Current Biology 23, 2434–2439, December 2, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.10.057 Report Heart-Specific Stiffening in Early Embryos Parallels Matrix and Myosin Expression to Optimize Beating Stephanie Majkut, 1,2 Timon Idema, 2,4 Joe Swift, 1 Christine Krieger, 1 Andrea Liu, 2 and Dennis E. Discher 1,2,3, * 1 Biophysical Engineering Laboratory, University of Pennsylvania, Philadelphia, PA 19104, USA 2 Physics and Astronomy Graduate Group, University of Pennsylvania, Philadelphia, PA 19104, USA 3 Cell and Molecular Biology Graduate Group, University of Pennsylvania, Philadelphia, PA 19104, USA 4 Department of Bionanoscience, Kavli Institute of Nanoscience, Delft University of Technology, Lorentzweg 1, 2628 CJ Delft, the Netherlands Summary In development and differentiation, morphological changes often accompany mechanical changes [1], but it is unclear whether or when cells in embryos sense tissue elasticity. The earliest embryo is uniformly pliable, while adult tissues vary widely in mechanics from soft brain and stiff heart to rigid bone [2]. However, cell sensitivity to microenvironment elasticity is debated based in part on results from complex three-dimensional culture models [3]. Regenerative cardiol- ogy provides strong motivation to clarify any cell-level sensi- tivities to tissue elasticity because rigid postinfarct regions limit pumping by the adult heart [4]. Here, we focus on the spontaneously beating embryonic heart and sparsely cultured cardiomyocytes, including cells derived from pluripotent stem cells. Tissue elasticity, E t , increases daily for heart to 1–2 kPa by embryonic day 4 (E4), and although this is w10-fold softer than adult heart, the beating contrac- tions of E4 cardiomyocytes prove optimal at wE t,E4 both in vivo and in vitro. Proteomics reveals daily increases in a small subset of proteins, namely collagen plus cardiac- specific excitation-contraction proteins. Rapid softening of the heart’s matrix with collagenase or stiffening it with enzy- matic crosslinking suppresses beating. Sparsely cultured E4 cardiomyocytes on collagen-coated gels likewise show maximal contraction on matrices with native E4 stiffness, highlighting cell-intrinsic mechanosensitivity. While an optimal elasticity for striation proves consistent with the mathematics of force-driven sarcomere registration, con- traction wave speed is linear in E t as theorized for excita- tion-contraction coupled to matrix elasticity. Pluripotent stem cell-derived cardiomyocytes also prove to be mechano- sensitive to matrix and thus generalize the main observation that myosin II organization and contractile function are opti- mally matched to the load contributed by matrix elasticity. Results and Discussion The heart is the first functional organ in vertebrate embryos. It beats spontaneously as a heart tube by w36 hr after fertiliza- tion (Figure 1A). Subsequent stiffening has been described thus far in terms of changes in cell volume, hyaluronic acid, and/or collagen I[5], but functional evaluations of such changes have been lacking. Cardiomyocytes isolated from either late embryos [6, 7] or neonates [8–10] and cultured on substrates of varied stiffness suggest that gels that are stiffer than adult heart suppress contraction. Extremely soft sub- strates suppress sarcomere organization and limit contractile ability, with additional evidence of altered cytoskeletal confor- mation and assembly [6] in the absence of changes in mecha- nosensitive degradation [11] or transcription [12] that are evident in other cells. Mature cells cultured on gels can thus exhibit an optimal stiffness for contraction, but relevance of matrix elasticity to intact heart remains unclear and controver- sial based on a handful of culture studies (e.g., [3]). Heart Stiffens with Expression of Excitation-Contraction and Collagen Proteins, whereas Brain Remains Soft Tissue aspiration into micropipettes (Figure 1B) of diameter sufficient to probe dozens of cells plus extracellular matrix [1] shows that heart at all stages behaves elastically (Figure 1C), whereas midbrain tissue and embryonic disk flow over minutes and fail to recover fully after release of the applied stress. The effective Young’s modulus of each tissue, E t , was calculated from the slope of aspiration pressure versus aspirated length (see Figure S1A available online) [13], and for brain and embry- onic disc, the (already large) aspirated length at 2 min was used. By E2, the presumptive ventricle is already 3-fold stiffer than undifferentiated embryonic disc and embryonic brain. The latter remains roughly constant through development at 0.3 6 0.2 kPa (Figure 1D) and matches adult brain stiffness [14]. Brain tissue is thus always soft, whereas heart stiffens up to about 10-fold to reach neonate and adult heart stiffness by wE14 [6]. Modest stiffness variations of 620% along the heart tube (Figure S1B) are also consistent with past reports [15]. Expression trends for at least some tissue proteins seemed likely to parallel the divergent trends in tissue mechanics and to confer tissue stiffness. Quantitative mass spectrometry of extracts from embryonic discs and E2–E4 and E10 heart and brain tissue identified over 200 diverse proteins (Table S1), of which fewer than 10% followed trends in expression similar to those of E t (Figures 1D–1H, S1C, and S1D). Most trend- following proteins related closely to the excitation-contraction coupling system, such as cardiac actomyosin contractile proteins, adhesion proteins, and the sarcoplasmic calcium channel SERCA. Mitochondrial proteins (relevant to ener- getics) also followed the trends, but with smaller differences between heart and brain. Proteins that were notably uncorre- lated with tissue stiffness included many nuclear proteins, intermediate filament proteins, and nonmuscle myosin. Of two extracellular matrix (ECM) proteins detected, fibronectin and collagen I, only collagen I follows the E t trends. To begin to assess stiffness contributions of the actomyosin cytoskeleton or collagen, we inhibited myosin contractility with the myosin II ATPase inhibitor blebbistatin or else disrup- ted the collagenous ECM with mild collagenase treatments and then measured tissue stiffness. With blebbistatin, heart tissue from E2 to E14 is softened by w25% and brain tissue by w50% (Figure S1E). In contrast, collagenase had no signif- icant effect on brain tissue but considerably softened both *Correspondence: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Heart-Specific Stiffening in E
Current Biology 23, 2434–2439, December 2, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.10.057
Reportarly
Embryos Parallels Matrix and MyosinExpression to Optimize Beating
Stephanie Majkut,1,2 Timon Idema,2,4 Joe Swift,1
Christine Krieger,1 Andrea Liu,2 and Dennis E. Discher1,2,3,*1Biophysical Engineering Laboratory, University ofPennsylvania, Philadelphia, PA 19104, USA2Physics and Astronomy Graduate Group, University ofPennsylvania, Philadelphia, PA 19104, USA3Cell and Molecular Biology Graduate Group, University ofPennsylvania, Philadelphia, PA 19104, USA4Department of Bionanoscience, Kavli Institute ofNanoscience, Delft University of Technology, Lorentzweg 1,2628 CJ Delft, the Netherlands
Summary
In development and differentiation, morphological changesoften accompany mechanical changes [1], but it is unclear
whether or when cells in embryos sense tissue elasticity.The earliest embryo is uniformly pliable, while adult tissues
vary widely in mechanics from soft brain and stiff heart torigid bone [2]. However, cell sensitivity to microenvironment
elasticity is debated based in part on results from complexthree-dimensional culture models [3]. Regenerative cardiol-
ogy provides strongmotivation to clarify any cell-level sensi-tivities to tissue elasticity because rigid postinfarct regions
limit pumping by the adult heart [4]. Here, we focus onthe spontaneously beating embryonic heart and sparsely
cultured cardiomyocytes, including cells derived from
pluripotent stem cells. Tissue elasticity, Et, increases dailyfor heart to 1–2 kPa by embryonic day 4 (E4), and although
this is w10-fold softer than adult heart, the beating contrac-tions of E4 cardiomyocytes prove optimal at wEt,E4 both
in vivo and in vitro. Proteomics reveals daily increases ina small subset of proteins, namely collagen plus cardiac-
specific excitation-contraction proteins. Rapid softening ofthe heart’s matrix with collagenase or stiffening it with enzy-
matic crosslinking suppressesbeating. Sparsely culturedE4cardiomyocytes on collagen-coated gels likewise show
maximal contraction on matrices with native E4 stiffness,highlighting cell-intrinsic mechanosensitivity. While an
optimal elasticity for striation proves consistent with themathematics of force-driven sarcomere registration, con-
traction wave speed is linear in Et as theorized for excita-tion-contraction coupled to matrix elasticity. Pluripotent
stemcell-derived cardiomyocytes alsoprove to bemechano-sensitive to matrix and thus generalize the main observation
that myosin II organization and contractile function are opti-mally matched to the load contributed by matrix elasticity.
Results and Discussion
The heart is the first functional organ in vertebrate embryos. Itbeats spontaneously as a heart tube by w36 hr after fertiliza-tion (Figure 1A). Subsequent stiffening has been describedthus far in terms of changes in cell volume, hyaluronic acid,
*Correspondence: [email protected]
and/or collagen I [5], but functional evaluations of suchchanges have been lacking. Cardiomyocytes isolated fromeither late embryos [6, 7] or neonates [8–10] and cultured onsubstrates of varied stiffness suggest that gels that are stifferthan adult heart suppress contraction. Extremely soft sub-strates suppress sarcomere organization and limit contractileability, with additional evidence of altered cytoskeletal confor-mation and assembly [6] in the absence of changes in mecha-nosensitive degradation [11] or transcription [12] that areevident in other cells. Mature cells cultured on gels can thusexhibit an optimal stiffness for contraction, but relevance ofmatrix elasticity to intact heart remains unclear and controver-sial based on a handful of culture studies (e.g., [3]).
Heart Stiffens with Expression of Excitation-Contraction
and Collagen Proteins, whereas Brain Remains SoftTissue aspiration into micropipettes (Figure 1B) of diametersufficient to probe dozens of cells plus extracellular matrix [1]shows that heart at all stages behaves elastically (Figure 1C),whereasmidbrain tissue and embryonic disk flowoverminutesand fail to recover fully after release of the applied stress. Theeffective Young’s modulus of each tissue, Et, was calculatedfrom the slope of aspiration pressure versus aspirated length(see Figure S1A available online) [13], and for brain and embry-onicdisc, the (already large) aspirated lengthat 2minwasused.By E2, the presumptive ventricle is already 3-fold stiffer thanundifferentiated embryonic disc and embryonic brain. Thelatter remains roughly constant through development at 0.3 60.2 kPa (Figure 1D) andmatches adult brain stiffness [14]. Braintissue is thus always soft, whereas heart stiffens up to about10-fold to reach neonate and adult heart stiffness by wE14[6]. Modest stiffness variations of 620% along the heart tube(Figure S1B) are also consistent with past reports [15].Expression trends for at least some tissue proteins seemed
likely to parallel the divergent trends in tissue mechanics andto confer tissue stiffness. Quantitative mass spectrometry ofextracts from embryonic discs and E2–E4 and E10 heart andbrain tissue identified over 200 diverse proteins (Table S1), ofwhich fewer than 10% followed trends in expression similarto those of Et (Figures 1D–1H, S1C, and S1D). Most trend-following proteins related closely to the excitation-contractioncoupling system, such as cardiac actomyosin contractileproteins, adhesion proteins, and the sarcoplasmic calciumchannel SERCA. Mitochondrial proteins (relevant to ener-getics) also followed the trends, but with smaller differencesbetween heart and brain. Proteins that were notably uncorre-lated with tissue stiffness included many nuclear proteins,intermediate filament proteins, and nonmuscle myosin. Oftwo extracellular matrix (ECM) proteins detected, fibronectinand collagen I, only collagen I follows the Et trends.To begin to assess stiffness contributions of the actomyosin
cytoskeleton or collagen, we inhibited myosin contractilitywith the myosin II ATPase inhibitor blebbistatin or else disrup-ted the collagenous ECM with mild collagenase treatmentsand then measured tissue stiffness. With blebbistatin, hearttissue from E2 to E14 is softened by w25% and brain tissuebyw50% (Figure S1E). In contrast, collagenase had no signif-icant effect on brain tissue but considerably softened both

Heart Brain
∆P
0
3
6
0 200
E4 Heart
∆P = 0.9 kPaLRp
0 200
E4 Brain
∆P = 0.6 kPa
Time (s)
Embryo (E3) BA
L
Rp
Pin Pout
C
E
Rel
ativ
e ex
pres
sion
Embryo age (days)
0.1
10
1000
Cardiac Myosin II α-Actin
α-Actinin-2
Desmo-plakin
SERCA2
Myo.Bind.Prot.C
Talin Vimentin α, -Spectrin
0 100 100.1
1
10
0 10
1
100
0.1
1
10
100
0 10
Collagen I
Rel
. exp
ress
ion
Embryo age (days)
E4 E14decellularized
GAdult range
Adult range
0.1
1
10
0 2 4 6 14
tissue stiffnessHeartBrainEmbryonic disc
Et(k
Pa)
Embryo age (days)
0.3 kPa/day
0.3
3
D F
Cardiac Myosin II
Lamin-B1
MW
Car
d. M
yosi
n II
Lam
in-B
1
p < 0.05
2
1
0
260
8060
17 proteins follow �ssue trends for Et
H
E4 E6 E14
209 proteins quan�fied
Figure 1. Mechanical Development of Heart and Brain Tissue Parallels Expression of Abundant Cell and Matrix Proteins
(A) E3 chick embryo with heart tube (white box) and midbrain (blue box) in situ and after isolation. The heart continues to beat ex vivo, with contraction and
flow propagating along the dashed line. Scale bar represents 100 mm.
(B) Micropipette aspiration of an E3 heart tube in phase-contrast microscopy using pipettes that are much larger in diameter than single cells. Scale bar
represents 10 mm.
(C) Representative aspiration and relaxation curves for E4 heart and brain demonstrate the respective elastic and inelastic responses.
(D) Et for heart and brain tissue throughout embryonic development, starting with day 1 embryonic disk, then E2, E4, E6, and E14 heart and brain (n R 3
measurements each). By the time beating starts, the heart is already 3-fold stiffer than early embryonic tissue and then stiffens at a rate of 0.3 kPa/day.
Due to the thick epicardium of E6 and older hearts relative to the inner diameter of our micropipettes, measurements likely underestimate stiffness of
the myocardium at those stages due to significant contribution of epicardium. Brain tissue does not stiffen during development and remains viscoelastic
with a mean Et = 0.3 kPa.
(E) Quantitative mass spectrometry (MS) of cellular proteins extracted from intact embryonic disc (Hamburger-Hamilton stages 3–4), E2, E3, E4, and E10
heart and brain tissue reveals a small set of detected proteins with expression patterns similar to heart or brain mechanics: namely, a general increase
in heart and relatively small increase in brain. Expression is relative to average in brain E2–E3 (n R 3 MS measurements).
(F) Immunoblot confirms that MS measurements of cardiac myosin II expression increase in heart development. Samples are pooled from three to four
embryos at each reported stage and were normalized to lamin-B1 (n R 3).
(G) MS indicates that collagen I expression increases during heart development, but not greatly during brain development. Inset images: 1% SDS-decellu-
larized E4 and E14 hearts. The insoluble matrices retain the shape of the embryonic hearts, but whereas E14 matrix (with 80% of MS ion current being
collagen I) appears solid, the E4 matrix appears more reticulated and porous, consistent with relatively less mass.
(H) Of the proteins identified byMS (Table S1), a small subset had expression levels across tissues and development that paralleledmechanics. Error bars in
all figures represent SEM.
Beating Hearts Have Optimal Matrix and Myosin2435
early and late heart (Figure S1F), without perturbing myosin IIlevels (Figure S1G). The stiffness of brain tissue thus seemscellular in nature, whereas heart tissue mechanics have majorextracellular matrix contributions at even the earliest func-tional stage of beating.
Modest Softening or Stiffening Impairs Beating
Embryonic heart tubes beat spontaneously atw1 Hz for up to1–2 days after isolation, and we could easily measure local tis-sue strain in heart tubes during beating by imaging GFP-trans-fected cells as fiducial markers (Figure 2A). This very visibleactivity is used to address the main question of our studies:whether cells in an intact living tissue are sensitive to
microenvironment elasticity. Controlled dose-time treatmentswith collagenase provided a simple means of softening tissuematrix (in only 30 min), while enzymatic crosslinking of ECMwith transglutaminase provided a similarly rapid method tostiffen tissue (Figures S2A and S2B). Enzyme permeated thetissue (Figure S2C), and for all but the most extreme softeningtreatment, embryonic heart behaved elastically in micropi-pette aspiration (Figure S2B, inset). By transfecting cells witha GFP membrane protein (SIRPA-GFP), we could also seethat the contours of beating cells were unaffected by collage-nase (Figures 2B and S2D). Tissue softening is thus dueprimarily to cleavage of ECM rather than disruption of cellconnections.

A
∆s12
∆s1’2’
B
Relaxed
Contracted
C
Strain in Ventricle = 0.10 – 0.15
0
1Local Heart contraction
Stra
intre
ated
/ Stra
inun
treat
ed
E
very lowEo
0
30
0 1 2 3
Velocity through ventricle
Vel
ocity
(mm
/s)
E4 Heart stiffness: Etreated / Euntreated
E4 hearts
ContractionBefore treatment
Collagenase
1 2 3 4
2 3 3 4
30 mini ii iii
iv
v
0
1
2
0 50
Rel
ativ
e st
iffne
ss
Softening time (min)10-30
D
Non
beat
ing
+ P
artia
l-be
atin
g fra
ctio
n of
hea
rtsPartial-beating0
1
Aspiration
heart contours
*
E6 hearts
Figure 2. Effect of Extracellular Matrix Softening
and Stiffening on Heart Tube Beating
(A) E4 heart imaged by phase contrast (i) or fluo-
rescence (ii) after sparse transfection with GFP.
Scale bar represents 100 mm. Each green spot
is a cell. Three cells used to calculate strain dur-
ing beating are tracked from their relaxed (iii) to
contracted (iv) positions. Scale bar represents
20 mm. Strain is schematized in (v).
(B) SIRPA-GFP-expressing cell in transfected
E4 heart tissue before and after softening.
Overlays of SIRPA-GFP-expressing cells over
time help visualize cell shape changes during
contraction and softening treatments. Overlay
of the same cell pretreatment while relaxed
(green) and contracted (red) (bottom left)
shows less overlap (yellow) than the relaxed
cell before and after tissue softening; cells
thus maintain morphology and adhesions dur-
ing softening.
(C) Tissue strain during beating of GFP-trans-
fected softened and stiffened E4 heart normal-
ized to that of untreated and the resulting relative
strain averaged for atria, ventricles, and outflow
tract: softened and stiffened tissues suppress
contractions. Typical peak strains throughout
the untreated heart tube were 10% 6 4%. The
dashed curve is a fit to Equation 1 with exponent
n = 4 6 1 and Em = 1.6 6 0.2 kPa.
(D) E6 hearts treated progressively with collage-
nase stop beating or beat only partially. Beating
is suppressed after 50 min softening treatment,
relative to untreated or briefly (10–30 min) treated
hearts (p = 0.016). Insets: representative aspira-
tion-relaxation curves for mildly softened or considerably softened hearts. Red indicates untreated tissue; blue indicates treated tissue.
(E) Velocity of the contraction wave through the ventricle versus normalized Et. Wave speeds in untreated ventricle, atria, and outflow tract of 226 4, 46 2,
and 2.8 6 0.7 mm/s, respectively, are consistent with past work [16]. For the most extreme stiffening treatment, contraction does not propagate past the
presumptive pacemaker. The dashed line is the theoretical prediction with a single adjustable parameter, namely the ratio of the stress threshold to the
magnitude of the force dipole corresponding to a contracting cell. Eo indicates the theoretically predicted stiffness below which a contraction wave should
not propagate. Error bars for all figures represent SEM (n R 3 hearts).
Current Biology Vol 23 No 232436
After the enzymatic treatments, hearts continue to beatrhythmically (Movie S1), but themagnitude of local contraction(calculated from GFP-expressing cells) was always affected.Each heart tube region was analyzed separately (FiguresS2E–S2I), and normalization to pretreatment measurementsaccounted for slight variations (w20%) in embryo age and/orlab temperature. Untreated tissue invariably showed thelargest contraction, which was typically w10% strain, whileboth softening and stiffening of the heart suppressed contrac-tile strain (Figures 2C and S2E). Stiffening of tissue should sup-press strain because any muscle cell has a finite capacity towork against a very high load, but softening of the tissuematrixalso decreased contractile strain. Consistent with these E4results, softening of E6 hearts likewise impeded beating(Figure 2D). A mathematical theory for striation [17] providesa basis for modeling contractile functions with the equation
fwhE=ðEm +EÞ2
in; (Equation 1)
which also fits contractile strains in experiments (Figure 2C,dashed line; Box S1). The optimal stiffness for heart contrac-tion is thus the stiffness of native heart.
The speed of the contraction wave in each heart regionincreases monotonically with tissue stiffness, except for themost extreme rigidity (Figure 2E). The linearity of wave speedcan be predicted from a viscoelastic model of active media(Box S2). For the rigidified heart, beating was still evident,but the contraction wave did not propagate past the
pacemaker region in the atrium (Figure S2I). Softening treat-ments also decreased the probability of contractions propa-gating out of the atrium.
E4 and Stem Cell-Derived Cardiomyocytes Are Highly
Sensitive to Matrix Elasticity
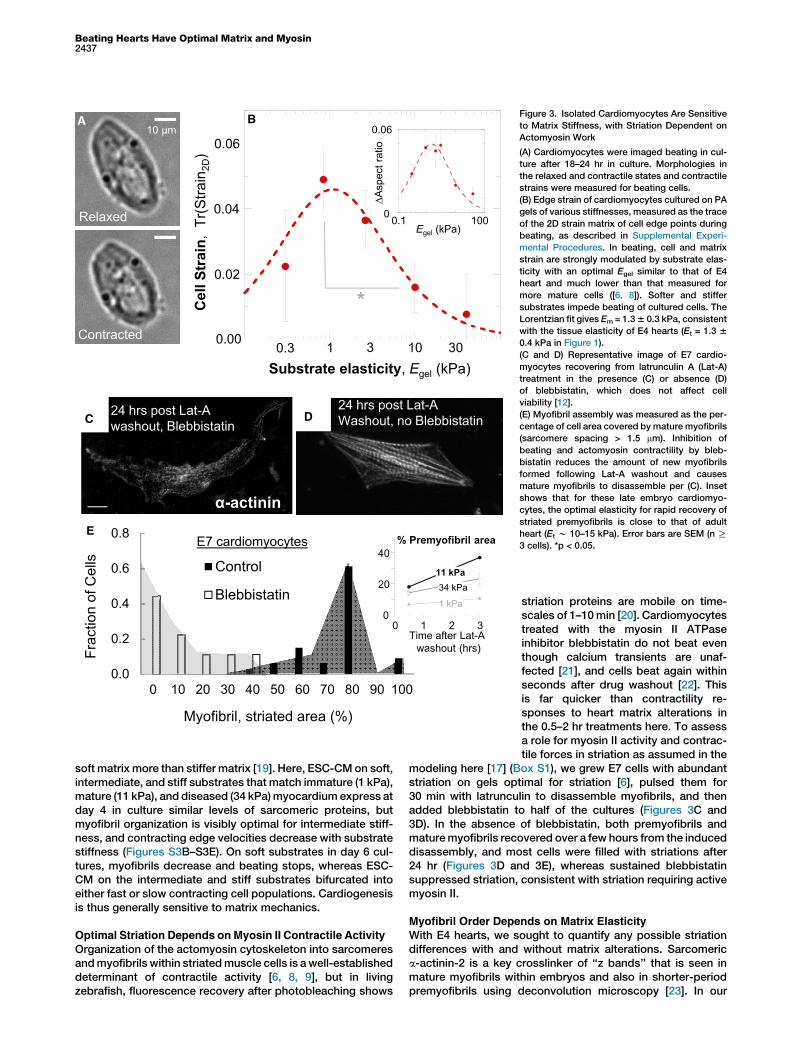
To assess whether variations in matrix elasticity affectE4-cardiomyocyte adhesion and beating, isolated cells andtheir properties were studied as sparse cultures on collagenI-coated polyacrylamide gels of varied stiffness (Figure 3A).Most of the cultured cells beat at 0.5–1.5 Hz, similar to theheart, indicating high viability as well as sustained adhesion.Relaxed morphologies were measured after 24 hr in cultureand showed that substrates stiffer than E4 heart tissue pro-mote spreading and elongation (Figure S3A), as is commonwith other mesenchymal cell types (e.g., [12]). Cells onmatrices of stiffness similar to that of the tissue of origin(w1–2 kPa) were relatively round and unspread compared tothe maximum achievable elongation and spreading. Nonethe-less, contractile deformation of an E4 cardiomyocyte and itslocal matrix proves optimal at the matrix elasticity of nativeE4 tissue (Figure 3B). In vitro contractions were measured interms of both 2D strains using cell edge displacements andchanges in aspect ratio.Cardiomyocytes derived from embryonic stem cells (ESC-
CM), induced pluripotent stem cells (iPS-CM), or directlytransdifferentiated cells hold great potential for regenerativetherapies [18], and human ESC-CM and iPS-CM displace
0.0
0.2
0.4
0.6
0.8
0 10 20 30 40 50 60 70 80 90 100
Myofibril, striated area (%)
Control
Blebbistatin
E7 cardiomyocytesE
α-actinin
24 hrs post Lat-AWashout, no Blebbistatin
24 hrs post Lat-A washout, Blebbistatin
D
0
20
40
0 1 2 3
% Premyofibril area
Time after Lat-A washout (hrs)
1 kPa
11 kPa34 kPa
Frac
tion
of C
ells
10 μm
Relaxed
Contracted
A B
Cel
l Str
ain,
Tr(S
train
2D)
0.00
0.02
0.04
0.06
0.3 1 10 30Substrate elasticity, Egel (kPa)
*
0
0.06
0.1 100
∆Asp
ect r
atio
Egel (kPa)
3
C
Figure 3. Isolated Cardiomyocytes Are Sensitive
to Matrix Stiffness, with Striation Dependent on
Actomyosin Work
(A) Cardiomyocytes were imaged beating in cul-
ture after 18–24 hr in culture. Morphologies in
the relaxed and contractile states and contractile
strains were measured for beating cells.
(B) Edge strain of cardiomyocytes cultured on PA
gels of various stiffnesses, measured as the trace
of the 2D strain matrix of cell edge points during
beating, as described in Supplemental Experi-
mental Procedures. In beating, cell and matrix
strain are strongly modulated by substrate elas-
ticity with an optimal Egel similar to that of E4
heart and much lower than that measured for
more mature cells ([6, 8]). Softer and stiffer
substrates impede beating of cultured cells. The
Lorentzian fit gives Em = 1.36 0.3 kPa, consistent
with the tissue elasticity of E4 hearts (Et = 1.3 6
0.4 kPa in Figure 1).
(C and D) Representative image of E7 cardio-
myocytes recovering from latrunculin A (Lat-A)
treatment in the presence (C) or absence (D)
of blebbistatin, which does not affect cell
viability [12].
(E) Myofibril assembly was measured as the per-
centage of cell area covered by mature myofibrils
(sarcomere spacing > 1.5 mm). Inhibition of
beating and actomyosin contractility by bleb-
bistatin reduces the amount of new myofibrils
formed following Lat-A washout and causes
mature myofibrils to disassemble per (C). Inset
shows that for these late embryo cardiomyo-
cytes, the optimal elasticity for rapid recovery of
striated premyofibrils is close to that of adult
heart (Et w 10–15 kPa). Error bars are SEM (n R
3 cells). *p < 0.05.
Beating Hearts Have Optimal Matrix and Myosin2437
soft matrix more than stiffer matrix [19]. Here, ESC-CM on soft,intermediate, and stiff substrates that match immature (1 kPa),mature (11 kPa), and diseased (34 kPa)myocardium express atday 4 in culture similar levels of sarcomeric proteins, butmyofibril organization is visibly optimal for intermediate stiff-ness, and contracting edge velocities decrease with substratestiffness (Figures S3B–S3E). On soft substrates in day 6 cul-tures, myofibrils decrease and beating stops, whereas ESC-CM on the intermediate and stiff substrates bifurcated intoeither fast or slow contracting cell populations. Cardiogenesisis thus generally sensitive to matrix mechanics.
Optimal Striation Depends onMyosin II Contractile Activity
Organization of the actomyosin cytoskeleton into sarcomeresandmyofibrils within striatedmuscle cells is a well-establisheddeterminant of contractile activity [6, 8, 9], but in livingzebrafish, fluorescence recovery after photobleaching shows
striation proteins are mobile on time-scales of 1–10min [20]. Cardiomyocytestreated with the myosin II ATPaseinhibitor blebbistatin do not beat eventhough calcium transients are unaf-fected [21], and cells beat again withinseconds after drug washout [22]. Thisis far quicker than contractility re-sponses to heart matrix alterations inthe 0.5–2 hr treatments here. To assessa role for myosin II activity and contrac-tile forces in striation as assumed in the
modeling here [17] (Box S1), we grew E7 cells with abundantstriation on gels optimal for striation [6], pulsed them for30 min with latrunculin to disassemble myofibrils, and thenadded blebbistatin to half of the cultures (Figures 3C and3D). In the absence of blebbistatin, both premyofibrils andmaturemyofibrils recovered over a few hours from the induceddisassembly, and most cells were filled with striations after24 hr (Figures 3D and 3E), whereas sustained blebbistatinsuppressed striation, consistent with striation requiring activemyosin II.
Myofibril Order Depends on Matrix Elasticity
With E4 hearts, we sought to quantify any possible striationdifferences with and without matrix alterations. Sarcomerica-actinin-2 is a key crosslinker of ‘‘z bands’’ that is seen inmature myofibrils within embryos and also in shorter-periodpremyofibrils using deconvolution microscopy [23]. In our

Sarcomere spacing
z-band breadth
A Bin vivo
α-Actinin 2 DNA F-actin
0
0.2
0.4
0 2 4
softeneduntreated
Frac
tion
z band breadth (μm)
0
0.2
0 1 2 3Fr
actio
nStriation spacing (μm)
F
1
2
0.1 1 10 100
in vivo
z ba
nd b
read
th (μ
m)
Egel or Et (kPa)
premyofibril
myofibril
**E
0
1
0.1 1 10 100
Frac
tion
Egel (kPa)
premyofibril
myofibril
α-Actinin 2DNA
F-actin
0.3 kPa
in vitro
10 kPa
3 kPa
10 μm
D
1
2
0.1 1 10 100
premyofibril
myofibril
Spa
cing
(μ m
)
Egel (kPa)
C
Figure 4. Sarcomere Breadth Changes in Soft-
ened Heart and in Isolated Cardiomyocytes on
Compliant Substrates
(A) Untreated and softened whole E4 hearts
were immunostained for sarcomeric a-actinin-2,
F-actin, and DNA and imaged by confocal micro-
scopy. Sarcomere spacing and z band breadth
(inset) were measured to assess any structural
changes.
(B) z band breadth is significantly decreased in
the 47%-softened heart relative to untreated con-
trols. The decreased registry of myofibrils sug-
gests a decreased coupling between adjacent
myofibrils during contraction. Sarcomere spacing
(lower plot) is consistent with mature myofibril
sarcomere spacing and is not significantly
different in softened and untreated hearts.
(C) E4 cardiomyocytes cultured on gels were
stained in the same manner as the whole hearts
in (A). Figure shows typical E4 cardiomyocytes
on gels with stiffnesses of 0.3, 3.0, and 10 kPa.
(D) Striation spacing was bimodal in distribution,
indicating mature myofibrils (sarcomere spac-
ing > 1.8 mm) as well as premyofibrils (sarcomere
spacing < 1.4 mm).
(E) Fraction of each type of striation per cell with
myofibrils maximal on gels where premyofibrils
are minimal. We fit the fraction of myofibrils with
fm = f (Equation 1) (blue dashed line) and premyo-
fibrils using fp = 1 2 fm with Em = 9 6 2 kPa.
(F) z band breadth for myofibrils and premyofi-
brils were maximized on substrates of intermedi-
ate stiffness. Fits to Equation 1 yield Em = 1.7 6
0.3 kPa for premyofibrils and Em = 4.2 6 0.6 kPa
for myofibrils.
Error bars are SEM (n R 3 hearts or cells).
Current Biology Vol 23 No 232438
confocal imaging, we measured sarcomere spacing lateral‘‘breadth’’ of z bands along in-plane sarcomeres as a keymetric of registry (Figure 4A, inset). Whereas striation spacingpeaked at 1.8 mm and appeared unaffected by w50% soft-ening of the E4 heart, z band breadth was reduced relative tountreated control (Figure 4B). This decreased registry of myo-fibrils shortly after softening of the matrix indicates adecreased coupling of sarcomeres and is consistent with thestriationmodel [17], highlighting amolecular-scalemechanismfor decreased contraction against decreased extracellularload.
Isolated cardiomyocytes beating on gels (Figure 4C) showstriation spacing of w1 mm for premyofibrils, which conformsto expectations [20], and also the typical w1.9 mm spacing ofmyofibrils (Figure 4D) evident in intact heart. Striation spacingshows no variation with matrix, but the abundance of myofi-brils relative to premyofibrils is maximized on matrices ofelasticity 2–10 kPa (Figure 4E). This is consistent with myofi-bril assembly from premyofibrils [20]. The z band breadth ofmyofibrils also exhibited a broad and significant (p < 0.05)maximum (at w2 mm breadth) within a similar range of matrixelasticities that promote myofibril formation (Figure 4F).The premyofibrils exhibited a somewhat narrower (w1.5 mm)z band breadth that decreased on the stiffest substrates.
Myofibril structural trends in responseto substrate stiffness in culture arethus consistent with intact E4 heartand suggest a common mechanism ofstiffness-dependent registration.
z band breadth results for both mature myofibrils andpremyofibrils (Figure 4F) were also fit by Equation 1 withrespective Em = 4.2 6 0.6 and 1.7 6 0.3 kPa (n = 0.24 6 0.1).Differences in Em suggest that myofibril organization favors astiffer matrix or higher load. However, z band breadth alsolikely underestimates registration order within a cell, as itonly includes immediately adjacent and perfectly registeredstriated fibers. Indeed, z band breadth trends for myofibrilsand premyofibrils in isolated cells are broader than in simula-tion [17] but are still consistent with trends for intact heart(Figures 4B and 4F). What emerges systematically from fittingto Equation 1 is that n increases with length scale: the smallestn is determined for z band breadth in culture, and the largestn is found for strain in the intact heart. High cell density, 3Dcell-matrix coupling, and cell-cell signaling in tissue (includingcalcium excitation waves) could all provide a basis for theenhanced sensitivity to matrix E of tissue.Protein interactions that govern molecular mobility are force
sensitive in living cardiomyocytes and vary with matrixelasticity [6]. An optimum stiffness for striation is thus under-standable: while contractile activity ‘‘massages’’ registration(Figures 3E and 4) and these forces increase with matrix stiff-ness [1], high forces on stiff matrix tend to break bonds [6].Myofibrils thereby misregister if the load is either too low or

Beating Hearts Have Optimal Matrix and Myosin2439
high, which largely explains why parallel and optimal increasesin actomyosin proteins and collagens (Figure 1) must be coor-dinated in the tissue development program. Invading andproliferating fibroblasts make and remodel the matrix thatstimulates cardiomyocyte proliferation [24] with increasedexpression of specialized contractile proteins (a-actinin-2,cardiac myosin II in Figures 1D and 1E), and so it is sensiblethat this program requires matrix engagement by integrins[24] and extends tomechanosensitive, adhesion complex pro-teins such as talin [25] that also increase (Figure 1D). More-over, since collagen synthesis and organization by fibroblastsare regulated by strain (as reviewed in [26]), heart matrix islikely to be optimized by the optimal stiffness for cardiomyo-cyte striation and contraction (Figures 2, 3, and 4). The factthat the optimum increases in development from 1 to 2 kPaat E4 toward the stiffness of adult heart (e.g., Figures 1C and3E, inset) [6, 8, 9] is also consistent with preliminary observa-tions that hearts that were stiffened and stopped beatingwould sometimes restart their beating 10 hr later. Last, excita-tion-contraction coupling (ECC) in muscle physiology is wellestablished [27], but the broad effects of matrix stiffness onindividual cells in sparse culture preclude the confoundingimpacts of cell-cell electrical communication and ultimatelysuggest that excitation-contraction-matrix coupling (ECMC)is required to truly understand muscle.
Experimental Procedures
Heart isolation (per IACUC-approved protocol 804455), enzyme treatments,
micropipette analyses, tissue strain analyses, mass spectrometry prote-
omics, cell isolation, and standard techniques are described in detail in
the Supplemental Experimental Procedures.
Supplemental Information
Supplemental Information includes three figures, two boxes, Supplemental
Analysis, Supplemental Experimental Procedures, one table, and onemovie
and can be found with this article online at http://dx.doi.org/10.1016/j.cub.
2013.10.057.
Acknowledgments
We appreciate comments from Jean and Joseph Sanger (Cell and Develop-
mental Biology, Upstate Medical University). Funding from the National
Institutes of Health (P01DK032094, R01HL062352, and NCATS-
8UL1TR000003) and the National Science Foundation (LRSM-MRSEC) is
gratefully acknowledged.
Received: April 16, 2013
Revised: September 5, 2013
Accepted: October 22, 2013
Published: November 21, 2013
References
1. Gonzalez-Rodriguez, D., Guevorkian, K., Douezan, S., and Brochard-
Wyart, F. (2012). Soft matter models of developing tissues and tumors.
Science 338, 910–917.
2. Swift, J., Ivanovska, I.L., Buxboim, A., Harada, T., Dingal, P.C., Pinter, J.,
Pajerowski, J.D., Spinler, K.R., Shin, J.W., Tewari, M., et al. (2013).
Nuclear lamin-A scales with tissue stiffness and enhances matrix-
directed differentiation. Science 341, 1240104.
3. Khetan, S., Guvendiren, M., Legant, W.R., Cohen, D.M., Chen, C.S., and
Burdick, J.A. (2013). Degradation-mediated cellular traction directs
stem cell fate in covalently crosslinked three-dimensional hydrogels.
Nat. Mater. 12, 458–465.
4. Chien, K.R., Domian, I.J., and Parker, K.K. (2008). Cardiogenesis and the
complex biology of regenerative cardiovascular medicine. Science 322,
1494–1497.
5. Butcher, J.T., McQuinn, T.C., Sedmera, D., Turner, D., and Markwald,
R.R. (2007). Transitions in early embryonic atrioventricular valvular func-
tion correspondwith changes in cushion biomechanics that are predict-
able by tissue composition. Circ. Res. 100, 1503–1511.
6. Engler, A.J., Carag-Krieger, C., Johnson, C.P., Raab, M., Tang, H.Y.,
Speicher, D.W., Sanger, J.W., Sanger, J.M., and Discher, D.E. (2008).
Embryonic cardiomyocytes beat best on amatrix with heart-like elastic-
ity: scar-like rigidity inhibits beating. J. Cell Sci. 121, 3794–3802.
7. Hersch, N., Wolters, B., Dreissen, G., Springer, R., Kirchgeßner, N.,
Merkel, R., andHoffmann, B. (2013). The constant beat: cardiomyocytes
adapt their forces by equal contraction upon environmental stiffening.
Biol. Open 2, 351–361.
8. Jacot, J.G., McCulloch, A.D., and Omens, J.H. (2008). Substrate
stiffness affects the functional maturation of neonatal rat ventricular
myocytes. Biophys. J. 95, 3479–3487.
9. Rodriguez, A.G., Han, S.J., Regnier, M., and Sniadecki, N.J. (2011).
Substrate stiffness increases twitch power of neonatal cardiomyocytes
in correlation with changes in myofibril structure and intracellular cal-
cium. Biophys. J. 101, 2455–2464.
10. McCain, M.L., Lee, H., Aratyn-Schaus, Y., Kleber, A.G., and Parker, K.K.
(2012). Cooperative coupling of cell-matrix and cell-cell adhesions in
cardiac muscle. Proc. Natl. Acad. Sci. USA 109, 9881–9886.
11. Ulbricht, A., Eppler, F.J., Tapia, V.E., van der Ven, P.F., Hampe, N.,
Hersch, N., Vakeel, P., Stadel, D., Haas, A., Saftig, P., et al. (2013).
Cellular mechanotransduction relies on tension-induced and chap-
erone-assisted autophagy. Curr. Biol. 23, 430–435.
12. Engler, A.J., Sen, S., Sweeney, H.L., and Discher, D.E. (2006). Matrix
elasticity directs stem cell lineage specification. Cell 126, 677–689.
13. Theret, D.P., Levesque, M.J., Sato, M., Nerem, R.M., and Wheeler, L.T.
(1988). The application of a homogeneous half-space model in the
analysis of endothelial cell micropipette measurements. J. Biomech.
Eng. 110, 190–199.
14. Georges, P.C., Miller, W.J., Meaney, D.F., Sawyer, E.S., and Janmey,
P.A. (2006). Matrices with compliance comparable to that of brain tissue
select neuronal over glial growth in mixed cortical cultures. Biophys. J.
90, 3012–3018.
15. Zamir, E.A., Srinivasan, V., Perucchio, R., and Taber, L.A. (2003).
Mechanical asymmetry in the embryonic chick heart during looping.
Ann. Biomed. Eng. 31, 1327–1336.
16. de Jong, F., Opthof, T., Wilde, A., Janse, M., Charles, R., Lamers, W.,
and Moorman, A. (1992). Persisting zones of slow impulse conduction
in developing chicken hearts. Circ. Res. 71, 240–250.
17. Friedrich, B.M., Buxboim, A., Discher, D.E., and Safran, S.A. (2011).
Striated acto-myosin fibers can reorganize and register in response to
elastic interactions with the matrix. Biophys. J. 100, 2706–2715.
18. Laflamme, M.A., and Murry, C.E. (2005). Regenerating the heart. Nat.
Biotechnol. 23, 845–856.
19. Hazeltine, L., Simmons, C., Salick, M., Lian, X., Badur, M., Han, W.,
Delgado, S., Wakatsuki, T., Crone, W., Pruitt, B., and Palecek, S.
(2012). Effects of substrate mechanics on contractility of cardiomyo-
cytes generated from human pluripotent stem cells. Int. J. Cell Cytol.
2012, 508294.
20. Sanger, J.W., Wang, J., Holloway, B., Du, A., and Sanger, J.M. (2009).
Myofibrillogenesis in skeletal muscle cells in zebrafish. Cell Motil.
Cytoskeleton 66, 556–566.
21. Skwarek-Maruszewska, A., Hotulainen, P., Mattila, P.K., and
Lappalainen, P. (2009). Contractility-dependent actin dynamics in
cardiomyocyte sarcomeres. J. Cell Sci. 122, 2119–2126.
22. Farman, G.P., Tachampa, K., Mateja, R., Cazorla, O., Lacampagne, A.,
and de Tombe, P.P. (2008). Blebbistatin: use as inhibitor of muscle
contraction. Pflugers Arch. 455, 995–1005.
23. Du, A., Sanger, J.M., and Sanger, J.W. (2008). Cardiac myofibrillogene-
sis inside intact embryonic hearts. Dev. Biol. 318, 236–246.
24. Ieda, M., Tsuchihashi, T., Ivey, K.N., Ross, R.S., Hong, T.T., Shaw, R.M.,
and Srivastava, D. (2009). Cardiac fibroblasts regulate myocardial
proliferation through beta1 integrin signaling. Dev. Cell 16, 233–244.
25. del Rio, A., Perez-Jimenez, R., Liu, R., Roca-Cusachs, P., Fernandez,
J.M., and Sheetz, M.P. (2009). Stretching single talin rod molecules ac-
tivates vinculin binding. Science 323, 638–641.
26. Baudino, T.A., Carver,W., Giles,W., andBorg, T.K. (2006). Cardiac fibro-
blasts: friend or foe? Am. J. Physiol. Heart Circ. Physiol. 291, H1015–
H1026.
27. Bers,D. (2001). Excitation-ContractionCoupling andCardiacContraction
Force, Second Edition (Boston: Kluwer Academic Publishers).
Related Documents











