Eur. J. Immunol. 2015. 00: 1–13 Innate immunity DOI: 10.1002/eji.201445440 1 Hematopoietic plakophilin-3 regulates acute tissue-specific and systemic inflammation in mice Tatyana Sklyarova 1,2 , Jolanda van Hengel 1,2,3 , Elien Van Wonterghem 1,2 , Claude Libert 1,2 , Frans van Roy 1,2 and Roosmarijn E. Vandenbroucke 1,2 1 Inflammation Research Center, VIB, Ghent, Belgium 2 Department of Biomedical Molecular Biology, Ghent University, Ghent, Belgium 3 Department of Basic Medical Sciences, Ghent University, Ghent, Belgium Plakophilin-3 (PKP3) is a member of the armadillo protein family, which is important in cell−cell contacts and signaling during development and tumorigenesis. In conventional facilities, PKP3-deficient mice (PKP3 −/− ) develop spontaneous dermatitis, indicating a pos- sible involvement of PKP3 in inflammatory responses. Here, we show that PKP3 deficiency sensitizes mice to irritant contact dermatitis induced by phorbol myristate acetate (PMA). This sensitization occurred in mice with PKP3 deficiency in the hematopoietic system (PKP3 −/−hem ), but not if the deficiency was specific to skin keratinocytes (PKP3 −/−ker ). In a model of dextran sulfate sodium induced colitis, ubiquitous PKP3 deletion, but not intestinal epithelial PKP3 deficiency (PKP3 −/−IEC ), impaired survival from disease. Inter- estingly, PKP3 −/−hem mice also displayed increased sensitivity to dextran sulfate sodium induced colitis. Finally, PKP3 −/− mice were more sensitive to the lethality of lipopolysac- charide (LPS) injection than wild-type (WT) mice, and this phenotype was associated with increased intestinal permeability. PKP3 −/−IEC mice did not reproduce the enhanced endo- toxin reactivity of PKP3 −/− mice, in contrast to PKP3 −/−hem mice. Finally, in vitro stim- ulation of WT neutrophils with LPS or PMA increased Pkp3 expression. In conclusion, our data highlight a novel role for hematopoietic PKP3 in the regulation of both locally and systemically induced immune responses. Nonetheless, further research is needed to unravel the underlying mechanism. Keywords: Acute inflammation Colitis Dermatitis Hematopoietic cell Intestine Plakophilin-3 Sepsis Skin Additional supporting information may be found in the online version of this article at the publisher’s web-site Introduction The interaction of the organism with its environment requires the presence of a specialized “danger sensing” system. This protec- tive system operates mostly at the surfaces between the outer and inner environments of the body, such as the skin, intestinal tract, and respiratory tract. From a classical point of view, the epithelial Correspondence: Mrs. Roosmarijn E. Vandenbroucke e-mail: [email protected] surfaces at these locations form a physical barrier that is imper- meable to most infectious agents and thereby provide a first line of defense. More recently, subtle interactions between microbes and these epithelial surfaces have been shown to determine local immune reactions and disease. The barriers consists of three parts: an extensive epithelial cell surface exposed to the environment, a basement membrane that, together with various cell-junction structures, provides an extra level of separation, and a highly reac- tive lymphoid tissue underneath composed of antigen-presenting cells [1–4]. C 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Immunol. 2015. 00: 1–13 Innate immunityDOI: 10.1002/eji.201445440 1
Hematopoietic plakophilin-3 regulates acutetissue-specific and systemic inflammation in mice
Tatyana Sklyarova1,2, Jolanda van Hengel1,2,3, Elien Van Wonterghem1,2,Claude Libert1,2, Frans van Roy1,2 and Roosmarijn E. Vandenbroucke1,2
1 Inflammation Research Center, VIB, Ghent, Belgium2 Department of Biomedical Molecular Biology, Ghent University, Ghent, Belgium3 Department of Basic Medical Sciences, Ghent University, Ghent, Belgium
Plakophilin-3 (PKP3) is a member of the armadillo protein family, which is important incell−cell contacts and signaling during development and tumorigenesis. In conventionalfacilities, PKP3-deficient mice (PKP3−/−) develop spontaneous dermatitis, indicating a pos-sible involvement of PKP3 in inflammatory responses. Here, we show that PKP3 deficiencysensitizes mice to irritant contact dermatitis induced by phorbol myristate acetate (PMA).This sensitization occurred in mice with PKP3 deficiency in the hematopoietic system(PKP3−/−hem), but not if the deficiency was specific to skin keratinocytes (PKP3−/−ker). Ina model of dextran sulfate sodium induced colitis, ubiquitous PKP3 deletion, but notintestinal epithelial PKP3 deficiency (PKP3−/−IEC), impaired survival from disease. Inter-estingly, PKP3−/−hem mice also displayed increased sensitivity to dextran sulfate sodiuminduced colitis. Finally, PKP3−/− mice were more sensitive to the lethality of lipopolysac-charide (LPS) injection than wild-type (WT) mice, and this phenotype was associated withincreased intestinal permeability. PKP3−/−IEC mice did not reproduce the enhanced endo-toxin reactivity of PKP3−/− mice, in contrast to PKP3−/−hem mice. Finally, in vitro stim-ulation of WT neutrophils with LPS or PMA increased Pkp3 expression. In conclusion,our data highlight a novel role for hematopoietic PKP3 in the regulation of both locallyand systemically induced immune responses. Nonetheless, further research is needed tounravel the underlying mechanism.
Keywords: Acute inflammation � Colitis � Dermatitis � Hematopoietic cell � Intestine �
Plakophilin-3 � Sepsis � Skin
� Additional supporting information may be found in the online version of this article at thepublisher’s web-site
Introduction
The interaction of the organism with its environment requires thepresence of a specialized “danger sensing” system. This protec-tive system operates mostly at the surfaces between the outer andinner environments of the body, such as the skin, intestinal tract,and respiratory tract. From a classical point of view, the epithelial
Correspondence: Mrs. Roosmarijn E. Vandenbrouckee-mail: [email protected]
surfaces at these locations form a physical barrier that is imper-meable to most infectious agents and thereby provide a first lineof defense. More recently, subtle interactions between microbesand these epithelial surfaces have been shown to determine localimmune reactions and disease. The barriers consists of three parts:an extensive epithelial cell surface exposed to the environment,a basement membrane that, together with various cell-junctionstructures, provides an extra level of separation, and a highly reac-tive lymphoid tissue underneath composed of antigen-presentingcells [1–4].
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
Several inflammatory conditions, such as skin inflammation,inflammatory bowel disease (IBD) and the systemic inflammatoryresponse syndrome (SIRS), are associated with disruption of therespective epithelial barriers. The skin is the largest organ of thebody and is continuously exposed to microorganisms, ultravio-let radiation, toxic agents, and mechanical stress. Skin inflamma-tion can be mimicked in the mouse by topical application of asingle dose of 12-O-tetradecanoylphorbol-13-acetate, also knownas phorbol-12-myristate-13-acetate (PMA). This induces an acuteinflammatory reaction called irritant contact dermatitis (ICD) con-sisting of erythema, edema, and polymorphonuclear leukocyte(PMN) infiltration [5]. Skin inflammation is accompanied by epi-dermal barrier disruption because tight junctions between ker-atinocytes are disrupted and cytokines induce apoptosis or necro-sis of keratinocytes [6]. Disruption of the epithelial barrier isalso a key feature of IBD [7, 8]. The intestinal barrier defectsactivate the underlying immune system, resulting in tissue dam-age. The dextran sulfate sodium (DSS) model of colitis correlateswell with human IBD and is deemed suitable for investigating itspathogenesis [9]. SIRS is a lethal inflammatory disease caused bytrauma, burns, infections, and other insults [10]. When SIRS isassociated with hypotension, it is known as sepsis or septic shock.Several mouse models of sepsis are known [10]. The endotox-emia model involves the injection of bacterial lipopolysaccharides(LPS), which binds to the pathogen-recognition receptor (PRR),Toll-like receptor 4 (TLR4), leading to activation of downstreamsignaling cascades such as the NF-κB and the MAP kinase (MAPK)pathways. The consequence is synthesis and secretion of proin-flammatory cytokines and costimulatory molecules. We and oth-ers have shown that LPS-induced systemic inflammation increasesintestinal permeability [11–13], which leads to leakage of gutflora, pathogen-associated molecular patterns (PAMPs), and/ordamage-associated molecular patterns (DAMPs) from the gut intothe peripheral blood and tissues, thereby accelerating systemicinflammation [11, 12, 14–16].
Invading organisms or tissue damage at epithelial barriers ini-tiate an inflammatory response mediated by surface-exposed orintracellular PRRs, expressed by both hematopoietic and non-hematopoietic cells. These PRRs detect PAMPs from the invad-ing organisms or DAMPs released from injured cells. Activation ofPRRs stimulates several signaling cascades that culminate in therelease of proinflammatory cytokines, chemokines, and mediatorsthat enhance vasodilation and neutrophil extravasation, as well asacquisition of plasma components such as complement and anti-bodies to destroy pathogens and damaged cells [17]. Additionally,phagocytes are stimulated to clear the noxious agents and furtheractivate inflammation in order to induce pathogen clearance andtissue repair, and restore immune homeostasis.
Plakophilin-3 (PKP3) is a member of the p120ctn subfamilyof armadillo proteins, which comprises adaptor proteins involvedin the regulation of cell−cell adhesion, morphogenesis, home-ostasis, and tumor development [18–20]. PKP3 is predominantlyexpressed in epithelial tissues and is an important integrator atcell–cell junctions as it contributes to correct maturation of bothdesmosomes and adherens junctions [18, 19, 21–24]. However,
it also has important nonjunctional functions, such as regulatingprotein synthesis, growth control, and transcription [18–20, 23].In the cytoplasm, PKP3 is a constituent of stress granules andother RNA-containing particles that influence the stability of spe-cific mRNAs and thus act as posttranscriptional regulators of geneexpression [25–27].
PKP3-deficient mice spontaneously develop chronic dermatitiswhen bred in conventional facilities but not in specific-pathogenicfree conditions [28]. In addition, two SNPs in the human PKP3gene are significantly associated with susceptibility to tuberculosis[29]. These findings indicate that PKP3 could play a role in theinflammatory response.
Here, we tested the hypothesis that PKP3 plays a role in orches-trating the inflammatory response in addition to its structuralfunction at desmosomes. We studied the effect of PKP3 defi-ciency in mouse models of PMA-induced ICD, DSS-induced col-itis, and endotoxemia. We demonstrate that ubiquitous as wellas hematopoietic loss of PKP3 instigates the development of ICDand DSS-induced colon inflammation. Additionally, endotoxemialethality was increased by ablation of PKP3 in the whole body orspecifically in hematopoietic cells.
Results
Total and hematopoietic PKP3 deficiency, not skinkeratinocyte deficiency, sensitize to PMA dermatitis
We previously reported the generation of PKP3−/− mice to studythe in vivo function of PKP3 [28]. We found that PKP3−/− micehoused in a conventional facility spontaneously develop dermatitisthat gradually leads to severe nonhealing ulceration [28]. Thisprompted us to investigate whether PKP3 modulates inflammatoryresponses.
In specific pathogen-free (SPF) conditions, PKP3−/− mice didnot develop spontaneous dermatitis. Additionally, comparison ofsystemic cytokine levels in PKP3+/+ and PKP3−/− mice revealedno difference in the inflammatory status of the mice (Support-ing Information Fig. 1). This enabled us to use an experimentalmodel of induced skin inflammation. In the model of ICD, a singleapplication of PMA elicits an acute inflammatory response by non-specific activation of protein kinase C and other PMA receptors;the result is inflammation manifested in immediate tissue ery-thema (redness) and edema (swelling) [30]. Auricles of PKP3+/+
and PKP3−/− mice were treated with PMA, skin thickness wasmeasured at different time points, and tissue samples were ana-lyzed histologically (Fig. 1). Our data revealed that PKP3−/− mice(right panels) react differently from PKP3+/+ mice (left panels).After PMA application, erythema and swelling were more pro-nounced in PKP3−/− mice than in PKP3+/+ mice (Fig. 1A). Auric-ular sections of PKP3+/+ and PKP3−/− mice were obtained 12 and24 h after PMA application, stained with hematoxylin and eosin(H&E), and examined histologically (Fig. 1B and C). These sec-tions, together with measurements of skin thickening (Fig. 1E),confirmed the macroscopically observed increased thickness of
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 3
Figure 1. PKP3 deficiency exacerbates erythema, tissue swelling, andepidermal hyperproliferation in a model of ICD. PMA was applied to theauricles of PKP3+/+ and PKP3−/− mice followed by analysis of erythema,tissue swelling and epidermal hyperproliferation. (A) Representativemacroscopic images of auricular erythema 45 min after PMA applica-tion on PKP3+/+ (left) and PKP3−/− (right) mice. The arrows point tothe PMA-treated left auricles. (B and C) Representative H&E-stainedsections of auricles from PKP3+/+ (left) and PKP3−/−mice (right) at(B) 12 h and (C) 24 h after PMA treatment respectively. The scale barsrepresent 50 μm. (D) Ki67 immunohistochemical staining of proliferat-ing keratinocytes 24 h after PMA application. The scale bars represent50 μm. (E) Time course of PMA-induced edema in PKP3+/+ (black bars; n= 4) and PKP3−/− (gray bars; n = 4) mice. Increase in auricular thicknesswas measured 2, 4, 9, and 24 h after PMA application. Data are shownas mean ± SEM; one of three representative experiments is shown.Statistical significance determined by unpaired t-test; *0.01 < p < 0.05.
PMA-treated PKP3−/− auricles compared to their wild-type (WT)counterparts. Skin thickening, the first hallmark of skin irritationand local inflammation, indicates enhanced vascular permeability,edema within the epidermis and dermis, and proliferation of epi-dermal keratinocytes. Indeed, Ki67 immunostaining revealed thatPMA-treated auricles of PKP3−/− mice show enhanced epidermalproliferation and hyperplasia 24 h after treatment in comparisonto PKP3+/+ littermates (Fig. 1D).
To investigate the cell type specific functions of PKP3, we gener-ated two different conditional knockout mice by crossing PKP3fl/fl
mice, generated in our lab [28], with mice containing Cre recombi-nase under control of either the hematopoietic-specific Vav1 pro-moter [21] or the basal epidermal layer specific keratin 5 (K5)promoter [32]. The resulting PKP3fl/fl × VavCre and PKP3fl/fl ×K5Cre mice, hereafter called PKP3−/−hem and PKP3−/−ker, wereviable and did not show signs of spontaneous skin inflammationunder SPF conditions. To determine if PKP3 deficiency affected theproduction of hematopoietic cells, we analyzed white blood cellsin peripheral blood from PKP3fl/fl and PKP3−/−hem mice using themouse Hemavet R© System. We observed no statistically significantdifferences in absolute and relative numbers of circulating neu-trophils, lymphocytes, monocytes, eosinophils, or basophils (datanot shown).
As shown in Figure 2, application of PMA to the auricles ofPKP3fl/fl and PKP3−/−ker mice resulted in mild edema and pre-dominantly dermal infiltration of granulocytes. In contrast, PMA-treated auricles of PKP3−/−hem mice developed edema earlier andit was significantly more pronounced (Fig. 2A). Moreover, exten-sive hemorrhage (arrowheads in Fig. 2C and D) and intraepithe-lial edema were evident in PKP3−/−hem but not in PKP3fl/fl andPKP3−/−ker auricular sections (Fig. 2B–D). Additionally, within 4h after challenge, PKP3−/−hem mice developed epidermal necrosismanifested in keratinocyte ballooning (Fig. 2C; arrows). Eventu-ally, this response developed into severe neutrophilic pustulosis24 h after PMA application (Fig. 2D; arrows).
In acute reactions, mast cells and neutrophils are the firstresponders to harmful stimuli [33]. Two hours after PMA treat-ment, mast cells numbers (counted in skin sections stained withtoluidine blue) were significantly higher in PKP3−/−hem than inPKP3fl/fl mice (Supporting Information Fig. 2A and B). Next, gran-ulocytes with characteristic polysegmented nuclear morphologywere counted in H&E-stained sections. This revealed significantlymore granulocytes in the dermis and epidermis of PKP3−/−hem
mice than in PKP3fl/fl and PKP3−/−ker mice (Fig. 2E and F).Consistent with these observations, skin from PKP3−/− mice
(Fig. 3A) and PKP3−/−hem mice (Fig. 3B) showed elevated expres-sion of mRNA of the proinflammatory cytokines and chemokinesIl6, Il1β, Tnf, Ifnγ, Il17, iNos, and Kc compared to their WT coun-terparts, while the levels of the macrophage chemokine attractant,Rantes (Ccl5), did not differ. These results indicate that deficiencyof hematopoietic PKP3 enhances PMA-induced skin inflammation,reflected by increased focal necrosis, ulceration of the epidermis,and increased neutrophil influx that is associated with skin irrita-tion responses in vivo.
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

4 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
Figure 2. Expression of PKP3 in the hematopoietic compartment protects skin from exaggerated irritant response. PMA was applied to ears ofPKP3fl/fl, PKP3−/−hem, and PKP3−/−ker mice as in Figure 1, and edema and histology were studied. (A) Time course of PMA-induced auricular edema inPKP3fl/fl mice (black bars; n = 4), PKP3−/−hem mice (dark gray bars; n = 4), and PKP3−/−ker mice (light gray bars; n = 4). Increase in auricular thicknesswas measured 2, 4, 9, and 24 h after PMA application. (B–D) Auricular skin histology (H&E stain) of PKP3fl/fl (left), PKP3−/−hem (middle), and PKP3−/−ker
(right) mice. (B) Auricles before PMA application, (C) 4 h after application, and (D) 24 h after application. Arrows point to (C) necrotic keratinocytesand (D) neutrophil-filled pustules; arrowheads (C and D) point to hemorrhage. The scale bars represent 50 μm. (E and F) Auricles from PKP3+/+
(black bars; n = 4) and PKP3−/− (gray bars; n = 4) mice were stained with H&E 24 h after PMA challenge. (E) Neutrophil counts per square millimeterwere enumerated and (F) a representative staining is shown. The scale bars represent 50 μm. Similar results were obtained when neutrophils werequantified in six auricles from two independent experiments. (A and E) Data are shown as mean ± SEM from single experiments representative oftwo performed. Data were analyzed by an unpaired t-test; *0.01 � p < 0.05; ****p < 0.0005.
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 5
Figure 3. PKP3 deficiency upregulates markers of immune activation inPMA-treated skin. PMA was applied to auricles of PKP3+/+ and PKP3−/−
mice as in Figure 1, followed by cytokine analysis. (A) PKP3+/+ (blackbars; n = 4−5) and PKP3−/− (gray bars; n = 3) mice were subjected to PMAtreatment and auricles were examined 24 h after application. Tran-script expression analysis of the indicated cytokines and chemokineswas determined by real-time qPCR. (B) Auricles of PKP3fl/fl (black bars; n= 3−4) and PKP3−/−hem (gray bars; n = 3−4) mice were subjected to PMAtreatment and examined 24 h after application. Transcript expressionanalysis of the indicated cytokines and chemokines was done by real-time qPCR. (A and B) Data are shown as mean ± SEM, and are from asingle experiment representative of two performed experiments. Datawere analyzed by an unpaired t-test; *0.01 � p < 0.05; ns, not signifi-cantly different.
Interestingly, PKP3 gene expression appears to be altered invarious human skin inflammatory conditions. According to thetoolbox Genevestigator, a database of Affymetrix microarray-based studies [34], two independent microarray experimentsrevealed that PKP3 gene expression in psoriatic skin lesions isupregulated up to 2.5-fold relative to healthy tissue (SupportingInformation Fig. 3A) [35, 36]. Additionally, PKP3 mRNA was sig-nificantly upregulated 1.7- and 1.35-fold relative to normal skin inlesional and nonlesional atopic dermatitis skin, respectively [37](Supporting Information Fig. 3B). These data suggest that PKP3upregulation in lesional skin might be a mechanism for protectingthe skin from excessive inflammation.
Hematopoietic but not intestinal epithelial PKP3restrains inflammation in DSS-induced colitis
Having found that PKP3 expression in leukocytes restrains skininflammation, we studied colon-specific inflammation. In basalconditions, PKP3+/+ and PKP3−/− mouse colon tissues did notshow any differences in morphology or in the number and pro-liferation/apoptosis rates of the different intestinal cell types(Supporting Information Fig. 4 and data not shown). To study
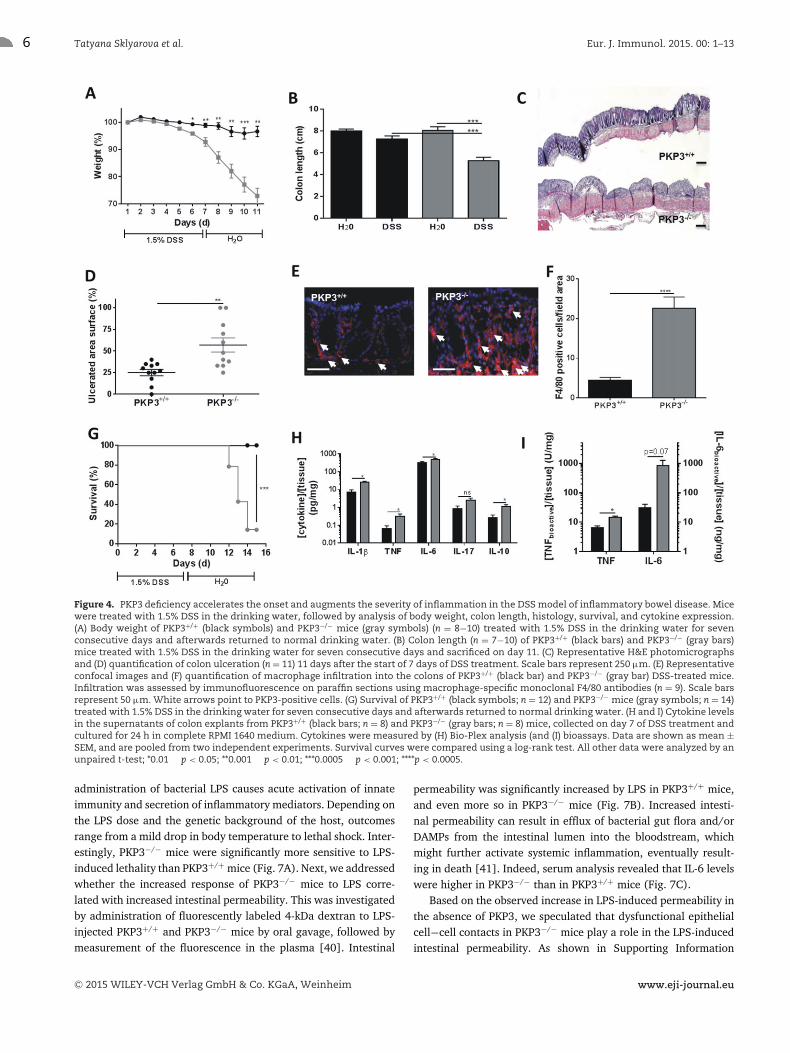
mouse colon inflammation, we induced colitis by oral administra-tion of DSS. Acute colitis induced by DSS administration in thedrinking water is characterized by mucosal ulceration and submu-cosal inflammation provoked by disruption of the epithelial barrierand subsequent translocation of luminal microbiota into the lam-ina propria [38]. In both PKP3+/+ and PKP3−/− mice, ingestion of1.5% DSS for 7 days induced symptoms of acute intestinal inflam-mation, such as weight loss, diarrhea, and rectal bleeding. How-ever, PKP3−/− mice were affected more by DSS in terms of weightloss (Fig. 4A), colon shortening (Fig. 4B), mucosal ulceration(Fig. 4C and D), and macrophage infiltration (Fig. 4E and F). Toevaluate the recovery from colitis, mice were returned to normaldrinking water after 7 days of DSS treatment. WT mice recoveredfully, but most of PKP3−/− mice did not recover from the colitisand had to be euthanized due to severe morbidity (Fig. 4G).
Accordingly, analysis of supernatant from colonic explants ofDSS-treated PKP3−/− mice revealed higher levels of TNF, IL-1β,IL-6, and IL-10 than those from the DSS-treated PKP3+/+ mice(Fig. 4H and I). Clearly, PKP3 deficiency leads to loss of thecontrol of proinflammatory responses, which leads to enhancedprogression and maintenance of the chemically induced intestinalinflammation.
To investigate cell type specific functions of PKP3 in coloninflammation, we generated another mutant mouse line by cross-ing PKP3fl/fl mice with mice expressing Cre recombinase underthe control of the intestinal-specific Villin promoter [39]. SuchPKP3−/−IEC mice were born at the expected Mendelian ratio, devel-oped normally, and did not show any gross morphological changesin the overall histology of small and large intestines (data notshown). To determine whether the results obtained using PKP3−/−
mice with total PKP3 deficiency could be reproduced in PKP3−/−IEC
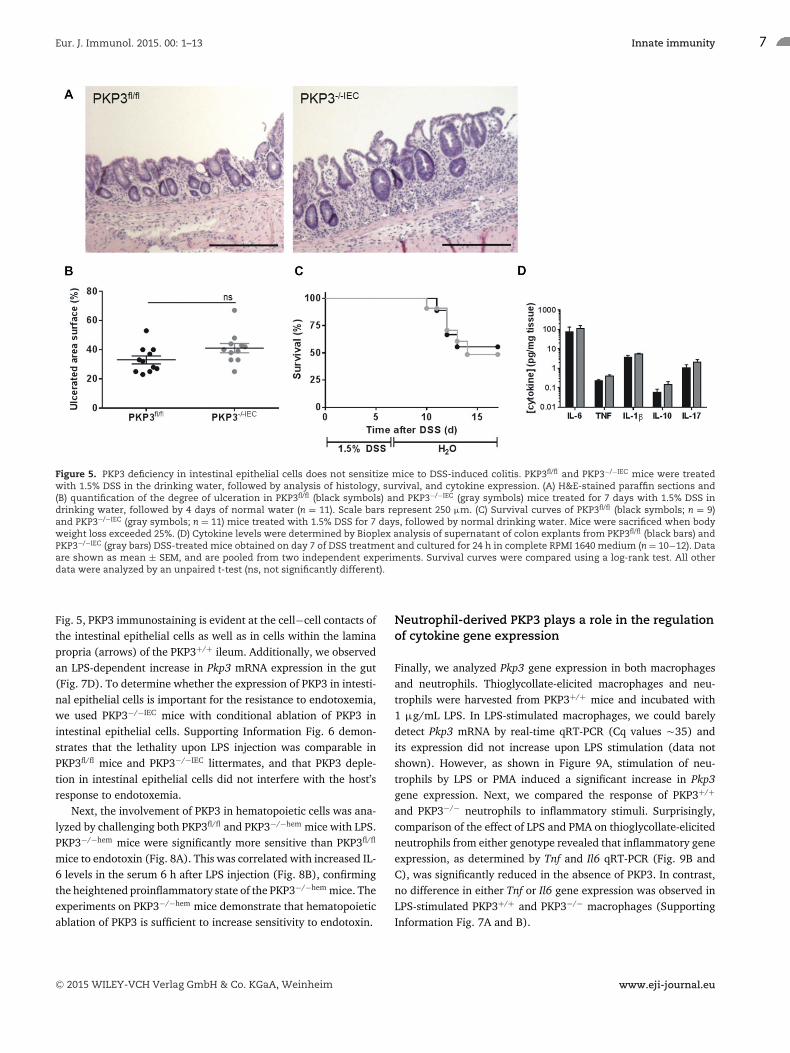
mice, the latter as well as control PKP3fl/fl littermates were treatedwith 1.5% DSS for 7 days. Surprisingly, the elimination of PKP3from intestinal epithelial cells did not result in significant changesin DSS-induced colon ulceration (Fig. 5A and B), mouse survival(Fig. 5C), or cytokine production by colon explants (Fig. 5D) rel-ative to PKP3fl/fl mice. However, colitis onset, severity, and pro-gression in PKP3−/−hem mice that ingested 2% DSS for 7 days wasaggravated compared to PKP3fl/fl mice (Fig. 6). Acute symptomsin PKP3−/−hem mice appeared earlier and were more severe, asreflected by increased weight loss and total clinical score (Fig. 6Aand B). Histologically, the increased disease activity in PKP3−/−hem
mice correlated with more colonic shortening (Fig. 6C) and withextensive epithelial ulceration (Fig. 6D). These results indicatethat absence of hematopoietic PKP3 worsens DSS-driven colonicinflammation and that PKP3 plays a protective role against colitisin WT animals.
PKP3−/− and PKP3−/−hem mice, but not PKP3−/−IEC mice,have increased sensitivity to endotoxemia
To examine the potential involvement of PKP3 in SIRS, endotox-emia was induced by intraperitoneal injection of LPS in PKP3+/+
and PKP3−/− mice. In this model of sterile septic shock, systemic
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
6 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
Figure 4. PKP3 deficiency accelerates the onset and augments the severity of inflammation in the DSS model of inflammatory bowel disease. Micewere treated with 1.5% DSS in the drinking water, followed by analysis of body weight, colon length, histology, survival, and cytokine expression.(A) Body weight of PKP3+/+ (black symbols) and PKP3−/− mice (gray symbols) (n = 8−10) treated with 1.5% DSS in the drinking water for sevenconsecutive days and afterwards returned to normal drinking water. (B) Colon length (n = 7−10) of PKP3+/+ (black bars) and PKP3−/− (gray bars)mice treated with 1.5% DSS in the drinking water for seven consecutive days and sacrificed on day 11. (C) Representative H&E photomicrographsand (D) quantification of colon ulceration (n = 11) 11 days after the start of 7 days of DSS treatment. Scale bars represent 250 μm. (E) Representativeconfocal images and (F) quantification of macrophage infiltration into the colons of PKP3+/+ (black bar) and PKP3−/− (gray bar) DSS-treated mice.Infiltration was assessed by immunofluorescence on paraffin sections using macrophage-specific monoclonal F4/80 antibodies (n = 9). Scale barsrepresent 50 μm. White arrows point to PKP3-positive cells. (G) Survival of PKP3+/+ (black symbols; n = 12) and PKP3−/− mice (gray symbols; n = 14)treated with 1.5% DSS in the drinking water for seven consecutive days and afterwards returned to normal drinking water. (H and I) Cytokine levelsin the supernatants of colon explants from PKP3+/+ (black bars; n = 8) and PKP3−/− (gray bars; n = 8) mice, collected on day 7 of DSS treatment andcultured for 24 h in complete RPMI 1640 medium. Cytokines were measured by (H) Bio-Plex analysis (and (I) bioassays. Data are shown as mean ±SEM, and are pooled from two independent experiments. Survival curves were compared using a log-rank test. All other data were analyzed by anunpaired t-test; *0.01 � p < 0.05; **0.001 � p < 0.01; ***0.0005 � p < 0.001; ****p < 0.0005.
administration of bacterial LPS causes acute activation of innateimmunity and secretion of inflammatory mediators. Depending onthe LPS dose and the genetic background of the host, outcomesrange from a mild drop in body temperature to lethal shock. Inter-estingly, PKP3−/− mice were significantly more sensitive to LPS-induced lethality than PKP3+/+ mice (Fig. 7A). Next, we addressedwhether the increased response of PKP3−/− mice to LPS corre-lated with increased intestinal permeability. This was investigatedby administration of fluorescently labeled 4-kDa dextran to LPS-injected PKP3+/+ and PKP3−/− mice by oral gavage, followed bymeasurement of the fluorescence in the plasma [40]. Intestinal
permeability was significantly increased by LPS in PKP3+/+ mice,and even more so in PKP3−/− mice (Fig. 7B). Increased intesti-nal permeability can result in efflux of bacterial gut flora and/orDAMPs from the intestinal lumen into the bloodstream, whichmight further activate systemic inflammation, eventually result-ing in death [41]. Indeed, serum analysis revealed that IL-6 levelswere higher in PKP3−/− than in PKP3+/+ mice (Fig. 7C).
Based on the observed increase in LPS-induced permeability inthe absence of PKP3, we speculated that dysfunctional epithelialcell−cell contacts in PKP3−/− mice play a role in the LPS-inducedintestinal permeability. As shown in Supporting Information
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 7
Figure 5. PKP3 deficiency in intestinal epithelial cells does not sensitize mice to DSS-induced colitis. PKP3fl/fl and PKP3−/−IEC mice were treatedwith 1.5% DSS in the drinking water, followed by analysis of histology, survival, and cytokine expression. (A) H&E-stained paraffin sections and(B) quantification of the degree of ulceration in PKP3fl/fl (black symbols) and PKP3−/−IEC (gray symbols) mice treated for 7 days with 1.5% DSS indrinking water, followed by 4 days of normal water (n = 11). Scale bars represent 250 μm. (C) Survival curves of PKP3fl/fl (black symbols; n = 9)and PKP3−/−IEC (gray symbols; n = 11) mice treated with 1.5% DSS for 7 days, followed by normal drinking water. Mice were sacrificed when bodyweight loss exceeded 25%. (D) Cytokine levels were determined by Bioplex analysis of supernatant of colon explants from PKP3fl/fl (black bars) andPKP3−/−IEC (gray bars) DSS-treated mice obtained on day 7 of DSS treatment and cultured for 24 h in complete RPMI 1640 medium (n = 10−12). Dataare shown as mean ± SEM, and are pooled from two independent experiments. Survival curves were compared using a log-rank test. All otherdata were analyzed by an unpaired t-test (ns, not significantly different).
Fig. 5, PKP3 immunostaining is evident at the cell−cell contacts ofthe intestinal epithelial cells as well as in cells within the laminapropria (arrows) of the PKP3+/+ ileum. Additionally, we observedan LPS-dependent increase in Pkp3 mRNA expression in the gut(Fig. 7D). To determine whether the expression of PKP3 in intesti-nal epithelial cells is important for the resistance to endotoxemia,we used PKP3−/−IEC mice with conditional ablation of PKP3 inintestinal epithelial cells. Supporting Information Fig. 6 demon-strates that the lethality upon LPS injection was comparable inPKP3fl/fl mice and PKP3−/−IEC littermates, and that PKP3 deple-tion in intestinal epithelial cells did not interfere with the host’sresponse to endotoxemia.
Next, the involvement of PKP3 in hematopoietic cells was ana-lyzed by challenging both PKP3fl/fl and PKP3−/−hem mice with LPS.PKP3−/−hem mice were significantly more sensitive than PKP3fl/fl
mice to endotoxin (Fig. 8A). This was correlated with increased IL-6 levels in the serum 6 h after LPS injection (Fig. 8B), confirmingthe heightened proinflammatory state of the PKP3−/−hem mice. Theexperiments on PKP3−/−hem mice demonstrate that hematopoieticablation of PKP3 is sufficient to increase sensitivity to endotoxin.
Neutrophil-derived PKP3 plays a role in the regulationof cytokine gene expression
Finally, we analyzed Pkp3 gene expression in both macrophagesand neutrophils. Thioglycollate-elicited macrophages and neu-trophils were harvested from PKP3+/+ mice and incubated with1 μg/mL LPS. In LPS-stimulated macrophages, we could barelydetect Pkp3 mRNA by real-time qRT-PCR (Cq values �35) andits expression did not increase upon LPS stimulation (data notshown). However, as shown in Figure 9A, stimulation of neu-trophils by LPS or PMA induced a significant increase in Pkp3gene expression. Next, we compared the response of PKP3+/+
and PKP3−/− neutrophils to inflammatory stimuli. Surprisingly,comparison of the effect of LPS and PMA on thioglycollate-elicitedneutrophils from either genotype revealed that inflammatory geneexpression, as determined by Tnf and Il6 qRT-PCR (Fig. 9B andC), was significantly reduced in the absence of PKP3. In contrast,no difference in either Tnf or Il6 gene expression was observed inLPS-stimulated PKP3+/+ and PKP3−/− macrophages (SupportingInformation Fig. 7A and B).
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

8 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
Figure 6. Ablation of PKP3 in hematopoietic cells exacerbates DSS-induced colitis. PKP3fl/fl and PKP3−/−hem mice were treated with 2% DSS in thedrinking water, followed by analysis of clinical and histological parameters. (A) Weight loss, (B) clinical score, (C) colon length on day 7, and (D)quantification of ulceration in histological sections of the colon for PKP3fl/fl (black symbols or bars) and PKP3−/−hem (gray symbols or bars) micetreated with 2% DSS in drinking water for 7 days (n = 8). Data are representative of two independent experiments. Data are shown as mean ± SEM,and are from a single experiment. All data were analyzed by an unpaired t-test; *0.01 � p < 0.05; ***0.0005 � p < 0.001.
Discussion
We identified a novel role for PKP3 in molecular processes associ-ated with skin and gut inflammation, mimicked by PMA-inducedskin inflammation, DSS-induced colitis and LPS-induced septicshock, called endotoxemia. In all three inflammatory mouse mod-els, PKP3−/− and PKP3−/−hem mice displayed increased sensitiv-ity compared to their PKP3 expressing counterparts. Surprisingly,PKP3−/−ker and PKP3−/−IEC mice with PKP3 deficiencies in epithe-lial cells of either skin or intestine did not display increasedsensitivity to, respectively, skin and colon inflammation. Thisimplies that in our in vivo models the disruption of eitherdesmosomes or adherens junctions, as observed recently inin vitro skin models with PKP3 knockdown [24] is insufficientlysevere to play a significant role in the observed inflammationphenotypes. As observed previously [28], this might be dueto compensatory upregulations of PKP1 and PKP2 in epithe-lial cells, whereas this might not be the case in hematopoieticcells.
Our in vitro analysis further showed that PKP3 is upregulatedin stimulated neutrophils, but not in macrophages, and plays arole in transcriptional activation or RNA stabilization of proin-flammatory cytokines. Although not previously linked to proin-flammatory mRNAs, similar activation mechanisms have beendescribed for members of the plakophilin protein family. Indeed,
PKP3 was shown to activate transcription via interaction with tran-scription factor ETV1 [20], PKP1 and PKP3 can stabilize mRNAwhen they are present in mRNA ribonucleoprotein particles [27],and PKP1 interacts with the eukaryotic translation initiation factor4A1 (eIF4A1), thereby stimulating translation [42]. The observedPKP3-dependent transcriptional activation or RNA stabilization ofproinflammatory cytokines in isolated neutrophils is seemingly incontrast to the increased systemic cytokine levels and sensitivityof the PKP3−/− and PKP3−/−hem mice in the different inflamma-tory mouse models. However, epithelial cell layers such as skinand gut epithelium are heavily colonized with bacteria. In caseof an inflammatory trigger, PKP3 upregulation might protect theepithelial barrier from excessive damage by playing a role ininflammatory cell activation and local cytokine production, pre-venting bacteria to enter the body and to induce a further increasein inflammation. Consequently, reduction in Tnf and/or Il6 mRNAtranscription or stabilization in the absence of PKP3 might sen-sitize the mice to skin and gut epithelial damage and inflamma-tion, as was observed in PMA-induced ICD, DSS-induced colitis,and endotoxemia. Interestingly, PKP3 was recently compared tothe protein HuR [27], an RNA-binding protein that affects targetmRNA stability and translation of, for example, Tnf mRNA [43].Moreover, myeloid HuR-deficient mice displayed enhanced sensi-tivity to endotoxemia and rapid progression of chemical-inducedcolitis [44].
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 9
Figure 7. Ubiquitous deletion of PKP3 sensitizes mice to LPS toxicity. PKP3+/+ and PKP3−/− mice were injected i.p. with LPS, followed by monitoringof survival, intestinal permeability and IL-6 serum levels. Additionally, PKP3+/+ ileal lysates were collected to analyze Pkp3 mRNA levels upon LPSstimulation. (A) Kaplan−Meier survival graph of PKP3+/+ (black symbols; n = 10) and PKP3−/− mice (gray symbols; n = 18) injected i.p. with LPS. (B)Intestinal permeability in basal conditions (n = 3) and 8 h after LPS injection (n = 7). PKP3+/+ (black bars) and PKP3−/− mice (gray bars) mice weregavaged with FITC-dextran solution and serum concentration was determined without treatment and 8 h after LPS challenge. (C) IL-6 serum levels1.5 h after LPS injection in PKP3+/+ (black bar; n = 7) and PKP3−/− mice (gray bar; n = 3). (D) Pkp3 mRNA expression in ileum lysates of WT micebefore and at different times after LPS injection (n = 4−5 for each time point). Data are shown as mean ± SEM. The survival graph is the resultof two pooled independent experiments. Intestinal permeability, IL-6 cytokine results and Pkp3 expression analysis are from a single experiment.Survival curves were compared using a log-rank test. All other data were analyzed by an unpaired t-test; *0.01 � p < 0.05; **0.001 � p < 0.01; ****p <
0.0005.
Clearly, further research is needed to unravel the underlyingmechanisms of the PKP3-ablation associated phenotypes in thethree different inflammatory models. Amongst others, the mech-anism by which PKP3 controls cytokine levels is still unclear. Incase of the skin phenotype, it would be interesting to identify therelevant cytokines that mediate the PMA-induced mast cell accu-mulation in the skin, and whether neutrophils are responsiblefor the production of these cytokines. We did not study in detailwhich hematopoietic cell type is responsible for the observed phe-notype in the colitis model, but neutrophilic PKP3 might play arole. Indeed, several reports have demonstrated that neutrophildepletion exacerbates inflammation in rat and mouse models ofcolitis [45]. Also the impact of hematopoietic PKP3 on epithelialbarrier integrity during endotoxemia needs further investigation.Our data suggest that PKP3 is essential in cytokine production toprevent bacteria to enter the body. Consequently, absence of PKP3might result in amplification of local intestinal inflammation andconsequent barrier dysfunction.
In conclusion, our study shows that PKP3 plays a protectiverole in both local and systemic inflammatory diseases, as shownby the increased sensitivity of PKP3−/− mice to PMA-induced ICD,DSS-induced colitis, and endotoxemia. Surprisingly, not epithelial
but hematopoietic derived PKP3 seems to play a central role inthe observed phenotypes. This is the first time that a prominentbiological function is attributed to a so-called desmosomal proteinexpressed in hematopoietic cells. Further research is needed tounravel the detailed mechanism by which hematopoietic PKP3induces an increase in inflammatory gene expression in vitro anda strikingly increased resistance to inflammatory triggers in vivo.
Materials and methods
Animals
PKP3fl/fl mice and derivative PKP3-deficient mice (PKP3−/−) weregenerated by gene targeting as described previously [28]. In thesemice, Pkp3 exons 2–4 were loxP-flanked, and their removal byCre action also generated a frame shift in the encoded mRNA.After generation, these mice were backcrossed 8 and 10 timesto C57/BL6 background, respectively. PKP3−/−hem, PKP3−/−ker,and PKP3−/−IEC mice were generated by crossing PKP3fl/fl micewith mice expressing Cre recombinase under the control of the
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

10 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
Figure 8. Hematopoietic deletion of PKP3 sensitizes mice to LPS toxicity. PKP3−/−hem and corresponding negative-control mice (PKP3fl/fl) wereinjected i.p. with LPS; survival was monitored and IL-6 serum levels were analyzed. (A) Survival of PKP3fl/fl (black symbols; n = 24) and PKP3−/−hem
mice (gray symbols; n = 20) in the endotoxemia model. Mice were injected i.p. with LPS, monitored for survival, and sacrificed when moribund.(B) IL-6 serum levels in PKP3fl/fl (black bars; n = 7) and PKP3−/−hem mice (gray bars; n = 6) 1.5 and 6 h after LPS injection. Data are shown as mean ±SEM, and are pooled from three independent experiments for generation of the survival curve, while a single experiment was done to determineIL-6 serum levels. Survival curves were compared using a log-rank test. The IL-6 data were analyzed by an unpaired t-test; *0.01 � p < 0.05; **0.001� p < 0.01.
Figure 9. Gene expression analysis in thioglycollate-elicited neutrophils stimulated with LPS and PMA. Primary neutrophils in culture werestimulated in vitro by LPS and PMA, followed by gene expression analysis. (A) Analysis of Pkp3 gene expression in thioglycollate-elicited untreatedprimary neutrophils or after incubation with LPS or PMA (n = 4). (B and C) Analysis of Il6 (B) and Tnf (C) gene expression in PKP3+/+ (black bars;n = 4) and PKP3−/− (gray bars; n = 3) neutrophils stimulated with LPS or PMA. Data are shown as mean ± SEM, and are representative for twoindependent experiments. All data were analyzed by an unpaired t-test; *0.01 � p < 0.05; ***0.0005 � p < 0.001.
hematopoietic-specific Vav, the basal epidermal layer specific ker-atin 5 (K5), or the intestinal epithelium specific Villin promoter[31, 32, 39]. The resultant PKP3fl/fl-VavCretg/+ mice were on amixed C57/BL6/Swiss background, PKP3fl/fl-K5Cretg/+ mice wereon a mixed 129P2/OlaHsd/C57/BL6/CBA/SJL background, andPKP3fl/fl/VillinCretg/+ mice were on a pure C57/BL6 background.All mice were housed in a SPF animal facility with ad libitum accessto food and water. Both male and female mice (8–12 weeks old)were used in the PMA-induced ICD model and DSS-induced colitismodel. Only female mice were used in the endotoxemia model.All experiments were approved by the Animal Ethics Committeeof the Faculty of Science of Ghent University.
PMA-induced ICD model
Female mice were used between 2 and 6 months of age. Twentymicroliters of containing 1 μg PMA (Sigma-Aldrich) dissolved in
acetone was applied to both sides of the left auricle, and 20 μLof vehicle was applied to both sides of the right auricle. Auricularedema was assessed by measuring with a Vernier caliper (Dyer)the auricular thickness before and 2, 4, 9, and 24 h after treat-ment. For histological and immunohistochemical analyses, micewere euthanized 12 or 24 h after PMA application, auricular tis-sue was fixed in 4% paraformaldehyde (PFA) and processed fur-ther for different applications. For gene expression analysis, tissuewas collected 24 h after PMA application and stored in RNALatersolution (Ambion) until used for RNA isolation.
DSS-induced colitis model
Acute colitis was induced by adding DSS (36–50 kDa; MP Biomed-icals) to the drinking water (1.5% for PKP3−/− and PKP3−/−IEC
mice; 2% for PKP3−/−hem mice; DSS concentrations optimized con-sidering strain differences in DSS sensitivity [46]). Body weight,
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 11
gross or occult blood in stool, and stool consistency were deter-mined daily. Fecal blood was determined using Hemoccult SENSA(Beckman Coulter) analysis. The baseline clinical score was deter-mined on day 0. Absence of weight loss was scored as 0, weightloss of 1–5% below baseline as 1, 5–10% as 2, 10–20% as 3, and>20% as 4. For bleeding, a score of 0 was assigned for no blood,2 for positive Hemoccult, and 4 for gross bleeding. For stool con-sistency, a score of 0 was assigned for well-formed pellets, 2 forpasty stool, and 4 for liquid stool. The scores were combined anddivided by three, resulting in an overall clinical score ranging from0 (healthy) to 4 (maximal colitis). Postmortem, the entire colonfrom cecum to anus was removed, its length was measured asan indicator of inflammation, and colon samples were taken forpreparing PFA-fixed, paraffin-embedded sections.
Endotoxemia model
Mice were injected intraperitoneally (i.p.) with the indicated doseof LPS from Salmonella enterica serotype abortus equi (Sigma).Rectal temperature was measured at different times after chal-lenge. Blood and tissue samples were collected at designated timesafter injection. Mice were bled by retro-orbital bleeding or heartpuncture, and serum was prepared and stored at –20°C until used.
Measurement of cytokines in serum and explantsupernatant
Cytokines and chemokines were measured with the Bio-Plexcytokine assays (Bio-Rad) according to the manufacturer’s instruc-tions. TNF bioactivity (in U/mg tissue) was determined by theL929 bioassay system. Serum IL-6 levels were determined using a7TD1 bioassay.
Real-time qRT-PCR
Organs were stored in RNALater (Ambion) and RNA was isolatedwith the RNeasy Mini Kit (Qiagen). cDNA was synthesized bythe iScript cDNA Synthesis Kit (BioRad). Real-time PCR was per-formed on the Light Cycler 480 system (Roche) using the Light-Cycler 480 SYBR Green I Master Mix (Roche). Expression levelswere normalized to the expression of the two most stable referencegenes, which were determined for each organ using the geNormHousekeeping Gene Selection Software [47].
Histopathology and immunostaining
Skin, small intestine, and colon tissues were fixed with 4% PFA,embedded in paraffin, and sectioned at 4 μm. For H&E staining,sections were dewaxed and stained with hematoxylin (Fluka) andeosin (Merck). Mast cells were visualized by using 0.1% tolui-dine blue (Sigma-Aldrich). Ten high-power fields were evaluated,
and a mean was determined. Neutrophil infiltration was quanti-fied by counting the number of PMN in 10 high-power fields. Todetect proliferating keratinocytes, immunostaining for Ki67 wasperformed with rat anti-mouse antibody (Dako). Immunohisto-chemistry was performed by using the indirect avidin−biotin com-plex (Vector) method as described in the manual. Macrophageswere detected by staining of citrate buffer retrieved sections withrat anti-F4/80 antibody (Serotec) and subsequent incubation witha secondary Alexa 564 goat anti-rat antibody. The total number ofcells was counted in ten different high-power fields from each sec-tion. Fluorescent images and light microscopy images were takenby a laser scanning confocal microscope (Leica TCS SP5) and anOlympus light microscope, respectively.
Gut permeability
FITC-labeled dextran (4 kDa, Sigma) was administered to mice bygavage at 150 mg/kg body weight. After 5 h, blood was collectedin EDTA-coated tubes (Sarstedt) by heart puncture, and plasmawas prepared. Leakage of FITC-labeled dextran into the circulationwas determined by measurement of the plasma fluorescence (λex/λem = 488/520 nm). Values were normalized to the lowest value.
Thioglycollate neutrophils and macrophages
Mice were injected i.p. with 4 mL of 3% thioglycollate followed byperitoneal lavage 4 days later to collect macrophages. To enrichfor neutrophils, mice were injected with 1 mL of 3% thioglycol-late, and peritoneal lavage was performed 4 h later with ice-coldPBS. To remove red blood cells, cell pellets were treated withammonium chloride potassium lysis buffer (Life Technologies),and remaining cells were counted and plated in 6-well plates ata density of 2 × 106 cells per well. Macrophages were washed4 h later to remove nonadherent cells and subsequently incubatedwith LPS (1 μg/mL). Neutrophils were incubated overnight withLPS (1 μg/mL) or PMA (1 μg/mL) 1 h after plating. At the indi-cated time points, cells were harvested and RNA was isolated.
Statistical analysis
Data are presented as means ± SEM. Data were analyzed by anunpaired t-test. Survival curves were compared using a log-ranktest. Significance levels are indicated as follows: *, 0.01 � p <
0.05; **, 0.001 � p < 0.01; ***, 0.0005 � p < 0.001; ****, p <
0.0005.
Acknowledgments: We thank Mara Vleeschouwers for techni-cal assistance and Dr. Amin Bredan for careful editing of the
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

12 Tatyana Sklyarova et al. Eur. J. Immunol. 2015. 00: 1–13
manuscript. This research was supported by the Research Founda-tion - Flanders (FWO), the Concerted Research Actions (GOA) ofGhent University, and the Belgian Science Policy (InteruniversityAttraction Pools - IAP7/07).
Conflict of interest: The authors declare no commercial or finan-cial conflict of interest.
References
1 Niessen, M. T., Iden, S. and Niessen, C. M., The in vivo function of mam-
malian cell and tissue polarity regulators–how to shape and maintain
the epidermal barrier. J. Cell Sci. 2012. 125: 3501–3510.
2 Peterson, L. W. and Artis, D., Intestinal epithelial cells: regulators of
barrier function and immune homeostasis. Nat. Rev. Immunol. 2014. 14:
141–153.
3 Actis, G. C. and Rosina, F., Inflammatory bowel disease: an archetype
disorder of outer environment sensor systems. World J. Gastrointest. Phar-
macol. Ther. 2013. 4: 41–46.
4 Grainge, C. L. and Davies, D. E., Epithelial injury and repair in airways
diseases. Chest 2013. 144: 1906–1912.
5 Petersen, T. K., In vivo pharmacological disease models for psoriasis and
atopic dermatitis in drug discovery. Basic Clin. Pharmacol. Toxicol. 2006. 99:
104–115.
6 Hanel, K. H., Cornelissen, C., Luscher, B. and Baron, J. M., Cytokines and
the skin barrier. Int. J. Mol. Sci. 2013. 14: 6720–6745.
7 John, L. J., Fromm, M. and Schulzke, J. D., Epithelial barriers in intestinal
inflammation. Antioxid. Redox Signal. 2011. 15: 1255–1270.
8 Turner, J. R., Intestinal mucosal barrier function in health and disease.
Nat. Rev. Immunol. 2009. 9: 799–809.
9 Solomon, L., Mansor, S., Mallon, P., Donnelly, E., Hoper, M., Loughrey,
M., Kirk, S. et al., The dextran sulphate sodium (DSS) model of colitis: an
overview. Comp. Clin. Pathol. 2010. 19: 235–239.
10 Bosmann, M. and Ward, P. A., The inflammatory response in sepsis.
Trends Immunol. 2013. 34: 129–136.
11 Vandenbroucke, R. E., Vanlaere, I., Van Hauwermeiren, F., Van Won-
terghem, E., Wilson, C. and Libert, C., Pro-inflammatory effects of matrix
metalloproteinase 7 in acute inflammation. Mucosal Immunol. 2014. 7:
579–588.
12 Vandenbroucke, R. E., Dejonckheere, E., Van Hauwermeiren, F., Lodens,
S., De Rycke, R., Van Wonterghem, E., Staes, A. et al., Matrix metallopro-
teinase 13 modulates intestinal epithelial barrier integrity in inflamma-
tory diseases by activating TNF. EMBO Mol. Med. 2013. 5: 932–948.
13 Guo, S., Al-Sadi, R., Said, H. M. and Ma, T. Y., Lipopolysaccharide causes
an increase in intestinal tight junction permeability in vitro and in vivo
by inducing enterocyte membrane expression and localization of TLR-4
and CD14. Am. J. Pathol. 2013. 182: 375–387.
14 Ilan, Y., Leaky gut and the liver: a role for bacterial translocation in non-
alcoholic steatohepatitis. World J. Gastroenterol. 2012. 18: 2609–2618.
15 Faries, P. L., Simon, R. J., Martella, A. T., Lee, M. J. and Machiedo, G.
W., Intestinal permeability correlates with severity of injury in trauma
patients. J. Trauma 1998. 44: 1031–1035; discussion 1035–1036.
16 Swank, G. M. and Deitch, E. A., Role of the gut in multiple organ failure:
bacterial translocation and permeability changes. World J. Surg. 1996. 20:
411–417.
17 Newton, K. and Dixit, V. M., Signaling in innate immunity and inflamma-
tion. Cold Spring Harb. Perspect. Biol. 2012. 4. pii: a006049. doi: 10.1101/csh-
perspect.a006049
18 Hatzfeld, M., Wolf, A. and Keil, R., Plakophilins in desmosomal adhesion
and signaling. Cell Commun. Adhes. 2014. 21: 25–42.
19 Bass-Zubek, A. E., Godsel, L. M., Delmar, M. and Green, K. J., Plakophilins:
multifunctional scaffolds for adhesion and signaling. Curr. Opin. Cell Biol.
2009. 21: 708–716.
20 Munoz, W. A., Lee, M., Miller, R. K., Ahmed, Z., Ji, H., Link, T. M., Lee, G. R.
et al., Plakophilin-3 catenin associates with the ETV1/ER81 transcription
factor to positively modulate gene activity. PLoS One 2014. 9: e86784.
21 Neuber, S., Muhmer, M., Wratten, D., Koch, P. J., Moll, R. and Schmidt,
A., The desmosomal plaque proteins of the plakophilin family. Dermatol.
Res. Pract. 2010. 2010: 101452. doi: 10.1155/2010/101452.
22 Bonne, S., Gilbert, B., Hatzfeld, M., Chen, X., Green, K. J. and van Roy,
F., Defining desmosomal plakophilin-3 interactions. J. Cell Biol. 2003. 161:
403–416.
23 Bonne, S., van Hengel, J., Nollet, F., Kools, P. and van Roy, F., Plakophilin-
3, a novel armadillo-like protein present in nuclei and desmosomes of
epithelial cells. J. Cell Sci. 1999. 112 (Pt 14): 2265–2276.
24 Todorovic, V., Koetsier, J. L., Godsel, L. M. and Green, K. J., Plakophilin 3
mediates Rap1-dependent desmosome assembly and adherens junction
maturation. Mol. Biol. Cell 2014. 25: 3749–3764.
25 Hofmann, I., Casella, M., Schnolzer, M., Schlechter, T., Spring, H. and
Franke, W. W., Identification of the junctional plaque protein plakophilin
3 in cytoplasmic particles containing RNA-binding proteins and the
recruitment of plakophilins 1 and 3 to stress granules. Mol. Biol. Cell 2006.
17: 1388–1398.
26 Yang, C., Strobel, P., Marx, A. and Hofmann, I., Plakophilin-associated
RNA-binding proteins in prostate cancer and their implications in tumor
progression and metastasis. Virchows Arch. 2013. 463: 379–390.
27 Fischer-Keso, R., Breuninger, S., Hofmann, S., Henn, M., Rohrig, T., Stro-
bel, P., Stoecklin, G. et al., Plakophilins 1 and 3 bind to FXR1 and thereby
influence the mRNA stability of desmosomal proteins. Mol. Cell. Biol. 2014.
34: 4244–4256.
28 Sklyarova, T., Bonne, S., D’Hooge, P., Denecker, G., Goossens, S., De
Rycke, R., Borgonie, G. et al., Plakophilin-3-deficient mice develop hair
coat abnormalities and are prone to cutaneous inflammation. J. Invest.
Dermatol. 2008. 128: 1375–1385.
29 Horne, D. J., Randhawa, A. K., Chau, T. T., Bang, N. D., Yen, N.
T., Farrar, J. J., Dunstan, S. J. et al., Common polymorphisms in the
PKP3-SIGIRR-TMEM16J gene region are associated with susceptibility to
tuberculosis. J. Infect. Dis. 2012. 205: 586–594.
30 Brose, N. and Rosenmund, C., Move over protein kinase C, you’ve got
company: alternative cellular effectors of diacylglycerol and phorbol
esters. J. Cell Sci. 2002. 115: 4399–4411.
31 de Boer, J., Williams, A., Skavdis, G., Harker, N., Coles, M., Tolaini, M.,
Norton, T. et al., Transgenic mice with hematopoietic and lymphoid spe-
cific expression of Cre. Eur. J. Immunol. 2003. 33: 314–325.
32 Ramirez, A., Page, A., Gandarillas, A., Zanet, J., Pibre, S., Vidal, M., Tusell,
L. et al., A keratin K5Cre transgenic line appropriate for tissue-specific or
generalized Cre-mediated recombination. Genesis 2004. 39: 52–57.
33 Kumar, V. and Sharma, A., Mast cells: emerging sentinel innate immune
cells with diverse role in immunity. Mol. Immunol. 2010. 48: 14–25.
34 Hruz, T., Laule, O., Szabo, G., Wessendorp, F., Bleuler, S., Oertle, L.,
Widmayer, P. et al., Genevestigator v3: a reference expression database
for the meta-analysis of transcriptomes. Adv. Bioinformatics 2008. 2008:
420747. doi: 10.1155/2008/420747.
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2015. 00: 1–13 Innate immunity 13
35 Nair, R. P., Duffin, K. C., Helms, C., Ding, J., Stuart, P. E., Goldgar, D., Gud-
jonsson, J. E. et al., Genome-wide scan reveals association of psoriasis
with IL-23 and NF-kappaB pathways. Nat. Genet. 2009. 41: 199–204.
36 Yao, Y., Richman, L., Morehouse, C., de los Reyes, M., Higgs, B. W.,
Boutrin, A., White, B. et al., Type I interferon: potential therapeutic target
for psoriasis? PLoS One 2008. 3: e2737.
37 Suarez-Farinas, M., Tintle, S. J., Shemer, A., Chiricozzi, A., Nograles, K.,
Cardinale, I., Duan, S. et al., Nonlesional atopic dermatitis skin is char-
acterized by broad terminal differentiation defects and variable immune
abnormalities. J. Allergy Clin. Immunol. 2011. 127: 954–964 e951–e954.
38 Pastorelli, L., De Salvo, C., Mercado, J. R., Vecchi, M. and Pizarro,
T. T., Central role of the gut epithelial barrier in the pathogen-
esis of chronic intestinal inflammation: lessons learned from ani-
mal models and human genetics. Front. Immunol. 2013. 4: 280. doi:
10.3389/fimmu.2013.00280.
39 el Marjou, F., Janssen, K. P., Chang, B. H., Li, M., Hindie, V., Chan, L., Lou-
vard, D. et al., Tissue-specific and inducible Cre-mediated recombination
in the gut epithelium. Genesis 2004. 39: 186–193.
40 Costantini, T. W., Eliceiri, B. P., Peterson, C. Y., Loomis, W. H., Putnam, J.
G., Baird, A., Wolf, P. et al., Quantitative assessment of intestinal injury
using a novel in vivo, near-infrared imaging technique. Mol. Imaging 2010.
9: 30–39.
41 Fink, M. P., Intestinal epithelial hyperpermeability: update on the patho-
genesis of gut mucosal barrier dysfunction in critical illness. Curr. Opin.
Crit. Care 2003. 9: 143–151.
42 Wolf, A., Krause-Gruszczynska, M., Birkenmeier, O., Ostareck-Lederer,
A., Huttelmaier, S. and Hatzfeld, M., Plakophilin 1 stimulates translation
by promoting eIF4A1 activity. J. Cell Biol. 2010. 188: 463–471.
43 Hinman, M. N. and Lou, H., Diverse molecular functions of Hu proteins.
Cell. Mol. Life Sci. 2008. 65: 3168–3181.
44 Yiakouvaki, A., Dimitriou, M., Karakasiliotis, I., Eftychi, C., Theocharis,
S. and Kontoyiannis, D. L., Myeloid cell expression of the RNA-binding
protein HuR protects mice from pathologic inflammation and colorectal
carcinogenesis. J. Clin. Invest. 2012. 122: 48–61.
45 Fournier, B. M. and Parkos, C. A., The role of neutrophils during intestinal
inflammation. Mucosal Immunol. 2012. 5: 354–366.
46 Stevceva, L., Pavli, P., Buffinton, G., Wozniak, A. and Doe, W. F., Dex-
tran sodium sulphate-induced colitis activity varies with mouse strain
but develops in lipopolysaccharide-unresponsive mice. J. Gastroenterol.
Hepatol. 1999. 14: 54–60.
47 Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De
Paepe, A. and Speleman, F., Accurate normalization of real-time quan-
titative RT-PCR data by geometric averaging of multiple internal control
genes. Genome Biol. 2002. 3: RESEARCH0034.
Abbreviations: DAMPs: damage-associated molecular patterns · DSS:
dextran sulfate sodium · IBD: inflammatory bowel disease · ICD: irritant
contact dermatitis · PFA: paraformaldehyde · PKP3: plakophilin-3 · SIRS:
systemic inflammatory response syndrome · SPF: specific pathogen-
free
Full correspondence: Dr. Roosmarijn E. Vandenbroucke, Mouse Geneticsin Inflammation, Inflammation Research Center, VIB & GhentUniversity, Technologiepark 927, B-9052 Ghent, Belgiume-mail: [email protected]: +32-92217673
Additional correspondence: Dr. Frans van Roy, Molecular Cell BiologyUnit, Inflammation Research Center, VIB & Ghent University,Technologiepark 927, B-9052 Ghent, Belgiume-mail: [email protected]
Received: 28/12/2014Revised: 29/5/2015Accepted: 8/7/2015Accepted article online: 14/7/2015
C© 2015 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Related Documents











