Hedgehog signaling and primary cilia are required for the formation of adult neural stem cells Young-Goo Han 1 , Nathalie Spassky 2 , Miriam Romaguera-Ros 3 , Jose-Manuel Garcia-Verdugo 3 , Andrea Aguilar 2 , Sylvie Schneider-Maunoury 4 & Arturo Alvarez-Buylla 1 Neural stem cells that continue to produce neurons are retained in the adult hippocampal dentate gyrus. The mechanisms by which embryonic neural progenitors expand and transform into postnatal neural stem cells, an essential process for the continual production of neurons throughout life, remain unknown. We found that radial astrocytes, the postnatal progenitors in the dentate gyrus, failed to develop after embryonic ablation of ciliary genes or Smoothened (Smo), an essential component for Sonic hedgehog (Shh) signaling. Postnatal dentate neurogenesis failed in these mutant mice, and the dentate gyrus became severely hypotrophic. In contrast, expression of a constitutively active Smo (SmoM2-YFP) resulted in a marked expansion of the dentate gyrus. Double-mutant analyses suggested that both wild-type Smo and SmoM2-YFP function through the primary cilia. We conclude that Shh signaling, acting through the primary cilia, has a critical role in the expansion and establishment of postnatal hippocampal progenitors. Granule neurons in the hippocampal dentate gyrus receive the primary input from the cortex and have crucial roles in learning and memory. The genesis of these neurons begins during mid-gestation and con- tinues throughout life. Granule neuron precursors (GNPs) are initially specified in the dentate neuroepithelium of the hippocampus, migrate from this region into the developing dentate gyrus, expand in number and assume final residence in the subgranular zone (SGZ) between the condensing granular layer and the hilus 1,2 . Production of new granule neurons continues postnatally, resulting in a substantial increase in the size of the dentate gyrus during the first 10 d of life in mice. During this period, GNPs acquire morphological features of astroglial cells, with radial processes spanning the entire granular layer. These radial astro- cytes, also known as type I progenitors, function as the SGZ stem cells and continue to generate new granule neurons throughout life 3–5 . Initial expansion of GNPs and their conversion into radial astrocytes are essential steps for proper dentate gyrus development and the maintenance of neurogenesis in the adult. The primary cilium is a hair-like appendage extending from the surface of a cell. This specialized structure, with a unique microtubular cytoskeleton (axoneme) and a surrounding membrane, is assembled and maintained by the intraflagellar transport (IFT) machinery 6 . IFT is a specialized intracellular trafficking between the cell body and the tip of the cilia, in which kinesin-II and cytoplasmic dynein motors move cargoes bound to multiprotein complexes called IFT particles along the ciliary axoneme. IFT is a highly conserved process, as mutations in the IFT motors or particles result in the loss of cilia from single-celled organisms to mammals. Recent work has shown that primary cilia concentrate receptors and signal transduction components that have vital roles in develop- ment 7–9 . In particular, evidence in the developing neural tube and limb bud has shown that genes encoding the IFT motors (Kif3a, a subunit of kinesin-II, and Dnchc2, a subunit of cytoplasmic dynein) and the IFT particle subunits (Ift52, Ift88 and Ift172) are required for Shh signal- ing 10–14 . Consistently, essential Shh signaling components, Patched (Ptc), Smo, Suppressor of fused and Gli transcription factors, concen- trate in primary cilia 10,15,16 . Defective primary cilia have been associated with diverse human diseases including Alstro ¨m, Bardet-Biedl, Joubert, Meckel-Gruber and Oral-facial-digital type 1 syndromes 17 . The pathology of these pleio- tropic disorders includes obesity, mental retardation and ataxia, suggesting that primary cilia are required for the proper development or function of the brain. A recent study showed that conditional ablation of primary cilia in a subset of hypothalamic neurons caused hyperphagia-induced obesity 18 . Neural stem cells in both embryonic and adult brains have a primary cilium 19 , but its function is not known. Using mutant mice that lack primary cilia in a subset of embryonic neural stem cells, we show here that primary cilia and Shh signaling are essential for the expansion and establishment of GNPs in the postnatal dentate gyrus. RESULTS Loss of primary cilia in hGFAP::Cre; Kif3a fl/fl mice Primary cilia are present in neural progenitor cells in the ventricular zone of the embryo and in the subventricular zone of the adult Received 19 December 2007; accepted 29 January 2008; published online 24 February 2008; doi:10.1038/nn2059 1 Department of Neurological Surgery, Institute for Regeneration Medicine, University of California San Francisco, San Francisco, California 94143, USA. 2 Universite ´ Pierre and Marie Curie-Paris 6, IFR des Neurosciences, Ho ˆpital Pitie ´-Salpe ˆtrie `re, INSERM, U711, Paris, F-75013 France. 3 Laboratorio de Morfologia Celular, Unidad asociada Universidad de Valencia and Centro de Investigacion Principe Felipe, Valencia, 46100 Spain. 4 CNRS UMR7622, Universite ´ Pierre et Marie Curie, 9 Quai Saint Bernard, 75005 Paris, France. Correspondence should be addressed to A.A.-B. ([email protected]). NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 3 [ MARCH 2008 277 ARTICLES © 2008 Nature Publishing Group http://www.nature.com/natureneuroscience

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hedgehog signaling and primary cilia are required forthe formation of adult neural stem cells
Young-Goo Han1, Nathalie Spassky2, Miriam Romaguera-Ros3, Jose-Manuel Garcia-Verdugo3, Andrea Aguilar2,Sylvie Schneider-Maunoury4 & Arturo Alvarez-Buylla1
Neural stem cells that continue to produce neurons are retained in the adult hippocampal dentate gyrus. The mechanisms by
which embryonic neural progenitors expand and transform into postnatal neural stem cells, an essential process for the continual
production of neurons throughout life, remain unknown. We found that radial astrocytes, the postnatal progenitors in the dentate
gyrus, failed to develop after embryonic ablation of ciliary genes or Smoothened (Smo), an essential component for Sonic
hedgehog (Shh) signaling. Postnatal dentate neurogenesis failed in these mutant mice, and the dentate gyrus became severely
hypotrophic. In contrast, expression of a constitutively active Smo (SmoM2-YFP) resulted in a marked expansion of the dentate
gyrus. Double-mutant analyses suggested that both wild-type Smo and SmoM2-YFP function through the primary cilia. We
conclude that Shh signaling, acting through the primary cilia, has a critical role in the expansion and establishment of postnatal
hippocampal progenitors.
Granule neurons in the hippocampal dentate gyrus receive the primaryinput from the cortex and have crucial roles in learning and memory.The genesis of these neurons begins during mid-gestation and con-tinues throughout life. Granule neuron precursors (GNPs) are initiallyspecified in the dentate neuroepithelium of the hippocampus, migratefrom this region into the developing dentate gyrus, expand in numberand assume final residence in the subgranular zone (SGZ) between thecondensing granular layer and the hilus1,2. Production of new granuleneurons continues postnatally, resulting in a substantial increase in thesize of the dentate gyrus during the first 10 d of life in mice. During thisperiod, GNPs acquire morphological features of astroglial cells, withradial processes spanning the entire granular layer. These radial astro-cytes, also known as type I progenitors, function as the SGZ stem cellsand continue to generate new granule neurons throughout life3–5.Initial expansion of GNPs and their conversion into radial astrocytesare essential steps for proper dentate gyrus development and themaintenance of neurogenesis in the adult.
The primary cilium is a hair-like appendage extending from thesurface of a cell. This specialized structure, with a unique microtubularcytoskeleton (axoneme) and a surrounding membrane, is assembledand maintained by the intraflagellar transport (IFT) machinery6. IFT isa specialized intracellular trafficking between the cell body and the tipof the cilia, in which kinesin-II and cytoplasmic dynein motors movecargoes bound to multiprotein complexes called IFT particles along theciliary axoneme. IFT is a highly conserved process, as mutations in theIFT motors or particles result in the loss of cilia from single-celledorganisms to mammals.
Recent work has shown that primary cilia concentrate receptorsand signal transduction components that have vital roles in develop-ment7–9. In particular, evidence in the developing neural tube and limbbud has shown that genes encoding the IFT motors (Kif3a, a subunit ofkinesin-II, and Dnchc2, a subunit of cytoplasmic dynein) and the IFTparticle subunits (Ift52, Ift88 and Ift172) are required for Shh signal-ing10–14. Consistently, essential Shh signaling components, Patched(Ptc), Smo, Suppressor of fused and Gli transcription factors, concen-trate in primary cilia10,15,16.
Defective primary cilia have been associated with diverse humandiseases including Alstrom, Bardet-Biedl, Joubert, Meckel-Gruber andOral-facial-digital type 1 syndromes17. The pathology of these pleio-tropic disorders includes obesity, mental retardation and ataxia,suggesting that primary cilia are required for the proper developmentor function of the brain. A recent study showed that conditionalablation of primary cilia in a subset of hypothalamic neurons causedhyperphagia-induced obesity18. Neural stem cells in both embryonicand adult brains have a primary cilium19, but its function is not known.Using mutant mice that lack primary cilia in a subset of embryonicneural stem cells, we show here that primary cilia and Shh signaling areessential for the expansion and establishment of GNPs in the postnataldentate gyrus.
RESULTS
Loss of primary cilia in hGFAP::Cre; Kif3afl/fl mice
Primary cilia are present in neural progenitor cells in the ventricularzone of the embryo and in the subventricular zone of the adult
Received 19 December 2007; accepted 29 January 2008; published online 24 February 2008; doi:10.1038/nn2059
1Department of Neurological Surgery, Institute for Regeneration Medicine, University of California San Francisco, San Francisco, California 94143, USA. 2Universite Pierreand Marie Curie-Paris 6, IFR des Neurosciences, Hopital Pitie-Salpetriere, INSERM, U711, Paris, F-75013 France. 3Laboratorio de Morfologia Celular, Unidad asociadaUniversidad de Valencia and Centro de Investigacion Principe Felipe, Valencia, 46100 Spain. 4CNRS UMR7622, Universite Pierre et Marie Curie, 9 Quai Saint Bernard,75005 Paris, France. Correspondence should be addressed to A.A.-B. ([email protected]).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 3 [ MARCH 2008 277
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
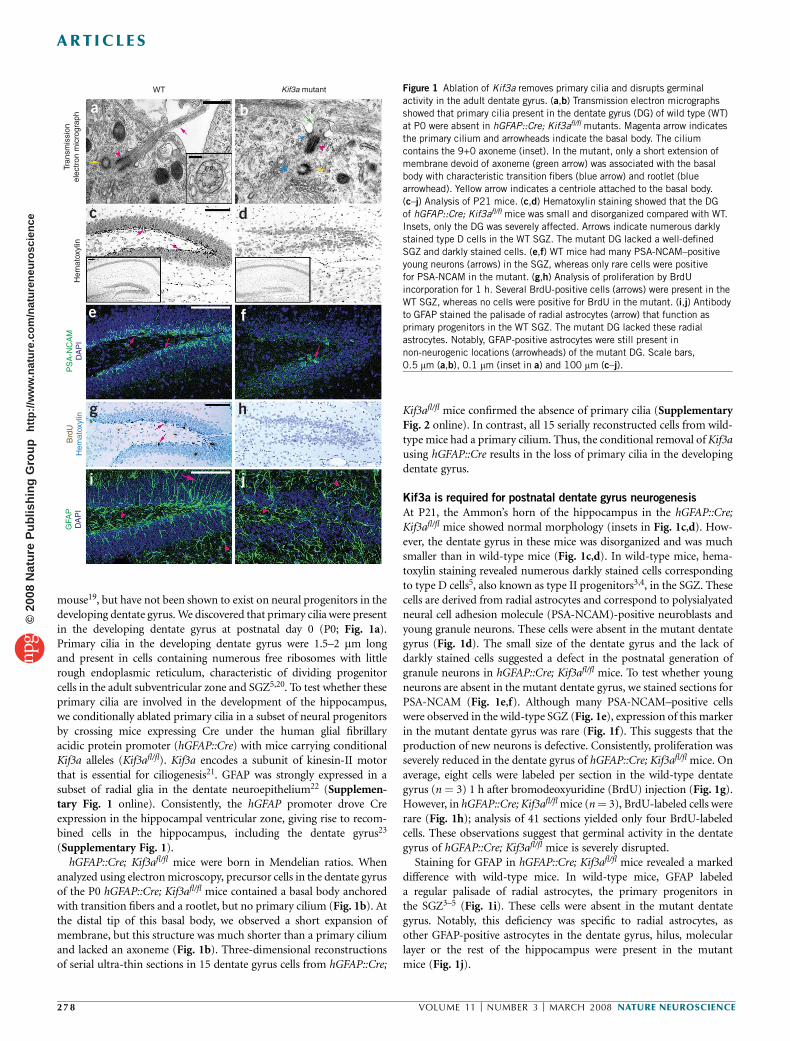
mouse19, but have not been shown to exist on neural progenitors in thedeveloping dentate gyrus. We discovered that primary cilia were presentin the developing dentate gyrus at postnatal day 0 (P0; Fig. 1a).Primary cilia in the developing dentate gyrus were 1.5–2 mm longand present in cells containing numerous free ribosomes with littlerough endoplasmic reticulum, characteristic of dividing progenitorcells in the adult subventricular zone and SGZ5,20. To test whether theseprimary cilia are involved in the development of the hippocampus,we conditionally ablated primary cilia in a subset of neural progenitorsby crossing mice expressing Cre under the human glial fibrillaryacidic protein promoter (hGFAP::Cre) with mice carrying conditionalKif3a alleles (Kif3afl/fl). Kif3a encodes a subunit of kinesin-II motorthat is essential for ciliogenesis21. GFAP was strongly expressed in asubset of radial glia in the dentate neuroepithelium22 (Supplemen-tary Fig. 1 online). Consistently, the hGFAP promoter drove Creexpression in the hippocampal ventricular zone, giving rise to recom-bined cells in the hippocampus, including the dentate gyrus23
(Supplementary Fig. 1).hGFAP::Cre; Kif3afl/fl mice were born in Mendelian ratios. When
analyzed using electron microscopy, precursor cells in the dentate gyrusof the P0 hGFAP::Cre; Kif3afl/fl mice contained a basal body anchoredwith transition fibers and a rootlet, but no primary cilium (Fig. 1b). Atthe distal tip of this basal body, we observed a short expansion ofmembrane, but this structure was much shorter than a primary ciliumand lacked an axoneme (Fig. 1b). Three-dimensional reconstructionsof serial ultra-thin sections in 15 dentate gyrus cells from hGFAP::Cre;
Kif3afl/fl mice confirmed the absence of primary cilia (SupplementaryFig. 2 online). In contrast, all 15 serially reconstructed cells from wild-type mice had a primary cilium. Thus, the conditional removal of Kif3ausing hGFAP::Cre results in the loss of primary cilia in the developingdentate gyrus.
Kif3a is required for postnatal dentate gyrus neurogenesis
At P21, the Ammon’s horn of the hippocampus in the hGFAP::Cre;Kif3afl/fl mice showed normal morphology (insets in Fig. 1c,d). How-ever, the dentate gyrus in these mice was disorganized and was muchsmaller than in wild-type mice (Fig. 1c,d). In wild-type mice, hema-toxylin staining revealed numerous darkly stained cells correspondingto type D cells5, also known as type II progenitors3,4, in the SGZ. Thesecells are derived from radial astrocytes and correspond to polysialyatedneural cell adhesion molecule (PSA-NCAM)-positive neuroblasts andyoung granule neurons. These cells were absent in the mutant dentategyrus (Fig. 1d). The small size of the dentate gyrus and the lack ofdarkly stained cells suggested a defect in the postnatal generation ofgranule neurons in hGFAP::Cre; Kif3afl/fl mice. To test whether youngneurons are absent in the mutant dentate gyrus, we stained sections forPSA-NCAM (Fig. 1e,f). Although many PSA-NCAM–positive cellswere observed in the wild-type SGZ (Fig. 1e), expression of this markerin the mutant dentate gyrus was rare (Fig. 1f). This suggests that theproduction of new neurons is defective. Consistently, proliferation wasseverely reduced in the dentate gyrus of hGFAP::Cre; Kif3afl/fl mice. Onaverage, eight cells were labeled per section in the wild-type dentategyrus (n ¼ 3) 1 h after bromodeoxyuridine (BrdU) injection (Fig. 1g).However, in hGFAP::Cre; Kif3afl/fl mice (n ¼ 3), BrdU-labeled cells wererare (Fig. 1h); analysis of 41 sections yielded only four BrdU-labeledcells. These observations suggest that germinal activity in the dentategyrus of hGFAP::Cre; Kif3afl/fl mice is severely disrupted.
Staining for GFAP in hGFAP::Cre; Kif3afl/fl mice revealed a markeddifference with wild-type mice. In wild-type mice, GFAP labeleda regular palisade of radial astrocytes, the primary progenitors inthe SGZ3–5 (Fig. 1i). These cells were absent in the mutant dentategyrus. Notably, this deficiency was specific to radial astrocytes, asother GFAP-positive astrocytes in the dentate gyrus, hilus, molecularlayer or the rest of the hippocampus were present in the mutantmice (Fig. 1j).
WT
Tran
smis
sion
elec
tron
mic
rogr
aph
Hem
atox
ylin
PS
A-N
CA
MD
AP
IB
rdU
Hem
atox
ylin
GFA
PD
AP
I
Kif3a mutant
a b
c d
e f
g h
i j
Figure 1 Ablation of Kif3a removes primary cilia and disrupts germinal
activity in the adult dentate gyrus. (a,b) Transmission electron micrographs
showed that primary cilia present in the dentate gyrus (DG) of wild type (WT)
at P0 were absent in hGFAP::Cre; Kif3afl/fl mutants. Magenta arrow indicates
the primary cilium and arrowheads indicate the basal body. The cilium
contains the 9+0 axoneme (inset). In the mutant, only a short extension of
membrane devoid of axoneme (green arrow) was associated with the basal
body with characteristic transition fibers (blue arrow) and rootlet (bluearrowhead). Yellow arrow indicates a centriole attached to the basal body.
(c–j) Analysis of P21 mice. (c,d) Hematoxylin staining showed that the DG
of hGFAP::Cre; Kif3afl/fl mice was small and disorganized compared with WT.
Insets, only the DG was severely affected. Arrows indicate numerous darkly
stained type D cells in the WT SGZ. The mutant DG lacked a well-defined
SGZ and darkly stained cells. (e,f) WT mice had many PSA-NCAM–positive
young neurons (arrows) in the SGZ, whereas only rare cells were positive
for PSA-NCAM in the mutant. (g,h) Analysis of proliferation by BrdU
incorporation for 1 h. Several BrdU-positive cells (arrows) were present in the
WT SGZ, whereas no cells were positive for BrdU in the mutant. (i,j) Antibody
to GFAP stained the palisade of radial astrocytes (arrow) that function as
primary progenitors in the WT SGZ. The mutant DG lacked these radial
astrocytes. Notably, GFAP-positive astrocytes were still present in
non-neurogenic locations (arrowheads) of the mutant DG. Scale bars,
0.5 mm (a,b), 0.1 mm (inset in a) and 100 mm (c–j).
278 VOLUME 11 [ NUMBER 3 [ MARCH 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
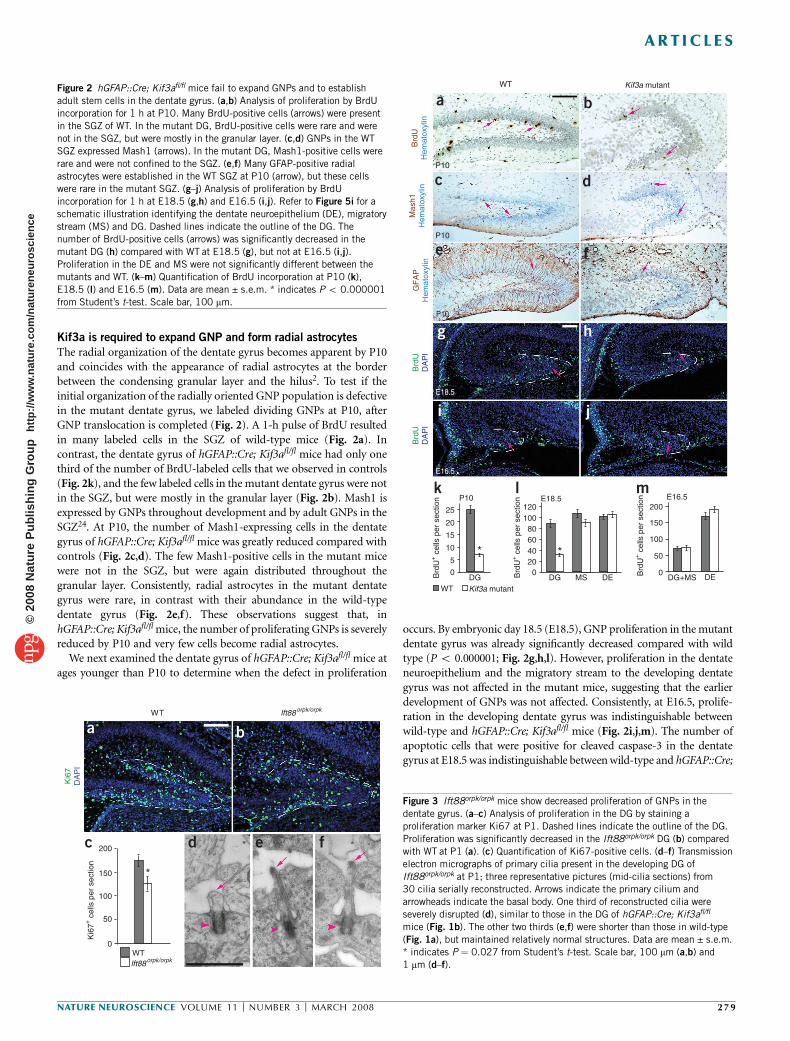
Kif3a is required to expand GNP and form radial astrocytes
The radial organization of the dentate gyrus becomes apparent by P10and coincides with the appearance of radial astrocytes at the borderbetween the condensing granular layer and the hilus2. To test if theinitial organization of the radially oriented GNP population is defectivein the mutant dentate gyrus, we labeled dividing GNPs at P10, afterGNP translocation is completed (Fig. 2). A 1-h pulse of BrdU resultedin many labeled cells in the SGZ of wild-type mice (Fig. 2a). Incontrast, the dentate gyrus of hGFAP::Cre; Kif3afl/fl mice had only onethird of the number of BrdU-labeled cells that we observed in controls(Fig. 2k), and the few labeled cells in the mutant dentate gyrus were notin the SGZ, but were mostly in the granular layer (Fig. 2b). Mash1 isexpressed by GNPs throughout development and by adult GNPs in theSGZ24. At P10, the number of Mash1-expressing cells in the dentategyrus of hGFAP::Cre; Kif3afl/fl mice was greatly reduced compared withcontrols (Fig. 2c,d). The few Mash1-positive cells in the mutant micewere not in the SGZ, but were again distributed throughout thegranular layer. Consistently, radial astrocytes in the mutant dentategyrus were rare, in contrast with their abundance in the wild-typedentate gyrus (Fig. 2e,f). These observations suggest that, inhGFAP::Cre; Kif3afl/fl mice, the number of proliferating GNPs is severelyreduced by P10 and very few cells become radial astrocytes.
We next examined the dentate gyrus of hGFAP::Cre; Kif3afl/fl mice atages younger than P10 to determine when the defect in proliferation
occurs. By embryonic day 18.5 (E18.5), GNP proliferation in the mutantdentate gyrus was already significantly decreased compared with wildtype (P o 0.000001; Fig. 2g,h,l). However, proliferation in the dentateneuroepithelium and the migratory stream to the developing dentategyrus was not affected in the mutant mice, suggesting that the earlierdevelopment of GNPs was not affected. Consistently, at E16.5, prolife-ration in the developing dentate gyrus was indistinguishable betweenwild-type and hGFAP::Cre; Kif3afl/fl mice (Fig. 2i,j,m). The number ofapoptotic cells that were positive for cleaved caspase-3 in the dentategyrus at E18.5 was indistinguishable between wild-type and hGFAP::Cre;
WT
P10
P10
P10
E18.5
E16.5
Brd
UH
emat
oxyl
in
Mas
h1H
emat
oxyl
inG
FA
PH
emat
oxyl
inB
rdU
DA
PI
Brd
UD
AP
I
Kif3a mutant
Kif3a mutant
25
P10 E18.5 E16.5
20
120
* *
200
150
100
50
0
100806040200B
rdU
+ c
ells
per
sec
tion
Brd
U+ c
ells
per
sec
tion
Brd
U+ c
ells
per
sec
tion
15
10
5
DGWT
DG MS DE0
DG+MS DE
a b
c d
e f
k l m
g h
i j
Figure 2 hGFAP::Cre; Kif3afl/fl mice fail to expand GNPs and to establish
adult stem cells in the dentate gyrus. (a,b) Analysis of proliferation by BrdU
incorporation for 1 h at P10. Many BrdU-positive cells (arrows) were present
in the SGZ of WT. In the mutant DG, BrdU-positive cells were rare and were
not in the SGZ, but were mostly in the granular layer. (c,d) GNPs in the WT
SGZ expressed Mash1 (arrows). In the mutant DG, Mash1-positive cells were
rare and were not confined to the SGZ. (e,f) Many GFAP-positive radial
astrocytes were established in the WT SGZ at P10 (arrow), but these cellswere rare in the mutant SGZ. (g–j) Analysis of proliferation by BrdU
incorporation for 1 h at E18.5 (g,h) and E16.5 (i,j). Refer to Figure 5i for a
schematic illustration identifying the dentate neuroepithelium (DE), migratory
stream (MS) and DG. Dashed lines indicate the outline of the DG. The
number of BrdU-positive cells (arrows) was significantly decreased in the
mutant DG (h) compared with WT at E18.5 (g), but not at E16.5 (i,j).
Proliferation in the DE and MS were not significantly different between the
mutants and WT. (k–m) Quantification of BrdU incorporation at P10 (k),
E18.5 (l) and E16.5 (m). Data are mean ± s.e.m. * indicates P o 0.000001
from Student’s t-test. Scale bar, 100 mm.
*
200
Ki6
7D
AP
I
150
Ki6
7+ c
ells
per
sec
tion
100
50
WTlft88orpk/orpk
lft88orpk/orpk
0
WT
a b
c d e f
Figure 3 Ift88orpk/orpk mice show decreased proliferation of GNPs in the
dentate gyrus. (a–c) Analysis of proliferation in the DG by staining a
proliferation marker Ki67 at P1. Dashed lines indicate the outline of the DG.
Proliferation was significantly decreased in the Ift88orpk/orpk DG (b) compared
with WT at P1 (a). (c) Quantification of Ki67-positive cells. (d–f) Transmission
electron micrographs of primary cilia present in the developing DG of
Ift88orpk/orpk at P1; three representative pictures (mid-cilia sections) from30 cilia serially reconstructed. Arrows indicate the primary cilium and
arrowheads indicate the basal body. One third of reconstructed cilia were
severely disrupted (d), similar to those in the DG of hGFAP::Cre; Kif3afl/fl
mice (Fig. 1b). The other two thirds (e,f) were shorter than those in wild-type
(Fig. 1a), but maintained relatively normal structures. Data are mean ± s.e.m.
* indicates P ¼ 0.027 from Student’s t-test. Scale bar, 100 mm (a,b) and
1 mm (d–f).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 3 [ MARCH 2008 279
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
Kif3afl/fl mice (Supplementary Fig. 3 online). Therefore, differences incell death do not appear to explain the reduction of the numbers ofGNPs. Instead, the loss of Kif3a in GNPs resulted in a marked decreasein GNP proliferation sometime between E16.5 and E18.5.
In hGFAP::Cre; Kif3afl/fl mice, expansion of dentate gyrus progeni-tors is specifically affected; the hippocampus and other telencephalicbrain regions appear to develop normally. This suggested that a lack ofproper kinesin-II function does not result in generalized defects inintracellular trafficking or cytokinesis, as has been suggested byexperiments in cultured cells using overexpression of dominant-negative subunits of kinesin-II25–27. We tested whether intracellulartrafficking and cytokinesis were affected in dentate gyrus progenitorsfrom hGFAP::Cre; Kif3afl/fl mutant dentate gyrus and found nodifferences from controls (Supplementary Fig. 4 online). Takentogether, these observations suggest that the loss of progenitor cellsand hypoplasia of the dentate gyrus in hGFAP::Cre; Kif3afl/fl mice areresults of the loss of primary cilia and not of defective intracellulartrafficking or cytokinesis.
Decreased GNP proliferation in Ift88 and Ftm mutants
To verify that the phenotype in the dentate gyrus of hGFAP::Cre;Kif3afl/fl mice is a result of the loss of cilia, we examined proliferationof GNPs in mice that were defective for Ift88, a gene that encodes anessential subunit of IFT particles. Mice with null alleles of Ift88 diearound E1028, which is much earlier than the development of thedentate gyrus. We therefore analyzed a hypomorphic insertionalmutant allele, Ift88orpk/orpk, which dies postnatally29, allowing us tostudy the early stages of dentate gyrus development. The prolifera-tion of GNPs in the dentate gyrus of Ift88orpk/orpk mice decreasedsignificantly by 38% compared with wild type at P1 (P ¼ 0.027;Fig. 3a–c). This decline was smaller compared with the 64% (atE18.5) or 72% (at P10) decreases that we observed in hGFAP::Cre;Kif3afl/fl mice. This is likely due to the hypomorphic nature of theIft88orpk/orpk mutation. Three-dimensional reconstructions of serialultra-thin sections revealed that one third of primary cilia (10 out of30 reconstructions) in the dentate gyrus of Ift88orpk/orpk mice wereseverely disrupted (Fig. 3d), as shown above for hGFAP::Cre; Kif3afl/fl
mice. The other 20 cilia reconstructed in the dentate gyrus ofIft88orpk/orpk mice were shorter than controls, but maintained rela-tively normal structures (Fig. 3e,f).
Fantom (Ftm; also known as Rpgrip1l, or retinitis pigmentosa GTPaseregulator interacting protein 1 like) encodes a basal body protein, andmutations in this gene are found in ciliary disorders including Joubertand Meckel syndromes30. Consistently, Ftm mutant mice (Ftm–/–) havemalformed cilia and reduced numbers of cilia31. The Ftm–/– phenotypeshows variable penetrance, with death beginning early during embryo-nic development in the severely affected mice and occurring at birth inless severe ones. We analyzed proliferation in the dentate gyrus in
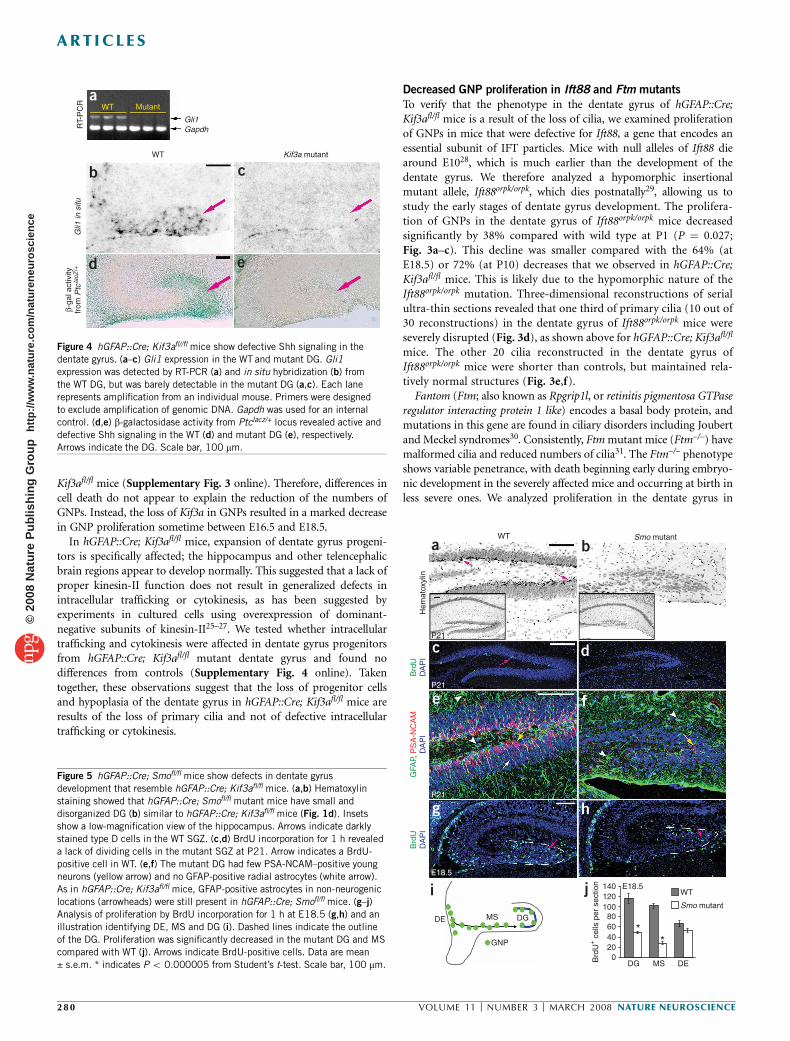
Gli1
WT
WT Mutant
RT-
PC
RG
li1 in
situ
β-ga
l act
ivity
from
Ptc
lacZ
/+
Gapdh
Kif3a mutant
a
b c
d e
Figure 4 hGFAP::Cre; Kif3afl//fl mice show defective Shh signaling in the
dentate gyrus. (a–c) Gli1 expression in the WT and mutant DG. Gli1
expression was detected by RT-PCR (a) and in situ hybridization (b) from
the WT DG, but was barely detectable in the mutant DG (a,c). Each lane
represents amplification from an individual mouse. Primers were designed
to exclude amplification of genomic DNA. Gapdh was used for an internal
control. (d,e) b-galactosidase activity from Ptclacz/+ locus revealed active and
defective Shh signaling in the WT (d) and mutant DG (e), respectively.
Arrows indicate the DG. Scale bar, 100 mm.
Hem
atox
ylin
Brd
UD
AP
IB
rdU
DA
PI
GFA
P, P
SA
-NC
AM
DA
PI
WT Smo mutant
140
DE MS DG
GNP
E18.5120100806040200B
rdU
+ c
ells
per
sec
tion
DG MS DE
WT
Smo mutant
**
P21
P21
P21
E18.5
a b
c d
e f
g h
i j
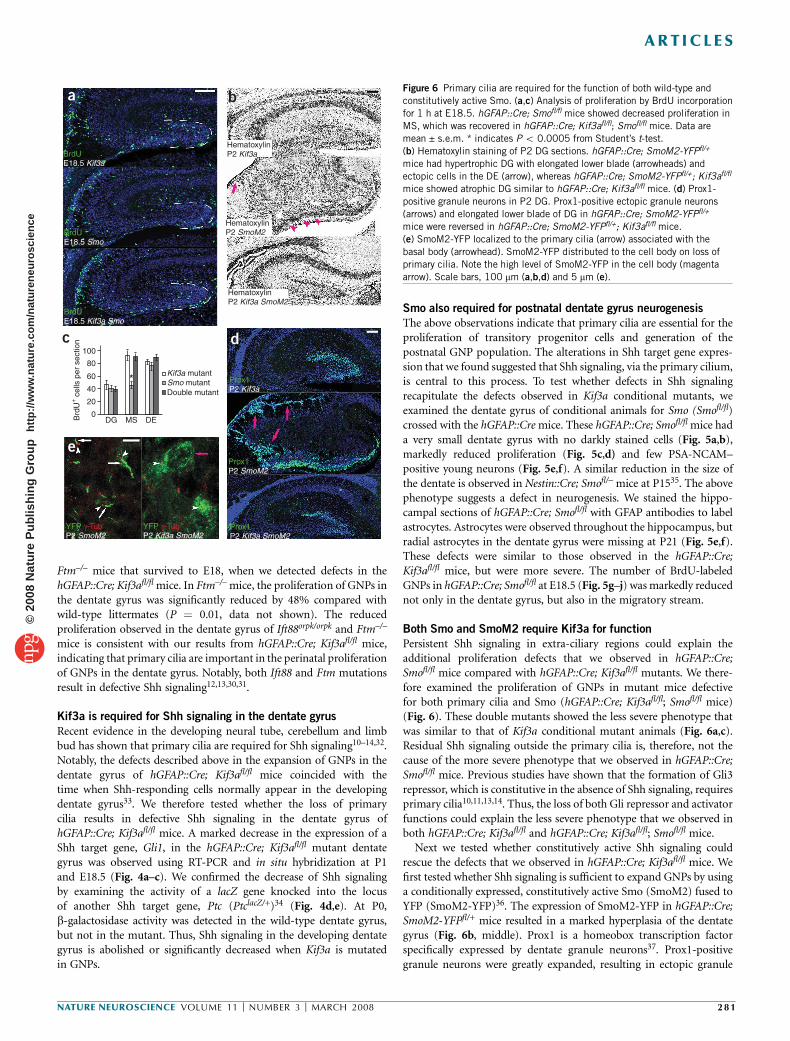
Figure 5 hGFAP::Cre; Smofl/fl mice show defects in dentate gyrus
development that resemble hGFAP::Cre; Kif3afl/fl mice. (a,b) Hematoxylin
staining showed that hGFAP::Cre; Smofl/fl mutant mice have small and
disorganized DG (b) similar to hGFAP::Cre; Kif3afl/fl mice (Fig. 1d). Insets
show a low-magnification view of the hippocampus. Arrows indicate darkly
stained type D cells in the WT SGZ. (c,d) BrdU incorporation for 1 h revealed
a lack of dividing cells in the mutant SGZ at P21. Arrow indicates a BrdU-
positive cell in WT. (e,f) The mutant DG had few PSA-NCAM–positive youngneurons (yellow arrow) and no GFAP-positive radial astrocytes (white arrow).
As in hGFAP::Cre; Kif3afl/fl mice, GFAP-positive astrocytes in non-neurogenic
locations (arrowheads) were still present in hGFAP::Cre; Smofl/fl mice. (g–j)
Analysis of proliferation by BrdU incorporation for 1 h at E18.5 (g,h) and an
illustration identifying DE, MS and DG (i). Dashed lines indicate the outline
of the DG. Proliferation was significantly decreased in the mutant DG and MS
compared with WT (j). Arrows indicate BrdU-positive cells. Data are mean
± s.e.m. * indicates P o 0.000005 from Student’s t-test. Scale bar, 100 mm.
280 VOLUME 11 [ NUMBER 3 [ MARCH 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
Ftm–/– mice that survived to E18, when we detected defects in thehGFAP::Cre; Kif3afl/fl mice. In Ftm–/– mice, the proliferation of GNPs inthe dentate gyrus was significantly reduced by 48% compared withwild-type littermates (P ¼ 0.01, data not shown). The reducedproliferation observed in the dentate gyrus of Ift88orpk/orpk and Ftm–/–
mice is consistent with our results from hGFAP::Cre; Kif3afl/fl mice,indicating that primary cilia are important in the perinatal proliferationof GNPs in the dentate gyrus. Notably, both Ift88 and Ftm mutationsresult in defective Shh signaling12,13,30,31.
Kif3a is required for Shh signaling in the dentate gyrus
Recent evidence in the developing neural tube, cerebellum and limbbud has shown that primary cilia are required for Shh signaling10–14,32.Notably, the defects described above in the expansion of GNPs in thedentate gyrus of hGFAP::Cre; Kif3afl/fl mice coincided with thetime when Shh-responding cells normally appear in the developingdentate gyrus33. We therefore tested whether the loss of primarycilia results in defective Shh signaling in the dentate gyrus ofhGFAP::Cre; Kif3afl/fl mice. A marked decrease in the expression of aShh target gene, Gli1, in the hGFAP::Cre; Kif3afl/fl mutant dentategyrus was observed using RT-PCR and in situ hybridization at P1and E18.5 (Fig. 4a–c). We confirmed the decrease of Shh signalingby examining the activity of a lacZ gene knocked into the locusof another Shh target gene, Ptc (PtclacZ/+)34 (Fig. 4d,e). At P0,b-galactosidase activity was detected in the wild-type dentate gyrus,but not in the mutant. Thus, Shh signaling in the developing dentategyrus is abolished or significantly decreased when Kif3a is mutatedin GNPs.
Smo also required for postnatal dentate gyrus neurogenesis
The above observations indicate that primary cilia are essential for theproliferation of transitory progenitor cells and generation of thepostnatal GNP population. The alterations in Shh target gene expres-sion that we found suggested that Shh signaling, via the primary cilium,is central to this process. To test whether defects in Shh signalingrecapitulate the defects observed in Kif3a conditional mutants, weexamined the dentate gyrus of conditional animals for Smo (Smofl/fl)crossed with the hGFAP::Cre mice. These hGFAP::Cre; Smofl/fl mice hada very small dentate gyrus with no darkly stained cells (Fig. 5a,b),markedly reduced proliferation (Fig. 5c,d) and few PSA-NCAM–positive young neurons (Fig. 5e,f). A similar reduction in the size ofthe dentate is observed in Nestin::Cre; Smofl/– mice at P1535. The abovephenotype suggests a defect in neurogenesis. We stained the hippo-campal sections of hGFAP::Cre; Smofl/fl with GFAP antibodies to labelastrocytes. Astrocytes were observed throughout the hippocampus, butradial astrocytes in the dentate gyrus were missing at P21 (Fig. 5e,f).These defects were similar to those observed in the hGFAP::Cre;Kif3afl/fl mice, but were more severe. The number of BrdU-labeledGNPs in hGFAP::Cre; Smofl/fl at E18.5 (Fig. 5g–j) was markedly reducednot only in the dentate gyrus, but also in the migratory stream.
Both Smo and SmoM2 require Kif3a for function
Persistent Shh signaling in extra-ciliary regions could explain theadditional proliferation defects that we observed in hGFAP::Cre;Smofl/fl mice compared with hGFAP::Cre; Kif3afl/fl mutants. We there-fore examined the proliferation of GNPs in mutant mice defectivefor both primary cilia and Smo (hGFAP::Cre; Kif3afl/fl; Smofl/fl mice)(Fig. 6). These double mutants showed the less severe phenotype thatwas similar to that of Kif3a conditional mutant animals (Fig. 6a,c).Residual Shh signaling outside the primary cilia is, therefore, not thecause of the more severe phenotype that we observed in hGFAP::Cre;Smofl/fl mice. Previous studies have shown that the formation of Gli3repressor, which is constitutive in the absence of Shh signaling, requiresprimary cilia10,11,13,14. Thus, the loss of both Gli repressor and activatorfunctions could explain the less severe phenotype that we observed inboth hGFAP::Cre; Kif3afl/fl and hGFAP::Cre; Kif3afl/fl; Smofl/fl mice.
Next we tested whether constitutively active Shh signaling couldrescue the defects that we observed in hGFAP::Cre; Kif3afl/fl mice. Wefirst tested whether Shh signaling is sufficient to expand GNPs by usinga conditionally expressed, constitutively active Smo (SmoM2) fused toYFP (SmoM2-YFP)36. The expression of SmoM2-YFP in hGFAP::Cre;SmoM2-YFPfl/+ mice resulted in a marked hyperplasia of the dentategyrus (Fig. 6b, middle). Prox1 is a homeobox transcription factorspecifically expressed by dentate granule neurons37. Prox1-positivegranule neurons were greatly expanded, resulting in ectopic granule
100
80
60
40
20
0DGB
rdU
+ c
ells
per
sec
tion
MS DE
*Kif3a mutantSmo mutantDouble mutant
BrdUE18.5 Kif3a
BrdUE18.5 Kif3a Smo
Prox1P2 Kif3a
Prox1P2 SmoM2
Prox1P2 Kif3a SmoM2
YFP γ-TubP2 Kif3a SmoM2
YFP γ-TubP2 SmoM2
BrdUE18.5 Smo
HematoxylinP2 Kif3a
HematoxylinP2 SmoM2
HematoxylinP2 Kif3a SmoM2
a b
c d
e
Figure 6 Primary cilia are required for the function of both wild-type and
constitutively active Smo. (a,c) Analysis of proliferation by BrdU incorporation
for 1 h at E18.5. hGFAP::Cre; Smofl/fl mice showed decreased proliferation in
MS, which was recovered in hGFAP::Cre; Kif3afl/fl; Smofl/fl mice. Data are
mean ± s.e.m. * indicates P o 0.0005 from Student’s t-test.
(b) Hematoxylin staining of P2 DG sections. hGFAP::Cre; SmoM2-YFPfl/+
mice had hypertrophic DG with elongated lower blade (arrowheads) and
ectopic cells in the DE (arrow), whereas hGFAP::Cre; SmoM2-YFPfl/+; Kif3afl/fl
mice showed atrophic DG similar to hGFAP::Cre; Kif3afl/fl mice. (d) Prox1-
positive granule neurons in P2 DG. Prox1-positive ectopic granule neurons
(arrows) and elongated lower blade of DG in hGFAP::Cre; SmoM2-YFPfl/+
mice were reversed in hGFAP::Cre; SmoM2-YFPfl/+; Kif3afl/fl mice.
(e) SmoM2-YFP localized to the primary cilia (arrow) associated with the
basal body (arrowhead). SmoM2-YFP distributed to the cell body on loss of
primary cilia. Note the high level of SmoM2-YFP in the cell body (magenta
arrow). Scale bars, 100 mm (a,b,d) and 5 mm (e).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 3 [ MARCH 2008 281
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

neurons in ventral and lateral hippocampus (Fig. 6d, middle). Further-more, morphogenesis of the dentate gyrus was abnormal, as granuleneurons expanded laterally and did not fold into the characteristicV-shaped granular layer. To our surprise, removal of Kif3a in cellsexpressing SmoM2-YFP (hGFAP::Cre; Kif3afl/fl; SmoM2-YFPfl/+)reversed the hypertrophic defects in histogenesis, resulting in anatrophic dentate gyrus similar to that observed in Kif3a conditionalmutants (Fig. 6b,d). Thus, constitutively active Smo was not able torescue the defects in hGFAP::Cre; Kif3afl/fl mice and required Kif3a forits action. A previous study showed that Smo localizes to primarycilia on Shh treatment, but SmoM2 localizes to primary cilia indepen-dent of Shh in cultured MDCK cells15. We found that SmoM2-YFP alsolocalized to primary cilia in hGFAP::Cre; SmoM2-YFPfl/+ mice in vivo,but distributed to the cell body on removal of primary cilia inhGFAP::Cre; Kif3afl/fl; SmoM2-YFPfl/+ mice (Fig. 6e).
DISCUSSION
Shh has been shown to be important for the proliferation andmaintenance of adult neural stem cells in the SGZ35,38. In the presentstudy, we have shown that Shh signaling is essential for expandingGNPs during perinatal development to establish the adult stem cellpopulation. These effects of Shh signaling required Kif3a, an essentialmotor for assembling primary cilia. The conditional removal of Kif3ain a subset of hippocampal progenitors resulted in defective Shhsignaling in GNPs, leading to defective proliferation. In these mice,the earliest populations of GNPs were specified, but during later stagesof perinatal development, GNPs failed to expand and to generate radialastrocytes that function as the SGZ stem cells. Notably, other types ofastrocytes in the dentate gyrus were produced. Thus, Kif3a is requiredfor the production or maintenance of radial astrocytes in the SGZ, butnot for the production or differentiation of other astrocytes in thehippocampus. This suggests a fundamental difference between non-germinal astrocytes and astrocytes that function as primary progenitorsin the adult brain.
The removal of Smo in the same GNP population resulted in asimilar, but more severe, phenotype, and this severe phenotype wasameliorated on removal of both Kif3a and Smo, confirming that Shhacts through primary cilia to expand GNPs and to convert them toadult progenitors. Proliferation in the other major adult germinal layer,the subventricular zone, also showed a reduction in the hGFAP::Cre;Kif3afl/fl mice (data not shown), but this reduction was not as large asthat observed in the SGZ of the hippocampus. Shh signaling andprimary cilia may be particularly important for the expansion andestablishment of germinal centers away from the ventricular zone, as isthe case in the SGZ. Consistently, in hGFAP::Cre; Kif3afl/fl andhGFAP::Cre; Smofl/fl mice, we did not observe major defects in neuronallayers that are derived directly from proliferating ventricular zone cells,such as the cortex and Ammon’s horn of the hippocampus, but we andothers32 observed a major deficit in the expansion of cerebellar granuleneuron precursors, which also occurs away from the ventricular zone(N.S., Y.-G.H., A.A., L. Strehl, M.R.-R., J.-M.G.-V. and A.A.-B.,unpublished data).
Analyses of double mutants for both Kif3a and Smo demonstratethat Kif3a acts downstream of Smo and that Kif3a is required for all Shhsignaling in the developing dentate gyrus. Consistent with our geneticdata, mutant Smo protein that fails to localize to cilia is unable toactivate Shh signaling15. Notably, our results show that even constitu-tively active Smo (SmoM2) requires primary cilia for its function. WhySmoM2 is constitutively active in the absence of Shh is unknown. Ourdata indicate that, similar to wild-type Smo, SmoM2 signals throughthe primary cilia. Recent work has shown that the Shh receptor, Ptc,
localizes to primary cilia, where it binds to Shh16. On binding to Shh,Ptc exits the primary cilia and Smo accumulates there, resulting insignaling. The mutation in SmoM2 may target this protein to thecilium independent of Ptc or Shh, and this localization may besufficient to promote constitutive signaling.
Why do vertebrate cells require this intricate localization of receptorsand other downstream components in a tiny projection from the cellsurface? The cilia’s small volume relative to the surface area couldallow high sensitivity for detection of low levels of extracellular signals.The primary cilium, although continuous from the cell body, is adistinct subcellular compartment, in which trafficking is restrictedby IFT. This restriction may allow the coordinated spread or move-ment of secondary messengers or effecter molecules. In addition, thejuxtaposition of the basal body and the Golgi complex at the baseof the cilia may facilitate rapid trafficking of molecules such as Smointo the cilia after receiving extracellular signals and could helpcoordinate the cell cycle.
We used a conditional mutant for Kif3a to remove primary cilia.Kinesin-II has been also suggested to participate in intracellulartrafficking and mitosis in cultured cells25–27. In cells dissociated fromthe dentate gyrus of hGFAP::Cre; Kif3afl/fl mice, we did not observeincreased incidence of intracellular trafficking or mitotic defects.Moreover, our genetic evidence indicates that kinesin-II function isessential for Shh signaling; loss of Kif3a partially rescued the severedefects that we observed when Smo was removed or when SmoM2 wasexpressed. This rescue cannot be explained if Kif3a had a generalfunction in mitosis or intracellular trafficking. Consistently, condi-tional removal of either Kif3a or Ift88 results in identical phenotypes inthree different organs, including brain18,32,39. In vivo, Kif3a appears tobe essential for the formation and function of primary cilia, but isdispensable for mitosis and intracellular trafficking. The discrepancybetween these and previous in vitro studies could be the result ofdifferences in the cell types used or of differences in the experimentalapproaches used to interfere with kinesin-II function. We andothers18,32,39 analyzed loss-of-function mutant cells, whereas previousin vitro studies used overexpression of dominant-negative subunits ofkinesin-II, which, as demonstrated in Chlamydomonas, could have off-target effects. In this flagellated unicellular organism, point mutationsin kinesin-II can cause defective mitosis, but null mutations causedefects only in flagellar assembly. Notably, mitotic defects inthese mutants are dominant, whereas flagellar assembly defects arerecessive40,41. This demonstrates that mitotic defects arise fromacquired aberrant or toxic functions of the mutant kinesin-II proteinin Chlamydomonas.
If kinesin-II had essential roles in mitosis or intracellular trafficking,generalized defects in our Kif3a mutants would be expected, but we didnot observe increased apoptosis or general hippocampal defects.Defects in the dentate gyrus were first detected 5 d after the onset ofCre activity (at E13) and were specific to GNPs in the dentate gyrus andnot in the dentate neuroepithelium or the migratory stream, suggestingthat the loss of primary cilia specifically affects GNP expansion in thedentate gyrus. This proliferation deficit coincides with the time whenShh signaling starts to occur in the developing dentate gyrus33. Notealso that mutations in Ftm and Ofd1, genes that are not linked to IFT,but are required for ciliogenesis, also cause defects in Shh signal-ing30,31,42. Lastly, we observed a substantial reduction in GNP prolife-ration in other ciliary mutants (Ift88orpk/orpk and Ftm–/–). Altogether,the results listed here indicate that the phenotype that we observed inhGFAP::Cre; Kif3afl/fl mice is not the result of defects in mitosis orintracellular transport, but of deficits in Shh signaling mediated bythe cilia.
282 VOLUME 11 [ NUMBER 3 [ MARCH 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

Our work does not eliminate the possibility that Kif3a, Ift88 or Ftm,could also have functions outside of the primary cilia. Kinesin-IIhas been shown to localize to axons and synapses in neurons43,44.Therefore, disruption of other cellular functions of kinesin-II couldhave contributed to the phenotypes observed in hGFAP::Cre; Kif3afl/fl
mice. The results from Kif3a and Smo double mutants indicate that anypossible nonciliary function for kinesin-II must also be involved in Shhsignaling. Mutations in different ciliogenic genes (Kif3a, Dnchc2, Ift52,Ift88, Ift172, Ftm and Ofd1)10–14,30,31,42 all result in Shh signalingdefects. Although we cannot totally exclude that Kif3a, Ift88 or Ftmcould have some role outside of the primary cilia, cumulative evidencepoints to this organelle as an essential component of Shh signaling.
Mental retardation is a prominent feature of several systemicdisorders associated with dysfunctional cilia17. Our findings showingthat primary cilia are essential for hippocampal development couldhelp to elucidate the molecular etiology of mental retardation found inthese diseases. The present results show that dentate gyrus progenitorsrequire primary cilia to mediate of Shh signaling at a critical stage oftheir development, when they are transitioning and expanding fromembryonic to postnatal neural stem cells.
METHODSMice. All animal procedures were approved by the University of California, San
Francisco Institutional Animal Care and Use Committee. The genetically
modified mice used in the study, hGFAP::Cre23, Kif3afl/fl (ref. 45), PtclacZ
(ref. 34), Smofl/fl (ref. 46), SmoM2-YFPfl/fl (ref. 36), Ift88orpk/orpk (ref. 29) and
Ftm–/– (ref. 30), have been described previously.
Tissue preparation and staining. Mice were injected with BrdU (50 mg
per kg of body weight) and perfused 1 h later with 4% paraformaldehyde
(w/v). The brains were postfixed overnight at 4 1C. Embryonic brains were
fixed in the same fixative overnight at 4 1C. For frozen sections, brains were
cryoprotected in 30% sucrose, embedded in Tissue-Tek Optimal Cutting
Temperature compound (Sakura) and cut into 12-mm sections. For poly-
ethylene glycol (PEG) sections, brains were dehydrated in an ethanol
series, embedded in a 2:1 mixture of PEG 1,000 and PEG 1,500, and cut into
10-mm sections. For immunostaining, sections were incubated with primary
antibodies overnight at 4 1C, followed by incubation with secondary antibodies
at 20–23.5 1C for 2 h. For X-gal staining and electron microscopy, brains were
perfused with 2% paraformaldehyde and 2.5% glutaraldehyde and postfixed in
2.5% glutaraldehyde overnight at 4 1C. Coronal 300-mm sections were cut on a
vibratome. The sections were postfixed in 2% osmium for 2 h, dehydrated and
embedded in Araldite (Durcupan, Fluka Biochemica). Semi-thin (1.5 mm) and
ultra-thin (0.06 mm) sections were cut with a diamond knife and stained with
1% toluidine blue and lead citrate, respectively.
Images were taken with an Olympus AX70 light microscope, a Leica
SP2 confocal microscope or a Jeol electron microscope, and processed using
Adobe Photoshop.
Antibodies and reagents. The following primary antibodies were used: rabbit
antibody to GFAP (1:300, DAKO), mouse antibody to Cre (1:500, Abcam), rat
antibody to BrdU (1:10, Oxford Biotech), rabbit antibody to Ki67 (1:1,000,
Novo Castra Labs), mouse antibody to PSA-NCAM (1:400, AbCys), mouse
antibody to Mash1 (1:200, Pharmingen), mouse antibody to acetylated tubulin
(1:1,000, Sigma), rabbit antibody to g-tubulin (1:1,000, Sigma), rabbit antibody
to Prox1 (1:2,500, gift from S. Pleasure, University of California San Francisco),
rabbit antibody to cleaved caspase-3 (1:50, Cell Signaling Technology), chicken
antibody to GFP (1:500, Aves Labs) and mouse antibody to Lamp1 (1:100,
Stressgen). Antibody to GFP was used to visualize the localization of SmoM2-
YFP. For fluorescence staining, sections were incubated with Alexa Fluor–
conjugated secondary antibodies (1:400, Molecular Probes) and counter stained
with DAPI (100 ng ml–1). For enzymatic staining, biotin-conjugated secondary
antibodies (1:200, Jackson Lab) and ABC reaction (Vector Laboratories) were
used. Standard hematoxylin staining was used for counterstaining. FITC-
conjugated HP lectin was used to stain Golgi (0.1 mg ml–1, Sigma).
Cell counting. The immunostained cells were counted in at least three
sections from each mouse at comparable rostrocaudal levels. Data from at
least three mice per each group were pooled for statistical analysis with
Student’s t-test.
RT-PCR. Total RNA was isolated from the dentate gyrus of individual P1 mice
using Qiagen RNeasy Mini Kit and was reverse transcribed by Superscript II
reverse transcriptase (Invitrogen). Both Gli1 and Gapdh cDNAs were amplified
from individual mice by PCR in a single reaction.
Cell culture. Cells were dissociated from the dentate gyrus of P1 mice using
papain (0.6 mg ml–1, Worthington), followed by trituration, resuspended in
DMEM and F12 (50:50) medium supplemented with N2 (Gibco) and B27
(Gibco), and plated in chamber slides coated with poly-D-lysine and laminin.
In situ hybridization. Standard in situ protocol and antisense riboprobe from
Gli1 cDNA (gift from A. Ruiz i Altaba, University of Geneva Medical School)
were used.
Note: Supplementary information is available on the Nature Neuroscience website.
ACKNOWLEDGMENTSWe are grateful to L.S. Goldstein for providing us with the Kif3a conditionalmutant mice, N. Murcia for Ift88orpk mice, and J. Reiter and D. Rowitch for Smoand SmoM2 conditional mutant mice and PtclacZ/+ mice. We thank R. Ihrie,D. Lim, S. Pleasure, J. Reiter and C. Yaschine on the manuscript. The work wassupported by a Mark Linder/American Brain Tumor Association Fellowship toY.-G.H. and by grants (NS28478 and HD32116) from the US National Institutesof Health and a grant from the Goldhirsh foundation to A.A.-B. N.S. wassupported by the Human Frontier Science Program and the Agence Nationale dela Recherche. Confocal microscopy at Diabetes Endocrinology Research CenterMicroscopy and Imaging Core was supported by an US National Institute ofHealth grant (P30 DK063720).
AUTHOR CONTRIBUTIONSY.-G.H. designed and performed most of the experiments. M.R.-R. andJ.-M.G.-V. carried out the electron microscopic analyses. N.S. assisted with theinitial analysis of hGFAP::Cre; Kif3afl/fl mice. N.S. and A.A. analyzed the Ftm–/–
mice. S.S.-M. provided the Ftm–/– mice. A.A.-B. supervised the project.Y.-G.H. and A.A.-B. wrote the manuscript.
Published online at http://www.nature.com/natureneuroscience
Reprints and permissions information is available online at http://npg.nature.com/
reprintsandpermissions
1. Altman, J. & Bayer, S.A. Migration and distribution of two populations of hippocampalgranule cell precursors during the perinatal and postnatal periods. J. Comp. Neurol.301,365–381 (1990).
2. Li, G. & Pleasure, S.J. Morphogenesis of the dentate gyrus: what we are learning frommouse mutants. Dev. Neurosci. 27, 93–99 (2005).
3. Filippov, V. et al. Subpopulation of nestin-expressing progenitor cells in the adult murinehippocampus shows electrophysiological and morphological characteristics of astro-cytes. Mol. Cell. Neurosci. 23, 373–382 (2003).
4. Fukuda, S. et al. Two distinct subpopulations of nestin-positive cells in adult mousedentate gyrus. J. Neurosci. 23, 9357–9366 (2003).
5. Seri, B., Garcia-Verdugo, J.M., McEwen, B.S. & Alvarez-Buylla, A. Astrocytes give rise tonew neurons in the adult mammalian hippocampus. J. Neurosci. 21, 7153–7160(2001).
6. Rosenbaum, J.L. & Witman, G.B. Intraflagellar transport. Nat. Rev. Mol. Cell Biol. 3,813–825 (2002).
7. Davis, E.E., Brueckner, M. & Katsanis, N. The emerging complexity of thevertebrate cilium: new functional roles for an ancient organelle. Dev. Cell 11, 9–19(2006).
8. Scholey, J.M. & Anderson, K.V. Intraflagellar transport and cilium-based signaling.Cell 125, 439–442 (2006).
9. Singla, V. & Reiter, J.F. The primary cilium as the cell’s antenna: signaling at a sensoryorganelle. Science 313, 629–633 (2006).
10. Haycraft, C.J. et al. Gli2 and Gli3 localize to cilia and require the intraflagellar transportprotein polaris for processing and function. PLoS Genet. 1, e53 (2005).
11. Huangfu, D. & Anderson, K.V. Cilia and Hedgehog responsiveness in the mouse.Proc. Natl. Acad. Sci. USA 102, 11325–11330 (2005).
12. Huangfu, D. et al. Hedgehog signaling in the mouse requires intraflagellar transportproteins. Nature 426, 83–87 (2003).
13. Liu, A., Wang, B. & Niswander, L.A. Mouse intraflagellar transport proteins regulate boththe activator and repressor functions of Gli transcription factors. Development 132,3103–3111 (2005).
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 3 [ MARCH 2008 283
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

14. May, S.R. et al. Loss of the retrograde motor for IFT disrupts localization of Smo to ciliaand prevents the expression of both activator and repressor functions of Gli. Dev. Biol.287, 378–389 (2005).
15. Corbit, K.C. et al. Vertebrate Smoothened functions at the primary cilium. Nature 437,1018–1021 (2005).
16. Rohatgi, R., Milenkovic, L. & Scott, M.P. Patched1 regulates hedgehog signaling at theprimary cilium. Science 317, 372–376 (2007).
17. Badano, J.L., Mitsuma, N., Beales, P.L. & Katsanis, N. The ciliopathies: an emergingclass of human genetic disorders. Annu. Rev. Genomics Hum. Genet. 7, 125–148(2006).
18. Davenport, J.R. et al. Disruption of intraflagellar transport in adult mice leads to obesityand slow-onset cystic kidney disease. Curr. Biol. 17, 1586–1594 (2007).
19. Alvarez-Buylla, A., Garcia-Verdugo, J.M. & Tramontin, A.D. A unified hypothesis on thelineage of neural stem cells. Nat. Rev. Neurosci. 2, 287–293 (2001).
20. Doetsch, F., Garcia-Verdugo, J.M. & Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalianbrain. J. Neurosci. 17, 5046–5061 (1997).
21. Kozminski, K.G., Beech, P.L. & Rosenbaum, J.L. The Chlamydomonas kinesin-likeprotein FLA10 is involved in motility associated with the flagellar membrane. J. CellBiol. 131, 1517–1527 (1995).
22. Rickmann, M., Amaral, D.G. & Cowan, W.M. Organization of radial glial cells during thedevelopment of the rat dentate gyrus. J. Comp. Neurol. 264, 449–479 (1987).
23. Zhuo, L. et al. hGFAP-cre transgenic mice for manipulation of glial and neuronal functionin vivo. Genesis 31, 85–94 (2001).
24. Pleasure, S.J., Collins, A.E. & Lowenstein, D.H. Unique expression patterns of cell fatemolecules delineate sequential stages of dentate gyrus development. J. Neurosci. 20,6095–6105 (2000).
25. Brown, C.L. et al. Kinesin-2 is a motor for late endosomes and lysosomes. Traffic 6,1114–1124 (2005).
26. Le Bot, N., Antony, C., White, J., Karsenti, E. & Vernos, I. Role of xklp3, a subunit of theXenopus kinesin II heterotrimeric complex, in membrane transport between the endo-plasmic reticulum and the Golgi apparatus. J. Cell Biol. 143, 1559–1573 (1998).
27. Haraguchi, K., Hayashi, T., Jimbo, T., Yamamoto, T. & Akiyama, T. Role of the kinesin-2family protein, KIF3, during mitosis. J. Biol. Chem. 281, 4094–4099 (2006).
28. Murcia, N.S. et al. The Oak Ridge Polycystic Kidney (orpk) disease gene is required forleft-right axis determination. Development 127, 2347–2355 (2000).
29. Moyer, J.H. et al. Candidate gene associated with a mutation causing recessivepolycystic kidney disease in mice. Science 264, 1329–1333 (1994).
30. Delous, M. et al. The ciliary gene RPGRIP1L is mutated in cerebello-oculo-renalsyndrome (Joubert syndrome type B) and Meckel syndrome. Nat. Genet. 39,875–881 (2007).
31. Vierkotten, J., Dildrop, R., Peters, T., Wang, B. & Ruther, U. Ftm is a novel basal bodyprotein of cilia involved in Shh signalling. Development 134, 2569–2577 (2007).
32. Chizhikov, V.V. et al. Cilia proteins control cerebellar morphogenesis by promotingexpansion of the granule progenitor pool. J. Neurosci. 27, 9780–9789 (2007).
33. Ahn, S. & Joyner, A.L. In vivo analysis of quiescent adult neural stem cells responding toSonic hedgehog. Nature 437, 894–897 (2005).
34. Goodrich, L.V., Milenkovic, L., Higgins, K.M. & Scott, M.P. Altered neural cell fates andmedulloblastoma in mouse patched mutants. Science 277, 1109–1113 (1997).
35. Machold, R. et al. Sonic hedgehog is required for progenitor cell maintenance intelencephalic stem cell niches. Neuron 39, 937–950 (2003).
36. Mao, J. et al. A novel somatic mouse model to survey tumorigenic potential applied tothe Hedgehog pathway. Cancer Res. 66, 10171–10178 (2006).
37. Bagri, A. et al. The chemokine SDF1 regulates migration of dentate granule cells.Development 129, 4249–4260 (2002).
38. Lai, K., Kaspar, B.K., Gage, F.H. & Schaffer, D.V. Sonic hedgehog regulates adult neuralprogenitor proliferation in vitro and in vivo. Nat. Neurosci. 6, 21–27 (2003).
39. Haycraft, C.J. et al. Intraflagellar transport is essential for endochondral bone formation.Development 134, 307–316 (2007).
40. Matsuura, K., Lefebvre, P.A., Kamiya, R. & Hirono, M. Kinesin-II is not essential formitosis and cell growth in Chlamydomonas. Cell Motil. Cytoskeleton 52, 195–201(2002).
41. Miller, M.S. et al.Mutant kinesin-2 motor subunits increase chromosome loss.Mol. Biol.Cell 16, 3810–3820 (2005).
42. Ferrante, M.I. et al. Oral-facial-digital type I protein is required for primary ciliaformation and left-right axis specification. Nat. Genet. 38, 112–117 (2006).
43. Kondo, S. et al. KIF3A is a new microtubule-based anterograde motor in the nerve axon.J. Cell Biol. 125, 1095–1107 (1994).
44. Muresan, V., Lyass, A. & Schnapp, B.J. The kinesin motor KIF3A is a componentof the presynaptic ribbon in vertebrate photoreceptors. J. Neurosci. 19, 1027–1037(1999).
45. Marszalek, J.R. et al. Genetic evidence for selective transport of opsin and arrestin bykinesin-II in mammalian photoreceptors. Cell 102, 175–187 (2000).
46. Long, F., Zhang, X.M., Karp, S., Yang, Y. & McMahon, A.P. Genetic manipulation ofhedgehog signaling in the endochondral skeleton reveals a direct role in the regulation ofchondrocyte proliferation. Development 128, 5099–5108 (2001).
284 VOLUME 11 [ NUMBER 3 [ MARCH 2008 NATURE NEUROSCIENCE
ART ICLES©
2008
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
Related Documents











