Hell’s Gate globin I: An acid and thermostable bacterial hemoglobin resembling mammalian neuroglobin Aik-Hong Teh a , Jennifer A. Saito a , Aida Baharuddin a,1 , Jason R. Tuckerman b , James S. Newhouse c , Masaomi Kanbe a , Elizabeth I. Newhouse c , Rashidah Abdul Rahim a , Frédérique Favier d , Claude Didierjean d , Eduardo H.S. Sousa e , Matthew B. Stott f , Peter F. Dunfield g , Gonzalo Gonzalez b , Marie-Alda Gilles-Gonzalez b , Nazalan Najimudin a , Maqsudul Alam a,h,i,⇑ a Centre for Chemical Biology, Universiti Sains Malaysia, 10 Persiaran Bukit Jambul, 11900 Bayan Lepas, Penang, Malaysia b Department of Biochemistry, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390-9038, United States c Maui High Performance Computing Center, 550 Lipoa Parkway, Kihei, Maui, HI 96753, United States d LCM3B, Equipe Biocristallographie, UMR 7036 CNRS-UHP, Faculté des Sciences et Techniques, Nancy Université, BP 239, 54506 Vandoeuvre Cedex, France e Department of Organic and Inorganic Chemistry, Universidade Federal do Ceará – Centro de Ciências, Campus do Pici, CEP 60455-760 Fortaleza-Ceará, Brazil f GNS Science, Extremophile Research Group, Private Bag 2000, Taupo, New Zealand g Department of Biological Sciences, University of Calgary, 2500 University Drive NW, Calgary, Alberta, Canada T2N 1N4 h Department of Microbiology, University of Hawaii, 2538 McCarthy Mall, Honolulu, HI 96822, United States i Advanced Studies in Genomics, Proteomics and Bioinformatics, University of Hawaii, 2565 McCarthy Mall, Honolulu, HI 96822, United States article info Article history: Received 18 May 2011 Revised 17 August 2011 Accepted 1 September 2011 Available online 9 September 2011 Edited by Stuart Ferguson Keywords: Hell’s Gate globin I Globin Crystal structure Oxidation resistance Molecular dynamics Oxygen binding abstract Hell’s Gate globin I (HGbI), a heme-containing protein structurally homologous to mammalian neu- roglobins, has been identified from an acidophilic and thermophilic obligate methanotroph, Meth- ylacidiphilum infernorum. HGbI has very high affinity for O 2 and shows barely detectable autoxidation in the pH range of 5.2–8.6 and temperature range of 25–50 °C. Examination of the heme pocket by X-ray crystallography and molecular dynamics showed that conformational move- ments of Tyr29(B10) and Gln50(E7), as well as structural flexibility of the GH loop and H-helix, may play a role in modulating its ligand binding behavior. Bacterial HGbI’s unique resistance to the sort of extreme acidity that would extract heme from any other hemoglobin makes it an ideal candidate for comparative structure–function studies of the expanding globin superfamily. Ó 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Hell’s Gate (Tikitere), a region of fuming geothermal vents and caustic soil in New Zealand, is home to Methylacidiphilum inferno- rum, an obligately aerobic methanotroph that grows optimally at 60 °C and pH 2.0 [1]. From the genome of M. infernorum [2], we identified five different globins including Hell’s Gate globin I (HGbI), a single-domain protein with 133 residues. Until the 1970s, the globin family only included the vertebrate hemoglobins, myoglobins, and symbiotic hemoglobins of legumi- nous plants. Since then, the globins have grown into a superfamily that includes the non-symbiotic plant hemoglobins [3], fungal and bacterial flavohemoglobins [4,5], invertebrate globins [6], and truncated hemoglobins [7]. Recent additions are the mammalian neuroglobin (Ngb) [8] and cytoglobin (Cgb) [9], and prokaryotic glo- bin-coupled sensors [10], protoglobins [11], and thermoglobin [12]. 0014-5793/$36.00 Ó 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2011.09.002 Abbreviations: HGbI, Hell’s Gate globin I; oxy-HGbI, oxygen-bound HGbI; act- HGbI, acetate-bound HGbI; Ngb, neuroglobin; Cgb, cytoglobin; CO-Ngb, CO-bound murine neuroglobin; swMb, sperm whale myoglobin; AsHb, Ascaris suum hemo- globin; AaTgb, Aquifex aeolicus thermoglobin; CjHb, Campylobacter jejuni hemoglo- bin; VsHb, Vitreoscilla stercoraria hemoglobin; ReFHb, Ralstonia eutropha flavohemoglobin; HMP, Escherichia coli flavohemoglobin; ClHb, Cerebratulus lacteus mini-hemoglobin; SDgb, three-over-three single-domain globin related to the N- terminal of flavohemoglobins ⇑ Corresponding author at: Centre for Chemical Biology, Universiti Sains Malay- sia, 10 Persiaran Bukit Jambul, 11900 Bayan Lepas, Penang, Malaysia. Fax: +60 4 6535514. E-mail addresses: [email protected], [email protected] (M. Alam). 1 Present address: Department of Pharmaceutical Chemistry, University of California-San Francisco, 600 16th Street, Genentech Hall, San Francisco, CA 94143- 2280, United States. FEBS Letters 585 (2011) 3250–3258 journal homepage: www.FEBSLetters.org

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FEBS Letters 585 (2011) 3250–3258
journal homepage: www.FEBSLetters .org
Hell’s Gate globin I: An acid and thermostable bacterial hemoglobinresembling mammalian neuroglobin
Aik-Hong Teh a, Jennifer A. Saito a, Aida Baharuddin a,1, Jason R. Tuckerman b, James S. Newhouse c,Masaomi Kanbe a, Elizabeth I. Newhouse c, Rashidah Abdul Rahim a, Frédérique Favier d,Claude Didierjean d, Eduardo H.S. Sousa e, Matthew B. Stott f, Peter F. Dunfield g, Gonzalo Gonzalez b,Marie-Alda Gilles-Gonzalez b, Nazalan Najimudin a, Maqsudul Alam a,h,i,⇑a Centre for Chemical Biology, Universiti Sains Malaysia, 10 Persiaran Bukit Jambul, 11900 Bayan Lepas, Penang, Malaysiab Department of Biochemistry, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390-9038, United Statesc Maui High Performance Computing Center, 550 Lipoa Parkway, Kihei, Maui, HI 96753, United Statesd LCM3B, Equipe Biocristallographie, UMR 7036 CNRS-UHP, Faculté des Sciences et Techniques, Nancy Université, BP 239, 54506 Vandoeuvre Cedex, Francee Department of Organic and Inorganic Chemistry, Universidade Federal do Ceará – Centro de Ciências, Campus do Pici, CEP 60455-760 Fortaleza-Ceará, Brazilf GNS Science, Extremophile Research Group, Private Bag 2000, Taupo, New Zealandg Department of Biological Sciences, University of Calgary, 2500 University Drive NW, Calgary, Alberta, Canada T2N 1N4h Department of Microbiology, University of Hawaii, 2538 McCarthy Mall, Honolulu, HI 96822, United Statesi Advanced Studies in Genomics, Proteomics and Bioinformatics, University of Hawaii, 2565 McCarthy Mall, Honolulu, HI 96822, United States
a r t i c l e i n f o a b s t r a c t
Article history:Received 18 May 2011Revised 17 August 2011Accepted 1 September 2011Available online 9 September 2011
Edited by Stuart Ferguson
Keywords:Hell’s Gate globin IGlobinCrystal structureOxidation resistanceMolecular dynamicsOxygen binding
0014-5793/$36.00 � 2011 Federation of European Biodoi:10.1016/j.febslet.2011.09.002
Abbreviations: HGbI, Hell’s Gate globin I; oxy-HGHGbI, acetate-bound HGbI; Ngb, neuroglobin; Cgb, cymurine neuroglobin; swMb, sperm whale myoglobinglobin; AaTgb, Aquifex aeolicus thermoglobin; CjHb, Cbin; VsHb, Vitreoscilla stercoraria hemoglobin;flavohemoglobin; HMP, Escherichia coli flavohemoglobmini-hemoglobin; SDgb, three-over-three single-domterminal of flavohemoglobins⇑ Corresponding author at: Centre for Chemical Bio
sia, 10 Persiaran Bukit Jambul, 11900 Bayan Lepas, P6535514.
E-mail addresses: [email protected], maqsudul@cc1 Present address: Department of Pharmaceutic
California-San Francisco, 600 16th Street, Genentech H2280, United States.
Hell’s Gate globin I (HGbI), a heme-containing protein structurally homologous to mammalian neu-roglobins, has been identified from an acidophilic and thermophilic obligate methanotroph, Meth-ylacidiphilum infernorum. HGbI has very high affinity for O2 and shows barely detectableautoxidation in the pH range of 5.2–8.6 and temperature range of 25–50 �C. Examination of theheme pocket by X-ray crystallography and molecular dynamics showed that conformational move-ments of Tyr29(B10) and Gln50(E7), as well as structural flexibility of the GH loop and H-helix, mayplay a role in modulating its ligand binding behavior. Bacterial HGbI’s unique resistance to the sortof extreme acidity that would extract heme from any other hemoglobin makes it an ideal candidatefor comparative structure–function studies of the expanding globin superfamily.� 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
chemical Societies. Published by E
bI, oxygen-bound HGbI; act-toglobin; CO-Ngb, CO-bound; AsHb, Ascaris suum hemo-
ampylobacter jejuni hemoglo-ReFHb, Ralstonia eutrophain; ClHb, Cerebratulus lacteusain globin related to the N-
logy, Universiti Sains Malay-enang, Malaysia. Fax: +60 4
busm.edu.my (M. Alam).al Chemistry, University ofall, San Francisco, CA 94143-
Hell’s Gate (Tikitere), a region of fuming geothermal vents andcaustic soil in New Zealand, is home to Methylacidiphilum inferno-rum, an obligately aerobic methanotroph that grows optimally at60 �C and pH 2.0 [1]. From the genome of M. infernorum [2], weidentified five different globins including Hell’s Gate globin I(HGbI), a single-domain protein with 133 residues.
Until the 1970s, the globin family only included the vertebratehemoglobins, myoglobins, and symbiotic hemoglobins of legumi-nous plants. Since then, the globins have grown into a superfamilythat includes the non-symbiotic plant hemoglobins [3], fungal andbacterial flavohemoglobins [4,5], invertebrate globins [6], andtruncated hemoglobins [7]. Recent additions are the mammalianneuroglobin (Ngb) [8] and cytoglobin (Cgb) [9], and prokaryotic glo-bin-coupled sensors [10], protoglobins [11], and thermoglobin [12].
lsevier B.V. All rights reserved.

A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258 3251
In silico prediction of globins in the M. infernorum genome poses achallenge to our classical understanding of hemoglobins and myo-globins, which are expected to be highly unstable at the low pH andhigh temperature that this microorganism optimally grows at.
Globins are typically composed of eight a-helices, labeled A–H,that fold into a three-over-three a-helical sandwich structure [13].Exceptions include the truncated hemoglobins which consist of foura-helices in a two-over-two arrangement [14]. The distinguishingproperties of globins, such as their widely varied ligand bindingaffinities, are due to the sequestration of a heme prosthetic groupby specific residues. In addition to the four nitrogen atoms of proto-porphyrin IX, the heme iron can coordinate axially with two other li-gands. One of these is the sixth coordination position of the heme,which is the binding site for ligands such as O2, CO, and NO. In certainglobins, this sixth coordination site may be occupied by a proteinside chain when it is not occupied by exogenous ligands [15]. Theother axial ligand is usually a proximal His that extends from theF-helix to coordinate to the heme iron. In heme proteins the Fe–His bond is quite stable at neutral pH because the pKa of the proximalHis is shifted drastically downward, to about 4.7, from the usuallyneutral pKa of imidazole side chains. Consequently, a drop in pH be-low 4.7 usually breaks the Fe–His bond. Indeed this is an effectiveway to extract heme from hemoglobins so as to convert them tothe non-functional apoproteins. Clearly, ordinary heme proteinswould be quite useless to an organism living in the acidic mud vol-canoes of Tikitere. Here we report the identification, cloning, expres-sion, purification, biophysical characterization, structural analysis,and molecular dynamics of the unusually acid-tolerant and thermo-stable HGbI from M. infernorum.
2. Materials and methods
2.1. Cloning, expression, and purification
The gene encoding HGbI from M. infernorum was amplified byPCR using primers containing NdeI and BamHI restriction sites.Alternatively, the HGbI gene was also engineered with a His-tag se-quence immediately before the stop codon. Both PCR productswere cloned into the pET-3a expression vector (Novagen), andthe resulting vectors were transformed into Escherichia coli Rosetta2(DE3)pLysS (Novagen). The cells were grown in LB medium con-taining 100 lg/ml ampicillin and 34 lg/ml chloramphenicol, andexpression was induced with 50 lM isopropyl-b-D-thiogalactopy-ranoside. The medium was further supplemented with 100 mg/lFeSO4�7H2O and 17 mg/l d-aminolevulinic acid at the time of pro-tein induction. After incubation for 16 h at 30 �C, the cells wereharvested by centrifugation and resuspended in buffer I (50 mMNa2HPO4, 200 mM NaCl, pH 7.2) containing 1 mM phenylmethyl-sulfonyl fluoride, sonicated, and centrifuged to remove cell debris.
His-tagged HGbI was applied to a POROS MC/20 column (Ap-plied Biosystem) charged with Co2+, and eluted with a linear gradi-ent (up to 250 mM) of imidazole in buffer I. Fractions containingHGbI were dialyzed at 4 �C against buffer I, then further purifiedon a HiLoad 16/60 Superdex 200 gel filtration column (GE Health-care). Untagged HGbI was first precipitated with 50–60%(NH4)2SO4 and resuspended in buffer I before being applied tothe HiLoad 16/60 Superdex 200 column. For greater homogeneity,the last purification step was repeated 3–4 times. The protein usedin the experiments was of sufficient quality to crystallize and yieldcrystal structures of high resolution.
2.2. Ligand binding and oxidation
Unless otherwise noted, all determinations of UV–Vis absorp-tion and ligand binding were carried out with 2–5 lM protein in
0.10 M sodium phosphate, pH 7.5 at 25 �C. Absorption spectrawere monitored with a Cary 4000 UV–Vis spectrophotometer (Var-ian). Measurements of the effect of pH on the deoxy state weredone by recording the absorption spectra at 25 �C in 0.20 M sodiumphosphate at pH 5.2, 7.0, and 8.6. Autoxidation measurementswere performed by periodically recording the spectra in air forthe samples in buffers without any reducing agent; these spectrawere deconvoluted into the met and oxy components to determinethe extent of oxidation at each time point. The effect of pH on themet state was measured by recording the absorption spectra at25 �C for protein in 0.20 M sodium phosphate at the pH valuesnoted above, or in 0.20 M potassium glutamate at pH 4.6, 7.0,and 9.0. Comparison of the behavior in glutamate versus acetatewas done for the ferric protein at 25 �C in 0.20 M potassium gluta-mate at pH 4.6 and 0.20 M potassium acetate at pH 4.6.
Laser-flash photolysis and stopped-flow measurements wereconducted with an LKS.60 laser kinetic spectrometer equippedwith a PiStar stopped-flow drive unit (Applied Photophysics). Forsample excitation, the LKS.60 spectrometer was coupled to a Quan-tel Brilliant B Nd:YAG laser with second-harmonic generation. Li-gand-binding kinetics was followed at the wavelength of themaximum difference between the starting and final species. Eachrate constant was calculated from a linear plot of kobs versus ligandconcentration, including at least four ligand concentrations.
Association rates for O2 (80–1280 lM) were measured by laser-flash photolysis at two ranges of wavelengths (412–417 and 433–435 nm), while those for CO (30–960 lM) were measured at419 nm. The O2 dissociation rate constant was measured by thestopped-flow method at both 412 and 433–435 nm by mixingthe oxy-protein (in 50–256 lM free O2) with a solution of2.0 mM sodium dithionite. The CO dissociation rate constant wasmeasured by ligand replacement in the stopped-flow method at419 nm by mixing the CO-protein in 10–30 lM free CO with a200 lM solution of NO. The O2 affinity was measured by directlytitrating the protein with O2 (10–1150 lM) in a background ofCO (10 lM) at either 25 or 50 �C. Titration data were analyzed bydeconvoluting the whole spectra and then fitting the data to a qua-dratic equation for single binding.
2.3. Crystallization and structure determination
The buffer of purified HGbI was exchanged with 50 mM Tris–HCl, 150 mM NaCl, pH 8.0, before crystallization. The acetate-bound HGbI (act-HGbI) crystals of both native and His-taggedforms were grown by vapor diffusion in drops containing 1.6–2.0 M (NH4)2SO4, 0.1 M sodium acetate, pH 4.6, and 10–28 mg pro-tein/ml at 4 �C. The crystals were cryoprotected with 30% glyceroland flash-cooled at 100 K prior to data collection. The oxygen-bound HGbI (oxy-HGbI) crystals were formed by transferring theact-HGbI crystals directly into drops containing 1.6–2.0 M(NH4)2SO4, 0.1 M Tris–HCl, pH 8.0.
A Fe-SAD diffraction data set for the His-tagged act-HGbI wascollected at the European Synchrotron Radiation Facility (ESRF),Grenoble, on beamline BM30A at Fe peak wavelength (1.7397 Å),and processed in space group C2221 to 2.0 Å resolution with XDS[16]. Phasing was carried out with the program Solve [17], and im-proved by density modification with Resolve [18]. The map wasreadily interpretable, and after the model was manually built withCoot [19], iterative refinement with Refmac [20] and Coot was car-ried out. Diffraction data for the native act-HGbI, His-tagged oxy-HGbI, as well as a second set of His-tagged act-HGbI were collectedon a Rigaku MicroMax-007 HF X-ray generator (k = 1.5418 Å)equipped with an R-AXIS IV++ area detector. All were processedin the same space group C2221 with CrystalClear (Rigaku). Usingthe Fe-SAD structure as an initial model, these structures werebuilt and refined using Refmac, Phenix [21], and Coot. Data

Table 1Data collection and refinement statistics.
oxy-HGbI act-HGbI
Data collectionSpace group C2221 C2221
Cell dimensionsa, b, c (Å) 70.1, 126.2, 147.2 70.0, 126.2, 148.4
Resolution (Å) 36.1–1.77 (1.83–1.77)a 38.9–1.80 (1.86–1.80)Rmerge 0.044 (0.31) 0.042 (0.26)I/r(I) 12.8 (3.4) 12.5 (2.8)Completeness (%) 99.9 (100.0) 97.2 (88.2)Redundancy 3.9 (3.8) 2.9 (2.0)
RefinementNo. reflections 63705 59417Rwork/Rfree 0.207/0.225 0.209/0.236R.M.S. deviations
Bond lengths (Å) 0.007 0.008Bond angles (�) 0.977 0.989
a Values in parentheses are for the highest resolution shell.
3252 A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258
collection and refinement statistics for the His-tagged oxy-HGbIand His-tagged act-HGbI are summarized in Table 1.
Structural alignments of HGbI with other heme proteins wereperformed and presented using the programs ClustalW [22], Dali[23], and ESPript [24]. The cavity and tunnel calculations were car-ried out with SURFNET [25] and Caver [26]. Analysis of proteininterfaces and buried areas were calculated with PISA [27]. All fig-ures were generated using PyMOL (http://www.pymol.org).
2.4. Molecular dynamics
Molecular dynamics of unliganded HGbI was carried out usingNAMD [28]. All other simulations were performed using Desmond(Schrödinger). Desmond had the advantage of the OPLS force field[29], which had parameters for acetate, and was significantly fasterin parallel runs. The unliganded HGbI model was embedded in awater box that extended at least 10 Å from protein to box edge.The protein was neutralized and NaCl was added to a concentra-tion of 0.15 M. 60000 cycles of minimization were carried out, thenthe model was heated to 300 K in 5000 steps and equilibrated inanother 5000 steps. MD was done for �28 ns with sampling everyps in the NPT ensemble. For the Desmond simulations the HGbImodel was built in the same way, but equilibrated using the de-fault settings in the Maestro/Desmond interface (Schrödinger) foran NPT ensemble.
3. Results and discussion
3.1. HGbI is a heme protein that is acid-stable in vitro, with pH-dependent hexacoordination in the deoxy state
HGbI was expressed and purified as the native and His-taggedproteins. The absorption spectra of the deoxy and met HGbI weretypical of hemoglobins (Fig. 1A and B). What was far from typicalwas the effect of pH on these proteins. The spectra of both themet and ferrous forms showed no loss of heme even after 12 h atpH 4.6, and the ferrous form did not oxidize detectably at thislow pH in air at 37 �C (Table 2). When bound to acetic acid, the fer-ric form could be kept overnight at pH 2.8 without ill effect (notshown). The only apparent effect of pH on HGbI was an intriguingchange in the coordination of the heme iron in the ferrous statefrom pentacoordinate at low pH (�4.6) to predominantly hexaco-ordinate at high pH (Fig. 1A). For the ferric HGbI, the heme ironwas predominantly hexacoordinate even at low pH, although thereappeared to be a slight increase in the hexacoordination at higherpH (Fig. 1B).
3.2. Binding of ligands to HGbI heme
The kon for CO binding to HGbI (39 lM�1 s�1) was comparableto the kon for O2 binding. This value for kon(CO) was comparable tothat reported for Ascaris suum hemoglobin (AsHb) [30], but 80times faster than that of sperm whale myoglobin (swMb) [31],and even faster than that of soybean leghemoglobin [32] (Table2). As expected, the CO affinity of HGbI decreased with increasingtemperatures (Table 2; Fig. 1D and E). The koff was more stronglydependent on temperature than the kon. As a result, CO affinity de-creased twofold when the temperature increased from 25 to 50 �C.
We expected that exogenous ligands would bind more slowly athigher compared to lower pH because an incoming ligand mustcompete against an internal sixth ligand at high pH but not atlow pH. Surprisingly, the effect of pH on CO binding appeared tobe small to insignificant (Fig. 1C). It is possible that we failed tosee an effect by flash photolysis because after flashing at all pHs,the CO rebinds relatively rapidly (kon � 39 lM�1 s�1 and about80-fold faster than for swMb) compared to the internal ligand.Ordinarily, possible shortcomings of flash photolysis experimentswould be circumvented by using stopped flow, but in the case ofHGbI an exceptionally high O2 affinity (discussed below) interfereswith stopped-flow measurements of CO binding. We thereforeconclude that the effects noted in Fig. 1C are the minimum obser-vable by flash photolysis, and an increased understanding of theimpact of the internal HGbI ligand will require special apparatiand additional study.
The affinity of HGbI for O2 was exceptionally high, with a Kd of0.5 nM at pH 7.0 and 25 �C (Table 2; Fig. 2A and B). This representsan affinity about ten-fold higher than that of the extremely O2-avidAsHb (Kd = 4.6 nM) [30]. A temperature rise from 25 to 50 �Ccaused the O2 affinity to drop 13-fold, and this was associated witha much faster koff at the higher temperature (Table 2 and Fig. 2C).The kinetic parameters for binding of O2 by HGbI at 25 �C resem-bled those of the Aquifex aeolicus thermoglobin (AaTgb) [12] (Table2).
3.3. Homology models of HGbI
Sequence analysis showed that HGbI is similar to a group ofbacterial and certain eukaryotic globins (Fig. 3). We built homologymodels of HGbI based on two top-scoring mammalian Ngb: humanNgb (PDB ID: 1OJ6) [33] and CO-bound murine neuroglobin (CO-Ngb; PDB ID: 1W92) [34]. Based on the homology model, the maindifferences between HGbI and Ngb were a shorter CD loop and theabsence of a D-helix in HGbI, and the presence of a distal Gln50(E7)in HGbI compared to the His(E7) of Ngb (Fig. S1). The backboneRMSD between the homology model and the subsequent HGbIcrystal structure is 1.5 Å, confirming that human Ngb is an excel-lent template for accurately predicting a 3D model of HGbI.
3.4. Crystal structure of HGbI
The crystal structures of native and His-tagged HGbI, at pH 4.6and 8.0, were solved in the space group C2221 and are highly iden-tical. HGbI features a fold of seven a-helices without the short D-helix, which nonetheless conforms to the classical three-over-threea-helical sandwich (Fig. 4A and B). A search with the Dali server[23] revealed that HGbI is structurally most similar to murineNgb (Z score 20.5) [34] and human Ngb (19.2) [33], as well as bac-terial globins containing Tyr(B10) and Gln(E7) such as Campylobac-ter jejuni hemoglobin (CjHb; 19.4) [35], Vitreoscilla stercorariahemoglobin (VsHb; 18.4) [36], Ralstonia eutropha flavohemoglobin(ReFHb; 18.1) [37], and Escherichia coli flavohemoglobin (HMP;16.9) [38]. In addition, swMb (17.5) [39], Cgb (16.3) [40,41], AsHb(14.4) [42], and Cerebratulus lacteus mini-hemoglobin (ClHb;

Fig. 1. Influence of pH and temperature on heme-iron coordination and CO binding of HGbI. (A) Deoxy-HGbI absorption spectra at pH 5.2 (gray), 7.0 (black), and 8.6 (brokenline) at 25 �C. (B) Met-HGbI absorption spectra in acetate at pH 4.6 (thick line), and in glutamate at pH 4.6 (broken line), pH 7 and 9 (thin lines). (C) CO binding to deoxy-HGbIat pH 5.2 (open triangles), 7.0 (open circles), and 8.6 (closed circles). (D) Determination of CO association rate constants at 25 (open circles) and 50 �C (closed circles) by laserflash photolysis at 419 nm. (E) Determination of CO dissociation rate constants at 25 and 50 �C.
Table 2Parameters for ligand binding and autoxidation of HGbI.a
O2 binding CO binding Autoxidation
kon (lM�1 s�1) koff (s�1) Kd (nM) kon (lM�1 s�1) koff (s�1) Kd (nM) kox (h�1)
HGbI (25 �C) 26b 0.014 0.53 39 0.026 0.67c NDd
HGbI (50 �C) 63b 0.43 6.8 81 0.10 1.2c 0.084AaTgbe 20 0.019 0.95 5.1 0.0066 1.3AsHbf 2.8 0.013 4.6 17 0.018 1.1swMbg 16 14 830 0.51 0.019 37 0.060
a All data were collected with HGbI in 0.10 M sodium phosphate, pH 7.0.b Calculated from directly measured koff and Kd.c Calculated from directly measured koff and kon.d No discernible oxidation at 25 or 37 �C after 24 h.e Values for Aquifex aeolicus thermoglobin (AaTgb) are from [12].f Values for Ascaris suum hemoglobin (AsHb) are from [30].g Values for sperm whale myoglobin (swMb) are from [31].
A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258 3253
11.0) [43] also share a high degree of overall structural similarity(Fig. 3).
There are three essentially identical subunits, I, II, and III, in theasymmetric unit (Fig. S2). Subunits I and II are related by a non-crystallographic twofold axis and form a dimer. The hydrophobicinterface is comprised mainly of the B-helices from each subunit,with a buried surface area of 748 Å2 (Fig. 4C). Subunits III fromtwo adjacent asymmetric units also form a similar dimer relatedby a crystallographic twofold axis. Meanwhile, the contacts be-tween subunits II and III are due purely to crystal packing, andhave a total surface area of 533 Å2. Analysis of cavities in HGbIindicates a ligand-binding cavity of about 100 Å3 in the distal site
(Fig. 4B), lined by mostly hydrophobic residues. Two main open-ings of the cavity lead to several tunnels for ligand migration,and are gated by highly conserved residues (Leu32, Phe43, andVal95 in the first opening and Leu58, Leu99, and Tyr123 in thesecond).
The heme is oriented similarly as in swMb, but is rotated 180�around the a–c meso axis with respect to murine CO-Ngb [34](Fig. 5A). The close contact between the heme’s B vinyl groupand Tyr92(G5) might have prevented the opposite orientation inHGbI. The heme also shares a similar degree of ruffling as in CO-Ngb; however, in metNgb where Ngb’s heme has slid to bind tothe distal His64(E7), it becomes more planar [44]. The heme iron

Fig. 2. Binding of O2 to HGbI. (A) Representative spectra from a titration at 25 �C in 10 lM CO (black), 10 lM CO + 4 lM O2 (broken black), 10 lM CO + 16 lM O2 (gray), and1150 lM O2 (dotted black). (B) Titration data analyzed by deconvoluting whole spectra as in (A), with analysis by fitting the data to a quadratic equation for single binding. (C)Determination of O2 koff at 25 (black) and 50 �C (gray).
Fig. 3. Alignment of HGbI with (1) bacterial and (2) eukaryotic globins. HGbI, as well as the bacterial globins, lacks the D-helix and contains Tyr(B10) and Gln(E7). All theglobins have been structurally determined except for Aquifex aeolicus thermoglobin (AaTgb): Campylobacter jejuni globin (CjHb; PDB code 2WY4), Vitreoscilla stercorariahemoglobin (VsHb; 3VHB), Ralstonia eutropha flavohemoglobin (ReFHb; 3OZW), Escherichia coli flavohemoglobin (HMP; 1GVH), murine CO-Ngb (mNgb; 1W92), human Ngb(hNgb; 1OJ6), sperm whale myoglobin (swMb; 1A6M), human cytoglobin (hCgb; 2DC3), Ascaris suum hemoglobin (AsHb; 1ASH), and Cerebratulus lacteus mini-hemoglobin(ClHb; 1KR7).
3254 A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258
is coordinated to His82(F8) in the proximal site, but the orientationof its imidazole ring differs by about 90� from that of the other bac-terial globins, which is hydrogen-bonded to the highly conserved
Tyr(G5) and Glu(H23) (Fig. 5B). HGbI’s sequence does not includeGlu(H23), and its His82(F8), hydrogen-bonded to the backbone Oof Leu78(F4) instead, may be prevented by Met130 from rotating
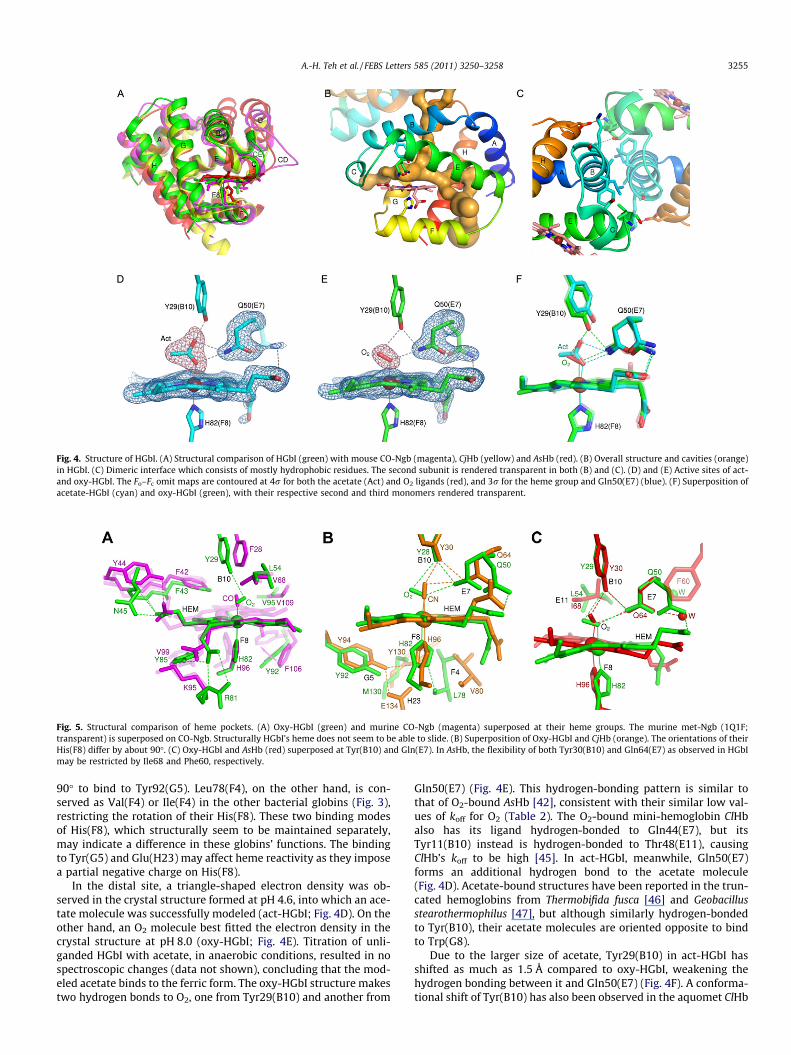
Fig. 4. Structure of HGbI. (A) Structural comparison of HGbI (green) with mouse CO-Ngb (magenta), CjHb (yellow) and AsHb (red). (B) Overall structure and cavities (orange)in HGbI. (C) Dimeric interface which consists of mostly hydrophobic residues. The second subunit is rendered transparent in both (B) and (C). (D) and (E) Active sites of act-and oxy-HGbI. The Fo–Fc omit maps are contoured at 4r for both the acetate (Act) and O2 ligands (red), and 3r for the heme group and Gln50(E7) (blue). (F) Superposition ofacetate-HGbI (cyan) and oxy-HGbI (green), with their respective second and third monomers rendered transparent.
Fig. 5. Structural comparison of heme pockets. (A) Oxy-HGbI (green) and murine CO-Ngb (magenta) superposed at their heme groups. The murine met-Ngb (1Q1F;transparent) is superposed on CO-Ngb. Structurally HGbI’s heme does not seem to be able to slide. (B) Superposition of Oxy-HGbI and CjHb (orange). The orientations of theirHis(F8) differ by about 90�. (C) Oxy-HGbI and AsHb (red) superposed at Tyr(B10) and Gln(E7). In AsHb, the flexibility of both Tyr30(B10) and Gln64(E7) as observed in HGbImay be restricted by Ile68 and Phe60, respectively.
A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258 3255
90� to bind to Tyr92(G5). Leu78(F4), on the other hand, is con-served as Val(F4) or Ile(F4) in the other bacterial globins (Fig. 3),restricting the rotation of their His(F8). These two binding modesof His(F8), which structurally seem to be maintained separately,may indicate a difference in these globins’ functions. The bindingto Tyr(G5) and Glu(H23) may affect heme reactivity as they imposea partial negative charge on His(F8).
In the distal site, a triangle-shaped electron density was ob-served in the crystal structure formed at pH 4.6, into which an ace-tate molecule was successfully modeled (act-HGbI; Fig. 4D). On theother hand, an O2 molecule best fitted the electron density in thecrystal structure at pH 8.0 (oxy-HGbI; Fig. 4E). Titration of unli-ganded HGbI with acetate, in anaerobic conditions, resulted in nospectroscopic changes (data not shown), concluding that the mod-eled acetate binds to the ferric form. The oxy-HGbI structure makestwo hydrogen bonds to O2, one from Tyr29(B10) and another from
Gln50(E7) (Fig. 4E). This hydrogen-bonding pattern is similar tothat of O2-bound AsHb [42], consistent with their similar low val-ues of koff for O2 (Table 2). The O2-bound mini-hemoglobin ClHbalso has its ligand hydrogen-bonded to Gln44(E7), but itsTyr11(B10) instead is hydrogen-bonded to Thr48(E11), causingClHb’s koff to be high [45]. In act-HGbI, meanwhile, Gln50(E7)forms an additional hydrogen bond to the acetate molecule(Fig. 4D). Acetate-bound structures have been reported in the trun-cated hemoglobins from Thermobifida fusca [46] and Geobacillusstearothermophilus [47], but although similarly hydrogen-bondedto Tyr(B10), their acetate molecules are oriented opposite to bindto Trp(G8).
Due to the larger size of acetate, Tyr29(B10) in act-HGbI hasshifted as much as 1.5 Å compared to oxy-HGbI, weakening thehydrogen bonding between it and Gln50(E7) (Fig. 4F). A conforma-tional shift of Tyr(B10) has also been observed in the aquomet ClHb

3256 A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258
structure [48]. As the Tyr30(B10) ? Phe mutation in AsHb in-creased its kon 14-fold [30], the shift of HGbI Tyr29(B10) could pos-sibly have a similar effect of increasing the cavity size, partlycontributing to a kon for O2 10-fold higher than that of AsHb. InAsHb, its corresponding Tyr30(B10) might be prevented from mov-ing by Ile68(E11), which is highly conserved as Leu(E11) in the bac-terial globins (Figs. 3 and 5C). The side chain of Gln50(E7) hasfurther been modeled with a minor alternate conformation, whichhas a lower occupancy in act-HGbI (averaging 0.25 in the threesubunits) than in oxy-HGbI (0.41) (Fig. 4D and E). Similar alternateconformations of the corresponding Gln44(E7) in the mini-hemo-globin ClHb have also been reported, but mutating it to alanine, va-line, or leucine had only small or moderate effect on O2 bindingrates, suggesting that in ClHb the E7 pathway was not the majorpathway for ligand migration [48,49]. The correspondingGln64(E7) in AsHb, however, may not be able to rotate likewisedue to steric hindrance from Phe60(E3) (Fig. 5C). In VsHb as wellas the flavohemoglobins ReFHb and HMP, on the other hand,Gln(E7) is not oriented into the distal site despite its prevalent con-servation [36–38].
Regarding the hexacoordinate species of the deoxy-HGbI ob-served at pH 8.6, structurally it is impossible for either Tyr29(B10)or Gln50(E7), at a distance of 5.3 or 4.3 Å away, to bind to Fe2+.Even if the heme of HGbI could slide and get nearer to them asin murine CO-Ngb [34], Tyr29(B10) would still be too far awaywhile the amide side chain of Gln50(E7) would chemically seemunable to bind to Fe2+. Furthermore, structural comparison withCO-Ngb has shown that in HGbI, such heme sliding would havebeen restricted by Phe43, Leu54, Tyr92, and Val95, as well as bythe larger number of hydrogen bonds between both the propio-nates of the heme and Gln44, Asn45, Arg81, and Tyr85.
3.5. Molecular dynamics behavior of HGbI
To gain further insights into the behavior of the HGbI hemepocket, molecular dynamics simulations over 100 ns were per-formed. The dissociation of O2 induces relatively large conforma-tional changes in the protein (Fig. 6). Compared to theunliganded form, oxy-HGbI has strikingly greater flexibility in theGH loop and H-helix, and less flexibility in the EF loop. Fewer avail-able conformations (less flexibility) in the EF loop with either O2 oracetate bound is expected, since interaction of the ligand withTyr29(B10) and Gln50(E7) will effectively anchor the E and F-heli-ces. More available conformations (greater flexibility) in the GHloop (O2 bound) and H-helix (O2 or acetate bound) may be a path-way for signaling. Bound O2 has more torsional freedom thanbound acetate (Fig. S3), in agreement with a higher B-factor for
Fig. 6. Flexibility of HGbI’s a-helices and loop regions. Data are from 80 ns MDsimulations of the unliganded (blue), acetate-bound (magenta) and O2-bound(cyan) structures. Alpha helices in HGbI are labeled at the top.
the O2 atom of O2 in the oxy-HGbI structure. In the unligandedform, the side chain torsions of Gln50(E7) occupy either one oftwo conformations (Fig. S4), as suggested in the crystal structures.Interconversion is slowed by the presence of either ligand, and inact-HGbI it is too slow to be detected in 100 ns. This agrees wellwith the lower occupancy for the minor alternate conformationof Gln50(E7) in act-HGbI than in oxy-HGbI. The solvent-accessiblesurface area of oxy-HGbI is 6000 Å2, while it is only 5300 Å2 for theunliganded HGbI. This change in size is primarily due to thechanges in the positions of the A and E-helices, as well as the CEand GH loops.
3.6. Speculations on the biological function of HGbI
To speculate on the possible biological function of HGbI, let usconsider three classic functions of heme proteins: O2 transport,heme-ligand sensing, and redox chemistry.
The extremely high O2 affinity and slow rate of O2 dissociationmake HGbI a poor candidate for an O2 transporter. While HGbIwould be a good O2 scavenger, it is not apparent why such a scav-enger would be needed by an organism like M. infernorum, whosemetabolism is probably based on combining O2 with methane, andwhose genome encodes proteins necessary for aerobic rather thananaerobic metabolism.
Likewise, heme-ligand sensing does not appear to be a promis-ing function for HGbI. Assuming a volume for a M. infernorum onthe order of a femtoliter, the nanomolar Kd value for binding ofO2 would imply strong binding even with only a few moleculesof O2 in the entire cell. Thus at low O2 concentrations the concen-tration of HGbI would severely perturb the concentration of O2,and at higher O2 concentrations the HGbI would be fully saturated.The possibility of sensing CO would also seem unlikely, because M.infernorum lacks the CO-processing enzymes that are regulated byCO sensors such as CooA.
This leaves redox chemistry, or else a wholly new class of hemefunction. M. infernorum is a methane-consuming organism thatcarries out some highly unusual redox reactions. The extreme sta-bility of the ferrous state may mean that ferric HGbI can acquire anelectron from some as yet unknown donor much more easily thanother heme proteins. Ferric HGbI has the ability, extremely unusualfor heme proteins, to bind acetate with fairly high affinity (Fig. 1B).In addition, HGbI features an oxidation-state and pH dependentbinding of an internal ligand. All of these observations suggest anunusual electrochemistry for HGbI.
3.7. Conclusion
Phylogenetic analysis has shown that the three-over-three sin-gle-domain globin related to the N-terminal of flavohemoglobins(SDgbs), and plant and metazoan globins form one of the three glo-bin lineages [50,51]. Furthermore, it was proposed that the meta-zoan globins evolved from a bacterial ancestor SDgb, which wasthen acquired by eukaryotes via lateral gene transfer [50,52].Strong evidence for this scenario comes from the fact that verte-brate Ngbs exhibit particularly high sequence similarity to bacte-rial SDgbs in PSI-BLAST searches, as compared to other globinclasses [50,52].
Here we have demonstrated that HGbI, a three-over-three bac-terial SDgb, has the closest structural resemblance to the verte-brate Ngbs. The high structural similarity provides a strongevolutionary link between the bacterial SDgbs and eukaryote Ngbs,and also indicates the possibility that the metazoan globins mayhave stemmed from the ancestral SDgb.
The HGbI protein is unusual due to an extremely strong resis-tance to oxidation in the face of low pH and high temperature,an O2 avidity that is even greater than that of AsHb, a CO on-rate

A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258 3257
so rapid as to be comparable to the O2 on-rate, and an ability tobind acetate with relatively high affinity in the ferric state. HGbIalso has interesting coordination structures, with the ferrous formundergoing a transition from penta- to hexacoordinate withincreasing pH, and the ferric form remaining predominantly hexa-coordinate at all pH. In the crystal structures of HGbI, conforma-tional movements were noted for Tyr29(B10) and Gln50(E7).Molecular dynamics studies predict an increased structural flexi-bility of Gln50(E7), the GH loop and the H-helix upon O2 binding.These structural features might play a role in HGbI’s exceptionallyhigh O2 affinity and extreme resistance to autoxidation. Additionalstudies will be needed to understand the unusual chemistry of thisinteresting hemoglobin and its importance to M. infernorum.
PDB accession numbers
The atomic coordinates and structure factors have been depos-ited in the Protein Data Bank as entries 3S1I (oxy-HGbI) and 3S1J(act-HGbI).
Acknowledgements
This work was supported by the U.S. Army Telemedicine and Ad-vanced Technology Research Center (W81XWH0520013) to M.A.,intramural funding from the Centre for Chemical Biology, UniversitiSains Malaysia (CCB@USM), and U.S. National Science FoundationGrant No. MCB620531 (M.A.G.G.). The authors acknowledge theTikitere Trust, the owners and guardians of Hell’s Gate.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2011.09.002.
References
[1] Dunfield, P.F., Yuryev, A., Senin, P., Smirnova, A.V., Stott, M.B., Hou, S., Ly, B.,Saw, J.H., Zhou, Z., Ren, Y., Wang, J., Mountain, B.W., Crowe, M.A., Weatherby,T.M., Bodelier, P.L., Liesack, W., Feng, L., Wang, L. and Alam, M. (2007) Methaneoxidation by an extremely acidophilic bacterium of the phylumVerrucomicrobia. Nature 450, 879–882.
[2] Hou, S., Makarova, K.S., Saw, J.H., Senin, P., Ly, B.V., Zhou, Z., Ren, Y., Wang, J.,Galperin, M.Y., Omelchenko, M.V., Wolf, Y.I., Yutin, N., Koonin, E.V., Stott, M.B.,Mountain, B.W., Crowe, M.A., Smirnova, A.V., Dunfield, P.F., Feng, L., Wang, L.and Alam, M. (2008) Complete genome sequence of the extremely acidophilicmethanotroph isolate V4, Methylacidiphilum infernorum, a representative ofthe bacterial phylum Verrucomicrobia. Biol. Direct 3, 26.
[3] Garrocho-Villegas, V., Gopalasubramaniam, S.K. and Arredondo-Peter, R.(2007) Plant hemoglobins: what we know six decades after their discovery.Gene 398, 78–85.
[4] Iwaasa, H., Takagi, T. and Shikama, K. (1992) Amino acid sequence of yeasthemoglobin. A two-domain structure. J. Mol. Biol. 227, 948–954.
[5] Vasudevan, S.G., Armarego, W.L., Shaw, D.C., Lilley, P.E., Dixon, N.E. and Poole,R.K. (1991) Isolation and nucleotide sequence of the hmp gene that encodes ahaemoglobin-like protein in Escherichia coli K-12. Mol. Gen. Genet. 226, 49–58.
[6] Weber, R.E. and Vinogradov, S.N. (2001) Nonvertebrate hemoglobins:functions and molecular adaptations. Physiol. Rev. 81, 569–628.
[7] Wittenberg, J.B., Bolognesi, M., Wittenberg, B.A. and Guertin, M. (2002)Truncated hemoglobins: a new family of hemoglobins widely distributed inbacteria, unicellular eukaryotes, and plants. J. Biol. Chem. 277, 871–874.
[8] Burmester, T., Weich, B., Reinhardt, S. and Hankeln, T. (2000) A vertebrateglobin expressed in the brain. Nature 407, 520–523.
[9] Burmester, T., Ebner, B., Weich, B. and Hankeln, T. (2002) Cytoglobin: a novelglobin type ubiquitously expressed in vertebrate tissues. Mol. Biol. Evol. 19,416–421.
[10] Hou, S., Freitas, T., Larsen, R.W., Piatibratov, M., Sivozhelezov, V., Yamamoto,A., Meleshkevitch, E.A., Zimmer, M., Ordal, G.W. and Alam, M. (2001) Globin-coupled sensors: a class of heme-containing sensors in Archaea and Bacteria.Proc. Natl. Acad. Sci. USA 98, 9353–9358.
[11] Freitas, T.A., Hou, S., Dioum, E.M., Saito, J.A., Newhouse, J., Gonzalez, G., Gilles-Gonzalez, M.A. and Alam, M. (2004) Ancestral hemoglobins in Archaea. Proc.Natl. Acad. Sci. USA 101, 6675–6680.
[12] Miranda, J.J., Maillett, D.H., Soman, J. and Olson, J.S. (2005) Thermoglobin,oxygen-avid hemoglobin in a bacterial hyperthermophile. J. Biol. Chem. 280,36754–36761.
[13] Holm, L. and Sander, C. (1993) Structural alignment of globins, phycocyaninsand colicin A. FEBS Lett. 315, 301–306.
[14] Pesce, A., Couture, M., Dewilde, S., Guertin, M., Yamauchi, K., Ascenzi, P.,Moens, L. and Bolognesi, M. (2000) A novel two-over-two a-helical sandwichfold is characteristic of the truncated hemoglobin family. EMBO J. 19, 2424–2434.
[15] Goodman, M.D. and Hargrove, M.S. (2001) Quaternary structure of ricenonsymbiotic hemoglobin. J. Biol. Chem. 276, 6834–6839.
[16] Kabsch, W. (1993) Automatic processing of rotation diffraction data fromcrystals of initially unknown symmetry and cell constants. J. Appl. Cryst. 26,795–800.
[17] Terwilliger, T.C. and Berendzen, J. (1999) Automated MAD and MIR structuresolution. Acta Crystallogr. D55, 849–861.
[18] Terwilliger, T.C. (2000) Maximum-likelihood density modification. ActaCrystallogr. D56, 965–972.
[19] Emsley, P. and Cowtan, K. (2004) Coot: model-building tools for moleculargraphics. Acta Crystallogr. D60, 2126–2132.
[20] Murshudov, G.N., Vagin, A.A. and Dodson, E.J. (1997) Refinement ofmacromolecular structures by the maximum-likelihood method. ActaCrystallogr. D53, 240–255.
[21] Adams, P.D., Afonine, P.V., Bunkóczi, G., Chen, V.B., Davis, I.W., Echols, N.,Headd, J.J., Hung, L.W., Kapral, G.J., Grosse-Kunstleve, R.W., McCoy, A.J.,Moriarty, N.W., Oeffner, R., Read, R.J., Richardson, D.C., Richardson, J.S.,Terwilliger, T.C. and Zwart, P. (2010) PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D66,213–221.
[22] Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A.,McWilliam, H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D.,Gibson, T.J. and Higgins, D.G. (2007) Clustal W and Clustal X version 2.0.Bioinformatics 23, 2947–2948.
[23] Holm, L. and Rosenström, P. (2010) Dali server: conservation mapping in 3D.Nucleic Acids Res. 38, W545–W549.
[24] Gouet, P., Courcelle, E., Stuart, D.I. and Metoz, F. (1999) ESPript: analysis ofmultiple sequence alignments in PostScript. Bioinformatics 15, 305–308.
[25] Laskowski, R.A. (1995) SURFNET: a program for visualizing molecular surfaces,cavities, and intermolecular interactions. J. Mol. Graph. 13, 323–330.
[26] Damborsky, J., Petrek, M., Banás, P. and Otyepka, M. (2007) Identification oftunnels in proteins, nucleic acids, inorganic materials and molecularensembles. Biotechnol. J. 2, 62–67.
[27] Krissinel, E. and Henrick, K. (2007) Inference of macromolecular assembliesfrom crystalline state. J. Mol. Biol. 372, 774–797.
[28] Phillips, J.C., Braun, R., Wang, W., Gumbart, J., Tajkhorshid, E., Villa, E., Chipot,C., Skeel, R.D., Kalé, L. and Schulten, K. (2005) Scalable molecular dynamicswith NAMD. J. Comput. Chem. 26, 1781–1802.
[29] Kaminski, G., Friesner, R.A., Tirado-Rives, J. and Jorgensen, W.L. (2001)Evaluation and reparametrization of the OPLS-AA force field for proteins viacomparison with accurate quantum chemical calculations on peptides. J. Phys.Chem. B 105, 6474–6487.
[30] De Baere, I., Perutz, M.F., Kiger, L., Marden, M.C. and Poyart, C. (1994)Formation of two hydrogen bonds from the globin to the heme-linked oxygenmolecule in Ascaris hemoglobin. Proc. Natl. Acad. Sci. USA 91, 1594–1597.
[31] Quillin, M.L., Arduini, R.M., Olson, J.S. and Philips, G.N.Jr. (1993) High-resolution crystal structures of distal histidine mutants of sperm whalemyoglobin. J. Mol. Biol. 234, 140–155.
[32] Hargrove, M.S., Barry, J.K., Brucker, E.A., Berry, M.B., Phillips, G.N.Jr., Olson, J.S.,Arredondo-Peter, R., Dean, J.M., Klucas, R.V. and Sarath, G. (1997)Characterization of recombinant soybean leghemoglobin a and apolar distalhistidine mutants. J. Mol. Biol. 266, 1032–1042.
[33] Pesce, A., Dewilde, S., Nardini, M., Moens, L., Ascenzi, P., Hankeln, T.,Burmester, T. and Bolognesi, M. (2003) Human brain neuroglobin structurereveals a distinct mode of controlling oxygen affinity. Structure 11, 1087–1095.
[34] Vallone, B., Nienhaus, K., Matthes, A., Brunori, M. and Nienhaus, G.U. (2004)The structure of carbonmonoxy neuroglobin reveals a heme-slidingmechanism for control of ligand affinity. Proc. Natl. Acad. Sci. USA 101,17351–17356.
[35] Shepherd, M., Barynin, V., Lu, C., Bernhardt, P.V., Wu, G., Yeh, S.R., Egawa, T.,Sedelnikova, S.E., Rice, D.W., Wilson, J.L. and Poole, R.K. (2010) The single-domain globin from the pathogenic bacterium Campylobacter jejuni: novel D-helix conformation, proximal hydrogen bonding that influences ligandbinding, and peroxidase-like redox properties. J. Biol. Chem. 285, 12747–12754.
[36] Tarricone, C., Galizzi, A., Coda, A., Ascenzi, P. and Bolognesi, M. (1997) Unusualstructure of the oxygen-binding site in the dimeric bacterial hemoglobin fromVitreoscilla sp.. Structure 5, 497–507.
[37] Ermler, U., Siddiqui, R.A., Cramm, R. and Friedrich, B. (1995) Crystal structureof the flavohemoglobin from Alcaligenes eutrophus at 1.75 Å resolution. EMBOJ. 14, 6067–6077.
[38] Ilari, A., Bonamore, A., Farina, A., Johnson, K.A. and Boffi, A. (2002) The X-raystructure of ferric Escherichia coli flavohemoglobin reveals an unexpectedgeometry of the distal heme pocket. J. Biol. Chem. 277, 23725–23732.

3258 A.-H. Teh et al. / FEBS Letters 585 (2011) 3250–3258
[39] Vojtechovsky, J., Chu, K., Berendzen, J., Sweet, R.M. and Schlichting, I. (1999)Crystal structures of myoglobin-ligand complexes at near-atomic resolution.Biophys. J. 77, 2153–2174.
[40] de Sanctis, D., Dewilde, S., Pesce, A., Moens, L., Ascenzi, P., Hankeln, T.,Burmester, T. and Bolognesi, M. (2004) Crystal structure of cytoglobin: thefourth globin type discovered in man displays heme hexa-coordination. J. Mol.Biol. 336, 917–927.
[41] Sugimoto, H., Makino, M., Sawai, H., Kawada, N., Yoshizato, K. and Shiro, Y.(2004) Structural basis of human cytoglobin for ligand binding. J. Mol. Biol.339, 873–885.
[42] Yang, J., Kloek, A.P., Goldberg, D.E. and Mathews, F.S. (1995) The structure ofAscaris hemoglobin domain I at 2.2 Å resolution: molecular features of oxygenavidity. Proc. Natl. Acad. Sci. USA 92, 4224–4228.
[43] Pesce, A., Nardini, M., Dewilde, S., Geuens, E., Yamauchi, K., Ascenzi, P., Riggs,A.F., Moens, L. and Bolognesi, M. (2002) The 109 residue nerve tissueminihemoglobin from Cerebratulus lacteus highlights striking structuralplasticity of the alpha-helical globin fold. Structure 10, 725–735.
[44] Vallone, B., Nienhaus, K., Brunori, M. and Nienhaus, G.U. (2004) The structureof murine neuroglobin: novel pathways for ligand migration and binding.Proteins 56, 85–92.
[45] Pesce, A., Nardini, M., Ascenzi, P., Geuens, E., Dewilde, S., Moens, L., Bolognesi,M., Riggs, A.F., Hale, A., Deng, P., Nienhaus, G.U., Olson, J.S. and Nienhaus, K.(2004) Thr-E11 regulates O2 affinity in Cerebratulus lacteus mini-hemoglobin.J. Biol. Chem. 279, 33662–33672.
[46] Bonamore, A., Ilari, A., Giangiacomo, L., Bellelli, A., Morea, V. and Boffi, A.(2005) A novel thermostable hemoglobin from the actinobacteriumThermobifida fusca. FEBS J. 272, 4189–4201.
[47] Ilari, A., Kjelgaard, P., von Wachenfeldt, C., Catacchio, B., Chiancone, E. andBoffi, A. (2007) Crystal structure and ligand binding properties of thetruncated hemoglobin from Geobacillus stearothermophilus. Arch. Biochem.Biophys. 457, 85–94.
[48] Pesce, A., Nardini, M., Dewilde, S., Capece, L., Martí, M.A., Congia, S., Salter,M.D., Blouin, G.C., Estrin, D.A., Ascenzi, P., Moens, L., Bolognesi, M. and Olson,J.S. (2011) Ligand migration in the apolar tunnel of Cerebratulus lacteus mini-hemoglobin. J. Biol. Chem. 286, 5347–5358.
[49] Salter, M.D., Nienhaus, K., Nienhaus, G.U., Dewilde, S., Moens, L., Pesce, A.,Nardini, M., Bolognesi, M. and Olson, J.S. (2008) The apolar channel inCerebratulus lacteus hemoglobin is the route for O2 entry and exit. J. Biol.Chem. 283, 35689–35702.
[50] Vinogradov, S.N., Hoogewijs, D., Bailly, X., Arredondo-Peter, R., Guertin, M.,Gough, J., Dewilde, S., Moens, L. and Vanfleteren, J.R. (2005) Three globinlineages belonging to two structural classes in genomes from the threekingdoms of life. Proc. Natl. Acad. Sci. USA 102, 11385–11389.
[51] Vinogradov, S.N., Hoogewijs, D., Bailly, X., Arredondo-Peter, R., Gough, J.,Dewilde, S., Moens, L. and Vanfleteren, J.R. (2006) A phylogenomic profile ofglobins. BMC Evol. Biol. 6, 31.
[52] Vinogradov, S.N., Hoogewijs, D., Bailly, X., Mizuguchi, K., Dewilde, S., Moens, L.and Vanfleteren, J.R. (2007) A model of globin evolution. Gene 398, 132–142.
Related Documents











