- modified heparin. A lOO-foid molar excess of N- desul N-AC heparin decreased the binding of label- ed heparin by 60%, which is in concordance with its antiproliferative activity.
3.4. Biological uctivity and internalization oj’FITC’- heparins
Internalization of heparin and N-desul N-Ac heparin was studied by using of FITC-derivatives. First, we measured the effect of FITC heparins on M3 cells growth. Fig. 6 shows that both FITC- heparins exhibited the same growth inhibition
*
i i
-. 3 5. /
TiME (Daysl
Fig. 6. Growth inhibition and reversibility of the antiprolifera- tive effect of FIX-heparin and FITC N-desulfared N- acetylated heparin. M3 celk were plated in MEM with 10% FCS. 24 h after, medium were changed for MEM with 2’% FCS without (A) or with 200 &ml of heparin (m, FITC heparin ( O), N-desulfated N-acetylated heparin (9 or FlTC N- desulfated .Wacetylated heparin (0). fn some cultures, (dashed line) the different heparins containing media were replaced after 2 days by MEM with 2% FCS. without GAGS. At dif- ferent times, subconfluent cell monolayers were washed with PBS and protein contents were measured. Data represent the mean of quadruplicate wells. Standard deviation was less than 8% of each mean.

G. Estehan Bertolesi et al. /Cancer Letters 90 (1995~ 123-131 129
Fig. 7. Internalization of FITC-heparin and FITC-N- desulfated N-acetylated heparin. MM3 ceils grown for 24 h without FCS were exposed to 25 @ml FITC-heparins for 30 min at 4’C. The FITC-heparins were removed and the cells were left to grow for another 6 h in medium without FCS at 37°C. They were then fixed and examined immediately by flu- orescence microscopy. (A) MM3 cells treated with FITC- heparin. (B) MM3 cells treated with FITC N-desulfated N- acetylated heparin. Bar = 10 pm.
effect as unlabeled ones and in a reversible fashion. Thus, M3 cells exposed to these GAGS for 2 days and then supplied with medium sup plemented with 2% FCS without GAGS showed the same population doubling time as controls (48-60 h), suggesting that the antiproliferative ac- tivity of either fluorescent or non-fluorescent GAGS was due to a cytostatic rather than a cyto- toxic effect.
FITC-heparin and FITC Ndesul N-AC heparin bound to cell membrane were internalized by M3 and MM3 cells. After 6 h, perinuclear and/or cyto- plasmic vesicles were seen in MM3 cells treated either with heparin or N-desul N-AC heparin (Fig. 7). Similar results were observed with M3 cells (data not shown).
4. Dlseussion
In this study, heparin binding to tumor cells with different metastatic capacity (M3 and MM3) was determined. Heparin binding was specific and saturable, suggesting a ligand-receptor type inter- action on the cell surface.
Only 1% of the ‘H-heparin used showed afti- nity for heparin receptors. This amount was lower than previously reported by other authors [4,5,17].
However, competition studies have shown that labeled and unlabeled heparin had similar receptor affinities. It is well known that the heparin molecu- lar structure is heterogenous and variable. A speci- fic sequence within heparin has high affinity for antithrombin and only a small fraction has an- ticoagulant activity [9,14]. Thus, heparin-receptor binding, as well as heparin-antithrombin interac- tion, may require a specific sequence within heparin.
Many extracellular matrix glycoproteins in- cluding laminin, libronectin, thrombospondin, and collagens bind to the GAG heparin [27], so heparin binding could be due to a heparin-extra- cellular matrix interaction. But, it has been previously shown that MM3 cells which had a high number of binding sites do not express fibronectin mRNA [18], while in M3 cells where fibronectin mRNA is expressed no Iibronectin is detected im- munohistochemically in FCS-free medium [ 161.
Otherwise, different extracellular matrix pro- teins differ in their binding affinity to heparin [27].
Both observations, together with the finding that the Scatchard plot showed only one site type, allow us to infer that the differences are probably due to a change in the receptor number rather than to the binding to extracellular matrix glyco- proteins.
As M3 and MM3 cells could also proliferate in serum-free medium, they could not be completely synchronized, so the doubling population time and the number of cells in each phase of the cell cycle was mostly dependent on the culture conditions (i.e. percentage of FCS) [l-3]. It is noteworthy that M3 cells growing in MEM with 2% FCS had a population doubling time of 48-60 h approx- imately and 3.5 x 10’ binding sites per cell, while M3 cells growing in FCS-free conditions had twofold more heparin binding sites (6.9 x lo5 sites per cell) and a twofold longer doubling time (90-100 h). Moreover, MM3 cells growing in FCS-free medium had the highest population doubling time (120- 130 h) and 11 x 10’ sites per cell. It is clear that heparin binding increases with the lengthening of the population doubling time. Studies on normal smooth muscle cells showed that growth arrested cells bound eightfold more heparin per cell than exponentially growing cells

[6], and that heparin inhibited cell proliferation by blocking mid- to late G, phase in the cell cycle [26]. Thus, it is possible that the number of heparin receptors on M3 and MM3 cell surfaces could be modified during the cell cycle, increasing in Go-G, phase.
The presence of different number of heparin receptors on several cell types, such as human monocytes (1.9 x lo6 sites per cell) [ 171. human endothelial cells (1 x 10” sites per cell) [IO]. two rhabdomyosarcoma with different metastatizing capacity (5 x 10” and 2.5 x 10h sites per cell) [24], B16 melanoma cells (5 x 10” sites per cell) [4] and smooth muscle cells ( 1 x 10’) may be due to intrinsic characteristics of each cell type and to the distribution of receptors in the different phases of the cell cycle (at the assay time).
M3 cells grown for 24 h without FCS presented twofold more heparin-receptors than those grown with 2% FCS. These results are in agreement with magnitude of the antiproliferative effect of heparin [3]. Furthermore, we observed that chemically modified heparins (0-desul, N-desul and OiN- desul N-AC) without antiproliferative activity, fail- ed to displace 3H-heparin bound to the cells. but N-desul N-AC heparin, which partially retained the antiproliferative effect, partially displaced the ‘H- heparin from the cell surface. Heparin and N-desul N-AC heparin were internalized, probably through a receptor-mediated process [6]. All these results suggest that the growth inhibition caused by heparin and N-desul N-AC heparin was media?ed by their binding to heparin receptor and/or inter- nalization.
Other molecular processes induced by heparin could be associated with its antiproliferative effect. Heparin inhibited the expression of early proto- oncogene c-fos and c-rnyc. in fibroblasts and smooth muscle cells [22,28] acting selectively on a protein kinase C-dependent pathways [7,23.28] and inhibited the activation of mitogenic activated protein kinase in response to FCS or phorbol myristate acetate [19]. Hepdrin also inhibited the induction or transcription of c-myh, another gene associated with cell cycle progression [26].
In this report, we observed a relationship be- tween cell growth inhibition by heparin and chemically modified heparins and their binding to the cell surface receptor
We are greatly indebted to Syntex S.A. (Buenos Aires, Argentina) and their scientific staff for pro- viding che@cally modified heparins. Critical reading of the manuscript by Dr Eduardo Cozza, Dr Elisa Bal de Kier Joff6 and Maria Adela Jasnis is also acknowledged. This study was supported by a grant from the University of Buenos Aires (UBA; EX-034), and by a grant from the Council for Scientific and Technical Research (CONICET: 1395’92). G.E.B. is a Research Fellow and L.L.C. is a professor of the University of Buenos Aires. A.M.E. and E.S.L. are members of the Scientific Career (CONICET).
References
Bal de Krcr JoBe. t.. Purrcelh. L.. vidal. MC’. .~nc
Sacerdote de Lustig. E. (1983) Characterization of two
murine mammary adenocarcinoma tumors with different
metastatic ability. J. Exp. Clin. Cancer Res.. 2. I51- 160
Bal de Kier Joffe, E., Puricelli. L. and Sacerdote dc
Lustig, E. (1986) Modified adhesion behaivour after in
vitro passage of two related murine mammary adenocar-
cinemas with different metastasizing ability. Invasion
Metastasis, 6. 302-312.
Bertolesi. GE.. Lauria de Cidre. 1.. and Eijan, A.M
(1994) Growth inhibition in vitro of murine mammary
adenocarcinoma cells by heparin and chemically modi-
fied heparins. Tumor Biol.. 15. 2?5-283.
Biswas, C (1988) Heparin and heparan sulfate binding
sites on 816 melanoma cells. J. Cell. Physiol.. I36
147-153.
Castellot, J.J.. Beeker. D.L., Rosenberg, R.D. and Kar-
novsky. N.J. (1984) Structural determinants of the capa-
city of heparin to inhibit the proliferation of vascular
smooth muscle cells. J. Cell. Physiol., 120. 315-320.
Castellot, J.J.. Wong, K., Herman, B.. Hoover. R.L.
Albertini, D.F.. Wrigth. ‘I’.(‘.. Caleb, 8.1.. and Karnov-
sky. M. (19X5) Binding and internalization of heparin bv
vascular smooth muscle cells. J. Cell. Physiol.. 124.
IL-20
Castcllot. J J Pukac. i A.. Caleb B.L.., Wqth. .J i’
and Karnovsky, M.J. (1989) Heparin selectively inhibits
a protein kinase C-dependent mechanism of ccl1 cycle
progression in calf aortic smooth muscle cells. J. Cell
Biol.. 109. 3147-3155.
C’lowes. A.W. and Karnovsy, M.J. ( 1977) Suppression by
heparin of smooth muscle cell proliferation in injured
arteries. Nature. 265. 625-626.
Evans, D.L.. Marshall. C.J.. Christey. P.B. and Carrel.
R.W. (1992) Heparin binding site. conformational
change and activition ofantithrombin. Biochemistry. 3!.
I2629- I2642

G. Esrehan Bertolesi et a/. /Cancer Letters 90 (1995) 123-131 131
[IO] Glimelius, B., Busch, C. and Hook, M. (1978) Binding of heparin on the surface of cultured human endothelial cells. Tromb. Res., 12, 773-782.
]I I] Halper, J. and Carter, B.J. (1989) Modulation of growth of human carcinoma SW-13 cells by heparin and growth factors. J. Cell. Physiol., 141, 16-23.
[ 121 Iozzo, R.V. (1988) Proteoglycans and neoplasia. Cancer Metastasis Rev., 7, 39-50
[13] Kjellen, L., Oldberg, A., Rubin, K. and Hook, M. (1977) Binding of heparin and heparan sulfate to rat liver cells. B&hem. Biophys. Res. Commun., 74, 126-133.
[14] Lindahl, U., Btickstrom, G. and Thunberg, L. (1983) The antithrombin-binding sequence in heparin. Identifi- cation of an essential 6-O-sulfate group. J. Biol. Chem., 16, 9826-9830.
(151 Labarca, C. and Paigen, K. (1980) A simple, rapid and sensitive DNA assay procedure. Anal. Biochem., 102, 344-352.
[16] Ladeda, V., Puricelli, L., Urtreger, A., Alonso, D., Korn- blihtt, A. and Ball de Kier Joffe, E. (1994) Action de1 Suero y de Inhibidores Proteasicos Sobre la Expresion in vitro de Fibronectina. VI Congreso de La Sociedad Argentina de Ciencias Morfolbgicas, 9la.
[I71 Leung, L., Katsuyasu, S. and Grant, D. (1989) Heparin binds to human monocytes and modulates their pro- coagulant activities and secretory phenotypes. Effect of histidine-rich glycoprotein. Blood, 73, l77- 184.
[I81 Muro, A., Puricelli, L., Komblihtt, A.R. and Bal de Kier Joffe, E. (1991) Inverse correlation between tibronectin mRNA levels and the metastatic potential of two murine mammary adenocarcinomas. Invation Metastasis, 11, 281-287.
[ 191 Ottlinger, M.E., Pukac, L.A. and Karnovsky, M.J. (1993) Heparin inhibits mitogenic actived protein kinase activation in intact rat vascular smooth muscle cells. J. Biol. Chem., 268, 19173-19176.
[20] Oyama, V. and Eagle, H. (1956) Measurament of cell growth in tissue culture with phenol reagent (folin- ciocalteau). Proc. Sot. Exp. Biol. Med., 91, 305-307.
[21] Pereyra-Alfonso, S., Haedo, A. and Bal de Kier Joffe, E. (1988) Correlation betwen urokinase-type plasminogen activator production and the metastasizing ability of two murine mammary adenocarcinomas. Int. J. Cancer, 42, 59-63.
[22] Pukac, L.A., Castellot, J.J., Wrigth, T.C., Caleb, B.L. and Karnovsky, M.J. (1990) Heparin inhibits c-fos and c- myc mRNA expression in vascular smooth muscle cells. Cell Regul., I, 435-443.
[23] Pukac, L.A., Ottlinger, M.E. and Kamovsky, M.J. (1992) Heparin supresses specific second messenger pathways for protooncogene expression in rat vascular smooth muscle cells. J. Biol. Chem., 267, 3707-371 I.
[24] Redini, F., Moczar, E., Antoine, E. and Poupon, M.F. (1989) Binding and internalization of exogenous glycosaminoglicans in weakly and highly metastatic rhabdomyosarcoma cells. Biochem. Biophys. Acta, 991, 359-366.
[25] Reilly, CF., Fritze, L.M.S. and Rosenberg, R.D. (1988) Heparin-Like molecules regulate the number of epider- mal growth factors on vascular smooth muscle cell. J. Cell Physiol., 136, 26-32.
1261 Reilly, F.C., Kindy, MS., Brown, K.E., Rosenberg, R.D. and Sonenshein, GE. (1989) Heparin prevents vas- cular smooth muscle cell progression through the GI phase of the cell cycle. J. Biol. Chem., 264, 6990-6995.
[27] San Antonio, J.D., Slaver, J., Lawler, J., Kamovsky, M.J. and Lander, A.D. (1993) Specificity in the interac- tions of extracellular matrix proteins with subpopula- tions of the glycosaminoglycan heparin. B&hem., 32, 4746-4755.
[28] Wrigth, T.C.Jr., Pukac, L.A., Castellot, J.J., Karnovsky, M.J., Levine, R.A., Kim-Park, H.Y. and Campisi, J. (1989) Heparin suppresses the induction of c-fos and c- myc mRNA in murine libroblasts by selective inhibition of a protein kinase C-dependent pathway. Proc. Nat. Acad. Sci. USA, 86, 3199-3203.
Related Documents











