Hemostatic status in long-term surviving xenografts David G. Molleví, PhD, a Mireia M. Ginestà, PhD, a Pere Domenech, PhD, b Teresa Serrano, PhD, c Joan Figueras, PhD, a and Eduard Jaurrieta, PhD, a Barcelona, Spain Background. Liver xenotransplantation presents, apart from immunologic problems, metabolic incompatibilities between species. The liver plays a key role in blood coagulation. The aim of this study is to describe the hemostatic status of long-term surviving xenografts in a hamster-to-rat liver xenotransplantation model. Methods. Orthotopic liver transplantation with Tacrolimus and MMF was carried out with Golden Syrian hamsters, Brown Norway, or Dark Agouti rats as donors and Lewis rats as recipients. Prothrombine time (PT), activated partial thromboplastin time (APTT), antithrombin, protein-C, free protein-S, TAT-complexes, and factors V and VIII were assessed using standard methods. Results. Protein-C was absent in rats, but values in xenotransplanted animals increased progressively toward those recorded in hamsters. Xenotransplanted animals also acquired PT, APTT, free protein-S, and antithrombin levels similar to those of donors and we observed a substantial activation of coagulation especially 7 days post-transplantation. Despite TAT high levels, we did not find thrombotic alterations in the histologic analysis of grafts. Conclusions. These results reflect a destabilization of the thrombotic-hemostatic balance, not associated with consumption coagulopathy, which gradually disappears. This deregulation is a general imbalance resulting from the replacement of all the components of hepatic synthesis. After 100 days of xenotransplantation, the absence of symptoms of thrombosis or hemorrhage suggests that the change of hemostatic status takes place under conditions of relative equilibrium. (Surgery 2006;139:775-81.) From the Department of Surgery and Surgical Specialties, University of Barcelona School of Medecine Campus Bellvitge, L’Hospitalet de Llobregat, Barcelona, Catalonia, a Hematology and Hemotherapy Service, Hospital Universitari de Bellvitge, b and Pathology Service, Hospital Universitari de Bellvitge, c Barcelona, Spain Hepatic xenotransplantation is of particular interest because of the role of the liver in this metabolic function and because the physiology of blood coagulation is closely related to hepatic func- tion. The liver synthesizes and controls most factors involved in coagulation and fibrinolysis, with the exception of von Willebrand factor, t-PA and u-PA. Furthermore, the clearance of activated clotting factors, fibrinolytic factors, activating complexes, and end-products of fibrinogen-to-fibrin conver- sion are mediated by the endothelial system of the liver. 1 Abnormal liver function may lead to hemo- static impairment either a lack of vitamin K, or by the synthesis of non-functional clotting factors, or an increase in consumption of these factors, or a disturbance in the clearance of circulating acti- vated components of the system. 2 Xenotransplantation, besides the problems caused by operative manipulation per se or rejec- tion, may led to metabolic incompatibilities be- tween donor and recipient species may therefore cause homeostatic imbalances in certain metabo- lites such as those involved in the coagulation cas- cade and fibrinolysis. Few studies have focused on metabolic aspects in long-surviving xenografts. 3,4 To date, most articles concerning coagulation in- compatibilities in xenotransplantation are devel- oped in vitro 5–8 and ex vivo models. 9 There is a noticeable lack of in vivo studies involving long- Supported by Ministry of Health, Spanish Government grant FIS 01/1199. David G. Molleví, PhD and Mireia M. Ginestà, PhD contributed equally to this work. 0039-6060/$ - see front matter Accepted for publication December 29, 2005. Reprint requests: David G. Molleví, Unitat de Recerca Quirúr- gica, Departament de Cirurgia i Especialitats Quirúrgiques, Universitat de Barcelona Campus de Bellvitge, Pavelló de Govern 4 a planta, laboratori 4120, 08907 L’Hospitalet de Llobregat, Barcelona, Catalonia. E-mail: [email protected] 0039-6060/$ - see front matter © 2006 Mosby, Inc. All rights reserved. doi:10.1016/j.surg.2005.12.007 SURGERY 775

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hemostatic status in long-termsurviving xenograftsDavid G. Molleví, PhD,a Mireia M. Ginestà, PhD,a Pere Domenech, PhD,b Teresa Serrano, PhD,c
Joan Figueras, PhD,a and Eduard Jaurrieta, PhD,a Barcelona, Spain
Background. Liver xenotransplantation presents, apart from immunologic problems, metabolicincompatibilities between species. The liver plays a key role in blood coagulation. The aim of this studyis to describe the hemostatic status of long-term surviving xenografts in a hamster-to-rat liverxenotransplantation model.Methods. Orthotopic liver transplantation with Tacrolimus and MMF was carried out with GoldenSyrian hamsters, Brown Norway, or Dark Agouti rats as donors and Lewis rats as recipients.Prothrombine time (PT), activated partial thromboplastin time (APTT), antithrombin, protein-C, freeprotein-S, TAT-complexes, and factors V and VIII were assessed using standard methods.Results. Protein-C was absent in rats, but values in xenotransplanted animals increased progressivelytoward those recorded in hamsters. Xenotransplanted animals also acquired PT, APTT, free protein-S,and antithrombin levels similar to those of donors and we observed a substantial activation ofcoagulation especially 7 days post-transplantation. Despite TAT high levels, we did not find thromboticalterations in the histologic analysis of grafts.Conclusions. These results reflect a destabilization of the thrombotic-hemostatic balance, notassociated with consumption coagulopathy, which gradually disappears. This deregulation is ageneral imbalance resulting from the replacement of all the components of hepatic synthesis. After100 days of xenotransplantation, the absence of symptoms of thrombosis or hemorrhage suggeststhat the change of hemostatic status takes place under conditions of relative equilibrium.(Surgery 2006;139:775-81.)
From the Department of Surgery and Surgical Specialties, University of Barcelona School of Medecine CampusBellvitge, L’Hospitalet de Llobregat, Barcelona, Catalonia,a Hematology and Hemotherapy Service, Hospital
Universitari de Bellvitge,b and Pathology Service, Hospital Universitari de Bellvitge,c Barcelona, SpainHepatic xenotransplantation is of particularinterest because of the role of the liver in thismetabolic function and because the physiology ofblood coagulation is closely related to hepatic func-tion. The liver synthesizes and controls most factorsinvolved in coagulation and fibrinolysis, with theexception of von Willebrand factor, t-PA and u-PA.
Supported by Ministry of Health, Spanish Government grant FIS01/1199.
David G. Molleví, PhD and Mireia M. Ginestà, PhD contributedequally to this work.
0039-6060/$ - see front matter
Accepted for publication December 29, 2005.
Reprint requests: David G. Molleví, Unitat de Recerca Quirúr-gica, Departament de Cirurgia i Especialitats Quirúrgiques,Universitat de Barcelona Campus de Bellvitge, Pavelló deGovern 4a planta, laboratori 4120, 08907 L’Hospitalet deLlobregat, Barcelona, Catalonia.E-mail: [email protected]
0039-6060/$ - see front matter
© 2006 Mosby, Inc. All rights reserved.
doi:10.1016/j.surg.2005.12.007
Furthermore, the clearance of activated clottingfactors, fibrinolytic factors, activating complexes,and end-products of fibrinogen-to-fibrin conver-sion are mediated by the endothelial system of theliver.1 Abnormal liver function may lead to hemo-static impairment either a lack of vitamin K, or bythe synthesis of non-functional clotting factors, oran increase in consumption of these factors, or adisturbance in the clearance of circulating acti-vated components of the system.2
Xenotransplantation, besides the problemscaused by operative manipulation per se or rejec-tion, may led to metabolic incompatibilities be-tween donor and recipient species may thereforecause homeostatic imbalances in certain metabo-lites such as those involved in the coagulation cas-cade and fibrinolysis. Few studies have focused onmetabolic aspects in long-surviving xenografts.3,4
To date, most articles concerning coagulation in-compatibilities in xenotransplantation are devel-oped in vitro5–8 and ex vivo models.9 There is a
noticeable lack of in vivo studies involving long-SURGERY 775

.
776 Molleví et al SurgeryJune 2006
term surviving xenografts, and no models has yetreached at least 100 days of survival.
However, hyperacute rejection and acute rejec-tion involve disturbances in the coagulation cas-cade and platelet aggregation. Even if these immu-nologic responses are inhibited completely, thehemostatic imbalances may persist as the conse-quence of metabolic incompatibilities between spe-cies. Therefore, the importance of our study focuson the fact that despite the known differences be-tween rats and hamsters in relation to haemosta-sis,10,11 in a short term there are no known meta-bolic disturbances that affect the survival of theanimal but it could be interesting to know. Theevolution of parameters of coagulation in the long-term surviving xenograft is of considerable interestbecause the coagulation system depends on anequilibrium in all components of the hemostaticpathway. In the hamster-to-rat xenotransplantmodel Valdivia and colleagues12 investigated coag-ulation parameters (Prothrombin time, bleedingtime, and protein C) demonstrating the acquisitionof donor levels in recipient animals. The aim of ourstudy was to describe, more accurately, the hemo-static status of long-term surviving hepatic xeno-grafts in the hamster-to-rat xenotransplantationmodel.
MATERIALS AND METHODSAnimals. Male adult Golden Syrian (GS) ham-
sters weighing greater than 100 g, Dark Agouti
Table I. Study groups*
Group (n) Animals Im
1 (10) Lewis rats No2 (8) BN rats No3 (6) Hamster No4 (6) DA to Lewis rats MMF: 50
Tacrolim5 (6) DA to Lewis rats MMF: 50
Tacrolim6 (7) BN to Lewis rats MMF: 50
Tacrolim31 to day
7 (7) Hamster to rat MMF: 50Tacrolim
8 (7) Hamster to rat MMF: 50Tacrolim
9 (6) Hamster to rat MMF: 50Tacrolim31 to 100
*All animals in groups 1, 2, 3, 6 and 9 were followed and sacrificedpost-transplantation and groups 5 and 8 at day �30 post-transplantation
(DA), and Brown Norway (BN) rats (200 to 270 g)
were used as donors and Lewis rats (200 to 270 g)as recipients. All animals were housed in pathogen-free conditions. The study was approved by theCommittee of Ethics of the University of Barcelona.
Experimental groups. The design involved con-trol allotransplant, and xenotransplant groups an-alyzed at different days post-transplantation (TableI). When we studied the 100-day allotransplantgroup (group 6), we noted that the Dark Agouti-to-Lewis combination had evidence of considerablerejection. Because the xenotransplant groups 100days post-transplant (GS Hamster to Lewis rat)were not experiencing problems, and because themain focus of the study was to compare allogenictransplantation versus xenogenic transplantation,we maintained the same immunosuppressive pro-tocol but used a different, fully allogenic straincombination, the Brown Norway–Lewis combina-tion, instead of increasing the immunosuppressionin the DA-to-Lewis combination. The protocol de-tailed below was sufficient to achieve survivals of100 days in the Hamster-to-Lew Rat model and inthe BN-to-Lew, but not in the DA-to-Lew transplan-tation model. Control groups were also observedfor 100 days.
Immunosuppressive protocol. Tacrolimus (Fuji-sawa-GmbH, Munich, Germany), was given intra-muscularly into rat hind legs at a dose of 0.5 mg/Kg/day from day 0 to �30 post-transplantation anda dose of 0.2 mg/Kg/day from day �31 to �100post-transplantation; mycophenyl mofetil (MMF)
uppressive treatmentDay of sacrifice
(postransplantation)
�100�100�100
g day; 8 days �7o day 7: 0.5 mg/kg dayg day; 8 days �30o day 30: 0.5 mg/kg dayg day; 8 days �100o day 30: 0.5 mg/kg day0.2 mg/kg dayg day �7o day 7: 0.5 mg/kg dayg day; 8 days �30o 30: 0.5 mg/kg dayg day; 8 dayso 30: 0.5 mg/kg day
�100
g/kg day
�100 post-transplantation. Groups 4 and 7 were sacrificed at day �7
munos
mg/kus: 0 tmg/k
us: 0 tmg/k
us: 0 t100:mg/k
us: 0 tmg/k
us: 0 tmg/k
us: 0 t: 0.2 m
at day
(CellCeptTM, Roche Laboratories, Hertfordshire,

Surgery Molleví et al 777Volume 139, Number 6
UK) was diluted in 5% glucose solution and admin-istered by gavage at a dose of 50 mg/kg for 8 days.Immunosuppression was scheduled as representedin Table I.
Operative procedure. All operative procedureswere conducted under isoflurane anesthesia. Or-thotopic liver transplantation was carried out withrevascularization without hepatic artery reconstruc-tion, as described previously by Kamada andCalne.13 After the abdominal wall was closed, 0.5ml of H2CO3 1 M, 2 mg ciprofloxacin, and 1 mlRinger lactate was injected through the penile vein.
Blood sampling procedure. Blood samples wereobtained by aortic puncture at sacrifice. Blood wascollected in 0.38% tri-sodium citrate (final concen-tration). Plasma fractions were obtained by centrif-ugation (15 min, 4°C, 1,800g), quickly frozen inliquid nitrogen, and stored at �80°C until use. Forliver function tests blood samples were harvested bytail bleeding on days 0, 7, 15, 30, 50, 75, and 100after transplantation.
Liver function tests. Albumin, alanine amino-transferase (ALT), and aspartate aminotransferase(AST) were determined from sera at days 0, �7,�15, �30, �50, �75, and 100, using standard spec-trophotometric methods (normal ranges in controlanimals: ALT, 30-77 UI/l; AST, 57-162 UI/l; albu-min, 35-45 g/l, the hamster animals).
Hemostatic parameters. Determinations of pro-thrombin and factor V (coagulative method; usinghuman factor V- deficient plasma from Instrumen-tation Laboratory, Milan, Italy) were carried outusing a rabbit thromboplastin, IL test PT-Fibrino-gen HS PLUS from IL. APTT and factor VIII (co-agulative method; using human factor VIII-defi-cient plasma from IL) were obtained using IL TestAPTT Lyophilized silica from IL. All these methodswere carried out with an automatic coagulometer(ACL 9000, Instrumentation Laboratory, Milan, It-aly). TAT complexes were evaluated using a quan-titative sandwich enzyme immunoassay for the invitro determination of thrombin/antithrombincomplex in plasma (Enzygnost TAT micro, DadeBehring, Marburg, Germany). Free protein S wasassayed using a latex ligand immunoassay for ci-trated plasma. The procedure is based on the mea-surement of the increase in turbidity produced bythe agglutination of 2 latex reagents induced byfree protein S. The degree of agglutination is di-rectly proportional to the free protein S concentra-tion in the test sample (Instrumentation Labora-tory). Protein C was measured by a chromogenicassay for quantitative determination in citrated
plasma (Instrumentation Laboratory).Histology. Two graft samples, harvested fromthe medial lobe and from the hepatic hilum, werefixed in formalin 10% and embedded in paraffin.Liver samples were sectioned (3 �m) and stainedwith hematoxylin-eosin and Masson trichrome. Allslices were evaluated by 2 observers in a blindedfashion. Three different hepatic areas were studied:periportal, midzonal, and centrilobular. The pa-rameters evaluated included: lymphocytic infil-trate, endothelialitis, cholangiolar proliferation,bile duct damage, hepatocellular necrosis, apopto-sis, dilatation and congestion of sinusoids; each wasscored as: 0 � no presence, 1 � mild, 2 � moder-ate, 3 � severe. Fibrosis was evaluated with Massontrichrome staining and scored as: 0 � absence, 1 �portal fibrous expansion, 2 � porto-portal bridges,3 � porto-centrolobular bridges.
Statistics. Descriptive statistics, Kruskal-Wallis,and Mann-Whitney U test were used (P � .05 wasconsidered significant).
RESULTSGraft survival rates. All transplanted animals
reached the end-point of the study (depending onthe group). The graft survival rate was 100% with-out signs of hemorrhages, even not observed aftertail cutting for blood collection.
Liver function test and histology. Most determi-nations of serum transaminases AST and ALT (Ta-ble II), measured on different days post-transplant,were within the range of normal values in rats,although in the allotransplant groups, some valueswere slightly higher but accompanied by a relativelynormal histology (see below). Serum concentra-tions of albumin were slightly below normal butstable and without statistically significant differ-ences in relation to normal hamster controls(range 31-37 g/L) (Table II). Regarding the histol-ogy, in the allotransplant groups on day 7 post-transplantation the grafts showed only a slightcholangiolar periportal proliferation that did notdistort the hepatic architecture. In group 5 (DA-to-Lewis) a moderate cholangiolar proliferation waspresent accompanied by an increase in fibrosis anda slight hepatic architectural distortion. Howeveron day 100 post-transplantation, in the BN-to-Lewcombination (group 6), the cholangiolar prolifer-ation had increased markedly and was consideredsevere, accompanied by fibrosis and a clear loss ofthe parenchyma architecture. No canalicular cho-lestasis or lesions in the primary biliary ducts wereobserved (Fig 1A). In the xenotransplant groups,no endothelialitis was observed in the portal orcentrolobular veins nor were lesions in the main
bile ducts. No ischemia lesions were seen. Group 7
ssed in
778 Molleví et al SurgeryJune 2006
(Hamster-to-rat, 7 days post xenotransplant)showed a moderate inflammatory infiltrate ofmononuclear cells in the portal spaces, withoutspillover to the parenchyma. The infiltrate com-prised both activated and mature lymphocytes. Byday 30 post-transplantation (group 8) the infiltratewas much less. At 100 days post xenotransplant(group 9) the parenchyma was completely pre-served, with a very slight infiltrate due to non-activated lymphocytes and a slight cholangiolarproliferation (Fig 1B).
Hemostatic parameters. There were differencesin hemostatic parameters between control Lewis ratsand control Golden Syrian hamsters (Table III).
Prothrombin time. This parameter evaluates theextrinsic pathway of coagulation. The control val-ues of the recipient rats (Lewis) and hamsters haddifferences (Table III). The prothrombin time(PT) was longer in the three allotransplant groupsthan in control rats, but shorter in the xenotrans-plant groups than in controls (Table III). Theselatter values were much closer to those of controlhamsters; there were no statistical differences be-tween the xenotransplant groups and the hamsterbaseline values, whereas all the allotransplantgroups were statistically different compared to thenormal values in rats. These differences were
Table II. Liver function tests†
Group 1
Allotrans
Group 4 Gro
ALT d 0 45.5 43 4ALT d � 7 39 63.5 15ALT d � 15 43 — 4ALT d � 30 47 — 4ALT d � 50 56.5 — —ALT d � 75 46 — —ALT d � 100 44 — —AST d 0 93 81 7AST d � 7 121 142.5 23AST d � 15 97 — 19AST d � 30 131 — 14AST d � 50 130 — —AST d � 75 124 — —AST d � 100 96 — —Albumin d 0 40* 43.5 4Albumin d � 7 39* 32.5 3Albumin d � 15 40.5* — 3Albumin d � 30 44* — 2Albumin d � 50 36* — —Albumin d � 75 41* — —Albumin d � 100 38* — —
*P � .05 all groups to control group with immunosuppression.†All values are median values; levels of transaminases ALT and AST expreMann-Whitney U test.
present on day 7 post-transplant.
Activated partial thromboplastin time. This pa-rameter evaluates the intrinsic pathway of coagula-tion. Normal values in hamsters and rats also dif-fered (Table III). The 3 allotransplant groups hadvalues similar to those of the donor species (rats).The 3 xenotransplant groups had values similar tothose in the hamsters and therefore were greaterthan those of the control and allotransplanted rats.
Factor V. Values of this factor were similar in theplasma of rat and hamster controls. Plasma levelswere significantly lower in the allotransplantgroups than in control rats (Table III). In thexenotransplant groups at 7 and 30 days the factor Vplasma levels were slightly greater than in both ratand hamster controls but the differences were notstatistically significant. Factor V plasma levels in thegroup at 100 days were significantly lower than incontrol rats, but not different from those of thecontrol hamster donors.
Factor VIII. Rat and hamster values for plasmafactor VIII were similar (Table III). All the trans-plant and xenotransplant groups presented greaterfactor VIII levels than the control values in the ratsalthough not all the differences were statisticallysignificant. We wish to point out that factor VIII isan acute phase protein and is not synthesized ex-clusively by the liver; the spleen, kidney and lungs
tion Xenotransplantation
Group 6 Group 7 Group 8 Group 9
44 48 49.5 40.523 43 40 6352 — 51.5 58.551 — 62 6842 — — 75.532 — — 7222.5 — — 3579 84 87 82.576 113.5 120 91.5
136 — 128 109224 — 100 148186 — — 94.5173.5 — — 11580 — — 5843 42 41.5 4433 24* 30 27.535 — 27 2637 — 23 2635 — — 24.534 — — 2630 — — 23
UI/l and quantities of albumin in serum expressed in g/l. Statistical test:
planta
up 5
6.57.528
5593
2402.5
also synthesize Factor VIII.

Surgery Molleví et al 779Volume 139, Number 6
Antithrombin. Control values were different be-tween rats and hamsters. The 3 allotransplantgroups showed slightly greater values than the con-trol rats, without reaching statistical significance. Incontrast, the 3 xenotransplant groups had signifi-cantly lesser levels than control rats (Table III) butthere were no differences compared to controlhamsters.
Free protein S. Although there were differencesbetween rats and hamsters, the xenotransplantgroups had values more similar to those of thecontrol rats. In the allotransplant groups at 7 and30 days, (groups 4 � 5), free protein S values weresimilar to those in Lewis rats, but were much higherat 100 days post-transplant. We therefore measuredthe levels of free protein S in the plasma of the BNrats, the donor strain in the allotransplant group at100 days, and found higher values than in the Lewis
Fig 1. A, Liver allograft (BN-to-Lew strain combination)at 100 days post-transplantation (H-E �100) showingmarked cholangiolar proliferation. B, Liver xenograft at100 days post-transplantation (hematoxylin-eosin, �100)showing a well-preserved hepatic architecture and ab-sence of lymphocyte infiltration.
rats (Table III).
Protein C. This protein was not detected in con-trol rat plasma, but hamsters had values of 22.1IU/dl. The allotransplanted rats, like donor rats, didnot have measurable values of plasma protein C.However, at 7 days post-xenotransplant, the xeno-transplanted rats began to present protein C; atdays 30 and 100 post-xenotransplant the valueswere practically identical to those of the hamstercontrols.
TAT complexes. The evolution of thrombin-an-tithrombin (TAT) complexes was similar to that ofplasma protein S. The allotransplant groups at 7and 30 days post-transplantation had no significantdifferences with respect to control Lewis rats, butby 100 days the differences had become signifi-cantly different. The results in the BN rats sug-gested that rather than an activation of coagula-tion, the allotransplant group had values similar tothose of the donor strain. In the xenotransplantgroups a substantial activation of coagulation wasobserved, especially 7 days post-transplantation(Table III). This activation of coagulation correctsitself with time, reaching values of 0.42 �g/dl, sub-stantially lower, though still statistically differentfrom control hamsters but not different from con-trol rats.
DISCUSSIONProgress is being made in overcoming the im-
munologic barriers to xenotransplantation, butslowly. However, there are other natural barriers toovercome before xenotransplant becomes a clinicalreality. Recently, Ramirez and colleagues demon-strated that baboons grafted with hDAF transgenicporcine livers maintained sufficient coagulation upto day 8 post-xenotransplantation.14 In hDAFgrafts, the clotting factors were replaced by thenewly synthesized proteins produced by the por-cine liver, and reached normal levels. In the ham-ster-to-rat liver xenotransplantation model it hasbeen established that the synthesis of proteins bythe donor liver is determined by its genetic config-uration. The recipient, thus, acquires progressivelythe protein profile of the donor species.15 Despitethe fact that the liver coordinates many metabolicprocesses and has been studied extensively fromthe physiologic standpoint, very few studies haveanalyzed the consequences of liver xenotransplantin long-term survivors.
The liver tests indicated that the graft functionwas adequate. Importantly the histology of bothallografts and xenografts reflected the absence ofrejection in all the study groups. The only charac-teristic histologic feature of the allotransplant
groups was a periportal cholangiolar proliferation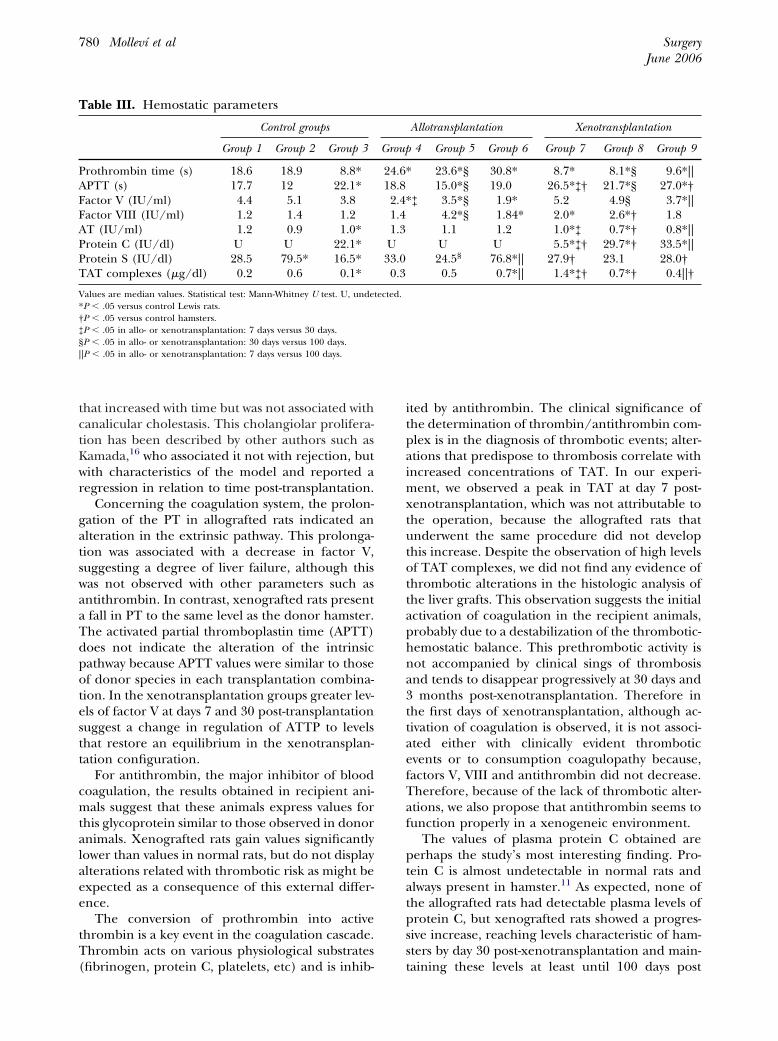
780 Molleví et al SurgeryJune 2006
that increased with time but was not associated withcanalicular cholestasis. This cholangiolar prolifera-tion has been described by other authors such asKamada,16 who associated it not with rejection, butwith characteristics of the model and reported aregression in relation to time post-transplantation.
Concerning the coagulation system, the prolon-gation of the PT in allografted rats indicated analteration in the extrinsic pathway. This prolonga-tion was associated with a decrease in factor V,suggesting a degree of liver failure, although thiswas not observed with other parameters such asantithrombin. In contrast, xenografted rats presenta fall in PT to the same level as the donor hamster.The activated partial thromboplastin time (APTT)does not indicate the alteration of the intrinsicpathway because APTT values were similar to thoseof donor species in each transplantation combina-tion. In the xenotransplantation groups greater lev-els of factor V at days 7 and 30 post-transplantationsuggest a change in regulation of ATTP to levelsthat restore an equilibrium in the xenotransplan-tation configuration.
For antithrombin, the major inhibitor of bloodcoagulation, the results obtained in recipient ani-mals suggest that these animals express values forthis glycoprotein similar to those observed in donoranimals. Xenografted rats gain values significantlylower than values in normal rats, but do not displayalterations related with thrombotic risk as might beexpected as a consequence of this external differ-ence.
The conversion of prothrombin into activethrombin is a key event in the coagulation cascade.Thrombin acts on various physiological substrates
Table III. Hemostatic parameters
Control groups
Group 1 Group 2 Group 3
Prothrombin time (s) 18.6 18.9 8.8*APTT (s) 17.7 12 22.1*Factor V (IU/ml) 4.4 5.1 3.8Factor VIII (IU/ml) 1.2 1.4 1.2AT (IU/ml) 1.2 0.9 1.0*Protein C (IU/dl) U U 22.1*Protein S (IU/dl) 28.5 79.5* 16.5*TAT complexes (�g/dl) 0.2 0.6 0.1*
Values are median values. Statistical test: Mann-Whitney U test. U, unde*P � .05 versus control Lewis rats.†P � .05 versus control hamsters.‡P � .05 in allo- or xenotransplantation: 7 days versus 30 days.§P � .05 in allo- or xenotransplantation: 30 days versus 100 days.||P � .05 in allo- or xenotransplantation: 7 days versus 100 days.
(fibrinogen, protein C, platelets, etc) and is inhib-
ited by antithrombin. The clinical significance ofthe determination of thrombin/antithrombin com-plex is in the diagnosis of thrombotic events; alter-ations that predispose to thrombosis correlate withincreased concentrations of TAT. In our experi-ment, we observed a peak in TAT at day 7 post-xenotransplantation, which was not attributable tothe operation, because the allografted rats thatunderwent the same procedure did not developthis increase. Despite the observation of high levelsof TAT complexes, we did not find any evidence ofthrombotic alterations in the histologic analysis ofthe liver grafts. This observation suggests the initialactivation of coagulation in the recipient animals,probably due to a destabilization of the thrombotic-hemostatic balance. This prethrombotic activity isnot accompanied by clinical sings of thrombosisand tends to disappear progressively at 30 days and3 months post-xenotransplantation. Therefore inthe first days of xenotransplantation, although ac-tivation of coagulation is observed, it is not associ-ated either with clinically evident thromboticevents or to consumption coagulopathy because,factors V, VIII and antithrombin did not decrease.Therefore, because of the lack of thrombotic alter-ations, we also propose that antithrombin seems tofunction properly in a xenogeneic environment.
The values of plasma protein C obtained areperhaps the study’s most interesting finding. Pro-tein C is almost undetectable in normal rats andalways present in hamster.11 As expected, none ofthe allografted rats had detectable plasma levels ofprotein C, but xenografted rats showed a progres-sive increase, reaching levels characteristic of ham-sters by day 30 post-xenotransplantation and main-
Allotransplantation Xenotransplantation
4 Group 5 Group 6 Group 7 Group 8 Group 9
* 23.6*§ 30.8* 8.7* 8.1*§ 9.6*||15.0*§ 19.0 26.5*‡† 21.7*§ 27.0*†
*‡ 3.5*§ 1.9* 5.2 4.9§ 3.7*||4.2*§ 1.84* 2.0* 2.6*† 1.81.1 1.2 1.0*‡ 0.7*† 0.8*||U U 5.5*‡† 29.7*† 33.5*||24.5§ 76.8*|| 27.9† 23.1 28.0†0.5 0.7*|| 1.4*‡† 0.7*† 0.4||†
Group
24.618.82.41.41.3U33.00.3
tected.
taining these levels at least until 100 days post

Surgery Molleví et al 781Volume 139, Number 6
xenotransplant when the animals were killed (end-point of study).
Both the active and the inactive forms of proteinC bind to its recipient, EPCR (endothelial proteinC recipient). Protein S, a vitamin K-dependent co-factor of protein C, is also necessary for the activa-tion of this protein. The binding between free pro-tein S and activated protein C, which in turn isbound to its receptor, is necessary to inactivatefactor V and factor VIII.17 Although free protein Sis not synthesized exclusively in the liver,18 we be-lieve the liver to be the main source of this protein,because the amounts detected in recipients of a BNgraft were similar to the levels observed in normalBN rats. The presence of free protein S in rats inthe absence of protein C raises doubts about therole of protein S in this animal. In humans, proteinS not only exerts its antithrombotic activity as acofactor of activated protein C but also possesses adirect effect, reducing the activation of prothrom-bin, the non-protein C-dependent formation of theprothrombinase complex.19 Conceivably, this ef-fect may represent the role of protein S in the rat.
Although in humans activated protein C is nec-essary to control the coagulation cascade or throm-botic tendency, in normal rats this protein is notdetected. Therefore, we hypothesize that there maybe an alternative mechanism at work in rats, be-cause in neither normal nor allografted rats werethrombotic anomalies observed. Furthermore wedo not believe that protein C of hamster origininduces hemostatic alterations, although we do notyet understand the role of protein C in xenograftedrats.
In conclusion, the hamster-rat xenotransplantmodel induces a large-scale activation of thrombo-sis in the first days post-transplantation, which isnot associated with a consumption coagulopathy.This deregulation is probably due to a generalimbalance as a result of the replacement of all therat components of hepatic synthesis by hamsterproteins, but that does not cause further problemsbecause it is corrected with time. In addition, liverxenotransplant causes the recipient animal toadopt the hemostatic profile of the donor species.Finally, the absence of thrombotic or hemorrhagicsymptoms suggests that the change of hemostaticstatus takes place in equilibrium conditions.
REFERENCES1. Joist JH. Hemostatic abnormalities in liver disease. In: Col-
man RW, Hirsh J, Marder VJ, et al, editors. Hemostasis andthrombosis. Basic principles and clinical practice. 3rd ed.
Philadelphia: JB Lippincott; 1993. p. 906-20.2. Mammen EF. Coagulation defects in liver disease. Med ClinNorth Am 1994;78:545-54.
3. Celli S, Valdivia LA, Fung JJ, Kelly RH, Pan F, Tsugita M, etal. Metabolic changes in the recipient after successful liverxenotransplantation in the rat. Transplant Proc 1994;26:1207.
4. Mollevi DG, Jaurrieta E, Ribas Y, Hurtado I, Serrano T,Gomez N, et al. Liver xenotransplantation: changes in lipidand lipoprotein concentration after long-term graft survival.J Hepatol 2000;32:655-660.
5. Kopp CW, Siegel JB, Hancock WW, Anrather J, Winkler H,Geczy CL, et al. Effect of porcine endothelial tissue factorpathway inhibitor on human coagulation factors. Transplan-tation 1997;63:749-58.
6. Siegel JB, Grey ST, Lesnikoski BA, Kopp CW, Soares M,Schulte J, et al. Xenogeneic endothelial cells activate humanprothrombin. Transplantation 1997;64:888-96.
7. Robson SC, Young VK, Cook NS, Metternich R, Kasper-Konig W, Lesnikoski BA, et al. Thrombin inhibition in an exvivo model of porcine heart xenograft hyperacute rejection.Transplantation 1996;61:862-8.
8. Lawson JH, Daniels LJ, Platt JL. The evaluation of throm-bomodulin activity in porcine to human xenotransplanta-tion. Transplant Proc 1997; 29:884-5.
9. Jurd KM, Gibbs RV, Hunt BJ. Activation of human pro-thrombin by porcine aortic endothelial cells—a potentialbarrier to pig to human xenotransplantation. Blood CoagulFibrinolysis 1996;7:336-43.
10. Tomson FN, Wardrop KJ. Clinical chemistry and hematol-ogy. In: Van Hoosier GL Jr, McPherson CW, editors. Labo-ratory hamsters. 1st ed. Orlando: Academic Press; 1987. p.43-59.
11. Ringler DH, Dabich L. Hematology and clinical biochemis-try. In: Baker HJ, Lindsey JR, Weisbroth SH, editors. Thelaboratory rat. Vol. I. Biology and disease. 1st ed. Orlando:Academic Press; 1979. p. 105-21.
12. Valdivia LA, Lewis JH, Celli S, Bontempo FA, Fung JJ, Dem-etris AJ, et al. Hamster coagulation and serum proteins inrat recipients of hamster xenografts. Transplantation1993;56:489-90.
13. Kamada N, Calne RY. Orthotopic liver transplantation inthe rat: technique using cuff for portal vein anastomosis andbiliary drainage. Transplantation 1979;28:47-50.
14. Ramirez P, Chavez R, Majado M, Munitiz V, Munoz A,Hernandez Q, et al. Life-supporting human complementregulator decay accelerating factor transgenic pig liverxenograft maintains the metabolic function and coagula-tion in the nonhuman primate for up to 8 days. Transplan-tation 2000;70:989-98.
15. Celli S, Valdivia LA, Fung JJ, Kelly RH. Early recipient-donorswitch of the complement type after liver xenotransplanta-tion. Immunol Invest 1997;26:589-600.
16. Kamada N. The process of rejection and non-rejection his-tology and functional competence of rat livers after trans-plantation. In: Experimental liver transplantation. Boca Ra-ton: CRC Press; 1988.
17. Esmon CT. Role of coagulation inhibitors in inflammation.Thromb Haemost 2001;86:51-6.
18. Jamison CS, McDowell SA, Marlar RA, Degen SJ. Develop-mental expression of protein C and protein S in the rat.Thromb Res 1995;78:407-19.
19. Daly ME, Beauchamp NJ. Inherited protein S deficiency:
from genotype to phenotype. Haematologica 2003;88:363-7.
Related Documents











