24 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro REVIEWS What drives the growth of the brain during develop- ment and how mammalian brains have evolved to reach their current level of complexity remain largely unknown. What are the molecular mechanisms involved in controlling the production of brain cells? What con- trols the balance between proliferation and cell death, which we assume, together with increased connectivity, to be the driving force behind brain growth? What con- trols the morphological plasticity of the brain during evolution and ontogeny? The brain expresses a large part of the genome, and the known actions of many secreted factors, such as FIBROBLAST GROWTH FACTORS (FGFs), insulin growth factors (IGFs), WNT PROTEINS, platelet-derived growth factors (PDGFs) and epidermal growth factor (EGF), tell us that a myriad of activities, which we are just beginning to understand, affect brain development. Brain cells can be viewed as being immersed in a sea of secreted factors, many of which affect cell proliferation and differentiation, and also influence the development of other body parts. Given the complexity of brain development, it is interesting that experiments with single factors have dis- cernible, and at times profound, effects on brain growth. Take, for example, the effects of FGFs on the prolifera- tion of brain cells 1–6 . Injections of FGF2 into the mouse brain at early embryonic stages result in an increase in the number of neurons that are generated, whereas injections at later stages, towards the end of embryo- genesis, result in an increase in the number of glial cells 7 . Although these changes are less than twofold, they are significant, as small variations in the proliferation of neocortical precursors have been proposed to account for the enormous growth of the cortex in mammals 8,9 . So, considering the many factors and effects involved in development, it is surprising (and encouraging) that experiments on the role of Sonic hedgehog (SHH) have shed light on the problem of brain growth. Indeed, recent findings 10 implicate the SHH–Gli signalling pathway in brain precursor proliferation, indicating mechanisms that might underlie brain growth during development and evolution, and giving a new twist to the evolving Hedgehog story (see REFS 11,12). Sonic hedgehog SHH is a member of the HEDGEHOG FAMILY of secreted glycoproteins, which affects the development of many organs and cell groups 13–17 . SHH acts through the Patched 1 (Ptc1)–Smoothened (Smo) receptor com- plex to trigger the activation of an intricate signal- transduction pathway (FIG. 1). Cells that respond to HEDGEHOG–GLI SIGNALLING AND THE GROWTH OF THE BRAIN Ariel Ruiz i Altaba*, Verónica Palma* and Nadia Dahmane ‡ The development of the vertebrate brain involves the creation of many cell types in precise locations and at precise times, followed by the formation of functional connections. To generate its cells in the correct numbers, the brain has to produce many precursors during a limited period. How this is achieved remains unclear, although several cytokines have been implicated in the proliferation of neural precursors. Understanding this process will provide profound insights, not only into the formation of the mammalian brain during ontogeny, but also into brain evolution. Here we review the role of the Sonic hedgehog–Gli pathway in brain development. Specifically, we discuss the role of this pathway in the cerebellar and cerebral cortices, and address the implications of these findings for morphological plasticity. We also highlight future directions of research that could help to clarify the mechanisms and consequences of Sonic hedgehog signalling in the brain. *The Skirball Institute, Developmental Genetics Program and Department of Cell Biology, New York University School of Medicine, 540 First Avenue, New York 10016, USA. ‡ Developmental Biology Institute of Marseille, Université de la Méditterannée–Centre National de la Recherche Scientifique, Campus de Luminy, Case 907, 13 288 Marseille Cedex 09, France. Correspondence to A.R.A. e-mail: [email protected] DOI: 10.1038/nrn704

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

24 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro
R E V I E W S
What drives the growth of the brain during develop-ment and how mammalian brains have evolved to reachtheir current level of complexity remain largelyunknown.What are the molecular mechanisms involvedin controlling the production of brain cells? What con-trols the balance between proliferation and cell death,which we assume, together with increased connectivity,to be the driving force behind brain growth? What con-trols the morphological plasticity of the brain duringevolution and ontogeny?
The brain expresses a large part of the genome, andthe known actions of many secreted factors, such asFIBROBLAST GROWTH FACTORS (FGFs), insulin growth factors(IGFs), WNT PROTEINS, platelet-derived growth factors(PDGFs) and epidermal growth factor (EGF), tell usthat a myriad of activities, which we are just beginning tounderstand, affect brain development. Brain cells can beviewed as being immersed in a sea of secreted factors,many of which affect cell proliferation and differentiation,and also influence the development of other body parts.
Given the complexity of brain development, it isinteresting that experiments with single factors have dis-cernible, and at times profound, effects on brain growth.Take, for example, the effects of FGFs on the prolifera-tion of brain cells1–6. Injections of FGF2 into the mouse
brain at early embryonic stages result in an increase inthe number of neurons that are generated, whereasinjections at later stages, towards the end of embryo-genesis, result in an increase in the number of glial cells7.Although these changes are less than twofold, they aresignificant, as small variations in the proliferation ofneocortical precursors have been proposed to accountfor the enormous growth of the cortex in mammals8,9.So, considering the many factors and effects involved indevelopment, it is surprising (and encouraging) thatexperiments on the role of Sonic hedgehog (SHH) haveshed light on the problem of brain growth. Indeed,recent findings10 implicate the SHH–Gli signallingpathway in brain precursor proliferation, indicatingmechanisms that might underlie brain growth duringdevelopment and evolution, and giving a new twist tothe evolving Hedgehog story (see REFS 11,12).
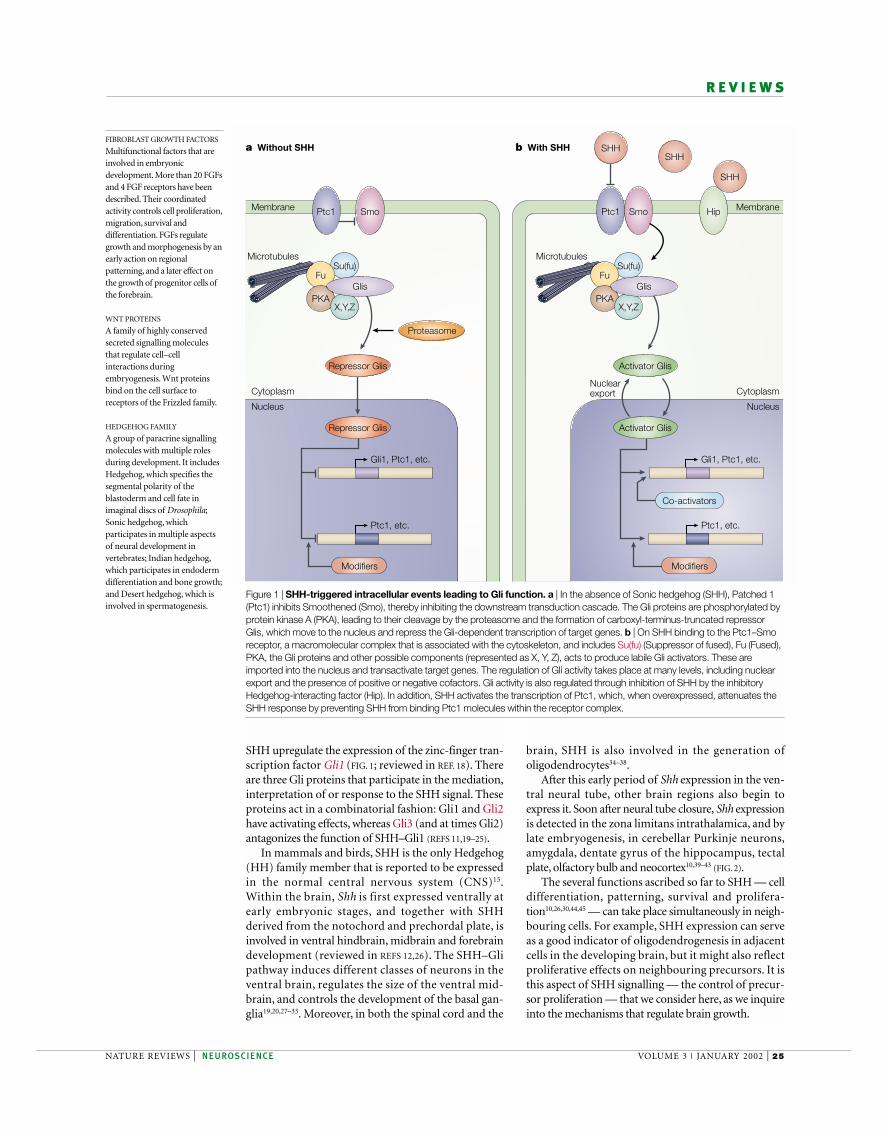
Sonic hedgehogSHH is a member of the HEDGEHOG FAMILY of secretedglycoproteins, which affects the development of manyorgans and cell groups13–17. SHH acts through thePatched 1 (Ptc1)–Smoothened (Smo) receptor com-plex to trigger the activation of an intricate signal-transduction pathway (FIG. 1). Cells that respond to
HEDGEHOG–GLI SIGNALLING ANDTHE GROWTH OF THE BRAINAriel Ruiz i Altaba*, Verónica Palma* and Nadia Dahmane‡
The development of the vertebrate brain involves the creation of many cell types in precise locationsand at precise times, followed by the formation of functional connections. To generate its cells in thecorrect numbers, the brain has to produce many precursors during a limited period. How this isachieved remains unclear, although several cytokines have been implicated in the proliferation ofneural precursors. Understanding this process will provide profound insights, not only into theformation of the mammalian brain during ontogeny, but also into brain evolution. Here we review therole of the Sonic hedgehog–Gli pathway in brain development. Specifically, we discuss the role ofthis pathway in the cerebellar and cerebral cortices, and address the implications of these findingsfor morphological plasticity. We also highlight future directions of research that could help to clarifythe mechanisms and consequences of Sonic hedgehog signalling in the brain.
*The Skirball Institute,Developmental GeneticsProgram and Department ofCell Biology, New YorkUniversity School ofMedicine, 540 First Avenue,New York 10016, USA.‡Developmental BiologyInstitute of Marseille,Université de laMéditterannée–CentreNational de la RechercheScientifique, Campus deLuminy, Case 907, 13 288Marseille Cedex 09, France.Correspondence to A.R.A.e-mail:[email protected]: 10.1038/nrn704
NATURE REVIEWS | NEUROSCIENCE VOLUME 3 | JANUARY 2002 | 25
R E V I E W S
FIBROBLAST GROWTH FACTORS
Multifunctional factors that areinvolved in embryonicdevelopment. More than 20 FGFsand 4 FGF receptors have beendescribed. Their coordinatedactivity controls cell proliferation,migration, survival anddifferentiation. FGFs regulategrowth and morphogenesis by anearly action on regionalpatterning, and a later effect onthe growth of progenitor cells ofthe forebrain.
WNT PROTEINS
A family of highly conservedsecreted signalling moleculesthat regulate cell–cellinteractions duringembryogenesis. Wnt proteinsbind on the cell surface toreceptors of the Frizzled family.
HEDGEHOG FAMILY
A group of paracrine signallingmolecules with multiple rolesduring development. It includesHedgehog, which specifies thesegmental polarity of theblastoderm and cell fate inimaginal discs of Drosophila;Sonic hedgehog, whichparticipates in multiple aspectsof neural development invertebrates; Indian hedgehog,which participates in endodermdifferentiation and bone growth;and Desert hedgehog, which isinvolved in spermatogenesis.
brain, SHH is also involved in the generation ofoligodendrocytes34–38.
After this early period of Shh expression in the ven-tral neural tube, other brain regions also begin toexpress it. Soon after neural tube closure, Shh expressionis detected in the zona limitans intrathalamica, and bylate embryogenesis, in cerebellar Purkinje neurons,amygdala, dentate gyrus of the hippocampus, tectalplate, olfactory bulb and neocortex10,39–43 (FIG. 2).
The several functions ascribed so far to SHH — celldifferentiation, patterning, survival and prolifera-tion10,26,30,44,45 — can take place simultaneously in neigh-bouring cells. For example, SHH expression can serveas a good indicator of oligodendrogenesis in adjacentcells in the developing brain, but it might also reflectproliferative effects on neighbouring precursors. It isthis aspect of SHH signalling — the control of precur-sor proliferation — that we consider here, as we inquireinto the mechanisms that regulate brain growth.
SHH upregulate the expression of the zinc-finger tran-scription factor Gli1 (FIG. 1; reviewed in REF. 18). Thereare three Gli proteins that participate in the mediation,interpretation of or response to the SHH signal. Theseproteins act in a combinatorial fashion: Gli1 and Gli2have activating effects, whereas Gli3 (and at times Gli2)antagonizes the function of SHH–Gli1 (REFS 11,19–25).
In mammals and birds, SHH is the only Hedgehog(HH) family member that is reported to be expressedin the normal central nervous system (CNS)15.Within the brain, Shh is first expressed ventrally atearly embryonic stages, and together with SHHderived from the notochord and prechordal plate, isinvolved in ventral hindbrain, midbrain and forebraindevelopment (reviewed in REFS 12,26). The SHH–Glipathway induces different classes of neurons in theventral brain, regulates the size of the ventral mid-brain, and controls the development of the basal gan-glia19,20,27–33. Moreover, in both the spinal cord and the
Smo
Ptc1, etc.
Co-activators
Repressor Glis
a Without SHH b With SHH
Nuclearexport
Gli1, Ptc1, etc.
Ptc1, etc.
Gli1, Ptc1, etc.
Ptc1 Ptc1 Hip
Cytoplasm
Membrane
Nucleus
Cytoplasm
Membrane
Nucleus
ModifiersModifiers
Smo
SHHSHH
SHH
Su(fu)Fu
PKAX,Y,Z
Glis
Microtubules MicrotubulesSu(fu)
Fu
PKAX,Y,Z
Glis
Activator Glis
Repressor Glis Activator Glis
Proteasome
Figure 1 | SHH-triggered intracellular events leading to Gli function. a | In the absence of Sonic hedgehog (SHH), Patched 1(Ptc1) inhibits Smoothened (Smo), thereby inhibiting the downstream transduction cascade. The Gli proteins are phosphorylated byprotein kinase A (PKA), leading to their cleavage by the proteasome and the formation of carboxyl-terminus-truncated repressorGlis, which move to the nucleus and repress the Gli-dependent transcription of target genes. b | On SHH binding to the Ptc1–Smoreceptor, a macromolecular complex that is associated with the cytoskeleton, and includes Su(fu) (Suppressor of fused), Fu (Fused),PKA, the Gli proteins and other possible components (represented as X, Y, Z), acts to produce labile Gli activators. These areimported into the nucleus and transactivate target genes. The regulation of Gli activity takes place at many levels, including nuclearexport and the presence of positive or negative cofactors. Gli activity is also regulated through inhibition of SHH by the inhibitoryHedgehog-interacting factor (Hip). In addition, SHH activates the transcription of Ptc1, which, when overexpressed, attenuates theSHH response by preventing SHH from binding Ptc1 molecules within the receptor complex.

26 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro
R E V I E W S
was the nature and mode of action of the moleculesinvolved in the growth of the cerebellar cortex duringontogeny. Similarly, it is unclear what mechanisms drivedifferential cerebellar growth during evolution. As cere-bellar cell types and their relative organization are con-served across vertebrate phylogeny, the mechanismunderlying cerebellar growth might have been conservedduring evolution.
In recent years, several factors have been found toparticipate in cerebellar growth. For example, BONE
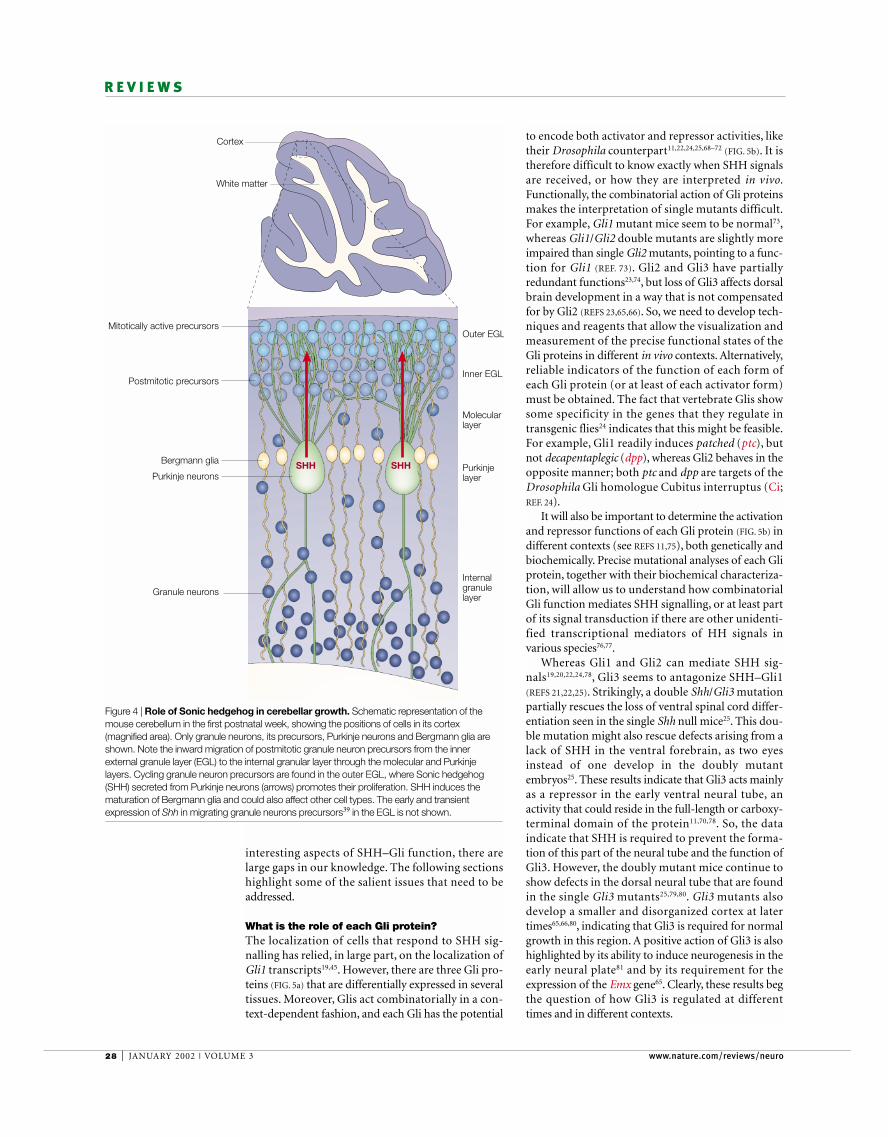
MORPHOGENETIC PROTEINS (BMPs) and an absence of SHHare required to specify embryonic cells of the rhombiclip as granule neuron precursors, which begin theirmigration after specification55. Moreover, FGFs not onlyaffect early embryonic cerebellar specification in themidbrain/hindbrain region54,56, but also late cerebellargrowth6,57. Importantly, the finding that SHH controlsgranule neuron precursor proliferation in embryo-genesis and postnatally39–41 highlighted a mechanismresponsible for modulating the proliferation of cerebel-lar cortical cells (the precursors of granule neurons) andtherefore the overall size of this structure.
Recombinant SHH causes purified granule neuronprecursors to proliferate, and inhibition of signalling in vitro results in a decrease in proliferation39 (FIG. 4).In vivo, inhibition of SHH signalling with a monoclonalantibody58 results in a marked decrease of proliferationin the external germinal layer (EGL)39,41, where granuleneuron precursors are found. Moreover, the Purkinjelayer (PL) is also disorganized in the brains of chicksinjected with cells that secrete SHH-blocking anti-bodies39. This could reflect a requirement of SHH inPurkinje neurons — the cells that produce SHH.Alternatively, this finding could also be explained by anindirect effect of the aberrant EGL, as a well-formed EGLis necessary for proper PL formation59. SHH is therefore acrucial factor in the growth of the cerebellar cortex.Moreover, its production by differentiated Purkinje neu-rons provides a mechanism by which differentiated cellsregulate the proliferation of later-differentiating neuronsat a distance, in germinative zones that express Gli1 (theEGL in this case). The fact that the proliferative effect ofSHH is antagonized by FGF2 (REF. 41) highlights theinvolvement of several pathways in the precise control ofprecursor proliferation.
Are the mechanisms that control cerebellar size alsoinvolved in controlling its characteristic foliated shape?Control of size could imply control of shape if the mech-anisms involved in modulating precursor-cell prolifera-tion show local variations. The idea that differentialSHH signalling could be responsible for the generationof the pattern of the cerebellum39 is in line with the abilityof SHH to re-specify early ventral midbrain pattern andsize33,60, but is in contrast to the view that physical forcesare the primary mediators of foliation52. Differential sig-nalling by SHH and other factors across the cerebellarcortex could similarly account for the generation of thedifferent cerebellar foliation patterns seen in variousspecies. It is possible that the response to SHH signallingis modulated by other factors, such as FGF2 (REF. 41),that normally restrict the duration of SHH effects, or,
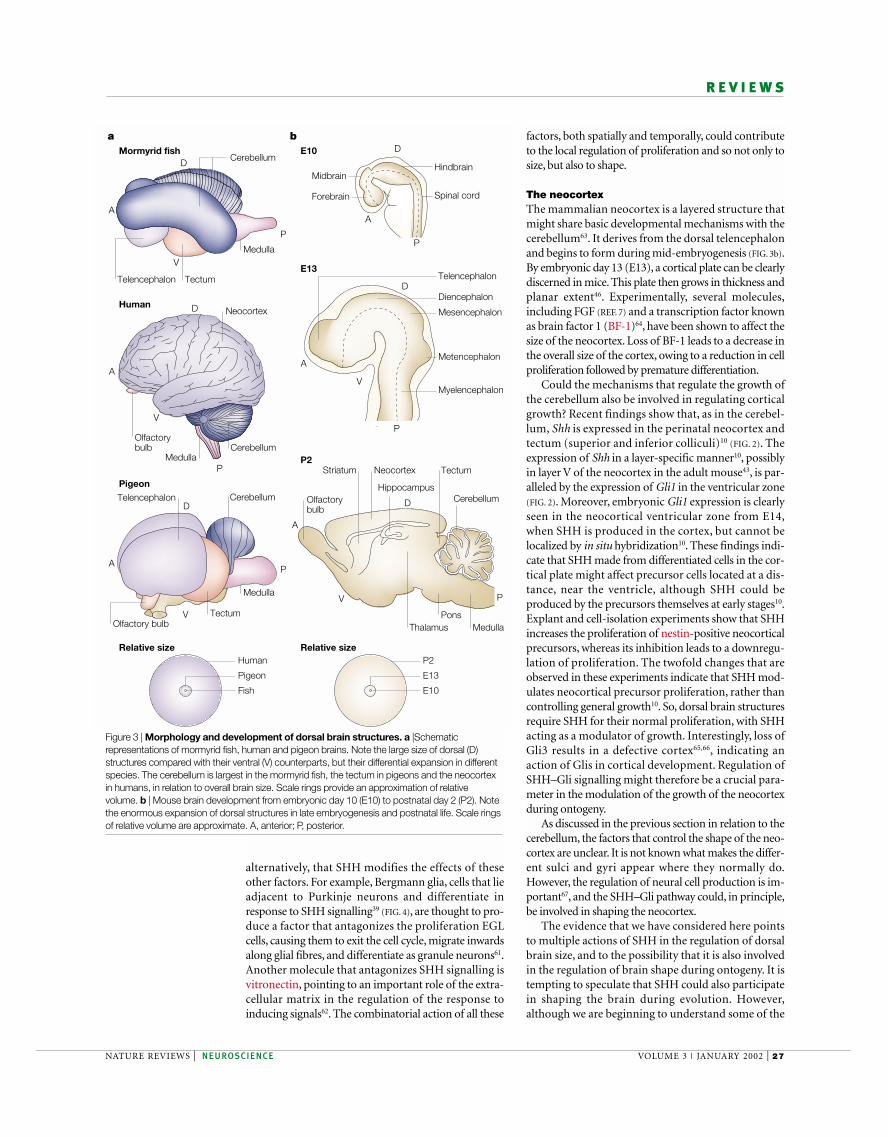
The dorsal brainThe development of the dorsal aspect of the brainachieves clear prominence towards mid-embryogenesisin mammals and birds. At this time, the three main lay-ered structures of the dorsal brain — the cerebral cortex,the cerebellum and the tectum (superior and inferiorcolliculi) — undergo rapid expansion46. The end result isthat these dorsal structures are larger than their ventralcounterparts. For example, most of the brain in humansis comprised of the dorsal cerebral cortex (FIG. 3a). Bycontrast, electroreceptive mormyrid fish have anenlarged cerebellum, and, in some birds, it is the optictectum that undergoes profound early growth (FIG. 3a).There seems to be a trend towards bigger dorsal struc-tures in relation to overall brain size during evolution,compared with, for example, the size of the striatum47,but the relative size of each of the dorsal structures isregulated independently in different animal groups48.
How is the size of dorsal brain structures regulatedwithin a species? Analyses of lineages49,50 and prolifera-tion parameters8,9,51 of precursor cells in the cortex indi-cate that the behaviour of these precursors, some ofwhich are stem cells (that can self-renew and give rise todifferentiated progeny), will determine the final numberof mature neurons and glia, and therefore determine thesize of the cortex. The question then becomes: what arethe molecular mechanisms that regulate precursorbehaviour? Here we focus on two dorsal structures —the cerebellum and the cortex — in relation to theSHH–Gli signalling pathway.
The cerebellumA large amount of work has been published on the cere-bellum52, starting with the initial detailed descriptions ofits nerve cells. We will not attempt to summarize it here,referring the reader instead to recent reviews53,54. It is fairto say, however, that despite all the cellular, histologicaland genetic work on the cerebellum, little was known,until recently, about the molecules that make the cere-bellum grow. Because the most significant growth of thecerebellum occurs in its cortex (the deep nuclei consti-tute a relatively small part), what remained unknown
BONE MORPHOGENETIC
PROTEINS
Multifunctional secretedproteins of the transforminggrowth factor-β superfamily. Inthe early embryo, theyparticipate in dorsoventralpatterning.
Medulla Spinal cord
Thalamus
Pretectum
Hippocampus
Neocortex
Olfactorybulb
Hypothalamus
Tegmentum
Mouse P0
Cerebellum
Inferior colliculus
Superior colliculus
Striatum
VZ/SVZ
VZ/SVZEGLPL
Figure 2 | Layer-specific expression of SHH in dorsal regions. A sagittal section through apostnatal day 0 (P0) mouse brain. Arrows represent the proposed action of Sonic hedgehog(SHH), at a distance from its source. SHH-producing cells are located in the neocortical and tectalplates, and in the Purkinje layer (PL). Responding cells are located in the ventricular/subventricularzones (VZ/SVZ) of the neocortex and tectum, and in the external germinal layer (EGL) in thecerebellum. Ventral sources and actions of SHH are not shown.
NATURE REVIEWS | NEUROSCIENCE VOLUME 3 | JANUARY 2002 | 27
R E V I E W S
factors, both spatially and temporally, could contributeto the local regulation of proliferation and so not only tosize, but also to shape.
The neocortexThe mammalian neocortex is a layered structure thatmight share basic developmental mechanisms with thecerebellum63. It derives from the dorsal telencephalonand begins to form during mid-embryogenesis (FIG. 3b).By embryonic day 13 (E13), a cortical plate can be clearlydiscerned in mice. This plate then grows in thickness andplanar extent46. Experimentally, several molecules,including FGF (REF. 7) and a transcription factor knownas brain factor 1 (BF-1)64, have been shown to affect thesize of the neocortex. Loss of BF-1 leads to a decrease inthe overall size of the cortex, owing to a reduction in cellproliferation followed by premature differentiation.
Could the mechanisms that regulate the growth ofthe cerebellum also be involved in regulating corticalgrowth? Recent findings show that, as in the cerebel-lum, Shh is expressed in the perinatal neocortex andtectum (superior and inferior colliculi)10 (FIG. 2). Theexpression of Shh in a layer-specific manner10, possiblyin layer V of the neocortex in the adult mouse43, is par-alleled by the expression of Gli1 in the ventricular zone(FIG. 2). Moreover, embryonic Gli1 expression is clearlyseen in the neocortical ventricular zone from E14,when SHH is produced in the cortex, but cannot belocalized by in situ hybridization10. These findings indi-cate that SHH made from differentiated cells in the cor-tical plate might affect precursor cells located at a dis-tance, near the ventricle, although SHH could beproduced by the precursors themselves at early stages10.Explant and cell-isolation experiments show that SHHincreases the proliferation of nestin-positive neocorticalprecursors, whereas its inhibition leads to a downregu-lation of proliferation. The twofold changes that areobserved in these experiments indicate that SHH mod-ulates neocortical precursor proliferation, rather thancontrolling general growth10. So, dorsal brain structuresrequire SHH for their normal proliferation, with SHHacting as a modulator of growth. Interestingly, loss ofGli3 results in a defective cortex65,66, indicating anaction of Glis in cortical development. Regulation ofSHH–Gli signalling might therefore be a crucial para-meter in the modulation of the growth of the neocortexduring ontogeny.
As discussed in the previous section in relation to thecerebellum, the factors that control the shape of the neo-cortex are unclear. It is not known what makes the differ-ent sulci and gyri appear where they normally do.However, the regulation of neural cell production is im-portant67, and the SHH–Gli pathway could, in principle,be involved in shaping the neocortex.
The evidence that we have considered here pointsto multiple actions of SHH in the regulation of dorsalbrain size, and to the possibility that it is also involvedin the regulation of brain shape during ontogeny. It istempting to speculate that SHH could also participatein shaping the brain during evolution. However,although we are beginning to understand some of the
alternatively, that SHH modifies the effects of theseother factors. For example, Bergmann glia, cells that lieadjacent to Purkinje neurons and differentiate inresponse to SHH signalling39 (FIG. 4), are thought to pro-duce a factor that antagonizes the proliferation EGLcells, causing them to exit the cell cycle, migrate inwardsalong glial fibres, and differentiate as granule neurons61.Another molecule that antagonizes SHH signalling isvitronectin, pointing to an important role of the extra-cellular matrix in the regulation of the response toinducing signals62. The combinatorial action of all these
CerebellumMedulla
Cerebellum
Neocortex
Relative size Relative size
Medulla
Medulla
Cerebellum
Human
Human
Mormyrid fish
Fish
a b
Pigeon
Pigeon
Midbrain
E10
E13
P2
Hindbrain
Spinal cord
Telencephalon
Metencephalon
Cerebellum
MedullaThalamus
Tectum
Olfactory bulb
Olfactory bulb
NeocortexStriatum
Pons
Hippocampus
Forebrain
Telencephalon
A
A
A
A
A
A
P
P
P
P
P
P
D
D
DD
D
D
V
V
V
V
V
Telencephalon
Tectum
Tectum
Olfactorybulb
P2
E10
E13
Diencephalon
Mesencephalon
Myelencephalon
Figure 3 | Morphology and development of dorsal brain structures. a |Schematicrepresentations of mormyrid fish, human and pigeon brains. Note the large size of dorsal (D)structures compared with their ventral (V) counterparts, but their differential expansion in differentspecies. The cerebellum is largest in the mormyrid fish, the tectum in pigeons and the neocortexin humans, in relation to overall brain size. Scale rings provide an approximation of relativevolume. b | Mouse brain development from embryonic day 10 (E10) to postnatal day 2 (P2). Notethe enormous expansion of dorsal structures in late embryogenesis and postnatal life. Scale ringsof relative volume are approximate. A, anterior; P, posterior.
28 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro
R E V I E W S
to encode both activator and repressor activities, liketheir Drosophila counterpart11,22,24,25,68–72 (FIG. 5b). It istherefore difficult to know exactly when SHH signalsare received, or how they are interpreted in vivo.Functionally, the combinatorial action of Gli proteinsmakes the interpretation of single mutants difficult.For example, Gli1 mutant mice seem to be normal73,whereas Gli1/Gli2 double mutants are slightly moreimpaired than single Gli2 mutants, pointing to a func-tion for Gli1 (REF. 73). Gli2 and Gli3 have partiallyredundant functions23,74, but loss of Gli3 affects dorsalbrain development in a way that is not compensatedfor by Gli2 (REFS 23,65,66). So, we need to develop tech-niques and reagents that allow the visualization andmeasurement of the precise functional states of theGli proteins in different in vivo contexts. Alternatively,reliable indicators of the function of each form ofeach Gli protein (or at least of each activator form)must be obtained. The fact that vertebrate Glis showsome specificity in the genes that they regulate intransgenic flies24 indicates that this might be feasible.For example, Gli1 readily induces patched (ptc), butnot decapentaplegic (dpp), whereas Gli2 behaves in theopposite manner; both ptc and dpp are targets of theDrosophila Gli homologue Cubitus interruptus (Ci;REF. 24).
It will also be important to determine the activationand repressor functions of each Gli protein (FIG. 5b) indifferent contexts (see REFS 11,75), both genetically andbiochemically. Precise mutational analyses of each Gliprotein, together with their biochemical characteriza-tion, will allow us to understand how combinatorialGli function mediates SHH signalling, or at least partof its signal transduction if there are other unidenti-fied transcriptional mediators of HH signals in various species76,77.
Whereas Gli1 and Gli2 can mediate SHH sig-nals19,20,22,24,78, Gli3 seems to antagonize SHH–Gli1(REFS 21,22,25). Strikingly, a double Shh/Gli3 mutationpartially rescues the loss of ventral spinal cord differ-entiation seen in the single Shh null mice25. This dou-ble mutation might also rescue defects arising from alack of SHH in the ventral forebrain, as two eyesinstead of one develop in the doubly mutantembryos25. These results indicate that Gli3 acts mainlyas a repressor in the early ventral neural tube, anactivity that could reside in the full-length or carboxy-terminal domain of the protein11,70,78. So, the dataindicate that SHH is required to prevent the forma-tion of this part of the neural tube and the function ofGli3. However, the doubly mutant mice continue toshow defects in the dorsal neural tube that are foundin the single Gli3 mutants25,79,80. Gli3 mutants alsodevelop a smaller and disorganized cortex at latertimes65,66,80, indicating that Gli3 is required for normalgrowth in this region. A positive action of Gli3 is alsohighlighted by its ability to induce neurogenesis in theearly neural plate81 and by its requirement for theexpression of the Emx gene65. Clearly, these results begthe question of how Gli3 is regulated at differenttimes and in different contexts.
interesting aspects of SHH–Gli function, there arelarge gaps in our knowledge. The following sectionshighlight some of the salient issues that need to beaddressed.
What is the role of each Gli protein?The localization of cells that respond to SHH sig-nalling has relied, in large part, on the localization ofGli1 transcripts19,45. However, there are three Gli pro-teins (FIG. 5a) that are differentially expressed in severaltissues. Moreover, Glis act combinatorially in a con-text-dependent fashion, and each Gli has the potential
c
Granule neurons
Bergmann glia
Mitotically active precursors
SHH
Outer EGL
Inner EGL
Purkinje layer
Molecular layer
White matter
Cortex
Internalgranulelayer
Postmitotic precursors
Purkinje neuronsSHH
Figure 4 | Role of Sonic hedgehog in cerebellar growth. Schematic representation of themouse cerebellum in the first postnatal week, showing the positions of cells in its cortex(magnified area). Only granule neurons, its precursors, Purkinje neurons and Bergmann glia areshown. Note the inward migration of postmitotic granule neuron precursors from the innerexternal granule layer (EGL) to the internal granular layer through the molecular and Purkinjelayers. Cycling granule neuron precursors are found in the outer EGL, where Sonic hedgehog(SHH) secreted from Purkinje neurons (arrows) promotes their proliferation. SHH induces thematuration of Bergmann glia and could also affect other cell types. The early and transientexpression of Shh in migrating granule neurons precursors39 in the EGL is not shown.

NATURE REVIEWS | NEUROSCIENCE VOLUME 3 | JANUARY 2002 | 29
R E V I E W S
How does SHH travel? Although SHH is transiently detected in EGL cells, andmight be expressed by early neocortical precursors,proliferation of later cells in the dorsal brain seems todepend on SHH that is secreted by differentiated cells.In the cerebellum, tectum and neocortex, SHH issecreted from postmitotic cells located at a distancefrom the responding Gli1-positive cycling precursors.The distance from the somata of the secreting cells —say, the Purkinje neurons — to the responding precur-sors in the EGL is more than ten cell diameters. SHHcould therefore be transported or diffuse from its siteof production to its site of action. Studies in flies and inother parts of the vertebrate nervous system have shedsome light on the mechanisms that regulate SHHactions at a distance from its source.
In principle, SHH could freely diffuse from thePurkinje soma or from cortical plate neurons to reachthe proliferating precursors89,90. Indeed, SHH hasrecently been visualized across a wide area, away fromits site of production91. Consistent with this observa-tion, HHs can act as morphogens over considerable dis-tances90,92–99. However, transport of HH in flies dependson the activity of several genes, including tout-velu (ttv)and dispatched (disp)100,101. Moreover, mature SHH isnormally tethered to the cytoplasmic membrane by acholesterol moiety17, and the transport of vesicles thatcontain the ligand and receptor complex is con-trolled102–105. HHs could therefore reach their desti-nation by acting as morphogens, the secretion andtransport of which is highly regulated. Indeed, SHHitself is required at a distance from its source93–97.
Do SHH concentration thresholds for proliferationexist in the brain ? Within the cerebellar EGL or the cor-tical ventricular/subventricular zone, the cells furthestfrom the source of SHH (for example, the outer EGL)show the highest mitogenic activity (FIG. 4). By contrast,cells in the inner EGL cease to cycle and begin their pro-gramme of differentiation. Although it is probable thatthis reflects the action of antagonizing signals close tothe inner EGL, it would be interesting to determinewhether different concentrations of SHH within theproliferative domain have different instructive effects.
Alternatively, SHH could act at its site of production,never leaving the membrane of the producing cell. Inthis case, Purkinje and cortical plate neurons might sig-nal through dendrites that reach the EGL and the ven-tricular zone, respectively. This is conceivable, as SHH istransported through retinal ganglion axons106–108.Alternatively, SHH might reach its target cells throughvery fine cellular protrusions known as cytonemes109.Such cytonemes could, in principle, reach targets situatedfar away from the soma of the producing cell.
SHH signalling and other factorsThe effects of several brain factors can vary dependingon the context. In some cases, different factors actingsimultaneously can have antagonistic or cooperativeeffects32,41,55,110. How does a brain precursor integratethese signals? Much is known about how BMPs affectdorsal neural tube patterning111, about how SHH
How does SHH affect the cell cycle?SHH increases the proliferation of responsive neuralprecursors. It is not yet completely clear how this isachieved. Its action could involve a shortening of thecell cycle, but also the induction of resting precursors tore-enter it. In primary cultures from neonatal mousecerebellum, SHH acts to maintain phosphorylation ofthe retinoblastoma protein (Rb) in a subset of granulecell precursors; however, it cannot recruit quiescentcells into the cell cycle after arrest82. It is clear that SHHsignalling can affect several cell-cycle parameters (FIG. 6),enhancing proliferation of brain and spinal cord pre-cursors10,44,58. We know that SHH signalling upregulatesthe transcription of G1-phase CYCLINS of types D and E(REF. 82; R. Tikoo, N.D., M. Chao and A.R.A., unpub-lished data). Cyclin D2 mutants have smaller cerebellaand show a loss of granule cells83. Ptc1 also seems to beable to act directly on G2-phase cyclin B (REF. 84). More-over, SHH can inhibit growth arrest by the cell-cycleinhibitor p21 in epithelial cells85, but not by p27 ingranule cell precursors of the cerebellum86. Induction ofmotor neuron cell fate by SHH seems to depend on twodistinct periods, one late in the S phase58. Multipleeffects of SHH signalling on cyclin-dependent kinases(Cdks), Cdk inhibitors, cyclins or the transcription fac-tor E2F at different points in the cell cycle could account,in principle, for its proliferative effects87. Resolving howSHH–Gli signalling affects the cell cycle at various stagesis crucial to understanding how two neighbouring cellsinterpret the same signal in opposite ways, either as pro-liferative or as differentiative information. Interestingly,SHH might function in the self-renewal process ofhaematopoietic stem cells88, raising the question ofwhether this is also the case in the CNS.
Activators
Repressors
a b
Wnts
Activators
Repressors
Activators
Repressors ?
SHH
?
?
SHH
Gli3Gli1
FGFGli2
Gli3
Gli2
Gli1
Figure 5 | Interactions between secreted signalling molecules and Gli function. a | Sonichedgehog (SHH) can induce Gli1 and Gli2 transcription, whereas it represses Gli3. Gli2 mightmediate the induction of Gli1 by SHH. Fibroblast growth factor (FGF) signalling can induce thetranscription of Gli2 and Gli3, and all three Gli proteins can induce Wnt transcription. Gli1/SHHand Gli3 antagonize each other. b | SHH affects the choice of repressor or activator Gli function.SHH signalling can repress the formation of Gli3 repressors, and influence the formation of Gli2and perhaps Gli1 activators.
CYCLINS
A family of proteins, the levels ofwhich fluctuate throughout thecell cycle. By activating cyclin-dependent kinases, they help toregulate several stages of celldivision.

30 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro
R E V I E W S
Later on, during corticogenesis, SHH acts as prolifer-ative agent10, and BMPs could counteract this effect byacting as differentiation-inducing, anti-proliferative fac-tors118–120. Intriguing possibilities are that Gli2/3 coulddirectly regulate BMP expression121, and that BMPscould induce the expression of Rab23, a negative regula-tor of the SHH pathway104. Clearly, more research isneeded to determine how these signalling pathways actand interact in the brain.
A similar situation occurs with FGFs. SHH cansometimes cooperate with FGF signalling; for example,in the induction of dopaminergic neurons in the ven-tral forebrain32 and in neurosphere proliferation118. Bycontrast, FGF2 signalling can inhibit the proliferativeaction of SHH in the cerebellum41. However, the mech-anisms by which FGFs and HHs interact remainunclear. Unexpectedly, Gli2 and Gli3 were found to beresponsive to FGF (FIG. 5a) and to participate in theFGF–brachyury regulatory loop that is involved in earlymesoderm formation122. It is therefore possible that thecombinatorial function of Glis is a potential site of sig-nal integration in cells that respond to both FGF andSHH. Such a relationship might be particularly rele-vant in the germinative zones of the brain, where bothFGF and SHH affect precursor behaviour downstreamof signal transduction: the effects of SHH on the cellcycle seem to be independent of mitogen-activated pro-tein kinase (MAPK) activity, whereas FGF signallingactivates MAPK82.
The Wnt family of proteins has been implicated inmany aspects of animal development. In the brain, dor-sal cells within the cortical hem require Wnts123. Fromhere, these proteins influence the development of thehippocampus124 and possibly the cortex, where someWnts are expressed in a nested fashion125,126. Moreover,some Wnts are targets and mediators of Gli function127,raising the possibility that some of the effects on brainprecursors that are attributable to Gli-, SHH- and pos-sibly FGF-mediated signalling are mediated by combi-nations of Wnt proteins127. Interestingly, inactivation ofβ-catenin, a mediator of Wnt signalling, results inabnormal brain formation128, and Wnts have beenimplicated in hippocampal cell proliferation124.
We should also consider the effects of direct inhibitorsof the SHH signalling pathway (FIG. 1). On SHH sig-nalling, a membrane protein known as Hedgehog-interacting factor (Hip) is produced, which attenuatesthe strength of the signal129. Similarly, Ptc1 transcrip-tion is increased in response to SHH. Ptc1 not onlyacts in the receptor complex to inhibit the action ofSmo, but can also act as a sponge for SHH, preventingit from reaching cells further away93. Increases in Hipand Ptc1 expression are ways in which a cell couldattenuate the response to SHH, allowing other signalsto act instead.
Other factors that need to be examined further inrelation to their interaction with the SHH pathwayinclude IGFs, neuregulins, pituitary-adenylyl-cyclase-activating polypeptide (PACAP), vitronectin, Notchand growth-arrest-specific protein 1 (Gas1). IGFsmight mediate growth regulation by SHH, as Igf2 is a
signalling affects ventral patterning26,97, and about thefact that these pathways antagonize the effect of eachother. However, little is known about the mechanismsthat integrate their inputs directly, although it has beenshown that SHH indirectly activates the expression ofBMP antagonists112–115. Interactions between SHH andBMPs might also occur at the level of transcriptionalcross-regulation, by the modulation of shared signallingpathways, and/or at the level of common target genes.
Gli repressors, truncated at their carboxyl terminus,bind inactive SMADS (REF. 116), the transcription factorsthat mediate BMP signals. However, the presence of Glirepressors did not affect the Smad-dependent tran-scriptional activation of reporter genes. Moreover, theGli repressor/Smad complex dissociated on ligand acti-vation of the BMP receptors and Smad phosphoryla-tion. As we lack methods to test for coordinating mech-anisms, we can only imagine that a possible function ofsuch mechanisms would be to act as switches fromSHH to BMP responsiveness. For example, SHH sig-nalling could proceed by activating Glis, inhibiting newrepressor formation and inducing labile activa-tors11,68,71,117. Any remaining Gli repressors could beinactivated through sequestration by inactive Smads.On BMP signalling, the Gli repressor/Smad complexwould dissociate, allowing Smads to activate BMP-responsive targets, and Gli repressors to begin to inhibitSHH targets.
SMADS
A family of transcription factorsthat mediate transforminggrowth factor-β signals. Theterm SMAD is derived from thefounding members of thisfamily, the Drosophila proteinMad (Mothers againstDecapentaplegic) and theCaenorhabditis elegans proteinSMA (small body size).
Cyclin D
Cdc2
Cyclin B
Cdc2
Cyclin A
Cdc2
Cyclin A
G0
G2 G1
S
M
?
E2F
p21
p27
Rb
Rb P
Cyclin B Ptc1SHH
Cdk4/6
Cyclin D
Cdk2
Cyclin E
Cyclin E
E2F
SHH
SHH
SHH
Figure 6 | SHH signalling and the cell cycle. Sonic hedgehog (SHH) seems to regulateprogression into late G1 phase. SHH induces the expression of D- and E-type cyclins, whichactivate cyclin-dependent kinases (Cdk4/6 and Cdk2, respectively). These Cdks promote thephosphorylation of retinoblastoma protein (Rb), causing it to dissociate from the transcriptionfactor E2F, which can then act to promote cycle progression. SHH also inhibits p21, an inhibitorof Cdk activity, which leads to a potentiation of Rb phosphorylation. SHH acts in late S phase insome precursors. Patched 1 (Ptc1) can normally inhibit cyclin B. On SHH signalling, inhibition ofPtc1 function could free cyclin B, allowing it to stimulate the cell to progress from G2 to M phase.Other points of action are possible.

NATURE REVIEWS | NEUROSCIENCE VOLUME 3 | JANUARY 2002 | 31
R E V I E W S
What is the role of SHH in the adult brain?The fact that neurogenesis and gliogenesis continue inthe adult mammalian brain50 might explain why SHHis expressed in some sites of the mature brain. However,SHH continues to be expressed in the cortex and in thegranule layer of the cerebellum, where little or no pro-liferation occurs. The prominent expression of genes ofthe SHH pathway in the adult brain (REFS 42,43; N.D.and A.R.A., unpublished observations) raises the ques-tion of whether SHH has survival roles in the brain, asit does in the spinal cord30, as well as additional func-tions. Although only the study of conditional mutantsin mice will begin to address this issue, it is tempting toconsider whether SHH could also have a role in experi-ence-dependent remodelling. It could also function inrepair mechanisms after brain injury, in addition tobeing involved in brain tumorigenesis10,39–41,140. Clearly,HHs and Glis hold secrets for understanding how thebrain develops and grows, and it might not be unrea-sonable to think that normal brain function involvestheir participation.
target of this pathway130, and FGF2 and IGF1 act syner-gistically to stimulate oligodendrocyte progenitors (acell type induced by SHH) to enter the cell cycle131.Neuregulins can regulate precursor growth132. PACAPinhibits the proliferation of cortical precursors133 and,as PACAP upregulates cyclic AMP levels, could coun-teract the effects of SHH signalling through proteinkinase A (PKA). Indeed, PACAP represses Gli1 tran-scription134. By contrast, vitronectin inhibits theresponse of EGL cerebellar precursors to SHH throughMAPK activity and phosphorylation of the cAMP-responsive-element-binding protein CREB62, but actssynergistically with SHH in motor neuron inductionin the early ventral neural tube135. Gas1 is a glyco-sylphosphatidylinositol (GPI)-linked protein thatinteracts with SHH and could antagonize its effects insome contexts136,137. In addition, BF-1 could interactwith SHH signalling, as loss of the former inhibitsexpression of the latter138. Last, SHH might affect Notchsignalling, which promotes granule neuron precursorproliferation by inducing His expression139.
1. Qian, X., Davis, A. A., Goderie, S. K. & Temple, S. FGF2concentration regulates the generation of neurons and gliafrom multipotent cortical stem cells. Neuron 18, 81–93(1997).
2. Qian, X., Goderie, S. K., Shen, Q., Stern, J. H. & Temple, S.Intrinsic programs of patterned cell lineages in isolatedvertebrate CNS ventricular zone cells. Development 125,3143–3152 (1998).
3. Raballo, R. et al. Basic fibroblast growth factor (Fgf2) isnecessary for cell proliferation and neurogenesis in thedeveloping cerebral cortex. J. Neurosci. 20, 5012–5023(2000).
4. Lillien, L. & Raphael, H. BMP and FGF regulate thedevelopment of EGF-responsive neural progenitor cells.Development 127, 4993–5005 (2000).
5. Tropepe, V. et al. Distinct neural stem cells proliferate inresponse to EGF and FGF in the developing mousetelencephalon. Dev. Biol. 208, 166–188 (1999).
6. Cheng, Y., Tao, Y., Black, I. B. & DiCicco-Bloom, E. A singleperipheral injection of basic fibroblast growth factor (bFGF)stimulates granule cell production and increases cerebellargrowth in newborn rats. J. Neurobiol. 46, 220–229 (2001).Shows the effects of FGF on cerebellar growth.
7. Vaccarino, F. M. et al. Changes in cerebral cortex size aregoverned by fibroblast growth factor during embryogenesis.Nature Neurosci. 2, 246–253 (1999).Shows that FGF signalling has an important role in thegrowth of the cortex.
8. Rakic, P. A small step for the cell, a giant leap for mankind: ahypothesis of neocortical expansion during evolution. TrendsNeurosci. 18, 383–388 (1995).
9. Rakic, P. Radial unit hypothesis of neocortical expansion.Novartis Found. Symp. 228, 30–42 (2000).
10. Dahmane, N. et al. The Shh–Gli pathway modulates thenormal and abnormal growth of the dorsal brain.Development 128 (in the press).Provides evidence for a role of SHH signalling incontrolling the growth of the dorsal brain, and theinitiation and possible maintenance of braintumorigenesis, including gliomagenesis.
11. Ruiz i Altaba, A. Gli proteins encode context-dependentpositive and negative functions: implications for developmentand disease. Development 126, 3205–3216 (1999).Indicates activator and repressor functions ofvertebrate Glis.
12. Ruiz i Altaba, A. Gli proteins and Hedgehog signaling:development and cancer. Trends Genet. 15, 418–425(1999).
13. Riddle, R. D., Johnson, R. L., Laufer, E. & Tabin, C. J. Sonichedgehog mediates the polarizing activity of the ZPA. Cell 75, 1401–1416 (1993).Reports the first insights into HH signalling invertebrates (see also references 14–16).
14. Krauss, S., Concordet, J. P. & Ingham, P. W. A functionallyconserved homolog of the Drosophila segment polarity
gene hh is expressed in tissues with polarizing activity inzebrafish embryos. Cell 75, 1431–1444 (1993).
15. Echelard, Y. et al. Sonic hedgehog, a member of a family ofputative signaling molecules, is implicated in the regulationof CNS polarity. Cell 75, 1417–1430 (1993).
16. Roelink, H. et al. Floor plate and motor neuron induction byvhh-1, a vertebrate homolog of Hedgehog expressed by thenotochord. Cell 76, 761–775 (1994).
17. Porter, J. A., Young, K. E. & Beachy, P. A. Cholesterolmodification of Hedgehog signaling proteins in animaldevelopment. Science 274, 255–259 (1996).
18. Ruiz i Altaba, A. The works of Gli and the power ofHedgehog. Nature Cell Biol. 1, 147–148 (1999).
19. Lee, J., Platt, K. A., Censullo, P. & Ruiz i Altaba, A. Gli1 is atarget of Sonic hedgehog that induces ventral neural tubedevelopment. Development 124, 2537–2552 (1997).
20. Hynes, M. et al. Control of cell pattern in the neural tube bythe zinc finger transcription factor and oncogene Gli-1.Neuron 19, 15–26 (1997).
21. Ruiz i Altaba, A. Catching a Gli-mpse of Hedgehog. Cell 90,193–196 (1997).
22. Ruiz i Altaba, A. Combinatorial Gli gene function in floor plateand neuronal inductions by Sonic hedgehog. Development125, 2203–2212 (1998).Elaborates on the regulation of Gli transcription bySHH signalling, showing an antagonism betweenSHH/Gli1 and Gli3, as well as providing evidence forthe combinatorial and context-dependent action ofthe Gli proteins.
23. Mo, R. et al. Specific and redundant functions of Gli2 andGli3 zinc finger genes in skeletal patterning anddevelopment. Development 124, 113–123 (1997).Genetic evidence that Gli2 and Gli3 have partiallyoverlapping functions in mice.
24. Aza-Blanc, P., Lin, H. Y., Ruiz i Altaba, A. & Kornberg, T. B.Expression of the vertebrate Gli proteins in Drosophilareveals a distribution of activator and repressor activities.Development 127, 4293–4301 (2000).Shows that vertebrate Gli proteins have specificfunctions in vivo; also shows the differential regulationof repressor and activator functions by HH signallingin flies.
25. Litingtung, Y. & Chiang, C. Specification of ventral neurontypes is mediated by an antagonistic interaction betweenShh and Gli3. Nature Neurosci. 3, 979–985 (2000).Reports the rescue of aspects of ventral differentiationin double SHH/Gli3 mutants, indicating that SHH actsat least in part by repressing Gli3 function.
26. Jessell, T. M. Neuronal specification in the spinal cord:inductive signals and transcriptional codes. Nature Rev.Genet. 1, 20–29 (2000).A superb and accessible review on a decade of ground-breaking work on cell specification in the spinal cord.
27. Hynes, M. et al. Induction of midbrain dopaminergic neuronsby Sonic hedgehog. Neuron 15, 35–44 (1995).
28. Chiang, C. et al. Cyclopia and defective axial patterning inmice lacking Sonic hedgehog gene function. Nature 383,407–413 (1996).A genetic demonstration that SHH is required fornormal ventral CNS development, as well as for thedevelopment of many other organs and tissues.
29. Ericson, J., Muhr, J., Jessell, T. M. & Edlund, T. Sonichedgehog: a common signal for ventral patterning along therostrocaudal axis of the neural tube. Int. J. Dev. Biol. 39,809–816 (1995).
30. Miao, N. et al. Sonic hedgehog promotes the survival of specific CNS neuron populations and protects these cells from toxic insult in vitro. J. Neurosci. 17, 5891–5899(1997).
31. Kohtz, J. D., Baker, D. P., Corte, G. & Fishell, G.Regionalization within the mammalian telencephalon ismediated by changes in responsiveness to SonicHedgehog. Development 125, 5079–5089 (1998).
32. Ye, W., Shimamura, K., Rubenstein, J. L., Hynes, M. A. &Rosenthal, A. FGF and Shh signals control dopaminergicand serotonergic cell fate in the anterior neural plate. Cell 93,755–766 (1998).
33. Agarwala, S., Sanders, T. A. & Ragsdale, C. W. SonicHedgehog control of size and shape in midbrain patternformation. Science 291, 2147–2150 (2001).Shows that early ectopic SHH can ventralize andrepattern the dorsal midbrain.
34. Poncet, C. et al. Induction of oligodendrocyte progenitors inthe trunk neural tube by ventralizing signals: effects ofnotochord and floor plate grafts, and of sonic hedgehog.Mech. Dev. 60, 13–32 (1996).
35. Pringle, N. P. et al. Determination of neuroepithelial cell fate:induction of the oligodendrocyte lineage by ventral midlinecells and sonic hedgehog. Dev. Biol. 177, 30–42 (1996).
36. Orentas, D. M., Hayes, J. E., Dyer, K. L. & Miller, R. H. Sonichedgehog signaling is required during the appearance ofspinal cord oligodendrocyte precursors. Development 126,2419–2429 (1999).
37. Lu, Q. R. et al. Sonic hedgehog-regulated oligodendrocytelineage genes encoding bHLH proteins in the mammaliancentral nervous system. Neuron 25, 317–329 (2000).
38. Soula, C. et al. Distinct sites of origin of oligodendrocytesand somatic motoneurons in the chick spinal cord:oligodendrocytes arise from Nkx2.2-expressing progenitorsby a Shh-dependent mechanism. Development 128,1369–1379 (2001).
39. Dahmane, N. & Ruiz i Altaba, A. Sonic hedgehog regulatesthe growth and patterning of the cerebellum. Development126, 3089–3100 (1999).Shows the requirement of SHH signalling in thedevelopment and growth of the cerebellum, providingthe basis for a model of morphological plasticity ofthis structure. Also provides a rational cellular contextfor the development of medulloblastomas (see alsoreferences 40 and 41).

32 | JANUARY 2002 | VOLUME 3 www.nature.com/reviews/neuro
R E V I E W S
40. Wallace, V. A. Purkinje-cell-derived Sonic hedgehogregulates granule neuron precursor cell proliferation in thedeveloping mouse cerebellum. Curr. Biol. 9, 445–448 (1999).
41. Wechsler-Reya, R. J. & Scott, M. P. Control of neuronalprecursor proliferation in the cerebellum by Sonic hedgehog.Neuron 22, 103–114 (1999).
42. Traiffort, E., Charytoniuk, D. A., Faure, H. & Ruat, M.Regional distribution of sonic sedgehog, patched, andsmoothened mRNA in the adult rat brain. J. Neurochem.70, 1327–1330 (1998).This work indicates the presence of SHH signalling inthe mature brain.
43. Traiffort E. et al. Discrete localizations of hedgehog signallingcomponents in the developing and adult rat nervoussystem. Eur. J. Neurosci. 11, 3199–3214 (1999).
44. Rowitch, D. H. et al. Sonic hedgehog regulates proliferationand inhibits differentiation of CNS precursor cells. J. Neurosci. 19, 8954–8965 (1999).Reports the early role of SHH as a mitogen in theembryonic spinal cord.
45. Dahmane, N., Lee, J., Robins, P., Heller, P. & Ruiz i Altaba, A.Activation of the transcription factor Gli1 and the Sonichedgehog signalling pathway in skin tumours. Nature 389,876–881 (1997).Shows the ability of Gli1 to induce tumours in vivo,and the expression of GLI1 in virtually all sporadichuman basal cell carcinomas (BCCs), the mostcommon human cancer. This work led to the proposalthat all sporadic human BCCs derive from activationof the SHH–GLI pathway, providing a basis for thistumour type and a potential rational therapy.
46. Bayer, S. A. & Altman, J. Neocortical Development(Raven Press, New York, 1991).
47. Clark, D. A., Mitra, P. P. & Wang, S. S. Scalable architecturein mammalian brains. Nature 411, 189–193 (2001).
48. Kornack, D. R. Neurogenesis and the evolution of corticaldiversity: mode, tempo, and partitioning duringdevelopment and persistence in adulthood. Brain Behav.Evol. 55, 336–344 (2000).
49. Temple, S. Stem cell plasticity — building the brain of ourdreams. Nature Rev. Neurosci. 2, 513–520 (2001).
50. Alvarez-Buylla, A., Garcia-Verdugo, J. M. & Tramontin, A. D.A unified hypothesis on the lineage of neural stem cells.Nature Rev. Neurosci. 2, 287–293 (2001).
51. Caviness, V. S. Jr & Takahashi, T. Proliferative events in thecerebral ventricular zone. Brain Dev. 17, 159–163 (1995).
52. Altman, J. & Bayer, S. A. Development of the CerebellarSystem. In Relation to its Evolution, Structure, and Functions(CRC Press, New York, 1997).
53. Hatten, M. E & Heintz, N. Mechanisms of neural patterningand specification in the developing cerebellum. Annu. Rev.Neurosci. 18, 385–408 (1995).
54. Wang, V. Y. & Zoghbi, H. Y. Genetic regulation of cerebellardevelopment. Nature Rev. Neurosci. 2, 484–491 (2001).
55. Alder, J., Lee, K. J., Jessell, T. M. & Hatten, M. E. Generationof cerebellar granule neurons in vivo by transplantation ofBMP-treated neural progenitor cells. Nature Neurosci. 2,535–540 (1999).Shows that the early specification and generation ofgranule neuron precursors in the rhombic lip is by aBMP-dependent, SHH-independent mechanism.
56. Sato, T., Araki, I. & Nakamura, H. Inductive signal and tissueresponsiveness defining the tectum and the cerebellum.Development 128, 2461–2469 (2001).
57. Xu, J., Liu, Z. & Ornitz, D. M. Temporal and spatial gradientsof Fgf8 and Fgf17 regulate proliferation and differentiation ofmidline cerebellar structures. Development 127, 1833–1843(2000).
58. Ericson, J., Morton, S., Kawakami, A., Roelink, H. & Jessell,T. M. Two critical periods of Sonic Hedgehog signalingrequired for the specification of motor neuron identity. Cell87, 661–673 (1996).Shows the requirement of SHH signalling at differenttimes in the cell cycle.
59. Ben-Arie, N. et al. Math1 is essential for genesis ofcerebellar granule neurons. Nature 390, 169–172 (1997).
60. Watanabe, Y. & Nakamura, H. Control of chick tectumterritory along dorsoventral axis by Sonic hedgehog.Development 127, 1131–1140 (2000).
61. Hatten, M. E. Central nervous system neuronal migration.Annu. Rev. Neurosci. 22, 511–539 (1999).
62. Pons, S., Trejo, J. L., Martinez-Morales, J. R. & Marti, E.Vitronectin regulates Sonic hedgehog activity duringcerebellum development through CREB phosphorylation.Development 128, 1481–1492 (2001).
63. Herrup, K. Thoughts on the cerebellum as a model forcerebral cortical development and evolution. NovartisFound. Symp. 228, 15–24 (2000).
64. Xuan, S. et al. Winged helix transcription factor BF-1 isessential for the development of the cerebral hemispheres.Neuron 14, 1141–1152 (1995).
65. Theil, T., Alvarez-Bolado, G., Walter, A. & Ruther, U. Gli3 isrequired for Emx gene expression during dorsaltelencephalon development. Development 126, 3561–3571(1999).
66. Toole, S. Ragsdale, C. W. & Grove, E. A. Dorsoventralpatterning of the mouse telencephalon is disrupted in themouse mutant extra-toes. Dev. Biol. 217, 254–265 (2000).
67. Kornack, D. R. & Rakic, P. Changes in cell-cycle kineticsduring the development and evolution of primate neocortex.Proc. Natl Acad. Sci. USA 95, 1242–1246 (1998).
68. Aza-Blanc, P., Ramirez-Weber, F.-A., Laget, M.-P.,Schwartz, C. & Kornberg, T. B. Proteolysis that is inhibited byHedgehog targets Cubitus interruptus protein to the nucleusand converts it to a repressor. Cell 89, 1043–1053 (1997).Demonstration of repressor activity by proteolyticcleavage of the Drosophila Gli.
69. Methot, N. & Basler, K. Hedgehog controls limbdevelopment by regulating the activities of distincttranscriptional activator and repressor forms of Cubitusinterruptus. Cell 96, 819–831 (1999).A detailed analysis of the control of activator andrepressor functions of Ci, the Drosophila Glihomologue.
70. Shin, S. H, Kogerman, P., Lindstrom, E., Toftgard, R. &Biesecker, L. G. GLI3 mutations in human disorders mimicDrosophila Cubitus interruptus protein functions andlocalization. Proc. Natl Acad. Sci. USA 96, 2880–2884(1999).Structure–function analysis of Gli3 and its repressoractivity.
71. Dai, P. et al. Sonic Hedgehog-induced activation of the Gli1promoter is mediated by GLI3. J. Biol. Chem. 274,8143–8152 (1999).
72. Wang, B., Fallon, J. F. & Beachy, P. A. Hedgehog-regulatedprocessing of Gli3 produces an anterior/posterior repressorgradient in the developing vertebrate limb. Cell 100,423–434 (2000).Shows the presence of smaller-than-full-length Gli3bands in chick limb bud western blots, indicating theexistence of putative repressor forms in vivo.
73. Park, H. L. et al. Mouse Gli1 mutants are viable but havedefects in SHH signaling in combination with a Gli2mutation. Development 127, 1593–1605 (2000).Shows that mouse mutants for Gli1 are viable.
74. Matise, M. P., Epstein, D. J., Park, H. L., Platt, K. A. &Joyner, A. L. Gli2 is required for induction of floor plate andadjacent cells, but not most ventral neurons in the mousecentral nervous system. Development 125, 2759–2770(1998).
75. Smith, M. J. et al. GLI-2 modulates retroviral geneexpression. J. Virol. 75, 2301–2313 (2001).
76. Krishnan, V. et al. Mediation of Sonic hedgehog-inducedexpression of COUP-TFII by a protein phosphatase.Science 278, 1947–1950 (1997).
77. Gallet, A., Angelats, C., Kerridge, S. & Therond, P. P. Cubitusinterruptus-independent transduction of the Hedgehogsignal in Drosophila. Development 127, 5509–5522 (2000).
78. Sasaki, H., Nishizaki, Y., Hui, C., Nakafuku, M. & Kondoh, H.Regulation of Gli2 and Gli3 activities by an amino-terminalrepression domain: implication of Gli2 and Gli3 as primarymediators of Shh signaling. Development 126, 3915–3924(1999).Evidence that the early mediation of SHH signalsinvolves Gli2 function.
79. Hui, C. C & Joyner, A. L. A mouse model of Greigcephalopolysyndactyly syndrome: the extra-toes mutationcontains an intragenic deletion of the Gli3 gene. NatureGenet. 3, 241–246 (1993).
80. Franz, T. Extra-toes (Xt) homozygous mice demonstrate arole for the Gli3 gene in the development of the forebrain.Acta Anat. 150, 38–44 (1994).
81. Brewster, R., Lee, J. & Ruiz i Altaba, A. Gli/Zic factorspattern the neural plate by defining domains of celldifferentiation. Nature 393, 579–583 (1998).
82. Kenney, A. M. & Rowitch, D. H. Sonic hedgehog promotesG(1) cyclin expression and sustained cell cycle progressionin mammalian neuronal precursors. Mol. Cell. Biol. 20,9055–9067 (2000).An exploration of the ways in which SHH affects thecell cycle, providing evidence that it acts on G1cyclins.
83. Huard, J. M., Forster, C. C., Carter, M. L., Sicinski, P. &Ross, M. E. Cerebellar histogenesis is disturbed in micelacking cyclin D2. Development 126, 1927–1935 (1999).
84. Barnes, E. A., Kong, M., Ollendorff, V. & Donoghue, D. J.Patched1 interacts with cyclin B1 to regulate cell cycleprogression. EMBO J. 20, 2214–2223 (2001).
85. Fan, H. & Khavari, P. A. Sonic hedgehog opposes epithelialcell cycle arrest. J. Cell Biol. 147, 71–76 (1999).
86. Miyazawa, K., Himi, T., Garcia, V., Yamagishi, H., Sato, S. &Ishizaki, Y. A role for p27/Kip1 in the control of cerebellar
granule cell precursor proliferation. J. Neurosci. 20,5756–5763 (2000).
87. Sherr, C. J. & Roberts, J. M. CDK inhibitors: positive andnegative regulators of G1-phase progression. Genes Dev.13, 1501–1512 (1999).
88. Bhardwaj, G. et al. Sonic hedgehog induces the proliferationof primitive human hematopoietic cells via BMP regulation.Nature Immunol. 2, 172–180 (2001).
89. Pepinsky, R. B. et al. Identification of a palmitic acid-modified form of human Sonic hedgehog. J. Biol. Chem.273, 14037–14045 (1998).
90. Zeng, X. et al. A freely diffusible form of Sonic hedgehogmediates long-range signalling. Nature 411, 716–720 (2001).
91. Gritli-Linde, A., Lewis, P., McMahon, A. P. & Linde, A. Thewhereabouts of a morphogen: direct evidence for short- andgraded long-range activity of hedgehog signaling peptides.Dev. Biol. 236, 364–386 (2001).Fantastic visualization of SHH protein far from itssources.
92. Heemskerk, J. & DiNardo, S. Drosophila Hedgehog acts asa morphogen in cellular patterning. Cell 76, 449–460 (1994).
93. Chen, Y. & Struhl, G. Dual roles for Patched in sequesteringand transducing Hedgehog. Cell 87, 553–563 (1996).
94. Mullor, J. L., Calleja, M., Capdevila, J. & Guerrero, I. Hedgehogactivity, independent of Decapentaplegic, participates inwing disc patterning. Development 124, 1227–1237 (1997).This work, together with references 97 and 98,provides evidence that HH acts as a morphogen.
95. Mullor, J. L. & Guerrero, I. A gain-of-function mutant ofpatched dissects different responses to Hedgehog gradient.Dev. Biol. 228, 211–224 (2000).
96. Strigini, M. & Cohen, S. M. A Hedgehog activity gradientcontributes to AP axial patterning of the Drosophila wing.Development 124, 4697–4705 (1997).
97. Briscoe, J., Chen, Y., Jessell, T. M. & Struhl, G. A hedgehog-insensitive form of patched provides evidence for directlong-range morphogen activity of sonic hedgehog in theneural tube. Mol. Cell 7, 1279–1291 (2001).
98. Hynes, M. et al. The seven-transmembrane receptorSmoothened cell-autonomously induces multiple ventral celltypes. Nature Neurosci. 3, 41–46 (2000).
99. Lewis, P. M. et al. Modification of Sonic hedgehog is requiredfor long-range signaling activity and effective modulation ofsignaling by Ptc1. Cell 105, 599–612 (2001).
100. Bellaiche, Y., The, I. & Perrimon, N. Tout-velu is a Drosophilahomologue of the putative tumour suppressor EXT-1 and isneeded for Hh diffusion. Nature 394, 85–88 (1998).
101. Burke, R. et al. Dispatched, a novel sterol-sensing domainprotein dedicated to the release of cholesterol-modifiedHedgehog from signaling cells. Cell 99, 803–815 (1999).
102. Martin, V., Carrillo, G., Torroja, C. & Guerrero, I. The sterol-sensing domain of Patched protein seems to controlSmoothened activity through Patched vesicular trafficking.Curr. Biol. 11, 601–607 (2001).Shows that HH signalling is regulated by the transportof vesicles (see also reference 103).
103. Strutt, H. et al. Mutations in the sterol-sensing domain ofPatched suggest a role for vesicular trafficking inSmoothened regulation. Curr. Biol. 11, 608–613 (2001).
104. Eggenschwiler, J. T., Espinoza, E. & Anderson, K. V. Rab23 is an essential negative regulator of the mouse Sonic hedgehog signalling pathway. Nature 412, 194–198(2001).Highlights the presence of inhibitors of SHH signallingin the dorsal neural tube, as well as the regulation ofSHH diffusion/signalling by vesicular components.
105. Incardona, J. P. et al. Receptor-mediated endocytosis ofsoluble membrane-tethered Sonic hedgehog by Patched-1.Proc. Natl Acad. Sci. USA 97, 12044–12049 (2000).
106. Huang, Z. & Kunes, S. Hedgehog, transmitted along retinalaxons, triggers neurogenesis in the developing visualcenters of the Drosophila brain. Cell 86, 411–422 (1996).Shows that HH can be transported through axons toaffect target cells.
107. Wallace, V. A. & Raff, M. C. A role for Sonic hedgehog inaxon-to-astrocyte signalling in the rodent optic nerve.Development 126, 2901–2909 (1999).
108. Traiffort, E., Moya, K. L., Faure, H., Hässig, R. & Ruat, M.High expression and anterograde axonal transport of animoterminal sonic hedgehog in the adult hamster brain. Eur. J.Neurosci. 14, 839–850 (2001).
109. Ramirez-Weber, F. A. & Kornberg, T. B. Cytonemes: cellularprocesses that project to the principal signaling center inDrosophila imaginal discs. Cell 97, 599–607 (1999).Cytonemes as an alternative to morphogens for long-range action of signalling molecules.
110. Dale, J. K. et al. Cooperation of BMP7 and SHH in theinduction of forebrain ventral midline cells by prechordalmesoderm. Cell 90, 257–269 (1997).
111. Liem, K. F. Jr, Tremml, G., Roelink, H. & Jessell, T. M. Dorsaldifferentiation of neural plate cells induced by BMP-

NATURE REVIEWS | NEUROSCIENCE VOLUME 3 | JANUARY 2002 | 33
R E V I E W S
mediated signals from epidermal ectoderm. Cell 82,969–979 (1995).An elegant demonstration that BMPs control dorsaldifferentiation in the early neural tube.
112. Zuniga, A., Haramis, A. P., McMahon, A. P. & Zeller, R. Signalrelay by BMP antagonism controls the SHH/FGF4 feedbackloop in vertebrate limb buds. Nature 401, 598–602 (1999).
113. Rodriguez Esteban, C. et al. The novel Cer-like proteinCaronte mediates the establishment of embryonic left–rightasymmetry. Nature 401, 243–251 (1999).
114. Yokouchi, Y., Vogan, K. J., Pearse, R. V. & Tabin, C. J.Antagonistic signaling by Caronte, a novel Cerberus-relatedgene, establishes left–right asymmetric gene expression.Cell 98, 573–583 (1999).
115. Liem, K. F. Jr, Jessell, T. M. & Briscoe, J. Regulation of theneural patterning activity of sonic hedgehog by secretedBMP inhibitors expressed by notochord and somites.Development 127, 4855–4866 (2000).
116. Liu, F., Massague, J. & Ruiz i Altaba, A. Carboxy-terminallytruncated Gli3 proteins associate with Smads. NatureGenet. 20, 325–326 (1998).
117. Ohlmeyer, J. T. & Kalderon, D. Hedgehog stimulatesmaturation of Cubitus interruptus into a labile transcriptionalactivator. Nature 396, 749–753 (1998).Evidence that HH is required to make a labile Ciactivator in flies. The nature of the modifications thatmake this activator remain unknown.
118. Zhu, G., Mehler, M. F., Zhao, J., Yu Yung, S. & Kessler, J. A.Sonic hedgehog and BMP2 exert opposing actions onproliferation and differentiation of embryonic neuralprogenitor cells. Dev. Biol. 215, 118–129 (1999).
119. Li, W. & LoTurco, J. J. Noggin is a negative regulator ofneuronal differentiation in developing neocortex. Dev.Neurosci. 22, 68–73 (2000).
120. Panchision, D. M. et al. Sequential actions of BMP receptorscontrol neural precursor cell production and fate. GenesDev. 15, 2094–2110 (2001).
121. Kawai, S. & Sugiura, T. Characterization of human bonemorphogenetic protein (BMP)-4 and -7 gene promoters:activation of BMP promoters by Gli, a sonic hedgehogmediator. Bone 29, 54–61 (2001).
122. Brewster, R., Mullor, J. L. & Ruiz i Altaba, A. Gli2 functions inFGF signaling during antero-posterior patterning.Development 127, 4395–4405 (2000).
123. Grove, E. A., Tole, S., Limon, J., Yip, L. & Ragsdale, C. W.The hem of the embryonic cerebral cortex is defined by the
expression of multiple Wnt genes and is compromised inGli3-deficient mice. Development 125, 2315–2325 (1998).
124. Lee, S. M., Tole, S., Grove, E. & McMahon, A. P. A localWnt-3a signal is required for development of the mammalianhippocampus. Development 127, 457–467 (2000).
125. Ragsdale, C. W. & Grove, E. A. Patterning the mammaliancerebral cortex. Curr. Opin. Neurobiol. 11, 50–58 (2001).
126. Kim, A. S., Lowenstein, D. H. & Pleasure, S. J. Wntreceptors and Wnt inhibitors are expressed in gradients inthe developing telencephalon. Mech. Dev. 103, 167–172(2001).
127. Mullor, J. L., Dahmane, N., Sun, T. & Ruiz i Altaba, A. Wntsignals are targets and mediators of Gli function. Curr. Biol.11, 769–773 (2001).
128. Brault, V. et al. Inactivation of the β-catenin gene byWnt1–Cre-mediated deletion results in dramatic brainmalformation and failure of craniofacial development.Development 128, 1253–1264 (2001).
129. Chuang, P. T. & McMahon, A. P. Vertebrate Hedgehogsignalling modulated by induction of a Hedgehog-bindingprotein. Nature 397, 617–621 (1999).Identification of Hip as a negative regulator of HHsignalling.
130. Hahn, H. et al. Patched target Igf2 is indispensable for theformation of medulloblastoma and rhabdomyosarcoma. J. Biol. Chem. 275, 28341–28344 (2000).
131. Jiang, F., Frederick, T. J. & Wood, T. L. IGF-I synergizes withFGF-2 to stimulate oligodendrocyte progenitor entry into thecell cycle. Dev. Biol. 232, 414–423 (2001).
132. Calaora V. et al. Neuregulin signaling regulates neuralprecursor growth and the generation of oligodendrocytes in vitro. J. Neurosci. 21, 4740–4751 (2001).
133. Suh, J., Lu, N., Nicot, A., Tatsuno, I. & DiCicco-Bloom, E.PACAP is an anti-mitogenic signal in developing cerebralcortex. Nature Neurosci. 4, 123–124 (2001).
134. Waschek, J. A. et al. Neural tube expression of pituitaryadenylate cyclase-activating peptide (PACAP) and receptor:potential role in patterning and neurogenesis. Proc. NatlAcad. Sci. USA 95, 9602–9607 (1998).
135. Pons, S. & Marti, E. Sonic hedgehog synergizes with theextracellular matrix protein vitronectin to induce spinal motorneuron differentiation. Development 127, 333–342 (2000).Highlights the importance of the extracellular matrixin the control of SHH signalling.
136. Lee, C. S., Buttitta, L. & Fan, C. M. Evidence that the WNT-inducible growth arrest-specific gene 1 encodes
an antagonist of sonic hedgehog signaling in the somite. Proc. Natl Acad. Sci. USA 98, 11347–11352 (2001).
137. Mullor, J. L. & Ruiz i Altaba, A. Growth, hedgehog and theprice of Gas. Bioessays (in the press).
138. Huh, S., Hatini, V., Marcus, R. C., Li, S. C. & Lai, E.Dorsal–ventral patterning defects in the eye of BF-1-deficient mice associated with a restricted loss of shhexpression. Dev. Biol. 211, 53–63 (1999).
139. Solecki, D. J., Liu, X. L., Tomoda, T., Fang, Y. & Hatten, M. E.Activated Notch2 signaling inhibits differentiation ofcerebellar granule neuron precursors by maintainingproliferation. Neuron 31, 557–568 (2001).
140. Goodrich, L. V., Milenkovic, L., Higgins, K. M. & Scott, M. P.Altered neural cell fates and medulloblastoma in mousepatched mutants. Science 277, 1109–1113 (1997).Reports a mouse model of medulloblastoma.
AcknowledgementsWe are grateful to J. L. Mullor, P. Sánchez and Y. Gitton for com-ments. V.P. is a Latin-American Pew Fellow. Work in A.R.A.’s labo-ratory is supported by grants from the March of Dimes, the NationalCancer Institute and National Institute for Neurological Disordersand Stroke. The laboratory of N.D. is funded by grants from theCentre National de la Recherche Scientifique, La Fondation pour laRecherche Médicale and L’Association pour la Recherche sur le Cancer. Due to space limitations, only a proportion of relevant references could be cited here.
Online links
DATABASESThe following terms in this article are linked online to:FlyBase: http://flybase.bio.indiana.edu/Ci | disp | dpp | ptc | ttvLocusLink: http://www.ncbi.nlm.nih.gov/LocusLink/BF-1 | BMP | brachyury | β-catenin | Cdk | CREB | cyclin B | cyclin D2 | E2F | EGF | Emx | FGF | Gas1 | Gli1 | Gli2 | Gli3 | Hip |IGF | nestin | neuregulin | Notch | PACAP | PDGF | Ptc1 | Rab23 |Rb | SHH | Smad | Smo | Su(fu) | vitronectin | Wnt
FURTHER INFORMATIONEncyclopedia of Life Sciences: http://www.els.net/cerebral cortex development | neuronal subtype identity regulationAccess to this interactive links box is free online.
Related Documents





![Regulation of Hedgehog Signalling Inside and Outside the Celleprints.whiterose.ac.uk/119986/1/jdb_04_00023.pdfJ. Dev. Biol. 2016, 4, 23 2 of 20 elsewhere [11,12]. This review will](https://static.cupdf.com/doc/110x72/6128f9d9bafd3e72b224dcda/regulation-of-hedgehog-signalling-inside-and-outside-the-j-dev-biol-2016-4.jpg)





![Gastric cancer and Hedgehog signaling pathway: emerging ...Hedgehog signalling as compared to the adjacent duodenal tissues [19], and also is proposed to maintain pit-gland asymmetry](https://static.cupdf.com/doc/110x72/60675b8841f1e547814bd087/gastric-cancer-and-hedgehog-signaling-pathway-emerging-hedgehog-signalling.jpg)