Brita Ngum Che PhD Thesis 1 Health promoting factors from milk of cows fed green plant material - The role of phytanic acid Brita Ngum Che MSc in Molecular Biology Department of Food Science Faculty of Science and Technology Aarhus University Denmark A thesis submitted for the degree of Doctor of Philosophy at Aarhus University September 2012

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

B r i t a N g u m C h e P h D T h e s i s
1
Health promoting factors from milk of cows fed green
plant material - The role of phytanic acid
Brita Ngum Che
MSc in Molecular Biology
Department of Food Science
Faculty of Science and Technology
Aarhus University
Denmark
A thesis submitted for the degree of Doctor of Philosophy at Aarhus University
September 2012

B r i t a N g u m C h e P h D T h e s i s
2
Title of PhD project: Health promoting factors from milk of cows fed green plant
material - The role of phytanic acid
PhD student: Brita Ngum Che, MSc in Molecular Biology, Department of Food Science,
Faculty of Science and Technology, Aarhus University, Denmark
Supervisor: Associate professor Jette F. Young, Department of Food Science, Faculty of
Science and Technology, Aarhus University, Denmark
Co-supervisor: Assistant professor Mette K. Larsen, Department of Food Science,
Faculty of Science and Technology, Aarhus University, Denmark
Opponents:
1. Professor Karsten Kristiansen, Department of Biology, University of Copenhagen
2. Professor Ian Givens, Animal Science Research Group, University of Reding, United
Kingdom

B r i t a N g u m C h e P h D T h e s i s
3
Preface
The work presented in the present PhD thesis titled “Health promoting factors from milk
of cows fed green plant material-the role of phytanic acid” centers around elucidating the
availability of phytanic acid in milk-fat and the relevance of this fatty acid on glucose and
fatty acid metabolism in humans. This work aimed at improving human health through the
differentiation of milk production enriched with phytanic acid.
This thesis is part of a “Green feed” project, where the metabolic impact of altered fatty
acid composition of milk-fat as affected by green feed was studied. I was specifically
interested in in vitro studies of glucose and fatty acid metabolism and the metabolic
impact of phytanic acid. The in vitro studies were conducted with a synthetic form of
phytanic acid, which is an isomeric mixture.
At the final stages of the project, the content of phytanic acid in milk-fat was accessed and
its diastereomers were observed to be differentially distribution under altered feeding
conditions. This was the reason behind the inclusion of the inclusion of studies on phytanic
acid diastereomers which was not part of the original setup of the project.
Regarding the setup of the thesis, an introduction to the thesis is presented, followed by a
literature review of factors that are relevant in understanding the work described in the
thesis. Results obtained from the study are discussed together, and some concluding
remarks are stated. Some future perspectives are included, and finally, an appendix of
manuscripts, which constitute work carried out in the project, is included.
This PhD project was based at the Department of Food Science at Aarhus University. Milk
analysis, in vitro cultures, and glucose metabolism studies were conducted at The
Department of Food Science, while the analyses of acylcarnitines were carried out at The

B r i t a N g u m C h e P h D T h e s i s
4
Department of Forensic Science, in collaboration with The Research unit of Molecular
Medicine at Aarhus University.
The work presented in this PhD project was funded by The Danish Council for Strategic
Research and The Danish Cattle Federation, and was part of the scientific network called
“Tailored milk and human health”.

B r i t a N g u m C h e P h D T h e s i s
5
Acknowledgements
I will commerce by expressing tremendous gratitude to my supervisor Jette F. Young,
Department of Food Science, for excellent supervision and guidance all through my PhD
studies. Thanks for the extreme understanding and support from you, especially during the
tough times. I am grateful for all the support right to the very last minute of completing my
thesis. To Lars I. Hellgren of The Technical University of Denmark, I am very grateful for
the fact that you conceptualized this project, thus making it at all possible. Your inputs at
every level have been indispensable for the realization of this thesis. Special thanks go to
my co-supervisor Mette K. Larsen, Department of Food Science, for her support both in
the laboratory and during the writing of manuscripts. I am grateful for the enthusiastic
discussion we have had concerning the milk-fat studies.
Special thanks to Niels Oksbjerg, Department of Food Science, for his contribution in the
isolation and culturing of porcine myotubes and the many discussions on glucose uptake
experiments and statistics. I will like to thank Jacob Holm Nielsen of Arla Foods, Braband,
Århus, for his fruitful input especially at the early stages of the project.
Special thanks to Mogens Johannsen and Rune Isaac Dupont Birkler at The Department of
Forensic Science at Aarhus University, for your help and support in the analysis of
acylcarnitines, and for you contribution in writing of manuscript. My gratitude also goes to
Niels Gregersen at The Research unit of Molecular Medicine at Aarhus University, thanks
for your inputs concerning the conception of acylcarnitine studies, but also for the warm
atmosphere and time spent at your laboratory.
Bente Andersen, Camilla Bjerg Kristensen, Caroline Nebel and Anne-Grethe Pedersen are
heartily thanked for contributing with excellent technical support.

B r i t a N g u m C h e P h D T h e s i s
6
Aase Sørensen and Anne Hjorth Balling, special thank you for your patience and excellent
help in proof-reading my thesis and manuscripts
My office mates Sumangala Bhattacharya and Bjørn Nielsen, thanks for the many
interesting discussions, and a good working atmosphere. My colleagues in Foulum, thank
you for creating an interesting working environment and for your support in one way or
the other.
To my friend Ngonidzashe Chirinda, I very much appreciate the interesting discussions all
through the years in Foulum, and your suggestions regarding the structure of my thesis. To
my friend Steen Pedersen, thank you so much for excellent support in preparing some of
the figures for my thesis.
To my friend Bülent Kocaman, thank you for giving me the courage and support eight
years ago to quit my cleaning job and continue on my academic career. I am grateful to my
friend Marceline Pirkaniemi for her moral support. To Ina Lindgård and family, thank you
so much for being such a caring and supporting family. Your unlimited help with my kids
Nicole and Victoria has been invaluable and I will always be grateful for that.
Special thanks to my family for their moral support and up-backing, and to you Dad
thanks for implanting the will and zeal in me; you knew I would make it, and I am almost
there…..
Last but not the least, I am grateful for the tolerance, understand and love of my two
daughters Nicole and Victoria. It is incredibly cool of you two. You do not stop to amaze
me; first, you came into my life, then you gave me the chance to accomplish what I have
always dreamt of. For that, and all the unsaid, this work is dedicated to you Nicole and
Victoria.

B r i t a N g u m C h e P h D T h e s i s
7
Abstract
Phytanic acid (PA) is a fatty acid (FA) that is present in ruminant products, and a natural
agonist of the peroxisome proliferator-activated receptors (PPAR). Because the PPARs are
pivotal in the regulation of glucose and FA homeostasis, it has been suggested that PA
could positively regulate these processes, in which case, it could be considered as a
bioactive compound with health-improving properties that can be used to fight against the
metabolic syndrome (MS). The objectives of this PhD study were to determine the impact
of total PA on glucose uptake and FA β-oxidation in skeletal muscles, and to elucidate the
total content of PA and the distribution of its diastereomers in milk as affected by feed
composition.
In this project, we established primary porcine myotubes as an efficient skeletal muscle
model for metabolic studies. Satellite cells (SC) derived from porcine muscles were
cultured to generate differentiated primary porcine myotubes. Viability studies were
performed to determine which concentrations or length of treatments could be tolerated
by the myotubes under glucose uptake, glycogen synthesis, and FA oxidation (FAO)
experiments. Optimization of glucose uptake assay using cytochalasin B revealed that both
the insulin-mediated and non-insulin mediated mechanisms of glucose uptake were
functioning in the myotubes. Exposures to myotubes of excess glucose during the analysis
of glucose uptake, and palmitate during the analysis of acylcarnitine, rendered the
myotubes insulin resistant and inhibited the oxidation of palmitoylcarnitine (C16),
phenomena of which can be expressed by skeletal muscles.
During the elucidation of the metabolic impact of PA, glucose uptake, glycogen synthesis
and FAO were analyzed by the use of tritiated 2-deoxyglucose, 14C- glucose and 13C-
palmitate, respectively. It was shown that physiological amounts of PA, which included 10

B r i t a N g u m C h e P h D T h e s i s
8
µM, enhanced glucose uptake, especially at low concentrations of insulin, but PA could not
activate the incorporation of glucose into glycogen, except in the presence of insulin. Thus,
PA seemed to regulate glucose uptake in both an insulin-dependent and insulin-
independent fashion. When the myotubes were rendered insulin resistant by exposure to
excess glucose, it was neither possible for PA nor insulin to stimulate glucose uptake or
glycogen synthesis. During the analysis of β-oxidation using acylcarnitine profiling, we
could show that PA enhanced the β-oxidation flux in the myotubes, as it caused an increase
in the content of acetylcarnitine (C2) and a decrease in the C16/C2 ratio. Regarding
stimulation of β-oxidation also 10 µM PA was an effective dose. However, we could not
conclude if the induction in FAO by PA was through the induction of PPARα, since a
PPARα agonist was necessary as a control to validate the changes observed.
It has previously been shown that cow breed and feed composition affect milk-output and
FA composition, respectively, and that green feed increases the content of PA in milk-fat.
In this project, milk was sampled from grazing Danish Holstein and Danish Jersey cows
May and September periods, and the total content of PA and its diastereomers (RRR PA
and SRR PA) in the milk-fat of the cows were studied by gas chromatography-mass-
spectrometry analysis. The milk yield was higher in the Danish Holstein than the Danish
Jersey cows, but the breed did not affect the total content of PA in milk-fat. The total
content of PA was higher during the grazing period of September than in May. However,
differences were small and the intake of green feed could not be related positively to the
total content of PA. The distribution of the diastereomers was affected by feeding, as the
content of the RRR PA was positively related to the intake of grazed legumes. This finding
indicates that it is possible to manipulate the PA isomer distribution through strategic
feeding.

B r i t a N g u m C h e P h D T h e s i s
9
In conclusion, a concentration of 10 µM PA achievable through dietary intake, is an
acceptable amount in in vitro studies, and can stimulate both glucose uptake and FAO.
The mechanisms behind these inductions need subsequent elucidations.

B r i t a N g u m C h e P h D T h e s i s
10
Dansk Sammendrag
Fytansyre (PA) er en fedtsyre (FA) der findes i produkter fra drøvtyggere og er en naturlig
agonist for peroxisomere proliferator-aktiverede receptorer (PPAR). Fordi PPAR’erne er
centrale for reguleringen af glukose og FA homeostase, er det blevet foreslået, at PA kan
regulere disse processer positivt, og i så fald betragtes som en bioaktiv komponent med
sundhedsfremmende egenskaber, der vil kunne bruges i forbindelse med forebyggelse af
metabolisk syndrom (MS). Formålene med dette PhD-projekt har været at fastslå
betydningen af det totale PA på glukoseoptaget og FA β-oxidation i skeletmuskulatur, og
undersøge det totale indhold af PA og distributionen af dets diastereomerer i mælk
afhængig af fodersammensætning.
I dette projekt har vi etableret primære svine myotubes som en effektiv model for
skeletmuskulatur til metaboliske studier. Satellitceller (SC) fra muskelceller fra grise er
blevet dyrket for at etablere differentierede primære svine myotubes. Levedygtigheden af
myotubes blev undersøgt, for at bestemme hvilke koncentrationer og hvilke varigheder af
behandlinger som kunne tolereres af myotubes i forbindelse med glukoseoptag,
glykogensyntese og FA oxidations (FAO) eksperimenter. Optimering af glukoseoptag ved
brug af cytochalasin B viste, at både det insulinmedierede og ikke-insulinmedierede
glukoseoptag fungerede i myotubes. Overeksponering af glukose til Myotubes i forbindelse
med analysen af glukoseoptag, og palmitat i forbindelse med analyser af acylcarnitin,
gjorde myotubes insulinresistente og hæmmede oxidationen af palmitoylcarnitin (C16),
fænomener som er velkendte i skeletmuskulatur.
I forbindelse med undersøgelsen af PA’s metaboliske betydning, blev glukoseoptag,
glycogensyntese og FAO analyseret ved hjælp af henholdsvis tritieret 2-deoxyglukose, 14C-
glukose og 13C-palmitat. Det blev vist, at den fysiologiske mængde af PA, inklusiv 10 µM,

B r i t a N g u m C h e P h D T h e s i s
11
forbedrede glukoseoptaget, især ved lave koncentrationer af insulin, men PA kunne ikke
aktivere dannelsen af glykogen fra glukose, medmindre der var insulin til stede. Dette
indikerer at PA regulerer glukoseoptag via både en insulin-afhængig og insulin-uafhængig
mekanisme. Når myotubes blev gjort insulinresistente ved overeksponering af glukose, var
det hverken muligt for PA eller insulin at stimulere glukoseoptag eller glykogensyntese. I
forbindelse med analysen af β-oxidation ved acylcarnitine profilering, kunne vi se, at PA
forbedrede β-oxidationen i myotubes, fordi det øgede indholdet af acetylcarnitin (C2) og
mindskede C16/C2 ratioen. Også i stimuleringen af β-oxidationen viste 10 µM PA sig at
være en effektiv dosis. Vi kunne dog ikke konkludere om PA-induceret FAO sket via
PPARα, fordi en veletableret PPARα agonisten ville være nødvendig som kontrol for at
kunne validere de ændringer, der blev observeret.
Det er tidligere vist, at kvægrace og fodersammensætning har betydning for såvel
mælkeudbytte som mælkens FA sammensætning, og at grønne fodermidler øger indholdet
af PA i mælkefedtet. I dette projekt blev der taget mælkeprøver fra græssende Holstein og
Jersey køer i maj og september, og det totale indhold af PA og dennes diastereomere (RRR
PA og SRR PA) i mælkefedtet fra køerne blev analyseret ved brug af gaschromatografi og
massespektrometri. Mælkeudbyttet var højere for Holstein end for Jersey køer, mens
racen ikke havde nogen betydning for det totale indhold af PA. Det totale indhold af PA var
højere i mælk fra september sammenlignet med maj. Forskellene var dog små, og det var
ikke muligt at relatere indtaget af grønne fodermidler positivt til mælkens PA indhold.
Fordelingen af diastereomererne var afhængig af fodringen, hvor indholdet af RRR PA var
positivt relateret til indtaget af bælgplanter i afgræsningen. Dette resultat indikerer, at det
er muligt at styrefordelingen af PA diastereomerer gennem strategisk fodring.

B r i t a N g u m C h e P h D T h e s i s
12
Det kan konkluderes, at koncentrationen på 10 µM PA, der kan opnås via kosten, er en
acceptabel mængde i in vitro studier, og at dette kan stimulere både glukoseoptag og FAO.
Mekanismerne bag disse aktiveringer kræver dog nærmere undersøgelser.

B r i t a N g u m C h e P h D T h e s i s
13
Key words
Primary porcine myotubes, phytanic acid, phytanic acid diastereomers, insulin, palmitic
acid, glucose uptake, fatty acid β-oxidation, acylcarnitine, pasture, legumes

B r i t a N g u m C h e P h D T h e s i s
14
Lists of disseminations:
A: Abstracts and posters
1. Separate and combined effects of phytanic acid and insulin on glucose uptake in
primary porcine myotubes; 2009.Poster presented at International Food Research Day,
Aarhus University, Aarhus, Denmark
2. Production of phytanic acid-rich milk by feeding cows green-plant material; 2009.
Poster presented at LMC PhD congress, Copenhagen, Denmark
3. Giver grønt foder sundere mælk? Forbedres de ernæringsmæssige egenskaber af
mælkefedt, når køerne fodres med grønt plantemateriale? Mælkeritidende, Nr.
16,2009, s. 344-345
4. Phytanic acid enhances glucose uptake in primary porcine myotubes; 2010. Abstract
for The 9th Conference of the International Society for the Study of Fatty Acids and
Lipids, Maastricht, Holland
5. Phytanic acid-induced glucose uptake in primary porcine myotubes may be mediated
by glucose transporter 4; 2011. Abstract presented at LMC Food Congress, Odense,
Denmark
6. The distribution of phytanic acid diastereomers in food; 2011. Poster presented at 3rd
COST Feed for Health Conference, Copenhagen Denmark

B r i t a N g u m C h e P h D T h e s i s
15
B: Manuscripts
1. Manuscript I; MI:
Phytanic acid stimulates glucose uptake in a model of skeletal muscles, the primary
porcine myotubes
2. Manuscript II; MII:
Effects of phytanic acid on beta-oxidation flux in a primary porcine myotube model as
detected by an UPLC-MS/MS acylcarnitine assay
3. Manuscript III; MIII:
Content and distribution of phytanic acid diastereomers in organic milk as affected by
feed composition

B r i t a N g u m C h e P h D T h e s i s
16
List of abbreviations
ACAD acyl-CoA dehydrogenase
Acyl-CoA acyl coenzyme A
AMCAR α-methylacyl-CoA racemase
AMPK adenosine monophosphate-activated protein kinase
AS160 Akt substrate 160
BMI body mass index
C16 palmitoylcarnitine
C2 acetylcarnitine
CACT carnitine-acetylcarnitine translocase
CAT carnitine acyltransferase
CLA conjugated linoleic acid
COT carnitine O-octanoyltransferase
CPT carnitine palmitoyl transferase
DBD DNA binding domain
EH enoyl-CoA hydratase
FA fatty acid
FAO fatty acid oxidation
GLUT glucose transporters
GS glycogen synthase
GSK glycogen synthase kinase
HAD 3-hydroxyacyl-CoA dehydrogenase
IGF insulin growth factor
IRAP insulin-responsive aminopeptidase
IRS insulin receptor substrate
KAT 3-ketoacyl-CoA thiolase

B r i t a N g u m C h e P h D T h e s i s
17
LBD ligand binding domain
LCEH long-chain enoyl-CoA hydratase
LCHAD long-chain hydroxyacyl-CoA dehydrogenase
LCKAT long-chain ketoacyl-CoA thiolase
MCAD medium-chain acyl-CoA dehydrogenase
MCKAT medium-chain ketoacyl-CoA thiolase
MS metabolic syndrome
mTOR rapamycin complexed with rictor
MTP mitochondrial trifunctional protein
MyoD myogenic differentiation
OCTN2 organic cation transporter number 2
PA phytanic acid
PAX paired box
PDK1 phosphoinositide dependent protein kinase-1
PI3K phosphatidyl inositol 3-kinase
PIP phosphatidyl inositol 3, 4, 5- triphosphate
PKB protein kinase B
PKC protein kinase C
PPAR peroxisome proliferator-activated receptor
PPRE peroxisome proliferator response element
RXR retinoid x receptor
SC satellite cells
SCAD short-chain acyl-CoA dehydrogenase
SCEH short-chain enoyl-CoA hydratase
SCHAD short-chain hydroxyacyl-CoA dehydrogenase
T2D type 2 diabetes

B r i t a N g u m C h e P h D T h e s i s
18
TAG triacylglyceride
UDP uridine diphosphate
VI vastus intermedius
VLCAD very long-chain acyl-CoA dehydrogenase

B r i t a N g u m C h e P h D T h e s i s
19
Table of contents
Title page………………………………………………………………………………………………………. 1
Authors addresses, supervisors, reviewers, opponents………………………………………. 2
Preface………………………………………………………………………………………………………….. 3
Acknowledgements………………………………………………………………………………………… 5
Abstract………………………………………………………………………………………………………… 7
Dansk Sammendrag……………………………………………………………………………………….. 10
Key words…………………………………………………………………………………………………….. 13
Lists of disseminations…………………………………………………………………………………… 14
Abbreviations………………………………………………………………………………………………… 16
Table of content……………………………………………………………………………………………… 19
1. INTRODUCTION………………………………………………………………………………………. 21
1.1 Problem description……………………………………………………………………….. 21
1.2 Hypotheses……………………………………………………………………………………. 23
1.3 Aims……………………………………………………………………………………………… 24
1.4 The metabolic syndrome………………………………………………………………… 25
1.4.1 Insulin resistance in skeletal muscles………………………………………………. 26
1.4.2 Obesity………………………………………………………………………………………….. 29
1.4.3 Type 2 diabetes (T2D)……………………………………………………………………… 30
1.5 The Satellite cell (SC)………………………………………………………………………. 31
1.5.1 From SCs to skeletal muscles…………………………………………………………… 33
1.6 Insulin signaling in skeletal muscles………………………………………………… 34
1.6.1 Glucose uptake……………………………………………………………………………… 36
1.6.2 Glycogen synthesis………………………………………………………………………… 37
1.7 The β- oxidation……………………………………………………………………………. 38

B r i t a N g u m C h e P h D T h e s i s
20
1.7.1 Acylcarnitine shuttling ……………………………………………………….………… 39
1.7.2 Β- oxidation enzymes……………………………………………………………………. 41
1.8 Peroxisome proliferator activated-receptors (PPAR)………………………… 43
1.8.1 PPARα…………………………………………………………………………………………. 44
1.8.2 PPARβ………………………………………………………………………………………….. 45
1.8.3 PPARγ………………………………………………………………………………………….. 45
1.9 Phytanic acid (PA)…………………………………………………………………………. 46
1.9.1 Origin of PA and its diastereomers………………………………………………….. 46
1.9.2 Metabolism of PA in humans………………………………………………………….. 48
1.9.3 Milk fat composition of PA as affected by cow feed…………………………… 51
2 RESULTS AND DISCUSSION……………………………………………………………………. 52
2.1 Primary porcine myotubes; a model of skeletal muscles, for studying glucose
and FA metabolism………………………………………………………………………… 52
2.2 The metabolic impact of PA……………………………………………………………. 55
2.3 Concerns regarding excess content of total PA in humans…………………. 58
2.4 PA in milk-fat as affected by cow feed……………………………………………… 58
2.5 Concerns regarding the distribution of PA diastereomers in humans…. 61
3 CONCLUDING REMARKS………………………………………………………………………... 63
4 PERSPECTIVES………………………………………………………………………………………… 64
5 REFERENCES…………………………………………………………………………………………… 66
6 APPENDIX………………………………………………………………………………………………. 83
6.1 Manuscript I; MI…………………………………………………………………………… 83
6.2 Manuscript II; MII………………………………………………………………………… 117
6.3 Manuscript III; MIII……………………………………………………………………… 152

B r i t a N g u m C h e P h D T h e s i s
21
1 INTRODUCTION
1.1 Problem description
Small changes in food components that can improve pathological conditions and leave the
population healthier are in focus. Milk is a global commodity and harbors numerous
bioactive compounds that have been suggested to improve diseases such as type 2 diabetes
(T2D). Besides proteins, minerals and vitamins, many fatty acids (FA) are constituents of
milk and share nutritional, functional and physiological importance in humans.
Accumulating data indicates that the intake of milk alleviates symptoms of the metabolic
syndrome (MS); an observation that is contrary to the fact that milk fat is highly saturated
and, thus, believed to be unhealthy. A milk-fat paradox has originated from these findings
and prompts the elucidation of potential bioactives responsible for the protective effect of
milk.
Phytanic acid (PA); 3, 7, 11, 15 tetramethylhexadecanoic acid, is a milk-fat- derived FA,
which is a natural ligand of the peroxisome proliferator-activated receptors (PPAR) and
the retinoid X receptor (RXR). The PPARs are known for their crucial role in the regulation
of glucose homeostasis and FA metabolism. Being natural agonists of these receptors, PA
has been suggested to be a bioactive compound with potential health-improving abilities
and might to some extent alleviate MS and T2D. Reviews on the metabolism and
physiological potentials of PA have thrown more light and focus on PA as an important
bioactive compound just like conjugated linoleic acid (CLA).
So far, one study in rat hepatocytes has shown that PA can improve glucose uptake, even
though very high concentrations of the FAs were utilized. Also, not much has been done
regarding the regulation of FA metabolism by PA. Thus, very little is known of the effect of
PA on glucose homeostasis and FA oxidation (FAO). Furthermore, works in this thesis

B r i t a N g u m C h e P h D T h e s i s
22
show that the content of the two naturally occurring stereomers of PA (RRR PA and SRR
PA) in milk fat is differentially affected by the feed composition of the cow. Not much is
known about the physiological activities of the diastereomers of PA. More so, in light of the
fact that enzymes involved in the metabolism of FA are stereo-specific, the distribution of
the PA diastereomers in milk fat becomes relevant.
T2D is a global epidemic that is fast-growing. People with diabetes live 12 years less than
the rest of the population, and the mortality rate in Denmark is 3-6 times higher amongst
people with diabetes ≤ 65 years than the rest of the population. Early and coordinated
treatment can reduce diabetic-related mortality by 50 %. To reduce the growth of the
diabetic epidemic and the use of medication, which is extremely expensive, the search for
solutions in food products with better health quality is of interest.

B r i t a N g u m C h e P h D T h e s i s
23
1.2 Hypotheses
Dairy fat is rich in PA, which exists naturally as SRR PA and RRR PA. PA is a natural
agonist of the peroxisome proliferator-activated receptors (PPAR). The PPARs are
regulators of glucose homeostasis and lipid metabolism, and have been documented to
improve insulin sensitivity and FAO. There is, thus, an incitement that PA can improve
these processes. Measurements in glucose uptake and acyl carnitines have been used to
follow changes in glucose homeostasis and FA metabolism, as they are indispensable
metabolites required for these processes. Skeletal muscles account for the bulk of insulin-
mediated glucose uptake and therefore chosen as the model for exploring the impact of PA
on glucose and FA metabolism. As PA diastereomers are derived from chlorophyll, their
content in milk fat is dependent on the feed composition, and increased amounts of green
feed are expected to affect the content of PA diastereomers. From these bases, four
hypotheses arose:
1. Primary porcine myotubes can be an effective model of skeletal muscles for studying
glucose and FA metabolism.
2. Glucose uptake in skeletal muscles can be improved by PA.
3. The exposure of skeletal muscles to PA will induce β-oxidation.
4. The total content of PA and its diastereomers is differentially affected by specific
feeding patterns.

B r i t a N g u m C h e P h D T h e s i s
24
1.3 Aims
The aims of this PhD project were to study the impact of total PA on glucose uptake and FA
β-oxidation in primary porcine myotubes, and to determine the total content of PA and the
distribution of its diastereomers in milk as affected by feed composition. The overall aim
was to produce differentiated milk of improved bioactive capacity.

B r i t a N g u m C h e P h D T h e s i s
25
1.4 The metabolic syndrome(MS)
The metabolic syndrome (MS) is a disease state consisting of abdominal obesity,
dyslipidemia, hypertension, pro-inflammatory and pro-thrombotic states, insulin
resistance and glucose intolerance (Table 1). These factors usually occur in clusters and
promote the prevalence of T2D and cardiovascular diseases directly [1]. Due to the inter-
mix of the various components constituting MS, it is very difficult to pin-point and treat.
Populations with MS are insulin resistant and at higher risk of developing cardiovascular
diseases than people without MS [2]. The prevalence of MS is about 20-30 % in normal
subjects and 70-80 % in diabetic subjects [2, 3]. In Denmark, the frequency of MS is 38 %
and 22 % amongst men and women ≥ 60 years of age, respectively [4]. Low physical
activity, excess dietary intake, maternal obesity and ethnicity are some of the main factors
that promote the prevalence of MS [5-10].
Table 1: Risk factors related to the metabolic syndrome (Adopted from [11])
Waist circumference* Men: ≥ 94 cm
Women: ≥ 80 cm
Triglyceride (TAG) levels >1.7 mM (150 mg/dL)
HDL-cholesterol Men: < 1.03 mM (40 mg/dL)
Women: < 1.29 mM (50 mg/dL)
Blood pressure systolic: ≥ 130 mm Hg
Diastolic: ≥ 85 mm Hg
Fasting plasma glucose ≥ 5.6 mM (100 mg/dL)
*The waist circumference is ethnic-specific and the values stated are for europids. It is also valid
for sub-Saharan Africans and eastern Mediterranean and Middle East populations until useful data
are available.

B r i t a N g u m C h e P h D T h e s i s
26
1.4.1 Insulin resistance in skeletal muscle
Insulin resistance (IR) in skeletal muscle is one of the early signs and a core phenomenon
in the pathophysiology of T2D [12, 13]. It occurs when normal concentrations of insulin are
not enough to execute effective uptake and disposal of glucose. The increase in blood
insulin levels as well as glucose levels, thus, characterize IR, and in muscles glucose uptake
is impaired with the down regulation of the glucose receptor transporters (GLUT) [14, 15].
Figure 1 illustrates the dysfunction in GLUT-4, which results in diminished glucose uptake
and IR. Since IR is not completely abolished by GLUT-1 and GLUT-4, it denotes that
players other than the glucose transporters are involved in the generation of IR [16, 17].
Decrease in the protein content of insulin receptor substrate-I (IRS-I) and impairment of
its phosphorylation, reduced activity of phosphatidyl inositol 3-kinase (PI3K) and
downstream enzymes are partly responsible for the defects of the insulin-signaling
pathway [18]. IR also results from a defect in glycogen synthesis, due to a reduced activity
of glycogen synthase (GS) originating from a drop in insulin-stimulated GS [19]. Defects in
the adenosine monophosphate-activated protein kinase (AMPK) pathway, has also been
shown to cause IR, in a manner that does not involve insulin [20].
Excess food intake overloads the muscles with glucose and lipids, which are known to
generate IR. High glucose levels have been shown to generate insulin resistance by
activation of the glucosamine pathway [21, 22], while accumulation of fats in the muscles
has also been linked to IR [23]. Lipid accumulation is thought to cause IR by inhibiting
mitochondrial function [24], and possibly the rate of FAO. Shulman et al proposed that
the mechanism by which FAs causes IR in the muscle lies in the presence of their
metabolites, generated through excess feeding or through defects in FAO, which cause the

B r i t a N g u m C h e P h D T h e s i s
27
phosphorylation of IRS at its serine residue, rendering it inactive [25]. When this occurs,
the translocation of GLUT 4 is obstructed and the result is reduced glucose uptake.
Figure 1: IR in muscles results in defect in GLUT 4 signaling (Adopted from [26])
Irresponsiveness of GLUT-4 to insulin hinders its expression and translocation to the plasma
membrane, thus, reducing the ability of glucose uptake in the muscles.
It was suggested that exercise and muscle contraction in the absence of insulin can
stimulate glucose uptake [27] and even restore insulin sensitivity in obese mice [28]. It has
actually been proven that glucose uptake in muscles can also occur in a non-insulin
dependent manner, and that IR could actually occur in muscles with an intact insulin-
signaling system [17]. The activation of AMPK activity has been suggested to be
responsible for this non-insulin control of glucose [17]. Moreover, lipids accumulating in

B r i t a N g u m C h e P h D T h e s i s
28
muscles generate ceramides and other intermediates, which have been linked to the
development of IR [29], not only through the inhibition of protein kinase B (PKB) or IRS-1
but also by the generation of oxidative stress and apoptosis [25, 30, 31]. Even so, defects in
fat tissue and liver are extended to skeletal muscles [32], and because of this cross-talk
between tissues, other pathways and organs are involved in the generation of IR in the
muscles. An illustration of the link between adipose tissues, liver and muscles in the
generation of IR is shown on Figure 2.
Figure 2: Defects in other organs contributes to IR in the muscles (Adapted from [25])
Dysfunction due to for example changes in the partitioning and storage of lipids in the liver adipose
tissue or muscle results in IR, through decreased oxidation capacity of the muscles

B r i t a N g u m C h e P h D T h e s i s
29
1.4.2 Obesity
Obesity correlates strongly with T2D and arises as a result of excess body fat [33, 34].
Excess food intake and inactivity are known to be principal causes of obesity. An extensive
review on the many other causes of obesity, which includes smoking, medication,
pollutants and genetic factors, have been done by Keith et al [35]. The body mass index
(BMI) is used as a generic measure of obesity as it is closely related to the total body fat
[36]. A person with a BMI ≥ 30 kg/m2 is considered obese, and once this is the case, their
waist circumference is not considered during diagnosis [11]; see Table 2. Obesity increases
the prevalence of many pathological factors that constitute MS, including diabetes [34]. In
Europe, it is responsible for 80 % of type 2 diabetic incidents and 55 % of diseases related
to high blood sugar [37]. As much as 1 million deaths in Europe each year are related to
excess body weight [37].
Table 2: Classification of obesity using BMI (adopted from [38])
BMI range – kg/m2 Health Risk
≥ 30 Obese (high risk of developing MS)
25-29.9 Overweight (moderate risk of developing MS)
18.5 to 24.9 Healthy range (low risk)
< 18.5 Underweight (risk of developing problems such as nutritional
deficiency and osteoporosis)

B r i t a N g u m C h e P h D T h e s i s
30
1.4.3 Type 2 diabetes (T2D)
Contrary to type 1 diabetes, which is insulin-dependent and affects 10 % of patients
suffering from diabetes [39], T2D is a non-insulin dependent disease that affects 90 % of
diabetic patients and is common in old age but is now spread to the entire population [40].
The incidence of cardiovascular diseases is elevated four-folds when T2D is involved [41],
and it has been estimated that the living costs of obese men and women are 15 % and 21 %
higher, respectively, when compared to healthy individuals, regardless of their shorter
lifespans [38].
In Europe, over 50 million people are affected by T2D, and in Denmark, every twelfth
person is diabetic, but only half of them are aware of it [42]. Furthermore, it is estimated
that about 0.5 million people in Denmark are pre-diabetic [42]. A central player and
predictor of T2D is IR [43]. Other disorders such as oxidative stress, hyperglycemia and
dyslipidemia and β-cell malfunction also result in T2D [21, 39, 44, 45]. Figure 3 illustrates
the metabolic progress of T2D.
Figure 3: The metabolic progress of T2D (adopted from [13] )
The onset of T2D starts with the presence of excess fat, gene defects and/or aging. These factors
cause the muscle to become intolerant to insulin. As a result, glucose levels become elevated and
the β-cells of the pancreas produce more insulin to cope with the high glucose level. Eventually,
other tissues such as adipose tissue and liver become defective due to either a change in their fat
content or rate of glucose production, respectively. They become IR and contribute to increase
plasma glucose. The stress on the β-cells of the pancreas is even more, which with time, becomes
ineffective in its insulin production. Eventually, the whole body becomes intolerant to glucose.

B r i t a N g u m C h e P h D T h e s i s
31
1.5 The Satellite cell (SC)
Skeletal muscle is one of the few adult tissues that possess the ability to regenerate itself,
and credit for this ability is given to a population of dormant cells residing within the
muscles and known as satellite cells (SC). SC were first described by Mauro A. in 1961 as
mono-nucleated cells situated between the basal membrane and cell membrane of fibers,
and coexisting alongside multinucleated muscle fibers, as the name implies [46]. They are
quiescent by nature and located between the sarcolemma and basement membrane of
terminally- differentiated muscle fibers [47, 48]. The SC provide nuclei to the growing
muscle fibers during postnatal development [49]. More so, it has been revealed that SC are
crucial in the repair of damaged fibers by facilitating their re-innervation and
B r i t a N g u m C h e P h D T h e s i s
32
vascularization [50]. Although SC retain the ability to grow and differentiate during adult
life, they however, acquire a quiescent nature and become active only upon response to
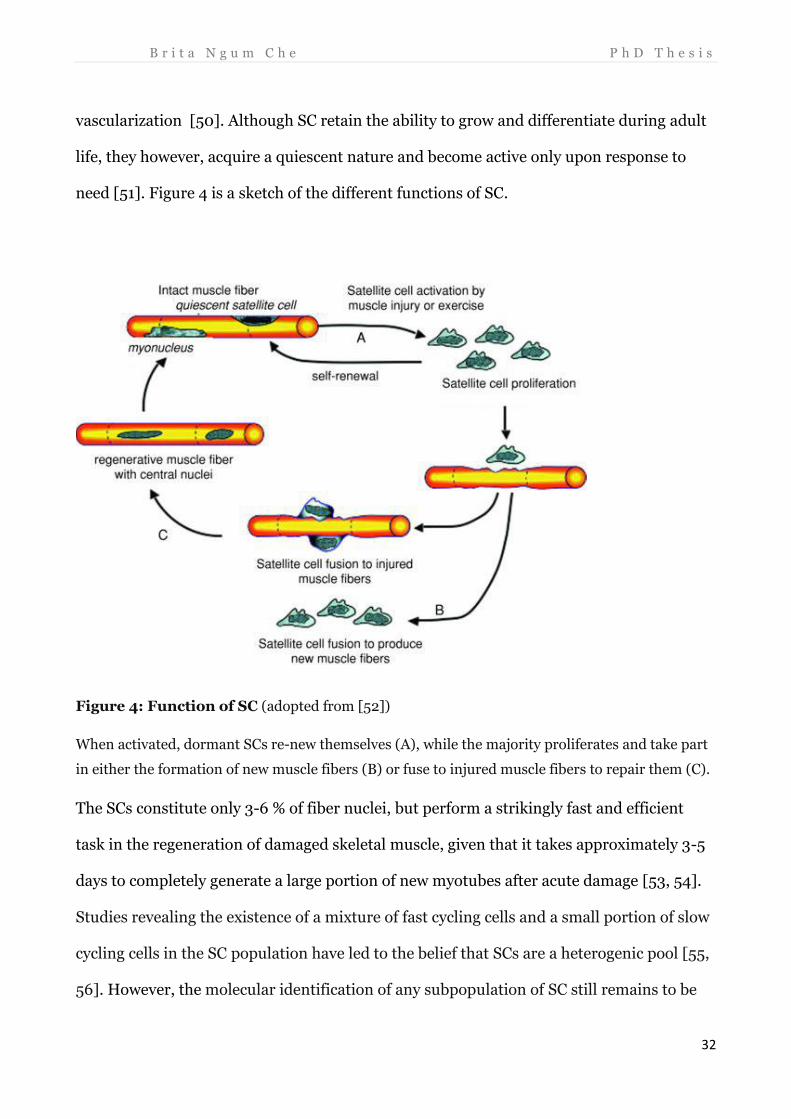
need [51]. Figure 4 is a sketch of the different functions of SC.
Figure 4: Function of SC (adopted from [52])
When activated, dormant SCs re-new themselves (A), while the majority proliferates and take part
in either the formation of new muscle fibers (B) or fuse to injured muscle fibers to repair them (C).
The SCs constitute only 3-6 % of fiber nuclei, but perform a strikingly fast and efficient
task in the regeneration of damaged skeletal muscle, given that it takes approximately 3-5
days to completely generate a large portion of new myotubes after acute damage [53, 54].
Studies revealing the existence of a mixture of fast cycling cells and a small portion of slow
cycling cells in the SC population have led to the belief that SCs are a heterogenic pool [55,
56]. However, the molecular identification of any subpopulation of SC still remains to be

B r i t a N g u m C h e P h D T h e s i s
33
achieved, and even though cells of non-muscular origin can contribute to muscle
production, it has not been possible to achieve efficient muscle regeneration using these
cells [57]. Much evidence points instead to a more homogeneous nature of SCs. It is
believed that following activation, SCs adopt a bifacial state, whereby the maintenance of a
satellite pool as well as proliferation to generate new nuclei is established [58]. This dual
fate is necessary for the SC to maintain a self-renewal pool of muscle progenitor cells [58].
1.5.1 From SCs to skeletal muscles
The transition of SCs to muscles occurs through the processes of activation, proliferation
and differentiation of the quiescent SCs (see Figure 4) and is effected by various
transcription and growth factors including insulin growth factor-1 (IGF-1) [59, 60]. While
a small portion of the activated SCs down-regulate the myogenic differentiation gene
(MyoD) but maintain the paired box (PAX)-7 gene to re-attain a quiescent state of renewed
SCs [58], the majority of the activated SCs loose PAX-7 and, thus, proliferate into
myoblasts [61]. Proliferating myoblasts are therefore characterized by a high expression of
MyoD, and in its absence, muscle regeneration is delayed [62]. The up- and down-
regulation of myogenin and myostatin, respectively, allows for the differentiation of
proliferating myoblasts [61, 63-66], while insulin growth factor-I (IGF-I) not only activates
proliferation, but is also involved in the innervation of the differentiated myoblasts [59, 60,
67]. The differentiation of myoblasts is also marked by the upregulation of IGF-II and IGF-
I and II receptors.[68], when compared to the proliferation process [68]. Skeletal muscle
actin and myosin heavy chain are typical markers of mature muscles, which are present at
the late stages of differentiation [54, 69]. Figure 5 shows the morphology of muscles cells
during myogenesis.

B r i t a N g u m C h e P h D T h e s i s
34
A B
C D
Figure 5: Transition from SCs to myotubes (adopted with modifications from Figure 1, MI)
Isolated porcine SCs cultured for 24 h on a matrigel matrix (A) proliferate into 50 % confluent
myoblasts after 3 days (B) and by day 5, myoblasts are 100 % confluent (C). The closely packed
myoblasts start to differentiate, and serum reduction enhances the differentiation process to
generate multinucleated myotubes (D).
1.6 Insulin signaling in skeletal muscles
Skeletal muscle is the main site of insulin action and accounts for over 70 % of glucose
disposal after a meal [70]. Insulin is a hormone that has a crucial role in the regulation of
whole body metabolism, and defects in its signaling generate IR, as discussed earlier. The
binding of insulin to its receptors initiates an auto-phosphorylation of the receptor and
sets off a signaling cascade, which eventually causes the translocation of GLUT 4 receptors

B r i t a N g u m C h e P h D T h e s i s
35
to the plasma membrane to ease the transportation of glucose [71, 72]. The
phosphorylated insulin receptor activates IRS-I and II by phosphorylating them at their
tyrosine residues [73]. Many signaling routes are affected by the phosphorylated IRS, but
the important pathway in the regulation of glucose is the phosphatidyl inositol 3-kinase
(PI3K) route [74]. PI3K, once activated catalyzes the formation of phosphatidylinositol 3,
4, 5- triphosphate (PIP) and the subsequent phosphorylation of the phosphoinositide
dependent protein kinase-1 (PDK1) enzyme [75]. It is the activated PDK1 that
phosphorylates protein kinase B (PKB) and a subtype of protein kinase C (PKCλ), and this
action effectuates the translocation of vesicles harboring GLUT4 [76, 77], via the action of
the enzyme Akt substrate 160 (AS160) [77]. PI3K has also been shown to activate PKB
through the mammalian target of rapamycin complexed with Rictor (mTORC2) [75]. The
action of insulin also causes PKB to catalyze the phosphorylation of Glycogen synthase
kinase (GSK3), which renders it inactive and allows for multiple dephosphorylation and,
hence, activation of GS [78]. Figure 6 is a sketch showing some of the enzymes involved in
insulin signaling.
Figure 6: Insulin signaling and glucose uptake in skeletal muscles (adopted from [77]).
The presence of glucose in plasma initiates the production of insulin by the beta cells in the islets of
the pancreas. The released insulin binds to its receptor on the plasma membrane and activates a
cascade of enzymes through phosphorylation. At the end of the reaction cascade is PKB, whose
activation leads to the transcription of GLUT 4 and translocation of GLUT 4 vesicles to the plasma
membrane, where they facilitate the transport of glucose into the cells.

B r i t a N g u m C h e P h D T h e s i s
36
1.6.1 Glucose uptake
Glucose uptake has been defined as the rate limiting step in the metabolism of glucose in
the muscles [79]. The regulation of glucose uptake in the muscles is accomplished by at
least GLUT 1 and GLUT 4 [80]. It has been established that GLUT 4 is the protein
responsible for the insulin-mediated glucose uptake in skeletal muscles while GLUT 1 is
responsible for basal glucose disposal, i.e. not dependent on insulin action [16, 81].
In the presence of insulin, the GLUT 4 vesicles, which are normally situated around the
nuclear space and in the cytosol, are activated to translocate and merge to the PM. The
insulin-responsive aminopeptidase (IRAP), which co-locates with insulin in the vesicles,

B r i t a N g u m C h e P h D T h e s i s
37
has been documented to assist insulin in the translocation process of GLUT 4 [82, 83].
GLUT 1 is found in the cytosol but also in the PM at basal conditions. It has been reported
that non-insulin-dependent mechanisms such as hypoxia and contraction also cause an
increase in the uptake of glucose, via the activation of AMPK [84, 85]. It has also been
revealed that hypoxia not only stimulates the expression and translocation of GLUT 1, but
also reduces the degradation of GLUT 1 [86, 87]. Therefore, one can conclude that it is
partly GLUT 1 that is responsible for the activation of glucose uptake in these instances.
1.6.2 Glycogen synthesis
The glucose taken up by the muscles can be used in many metabolic processes. About
50,000 glucose moieties make a single glycogen molecule, and 80 % of the glycogen in the
body is stored in skeletal muscles [88]. The principal role of glycogen in the muscle is to
produce energy during high level activity [89], whereas glycogen in the liver is utilized for
gluconeogenesis during fasting [77]. During glycogen synthesis, glucose taken up by the
cells is converted to glucose-6-phodphste, which is further converted into uridine
diphosphate (UDP)-glucose and finally to glycogen [25]. The enzymes hexokinase II and
GS are responsible for these processes, which are rate limiting in the synthesis of glycogen
[25]. Figure 7 shows the enhancing action of insulin on glycogen synthesis in primary
porcine cultures.
Other factors involved in the synthesis and regulation of glycogen in the muscles include
the auto-glucosylating protein; glycogenin, which is responsible for forming
oligosaccharide necessary for the further build-up of glycogen [90]. The glycogen
degrading enzymes; glycogen debranching enzyme and glycogen phosphorylase, which are

B r i t a N g u m C h e P h D T h e s i s
38
highly expressed in fast-twitched glycolytic fibers also regulate the synthesis of glycogen
[91].
b
*
Insulin concentration (nM)
0 1 10 100Fo
ld c
han
ge i
n g
lyco
gen
syn
thesis
(d
pm
/mg
pro
tein
)
0.9
1.0
1.1
1.2
1.3
1.4
1.5
a
ab
bc
cd
Figure 7: Glycogen synthesis is regulated by insulin (Adopted with modifications from M I,
Figure 4b)
Glycogen synthesis in primary porcine myotubes is enhanced by increasing amount of insulin. Data
points with different letters (a-d) denote significant difference.
1.7 β-oxidation
As skeletal muscles are not tuned for storing fats, they need a constant supply of energy,
which is provided by FAs generated from lipolysis in adipocytes. β-oxidation is the prime
route for the degradation of FAs [92] and is a core energy producing process of the cell,
which occurs in both the peroxisomes and mitochondria [92, 93]. Long-chain FAs once

B r i t a N g u m C h e P h D T h e s i s
39
transported into the cell for oxidation are directed towards the mitochondria, and these
processes are aided by transport and binding proteins [94-96]. The enzyme acyl-Coenzyme
A (acyl-CoA) synthase converts the FAs into acyl-CoAs, which are active intermediates of
FAs that are recognizable by enzymes involved in FA oxidation (FAO).
1.7.1 Acylcarnitine shuttling
The acyl-CoAs have to be shuttled into the mitochondria or the peroxisomes, where the
enzymes responsible for β-oxidation are situated [97]. L-carnitines (referred to hereafter
as carnitines) and their esters; acylcarnitines, are indispensable compounds in the β-
oxidation of FAs, with high levels present in the kidney, heart, and skeletal muscles [98,
99]. External sources of carnitines include dairy products and fish, but they can also be
produced endogenously by the kidney, heart and brain [99]. Carnitines are transported
into the cells by the aid of the sodium-dependent carnitine transporter; organic cation
transporter number 2 (OCTN2), and assist in the transportation of FAs by forming esters
of the FAs (acylcarnitines) that can easily be transported across the mitochondrial matrix
to undergo FAO [100]. See Figure 8 for a sketch describing acylcarnitine shuttling.
The esterification process is catalyzed by the enzyme carnitine-palmitoyl transferase I
(CPT I), using free Coenzyme A (CoASH), and occurs at the hydroxyl group of L-carnitine.
Unlike acetyl- and palmitoyl-transferases, the enzyme carnitine-acylcarnitine translocase
(CACT), has a wide specificity, ranging from free L-carnitine to acylcarnitine of all chain
lengths [101]. CACT assists in the transportation of acylcarnitines across the inner
mitochondrial matrix [101].

B r i t a N g u m C h e P h D T h e s i s
40
Once the acylcarnitine is inside the mitochondria, the enzyme carnitine-palmitoyl
transferase II (CPT II) helps in the release of L-carnitine and regeneration of acyl-CoA. The
reversible nature of the esterification of FAs with carnitines, not only enables
transportation of the FAs across the mitochondrial matrix, but also holds the content of
acyl-CoA in the cell at a non-toxic level [102, 103]. With the aid of short chain- acyl-
transferases, the carnitine system also assists in the transportation of short-chain FAs
from the peroxisome into the mitochondria [104, 105].
Figure 8: Acylcarnitine shuttling and FAO in the mitochondria (adopted with
modifications from [97])

B r i t a N g u m C h e P h D T h e s i s
41
Activated FAs in the form of acyl-CoAs are shuttled into the mitochondria in the form of
acylcarnitine and by the help of the enzymes CPT I, CACT and CPT II. Acyl-CoA dehydrogenases
and the trifunctional proteins execute a chronological degradation of the FAs to release acetyl-CoA
as the end-product of β-oxidation, which is shuttled into the kreb’s cycle
.
1.7.2 The β-oxidation enzymes
Many enzymes with specificity to either the long, medium, or short acyl-CoAs are involved
in the process of β-oxidation [106]; see Figure 9. Four main enzyme groups namely acyl-
CoA dehydrogenase (ACAD), enoyl-CoA hydratase (EH), 3-hydroxyacyl-CoA
dehydrogenase (HAD) and 3-ketoacyl-CoA thiolase (KAT), are responsible for the cyclic
degradation of acyl-CoAs by reducing a two carbon atom during each cycle [97, 107]. A
thorough review of ACADs has been achieved [108]. EHs, HADs and KATs are members of
the mitochondria trifunctional protein (MTP), which is a hetero-octomer consisting of four
α- and β- units, each [109]. The activities of EHs and HADs are situated within the α- unit,
while that of KAT is found in the β-unit [97, 110].
ACADs are known to exist in many forms in humans and depending on their chain-length
specificity to acyl-CoA, they are called short-chain acyl-CoA dehydrogenase (SCAD),
medium-chain acyl-CoA dehydrogenase (MCAD) or very long-chain acyl-CoA
dehydrogenase (VLCAD)[97]. ACAD are responsible for the first step of β-oxidation, where
FAD is necessary to dehydrogenate the long-chain acyl-CoA conjugate, producing a trans-
2-enoyl-CoA as intermediate [97] see Equation 1.
EHs exist in many forms in the mitochondria and include long-chain enoyl-CoA hydratase
(LCEH) and short-chain enoyl-CoA hydratase (SCEH), and their activities are different
from those in the peroxisomes [111]. LCEH and SCEH catalyze the addition of water to the

B r i t a N g u m C h e P h D T h e s i s
42
long-chain trans-2-enoyl-CoA and short-chain trans-2-enoyl-CoA substrates, respectively,
to generate L-3-hydroxyacyl-CoA [97, 112]; see Equation 2.
With regards to HAD, long-chain hydroxyacyl-CoA dehydrogenase (LCHAD) and short-
chain hydroxyacyl-CoA dehydrogenase (SCHAD) are two of the many forms that exist [97].
While LCHAD is specific only for long-chain L-3- hydroxyacyl-CoA, SCHAD has a broader
specificity and accepts substrates in the range of short chain (C4) to long chain (C16),
emphasizing on its importance in β-oxidation. HAD catalyzes the formation of 3-ketoacyl-
CoA from L-3-hydroxyacyl-CoA, using the cofactor NAD+ [113] see Equation 3.
Three types of KATs have been detected in the mitochondria and are responsible for the
conversion of 3-ketoacyl-CoA to an acyl-CoA that is short of two carbon atoms [114, 115];
see Equation 4. Medium-chain ketoacyl-CoA thiolase (MCKAT) effectively degrade
medium-chain FAs, as defects in this enzyme lead to fatal oxidation disorders [116]. Long-
chain ketoacyl-CoA thiolase (LCKAT) is crucial for the efficient degradation of LCFAs
[117], and utilizes substrates ranging from 3-ketohexanoyl-CoA to 3-ketopalmitoyl-CoA
[97]. The end-product of β-oxidation; acetyl-CoA, is either shuttled into the Krebs cycle to
produce ATP, or converted into acetylcarnitine and exported out of the mitochondria [97].

B r i t a N g u m C h e P h D T h e s i s
43
1.8 Peroxisome proliferator-activated receptors (PPAR)
The peroxisome proliferator-activated receptors (PPAR ) are members of the nuclear
receptor hormone family and consist of three isoforms (PPARα, PPARβ and PPARγ),
which control the expression of a well of genes by binding to peroxisome proliferator
response elements (PPRE) on the DNA strand [118]. Just like all nuclear receptors, the
PPARs consist of a DNA-binding domain (DBD) and a ligand-binding domain (LBD).
When activated by the binding of FAs or synthetic ligands to the LBD, PPAR forms
heterodimers with the retinoid X receptors (RXR), and this action causes their release
from the co-repressor [119]. The PPAR: RXR complex binds to PPRE via the DBD, and
initiates the expression of genes involved in lipid and glucose metabolism [120-122]. See
Figure 9 for a sketch of the mode of action of PPARs. Several unsaturated FAs, long-chain
FAs and branched-chain FAs are natural ligands of PPARs. As discussed earlier, PA is also
a natural ligand of the PPARs, especially PPARα [123-125].

B r i t a N g u m C h e P h D T h e s i s
44
Figure 9: Sketch of mode of action of PPARs (Adopted from [126] )
1.8.1 PPARα
PPARα is highly expressed in many organs, including skeletal muscle, liver, heart brown
adipose tissue and kidney [127]. PPARα activates FA uptake, transport and oxidation, as
well as regulates lipolysis and fat storage [128]. Attempts to use PPARα as targets of
treating insulin resistance and T2D have been made [129, 130]. Synthetic ligands of
PPARα include fibrates and the drug WY-14643 [131, 132], and they have been proven
effective to mend metabolic malfunction related to MS [133]. One possible mechanism of
action of PPARα is by a stimulation of glucose utilization in the muscles and adipose
tissue, resulting in a reduction in the glucose and insulin plasma levels [134, 135].

B r i t a N g u m C h e P h D T h e s i s
45
1.8.2 PPARβ
PPARβ is also involved in FAO [136]. Recent findings about PPARβ show that it helps
against heat injury in fibroblasts by increasing cell proliferation [137]. It has also been
shown to improve myogenesis through the down-regulation of myostatin expression [138],
and it is involved in the prostaglandin-induced activation of blastocyst development in
mice [139]. Synthetic agonists of PPARβ include GW501516, which has been reported to
inhibit dyslipidemia, by slowing down the activity of cholesterol ester transfer protein, and
reducing the content of very low-density lipoprotein [140].
1.8.3 PPARγ
PPARγ is mainly expressed in adipose tissues but it is also found in other tissues such as
the liver [127]. The isoform exists in two forms (PPARγ1 and PPARγ2), and it is PPARγ2
that predominates in adipocytes [141]. It activates protein synthesis and adiponectin
secretion in adipocytes, and it is crucial for efficient white and brown adipocyte
differentiation [142]. PPARγ also regulates lipogenesis in the liver [143] and increases
insulin sensitivity in adipocytes, thus, ameliorating the dysfunction of fat cells in T2D
[144]. In muscles, PPARγ also increases insulin sensitivity and improves FAO [121, 145].
Targeting PPARγ for the modulation of IR is common [145, 146]. The thiazolidinedione
(TZDs) are well known synthetic ligands of PPARγ, and they are important in the
treatment of T2D [147]. Some mechanisms employed by TZDs in improving IR are by
stimulating the production of adiponectin, which suppresses gluconeogenesis, and
reducing the level of circulating free FAs through their esterification to glycerol [148, 149].
Nevertheless, PPARγ has shown to induce the production of adiposites [150, 151].

B r i t a N g u m C h e P h D T h e s i s
46
1.9 Phytanic acid (PA)
1.9.1 Origin of PA and its diastereomers
The alcohol unit of chlorophyll; phytol, is the substrate for PA production (Figure 10). In
ruminants and in marine environments, oxidative processes of bacteria lead to the first of
the four steps in the degradation of chlorophyll; cleavage and oxidation of phytol from the
porphyrin ring of chlorophyll [155-157]. This step is the rate limiting step in chlorophyll
catabolism of chlorophyll and is controlled by the enzyme chlorophyllase [157]. The
generated phytol exists in two forms; E-phytol and Z-phytol, and when ingested by animals
or humans, can undergo a series of enzymatic reactions, including biohydrogenation, to
generate phytanic acid and afterwards, phytanoyl-CoA [158, 159], which can be
metabolized further. It was observed that E-phytol is the preferable substrate for the
PPAR-related metabolism of phytol [159, 160]. This conclusion was drawn from the fact
that E-phytol accumulates in the liver of PPARα-knockout mice fed phytol-rich diets [159].
More so, the enzymes responsible for phytol conversion to phytanoyl-CoA turned out to be
stereospecific [159]. It has also been shown that the aerobic and anaerobic activity of
bacteria on phytol generate both E- and Z-phytenic acid [161]. Thus, the role of bacteria in
the degradation of phytol is crucial, as these bacteria possess enzymes that are required for
the decarboxymethylation and oxidation of methyl-branched metabolites, without the
need of oxygen or the enzymes normally involved in methyl-branched FAs [162].

B r i t a N g u m C h e P h D T h e s i s
47
Figure 10: Production of PA from phytol (Adopted with modifications from [158])
The generation of Z- and E-phytol from chlorophyll requires bacterial action. Both forms of phytol
can be differentially processed to phytenic acid, which in turn, is converted to either RRR PA or
SRR PA by an enoyl-CoA reductase. The esterification of both forms of PA generates RRR- and
SRR-phytanoyl-CoA.
PA is 16 C- long and has four methyl groups positioned at carbons 3, 7, 11 and 15, with the
carbon 3, 7 and 11 being chiral centers. Carbons 7 and 11 have R configurations, just like in
phytol, while carbon 3 is either R- or S- configured owing to the biohydrogenation of

B r i t a N g u m C h e P h D T h e s i s
48
oxidized phytol. For this reason, PA occurs naturally as two diastereomers; SRR PA and
RRR PA [163]; see Figure 10. The SRR form has been shown to predominate in marine
animals while some terrestrial mammals have more of the RRR form [163-165]. Little is
known about the production and distribution of the two forms of PA in ruminal products
or about their physiological role. PA can be stored in the phospholipid and neutral
fractions of lipids in tissues and in milk [166]. Since humans lack the ability to cleave
phytol from chlorophyll [167], PA can only be obtained through the consumption of
ruminant or marine products [168, 169]. Differential deposition in lipid fractions of tissues
and metabolic rates of the diastereomers insinuates possible differences in the biological
activities of the two natural diastereomers of PA in humans [166, 170, 171].
1.9.2 Metabolism of PA in humans
After a phytol-rich meal, phytol is usually transported to the liver for catabolism, where
phytenic, phytanic and pristanic acids are formed [160]. Free phytol or PA taken up from
the diet can be converted into phytanoyl-CoA by the enzyme phytanoyl-CoA hydroxylase,
and further esterified and stored in lipids, hydrolyzed back to PA or used as a substrate for
alpha (α)- oxidation [172, 173]. Phytanoyl-CoA can also come directly from the diet, since it
is produced in ruminants. β-oxidation of PA is not directly possible due to the presence of
a methyl group on carbon 3 of PA ( see structure of PA in Figure 10).
The α- oxidation of PA occurs in the peroxisome [174] and the enzymes involved in the
process are illustrated in Figure 11. The actions of the hydroxylase and lyase enzymes on
phytanoyl-CoA and 2-hydroxylphytanoyl-CoA, respectively, are involved to produce
pristanic acid, with the release of formic acid as the primary product of α- oxidation [173,

B r i t a N g u m C h e P h D T h e s i s
49
175, 176]. Pristanic acid is further converted by a synthase reaction to pristanoyl-CoA, and
is used for β- oxidation. Phytanoyl-CoA and pristanoyl-CoA, thus, are the substrates for α-
and β- oxidations, respectively [173].
Another bottle neck in the degradation of PA is the presence of an R-form [177]. The
enzymes responsible for β- oxidation are stereospecific [178], and as such, an α-
methylacyl-CoA racemase (AMCAR) is required to convert 2R-pristanoyl-CoA to its 2S
isomer before it can be oxidized [93]; see Figure 11. At first, two cycles of β-oxidation, give
rise to 2, 6, 10 trimethylundecanoyl-CoA, which is R-configured. A racemase action
converts this product into its S-isomer, and a third β-oxidation is performed to generate 4
,8 dimethylnonanoyl CoA and short-chain acyl-CoAs [179]. These products of peroxisomal
β-oxidation are either hydrolyzed to acids (route 1) or esterified to carnitine esters by the
enzymes carnitine O-octanoyltransferase (COT) and carnitine acyltransferase (CAT), and
eventually transported to the mitochondria for further β-oxidation or processing (route 2)
[159, 173]. Besides acetate, propionate has been identified as the main degradation product
of PA oxidation [173, 180].
PA can also be degraded via omega (ω)-oxidation. For more information on ω-oxidation of
PA, see the reviews by Komen et al. (2004) [181] and Wanders et al. (2011) [182].

B r i t a N g u m C h e P h D T h e s i s
50
Figure 11: Peroxisomal α- oxidation and subsequent β-oxidation of PA (Adopted with
modifications from [173])
Phytanoyl-CoA and pristanoyl-CoA are the substrates for α- and β- oxidation, respectively.
Hydrolysis or esterification of the final peroxisomal oxidation products are transport to the
mitochondria for further processing.

B r i t a N g u m C h e P h D T h e s i s
51
1.9.3 Milk fat composition of PA as affected by cow feed
Organic farming practices and consequently milk fat composition vary with climate and
country [183]. More so, differential feeding strategies have been shown to improve the
quality of milk [184, 185]. Grazing or green grass feeding is an obligatory part of organic
farming in Denmark but other components of feed include maize and concentrates. Being
a by-product of chlorophyll, the content of PA in milk fat ought to correlate with greed feed
intake and has been reported so in various studies [169, 186]. As such, the content of PA in
organic milk has been proposed as a marker for organic dairy products [169]. In addition,
the distribution of PA diastereomers can be used for a better authentication of organic milk
[187]. It was recently shown that the distribution of PA diastereomers changes from more
to less RRR PA, when feeding shifts from green feed (organic) to concentrates
(conventional) [165, 188]. However, very little is known of the physiological roles of the
diastereomers of PA.

B r i t a N g u m C h e P h D T h e s i s
52
2 RESULTS AND DISCUSSION
In this section, the results regarding the use of primary porcine myotubes as a model for
skeletal muscles to study glucose and FA metabolism will be discussed. Furthermore, the
results of the effects of PA on the metabolic parameters, glucose uptake, glycogen
synthesis, and FA β-oxidation, shall be discussed in relation to glucose and FA
homeostasis. The effects of excess PA in humans shall be adressed and the impact of cow
feed on the content of PA in milk-fat will be elaborated. Finally, the metabolic implications
of the distribution of PA diastereomers in humans shall be discussed.
2.1 Primary porcine myotubes; a model of skeletal muscles, for studying
glucose and FA metabolism
The muscle is the major site for insulin-dependent glucose uptake [70, 189] and the initial
site in the generation of IR observed in T2D [189]. Therefore, the muscle represents an
ideal model to study the metabolic effects on glucose and FA homeostasis. The porcine
primary myotube model is similar to human myocytes in many ways [190], making it an
excellent choice. More so, because they are primary cells, they have not been modified in
any way other than enzymatic or physical dissociation from tissues, during purification.
Using well-established methods [191], SCs obtained from the semi-membranosus of
piglets were successfully proliferated and differentiated into multinucleated myotubes in
culture for use in this project (Figure 1, M I). As hypothesized, the porcine myotubes were
efficient for use in the analyses of glucose and FA metabolism.
In the analysis of glucose uptake, the myotubes were observed to harbor functional
insulin- mediated and non-mediated pathways, as revealed by their response to insulin

B r i t a N g u m C h e P h D T h e s i s
53
and by the inhibiting action of cytochalasin B in both pathways (Figures 2, M I). GLUT-1
and 4 were responsible for the basal and insulin-stimulated glucose uptake, respectively,
an observation that is common in human muscles [192], but the fact that 20 % of the
glucose taken up by the myotubes could not be accounted for by neither GLUT-1 nor
GLUT-4 denotes that there might be other transporters in the pig model that aid in the
uptake of glucose. Actually, GLUT-10, 11, and 12 are, for example, known to be expressed
in mammalian skeletal muscles, and may be involved in glucose uptake, as they bind to
glucose with different affinities [193-195]. So there is a possibility that these GLUTs are
also present in pigs. It should be noted that cytochalasin B independent glucose uptake of
about the same magnitude (20 %) has been observed before [196].
GLUT-1 and 4 each activated glucose uptake in the porcine myotubes by about 4o %
(Figure 2b, M I).The GLUT-4-mediated uptake seem low when compared to a study by
Nedachi et al. (2006), where 70 % of glucose uptake was insulin-mediated [70]. This
disparity could be due to the fact that Nedachi et al used a murine cell line for the glucose
uptake analysis, and furthermore, cell lines are known to produce more proteins and
GLUT-4 than primary cells, probably because cell lines do not age or become senescent as
normal cells [197].
Another explanation of the low level of insulin-mediated uptake registered could lie in the
fact that the myotubes generated for this study were isolated from the semi-membranosus
muscles (SM) of piglets, which consists predominantly of white-type fibers. White-type
fibers are known to be low in their contents of GLUT 4 and adiponectin [198, 199], both
factors of which are necessary for the insulin-mediation of glucose uptake in the muscles.
Actually, we have seen in our lab that red-type vastus intermedius muscles (VI) show 30 %
increased insulin-mediated glucose uptake, when compared to SM (unpublished data).

B r i t a N g u m C h e P h D T h e s i s
54
In muscles, up to 80 % of the insulin-activated glucose uptake is converted into glycogen
[200], underlining the importance of glycogen synthesis in the regulation of glucose
metabolism. In this study, the induction of glycogen synthesis in the porcine myotubes by
insulin was also effective (Figure 4b, M I), and hinted the up-regulation and activation of
GS [78]. In a previous study on insulin signaling in primary porcine myotubes, insulin
administration did not activate GS [201]. This controversial finding probably lies in
experimental methodology, rather than in the pig culture as a model, since cultures in the
latter example were exposed to supra-physiological concentrations of insulin for several
days before the analysis was performed [201]. The long-term exposure to insulin is the
likely cause of insulin-insensitivity, as hyperinsulinemia has been shown to cause IR [202].
More so, it is possible, that the chosen pigs for SC isolation was generally insulin
insensitive, especially as glycogenin which is also a rate limiting factor in glycogen
synthesis [203], was unaffected in the study [201]. We have seen in our lab that myotubes
isolated from different pigs vary in their insulin-regulated glucose uptake, with some
myotubes being completely insulin-resistant (unpublished data).
The porcine myotubes were also able to metabolize FAs. Their regulation of FA metabolism
seemed to lie in both the areas of uptake and oxidation of FAs. This is because the same
conditions that caused the generation of acetylcarnitine (C2); the end product of β-
oxidation, did not change or even reduced the degradation of palmitoylcarnitine (C16)
(Figures 4a and b, M II).
The myotubes could also be rendered resistant in both the uptake of glucose (Figure 5a, M
I) and the oxidation of FAs (Figure 3c, M II) using high concentration of glucose and
palmitate, respectively; a feature that is characteristic of human muscle cultures.

B r i t a N g u m C h e P h D T h e s i s
55
2.2 The metabolic impact of PA
During glucose uptake analysis, it was shown that physiological amounts of PA enhanced
glucose uptake in the primary porcine myotubes, especially at low concentrations of
insulin (Figure 4a, MI), while further increments in the concentration of PA did not further
improve glucose uptake (Table 2, MI). This finding revealed that PA and insulin may have
similar mode of action in activating glucose uptake, as a combination of both compounds
did not have any synergistic effect on the activation of glucose uptake. In a human
intervention study, where PA-rich dairy fat was administered, there were no significant
changes in the risk markers of MS (Cholesterol, triglycerides, C-reactive protein, serum
insulin and serum glucose) [204]. Especially the observation of an unchanged serum
glucose level after the intake of PA-rich fat [204], was controversial to the results in our
project, and could be attributed to other involved factors that are present at the organism
level. As shown by the involvement of many tissues in IR [25, 32], the regulation of glucose
involves many organs, and studies of the metabolic effect of PA in an organism, as a whole,
presents an intermix of many pathways whose results can be different from those obtained
from studies in single tissue (myotubes).
It was observed that the time-dependent activation of glucose uptake by PA was biphasic,
as incubation of myotubes with PA for 4-8 h turned to increase glucose uptake, while
incubation for 12-16 h reduced glucose uptake and by 24-h incubation, there was an
increase again in glucose uptake (Figure 3a, MI). It was suggested that depletion of GLUTs
was the reason for the drop in glucose uptake, but given that the preceding rise in glucose
uptake was mild, other factors must have been involved in the biphasic effect.
PA-induced glucose uptake was significant after 60 min of incubation with glucose,
although at this time, the glucose uptake process seemed saturated (Figure 3b, MI);

B r i t a N g u m C h e P h D T h e s i s
56
glucose uptake during the first 15 min was steepest when compared to the last 15 min. This
observation showed that other factors are involved in the regulation of glucose uptake,
when PA is involved.
We showed that the glucose taken up by the myotubes in the presence of PA could not be
incorporated into glycogen, unless insulin was available (Figure 4b, MI). This observation
insinuates that the mechanism, of which PA activates glucose uptake, is different from that
which regulates glycogen synthesis. Thus, PA regulates glucose homeostasis in both an
insulin-dependent and insulin-independent manner. Our results also show that pathways,
other than glycogen synthesis are involved in the metabolism of the PA-incorporated
glucose.
When the myotubes were rendered insulin resistant by exposure to excess glucose (Figure
5b, MI), it was not possible for PA or insulin to stimulate glucose uptake or glycogen
synthesis (Figures 5c and d). One could, therefore, suggest PA to be beneficial in the
regulation of glucose in normal individuals, who generally have very low or insignificant
content of PA in their serum, or in persons that have inadequate amounts of insulin in
their serum.
During the analysis of β-oxidation in the myotubes using acylcarnitine profiling, PA
enhanced the content of C2 generated in the myotubes, as well as reduced the content of
C16, as shown by a reduced C16/C2 level (Figures 5a and b, MII). These findings denote
that PA, possibly in a PPAR-manner [133, 205, 206] improves β-oxidation by both
enhancing the degradation of C16 and increasing the content of C2.
The effect of PA on β-oxidation was not dose-dependent but longer incubation periods
with PA increased β-oxidation. During 4 h incubation of myotubes with PA, no change was

B r i t a N g u m C h e P h D T h e s i s
57
observed in the content of C16 (shown as C16/C2) and C2, but after 24 h incubation, the
content of C2 increased as the level of C16 fell and the tendency continued even when the
exposure to PA was increased to 48 h (Figure 5a and b, MII). PA has been shown to
improve FA uptake [207], which could be a reason for the unchanged level of C16, during 4
h incubation. To concretize the time-dependent effect, a lower concentration of PA (5 µM ),
which could not reduce the content of C16 or increase the level of C2 during exposure for 4
h, actually reduced and increased the levels of C16 and C2, respectively, after exposure for
48 h. These findings insinuate that PA needs time (possibly to generate more potent
metabolites [208]) to exert its effect on β-oxidation.
The fact that 5 µM PA exposed to the myotubes for 24 h caused a reduction in the content
of C16, but not an elevation of C2, suggests that PA might affects β-oxidation at different
levels and different magnitudes. Experiments with oleic acid (OLA); a saturated FA that
has been shown to have PPAR characteristics [209], revealed that it also improves β-
oxidation, at least at two levels, just like PA; at the level of uptake of FAs into the cell
and/or mitochondria, and at the level of β-oxidation of FAs. The regulation by PA, of
intermediate carnitines at the level of acyl-dehydeogenases, carnitine acyl-transferases and
MTPs cannot be excluded as a means of regulating FAO. In fact, the metabolism of PA is
known to up-regulate octanoyl-transferase [210], and possibly propionyl-transferase, given
that propionate is one of the main products of PA degradation [104, 173]. Octonoyl-
transferases have been implicated in the induction of β-oxidation of VLCFA and MCFA
[211].
Despite the observations regarding the induction of β-oxidation by PA, we could not
unequivocally conclude if it was via a PPAR-manner, since a PPAR control agonist was not
included in the study. Thus, the use of a positive control, for example a PPARα agonist

B r i t a N g u m C h e P h D T h e s i s
58
such as bezafibrates or the well-known synthetic agonist; WY-14643 [133, 205, 212], is
necessary to validate the results.
2.3 Concerns regarding excess content of total PA in humans
It is a desire to increase the total content of PA in milk fat, to achieve levels that are
physiologically advantageous. The PA content in normal human subjects that consume
ruminant products has been documented to range from 0.04-11.5 µM, and consumers of
non-ruminant products have negligible amounts of PA in their plasma, compared to
consumers of ruminant products, whose total content of PA is in the upper range [204,
213, 214]. In our studies, 10 µM PA was utilized as standard concentration as it lies within
the physiological range. More so, the viability of the myotubes were negatively affected
when the concentration of PA was over 10 µM (table 1, M I), and results from FAO
experiments with 20 µM PA where not generally better, when compared to results
achieved with 10 µM (Figure 5 b and c, M II).
2.4 PA content in milk-fat as affected by cow feed
As shown in Figure 1, M III, two naturally-occurring stereomers of PA were identified in
milk-fat, with the SRR form dominating, just as in previous findings [165]. Milk-fat
produced organically or even by conventional feeding has been shown to have higher
contents of PA than in our study [187, 188]. This discrepancy in the content of PA lies in
different farming practices between farms or regions/countries.

B r i t a N g u m C h e P h D T h e s i s
59
Surprisingly, we found that an increase in the total content of PA was attributed to feed
concentrates, while the reverse was attributed to the content of pasture in feed (Table 2, M
III). These correlations were, however not very strong and could be attributed to random
variation. Nevertheless, our finding could actually explain the difference in PA contents
between the grazing seasons. The total content of PA was higher in September than in
May, and this is probably because the use of concentrates was generally augmented in
September to compromise for the weather-mediated low dry matter intake (DMI) from
grazing in September, and not because of the content of legumes that constituted a larger
part of pastures in September. Nevertheless, it could be that the availability of light during
the May season (there is generally more light in the May than September season), also
affected the results. The degradation of chlorophyll in leaves have been shown to be
enhanced by photo-oxidation [215]. In this case, the production of phytol should be
enhanced and, thereby generating more PA during the May season. But given our findings,
it is obvious that other factors (e.g. the developmental state of the plants) affect the
production and content of PA in milk-fat more.
We suggested that the use of grass pellets in concentrates was the cause for the elevated
content of total PA, since grass feed has been shown to generate higher contents of PA
[186]. Actually, dried grass has been shown to increase the degradation of chlorophyll in
sheep rumen [216]. As PA can only be produced from free phytol, chlorophyll degradation,
therefore, will increase the production of PA.
The inverse correlation of pasture to the total content of PA observed in our studies
contradicts previous conclusions that green feeding increase the content of PA [169, 217].
In our experiments, pasture was dominated mostly by legumes (ryegrass and white clover).
Howard et al. (1993) found that legumes like soya beans present a “stay green”

B r i t a N g u m C h e P h D T h e s i s
60
phenomenon, whereby chlorophyll degradation is resisted [218]; maybe this phenotype is
common for the legumes in the pasture used in our study, and would thereby explain the
decrease in the total content of PA recorded. Nevertheless, as we found no correlation
between the total content of PA and legumes, it is obvious that other unknown factors are
involved in the production of PA. A study conducted by Lee et al. (2006) showed that the
total content of PA in duodenal flow was reduced in line with a reduction in
biohydrogenation [186]. Although this trend was affected by the intake of red clover, it is
likely that, partly the influence of legumes on biohydrogenation is causing the reduction in
the total content of PA after pasture intake, observed in our study.
Many factors other than proteolytic activities in the pasture might alter ruminal micro
flora [219], and, thus, responsible for driving the mechanisms behind our observations of
altered PA content with feed. The rate limiting step in the catabolism of chlorophyll, which
generates free phytol, is controlled by the chlorophyllase and has been showed to be post-
translationally regulated [157], possibly by a well of factors, some of which can originate
from the rumen of the cow. In that case, the feed can have an influence on chlorophyllase.
Phytol exists in two forms (E- and Z- form) and the degradation and production of each
form from chlorophyll depend on the bacteria community [161, 220], whose population
and action can be affected differently by the feed of the cow. Moreover, the enzymes
involved in the degradation of phytol are stereospecific, favoring the degradation of E-
phytol, and these enzymes can be altered by the feed [159].
The values of the total content of PA obtained in our study do not necessarily indicate the
lack or limited use of green feed in organic farming, but merely indicate that the type of
(green) feed can alter the acquired content of total PA in milk-fat. It is apparent, therefore,

B r i t a N g u m C h e P h D T h e s i s
61
to suggest that it is the feed composition that in many levels and ways controls the total
content of PA.
To support this suggestion, we also showed that although pasture did not affect the total
content of PA, the increasing content of legumes in the pasture caused an elevation in the
content of RRR PA (Table 2, M III). Schröder et al. (2011 and 2012) also reported that
organic feeding, which is dominated by green feed (grass silage) or conventional feeding
rich in concentrates, increased the content of SRR PA in milk-fat, while organic feeding
dominated by hay feeding increased the content of RRR PA instead [187, 188]. Likely
reasons for the altered content of PA diastereomers are similar to some of those that
caused an alteration in the total content of PA; namely, the altered distribution or action of
ruminal micro flora bacteria in response to feed, and also the stereospecificity of
degradation enzymes. As mentioned earlier, β-oxidation enzymes prefer SRR-pristanic
acid (generated from SRR PA) as substrate, and in that case, one would expect to have
different contents of the diastereomers in milk. More so, effects on AMCAR (enzyme
necessary to convert RRR PA to SRR PA) by feed may be possible.
2.5 Concerns regarding the distribution of PA diastereomers in
humans
The notion to elevate the total PA content of milk fat was borne with little knowledge of the
distribution of the individual stereomers of PA and what impact the diastereomers might
have on the physiology. Studies carried out by Tsai et al. (1973) showed that SRRR PA
instead of RRR PA was preferred for oxidation [170], but this conclusion was drawn from
experiments with very high concentrations of PA, like those found in a diseased state, and

B r i t a N g u m C h e P h D T h e s i s
62
revealed nothing about the PPAR-effects of PA. Nevertheless, Heim et al. (2002) later
showed that both forms of PA affected PPARs equally [124], but this study was performed
just in one tissue; the liver. Results from other tissues or organs would give more
convincing observations and information on the effects of PA diastereomers.
The knowledge that β-oxidation of FAs is stereospecific [178] with preference towards the
SRR form generates interests in the diastereomatic distribution of PA in milk fat. Because
of a methyl side chain on position C3, the metabolism of PA occurs through α- oxidation in
the peroxisome, resulting in a more potent PPAR-inducer; pristanic acid [208]. Pristanic
acid existing in the RRR-form has to be converted by AMCAR to the SRR form, for it to be
β-oxidized. These bottle-necks in the metabolism of PA create a scenario with regards to
what feeding strategy to embark on. On the one hand, feed composition that will give a
higher ratio of SRR/RRR PA could be thought desirable, as its metabolism would be
preferably faster and less dependent on AMCAR. On the other hand, it is hard to conclude
that milk-fat with higher RRR/SRR PA ratio is undesirable; one can speculate that a slower
degradation of RRR PA may cause an increase in the level of pristanic acid, and since
pristanic acid is a more potent PPARα agonist than PA, the expression of PPARα would be
up-regulated. However, it is important that the elevated level of PA or pristanic acid is
within a physiological range.

B r i t a N g u m C h e P h D T h e s i s
63
3 CONCLUDING REMARKS
As illustrated in Figure 12, we show in this study that manipulating the feeding strategy of
the cow can alter the total content and distribution of PA diastereomers in the milk-fat of
the cow. More so, we could show that the exposure of a physiologically relevant
concentrations of a mixture of PA isomers induced glucose uptake and β-oxidation in
primary porcine myotubes, which we established as an efficient skeletal muscle model.
However, the mechanisms behind the control of glucose and FA metabolism by PA remain
to be investigated.
Figure 12: An illustration of the effects of cow feed on the total content of PA and its
diastereomers, and the metabolic impact of PA on skeletal muscles

B r i t a N g u m C h e P h D T h e s i s
64
4 PERSPECTIVES
During glucose uptake analysis, we found that PA at physiological concentrations
enhanced glucose uptake but not glycogen synthesis. It would, thus, be relevant to
investigate the fate of the glucose after uptake and whether other metabolic pathways of
glucose, than glycogen synthesis are affected by PA. While PA, in the presence of insulin,
generated a synergistic effect on the activation of glycogen synthesis, no such effect was
observed during glucose uptake. These observations insinuate that more than one
mechanism is involved in the PA-mediated metabolism of glucose, and needs to be
analyzed further.
PA was observed to enhance the degradation of C16 and increase the production of C2.
However, a positive PPARα control, such as bezafibrates, needs to be investigated in
parallel with PA, before we can conclude on what mechanism is responsible for this effect.
More so, the effect of PA on intermediate acylcarnitines is relevant to get an overview of
the general impact of PA on FA metabolism.
In our studies, synthetic PA was used. Given that this product is a mixture of up to eight
isomers of unknown distribution, it is hard to tell whether our in vitro finding concerning
the metabolic impact of PA could have been better, if a pure PA product consisting only of
the two naturally known isomers had been used. Thus, studies with the purified naturally
occurring forms of PA are of interest. The finding that RRR PA can be enhanced by cow
feed makes it even more relevant to elucidate the metabolic effects of this form. Also,
analysis of other tissues than muscles would give more information about the distribution
and effects of PA diastereomers.

B r i t a N g u m C h e P h D T h e s i s
65
Gene expression experiments and protein determination of specific markers of glucose
uptake and FAO are necessary to fully concretize the findings in our studies. In this regard,
human muscle cell lines could be used as a supplement to the primary porcine model,
since cell lines generate enough proteins for biochemical and histological studies. More so,
the human muscles express uncoupling proteins that regulate thermogenesis, and this
process has been shown to be regulated by PA. Even more interesting is the fact that the
use of cell line would exclude constrains of dealing with variation between pigs, although
this factor is a natural aspect that needs to be kept in mind when dealing with cell lines.
Which of the natural forms of PA, that influence this metabolic process could be of
relevance.
In our analysis with Jersey and Holstein cows, we could not link breed to the total content
of PA or distribution of its diastereomers. As the composition of feed intake varied between
breeds, one can speculate whether the conclusion would have been the same, had it been
the same variation in feed composition applied to both breeds. Such an experiment could
be of relevance.
Drawing from the observations in this project, the desired content of PA and distribution
of its diastereomers in milk-fat can be achieved by manipulating the composition of the
cow feed. Therefore, it will be interesting to analyze the impact of the legumes and other
feed components commonly used in Denmark, on the total content PA and its
diastereomatic distribution. Analysis from the level of chlorophyll degradation, through
the breakdown of the two forms of phytol, micro flora involvement, and ruminal
production and degradation of the two forms of PA would be relevant.

B r i t a N g u m C h e P h D T h e s i s
66
5. REFERENCES
1. Zimmet P, Alberti KGMM, Shaw J. Global and societal implications of the diabetes epidemic. Nature 2001; 414: 782-87.
2. Meigs JB, Wilson PWF, Nathan DM, et al. Prevalence and characteristics of the metabolic syndrome in the San Antonio Heart and Framingham Offspring Studies. Diabetes 2003; 52: 2160-67.
3. Ford ES, Giles WH. A comparison of the prevalence of the metabolic syndrome using two proposed definitions. Diabetes Care 2003; 26: 575-81.
4. Balkau B, Drivsholm T, Borch-Johnsen K, et al. Frequency of the who metabolic syndrome in European cohorts, and an alternative definition of an insulin resistance syndrome. Diabetes and Metabolism 2002; 28: 364-76.
5. Ekelund U, Anderssen S, Andersen LB, et al. Prevalence and correlates of the metabolic syndrome in a population-based sample of European youth. Am J Clin Nutr 2009; 89: 90-96.
6. Eckel RH, Alberti KGMM, Grundy SM, et al. The metabolic syndrome. The Lancet 2010; 375: 181-83.
7. Sale MM, Woods J, Freedman BI. Genetic determinants of the metabolic syndrome. Current Hypertension Reports 2006; 8: 16-22.
8. Kuczmarski RJ, Flegal KM, Campbell SM, Johnson CL. Journal of the American Medical Association 1994; 272: 205-11.
9. Kuczmarski RJ, Flegal KM, Campbell SM, et al. Increasing prevalence of overweight among US adults: The National Health and Nutrition Examination Surveys, 1960 to 1991. Journal of the American Medical Association 1994; 272: 205-11.
10. Nielsen J, Christensen DL. Glucose intolerance in the West African diaspora: A skeletal muscle fibre type distribution hypothesis. Acta Physiologica 2011; 202: 605-16.
11. Alberti KGMM, Zimmet P, Shaw J. The metabolic syndrome - A new worldwide definition. Lancet 2005; 366: 1059-62.
12. Gallagher EJ, Leroith D, Karnieli E. Insulin resistance in obesity as the underlying cause for the metabolic syndrome. Mount Sinai Journal of Medicine 2010; 77: 511-23.
13. Saltiel AR. New perspectives into the molecular pathogenesis and treatment of type 2 diabetes. CELL 2001; 104: 517-29.

B r i t a N g u m C h e P h D T h e s i s
67
14. Shepherd PR, Karhn BB. Glucose Transporters and Insulin Action - Implications for
Insulin Resistance and Diabetes Mellitus. The New England Journal of Medicine 1999; 341: 248-57.
15. Stenbit AE, Tsao TS, Li J, et al. GLUT4 heterozygous knockout mice develop muscle insulin resistance and diabetes. Nature Medicine 1997; 3: 1096-101.
16. Marshall BA, Hansen PA, Ensor NJ, Ogden MA, Mueckler M. GLUT-1 or GLUT-4 transgenes in obese mice improve glucose tolerance but do not prevent insulin resistance. American Journal of Physiology - Endocrinology and Metabolism 1999; 276: E390-400.
17. Zierath JR, Krook A, Wallberg-Henriksson H. Insulin action and insulin resistance in human skeletal muscle. Diabetologia 2000; 43: 821-35.
18. Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001; 414: 799-806.
19. Freymond D, Bogardus C, Okubo M, et al. Impaired insulin-stimulated muscle glycogen synthase activation in vivo in man is related to low fasting glycogen synthase phosphatase activity. Journal of Clinical Investigation 1988; 82: 1503-9.
20. Saha AK, Xu XJ, Balon TW, et al. Insulin resistance due to nutrient excess: Is it a consequence of AMPK downregulation? Cell Cycle 2011; 10: 3447-51.
21. Bailey CJ, Turner SL. Glucosamine-induced insulin resistance in L6 muscle cells. Diabetes, Obesity and Metabolism 2004; 6: 293-98.
22. Monauni T, Zenti MG, Cretti A, et al. Effects of glucosamine infusion on insulin secretion and insulin action in humans. Diabetes 2000; 49: 926-35.
23. Denis McGarry J. Dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes 2002; 51: 7-18.
24. Hirabara SM, Curi R, Maechler P. Saturated fatty acid-induced insulin resistance is associated with mitochondrial dysfunction in skeletal muscle cells. Journal of Cellular Physiology 2010; 222: 187-94.
25. Shulman GI. Unraveling the cellular mechanism of insulin resistance in humans: New insights from magnetic resonance spectroscopy. Physiology 2004; 19: 183-90.
26. Zierath JR. In vitro studies of human skeletal muscle: Hormonal and metabolic regulation of glucose transport. Acta Physiologica Scandinavica, Supplement 1995; 626: 1-96.
27. Goodyear LJ, King PA, Hirshman MF, et al. Contractile activity increases plasma membrane glucose transporters in absence of insulin. American Journal of Physiology - Endocrinology and Metabolism 1990; 258: E667-72.

B r i t a N g u m C h e P h D T h e s i s
68
28. Dolan PL, Tapscott EB, Dorton PJ, et al. Contractile activity restores insulin responsiveness in skeletal muscle of obese Zucker rats. Biochem J 1993; 289: 423-26.
29. Haus JM, Kashyap SR, Kasumov T, et al. Plasma ceramides are elevated in obese subjects with type 2 diabetes and correlate with the severity of insulin resistance. Diabetes 2009; 58: 337-43.
30. Pickersgill L, Litherland GJ, Greenberg AS, et al. Key role for ceramides in mediating insulin resistance in human muscle cells. J Biol Chem 2007; 282: 12583-89.
31. Summers SA. Ceramides in insulin resistance and lipotoxicity. Progress in Lipid Research 2006; 45: 42-72.
32. Abel ED, Peroni O, Kim JK, et al. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001; 409: 729-33.
33. Friedman JM. Obesity: Causes and control of excess body fat. Nature 2009; 459: 340-42.
34. Golay A, Ybarra J. Link between obesity and type 2 diabetes. Best Practice and Research: Clinical Endocrinology and Metabolism 2005; 19: 649-63.
35. Keith SW, Redden DT, Katzmarzyk PT, et al. Putative contributors to the secular increase in obesity: Exploring the roads less traveled. International Journal of Obesity 2006; 30: 1585-94.
36. Gray DS, Fujioka K. Use of relative weight and body mass index for the determination of adiposity. Journal of Clinical Epidemiology 1991; 44: 545-50.
37. Tsigos C, Hainer V, Basdevant A, et al. Management of obesity in adults: European clinical practice guidelines. Obesity Facts 2008; 1: 106-16.
38. Nagai M, Kuriyama S, Kakizaki M, et al. Impact of obesity, overweight and underweight on life expectancy and lifetime medical expenditures: The Ohsaki Cohort Study. British Medical Journal, Open access 2012; 2.
39. Niedowicz DM, Daleke DL. The role of oxidative stress in diabetic complications. Cell Biochemistry and Biophysics 2005; 43: 289-330.
40. Alberti G, Zimmet P, Shaw J, et al. Type 2 diabetes in the Young: The Evolving Epidemic. The International Diabetes Federation Consensus Workshop. Diabetes Care 2004; 27: 1798-811.
41. Beckman JA, Creager MA, Libby P. Diabetes and atherosclerosis epidemiology, pathophysiology, and management. Journal of the American Medical Association 2002; 287: 2570-81.

B r i t a N g u m C h e P h D T h e s i s
69
42. The Danish Diabetes association. How to stop the diabetes epidemic in Denmark. Charlotte Rulffs Clausen, 2008.
43. Cahill J. Beta-cell deficiency, insulin resistance, or both? New England Journal of Medicine 1988; 318: 1268-70.
44. Watts GF, O'Brien SF, Silvester W, et al. Impaired endothelium-dependent and independent dilatation of forearm resistance arteries in men with diet-treated non-insulin-dependent diabetes: Role of dyslipidaemia. Clinical Science 1996; 91: 567-73.
45. Robertson RP. Chronic oxidative stress as a central mechanism for glucose toxicity in pancreatic islet beta cells in diabetes. Journal of Biological Chemistry 2004; 279: 42351-54.
46. Mauro A. Satellite cell of skeletal muscle fibers. Journal of biophysical and biochemical cytology 1961; 9: 493-95.
47. Morgan JE, Partridge TA. Muscle satellite cells. International Journal of Biochemistry and Cell Biology 2003; 35: 1151-56.
48. Bischoff R. Proliferation of muscle satellite cells on intact myofibers in culture. Developmental Biology 1986; 115: 129-39.
49. Moss FP, Leblond CP. Satellite cells as the source of nuclei in muscles of growing rats. Anatomical Record 1971; 170: 421-35.
50. Bischoff R, Heintz C. Enhancement of skeletal muscle regeneration. Developmental Dynamics 1994; 201: 41-54.
51. Bischoff R. A satellite cell mitogen from crushed adult muscle. Developmental Biology 1986; 115: 140-7.
52. Tidball JG. Mechanisms of muscle injury, repair, and regeneration. Comprehensive Physiology 2011; 1: 2029-62.
53. Zammit PS, Heslop L, Hudon V, et al. Kinetics of myoblast proliferation show that resident satellite cells are competent to fully regenerate skeletal muscle fibers. Experimental Cell Research 2002; 281: 39-49.
54. Whalen RG, Harris JB, Butler-Browne GS, et al. Expression of myosin isoforms during notexin-induced regeneration of rat soleus muscles. Developmental Biology 1990; 141: 24-40.
55. Schultz E. Satellite cell proliferative compartments in growing skeletal muscles. Developmental Biology 1996; 175: 84-94.
56. Rantanen J, Hurme T, Lukka R, et al. Satellite cell proliferation and the expression of myogenin and desmin in regenerating skeletal muscle: Evidence for two different populations of satellite cells. Laboratory Investigation 1995; 72: 341-47.

B r i t a N g u m C h e P h D T h e s i s
70
57. Grounds MD, White JD, Rosenthal N, Bogoyevitch MA. The role of stem cells in skeletal and cardiac muscle repair. Journal of Histochemistry and Cytochemistry 2002; 50: 589-610.
58. Zammit PS, Golding JP, Nagata Y, et al. Muscle satellite cells adopt divergent fates: A mechanism for self-renewal? Journal of Cell Biology 2004; 166: 347-57.
59. Oksbjerg N, Gondret F, Vestergaard M. Basic principles of muscle development and growth in meat-producing mammals as affected by the insulin-like growth factor (IGF) system. Domestic Animal Endocrinology 2004; 27: 219-40.
60. Rabinovsky ED. The multifunctional role of IGF-1 in peripheral nerve regeneration. Neurological Research 2004; 26: 204-10.
61. Zammit PS, Relaix F, Nagata Y, et al. Pax7 and myogenic progression in skeletal muscle satellite cells. Journal of Cell Science 2006; 119: 1824-32.
62. Sabourin LA, Girgis-Gabardo A, Scale P, et al. Reduced differentiation potential of primary MyoD-/- myogenic cells derived from adult skeletal muscle. Journal of Cell Biology 1999; 144: 631-43.
63. Sabourin LA, Rudnicki MA. The molecular regulation of myogenesis. Clinical Genetics 2000; 57: 16-25.
64. Knight JDR, Kothary R. The myogenic kinome: Protein kinases critical to mammalian skeletal myogenesis. Skeletal Muscle 2011; 1: 29
65. Joulia-Ekaza D,Cabello G. Myostatin regulation of muscle development: Molecular basis, natural mutations, physiopathological aspects. Experimental Cell Research 2006; 312: 2401-14.
66. Jin EJ, Kim I, Lee CY, et al. Suppressed cell proliferation and differentiation following an over-expression of myostatin is associated with inhibited expression of insulin-like growth factor II and myogenin in rat L6 myoblasts. Asian-Australian Journal of Animal Sciences 2006; 19: 1508-13.
67. Rabinovsky ED, Gelir E, Gelir S, et al. Targeted expression of IGF-1 transgene to skeletal muscle accelerates muscle and motor neuron regeneration. Federation of American Societies for experimental Biology 2003; 17: 53-55.
68. Theil PK, Sorensen IL, Nissen PM, et al. Temporal expression of growth factor genes of primary porcine satellite cells during myogenesis. Animal Science Journal 2006; 77: 330-37.
69. Zammit PS, Carvajal JJ, Golding JP, et al. Myf5 expression in satellite cells and spindles in adult muscle is controlled by separate genetic elements. Developmental Biology 2004; 273: 454-65.

B r i t a N g u m C h e P h D T h e s i s
71
70. Nedachi T, Kanzaki M. Regulation of glucose transporters by insulin and extracellular glucose in C2C12 myotubes. American Journal of Physiology-Endocrinology and Metabolism 2006; 291: E817-28.
71. Slot JW, Geuze HJ, Gigengack S, James DE, Lienhard GE. Translocation of the glucose transporter GLUT4 in cardiac myocytes of the rat. Proceedings of the National Academy of Sciences, USA 1991; 88: 7815-19.
72. Furtado LM, Somwar R, Sweeney G, et al. Activation of the glucose transporter GLUT4 by insulin. Biochemistry and Cell Biology 2002; 80: 569-78.
73. White MF. The IRS-signalling system: A network of docking proteins that mediate insulin action. Molecular and Cellular Biochemistry 1998; 182: 3-11.
74. Yeh JI, Gulve EA, Rameh L, Birnbaum MJ. The effects of wortmannin on rat skeletal muscle. Dissociation of signaling pathways for insulin- and contraction-activated hexose transport. J Biol Chem 1995; 270: 2107-11.
75. Alessi DR, Cohen P. Mechanism of activation and function of protein kinase B. Current Opinion in Genetics and Development 1998; 8: 55-62.
76. Berwick DC, Dell GC, Welsh GI, et al. Protein kinase B phosphorylation of PIKfyve regulates the trafficking of GLUT4 vesicles. Journal of Cell Science 2004; 117: 5985-93.
77. Jensen J, Rustad PI, Kolnes AJ, et al. The role of skeletal muscle glycogen breakdown for regulation of insulin sensitivity by exercise. Frontiers in physiology 2011;2.
78. Bouskila M, Hirshman MF, Jensen J, et al. Insulin promotes glycogen synthesis in the absence of GSK3 phosphorylation in skeletal muscle. American Journal of Physiology - Endocrinology and Metabolism 2008; 294: E28-35.
79. Ren JM, Marshall BA, Gulve EA, et al. Evidence from transgenic mice that glucose transport is rate-limiting for glycogen deposition and glycolysis in skeletal muscle. Journal of Biological Chemistry 1993; 268: 16113-15.
80. Mueckler M. Family of glucose-transporter genes. Implications for glucose homeostasis and diabetes. Diabetes 1990; 39: 6-11.
81. Watson RT, Pessin JE. Intracellular organization of insulin signaling and GLUT4 translocation. Recent Progress in Hormone Research 2001; 56: 175-93.
82. Yeh TYJ, Sbodio JI, Tsun ZY, et al. Insulin-stimulated exocytosis of GLUT4 is enhanced by IRAP and its partner tankyrase. Biochemical Journal 2007; 402: 279-90.
83. Keller SR. Role of the insulin-regulated aminopeptidase IRAP in insulin action and diabetes. Biological and Pharmaceutical Bulletin 2004; 27: 761-64.

B r i t a N g u m C h e P h D T h e s i s
72
84. Azevedo J, Carey JO, Pories WJ, Morris PG, Dohm GL. Hypoxia stimulates glucose transport in insulin-resistant human skeletal muscle. Diabetes 1995; 44: 695-8.
85. Nesher R, Karl IE, Kipnis DM. Dissociation of effects of insulin and contraction on glucose transport in rat epitrochlearis muscle. The American journal of physiology 1985; 249: C226-32.
86. Zhang JZ, Behrooz A, Ismail-Beigi F. Regulation of glucose transport by hypoxia. American Journal of Kidney Diseases 1999; 34: 189-202.
87. Chen C, Pore N, Behrooz A, Ismail-Beigi F, Maity A. Regulation of glut1 mRNA by hypoxia-inducible factor-1: Interaction between H-ras and hypoxia. Journal of Biological Chemistry 2001; 276: 9519-25.
88. Meléndez R, Meléndez-Hevia E, Canela EI. The fractal structure of glycogen: A clever solution to optimize cell metabolism. Biophysical Journal 1999; 77: 1327-32.
89. Hermansen L, Hultman E, Saltin B. Muscle glycogen during prolonged severe exercise. Acta Physiologica Scandinavica 1967; 71: 129-39.
90. Skurat AV, Dietrich AD, Zhai L, et al. A novel protein that binds and activates glycogenin, the self-glucosylating initiator of glycogen biosynthesis. Journal of Biological Chemistry 2002; 277: 19331-38.
91. Ylä-Ajos M, Ruusunen M, Puolanne E. The significance of the activity of glycogen debranching enzyme in glycolysis in porcine and bovine muscles. Meat Science 2006; 72: 532-38.
92. Bartlett K, Eaton S. Mitochondrial beta-oxidation. European Journal of Biochemistry 2004; 271: 462-69.
93. Ferdinandusse S, Rusch H, Van Lint AEM, et al. Stereochemistry of the peroxisomal branched-chain fatty acid alpha- and beta-oxidation systems in patients suffering from different peroxisomal disorders. Journal of Lipid Research 2002; 43: 438-44.
94. Glatz JFC, Luiken JJFP, Van Bilsen M, et al. Cellular lipid binding proteins as facilitators and regulators of lipid metabolism. Molecular and Cellular Biochemistry 2002; 239: 3-7.
95. Meunier-Durmort C, Poirier H, Niot I, et al. Up-regulation of the expression of the gene for liver fatty acid-binding protein by long-chain fatty acids. Biochemical Journal 1996; 319: 483-87.
96. Dutta-Roy AK. Cellular uptake of long-chain fatty acids: Role of membrane-associated fatty-acid-binding/transport proteins. Cellular and Molecular Life Sciences 2000; 57: 1360-72.
97. Wanders RJA, Ruiter JPN, Ijlst L, et al. The enzymology of mitochondrial fatty acid beta-oxidation and its application to follow-up analysis of positive neonatal screening results. Journal of Inherited Metabolic Disease 2010; 33: 479-94.

B r i t a N g u m C h e P h D T h e s i s
73
98. Bremer J. Carnitine-Metabolism and functions. Physiological Reviews 1983; 63: 1420-80.
99. Plessas ST, Plessas CT. L-Carnitine and its role in medicine: A current consideration of its pharmacokinetics, its role in fatty acid metabolism and its use in ischaemic cardiac disease and primary and secondary L-carnitine deficiencies. Epitheorese Klinikes Farmakologias kai Farmakokinetikes 1996; 14: 11-64.
100. McGarry JD, Brown NF. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. European Journal of Biochemistry 1997; 244: 1-14.
101. Kerner J, Hoppel C. Fatty acid import into mitochondria. Biochemica et Biophysica Acta-Molecular and Cell Biology of Lipids 2000; 1486: 1-17.
102. Ventura FV, Ijlst L, Ruiter J, et al. Carnitine palmitoyltransferase II specificity towards bata-oxidation intermediates: Evidence for a reverse carnitine cycle in mitochondria. European Journal of Biochemistry 1998; 253: 614-8.
103. Brass EP. Overview of coenzyme A metabolism and its role in cellular toxicity. Chemico-Biological Interactions 1994; 90: 203-14.
104. Jakobs BS, Wanders RJA. Fatty acid beta-oxidation in peroxisomes and mitochondria: The first, unequivocal evidence for the involvement of carnitine in shuttling propionyl-CoA from peroxisomes to mitochondria. Biochemical and Biophysical Research Communications 1995; 213: 1035-41.
105. Ramsay RR, Arduini A. The carnitine acyltransferases and their role in modulating acyl-CoA pools. Achives of Biochemistry and Biophysics 1993; 302: 307-14.
106. Kunau WH, Dommes V, Schulz H. Beta-oxidation of fatty acids in mitochondria, peroxisomes, and bacteria: A century of continued progress. Progress in Lipid Research 1995; 34: 267-342.
107. Gregersen N, Andersen BS, Pedersen CB, et al. Mitochondria fatty acid oxidation defects-remaining challenges. Journal of Inherited Metabolic Disease 2008; 31: 643-56.
108. Kim JJP, Gregersen N, Olsen RKJ, Ghisla S. The acyl-CoA dehydrogenases. In: Hille et al., ed. Flavoproteins. Berlin: Walter de Gruyter, 2012 (in press).
109. Hashimoto T, Shindo Y, Souri M, et al. A new inhibitor of mitochondrial fatty acid oxidation. Journal of Biochemistry 1996; 119: 1196-201.
110. Weinberger MJ, Rinaldo P, Strauss AW, et al. Intact alpha-subunit is required for membrane-binding of human mitochondrial trifunctional beta-oxidation protein, but is not necessary for conferring 3-ketoacyl-CoA thiolase activity to the beta-subunit. Biochemical and Biophysical Research Communications 1995; 209: 47-52.

B r i t a N g u m C h e P h D T h e s i s
74
111. Furuta S, Miyazawa S, Osumi T, et al. Properties of mitochondrial and peroxisomal enoyl-CoA hydratases from rat liver. Journal of Biochemistry 1980; 88: 1059-70.
112. Malila LH, Siivari KM, Makela MJ, et al. Enzymes converting D-3-hydroxyacyl-CoA to trans-2-enoyl-CoA. Microsomal and peroxisomal isoenzymes in rat liver. J Biol Chem 1993; 268: 21578-85.
113. Stern JR. Crystalline beta-hydroxybutyryl dehydrogenase from pig heart. Biochimica et Biophysica Acta 1957; 26: 448-49.
114. Uchida Y, Izai K, Orii T, Hashimoto T. Novel fatty acid beta-oxidation enzymes in rat liver mitochondria: II. Purification and properties of enoyl-coenzyme A (CoA) hydratase/3-hydroxyacyl-CoA dehydrogenase/3-ketoacyl-CoA thiolase trifunctional protein. J Biol Chem 1992; 267: 1034-41.
115. Izai K, Uchida Y, Orii T, et al. Novel fatty acid beta-oxidation enzymes in rat liver mitochondria: I. Purification and properties of very-long-chain acyl-coenzyme A dehydrogenase. Journal of Biological Chemistry 1992; 267: 1027-33.
116. Kamijo T, Indo Y, Souri M, et al. Medium chain 3-ketoacyl-coenzyme A thiolase deficiency: A new disorder of mitochondrial fatty acid beta-oxidation. Pediatric Research 1997; 42: 569-76.
117. Das AM, Illsinger S, Lücke T, et al. Isolated mitochondrial long-chain ketoacyl-CoA thiolase deficiency resulting from mutations in the HADHB gene. Clinical Chemistry 2006; 52: 530-4.
118. Berger J, Moller DE. The mechanisms of action of PPARs. Annual Review of Medicine 2002; 53: 409-35.
119. Guri AJ, Hontecillas R, Bassaganya-Riera J. Peroxisome proliferator-activated receptors: Bridging metabolic syndrome with molecular nutrition. Clinical Nutrition 2006; 25: 871-85.
120. Grimaldi PA. Peroxisome proliferator-activated receptors as sensors of fatty acids and derivatives. Cellular and Molecular Life Sciences 2007; 64: 2459-64.
121. Verma NK, Singh J, Dey CS. PPAR-gamma expression modulates insulin sensitivity in C2C12 skeletal muscle cells. Bristish Journal of Pharmacology 2004; 143: 1006-13.
122. Mascaró C, Acosta E, Ortiz JA, et al. Control of human muscle-type carnitine palmitoyltransferase I gene transcription by peroxisome proliferator-activated receptor. Journal of Biological Chemistry 1998; 273: 8560-63.
123. Ellinghaus P, Wolfrum C, Assmann G, et al. Phytanic acid activates the peroxisome proliferator-activated receptor alpha (PPAR alpha) in sterol carrier protein 2- sterol carrier protein x-deficient mice. Journal of Biological Chemistry 1999; 274: 2766-72.

B r i t a N g u m C h e P h D T h e s i s
75
124. Heim M, Johnson J, Boess F, et al. Phytanic acid, a natural peroxisome proliferator-activated receptor (PPAR) agonist, regulates glucose metabolism in rat primary hepatocytes. The Federation of American Societies for Experimental Biology 2002; 16: 718-20.
125. Zomer AWM, van der Burg B, Jansen GA, Wanders RJA, Poll-The B, Van der Saag PT. Pristanic acid and phytanic acid: naturally occurring ligands for the nuclear receptor peroxisome proliferater-activated receptor alpha. Journal of Lipid Research 2000; 41: 1801-07.
126. Kota BP, Huang THW, Roufogalis BD. An overview on biological mechanisms of PPARs. Pharmacological Research 2005; 51: 85-94.
127. Braissant O, Foufelle F, Scotto C, et al. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-alpha, beta, and -gamma in the adult rat. Endocrinology 1996; 137: 354-66.
128. Brandt JM, Djouadi F, Kelly DP. Fatty acids activate transcription of the muscle carnitine palmitoyltransferase I gene in cardiac myocytes via the peroxisome proliferator-activated receptor -alpha. Journal of Biological Chemistry 1998; 273: 23786-92.
129. Azhar S, Kelley G. PPAR-alpha: Its role in the human metabolic syndrome. Future Lipidology 2007; 2: 31-53.
130. Lenhard JM. PPAR-gamma/RXR as a molecular target for diabetes. Receptors and Channels 2001; 7: 249-58.
131. Panadero MI, González MC, Herrera E, et al. Factors modulating fibrates response: Therapeutic implications and alternative strategies. Endocrine, Metabolic and Immune Disorders - Drug Targets 2009; 9: 219-36.
132. Ip E, Farrell G, Hall P, et al. administration of the potent PPAR-alpha agonist, Wy-14,643, reverses nutritional fibrosis and steatohepatitis in mice. Hepatology 2004; 39: 1286-96.
133. Djouadi F, Aubey F, Schlemmer D, et al. Bezafibrate increases very-long-chain acyl-CoA dehydrogenase protein and mRNA expression in deficient fibroblasts and is a potential therapy for fatty acid oxidation disorders. Human Molecular Genetics 2005; 14: 2695-703.
134. Muoio DM, MacLean PS, Lang DB, et al. Fatty acid homeostasis and induction of lipid regulatory genes in skeletal muscles of peroxisome proliferator-activated receptor (PPAR)-alpha knock-out mice. Evidence for compensatory regulation by PPAR-delta. Journal of Biological Chemistry 2002; 277: 26089-97.
135. Kersten S, Seydoux J, Peters JM, et al. Peroxisome proliferator-activated receptor -alpha mediates the adaptive response to fasting. Journal of Clinical Investigation 1999; 103: 1489-98.

B r i t a N g u m C h e P h D T h e s i s
76
136. Evans RM, Barish GD, Wang YX. PPARs and the complex journey to obesity. Nature Medicine 2004; 10: 355-61.
137. Zuo C, Liang P, Huang X. Role of PPAR-beta in fibroblast response to heat injury. Indian Journal of Biochemistry and Biophysics 2012; 49: 219-27.
138. Bonala S, Lokireddy S, Arigela H, et al. Peroxisome proliferator-activated receptor beta/delta induces myogenesis by modulating myostatin activity. Journal of Biological Chemistry 2012; 287: 12935-51.
139. Kang HJ, Hwang SJ, Yoon JA, et al. Activation of peroxisome proliferatorsactivated receptor delta (PPAR-delta) promotes blastocyst hatching in mice. Molecular Human Reproduction 2011; 17: 653-60.
140. Ooi EMM, Watts GF, Sprecher DL, et al. Mechanism of action of a Peroxisome Proliferator-Activated Receptor (PPAR)-delta agonist on lipoprotein metabolism in dyslipidemic subjects with central obesity. Journal of Clinical Endocrinology and Metabolism 2011; 96: 1568-76.
141. Saraf N, Sharma PK, Mondal SC, et al. Role of PPAR-gamma 2 transcription factor in thiazolidinedione-induced insulin sensitization. Journal of Pharmacy and Pharmacology 2012;64:161-71.
142. Rosen ED, Spiegelman BM. Molecular regulation of adipogenesis., Annual Review of Cell and Developmental Biology 2000; 16: 145-61
143. Schadinger SE, Bucher NLR, Schreiber BM, et al. PPAR-gamma 2 regulates lipogenesis and lipid accumulation in steatotic hepatocytes. American Journal of Physiology - Endocrinology and Metabolism 2005; 288: E1195-1205.
144. Hammarstedt A, Rotter Sopasakis V, Gogg S, Jansson PA, Smith U. Improved insulin sensitivity and adipose tissue dysregulation after short-term treatment with pioglitazone in non-diabetic, insulin-resistant subjects. Diabetologia 2005; 48: 96-104.
145. Cha BS, Ciaraldi TP, Park KS, et al. Impaired fatty acid metabolism in type 2 diabetic skeletal muscle cells is reversed by PPAR-gamma agonists. American Journal of Physiology - Endocrinology and Metabolism 2005; 289: E151-59.
146. McCarty MF. Up-regulation of PPAR-gamma coactivator-1 alpha as a strategy for preventing and reversing insulin resistance and obesity. Medical Hypotheses 2005; 64: 399-407.
147. Dumasia R, Eagle KA, Kline-Rogers E, et al. Role of PPAR- gamma agonist thiazolidinediones in treatment of pre-diabetic and diabetic individuals: A cardiovascular perspective. Current Drug Targets- Cardiovascular and Haematological Disorders 2005; 5: 377-86.
148. Guan HP, Yong L, Jensen MV, et al. A futile metabolic cycle activated in adipocytes by antidiabetic agents. Nature Medicine 2002; 8: 1122-28.

B r i t a N g u m C h e P h D T h e s i s
77
149. Tordjman J, Chauvet G, Quette J, Beale EG, Forest C, Antoine B. Thiazolidinediones block fatty acid release by inducing glyceroneogenesis in fat cells. Journal of Biological Chemistry 2003; 278: 18785-90.
150. Kersten S. Peroxisome proliferator activated receptors and obesity. European Journal of Pharmacology 2002; 440: 223-34.
151. Kersten S, Desvergne B, Wahli W. Roles of PPARs in health and disease. Nature 2000; 405: 421-4.
152. Hevener AL, He W, Barak Y, et al. Muscle-specific PPAR gamma deletion causes insulin resistance. Nature Medicine 2003; 9: 1491-97.
153. Norris AW, Chen L, Fisher SJ, et al. Muscle-specific PPAR gamma-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. Journal of Clinical Investigation 2003; 112: 608-18.
154. Yamauchi T, Kadowaki T. The molecular mechanisms by which PPAR gamma/RXR inhibitors improve insulin resistance. Japanese journal of clinical medicine 2001; 59: 2245-54.
155. Hansen RP. Occurrence of Phytanic Acid in Rumen Bacteria. Nature 1966; 210: 841.
156. Patton S, Benson AA. Phytol metabolism in the bovine. Biochemica et Biophysica Acta-Lipids and Lipid Metabolism 1966; 125: 22-32.
157. Harpaz-Saad S, Azoulay T, Arazi T, et al. Chlorophyllase is a rate-limiting enzyme in chlorophyll catabolism and is posttranslationally regulated. Plant Cell 2007; 19: 1007-22.
158. Van den Brink DM, Van Miert JNI, Dacremont G, et al. Characterization of the final step in the conversion of phytol into phytanic acid. Journal of Biological Chemistry 2005; 280: 26838-44.
159. Gloerich J, van den Brink DM, Ruiter JPN, et al. Metabolism of phytol to phytanic acid in the mouse, and the role of PPAR alpha in its regulation. Journal of Lipid Research 2007; 48: 77-85.
160. Gloerich J, van Vlies N, Jansen GA, et al. A phytol-enriched diet induces changes in fatty acid metabolism in mice both via PPAR alpha-dependent and -independent pathways. Journal Lipid Research 2005; 46: 716-26.
161. Rontani JF, Bonin PC, Volkman JK. Biodegradation of free phytol by bacterial communities isolated from marine sediments under aerobic and denitrifying conditions. Applied Enviromental Microbiology 1999; 65: 5484-92.
162. Hylemon PB, Harder J. Biotransformation of monoterpenes, bile acids, and other isoprenoids in anaerobic ecosystems. FEMS Microbiology Reviews 1998; 22: 475-88.

B r i t a N g u m C h e P h D T h e s i s
78
163. Ackman RG, Hansen RP. Occurrence of diastereomers of phytanic and pristanic acids and their determination by gas-Liquid chromatography. Lipids 1967; 2: 357-62
164. Markus S, Walter V. Distribution of the diastereomer distribution of phytanic acid in food samples. Journal of the American Oil Chemists Society 2011; 88: 341-9.
165. Schröder M, Vetter W. GC/EI-MS determination of the diastereomer distribution of phytanic acid in food samples. Journal of the American Oil Chemists' Society 2011; 88: 341-49.
166. Lough AK. Phytanic Acid Content of Lipids of Bovine Tissues and Milk. Lipids 1977; 12: 115-19.
167. Wanders RJA, Komen J, Ferdinandusse S. Phytanic acid metabolism in health and disease. Biochemica et Biophysica Acta-Molecular and Cellular Biology of Lipids 2011;1811:498-507.
168. Steinberg D, Herndon J, William Uhlendorf B, et al. Refsum's disease: Nature of the enzyme defect. Science 1967; 156: 1740-42.
169. Vetter W, Schröder M. Concentrations of phytanic acid and pristanic acid are higher in organic than in conventional dairy products from the German market. Food Chemistry 2010; 119: 746-52.
170. Tsai SC, Steinberg D, Avigan J, et al. Studies on the stereospecificity of mitochondrial oxidation of phytanic acid and of -hydroxyphytanic acid. Journal of Biological Chemistry 1973; 248: 1091-97.
171. Eldjarn L, Try K, Ackman RG, et al. Different ratios of the IDD and DDD diastereoisomers of phytanic acid in patients with Refsum's disease. Biochemica et Biophysica Acta- Lipids and Lipid Metabolism 1968; 164: 94-100.
172. Van Veldhoven PP. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. Journal of Lipid Research 2010; 51: 2863-95.
173. Jansen GA, Wanders RJA. Alpha-Oxidation. Biochimica et Biophysica Acta - Molecular Cell Research 2006; 1763: 1403-12.
174. Mihalik SJ, Rainville AM, Watkins PA. Phytanic acid alpha-oxidation in rat liver peroxisomes. Production of alpha-hydroxyphytanoyl-CoA and formate is enhanced by dioxygenase cofactors. European Journal of Biochemistry 1995;232:545-51.
175. Verhoeven NM, Schor DSM, Previs SF, et al. Stable isotope studies of phytanic acid alpha-oxidation: in vivo production of formic acid. European Journal of Pediatrics 1997; 156: 83-87.
176. Huang S, Van Veldhoven PP, Vanhoutte F, et al. Alpha-oxidation of 3-methyl-substituted fatty acids in rat liver. Archives of Biochemistry and Biophysics 1992; 296: 214-23.

B r i t a N g u m C h e P h D T h e s i s
79
177. Wanders RJA, Jansen GA, Lloyd MD. Phytanic acid alpha-oxidation, new insights into an old problem: A review. Biochemica et Biophysica Acta-Molecular and Cellular Biology of Lipids 2003; 1631: 119-35.
178. Battaile KP, McBurney M, Van Veldhoven PP, et al. Human long chain, very long chain and medium chain acyl-CoA dehydrogenases are specific for the S-enantiomer of 2-methylpentadecanoyl-CoA. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism 1998; 1390: 333-38.
179. Verhoeven NM, Roe DS, Kok RM, et al. Phytanic acid and pristanic acid are oxidized by sequential peroxisomal and mitochondrial reactions in cultured fibroblasts. Journal of Lipid Research 1998; 39: 66-74.
180. Hutton D, Steinber D. Identification of propionate as a degradation product of phytanic acid oxidation in rat and human tissues. Journal of Biological Chemistry 1973; 248: 6871-5.
181. Komen JC, Duran M, Wanders RJA. Omega-hydroxylation of phytanic acid in rat liver microsomes: Implications for Refsum disease. Journal of Lipid Research 2004; 45: 1341-46.
182. Wanders RJA, Komen J, Kemp S. Fatty acid omega-oxidation as a rescue pathway for fatty acid oxidation disorders in humans. Federation of European Biochemical Societies 2011; 278: 182-94.
183. Larsen MK, Nielsen JH, Butler G, et al. Milk quality as affected by feeding regimens in a country with climatic variation. Journal of Dairy Sciences 2010; 93: 2863-73.
184. Doreau M, Bauchart D, Chilliard Y. Enhancing fatty acid composition of milk and meat through animal feeding. Animal Production Sciences 2011; 51: 19-29.
185. Slots T, Butler G, Leifert C, et al. Potentials to differentiate milk composition by different feeding strategies. Journal of Dairy Sci ence 2009; 92: 2057-66.
186. Lee MRF, Connelly PL, Tweed JKS, et al. Effects of high-sugar ryegrass silage and mixtures with red clover silage on ruminant digestion. 2. Lipids. Journal of Animal Sciences 2006; 84: 3061-70.
187. Schröder M, Lutz NL, Tangwan EC, et al. Phytanic acid concentrations and diastereomer ratios in milk fat during changes in the cow's feed from concentrate to hay and back. European Food Research and Technology 2012; 234: 955-62.
188. Schröder M, Yousefi F, Vetter W. Investigating the day-to-day variations of potential marker fatty acids for organic milk in milk from conventionally and organically raised cows. European Food Research and Technology 2011; 232: 167-74.
189. DeFronzo RA, Tripathy D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2009;32 Suppl 2: 157-63.

B r i t a N g u m C h e P h D T h e s i s
80
190. Carey GB. The swine as a model for studying exercise-induced changes in lipid metabolism. Medicine and Sciences in Sports and Exercise 1997; 29: 1437-43.
191. Mau M, Oksbjerg N, Rehfeldt C. Establishment and conditions for growth and
differentiation of a myoblast cell line derived from the semimembranosus muscle of
newborn piglets. In Vitro Cellular and Developmental Biology- Animal 2008; 44: 1-5.
192. Al-Khalili L, Chibalin AV, Kannisto K, et al. Insulin action in cultured human skeletal muscle cells during differentiation: assessment of cell surface GLUT4 and GLUT1 content. Cellular and Molecular Life Sciences 2003; 60: 991-8.
193. Zhao FQ, Keating AF. Functional properties and genomics of glucose transporters. Current Genomics 2007; 8: 113-28.
194. Wu X, Li W, Sharma V, Godzik A, et al. Cloning and characterization of glucose transporter 11, a novel sugar transporter that is alternatively spliced in various tissues. Molecular Genetics and Metabolism 2002; 76: 37-45.
195. Rogers S, Macheda ML, Docherty SE, et al. Identification of a novel glucose transporter-like protein - GLUT-12. American Journal of Physiology - Endocrinology and Metabolism 2002; 282: E733-38.
196. Sarabia V, Lam L, Burdett E, et al. Glucose-Transport in Human Skeletal-Muscle Cells in Culture - Stimulation by Insulin and Metformin. Journal of Clinical Investigation 1992; 90: 1386-95.
197. Wang Z, Yi J, Li H, et al. Extension of life-span of normal human fibroblasts by reconstitution of telomerase activity. Journal of Experimental Biology 2000; 33: 129-40.
198. Gaster M, Poulsen P, Handberg A, et al. Direct evidence of fiber type-dependent GLUT-4 expression in human skeletal muscle. American Journal of Physiology- Endocrinology and Metabolism 2000; 278: E910-16.
199. Ingelsson E, Arnlöv J, Zethelius B, et al. Associations of serum adiponectin with skeletal muscle morphology and insulin sensitivity. Journal of Clinical Endocrinology and Metabolism 2009; 94: 953-57.
200. DeFronzo RA. Pathogenesis of type 2 diabetes: Metabolic and molecular implications for identifying diabetes genes. Diabetes Reviews 1997; 5: 177-269.
201. Henckel PR, Theil PK, Sørensen IL, et al. Temporal changes in glycogenolytic enzyme mRNAs during myogenesis of primary porcine satellite cells. Meat Science 2007; 75: 248-55.
202. Yang L, Chi G, Zhang J, et al. Skeletal muscle models of insulin resistance induced by L6 cell line. Journal of Clinical Rehabilitative Tissue Engineering Research 2009; 13: 248-51.

B r i t a N g u m C h e P h D T h e s i s
81
203. Mamedova LK, Shneyvays V, Katz A, et al. Mechanism of glycogen supercompensation in rat skeletal muscle cultures. Molecular and Cellular Biochemistry 2003; 250: 11-9.
204. Werner LB, Hellgren LI, Raff M, et al. Effect of dairy fat on plasma phytanic acid in healthy volunteers - A randomized controlled study. Lipid in Health and Dis ease 2011; 10.
205. Gloerich J, van den Brink DM, Ruiter JPN, et al. Metabolism of phytol to phytanic acid in the mouse, and the role of PPAR alpha in its regulation. Journal of Lipid Research 2007; 48: 77-85.
206. Ciaraldi TP, Cha BS, Park KS, et al. Free fatty acid metabolism in human skeletal muscle is regulated by PPAR gamma and RXR agonists. Annals of the New York Academy of Sciences 2002; 967: 66-70.
207. Wolfrum C, Ellinghaus P, Fobker M, et al. Phytanic acid is ligand and transcriptional activator of murine liver fatty acid binding protein. Journal of Lipid Research 1999; 40: 708-14.
208. Hellgren LI. Phytanic acid-an overlooked bioactive fatty acid in dairy fat? Annals of the New York Academy of Sciences 2010; 1190: 42-49.
209. Coll T, Eyre E, Rodr¡guez-Calvo R, et al. Oleate reverses palmitate-induced insulin resistance and inflammation in skeletal muscle cells. Journal of Biological Chemistry 2008; 283: 11107-16.
210. Gloerich J, van Vlies N, Jansen GA, et al. A phytol-enriched diet induces changes in fatty acid metabolism in mice both via PPAR alpha-dependent and -independent pathways. Journal of Lipid Research 2005; 46: 716-26.
211. Le Borgne F, Ben Mohamed A, Logerot M, Garnier E, Demarquoy J. Changes in carnitine octanoyltransferase activity induce alteration in fatty acid metabolism. Biochemical and Biophysical Research Communications 2011;409:699-704.
212. Cabrero A, Alegret M, Sanchez RM, et al. Bezafibrate Reduces mRNA Levels of Adipocyte Markers and Increases Fatty Acid Oxidation in Primary Culture of Adipocytes. Diabetes 2001; 50: 1883-90.
213. Al-Dirbashi OY, Santa T, Rashed MS, et al. Rapid UPLC-MS/MS method for routine analysis of plasma pristanic, phytanic, and very long chain fatty acid markers of peroxisomal disorders. Journal of Lipid Research 2008; 49: 1855-62.
214. Allen NE, Grace PB, Ginn A, et al. Phytanic acid: measurement of plasma concentrations by gas-liquid chromatography-mass spectrometry analysis and associations with diet and other plasma fatty acids. British Journal of Nutrition 2008; 99: 653-9.
215. Rontani JF, Rabourdin A, Marchand D, et al. Photochemical oxidation and autoxidation of chlorophyll phytyl side chain in senescent phytoplanktonic cells:

B r i t a N g u m C h e P h D T h e s i s
82
Potential sources of several acyclic isoprenoid compounds in the marine environment. Lipids 2003; 38: 241-54.
216. Davidson J. A comperative study of the pigments in microbial fractions from the sheeps rumen and in the corresponding diet. Journal of the Science of Food and Agriculture 1954; 5: 86-92.
217. Leiber F, Kreuzer M, Nigg D, Wettstein HR, Scheeder MRL. A study on the causes for the elevated n-3 fatty acids in cows' milk of alpine origin. Lipids 2005; 40: 191-202.
218. Howard T, Catherine MS. Crops that stay green. Annals of Applied Biology 1993; 123: 193-219.
219. Lee MRF, Winters AL, Scollan ND, et al. Plant-mediated lipolysis and proteolysis in red clover with different polyphenol oxidase activities. Journal of the Science of Food and Agriculture 2004; 84: 1639-45.
220. Rajab MS, Cantrell CL, Franzblau SG, et al. Antimycobacterial activity of (E)-phytol and derivatives: A preliminary structure-activity study. Planta Medica 1998; 64: 2-4.

B r i t a N g u m C h e P h D T h e s i s
83
6. APPENDIX
6.1 Manuscript I; MI

B r i t a N g u m C h e P h D T h e s i s
84
Phytanic acid stimulates glucose uptake in a model of skeletal
muscles, the primary porcine myotubes
Brita N. Che1, Niels Oksbjerg1, Lars I. Hellgren2, Jacob H. Nielsen3, Jette F. Young1*
1: Department of Food Science, Aarhus University, Blichers Allé 20, 8830 Tjele, Denmark
2: Department of System Biology, Technical University of Denmark, 2800 Kgs. Lyngby,
Denmark
3: Arla Foods, Sønderhøj 14, DK-8260 Viby J, Denmark
*Corresponding author: Jette Feveile Young
Tel: +45 87 15 80 51
Fax: +45 87 15 48 91

B r i t a N g u m C h e P h D T h e s i s
85
Abstract 1
Objectives: Phytanic acid (PA) is a chlorophyll metabolite with potentials in regulating 2
glucose metabolism, as it is a natural ligand of the peroxisome proliferator-activated 3
receptor (PPAR) that is known to regulate hepatic glucose homeostasis. This study aimed 4
to establish primary porcine myotubes as a model for measuring glucose uptake and 5
glycogen synthesis, and to examine the impact of physiological amounts of PA on glucose 6
uptake and glycogen synthesis either alone or in combination with insulin. 7
Methods: Porcine satellite cells were cultured into differentiated myotubes and tritiated 8
2-deoxyglucose (2-DOG) was used to measure glucose uptake, in relation to PA and 2-DOG 9
exposure times and also in relation to PA and insulin. The MIXED procedure model of SAS 10
was used for statistical analysis of data. 11
Results: PA increased glucose uptake by approximately 35%, and the presence of insulin 12
further increased the uptake, but this further increase in uptake was non- additive and less 13
pronounced at high insulin concentrations. There was no effect of PA alone on glycogen 14
synthesis, while the insulin stimulation of glycogen was increased by 20% in the presence 15
of PA. PA neither stimulated glucose uptake nor glycogen synthesis in insulin-resistant 16
myotubes generated by excess glucose exposure. 17
Conclusions: Primary porcine myotubes were established as a model of skeletal muscles 18
for measuring glucose uptake and glycogen synthesis, and we showed that PA can play a 19
role in stimulating glucose uptake at no or inadequate insulin concentrations. 20
Keywords: Phytanic acid, palmitic acid, insulin, glucose uptake, glycogen synthesis, 21
viability, primary porcine myotubes, excess glucose, free fatty acids 22
23

B r i t a N g u m C h e P h D T h e s i s
86
Background 24
Due to its high concentration of saturated fatty acids, dairy fat intake has been associated 25
with increased risk of cardiovascular diseases and type 2 diabetes in humans [1,2]. 26
However, recent meta-analysis of data from prospective cohort-studies shows an inverse 27
association between intake of dairy products and both cardiovascular diseases and 28
incidence of type 2 diabetes [3], and available data do not indicate any increased risk of 29
high-fat dairy products compared to low-fat dairy products [4]. Similarly, studies using the 30
odd-chained fatty acids C15:0 and C17:0 as validated biomarkers of dairy fat intake showed 31
that a higher intake of dairy fat was associated with a decreased risk of myocardial-32
infarction and development of the metabolic syndrome [5-8]. These findings suggest the 33
existence of a milk-fat paradox; despite the saturated profile of milk-fat, it does not seem 34
to increase metabolic risks, but might actually be protective. Hence, further research is 35
necessary to elucidate this controversy and to identify potential components in dairy fat 36
that might be responsible for the protective effect. 37
Milk is a very rich source of potentially bioactive lipids such as short-chained fatty acids 38
and conjugated linoleic acid (CLA) [9,10]. Another fatty acid of interest in milk is phytanic 39
acid (PA); a C20 saturated fatty acid with four methyl-branches, which is known to be a 40
natural ligand of the nuclear receptors peroxisome proliferator- activated receptors 41
(PPAR) and retinoid-X- receptors (RXR) [11-13]. The PPARs are lipid sensors, which act by 42
regulating body glucose and lipid homeostasis [14], and it has been shown that PA 43
enhances glucose uptake in rat hepatocytes, possibly in a PPAR-dependent manner, at 44
least at high concentrations [15]. Based on these observations, it was suggested that PA 45
might improve glucose- homeostasis, and thereby protect against the development of the 46
metabolic syndrome [16]. Dairy products or meat from ruminants are the major sources of 47

B r i t a N g u m C h e P h D T h e s i s
87
PA in the human diet [17]. The concentration of PA in plasma from healthy humans vary 48
between 0.04 and 11.5 µM [18], and is strongly dependent on the intake of ruminant 49
products [19]. 50
Skeletal muscle fibers are the major site for insulin-regulated glucose uptake, accounting 51
for over 70% of glucose disposal after a meal [20]. Furthermore, excess glucose and free 52
fatty acid exposure lead to insulin resistance in these muscle fibers [21,22]. Hence, effects 53
of PA on skeletal muscle glucose metabolism are expected to have an impact on total 54
glucose homeostasis at the organism level. The main aim of this study was therefore to 55
establish primary porcine myotubes as a model for glucose uptake and glycogen synthesis, 56
and to examine whether physiological concentrations of PA alter glucose uptake and the 57
rate of glycogen synthesis in primary porcine myotube cultures, either alone or in 58
combination with insulin. 59
60
Results 61
Viability assay 62
Primary porcine myotubes were incubated with doses of PA ranging from 1-500 µM (table 63
1). The viability of the myotubes was not affected by 1-10 µM PA. Exposure of the myotubes 64
to 20 µM PA markedly reduced their viability by about 20%. This viability dropped by 36% 65
after exposure to 500 µM PA. For comparative reasons, the effect of palmitic acid (PAM) 66
on viability was also assayed (table 1). Exposure of myotubes to 1 or 10 µM PAM had no 67
effect on viability while 50 µM PAM reduced their viability by about 15% and by 500 µM 68
PAM the viability of the myotubes were reduced to only 54 %. 69

B r i t a N g u m C h e P h D T h e s i s
88
Optimization in relation to glucose uptake assay 70
Glucose uptake in primary porcine myotubes was insulin-dose dependent (fig 2a). At 0.1 71
nM insulin, a significant increase in glucose uptake was recorded and at a 72
supraphysiological concentration of 500 nM insulin, a 70% increase in uptake was 73
obtained. To verify the presence of carrier-mediated glucose uptake in the primary porcine 74
myotube model, different doses of cytochalasin B (cyto B) (0-50 µM) were added to the 75
differentiated myotubes (fig. 2b). At 10 µM cyto B, the glucose uptake in myotubes with 76
and without insulin was reduced to 54 and 40%, respectively, when compared to that of 77
control cells without insulin or cyto B addition. In the presence of 20-50 µM cyto B, the 78
glucose uptake was reduced to approximately 70-90% of controls without cyto B, 79
irrespective of insulin addition. 80
Glucose uptake as a function of PA incubation time and 2-DOG incubation 81
time 82
Based on the results from the viability studies and the concentration of PA in human 83
plasma, glucose uptake assays were performed in myotubes that were incubated with 10 84
µM PA over a period of 24 h to define an effective incubation time with PA (fig. 3a). No 85
significant increase in uptake was observed after 1 h incubation with PA. There was, 86
however, a tendency of elevated glucose uptake between 4-8 h of incubation with PA 87
followed by a temporary marked decrease between 12-16 h of incubation, when compared 88
to 1 h of incubation. 89
To estimate an optimal incubation time for 2-DOG to achieve effective uptake, myotubes 90
were exposed to 2-DOG for 5-60 min with or without 10 µM PA (fig. 3b). There was a 91
general, steady increase in glucose uptake with increasing time of incubation with 2-DOG. 92

B r i t a N g u m C h e P h D T h e s i s
89
The steepest increase in 2-DOG uptake was observed during the first 15 min, while the 93
process was slowest during the last 10 min of incubation. In the presence of PA, glucose 94
uptake tended to be higher at all the different incubation times, but this difference was 95
significant only after 60 min of incubation. 96
Glucose uptake in response to PA and PAM 97
When testing the effect of PA on glucose uptake (table 2), a 23% increase in uptake was 98
noted even at the sub-physiological concentration of 0.5 µM PA. Increasing the PA 99
concentration further did not significantly increase the uptake, although average uptake 100
increased to between 30-35%. The effect of 1-10 µM PA could not be mimicked by same 101
concentrations of PAM (table 2). However, when myotubes were exposed to higher doses 102
of PAM, there were marked changes in glucose uptake. Glucose uptake dropped to 75% 103
when the myotubes were exposed to 100 µM PAM and at 200 µM and 400 µM PAM, the 104
glucose uptake ability of the myotubes was only 50 and 30%, respectively, when compared 105
to controls 106
Glucose uptake and glycogen synthesis with or without PA as a function of 107
insulin 108
The impact of PA on glucose uptake and glycogen synthesis was also studied in relation to 109
various insulin concentrations. Addition of 10 µM PA caused a further increase in glucose 110
uptake at all levels of insulin(fig.4a), but a significant improvement of glucose uptake by 111
addition of PA was only achieved at lower insulin concentrations (0 and 0.1nM). The 112
incorporation of glucose into glycogen increased in response to insulin treatment from 0-113
100 nM, with a significant increase at 10 and 100 nM insulin (fig.4b). Overall, glycogen 114
synthesis was not significantly affected by PA alone, regardless of the increase tendency. 115

B r i t a N g u m C h e P h D T h e s i s
90
However, in the presence of 10 nM insulin, PA caused an additional increase in glycogen 116
synthesis of about 20%. 117
Exposure of myotubes to extracellular glucose 118
The viability of the myotubes after exposure to excess glucose (12mM) was not significantly 119
affected (fig. 5a). However, addition of 7, 10 and 15 mM of extracellular glucose reduced 120
glucose uptake to about 80, 65 and 23%, respectively, when compared to myotubes 121
exposed to 4 mM of extracellular glucose (fig. 5b). Higher amounts of glucose did not cause 122
further drops in glucose uptake. While 10 µM PA and10 nM insulin caused an approximate 123
40 and 75% increase, respectively, in glucose uptake in myotubes exposed to 6 mM 124
extracellular glucose, no effect of neither PA nor insulin was observed when myotubes 125
were exposed to 14 mM extracellular glucose (fig. 5c). Glycogen synthesis was reduced to 126
25% when myotubes were treated with 14 mM glucose, compared to myotubes treated with 127
6 mM glucose (fig. 5d). The marked effect of 10 nM insulin on glycogen synthesis in the 128
presence of 6 mM glucose was abolished by exposing the myotubes to 14 mM glucose (fig. 129
5d). 130
131
Discussion 132
Phytanic acid (PA) is a natural ligand and activator of the PPARs, specifically PPAR-α and 133
γ [23]. The PPARs have been implicated not only in the regulation of adipose tissue 134
development and insulin signaling [24] but also in modulating insulin sensitivity of 135
muscles [25,26] and increasing exercise endurance [27]. It has been hypothesized that PA 136
can function as a natural PPAR agonist and be useful in the prevention and treatment of 137
diabetes [28]. Hence, PA may have a positive influence on the regulation of glucose 138

B r i t a N g u m C h e P h D T h e s i s
91
metabolism. As skeletal muscle is the major site of insulin-dependent glucose uptake [20], 139
we established a primary myotube model to study the effect of PA on glucose-homeostasis. 140
The porcine primary myotube model shares many similarities to the human myocytes and 141
has fewer artifacts when compared to immortal cell line systems [29]. Satellite cells were 142
grown (fig.1 a and c) and successfully differentiated into characteristic multinucleated 143
myotubes (fig.1 b and d), just as in previous findings [30]. 144
Initially, the suitability of the primary porcine myotube model was tested, and assay 145
conditions optimized in relation to both glucose (2-DOG) and PA exposure. Viability 146
assays were performed to determine an appropriate concentration of PA that did not 147
compromise the cells metabolic activities. The viability of the myotubes was noticeably 148
reduced when exposed to concentrations of PA equal to or above 20 µM (table 1). The toxic 149
effect of PA at 20 µM or above may be attributed to apoptosis of the cells, as shown 150
previously by studies in human skin fibroblasts and rat liver [31,32]. The average 151
concentration of PA in human plasma has been reported to be between 0.04-11.5 µM 152
[18,19,33], with higher values found predominantly in meat eaters and dairy product 153
consumers [18,19]. Based on this, and the results from our viability studies, the working 154
concentration of PA was chosen as 10 µM. Palmitic acid (PAM); a fatty acid of same chain 155
length as PA but without the branching methyl-groups was used as a control, and we 156
showed that 1 and 10 µM PAM had no effect on myotube viability. At concentrations of 100 157
µM PAM and above, the viability of the myotubes were reduced, possibly in a manner 158
similar to that of higher amounts of PA [31,32]. 159
The non-metabolizable analogue of glucose; 2-deoxyglucose (2-DOG), is a suitable marker 160
for the measurement of muscle glucose transport [34], and it was used in this study to 161
measure glucose uptake. Glucose transport in the muscles is insulin-dependent and 162

B r i t a N g u m C h e P h D T h e s i s
92
mediated by GLUT-4 transporters, as well as insulin-independent, mediated by GLUT-1 163
[35]. Insulin treatment of varying concentrations caused a dose-dependent increase in 164
glucose uptake in the differentiated myotubes cultures (fig. 2a), proving that the primary 165
porcine myotubes have a functional insulin-stimulated GLUT-4 translocation to the 166
plasma membrane [36]. The action of both GLUT-1 and GLUT-4 in the carrier-mediated 167
glucose uptake was checked by treatment of the myotubes with cytochalasin B (cyto B); a 168
drug known to inhibit glucose transport by binding to glucose transporters with a higher 169
affinity than glucose [37,38]. From our observation, cyto B inhibited both the insulin- and 170
non-insulin-mediated glucose uptake to a similar magnitude in primary porcine myotubes 171
(fig. 2b). The reduction in glucose uptake by cyto B confirms that both the insulin- and the 172
non-insulin-mediated transport systems are functional in our primary porcine myotube 173
model. Our observation with cyto B also showed that about 20% of glucose uptake in the 174
myotubes was not affected by cyto B. One can speculate that a minimal steady state of non-175
specific glucose uptake is maintained in the porcine cultures. It is also likely that other 176
GLUT receptors than GLUT 1 and 4 are present in the myotubes [39], and cyto B cannot 177
bind to them. In that case glucose transport is possible even in the presence of cyto B. 178
A biphasic effect on glucose uptake was observed during the 24 h incubation of myotubes 179
with 10 µM PA (fig. 3a). Since only a subtle increase in glucose uptake was noticed after 8 h 180
exposure to PA, it is less likely that the subsequent fall in glucose uptake is due to depletion 181
of GLUT depots. Other unknown factors could, thus, be responsible for this drop in glucose 182
uptake. A rise in glucose uptake at 24 h could be linked to consequent up-regulation of 183
transcripts responsible for the translation of proteins that skew up glucose uptake. 184
Glucose uptake has been shown to be linear for over 20 min in a human skeletal muscle 185
cell line study [40], and in the present study, the steepest increase in glucose uptake was 186

B r i t a N g u m C h e P h D T h e s i s
93
observed during the first 15 min of incubation with 2-DOG and by 45 min, the curve 187
started to level off (fig. 3b). It is possible that after 45 min incubation with 2-DOG, a 188
partial saturation of the normal glucose pathway is attained. Following these observations, 189
the incubation time with 2-DOG was set at 30 min. 190
Considering the effects of PA on glucose uptake, it is noteworthy that a concentration as 191
low as 1 µM caused a 25% increase in glucose uptake. Increasing the PA concentration to 192
10 µM slightly increased glucose uptake further (table 2), while equivalent amounts of 193
PAM had no effect on glucose uptake. The effect of PA was, thus, a specific effect, which is 194
in agreement with a similar study in rat hepatocytes, in which 100 µM PA markedly 195
increased glucose uptake following incubation with 2-DOG, when compared to 196
docosahexaenoic acid [15]. The reduction in glucose uptake observed at PAM 197
concentration of over 100 µM (table 2) was partly due to inhibition of glucose transport 198
activity [41], but also most likely a consequence of stress [31]. 199
The effect of PA diminished with increasing insulin stimulation of glucose uptake (fig. 4a). 200
This could indicate a similar mode of action between insulin and PA. Our results also 201
indicate that the PA concentrations normally found in plasma (> 1 µM), are sufficient to 202
enhance non-insulin dependent glucose uptake and that alteration of plasma PA 203
concentrations within normal physiological range, only have minor effects. Hence, we 204
suggest that normal levels of PA is a relevant factor in controlling fasting glucose 205
homeostasis in vivo and that increased intake of PA only will affect glucose uptake in 206
subjects with extremely low PA-levels (for example strict vegetarians with no intake of 207
dairy products or ruminant meat). 208
Despite the fact that 10 µM PA induced an increased glucose uptake in the absence of 209
insulin (fig 4a), it did not enhance the rate of glycogen synthesis (fig. 4b). Thus, PA at 210

B r i t a N g u m C h e P h D T h e s i s
94
physiological concentrations increases glucose uptake in the absence of insulin, without 211
causing a concomitant increase in glycogen synthesis, why the absorbed glucose must be 212
shunted into other metabolic pathways. However, the combination of 10 µM PA and 10 nM 213
insulin more than doubled the rate of glycogen synthesis, compared to the effect of insulin 214
alone (45% vs. 20%). This creates a somewhat paradoxical situation that increasing insulin 215
concentrations attenuate the effect of PA on glucose uptake, but enhance the effect on 216
glycogen synthesis. Hence, the two effects must be explained by different mechanisms of 217
action of PA. It should be noted, that the effect of PA on both glucose uptake and glycogen 218
synthesis is not a general effect of fatty acids, as the same concentration of PAM did not 219
cause these effects, neither with nor without insulin (data not shown). 220
Previous studies have shown that excess glucose causes glucose toxicity that is manifested 221
by the inhibition of glucose uptake and induction of insulin resistance in the muscles [42]. 222
The toxicity of excess glucose is due to the activation of the hexamine pathway and 223
production of glucosamine, which inhibits insulin-stimulated glucose uptake and 224
subsequently glycogen synthesis [43]. Our observations that increasing amounts of glucose 225
inhibit glucose uptake (fig. 5b and c) and glycogen synthesis (fig. 5d) are in agreement with 226
these findings. Excess glucose did not affect the viability of the myotubes (fig 5a), ruling 227
out the possibility that the fall in glucose uptake was due to decreased viability of the 228
myotubes. These findings stress the role of excess glucose in generating insulin resistance; 229
it is therefore also noteworthy that neither PA nor insulin had any effect on glucose uptake 230
or glycogen synthesis in myotubes that had been exposed to excess glucose. 231
232
Conclusions 233

B r i t a N g u m C h e P h D T h e s i s
95
In the present study, we have been able to generate a suitable primary porcine cell model 234
for studying both insulin and non- insulin-stimulated glucose uptake at physiological 235
levels of insulin, as 0.1 nM insulin induces a significant increase in glucose uptake. We 236
have shown that PA alone can improve glucose uptake but not glycogen synthesis and that 237
PA probably competes with insulin in insulin-signaling. PA can neither stimulate glucose 238
uptake nor glycogen synthesis in insulin-resistant cells generated by exposure to excess 239
glucose. A potential role for PA may, thus, be stimulating glucose uptake in muscle cells at 240
inadequate insulin concentrations, although the consequences of increasing glucose uptake 241
without a concomitant increase in glycogen synthesis need to be studied further. 242
243
Methods 244
Materials and reagents 245
Palmitic acid (PAM), PA, human insulin, dimethyl sulfoxide (DMSO), cytochalasin B (cyto 246
B), essentially fatty acid-free bovine serum albumin (defatted BSA), glycogen, 247
haematoxylin, chloral hydrate, cytosine arabinoside and antibiotics were all purchased 248
from Sigma. Dulbecco's Modified Eagle's Medium (DMEM), fetal bovine serum (FBS), 249
horse serum (HS) and phosphate buffered saline (PBS) were obtained from Life 250
Technology. The WST-1 reagent was obtained from Boehringer Mannheim. The matrigel 251
reagent was from Becton Dickinson, and the BCA kit was from Pierce Rockford. [1-3H] - 2-252
deoxyglucose (2-DOG) was purchased from GE Healthcare and [1-14C]-D-glucose from 253
PerkinElmer. 254
Buffers and media 255

B r i t a N g u m C h e P h D T h e s i s
96
The proliferation growth media (PGM) consisted of DMEM containing 22 mM D-glucose, 256
10% FBS, 10% HS, 1% penicillin-streptomycin, 1.2% amphotericin B and 0.2% gentamycin. 257
The first differentiation media (DM1) was as PGM but without HS and D-glucose was 258
reduced to 6 mM. The second differentiation media (DM2) was as DM1, except for a 259
reduction of FBS to 5% and addition of cytosine arabinoside to stop proliferation. The 260
third differentiation media (DM3) was as DM2 but lacked serum. Hepes-buffered saline 261
(HBS) pH 7.4 consisted of 20mM Hepes, 140 mM NaCl, 5 mM KCl, 2.5 mM MgSO4 and 1 262
mM CaCl2. The storage medium was made up of PGM and 10% DMSO. To haematoxylin 263
solution (5 g of hydrated potassium sulphate, 0.01 g sodium iodate and 0.1 g haematoxylin 264
dye in 100 mL of distilled water) were added 5 g chloral hydrate and 0.1 g acetic acid to 265
improve shelf-life of the solution. 266
The experimental media normally consisted of the stock media (2.5 mM D-glucose and 267
defatted BSA) but where indicated, included different doses of insulin and/or glucose and 268
fatty acids. Fatty acids were solubilized either in DMSO or ethanol. Ethanol was 269
evaporated under a stream of liquid nitrogen while the final amount of DMSO in the 270
experimental media was less than 0.1%. The experimental media with fatty acid was 271
sterilized using a sterile filter and left overnight at room temperature to ensure efficient 272
binding of defatted BSA to free fatty acid. In this study, the fatty acid:BSA ratio was limited 273
to 5:1. The amount of glucose, DMSO and defatted BSA in control samples was equivalent 274
to that of treated samples, unless stated otherwise. 275
Media for glycogen analysis consisted of 5 mM D-glucose, 0.2 µCi [1-14C]-D-glucose and 276
0.1% defatted BSA in HBS. Glycogen carrier stock solution consisted of 0.5 g glycogen in 277
50 ml 30% KOH solution. 278
Isolation and culture of primary porcine satellite cells 279

B r i t a N g u m C h e P h D T h e s i s
97
Primary satellite cells were obtained from the left semi-membranosus muscle of six-week 280
old female pigs, weighing between 12-13.5 kg following a method described by Theil et al 281
[44]. Isolated cells were placed in storage medium and stored in liquid nitrogen until use. 282
Cells were thawed in a 37oC water bath and seeded on 4% matrigel-coated plates 283
containing PGM. After 2 days, cells were washed with 1xPBS, pH 7.4 (with Ca2+, Mg2+) and 284
allowed to proliferate for 3-4 days to 80% confluence (fig. 1 a and c). Cells were given DM1 285
until 100% confluent, and thereafter DM2 was given to stop proliferation and initiate 286
differentiation. At this stage, the cells were denoted to be at their first day of 287
differentiation. Cells were given DM2 until day 2 of differentiation, where fully developed 288
myotubes were observed microscopically (fig. 1 b and d). Myotubes were given DM3 16 h 289
prior to experimental treatment. When the experimental treatment was administered for 290
more than 4 h, DM3 was omitted. Cell culturing was performed at 37°C with 5% CO2 and at 291
100% humidity. 292
Haematoxylin staining 293
Cells were rinsed twice with 1xPBS (without Ca2+, Mg2+) and fixed with ice-cold methanol 294
for 5 min. The methanol was replaced by haematoxylin solution for 10 min, after which the 295
dye was aspirated and cells were rinsed several times to discard of unwanted stain. Cells 296
were viewed for appropriate staining on a microscope at 100 x magnification. 297
Viability test 298
After cell growth, differentiation and incubation with experimental media for 24 h, 299
myotubes were rinsed with 1xPBS (with Ca2+, Mg2+) and WST-1 reagent was used as 300
described by Okura et al [45]. The generation of a formazan dye by cleavage of a 301
tetrazolium salt present in the reagent reflects the amount of active cells present in culture. 302

B r i t a N g u m C h e P h D T h e s i s
98
The absorbance of the culture media, which was corrected for blank readings in the 303
presence of medium only, was measured at A450-630 using the Envision Multilabel Plate 304
Reader model 2104 PerkinElmer. 305
Protein determination 306
The protein concentration of myotube samples was determined using the bicinchroninic 307
acid (BCA) assay as described earlier by Smith et al [46]. 308
2-deoxyglucose uptake 309
Cells seeded in 48-well plates were cultured as described earlier until the 2nd or 3rd day of 310
differentiation. They were rinsed once with 1xPBS, pH 7.4 (with Ca2+, Mg2+) and incubated 311
at given time points with experimental media (see figure legend). Cells were washed 3 x 312
with HBS solution, and glucose uptake was carried out using [1-3H] 2-DOG as described by 313
Klip et al [47]. Unless stated otherwise, cells were incubated with glucose for 30 min. 314
Carrier-dependent glucose uptake was checked using [1-3H]2-DOG solution containing 0-315
50 µM of cyto B [40]. Samples were assayed for [1-3H] 2-DOG-uptake as disintegrations 316
per min and normalized to the basal level of glucose uptake in control samples. 317
Glycogen synthesis 318
Glycogen analysis was carried out on cells grown in 48-well plates as described by Al-319
Khalili et al [36] with some modifications. On day 2 or 3 of differentiation, DM3 media was 320
replaced with experimental media. Cells were rinsed afterwards with HBS solution and 321
then incubated with fresh HBS solution containing 0.1 mM defatted BSA (and insulin 322
where indicated) for 2h. Media for glycogen analysis was added to the cells during the last 323
1.5 h of incubation. Cells were washed 3 times with ice-cold 1xPBS, pH 7.4 (without Ca2+, 324

B r i t a N g u m C h e P h D T h e s i s
99
Mg2+) and lysed with 200 µL 30% KOH per well for at least 30 min. The cell lysate was 325
transferred into eppendorf tubes, heated for 10 min at 96oC and 3 mg/ml of carrier 326
glycogen was added to the cell lysate. A portion of the cell lysate was used for protein 327
determination. Glycogen was precipitated by overnight incubation with 5 volumes of 328
ethanol and 5% sodium sulfate at -20oC. The precipitated glycogen was collected by 329
centrifugation at 20.800 x g for 5 min, and re-dissolved in distilled water. Radioactivity 330
was measured in a scintillation mix as disintegrations per min and the values were 331
corrected to the protein content of the cells. 332
Statistical analysis 333
Statistical analyses were performed using the MIXED procedure in SAS version 9.2 (SAS 334
Institute Inc., Cary, NC, USA). The results are presented as least square mean estimates 335
(LSmeans) and the standard error of mean (SEM). When the main effects were significant, 336
the least square mean estimates were separated by least significant difference (p < 0.05). 337
All data are expressed as fold changes normalized to control samples. 338
339
List of abbreviations 340
2-DOG: 2-deoxyglucose, BCA: bicinchroninic acid, CLA: conjugated linoleic acid, cyto B: 341
cytochalasin B, defatted BSA: fatty acid-free bovine serum albumin, DM1:first 342
differentiation media, DM2: second differentiation media, DM3: third differentiation 343
media, DMEM: Dulbecco's Modified Eagle's Medium, DMSO: dimethyl sulfoxide, FBS: 344
fetal bovine serum, GLUT: glucose transporter, HBS: Hepes-buffered saline, HS: horse 345
serum, LSmeans: least square mean estimates, PA: phytanic acid, PAM: palmitic acid, 346

B r i t a N g u m C h e P h D T h e s i s
100
PBS: phosphate buffered saline, PGM: proliferation growth media, PPAR: peroxisome 347
proliferator-activated receptor, RXR: retinoid-X-receptor, SEM: standard error of mean 348
349
Competing interests 350
The authors declare that they have no competing interests. 351
352
Authors’ contributions 353
B.N.C designed and performed experiments, analyzed data and wrote the manuscript. N.O. 354
contributed to statistical analysis of data, discussion, review and editing of manuscript. 355
L.I.H contributed to the conception of the project, discussion, interpretation, review and 356
editing of the manuscript. J.H.N. contributed to the conception of the project. J.F.Y 357
contributed to the experimental design, discussion, review and editing of manuscript. 358
359
Acknowledgements 360
This study is part of the project; Tailored milk for human health and was supported by the 361
Danish Council for Strategic Research and The Danish Cattle Federation. The authors 362
thank Aase Sorensen for proof reading of the manuscript and Inge-Lise Sørensen and 363
Anne-Grete Dyrvig Pedersen for technical support. 364
365
366

B r i t a N g u m C h e P h D T h e s i s
101
Reference List 367
1. Steyn NP, Mann J, Bennett PH, Temple N, Zimmet P, Tuomilehto J, Lindstrom J, Louheranta 368
A: Diet, nutrition and the prevention of type 2 diabetes. Public Health Nutr 2004, 369
7:147-165. 370
2. Warensjo E, Jansson JH, Berglund L, Boman K, Ahren B, Weinehall L, Lindahl B, Hallmans G, 371
Vessby B: Estimated intake of milk fat is negatively associated with cardiovascular 372
risk factors and does not increase the risk of a first acute myocardial infarction. A 373
prospective case-control study. Brit J Nutr 2004, 91:635-642. 374
3. Elwood PC, Pickering JE, Givens DI, Gallacher JE: The Consumption of Milk and Dairy 375
Foods and the Incidence of Vascular Disease and Diabetes: An Overview of the 376
Evidence. Lipids 2010, 45:925-939. 377
4. Soedamah-Muthu SS, Ding EL, Al-Delaimy WK, Hu BF, Engberik FM, Willett WC, Johanna 378
GM: Milk and dairy consumption and incidence of cardiovascular diseases and all-379
cause mortality: dose-response meta-analysis of prospective cohort studies. Am J 380
Clin Nutr 2011, 93:158-178. 381
5. Biong AS, Rebnord HM, Fimreite RL, Trygg KU, Ringstad J, Thelle DS, Pedersen JI: Intake of 382
dairy fat and dairy products, and risk of myocardial infarction: A case-control 383
study. Int J Food Sci Nutr 2008, 59:155-165. 384
6. Warensjö E, Smedman A, Stegmayr B, Hallmans G, Weinehall L, essby B, Johasson I: Stroke 385
and plasma markers of milk fat intake - a prospective nested case-control study. J 386
Nutr 2009, 8: 387
7. Warensjö E, Jansson J-H, Cederholm T, Boman K, Eliasson M, Hallmans G, Johansson I, 388
Sjögren P: Biomarkers of milk fat and the risk of myocardial infarction in men and 389
women: a prospective, matched case-control study. Am J Clin Nutr 2010, 92:194-202. 390
8. Wolk A, Furuheim M, Vessby B: Fatty acid composition of adipose tissue and serum 391
lipids are valid biological markers of dairy fat intake in men. J Nutr 2001, 131:828-392
833. 393
9. Belury MA: Conjugated Dienoic Linoleate: A Polyunsaturated Fatty Acid with 394
Unique Chemoprotective Properties. Nutr Rev 1995, 53:83-89. 395

B r i t a N g u m C h e P h D T h e s i s
102
10. Parodi PW: Milk fat in human nutrition. Aust J Dairy Technol 2004, 59:3-59. 396
11. Ellinghaus P, Wolfrum C, Assmann G, Spener F, Seedorf U: Phytanic acid activates the 397
peroxisome proliferator-activated receptor alpha (PPAR alpha) in sterol carrier 398
protein 2- sterol carrier protein x-deficient mice. J Biol Chem 1999, 274:2766-2772. 399
12. Kitareewan S, Burka LT, Tomer KB, Parker CE, Deterding LJ, Stevens RD, Forman BM, Mais 400
DE, Heyman RA, McMorris T, Weinberger C: Phytol metabolites are circulating dietary 401
factors that activate the nuclear receptor RXR. Mol Biol Cell 1996, 7:1153-1166. 402
13. Lemotte PK, Keidel S, Apfel CM: Phytanic acid is a retinoid X receptor ligand. Eur J 403
Biochem 1996, 236:328-333. 404
14. Grimaldi PA: Peroxisome proliferator-activated receptors as sensors of fatty acids 405
and derivatives. Cell Mol Life Sci 2007, 64:2459-2464. 406
15. Heim M, Johnson J, Boess F, Bendik I, Weber P, Hunziker W, Fluhmann B: Phytanic acid, a 407
natural peroxisome proliferator-activated receptor agonist, regulates glucose 408
metabolism in rat primary hepatocytes. FASEB J 2002, 16:718-720. 409
16. Hellgren LI: Phytanic acid-an overlooked bioactive fatty acid in dairy fat? Ann Ny 410
Acad Sci 2010, 1190:42-49. 411
17. Brown PJ, Mei G, Gibberd FB, Burston D, Mayne PD, Mcclinchy JE, Sidey M: Diet and 412
Refsums Disease - the Determination of Phytanic Acid and Phytol in Certain 413
Foods and the Application of This Knowledge to the Choice of Suitable 414
Convenience Foods for Patients with Refsums Disease. J Hum Nutr Diet 1993, 6:295-415
305. 416
18. Al-Dirbashi OY, Santa T, Rashed MS, Al-Hassnan Z, Shimozawa N, Chedrawi A, Jacob M, Al-417
Mokhadab M: Rapid UPLC-MS/MS method for routine analysis of plasma pristanic, 418
phytanic, and very long chain fatty acid markers of peroxisomal disorders. J Lipid 419
Res 2008, 49:1855-1862. 420
19. Allen NE, Grace PB, Ginn A, Travis RC, Roddam AW, Appleby PN, Key T: Phytanic acid: 421
measurement of plasma concentrations by gas-liquid chromatography-mass 422
spectrometry analysis and associations with diet and other plasma fatty acids. Brit 423
J Nutr 2008, 99:653-659. 424

B r i t a N g u m C h e P h D T h e s i s
103
20. Nedachi T, Kanzaki M: Regulation of glucose transporters by insulin and 425
extracelluar glucose in C2C12 myotubes. Am J Physiol-Endoc M 2006, 291:E817-E828. 426
21. Cooksey RC, Hebert J, Zhu JH, Wofford P, Garvey WT, McClain DA: Mechanism of 427
hexosamine-induced insulin resistance in transgenic mice overexpressing 428
glutamine: fructose-6-phosphate amidotransferase: Decreased glucose 429
transporter GLUT4 translocation and reversal by treatment with 430
thiazolidinedione. Endocrinology 1999, 140:1151-1157. 431
22. Pulawa LK, Eckel RH: Overexpression of muscle lipoprotein lipase and insulin 432
sensitivity. Curr Opin Clin Nutr Metab Care 2002, 5:569-574. 433
23. Zomer AWM, van der Burg B, Jansen GA, Wanders RJA, Poll-The B, van der Saag PT: 434
Pristanic acid and phytanic acid: naturally occurring ligands for the nuclear 435
receptor peroxisome proliferater-activated receptor alpha. J Lipid Res 2000, 436
41:1801-1807. 437
24. He WM, Barak Y, Hevener A, Olson P, Liao D, Le J, Nelson M, Ong E, Olefsky JM, Evans 438
RM: Adipose-specific peroxisome proliferator-activated receptor gamma knockout 439
causes insulin resistance in fat and liver but not in muscle. P Natl Acad Sci USA 2003, 440
100:15712-15717. 441
25. Verma NK, Singh J, Dey CS: PPAR-gamma expression modulates insulin sensitivity in 442
C2C12 skeletal muscle cells. Brit J Pharmacol 2004, 143:1006-1013. 443
26. Liao W, Nguyen MTA, Yoshizaki T, Favelyukis S, Patsouris D, Imamura T, Verma IM, Olefsky 444
JM: Suppression of PPAR-gamma attenuates insulin-stimulated glucose uptake by 445
affecting both GLUT1 and GLUT4 in 3T3-L1 adipocytes. Am J Physiol-Endoc M 2007, 446
293:E219-E227. 447
27. Wang YX, Zhang CL, Yu RT, Cho HK, Nelson MC, Bayuga-Ocampo CR, Ham J, Kang H, Evans 448
RM: Regulation of muscle fiber type and running endurance by PPAR delta. Plos 449
Biol 2004, 2:1532-1539. 450
28. McCarty MF: The chlorophyll metabolite phytanic acid is a natural rexinoid - 451
Potential for treatment and prevention of diabetes. Med Hypotheses 2001, 56:217-219. 452
29. Carey GB: The swine as a model for studying exercise-induced changes in lipid 453
metabolism. Med Sci Sport Exer 1997, 29:1437-1443. 454

B r i t a N g u m C h e P h D T h e s i s
104
30. Mau M, Oksbjerg N, Rehfeldt C: Establishment and conditions for growth and 455
differentiation of a myoblast cell line derived from the semimembranosus muscle 456
of newborn piglets. In Vitro Cell Dev- An 2008, 44:1-5. 457
31. Schönfeld P, Wojtczak L: Fatty acids as modulators of the cellular production of 458
reactive oxygen species. Free Radical Bio Med 2008, 45:231-241. 459
32. Schönfeld P, Reiser G: Comment concerning the article: 'Phytanic acid impairs 460
mitochondrial respiration through protonophoric action' by Komen et al.: 461
Branched chain phytanic acid inhibits the activity of the mitochondrial 462
respiratory chain. Cell Mol Life Sci 2008, 65:2266-2269. 463
33. Werner LB, Hellgren LI, Raff M, Jensen SK, Petersen RA, Drachmann T, Tholstrup T: Effect 464
of dairy fat on plasma phytanic acid in healthy volunteers - A randomized 465
controlled study. Lipid Health Dis 2011, 10: 466
34. Wickan DR, Morita TN: 2-Deoxyglucose; a metabolic block for glucose. Proc Soc Exp 467
Biol Med 1955, 89:579-582. 468
35. Mueckler M: Family of glucose-transporter genes. Implications for glucose 469
homeostasis and diabetes. Diabetes 1990, 39:6-11. 470
36. Al-Khalili L, Chibalin AV, Kannisto K, Zhang BB, Permert J, Holman GD, Ehrenborg E, Ding 471
VDH, Zierath JR, Krook A: Insulin action in cultured human skeletal muscle cells 472
during differentiation: assessment of cell surface GLUT4 and GLUT1 content. Cell 473
Mol Life Sci 2003, 60:991-998. 474
37. Ebstensen RD, Plagemann PG: Cytochalasin B: inhibition of glucose and glucosamine 475
transport. P Natl Acad Sci USA 1972, 69:1430-1434. 476
38. Chan YJ, Amrit LR: Cytochalasin B binding sites and glucose transport carrier in 477
human erythrocytes ghosts. J Biol Chem 1977, 252:5456-5463. 478
39. Schürmann A: Insight into the "odd" hexose transporters GLUT3, GLUT5, and 479
GLUT7. Am J Physiol-Endoc M 2008, 295:E225-E226 480
40. Sarabia V, Lam L, Burdett E, Leiter LA, Klip A: Glucose-Transport in Human Skeletal-481
Muscle Cells in Culture - Stimulation by Insulin and Metformin. J Clin Invest 1992, 482
90:1386-1395. 483

B r i t a N g u m C h e P h D T h e s i s
105
41. Archuleta TL, Lemieux AM, Saengsirisuwan V, Teachey MK, Lindborg KA, Kim JS, Henriksen 484
EJ: Oxidant stress-induced loss of IRS-1 and IRS-2 proteins in rat skeletal muscle: 485
Role of p38 MAPK. Free Radical Bio Med 2009, 47:1486-1493. 486
42. Kim JK, Zisman A, Fillmore JJ, Peroni OD, Kotani K, Perret P, Zong H, Dong J, Kahn CR, Kahn 487
BB, Shulman GI: Glucose toxicity and the development of diabetes in mice with 488
muscle-specific inactivation of glut4. J Clin Invest 2001, 108:153-160. 489
43. Shepherd PR, Kahn BB: Mechanisms of disease - Glucose transporters and insulin 490
action - Implications for insulin resistance and diabetes mellitus. New Engl J Med 491
1999, 341:248-257. 492
44. Theil PK, Sorensen IL, Nissen PM, Oksbjerg N: Temporal expression of growth factor 493
genes of primary porcine satellite cells during myogenesis. J Anim Sci 2006, 77:330-494
337. 495
45. Okura T, Igase M, Kitami Y, Fukuoka T, Maguchi M, Kohara K, Hiwada K: Platelet-derived 496
growth factor induces apoptosis in vascular smooth muscle cells: roles of the Bcl-497
2 family. BBA-Mol Cell Res 1998, 1403:245-253. 498
46. Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, 499
Goeke NM, Olson BJ, Klenk DC: Measurement of protein using bincinchoninic acid. 500
Anal Biochem 1985, 150:76-85. 501
47. Klip A, Ramlal T: Protein-Kinase-C Is Not Required for Insulin Stimulation of 502
Hexose Uptake in Muscle-Cells in Culture. Biochem J 1987, 242:131-136. 503
504
505
506
507
508
509

B r i t a N g u m C h e P h D T h e s i s
106
Table legends 510
Table 1: Effect of fatty acids on the viability of primary porcine myotubes 511
Fatty acid concentration (µM)
Fold change in viability
PA PAM
Control (0) 1a ± 0.023 1a ± 0.036
1 1.01a ± 0.030
10 0.97a ± 0.022 0.95a ± 0.031
20 0.81b ± 0.036
50 0.74bc ± 0.042 0.85b ± 0.032
100 0.70c ± 0.046 0.82b ± 0.035
200 0.74c ± 0.029
300 0.61d ± 0.051
500 0.64c ± 0.025 0.54d ± 0.030
512
Differentiated porcine myotubes were incubated with different concentrations of fatty 513
acids (PA or PAM) for 24 h, and then treated with WST-1 reagent as mentioned in the 514
methods. Control samples contained experimental media without fatty acids. Data are 515
expressed as LSmean values from five-eight replicate wells, carried out in three separate 516
experiments with cells isolated from a different pig each time. LSmeans with different 517
letters (a-d) within each row denote significantly different responses related to fatty acid 518
treatments; (P < 0.05). 519

B r i t a N g u m C h e P h D T h e s i s
107
520
Table 2: Glucose uptake in response to PA and PAM 521
Fatty acid concentration (µM) Glucose uptake
PA PAM
Control (0) 1.00a ± 0.045 1.00a ± 0.067
1 1.23b ± 0.105 1.09a ±.051
5 1.29b ± 0.071
10 1.32b ± 0.075 1.14a ± 0.061
20 1.34b ± 0.166
40 1.37b ± 0.076
50 0.92a ± 0.032
60 1.37b ± 0.079
100 0.75b ± 0.032
200 0.48c ± 0.04
400 0.32d ± 0.028
522
Glucose uptake was carried out on differentiated myotubes that had been incubated with 1-523
60 µM PA or 1and 10 µM PAM for 4 h. PAM from 50-400 µM was administered for 24 h. 524
Control samples lacked fatty acids. Data are expressed as LSmean values of triplicate wells, 525
carried out in three separate experiments with cells isolated from a different pig each time. 526

B r i t a N g u m C h e P h D T h e s i s
108
LSmeans with different letters (a-d) within each row denote significantly different 527
responses related to fatty acid treatments; (P < 0.05). 528
529
Figure legends 530
Fig. 1: Microscopic view of proliferating and differentiating porcine satellite 531
cells 532
Proliferating myoblasts at 80 % confluence (a) or myotubes at day 2 of differentiation (b) 533
were stained as described in methods to reveal the single nuclei cells of myoblasts (c) and 534
multi-nuclei tubular structure of the myotubes (d). Picture magnification was 100 x. 535
536
Fig. 2: Glucose uptake in porcine myotubes in response to insulin and cyto B 537
Glucose uptake was performed on differentiated porcine myotubes in the presence of 538
varying concentrations of insulin (a), or varying concentrations of cyto B with (open 539
circles) or without (full circles) 500 µM insulin (b). Control samples lacked cyto B. Data 540
are expressed as LSmean values of four replicate wells, carried out in two separate 541
experiments with cells isolated from a different pig each time. LSmeans with different 542
letters (a-f) denote significantly different responses related to the different treatments; (P 543
< 0.05). 544
545
546

B r i t a N g u m C h e P h D T h e s i s
109
Fig. 3: Glucose uptake with or without PA as a function of 2-DOG incubation 547
time 548
Glucose uptake was performed on differentiated myotubes that had been incubated with 549
(a) 10 µM PA for different times and then treated with 2-DOG for 30 min, or (b) with 550
(dashed line) or without (full line) 10 µM PA for 4 h and afterwards treated with 2-DOG for 551
5-60 min. Data are expressed as LSmean values of triplicate wells carried out in three 552
separate experiments with cells isolated from a different pig each time. LSmeans with 553
different letters (a-e) denote significantly different responses related to incubation times, 554
while * denotes LSmeans with significantly different responses with or without PA; (P < 555
0.05). 556
557
Fig. 4: Glucose uptake and glycogen synthesis with or without PA as a function 558
of insulin 559
Glucose uptake (a) was carried out on differentiated myotubes that had been incubated 560
with no (full line) or 10 µM (dashed line) PA for 4 h, followed by different concentrations 561
of insulin during the last 1 h. Glycogen synthesis (b) were performed on myotubes treated 562
with no (full line) or 10 µM PA (dashed line) for 4 h, followed by 0-100 nM insulin for 2 h. 563
Control samples lacked PA and insulin. Data are expressed as LSmean values collected 564
from three-four wells, carried out in three separate experiments with cells isolated from a 565
different pig each time. LSmeans with different letters (a-d) denote significantly different 566
responses related to the insulin concentration, while * denotes LSmeans with significantly 567
different responses with or without PA; (P < 0.05). 568
569

B r i t a N g u m C h e P h D T h e s i s
110
Fig. 5: Exposure of myotubes to extracellular glucose 570
Differentiated myotubes treated with two different concentrations of glucose for 24 h were 571
subjected to viability assay (5a).Glucose uptake was also performed on myotubes exposed 572
to various gluose concentrations (5b). Myotubes exposed to 6mM glucose (controls) or 14 573
mM glucose for 24 h in the absence (open bars) or presence of PA (black bars) or insulin 574
(grey bars) were subjected either to (5c) glucose uptake or (5d) glycogen synthesis 575
measurements. PA was administered for 24 h while insulin was given during the last 1 h of 576
incubation for glucose uptake measurements and 2 h for glycogen synthesis. Data are 577
expressed as LSmean values of four replicate wells, carried out in three separate 578
experiments with cells isolated from a different pig each time. LSmeans with different 579
letters (a and b) denote significantly different responses related to PA or insulin 580
treatments, while statistical difference in LSmeans between controls and myotubes given 581
14 mM glucose is denoted with *; (P < 0.05). 582
583
584
585
586
587
588
589
590

B r i t a N g u m C h e P h D T h e s i s
111
Figures 591
a b 592
c d 593
Fig. 1 594
595

B r i t a N g u m C h e P h D T h e s i s
112
Insulin concentration (nM)
0 0.01 0.1 0.5 1 10 100 500
Fold
change in g
lucose u
pta
ke (
dpm
)
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2
a
ab
bcbc
cdcd
cd
d
596
Fig. 2a 597
cyto B concentration (µM)
control 10 20 30 40 50
Fold
change in g
lucose u
pta
ke (
dpm
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
a
c
b
dde
defdef
efef
fef
f
598
Fig. 2b 599

B r i t a N g u m C h e P h D T h e s i s
113
PA incubation time (h)
1 2 4 8 12 16 24
Fo
ld c
ha
ng
e in
glu
co
se
up
take
(d
pm
)
0.6
0.8
1.0
1.2
1.4
aa
a
a
bb
a
600
Fig. 3a 601
2-DOG incubation time (min)
0 10 20 30 40 50 60 70
Fo
ld c
ha
nge
in
glu
co
se
up
take
(d
pm
)
0
2
4
6
8
10
12
14
e
d
d
d
c
bab
a
a
ab
b
c
*
602
Fig. 3b 603

B r i t a N g u m C h e P h D T h e s i s
114
Insulin concentration(nM)0 0.1 1 10
Fo
ld c
ha
nge
in
glu
co
se
up
take
(dp
m)
1.0
1.2
1.4
1.6
1.8
2.0
bc
c
b
a
b
a
a
a
*
*
604
Fig. 4a 605
b
*
Insulin concentration (nM)
0 1 10 100
Fold
change in g
lycogen s
ynth
esis
(dpm
/mg p
rote
in)
0.9
1.0
1.1
1.2
1.3
1.4
1.5
1.6
1.7
a
a
ab
a
bc
b
cd
b
*
*
606
Fig. 4b 607

B r i t a N g u m C h e P h D T h e s i s
115
Glucose concenteation (mM)5.6 12
Fo
ld c
ha
nge
in v
iab
ility
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
a
a
608
Fig. 5a 609
Glucose concentration (mM)
4 7 10 15 25
Fold
change in g
lucose u
pta
ke (
dpm
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
a
b
c
d
d
610
Fig. 5b 611

B r i t a N g u m C h e P h D T h e s i s
116
Glucose concentration (mM)
6 14
Fo
ld c
hang
e in
glu
cose u
pta
ke (
dp
m)
0.0
0.5
1.0
1.5
2.0
2.5
control
10 µM PA
10 nM Ins
*a
b
b
612
Fig. 5c 613
Glucose concentration (mM)
6 14
Fold
change in g
lycogen s
ynth
esis
(d
pm
/mg p
rote
in)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
control
10 µM PA
10 nM Ins
b
aa
*
614
Fig. 5d 615

B r i t a N g u m C h e P h D T h e s i s
117
6.2 Manuscript II; MII

B r i t a N g u m C h e P h D T h e s i s
118
Effects of phytanic acid on beta-oxidation flux in a primary porcine
myotube model as detected by an UPLC-MS/MS acylcarnitine assay
Brita Ngum Chea, Rune Isak Dupont Birklerc, Mogens Johannsenc, Niels Gregersenb, Lars
I. Hellgrend, Jette Feveile Younga*
a) Department of Food Science, Aarhus University, Blichers Allé 20, 8830 Tjele, Denmark
b) Department of Clinical Medicine, Research Unit for Molecular Medicine, Aarhus University,
Brendstrupgårdsvej 100, 8200 Aarhus N Denmark
c) Department of Forensic Medicine, Aarhus University, Brendstrupgårdsvej 100, 8200 Aarhus N
Denmark
d) Department of System Biology, Technical University of Denmark, 2800 Kgs. Lyngby, Denmark
*Corresponding author: Jette Feveile Young
Tel: +45 87 15 80 51
Fax: +45 87 15 48 91

B r i t a N g u m C h e P h D T h e s i s
119
Abstract 1
Primary porcine myotubes were successfully established as an efficient model for the 2
determination of β- oxidation flux, as a simple underivatized and sensitive UPLC-MS/MS 3
method was utilized. It was revealed that the myotubes could easily take up and oxidize 4
labeled palmitate, since they generated palmitoylcarnitine (C16) and acetylcarnitine (C2). 5
Phytanic acid (PA) is a natural ligand of the peroxisome proliferator-activated receptors 6
(PPAR) and has been suggested to have health-improving properties. If PA can improve 7
FAO just like PPARα ligands do, then it will have a positive implications in relation to e.g. a 8
fight against the metabolic syndrome (MS). PA was shown to improve β-oxidation in the 9
myotubes, as it significantly reduced the content of C16 and increased the content of C2 in 10
a time-dependent manner. However, the impact of PA on the myotubes can only be 11
confirmed if the findings are validated with a known PPARα agonist. 12
Keywords: fatty acid β-oxidation, phytanic acid, oleic acid, acylcarnitine, primary porcine 13
myotubes, viability, palmitic acid 14
15
16
17
18
19
20
21
22
23
24

B r i t a N g u m C h e P h D T h e s i s
120
1. Introduction 25
One of the most important metabolic functions of fatty acids (FAs) is to supply energy to 26
the body; FAs account for up to 90% of the cellular energy requirement during fasting 1. 27
Lipid oxidation and utilization of FAs during fasting are reduced in obese and diabetic 28
patients2;3 and this reduced ability of muscles to oxidize fats has been linked to the 29
accumulation of intramuscular triacylglycerides (TAG) in the muscles 4;5. The rate of FAO 30
is one of the many factors that influence the content of TAG. These observations direct 31
focus towards the search of components and mechanisms that might improve FAO. 32
The peroxisome proliferator-activated receptors (PPARs) are nuclear receptors, their 33
activation of which initiate the transcriptional regulation of genes that improve FAO, 34
insulin signaling and glucose metabolism 6;7. Three forms of PPAR exists (PPAR α, β, and 35
γ), and especially synthetic ligands of PPAR α such as benzofibrates have been 36
documented to improve FAO 8-10. More so, some unsaturated FAs such as oleic acid (OLA), 37
conjugated linoleic acid (CLA) and branched-chain FAs like phytanic acid (PA) have been 38
established as natural ligands of PPARs11-13. OLA and CLA have already been shown to 39
improve FOA or have therapeutic potentials, respectively11;14. 40
PA, which is of ruminant origin and present in dairy products15 has in our lab recently been 41
shown to improve glucose uptake in primary porcine myotubes at physiological 42
concentrations (unpublished data). Phytol-rich diets have been shown to increase 43
peroxisomal and mitochondrial oxidation enzymes in the liver of mice, in both a PPAR α-44
dependent and independent fashion16. To our knowledge, it is unknown whether PA can 45
improve FAO in the muscles, but given that it is a natural PPAR ligand, we hypothesize 46
that it is capable of mediating the metabolism of FAs, by improving β-oxidation in the 47
muscles. 48

B r i t a N g u m C h e P h D T h e s i s
121
In order to undergo FAO, long-chain FAs have to be taken up into the cells and shuttled 49
through the carnitine system17-19. FAs taken up into the cells are converted into acyl-50
coenzyme A (acyl-CoA) by the enzyme acyl-CoA synthetase, which is located on the outer 51
membrane of the mitochondria. By using free carnitine taken up into the cells through 52
carnitine transporters, carnitine palmitoyl transferase (CPT I) catalyzes the esterification 53
of acyl-CoA into acylcarnitines. Acylcarnitines are transported through the inner 54
mitochondria membrane by carnitine-acylcarnitine translocase (CACT), and inside the 55
mitochondria CPT II catalyzes the reformation of acyl-CoA, which is the substrate for β-56
oxidation. The β-oxidation of acyl-CoA to acetyl-CoA requires the action of acyl-57
dehydrogenases and the mitochondria trifunctional proteins (MTP) 19. 58
Assays using labeled palmitate to track changes in FAO in vitro is common 20;21. In line 59
with such assays, in vitro acylcarnitine profiling is performed in cells that are incubated in 60
a medium supplemented with labeled palmitate and excess of free L-carnitine, resulting in 61
an elevation and extracellular accumulation of the acylcarnitines derived from FAO. The 62
analyzed content of the produced acylcarnitines reflects the content of acyl-CoA in the 63
mitochondria. 64
In this study, primary porcine myotubes with similar characteristics to human muscle 65
fibers 22, were established as a model for analyses of β-oxidation flux in the mitochondria, 66
using the acylcarnitine assay. The time- and dose-dependent impact of PA on FAO was 67
tested by using a simple underivatized and sensitive UPLC-MS/MS method that measured 68
palmitoylcarnitine and acetylcarnitine pools in cell media. 69
70
71

B r i t a N g u m C h e P h D T h e s i s
122
72
2. Methods 73
2.1. Materials and reagents 74
Dulbecco's Modified Eagle's Medium (DMEM), fetal bovine serum (FBS), horse serum 75
(HS) and 1x phosphate buffered saline (Ca 2+ and Mg 2+ (PBS)) were obtained from Life 76
Technology. Essentially fatty acid-free bovine serum albumin, Cytosine arabinoside and 77
antibiotics were purchased from Sigma Aldrich, Denmark. PA, OLA and L-Carnitine were 78
also from Sigma Aldrich, Denmark, while [U-13 C]-Potassium palmitate (13C-PAM) was 79
from Cambridge Isotopes. WST-1 reagent, matrigel and BCA kit were bought from 80
Boehringer Mannheim, Becton Dickinson and Pierce Rockford, respectively. HPLC-grade 81
methanol, HPLC-grade acetonitrile, and formic acid 98-100% were purchased from Sigma 82
Aldrich, Denmark. Acetyl- and palmitoyl-L-carnitine.HCl were purchased from 83
Departamento de Química Orgánica, Universidad Autónoma de Madrid, Cantoblanco, 84
Madrid, España. Acylcarnitine Internal standard (IS) solution was purchased from 85
Cambridge Isotope Laboratories, Inc., Andover, MA, USA. The IS stock solution contained 86
L-carnitine-d9, L-acetylcarnitine-d3 and L-palmitoylcarnitine-d3; deuterium labelled 87
acylcarnitine analogues. 88
2.2. Media and buffers for cell culture 89
The proliferation growth media (PGM) was made up of DMEM containing 22mM D-90
glucose, 1% penicillin-stretomyocin, 1.2% Amphotericin B, 0.2% Gentamycin, 10% FBS 91
and 10% HS. The storage medium (SM) contained PGM and 10 % DMSO. The first fusion 92
media (FM1) was as PGM, but lacked HS and D-glucose was reduced to 6mM. The second 93
fusion media (FM2) was as FM1, however, it contained 10 µM Cytosine arabinoside and 94

B r i t a N g u m C h e P h D T h e s i s
123
FBS was only 5%. Hepes buffered saline (HBS) pH 7.4 consisted of 20 mM Hepes, 140 mM 95
NaCl, 5 mM KCl, 2.5 mM MgSO4 and 1 mM CaCl2. 96
2.3. Preparation of experimental media 97
A stock of 10% defatted BSA in PBS, 400 mM L-Carnitine in H2O and DMEM (25 mM 98
glucose) was used to prepare a stock medium containing 4 or 60 µM BSA, 400 µM L-99
carnitine and 5mM D-glucose. Experimental medium I (EM I) consisted of the stock 100
medium (with 60 µM BSA) and varying concentrations of 13C-PAM, while experimental 101
medium II (EM II) consisted of stock medium (with 4 µM BSA) and different 102
concentrations of FAs. When EM I was used in the second experimental set up, the 103
concentration of 13C-PAM was kept constant at 50 µM (see Figure 1b). To prepare the 104
experimental media, 13C-PAM, FAs were dissolved in ethanol and dried under a stream of 105
liquid nitrogen. Stock medium was then added and vigorously mixed for 5 min. The 106
experimental medium was left overnight at room temperature with gentle shaking and was 107
afterwards sterilized using a sterile filter. Experimental medium with lower concentrations 108
of 13C-PAM or PA were made by titration of higher concentrations with the stock media. 109
The minimum ratio of FAs to defatted BSA was 5:1 in EM I, EM II and stock media. 110
2.4. Isolation and culture of primary porcine satellite cells 111
Cell culturing was carried out at 37° C with 5% CO2. Satellite cells obtained from the semi-112
membranous muscle of young pigs 23 were placed in storage medium and stored in liquid 113
nitrogen until use. Cells were cultured as described earlier in Che et al (unpublished data). 114
Briefly, cells were thawed at 37o C and seeded on 4% matrigel coated plates containing 115
PGM. The cells were rinsed with PBS after two days, and allowed to proliferate into 116
myoblast. PGM were given to myoblasts every second day until they were 80 % confluent. 117

B r i t a N g u m C h e P h D T h e s i s
124
Then, FM1 was given to myoblasts until they were 100% confluent. At this point, FM2 was 118
given to the confluent cells to stop proliferation and initiate differentiation. When the cells 119
in FM2 were fully differentiated, as confirmed by microscopic observations, they were 120
rinsed with PBS and ready for experimental treatments. 121
2.5. Exposure of experimental media to myotubes 122
Two experimental setups (Figures 1a and b) were established to study the acylcarnitine 123
profiles in the primary porcine myotubes. On day 3 of differentiation, FM3 was discarded 124
and the myotubes were rinsed and incubated with stock media (white bars). Stock media 125
was replaced with EM I (black bars) for up to 4 days or EM II (grey bars) for up to 2 days 126
followed by a short rinse and then EM I for 1 day. EM I collected from the myotubes was 127
stored at -20 oC until use for acyl carnitine analysis, and the myotubes were used for 128
protein determination using the BCA method 24. 129
2.6. Preparation of collected media for acylcarnitine analysis 130
Cell growth media (50 µL) was mixed with 10 µL internal standard solution. 440 µL cold (-131
18° C) ACN was afterwards added and the solution was subsequently centrifuged at 20.800 132
g for 1 minute to separate the precipitate from the solution. 200 µL of the supernatant was 133
transferred to HPLC vials for further acylcarnitine analysis. 134
2.7. Acylcarnitine analysis 135
Acylcarnitines were analysed without derivatization prior to analysis using a Waters 136
ACQUITY UPLC coupled to a Waters Xevo TQS tandem mass spectrometer. Acylcarnitines 137
were separated using gradient elution on a HILIC (hydrophilic Interaction 138
Chromatography) column (ACQUITY BEH Amide column, 2.1 mm i.d. x 100 mm, particle 139

B r i t a N g u m C h e P h D T h e s i s
125
size 1.7 µm) equipped with an ACQUITY UPLC column inline filter (0.2 µm). 140
Acylcarnitines were analysed in positive ionization mode using the common acylcarnitine 141
fragments m/z 85 and [MH-59]+. The product ion m/z 85 were used as the quantifier ion 142
and the [MH-59]+ ion as qualifier when quantifying and verifying detected peaks of the 143
acylcarnitines respectively. Quantification of acylcarnitines in the cell growth media was 144
calculated using external calibration samples. Linear regression analysis of the peak area 145
ratio analyse/IS (weighed 1/X) versus concentration was used for each of the 146
acylcarnitines. 147
2.8. Viability test 148
Cells grown in 96-well plates were used for the viability test. After incubation of 149
differentiated myotubes with EM I, they were rinsed with PBS and exposed to the WST-1 150
reagent following a method described by Okura et al 25, with a few modifications. The cells 151
were incubated with WST-1 media for 4 h with a short shaking every hour. The action of 152
mitochondria on tetrazolium salts present in the WST-1 reagent generates a formazan dye, 153
the concentration of which reflects the number of active cells available. The absorbance at 154
A450-630 of the WST-1 media was recorded in the absence of cells, immediately after WST-1 155
addition and after 4 h of incubation, using the Envision Multi label Plate Reader model 156
2104 PerkinElmer. 157
158
3. Results 159
3.1. Viability assay 160
The viability of the primary porcine myotubes was affected by both time and dose of 13C-161
PAM (Figure 2). With doses of 13C-PAM ranging from 0-100 µM, there was a general 162

B r i t a N g u m C h e P h D T h e s i s
126
increase in the viability of the myotubes as the time of incubation with 13C-PAM increased 163
(from 1 day - 4 days). Incubation of myotubes with 25 µM or more of 13C-PAM for up to 2 164
days reduced the viability of the myotubes relative to controls without 13C-PAM. At 3 days 165
of incubation, the viability of the cells decreased only when 13C-PAM ≥ 50 µM was used. 166
Cell viability was significantly reduced by 200 and 300 µM 13C-PAM during all incubation 167
periods. Based on these results, subsequent experiments on determining the flux of β-168
oxidation in the myotubes were performed with concentrations of 13C-PAM ranging from 169
0-100 µM. 170
3.2. Acylcarnitine quantification using UPLC-MS/MS 171
Acylcarnitine concentrations in the cell growth media were measured using UPLC-MS/MS. 172
The concentrations were afterwards correlated to the protein content of the cells. An 173
example of 13C-C16 is shown in a chromatogram in Figure 3. Together with the two 174
transitions shown in the chromatogram, the retention time of 0.84 minutes is used for the 175
identification of 13C-palmitoylcarnitine. The first entry in the chromatogram shows the 176
transition 416.34 > 85.05, which is used as quantifier. The transition 416.34 > 357.34 177
(common loss of 59) is used as qualifier. For both C2 and C16 analogues, retention time, 178
quantifier, and qualifier transitions were used for exact identification. 179
3.3. Acylcarnitine profile as affected by incubation period and dose of 180
13C-PAM 181
182
3.3.1. Acetylcarnitine (C2) 183
Generally, the contents of C2 increased with increasing dose of 13C-PAM, regardless of the 184
duration of exposure of the myotubes to 13C-PAM (Figure 4a). At doses of 13C-PAM ≤ 25 185
µM, treatment for 4 days caused the highest increase in the content of C2, while treatments 186

B r i t a N g u m C h e P h D T h e s i s
127
for 1-3 days with 13C-PAM ≤ 10 µM resulted in amounts that were not significantly 187
different from each other. However, incubation for 2 or 3 days, with 25 µM 13C-PAM 188
resulted in similar C2 contents. When 50 µM 13C-PAM was administered to the myotubes 189
for 4 days, the content of C2 was the highest, although not significantly different from that 190
obtained after 3 days of incubation. More so, the level of C2 after 1 day of treatment with 191
50 µM 13C-PAM was lowest, even though it was not significantly different from that 192
obtained after 2 days of treatment. Exposure of the myotubes to 100 µM 13C-PAM for 1 day 193
resulted in a lowest C2-content, even though it was not significantly different from that 194
generated after 4 days of incubation. Nevertheless, treatments with 100 µM 13C-PAM for 195
over 1 day resulted in contents of C2 that were not significantly different from each other 196
(Figure 4a). 197
3.3.2. Palmitoylcarnitine (C16) 198
The exposure of 13C-PAM ≥ 25 µM generated C16-levels, which were bi-symmetrically 199
grouped, with respect to the duration of exposures; incubation for 1 and 2 days generated 200
higher levels, when compared to 3 and 4 days of incubation (Figures 4b). C16 generally 201
increased with increasing dose of 13C-PAM, even though no clear increase was noticed after 202
2 days of incubation with 0- 10 µM 13C-PAM, and unlike with C2, the content of C16 was 203
highest after 1 day of incubation with 13C-PAM, although they were not significantly 204
different from levels generated after 2 days of incubation (Figure 4b). 205
3.3.3. C16/C2 206
The ratio of C16/C2 was calculated to get a clear picture of inhibition or activation of β-207
oxidation during varying exposure of myotubes to different doses of 13C-PAM. Increasing 208
the incubation time of myotubes with 13C-PAM clearly increased the content of C16, as the 209

B r i t a N g u m C h e P h D T h e s i s
128
C16/C2 ratio dropped from 1 - 3 days of incubation (Figure 4c). Incubation for more than 3 210
days did not significantly increase the content of C16 except when a dose of 5 µM 13C-PAM 211
was used. With regards to the effect of 13C-PAM concentration, treatment for just 1 day 212
had no overall impact on the generation of C16, as the C16/C2 content was unchanged 213
when myotubes were exposed to all doses of 13C-PAM (Figure 3c). Treatments for 2 days 214
with U-13 C-PAM tended to increase the content of C16/C2 and by 50 and 100 µM 13C-PAM, 215
the content of C16/C2 was significantly increased. When myotubes were treated for over 2 216
days with 13C-PAM, the content of C16/C2 also tended to increase, but only at 100 µM was 217
the increase significant (Figure 4c). 218
As the content of C16/C2 showed that treatments of myotubes for 1 day with 50 µM 13C-219
PAM did not have any overall effect on the content of C16, this concentration and time was 220
used for further analysis of the effects of treatment of PA on β-oxidation. 221
3.4. PA effects on acylcarnitine contents 222
As shown in Figure 5a, the incubation of myotubes with PA enhanced the production of 223
C2, which was dependent on the amount of PA and the duration of exposure. No effect of 224
PA was noticed during an exposure of 4 h, but by 24 h, PA ≥ 10 µM increased the content 225
of C2 while at 48 h exposure of the myotubes also 5 µM PA caused an increase in the 226
content of C2. For comparison, OLA caused a significant increase in the content of C2 as 227
from 4 h of incubation. 228
The content of C16/C2 dropped after myotubes were exposed to PA ≥ 5 µM or OLA for 24 229
h, and 48 h exposure resulted in a further reduction of C16/C2 (figure 5b). 230
231

B r i t a N g u m C h e P h D T h e s i s
129
4. Discussion 232
4.1 Optimization of myotubes for acylcarnitine analysis 233
Optimization experiments based on viability and acylcarnitine measurements, after 234
exposure of myotubes to different concentrations of 13C-PAM at varying time points, were 235
performed to establish the myotubes as an efficient model for the analysis of β-oxidation. 236
To determine an appropriate concentration range of 13C-PAM and duration of 13C-PAM 237
exposure to the myotubes, viability studies were performed. Doses of 13C-PAM above 10 238
µM and administered for just 1 day significantly reduced the viability of the cells, and 200 239
and 300 µM 13C-PAM reduced the viability of myotubes, regardless of the exposure 240
periods (Figure 2). The reduction in viability by palmate has been observed before, and 241
suggested to be caused by stress-induced apoptosis, or through the accumulation of 242
ceramides, which are intermediates produced by palmitate metabolism 26. The reduction 243
of viability of the myotubes by 200 and 300 µM of 13C-PAM was extreme, and therefore, 244
these concentrations were excluded during further analysis. 245
During longer exposures of myotubes to 0-100 µM 13C-PAM, we observed that the viability 246
of the cells was increased (Figure 2). This could be attributed to the fact that myotubes 247
adapted to 13C-PAM as an extra energy source; FAs are predominant primary sources of 248
energy 1. 249
A concentration range of 0-100 µM 13C-PAM was exposed to the myotubes for up to 4 250
days, and the flux in β- oxidation was followed by analyzing the contents of C2 and C16 251
(discussed later). In the analysis of C2 and C16, the retention times, quantifications and 252
qualifications of their analogues were used in combination, to exactly identify each 253
acetylcarnitine. 254

B r i t a N g u m C h e P h D T h e s i s
130
Following the observations, a concentration of 50 µM 13C-PAM administered for just 1 day 255
was chosen for further analysis of the effects of PA on β- oxidation. The chosen 256
concentration and duration of incubation, regardless of a 10 % drop in viability, was based 257
predominantly on the fact that the content of C16 was unchanged under these conditions 258
(discussed later). More so, long-term incubation with palmitate could affect the general 259
metabolism of the myotubes as palmitate has been implicated in the inhibition of glucose 260
uptake and FAO 27-29. Thus, it was of interest not to expose the myotubes to palmitate for a 261
longer period. Furthermore, muscle cells can undergo senescence when cultured for 262
longer periods and this may also dysregulate their metabolism 30. Actually, myotubes 263
cultured for longer periods are known to exhibit physiological, morphological and 264
mechanical changes31-34. 265
266
267
4.2 Dose and time-dependent generation of acylcarnitines by 13C- PAM 268
The dose of 13C-PAM and duration of incubation with myotubes affected the content of C2, 269
possibly through the extent to which it was generated (Figures 4a). The content of C2 270
increased with increasing dose of 13C-PAM and this indicated that FAO was enhanced with 271
increased 13C-PAM (Figure 4a). More so, incubation of the myotubes for 4 days with 13C-272
PAM < 25 µM, resulted in the highest level of FAO, as the content of C2 was highest. 273
Studies on the effect of palmitate in rat muscles have shown that increasing the 274
concentration of palmitate and the duration of exposure to the muscles, led to an increase 275
in FAO 35. As the same palmitate has also been shown to inhibit FAO, as previously 276

B r i t a N g u m C h e P h D T h e s i s
131
mentioned, it seems apparent that a certain amount of palmitate available in a specific 277
duration is beneficial to the cells. 278
Although in the work of Rasmussen et al 35, the duration of exposure of muscles to 279
palmitate was in the range of minutes, and CO2 was measured as the oxidation product, 280
one can suggest the extrapolation of the physiological changes necessary to cause these 281
effects, e.g. changes in malonyl-CoA35, to also affect the content of C16. Analysis of the 282
content of C16 showed that it was increased during the first two days of incubation with 283
13C-PAM > 10 µM, when compared to the third and fourth day of incubation (Figure 4b), 284
but as observed in the level of C16/C2, the actual content of C16 was markedly reduced 285
when the duration of exposure of 13C-PAM ≤ 50 µM was more than 1 day (Figure 4c). This 286
disparity in the effect of dose and duration could lie in the fact that the contents of C16 287
and C2 generated in the myotubes are controlled possibly by same mechanisms in varying 288
magnitudes, or by different mechanisms. 289
The reduction in the content of C16 denotes part of the increase in the β-oxidation flux, 290
and reasons for this could be the same that caused an increase in the content of C2. The 291
activation of PPARα and γ, as well as the up-regulation of the CPT-1 have been shown to 292
enhance β-oxidation36-38. The fact that exposures to 100 µM 13C-PAM for more than 2 days 293
did not significantly change the level of C2 generated in the myotubes, denotes that during 294
these conditions, FAO is limited or even inhibited. This suggestion was confirmed by the 295
observation of an increased content of C16/C2 at 100 µM 13C-PAM, during 2-4 days of 296
incubation (Figure 4c). FAO is known to be inhibited by high doses of palmitate 39. Thus, 297
as the content of 13C-PAM in the myotubes is elevated by increasing the dose from 5-100 298
µM, there is a gradual tendency of reduction in the flux of C16, which eventually becomes 299
significant, regardless of the length of incubation (Figure 4c) 300

B r i t a N g u m C h e P h D T h e s i s
132
Many factors can account for the reduction in β-oxidation flux in the myotubes at high 301
palmitate concentrations. It is likely, that at higher concentrations of 13C-PAM, its uptake 302
into cells is either increased, or unaffected, but its oxidation is reduced through 303
increments and drops in the activities of malonyl CoA and CPT-1, respectively40. High 304
amounts of C2 generated in the presence of high doses of 13C-PAM can also be converted 305
to malonyl-CoA, which will, in turn suppress the generation of C16 by suppressing the 306
action of CPT-1 307
4.3 Impact of PA on β- oxidation 308
Myotubes were treated with different doses of the potential PPAR agonist PA together with 309
20 µM of the well-known PPAR agonists OLA 11 for 4h, 24 h or 48 h. Afterwards, the 310
myotubes were analyzed for β- oxidation capacity by exposure to 50 µM 13C-PAM for 24 h, 311
and analysis of the C2 and C16/C2 contents of the cell culture media, reflecting the 312
myotube content (Figures 5a and b). By relating the contents of C16 to that of C2, it was 313
possible to observe induced inhibition (higher C16/C2 ratios, when compared to controls) 314
or activation (lower C16/C2 ratios, when compared to controls) of β- oxidation. 315
The effect of PA on the content of C16 or C2 was not dose-dependent, and when there was 316
a significant effect of treatment, 10 µM PA activated β-oxidation either equally or even 317
more than 20 µM (Figures 5a and b). The used of PA over 10 µM compromises the viability 318
of the myotubes (unpublished data) and could be a reason for these observations. 319
After 24 h of incubation with 10 µM PA or OLA, FAO in the myotubes was increased, and 320
during further incubation, even 5 µM PA enhanced FAO, as observed in the increase in the 321
content of C2 (Figure 5a) and in the reduction in the content of C16/C2 (Figure 5b). Both 322
OLA and PA are PPAR agonists, and thus have FAO-inducing potentials11;41. The activation 323

B r i t a N g u m C h e P h D T h e s i s
133
of PPAR α and γ, as well as the up-regulation of the CPT-1 are possible causes to this 324
increase in C16 oxidation 36-38. More so, being PPARα agonists, one can anticipate that PA 325
and OLA, just like bezafibrates activate very long chain acyl coenzyme A dehydrogenases 326
42, thus, enhancing the oxidation of long-chain FAs. 327
Exposure time also affected FAO, as longer incubation periods with PA or OLA increased 328
the content of C2 (Figure 4a), as well as reduced the content of C16/C2 (Figure 5b). 329
Already after 4h of incubation with OLA, the content of C2 was elevated, showing that OLA 330
is possibly a more potent enhancer of FAO, being an unsaturated FA 43. The metabolism of 331
PA involves peroxisomal α- oxidation and racemase activity, in order to generate active 332
metabolites that are involved in FA metabolism, through the activation of PPARs 44;45. This 333
might account for a delay in its activation of FAO, as seen by the increase in the content of 334
C2, or reduction of C16/C2 only after 24h of incubation (Figures 5a and b). 335
As shown by the contents of C16/C2, no changes in the degradation of C16 by the 336
treatments were recorded during the first 4 h of incubation (Figure 5b). This could be 337
attributed to minimal oxidation rates during this time, but possibly because the treatments 338
simultaneously enhanced the uptake of U-13 C-PAM and eventually its activated form 339
(palmitoyl-CoA) into the cell and mitochondria, respectively. Both PA and OLA have been 340
reported to enhance FA binding proteins46;47, and as such, FA uptake. One could therefore 341
suggest that both PA and OLA affect β-oxidation by affecting both the uptake and the 342
degradation of 13C-PAM. More so, during 4 h treatment with OLA, the content of C2 was 343
increased but that of C16/C2 was unchanged, insinuating that OLA affects the rate of FAO 344
and that of FA uptake into the cell and/or mitochondria, differently. 345
5. Conclusions 346

B r i t a N g u m C h e P h D T h e s i s
134
In this study, primary porcine myotubes were established as an effective model for 347
determining the flux of β- oxidation, as they could successfully take up 13C-PAM into the 348
cell, generate and incorporate C16 into the mitochondria, and eventually degrade the 349
incorporated C16 into C2. Furthermore, the time and dose-dependent effect of palmitate 350
on FAO revealed that, longer exposures of myotubes to palmitate induced FAO by 351
enhancing the generation of both C2 and C16, while only the generation of C2 was induced 352
with different doses of palmitate. Thus, the FA metabolism of the myotubes was regulated 353
at both the uptake and oxidation levels, as same experimental conditions enhanced the 354
generation of C2 but not of C16. The treatments of myotubes with PA or OLA enhanced 355
FAO in a time-dependent fashion, and possibly in a PPARα- dependent manner. However, 356
concrete conclusions on our observations of PA on β-oxidation, awaits further analysis 357
with the inclusion of a PPARα control, e.g. bezafibrates, which will give a more convincing 358
understanding of the impact of PA on β- oxidation in primary porcine myotubes. 359
Abbreviations 360
Acyl-CoA, acylcoenzyme A; 13C-PAM, [U-13 C]-Potassium palmitate;C2, acetylcarnitine; 361
C16, palmitoylcarnitine; CACT, carnitine-acylcarnitine translocase; CLA, conjugated 362
linoleic acid; CPT, carnitine palmitoyl transferase; DMEM, Dulbecco's Modified Eagle's 363
Medium; EM, experimental media; FA, fatty acid; FAO, fatty acid oxidation; FBS, fetal 364
bovine serum; HS, horse serum; IS, internal standard; MS, metabolic syndrome; MTP, 365
mitochondria trifunctional protein; OLA, oleic acid; PA, phytanic acid; PGM, 366
proliferation/growth medium; PPAR, peroxisome proliferator-activated receptor; 367
368
369

B r i t a N g u m C h e P h D T h e s i s
135
Acknowledgments 370
The authors thank Aase Sørensen and Anne Hjorth Bailing for proof reading of the 371
manuscript, and Niels Oksbjerg for his help in statistical analysis. 372
Funding 373
This study is part of the project; Tailored milk for human health, and was supported by 374
the Danish Council for Strategic Research and The Danish Cattle Federation. 375
Disclosure of statement 376
The authors declare no competition in interests 377
Authors’ contribution 378
B.N.C. designed and performed experiments, analyzed data and wrote the manuscript. 379
R.I.D.R. contributed in the analysis of acylcarnitine data. M.J. contributed in providing 380
instrumentation for acylcarnitine measurements, discussion and interpretation of 381
manuscript. N.G. contributed in the conception, discussion, interpretation, review and 382
editing of manuscript. L.I.H. contributed to the conception of the project, discussion, 383
interpretation, review and editing of the manuscript. J.F.Y. contributed to the 384
experimental design, discussion, review and editing of manuscript. 385
386

B r i t a N g u m C h e P h D T h e s i s
136
Reference List 387
388
1. Zhang L, Keung W, Samokhvalov V, et al: Role of fatty acid uptake and fatty acid 389
beta-oxidation in mediating insulin resistance in heart and skeletal muscle. 390
Biochimica et Biophysica Acta - Molecular and Cell Biology of Lipids 1801:1-391
22, 2010 392
2. Simoneau JA, Veerkamp JH, Turcotte LP, et al: Markers of capacity to utilize fatty 393
acids in human skeletal muscle: Relation to insulin resistance and obesity and 394
effects of weight loss. Faseb Journal 13:2051-2060, 1999 395
3. Kelley DE, Simoneau JA: Impaired free fatty acid utilization by skeletal muscle in 396
non-insulin- dependent diabetes mellitus. Journal of Clinical Investigation 397
94:2349-2356, 1994 398
4. Denis McGarry J: Dysregulation of fatty acid metabolism in the etiology of type 2 399
diabetes. Diabetes 51:7-18, 2002 400
5. Hulver MW, Dohm GL: The molecular mechanism linking muscle fat accumulation to 401
insulin resistance. Proceedings of the Nutrition Society 63:375-380, 2004 402
6. Grimaldi PA: Peroxisome proliferator-activated receptors as sensors of fatty acids and 403
derivatives. Cellular and Molecular Life Sciences 64:2459-2464, 2007 404
7. Gilde AJ, Van Bilsen M: Peroxisome proliferator-activated receptors (PPARS): 405
Regulators of gene expression in heart and skeletal muscle. Acta Physiologica 406
Scandinavica 178:425-434, 2003 407
8. Panadero MI, González MC, Herrera E, et al: Factors modulating fibrates response: 408
Therapeutic implications and alternative strategies. Endocrine, Metabolic and 409
Immune Disorders - Drug Targets 9:219-236, 2009 410
9. Minnich A, Tian N, Byan L, et al: A potent PPAR alpha agonist stimulates 411
mitochondrial fatty acid beta-oxidation in liver and skeletal muscle. American 412
Journal of Physiology - Endocrinology and Metabolism 280:E270-E279, 2001 413
10. Oosterveer MH, Grefhorst A, van Dijk TH, et al: Fenofibrate simultaneously induces 414
hepatic fatty acid oxidation, synthesis, and elongation in mice. Journal of 415
Biological Chemistry 284:34036-34044, 2009 416
11. Coll T, Eyre E, Rodriguez-Calvo R, et al: Oleate reverses palmitate-induced insulin 417
resistance and inflammation in skeletal muscle cells. Journal of Biological 418
Chemistry 283:11107-11116, 2008 419

B r i t a N g u m C h e P h D T h e s i s
137
12. Houseknecht KL, Heuvel JPV, Moya-Camarena SY, et al: Dietary conjugated linoleic 420
acid normalizes impaired glucose tolerance in the Zucker diabetic fatty fa/fa 421
rat. Biochemical and Biophysical Research Communications 244:678-682, 422
1998 423
13. Zomer AWM, van der Burg B, Jansen GA, et al: Pristanic acid and phytanic acid: 424
naturally occurring ligands for the nuclear receptor peroxisome proliferater-425
activated receptor alpha. Journal of Lipid Research 41:1801-1807, 2000 426
14. Roche HM, Noone E, Nugent A, et al: Conjugated linoleic acid: A novel therapeutic 427
nutrient? Nutrition Research Reviews 14:173-187, 2001 428
15. Hellgren LI: Phytanic acid-an overlooked bioactive fatty acid in dairy fat? Annals of 429
the New York Academy of Sciences 1190:42-49, 2010 430
16. Gloerich J, van Vlies N, Jansen GA, et al: A phytol-enriched diet induces changes in 431
fatty acid metabolism in mice both via PPAR alpha-dependent and -432
independent pathways. Journal of Lipid Research 46:716-726, 2005 433
17. Ramsay RR, Arduini A: The carnitine acyltransferases and their role in modulating 434
acyl-CoA pools. Archives of Biochemistry and Biophysics 302:307-314, 1993 435
18. Wanders RJA, Ruiter JPN, Ijlst L, et al: The enzymology of mitochondrial fatty acid 436
beta-oxidation and its application to follow-up analysis of positive neonatal 437
screening results. Journal of Inherited Metabolic Disease 33:479-494, 2010 438
19. Gregersen N, Andersen BS, Pedersen CB, et al: Mitochondria fatty acid oxidation 439
defects-remaining challenges. Journal of Inherited Metabolic Disease 31:643-440
656, 2008 441
20. Ventura FV, Costa CG, Struys EA, et al: Quantitative acylcarnitine profiling in 442
fibroblasts using [U- 13C] palmitic acid: An improved tool for the diagnosis of 443
fatty acid oxidation defects. Clinica Chimica Acta 281:1-17, 1999 444
21. Young SP, Matern D, Gregersen N, et al: A comparison of in vitro acylcarnitine 445
profiling methods for the diagnosis of classical and variant short chain acyl-446
CoA dehydrogenase deficiency. Clinica Chimica Acta 337:103-113, 2003 447
22. Carey GB: The swine as a model for studying exercise-induced changes in lipid 448
metabolism. Medicine and Science in Sports and Exercise 29:1437-1443, 1997 449
23. Theil PK, Sorensen IL, Nissen PM, et al: Temporal expression of growth factor genes 450
of primary porcine satellite cells during myogenesis. Animal Science Journal 451
77:330-337, 2006 452
24. Smith PK, Krohn RI, Hermanson GT, Mallia AK et al: Measurement of protein using 453
bincinchoninic acid. Analytical Biochemistry 150:76-85, 1985 454

B r i t a N g u m C h e P h D T h e s i s
138
25. Okura T, Igase M, Kitami Y, et al: Platelet-derived growth factor induces apoptosis in 455
vascular smooth muscle cells: roles of the Bcl-2 family. Biochimica et 456
Biophysica Acta-Molecular Cell Research 1403:245-253, 1998 457
26. Schönfeld P, Wojtczak L: Fatty acids as modulators of the cellular production of 458
reactive oxygen species. Free Radical Biology and Medicine 45:231-241, 2008 459
27. Schmitz-Peiffer C, Craig DL, Biden TJ: Ceramide generation is sufficient to account 460
for the inhibition of the insulin-stimulated PKB pathway in C2C12 skeletal 461
muscle cells pretreated with palmitate. Journal of Biological Chemistry 462
274:24202-24210, 1999 463
28. Yuzefovych L, Wilson G, Rachek L: Different effects of oleate vs. palmitate on 464
mitochondrial function, apoptosis, and insulin signaling in L6 skeletal muscle 465
cells: Role of oxidative stress. American Journal of Physiology - Endocrinology 466
and Metabolism 299:E1096-E1105, 2010 467
29. Cha BS, Ciaraldi TP, Park KS, et al: Impaired fatty acid metabolism in type 2 diabetic 468
skeletal muscle cells is reversed by PPAR gamma agonists. American Journal 469
of Physiology - Endocrinology and Metabolism 289:E151-E159, 2005 470
30. Thompson DB, Pratley R, Ossowski V: Human primary myoblast cell cultures from 471
non-diabetic insulin resistant subjects retain defects in insulin action. Journal 472
of Clinical Investigation 98:2346-2350, 1996 473
31. Lieth E, Cardasis CA, Fallon JR: Muscle-derived agrin in cultured myotubes: 474
Expression in the basal lamina and at induced acetylcholine receptor clusters. 475
Developmental Biology 149:41-54, 1992 476
32. Gopinath SD, Rando TA: Stem Cell Review Series: Aging of the skeletal muscle stem 477
cell niche. Aging Cell 7:590-598, 2008 478
33. Oksbjerg N, Gondret F, Vestergaard M: Basic principles of muscle development and 479
growth in meat-producing mammals as affected by the insulin-like growth 480
factor (IGF) system. Domestic Animal Endocrinology 27:219-240, 2004 481
34. Mann C, Leckband D: Measuring traction forces in long-term cell cultures. Cellular 482
and Molecular Bioengineering 3:40-49, 2010 483
35. Merrill GF, Kurth EJ, Rasmussen BB, et al: Influence of malonyl-CoA and palmitate 484
concentration on rate of palmitate oxidation in rat muscle. Journal of Applied 485
Physiology 85:1909-1914, 1998 486
36. Guan Y, Zhang Y, Breyer MD: The role of PPARs in the transcriptional control of 487
cellular processes. Drug News and Perspectives 15:147-154, 2002 488

B r i t a N g u m C h e P h D T h e s i s
139
37. Jay MA, Ren J: Peroxisome proliferator-activated receptor (PPAR) in metabolic 489
syndrome and type 2 diabetes mellitus. Current Diabetes Reviews 3:33-39, 490
2007 491
38. Ciaraldi, T. P., Cha, B. S., Park, K. S., et al: Free fatty acid metabolism in human 492
skeletal muscle is regulated by PPAR gamma and RXR agonists. Annals of the 493
New York Academy of Sciences 967, 66-70. 2002. 494
39. Molé PA, Oscai LB, Holloszy JO: Adaptation of muscle to exercise. Increase in levels 495
of palmityl Coa synthetase, carnitine palmityltransferase, and palmityl Coa 496
dehydrogenase, and in the capacity to oxidize fatty acids. Journal of Clinical 497
Investigation 50:2323-2330, 1971 498
40. Rasmussen BB, Holmbäck UC, Volpi E, et al: Malonyl coenzyme A and the regulation 499
of functional carnitine palmitoyltransferase-1 activity and fat oxidation in 500
human skeletal muscle. Journal of Clinical Investigation 110:1687-1693, 2002 501
41. Gloerich J, van den Brink DM, Ruiter JPN, et al: Metabolism of phytol to phytanic 502
acid in the mouse, and the role of PPAR alpha in its regulation. Journal of 503
Lipid Research 48:77-85, 2007 504
42. Djouadi F, Aubey F, Schlemmer D, et al: Bezafibrate increases very-long-chain acyl-505
CoA dehydrogenase protein and mRNA expression in deficient fibroblasts and 506
is a potential therapy for fatty acid oxidation disorders. Human Molecular 507
Genetics 14:2695-2703, 2005 508
43. Rivellese AA, Maffettone A, Vessby B, et al: Effects of dietary saturated, 509
monounsaturated and n-3 fatty acids on fasting lipoproteins, LDL size and 510
post-prandial lipid metabolism in healthy subjects. Atherosclerosis 167:149-511
158, 2003 512
44. Jansen GA, Wanders RJA: Alpha-Oxidation. Biochimica et Biophysica Acta - 513
Molecular Cell Research 1763:1403-1412, 2006 514
45. Wanders RJA, Jansen GA, Lloyd MD: Phytanic acid alpha-oxidation, new insights 515
into an old problem: A review. Biochimica et Biophysica Acta - Molecular and 516
Cell Biology of Lipids 1631:119-135, 2003 517
46. Wolfrum C, Ellinghaus P, Fobker M, et al: Phytanic acid is ligand and transcriptional 518
activator of murine liver fatty acid binding protein. Journal of Lipid Research 519
40:708-714, 1999 520
47. Meunier-Durmort C, Poirier H, Niot I, et al: Up-regulation of the expression of the 521
gene for liver fatty acid-binding protein by long-chain fatty acids. Biochemical 522
Journal 319:483-487, 1996 523

B r i t a N g u m C h e P h D T h e s i s
140
524
525
526

B r i t a N g u m C h e P h D T h e s i s
141
Figure legends 527
Figure 1: Experimental setups 528
Two experimental setups were established to study the effects of time and dose of 13C-PAM 529
(1a), and the effects of time-dependent treatments (1b) on the palmitoyl-and 530
acetylcarnitine profiles in primary porcine myotubes. Myotubes in stock media (white 531
bars) were given either EMI (black bars) on a daily basis for 1 - 4 days (Figure 1a) or EM II 532
(grey bars) for 4 h, 24 h or 48 h, followed by a quick rinse and afterwards EM I for 24 h 533
(Figure 1b). In both setups, the EM I collected from the cells was stored at -20oC until use 534
for acyl carnitine analysis. 535
536
Figure 2: Viability of porcine primary myotubes 537
Differentiated porcine myotubes were exposed to varying doses of 13C-PAM (0-300 µM) for 538
up to 4 days and afterwards subjected to viability assay as described in the methods. 539
Control samples (0) had the same media (EM I) as other samples, but lacked 13C- PAM. 540
Data are expressed as mean values with SEM from triplicate wells, carried out in three 541
separate experiments with cells isolated from a different pig for every experiment. * denote 542
significant difference between doses of U-13 C-PAM used within a time period, with regards 543
to their respective controls (P < 0.05). 544
545
Figure 3: Extracted UPLC-MS/MS chromatogram of 13C-palmitoylcarnitine 546 (C16). 547
A peak specific to C16 is detected by use of the retention time, a quantifier and a qualifier. 548

B r i t a N g u m C h e P h D T h e s i s
142
549
Figure 4: Acylcarnitine contents of C2 and C16 as affected by incubation time 550
and dose of 13C- PAM 551
Differentiated porcine myotubes were exposed to varying doses of 13C- PAM (0-100 µM) 552
for 1 day (closed circles), 2 days (open circles), 3 days (closed triangles) or 4 days (open 553
triangles) and the media collected for acyl carnitine analysis. The contents of C2 (a) and 554
C16 (b) were measured using a fast and simple extraction procedure followed by UPLC-555
MS/MS analysis.The ratio of C16/C2 (c) was calculated in relation to the content generated 556
after 1-day incubation with 5 µM 13 C-PAM. Data are expressed as mean values with SEM, 557
and are generated from samples collected from triplicate wells, carried out in three 558
separate experiments, with cells isolated from a different pig each time. * applies only to 559
figure 4c and denotes significant difference in relation to treatments with 5 µM 13C-PAM 560
during each incubation period. 561
Figure 5: Contents of palmitoylcarnitine and acetylcarnitine as affected by 562
time-dependent treatments of myotubes 563
Differentiated porcine myotubes were exposed to varying doses of PA (0-20 µM) or 20 µM 564
OLA for 4 h, 24 h or 48 h and afterwards to EM II containing 50 µM U-13 C-PAM for 24 h. 565
Media was collected and analyzed for their C2 (a) and C16/C2 contents (b). Control 566
samples were only exposed to stock media before EM II. Data are expressed as mean 567
values with SEM and are related to control samples. The data are generated from samples 568
collected from 6-9 different wells, carried out in two separate experiments, with cells 569
isolated from a different pig for every experiment. Significant differences with regards to 570
controls for each time period (4h, 24 h or 48 h) are denoted with *; P < 0.05. 571

B r i t a N g u m C h e P h D T h e s i s
143
572
573
574

B r i t a N g u m C h e P h D T h e s i s
144
Figures 575
576
577
Figure 1a 578
579
Figure 1b 580

B r i t a N g u m C h e P h D T h e s i s
145
Incubation time with varying doses of µM 13
C-PAM (days)
1 2 3 4
Fo
ld c
ha
ng
e in
via
bil
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0
5
10
25
50
100
200
300
***
*
*
* *
*
*
*
*
*
*
*
*
*
581 582
Figure 2 583
584

B r i t a N g u m C h e P h D T h e s i s
146
585
Figure 3 586
min 0.620 0.640 0.660 0.680 0.700 0.720 0.740 0.760 0.780 0.800 0.820 0.840 0.860 0.880 0.900 0.920 0.940
%
0
100 F11:MRM of 5 channels,ES+
416.34 > 357.34
13C-Palmitoylcarnitine (C16)
0.84 min.
min
%
0
100 F11:MRM of 5 channels,ES+
416.34 > 85.05
13C-Palmitoylcarnitine (C16)
0.84 min.

B r i t a N g u m C h e P h D T h e s i s
147
C2
13C-PAM concentration (µM)
control 5 10 25 50 100
Ace
tylc
arn
itin
e co
nte
nt
x10
(µ
M/m
g p
rote
in)
0
2
4
6
C2
587
Figure 4a 588

B r i t a N g u m C h e P h D T h e s i s
148
13C-PAMconcentration (µM)
control 5 10 25 50 100
Pa
lmit
oy
lca
rnit
ine
con
ten
t x
10 (
µM
/mg
pro
tein
)
0
1
2
3
4
5
6
C16
589
Figure 4b 590

B r i t a N g u m C h e P h D T h e s i s
149
C16/C2
13C PAM concentration (µM)
5 10 25 50 100
Fo
ld c
han
ge
in C
16/C
2 c
on
ten
t (µ
M/m
g p
rote
in)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
*
*
*
*
591 592
Figure 4c 593

B r i t a N g u m C h e P h D T h e s i s
150
594
C2
Incubation time time (h)
4 24 48
Fo
ld c
han
ge i
n C
2 c
on
ten
t (µ
M/m
g p
rote
in)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
*
* **
*
*
*
*
595
Figure 5a 596
597

B r i t a N g u m C h e P h D T h e s i s
151
C16/C2
Incubation time (h)
4 24 48
Fo
ld c
ha
ng
e in
C16
/C2
co
nte
nt
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
**
*
* **
**
598
Control 1 µM PA 5 µM PA 10 µM PA 20 µM PA 20 µM OLA 599
Figure 5b 600

B r i t a N g u m C h e P h D T h e s i s
152
6.3 Manuscript III; MIII
Content and distribution of phytanic acid diastereomers in organic
milk as affected by feed composition
Brita N. Che†, Troels Kristensen‡, Caroline Nebel†, Trine K. Dalsgaard†, Lars I.
Hellgren§, Jacob H. Nielsen#, Jette F. Young† Mette K. Larsen†*
† Department of Food Science, ‡ Department of Agroecology, Aarhus University, AU
Foulum, Blichers Allé 20, DK-8830 Tjele, Denmark
§ Department of System Biology, Center for Biological Sequence Analysis, Technical
University of Denmark, Kemitorvet, Building 208, DK-2800 Kgs. Lyngby, Denmark
# Arla Foods, Sønderhøj 14, DK-8260 Viby J, Denmark
* Corresponding author: Mette K. Larsen
Tel: +45 87 15 80 62
Fax: +45 87 15 48 91

B r i t a N g u m C h e P h D T h e s i s
153
ABSTRACT: Phytanic acid (PA) is a bioactive compound of milk, which is derived from 1
the phytol chain of chlorophyll, and the content of PA in milk-fat depends on the 2
availability of phytol from feed. In this study, the content of PA diastereomers was 3
analyzed in milk sampled from five organic herds, twice during the grazing season (May 4
and September). The total content of PA was higher in September, when compared to May, 5
but was not affected by breed (Danish Holstein or Danish Jersey). Total PA could not be 6
directly related to intake of green feed items. The distribution between PA diastereomers 7
was closely related to the amount of grazed legumes, where a higher intake resulted in a 8
higher share of the RRR isomer. 9
10
KEYWORDS: Organic milk, phytanic acid diastereomers, pasture, legumes, silage 11

B r i t a N g u m C h e P h D T h e s i s
154
INTRODUCTION 12
Recent findings point to an inverse correlation between increased dairy consumption 13
and the metabolic syndrome (MS), 1, 2 even though dairy products are a rich source of 14
saturated fatty acids. This controversy has directed the focus towards the search for 15
components in milk that are responsible for neutralizing the adverse effects of the 16
metabolically detrimental saturated fatty acids. One such component is 3, 7, 11, 15 17
tetramethylhexadecanoic acid (phytanic acid (PA)). In the human food chain, PA is 18
primarily found in dairy products, ruminant meat and some marine fats. PA is formed 19
from phytol, which is cleaved from chlorophyll and subsequently oxidized by the rumenal 20
microbiota and certain marine organisms.3, 4 21
As PA is derived from chlorophyll, the content in milk-fat is dependent on the feed 22
composition, where increased amounts of green feed increase the content of PA. Due to the 23
higher use of grass-based products, PA content is higher in organic milk, when compared 24
to conventional milk, and a threshold value of the PA content in milk-fat of at least 200 25
mg/g has been suggested as a marker of organic milk products.5 26
PA, as well as its primary metabolite pristanic acid, are natural agonists of the 27
peroxisome proliferator activated receptor-α (PPARα) and the retinoid X receptor (RXR).6 28
As RXRs and the PPARα are known to control fatty acid and glucose metabolism,7 PA has 29
been suggested to have health-improving properties and a protective effect on MS.6, 8 30
Recently, we have shown that PA can increase glucose uptake in primary porcine myotubes 31
(unpublished data), although there was no concomitant increase in glycogen synthesis. 32
PA has 3 chiral centers positioned on carbons 3, 7 and 11. Carbons 7 and 11 are R- 33
configured as in phytol, whereas carbon 3 is either R- or S- configured as a result of the 34

B r i t a N g u m C h e P h D T h e s i s
155
biohydrogenation of oxidized phytol. As such, PA occurs naturally as two diastereomers; 35
SRR PA and RRR PA.3 The distribution profile of the PA diastereomers in lipid fractions in 36
serum varies, with higher SRR/RRR ratio in phospholipids than in other lipid fractions.9 37
More so, the biological activity of the diastereomers of PA in humans differ. At higher PA 38
concentrations, the oxidation of SRR PA is preferred by up to 50%, compared to the RRR 39
form,10 whereas at lower PA concentrations, both forms exhibit similar oxidation rates.10, 11 40
The initial step in PA oxidation is peroxisomal α-oxidation to pristanic acid (2,6,10,14 41
tetramethylpentadecanoic acid) which is further degraded by β-oxidation.12 The β-42
oxidation is stereospecific and requires α-methylacyl-CoA racemase (AMCAR) to convert 43
2R-pristanoic acid to its 2S isomer.12 These differences in the oxidation and distribution 44
profiles of PA diastereomers hint on potential variations in their physiological functions 45
and warrant an elucidation of their origin in ruminants. 46
The distribution of the two diastereomers of PA varies in food, with the SRR form 47
predominant in marine animals and some terrestrial mammals having more of the RRR 48
form.3, 13 The distribution between the two isomers varies between bovine milks of 49
different origin where organic farming gives higher shares of the RRR isomer.13, 14 Based on 50
these results it has been suggested that the threshold value of 200 mg/g PA in organic milk 51
fat5 should be replaced by a combination of total PA content and the ratio between the 52
diastereomers for authentication of organic milk.14 53
Organic dairy management varies between countries due to tradition as well as climatic 54
differences.15 In Danish organic dairy farming there is a compulsory use of grazing when 55
climatic conditions allow, which is in accordance with the definition in the European 56
Council Regulations,16 however, the extent of grazing varies considerably between farms. 57
Other main feed items include grass and maize silage, and concentrates.15 The feed 58

B r i t a N g u m C h e P h D T h e s i s
156
composition differs significantly from the ratios of pure hay or pure grass silage 59
investigated by Schröder et al (2012),14 and PA concentrations in milk fat may be lower as 60
the content of grass based feed is lower. 61
The present study was carried out as a survey of five Danish commercial organic herds 62
in the grazing season, and the purpose was to test whether the suggested target value of 63
200 mg/g PA in milk fat5 could be met, to investigate how the content of the two 64
diastereomers of PA varied in bulk milk, and to test the hypothesis that these differences 65
could be related to specific feeding patterns. 66
67
MATERIALS AND METHODS 68
The experiment was conducted at five Danish commercial organic dairy herds. 69
Registration of feed consumption and milk production was made during two periods of 14 70
days within the grazing season, where the first period started in spring about two weeks 71
after start of grazing, and the second period started in late summer at the end of August. 72
Milk was sampled from the bulk tank milk on two days at the end of each period. 73
Herds and feeding. Three herds (DH1, DH2 and DH3), were Danish Holstein cows 74
(DH), while the two other herds (DJ1 and DJ2) were Danish Jersey (DJ). All farms 75
practiced calving all year round, and all lactating cows participated in the experiment, as 76
the herd was the experimental unit. Feeding of supplemental feed as roughage (grass 77
silage, maize silage, hay or straw), or a mix of roughage and concentrate were applied 78
indoors at the feeding lane with one feeding space to each cow, whereas concentrate 79
(commercial mix and cereals) in all herds was applied in the milking parlour, divided in 80
equal amounts and given twice daily. 81

B r i t a N g u m C h e P h D T h e s i s
157
Pastures. At all farms, pastures dominated by perennial ryegrass (Lolium perenne) 82
and white clover (Trifolium repens) were part of a cropping rotation with arable crops. 83
Rotations were typically based on 3-year leys and grassland was established by seeding 84
cereals with grass clover mixtures. 85
Registration and calculation of feed intake. Milk yield was recorded once in 86
each period for each cow and the contents of fat, protein and urea were analysed 87
(Milkoscan 4000, Foss Electric A/S, Hillerød, Denmark). Milk yield as energy-corrected 88
milk (ECM) was calculated according to Sjaunja et al. (1990)17, and average contents of fat, 89
protein and urea at herd level were calculated based on the milk recordings from each 90
lactating cow. 91
Registration of the pasture was made three times during each period. Samples were 92
collected in 25 places at each farm by hand-plucking the pasture at the height at which 93
cows were observed to graze. The botanical composition of the dry matter (DM) was 94
determined after drying at 60°C to constant weight. The relative amounts of the individual 95
species were recorded, and the relative amount of legumes calculated as the sum of white 96
clover and red clover. 97
Three times during each period, intake of supplemental feed was determined by weight 98
over one day at herd level. Feed samples of concentrates and roughage were taken once 99
during each period. Each sample of roughage and supplement feed was analysed by near 100
infrared reflectance spectroscopy. Net energy value of lactation (NE) was estimated based 101
on Hvelplund et al (2007).18 102
The energy requirements for maintenance, grazing activity, lactation and live weight 103
change was calculated as described by Macoon et al. (2003)19 based on animal 104

B r i t a N g u m C h e P h D T h e s i s
158
performance data at group level, as the intake of the indoor feed was registered at group 105
level. Pasture DM intake (DMI) was estimated as the difference between the requirement 106
and NE in supplemental feed at group level divided by the NE concentration of the hand-107
plucked samples. 108
The DMI of the different species in the pasture was estimated proportionally to the 109
botanical analysis of the hand-plucked samples assuming no selectivity. DMI of individual 110
feed items was expressed as average relative distribution (kg DM/kg total DMI) at herd 111
level for each period. 112
Fatty acid extraction from milk. Milk (10 mL) was centrifuged at 1700 g at 4oC for 113
20 min and the cream phase transferred to a new tube. The cream was centrifuged for 10 114
min at 13000 g at 20°C and afterwards placed on a 60oC heater for 10 min. The heated 115
cream was then centrifuged for 10 min at 13000 g at 40oC and fat was sampled from the 116
liquid fat layer. A volume of 15 mg of fat was weighed out and mixed with 1 ml heptane 117
containing 0.4 mg/mL of C12:1 cis 11 triglyceride (Nu-Chek-Prep Inc, Elysian, MN, USA) 118
as an internal standard. For methylation 10 µl of a 2.2M sodium methylate was added. The 119
mixture was vortexed for 1 min and allowed to stand for 10 min at room temperature. The 120
methylated mixture was centrifuged for 5 min at 13000 g at 4oC and the heptane phase 121
collected for GC-MS analysis. 122
GC-MS analysis of methyl esters of PA diastereomers. Methylated milk fat 123
extracts were analyzed in details for PA isomers on a GC-MS (GC 6890N from Agilent 124
Technologies (Waldbronn, Germany)) equipped with a Restek 2560 column (100 m x 0.25 125
mm x 0.20 µm, Supelco, Bellafonte, PA, USA). A 1 μL aliquot of the extracts was injected in 126
split mode (20:1) into an inlet with a temperature of 250° C. The programmed column 127
temperature was isothermal at 100° C for 5 min followed by an increase to 140° C with a 128

B r i t a N g u m C h e P h D T h e s i s
159
rate of 3°C/min, a hold time of 5 min, followed by a raise to 160° C with a rate of 5° C/min 129
and a hold time for 20 min. Subsequently, the temperature was raised to 220° C with a rate 130
of 12° C/min and held there for 8 min. Finally, an increase to 240 ° C was performed with a 131
rate of 25° C/min and held there for 10 min before the next injection. Mass spectral 132
analysis was performed on a quadrupole MSD 5975 (Agilent Technologies, Germany) using 133
an external standard curve in selected ion monitoring using the ion 101 m/z as target and 134
the ions 57, 74, and 171 m/z as qualifier ions for the quantification of the PA isomers (SRR 135
and RRR). For the internal standard (C12:1) the ion 96 m/z was used as target ion, and 136
110, 123, 138 m/z were used as qualifier ions. The analysis was performed keeping the 137
quadrupole temperature of 150° C and a fragmentation voltage of 70 eV. The ion source 138
temperature was 230° C, and the interface was 280° C. The flow rate for helium carrier gas 139
was set at 1.0 mL/min. 140
Content of PA in the milk samples was calculated based on a standard curve made from 141
synthetic PA methyl esters (Sigma-Aldrich A/S, Brøndby, Denmark) and variation in 142
response between different samples was corrected using the internal standard. 143
Calculations and statistical treatment. The total amount of PA in milk fat was 144
calculated as the sum of the two isomers and the share of RRR isomer in total PA was 145
calculated as a measure of the distribution of isomers. Data were treated statistically by use 146
of SAS, version 9.2 for Windows (SAS Institute Inc, Cary, NC). The first model investigated 147
the main effects of breed and period, the second model was a one-way ANOVA of the effect 148
of combinations of herd and period. Pearson correlation coefficients between PA results 149
and share of individual feed items were calculated to determine relations between feed 150
components and PA. 151
152

B r i t a N g u m C h e P h D T h e s i s
160
RESULTS 153
Identification of PA diastereomers in milk fat. GC-MS analysis showed two PA 154
peaks of milk samples and three peaks of the synthetic standard (see Figure 1). A constant 155
ratio between the target (m/z101) and the qualifier ions (m/z 57, m/z 74, m/z 171) over the 156
whole peak was used for peak authentication and the first peak of the milk samples was 157
assigned as the SRR isomer of PA whereas the second peak of the milk samples was 158
assigned as the RRR isomer of PA according to literature.3, 13 159
Effect of breed and period. The concentration of total PA in milk fat was generally 160
higher in September compared to May (1.21 and 0.96 mg/g fat, respectively, p < 0.001), 161
and this difference could be ascribed to results for September of one DH herd and one DJ 162
herd (DH3 and DJ1) (see Figure 2). No overall breed difference was detected. There were 163
no significant effects of period or breed on the share of RRR isomer in total PA, despite 164
large variations (0.24 to 0.49) between different herds at different periods as shown in 165
Figure 3. 166
Effect of feed composition. The milk yield and feed consumption data are 167
presented in Table 1. Concentrations of fat and protein were higher and milk yields lower 168
for DJ compared to DH as normally observed for these breeds. There was a large variation 169
in feed composition between herds and the share of pasture ranged from 0.21 to 0.76, the 170
share of maize silage ranged from 0 to 0.48, the share of grass silage including hay and 171
straw ranged from 0 to 0.31, whereas the share of concentrate ranged from 0.10 to 0.34 172
corresponding to a total share of roughage from 0.66 to 0.90. Due to weather conditions 173
DMI from grazing was lower in September which was accompanied by a higher DMI of 174
silage as well as concentrate. Legumes constituted a larger part of pastures in September. 175

B r i t a N g u m C h e P h D T h e s i s
161
Pearson correlation coefficients of total PA as well as share of RRR isomer in total PA, 176
on individual feed groups are given in Table 2. Total PA was negatively correlated to intake 177
of pasture and positively correlated to intake of concentrate, whereas the share of RRR 178
isomer was positively correlated to the grazed amount of legumes. This latter correlation 179
coefficient was most significant and the share of RRR isomer in milk fat from different 180
herds at different periods related to the grazed amount of legumes is presented in Figure 4. 181
182
DISCUSSION 183
The concentrations of total PA in milk fat were only about half of the suggested 184
threshold value of 200 mg PA per100 g milk fat for organic milk.5 This could be due to 185
differences in Danish and German organic feeding practice; however, differences in 186
analytical procedures may have also affected results. The impact of different feeding 187
regimes on the content of PA in milk fat has been demonstrated by Schröder et al (2012)14 188
where cows have been switched from a conventional total mixed ratio to a ratio consisting 189
entirely of hay and afterwards back to the mixed ratio or to a ratio of only grass silage. 190
Average PA content in milk fat have been reported as 98-116 mg/100g after feeding the 191
mixed ratio, 153 mg/100 g at hay feeding, and 259 mg/100 g at grass silage feeding.14 192
These results demonstrate a positive relationship between the consumption of green 193
fodder and the PA content in milk fat; such relationship has also been documented by 194
Schröder et al (2011)20 where 50% or 87.5% green fodder has resulted in PA contents in 195
milk fat of 146 mg/100 g and 314 mg/100 g, respectively. Leiber et al (2005)21 have shown 196
a three-fold increase in PA content of milk-fat (from 0.15 to 0.45 g/100 g), when cows were 197
shifted from a mixed ratio based on hay, grass silage, maize silage and concentrate to a 198
ratio based entirely on fresh grass (either barn fed or pasture). Contrarily, in our present 199

B r i t a N g u m C h e P h D T h e s i s
162
results, grazing was negatively correlated and concentrates positively correlated to the 200
concentration of PA in milk fat. The negative correlation with grazing may be ascribed to 201
fatty acid oxidation products formed during the destruction of fresh plant material, which 202
alter the activity and composition of rumen microorganisms.22 The positive effect of 203
concentrate could be due to the use of grass pellets in concentrate mixtures which is 204
common in Danish organic farming. However, as differences in PA content in milk samples 205
were small and most samples did not differ significantly, the observed correlations may to 206
some extent reflect random variation. 207
The distribution between the diastereomers of PA depends on dairy management and 208
the share of SRR isomer has been reported as 39-71% in organic cheeses and 51-84% in 209
conventional cheeses.13 In milk from one organically and one conventionally fed cow the 210
share of SRR isomer has been reported as 47% and 85% , respectively.13 In the experiment 211
where feed ratios was shifted between a mixed ratio, pure hay and pure grass silage (see 212
above)14 the share of SRR isomer was reported as 50% during hay feeding and 70-80% 213
during feeding the mixed ratio as well as during grass silage feeding. These results 214
demonstrate that feed composition regulates the distribution of isomers, and hay favors 215
the formation of the RRR isomer. Our present results showed a high positive correlation 216
between the amount of grazed legumes and the share of RRR isomer. This relation 217
between legumes and isomer distribution could be an explanation of the higher share of 218
RRR isomer in organic products as the use of legumes is increased in organic agriculture to 219
ensure nitrogen fertilization. 220
The mechanisms involved in the formation of either SRR or RRR isomers have to our 221
knowledge not been previously reported. The R or S configuration of carbon 3 is formed 222
during the biohydrogenation of oxidized phytol, and possibly different microbial strains 223

B r i t a N g u m C h e P h D T h e s i s
163
form either predominantly SRR or RRR isomers. The biohydrogenation of most 224
unsaturated fatty acids takes place at carbon 9 or higher and different microorganisms are 225
involved in the hydrogenation step depending on the position and number of double 226
bonds.23 Similarly, the final hydrogenation of phytenic acid may involve other groups of 227
microorganisms. The hydrogenation of unsaturated fatty acids from feed has been 228
reported to be reduced by secondary plant metabolites from legumes.21 Likewise, these 229
secondary plant metabolites may favor strains forming predominantly RRR isomer or 230
suppress strains forming predominantly SRR isomer. 231
Very little is known on whether the intake of PA with different isomeric composition 232
has any physiological relevance. There is no difference in the agonist activity on PPARα, 233
between the isomeric forms of PA.11 However, pristanic acid, which is produced by α-234
oxidation of PA , is a considerable more potent PPARα agonist than PA,24 hence would a 235
shift towards more pristanic acid be expected to enhance PPARα activation. 236
It has been shown that the SRR-form is more rapidly oxidized in mitochondrial 237
preparations at high PA concentrations.10 However, since peroxisomal α-oxidation is 238
required for efficient β-oxidation of PA, the in vivo relevance of this is uncertain. But, as 239
the RRR-form requires isomerization by the AMCAR prior to β-oxidation,12 it is reasonable 240
to assume that the SRR-form of pristanic acid is more rapidly oxidized in the β-oxidation, 241
than the RRR-form, hereby decreasing the PA/pristanic acid ratio. 242
Thus, the intake of PA with a greater portion of SRR might be desirable as its oxidation 243
would be faster and less dependent on AMCAR, but on the other hand, a greater portion of 244
RRR might lead to higher concentrations pristanic acid, and hence more efficient 245
activation of PPARα. Which of these situations that is most advantageous for the 246
consumer cannot be concluded at present. In either case, feeding strategies can be 247

B r i t a N g u m C h e P h D T h e s i s
164
manipulated, for example by changing the content of legumes in the feed, to achieve an 248
altered content of RRR share of PA in dairy products. 249
In conclusion this study shows that total PA content in milk fat cannot be directly 250
related to the intake of green fodder items and other elements in the feed composition may 251
affect PA synthesis as well. The distribution of isomers was highly dependent on the 252
amount of legumes in feed and the results suggest that it should be possible to increase the 253
share of RRR isomer of PA by increasing the amount of legumes in feed. However, results 254
do not show whether this effect only relates to grazed legumes or similar effects could be 255
obtained by preserved legumes. The physiological relevance of the distribution of PA 256
isomers remain to be investigated. 257
258
ABBREVIATIONS USED 259
PA, phytanic acid; PPARα, peroxisome proliferator activated receptor-α; RXR, retinoid 260
X receptor; AMCAR, α-methylacyl-CoA racemase; DH, Danish Holstein; DJ, Danish 261
Jersey; ECM, energy corrected milk; DM, dry matter; metabolic syndrome, MS; NE, net 262
energy value of lactation; DMI, dry matter intake; GC-MS, gas chromatography mass 263
spectroscopy. 264
265
266
267
268
269

B r i t a N g u m C h e P h D T h e s i s
165
REFERENCES 270
(1) Azadbakht, L.; Mirmiran, P.; Esmaillzadeh, A.; Azizi, F. Dairy consumption is 271
inversely associated with the prevalence of the metabolic syndrome in Tehranian adults. 272
Am. J. Clin. Nutr. 2005, 82, 523-530. 273
(2) Pereira, M. A.; Jacobs, D. R.; Van Horn, L.; Slattery, M. L.; Kartashov, A. I.; Ludwig, 274
D. S. Dairy consumption, obesity, and the insulin resistance syndrome in young adults - 275
The CARDIA study. J. Am. Med. Ass. 2002, 287, 2081-2089. 276
(3) Ackman, R. G.; Hansen, R. P. Occurrence of Diastereomers of Phytanic and Pristanic 277
Acids and Their Determination by Gas-Liquid Chromatography. Lipids 1967, 2, 357-362. 278
(4) Patton, S.; Benson, A. A. Phytol Metabolism in Bovine. Biochim. Biophys. Acta 1966, 279
125 (1), 22-32. 280
(5) Vetter, W.; Schroder, M. Concentrations of phytanic acid and pristanic acid are higher 281
in organic than in conventional dairy products from the German market. Food Chem. 282
2010, 119, 746-752. 283
(6) Zomer, A. W. M.; van der Burg, B.; Jansen, G. A.; Wanders, R. J. A.; Poll-The, B.; van 284
der Saag, P. T. Pristanic acid and phytanic acid: naturally occurring ligands for the nuclear 285
receptor peroxisome proliferater-activated receptor alpha. J. Lipid Res. 2000, 41, 1801-286
1807. 287
(7) Grimaldi, P. A. Peroxisome Proliferator-Activated Receptors as sensors of fatty acids 288
and derivatives. Cell. Mol.Life Sci. 2007, 64, 2459-2464. 289
(8) Hellgren, L. I. Phytanic acid - an overlooked bioactive fatty acid in dairy fat? Ann. 290
N.Y. Acad. Sci. 2010, 1190 (1), 42-49. 291
(9) Eldjarn, L.; Try, K. Different Ratios of Ldd and Ddd Diastereoisomers of Phytanic 292
Acid in Patients with Refsums Disease. Biochim. Biophys. Acta 1968, 164 (1), 94-100. 293
(10) Tsai, S. C.; Steinber, D.; Avigan, J.; Fales, H. M. Studies on Stereospecificity of 294
Mitochondrial Oxidation of Phytanic Acid and of Alpha-Hydroxyphytanic Acid. J. Biol. 295
Chem. 1973, 248, 1091-1097. 296

B r i t a N g u m C h e P h D T h e s i s
166
(11) Heim, M.; Johnson, J.; Boess, F.; Bendik, I.; Weber, P.; Hunziker, W.; Fluhmann, B. 297
Phytanic acid, a natural peroxisome proliferator-activated receptor agonist, regulates 298
glucose metabolism in rat primary hepatocytes. FASEB J. 2002, 16, 718-720 299
(12) Ferdinandusse, S.; Rusch, H.; van Lint, A. E. M.; Dacremont, G.; Wanders, R. J. A.; 300
Vreken, P. Stereochemistry of the peroxisomal branched-chain fatty acid +¦- and +¦-301
oxidation systems in patients suffering from different peroxisomal disorders. J. Lipid Res. 302
2002, 43, 438-444. 303
(13) Schroder, M.; Vetter, W. GC/EI-MS determination of the diastereomer distribution of 304
phytanic acid in food samples. J. Am. Oil Chem. Soc. 2011, 88, 341-349. 305
(14) Schroder, M.; Lutz, N. L.; Tangwan, E. C.; Hajazimi, E.; Vetter, W. Phytanic acid 306
concentrations and diastereomer ratios in milk fat during changes in the cow's feed from 307
concentrate to hay and back. Eur. Food Res. Technol. 2012, 234, 955-962. 308
(15) Butler, G.; Nielsen, J. H.; Larsen, M. K.; Rehberger, B.; Stergiadis, S.; Canever, A.; 309
Leifert, C. The effects of dairy management and processing on quality characteristics of 310
milk and dairy products. NJAS - Wageningen J. Life Sci. 2011, 58 (3-4), 97-102. 311
(16) Council Regulation (EC) No 834/2007 of 28 June 2007 on organic production and 312
labelling of organic products and repealing Regulation (EEC) No 2092/91. http://eur-lex. 313
europa. eu/LexUriServ/LexUriServ. do?uri=OJ:L:2007:189:0001:0023:EN:PDF 2007. 314
(17) Sjaunja, L. O.; Baevre, L.; Junkkarinen, L.; Pedersen, J.; Setala, J. A Nordic Proposal 315
for An Energy Corrected Milk (Ecm) Formula. EAAP European Association for Animal 316
Production Publication 1991, 50, 156-157. 317
(18) Hvelplund, T.; Weisbjerg, M. R.; Lund, P. Energy and protein evaluation systems 318
used for dairy cows in Denmark. Proc. Soc. Nutr. Physiol. 2007, 16, 129-131. 319
(19) Macoon, B.; Sollenberger, L. E.; Moore, J. E.; Staples, C. R.; Fike, J. H.; Portier, K. M. 320
Comparison of three techniques for estimating the forage intake of lactating dairy cows on 321
pasture. J. Anim. Sci. 2003, 81, 2357-2366. 322

B r i t a N g u m C h e P h D T h e s i s
167
(20) Schroder, M.; Yousefi, F.; Vetter, W. Investigating the day-to-day variations of 323
potential marker fatty acids for organic milk in milk from conventionally and organically 324
raised cows. Eur. Food Res. Technol. 2011, 232, 167-174. 325
(21) Leiber, F.; Kreuzer, M.; Nigg, D.; Wettstein, H. R.; Scheeder, M. R. L. A study on the 326
causes for the elevated n-3 fatty acids in cows' milk of alpine origin. Lipids 2005, 40, 191-327
202. 328
(22) Lee, M. R. F.; Huws, S. A.; Scollan, N. D.; Dewhurst, R. J. Effects of fatty acid 329
oxidation products (green odor) on rumen bacterial populations and lipid metabolism in 330
vitro. J. Dairy Sci. 2007, 90, 3874-3882. 331
(23) Harfoot, C. G.; Hazlewood, G. P. Lipid metabolism in the rumen. In The rumen 332
microbial ecosystem, Hobson, P. N.; Stewart, C. S., Eds.; Chapman & Hall: London, UK, 333
1997; pp 382-426. 334
(24) Hanhoff, T.; Benjamin, S.; Borchers, T.; Spener, F. Branched-chain fatty acids as 335
activators of peroxisome proliferator-activated receptors. Eur. J. Lipid Sci.Technol. 2005, 336
107, 716-729 337
338

B r i t a N g u m C h e P h D T h e s i s
168
Table 1: Milk production, milk content of fat and protein, and feed consumption and composition of five herds in May and
September.
Period May September
Herd DH1 DH2 DH3 DJ1 DJ2 DH1 DH2 DH3 DJ1 DJ2
ECM kg cow-1day-1 29.4 29.8 27.1 22.3 26.5 30.9 26.7 25.6 21.6 23.1
Fat g kg milk-1 39.0 38.4 38.5 58.0 55.5 42.1 35.1 40.2 56.8 51.7
Protein g kg milk-1 32.7 32.1 33.0 41.0 40.0 34.7 32.6 36.1 41.1 38.1
Feed dry matter intake (DMI) kg
day-1
20.0 19.2 19.6 15.7 17.7 20.5 20.2 19.8 15.8 16.8
Feed composition kg kg DMI-1
Total pasture 0.47 0.76 0.52 0.66 0.33 0.42 0.21 0.28 0.43 0.29
Grazed grass 0.33 0.50 0.29 0.46 0.20 0.17 0.09 0.13 0.25 0.17
Grazed legumes 0.11 0.20 0.18 0.14 0.01 0.23 0.11 0.13 0.16 0.09
Grazed other species 0.03 0.06 0.05 0.06 0.12 0.02 0.01 0.02 0.02 0.03

B r i t a N g u m C h e P h D T h e s i s
169
Maize silage 0.23 0.06 0.13 0.00 0.15 0.17 0.48 0.20 0.00 0.11
Grass silage, hay and straw 0.04 0.07 0.06 0.14 0.30 0.17 0.00 0.19 0.23 0.31
Total concentrates 0.25 0.10 0.30 0.19 0.22 0.24 0.31 0.32 0.34 0.29

Brita Ngum Che PhD Thesis 2012
170
Table 2: Pearson correlation coefficients and corresponding p-values for the relations
between total relative amount of DMI of individual feed groups, and content of total PA in
milk-fat and share of RRR isomer of PA, respectively.
Total PA in milk fat Share of RRR isomer in
total PA
Correlation
coefficient p-value
Correlation
coefficient p-value
Feed
Total pasture -0.41 0.019 0.33 0.060
Grazed grass -0.42 0.014 0.11 0.526
Grazed legumes 0.07 0.697 0.85 <0.001
Grazed other species -0.52 0.002 -0.32 0.070
Maize silage 0.08 0.650 -0.28 0.110
Grass silage and hay
and straw
0.15 0.401 -0.02 0.898
Total concentrates 0.58 <0.001 -0.11 0.529

Brita Ngum Che PhD Thesis 2012
171

Brita Ngum Che PhD Thesis 2012
172
Figure 1: GC-MS-SIM chromatograms (m/z 101) of methyl esters of a sythetic phytanic
acid standard and a milk sample.
Elution time [min]
46.1 46.2 46.3 46.4 46.5 46.6 46.7
Ab
un
da
nc
e m
/z1
01
0
1e+5
2e+5
3e+5
4e+5
5e+5
standard
milk
SRR
RRR

Brita Ngum Che PhD Thesis 2012
173
Herd
DH1 DH2 DH3 DJ1 DJ2
To
tal
ph
yta
nic
ac
id i
n m
ilk
fa
t [m
g/g
]
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
May
September
bc bc
bc
b
bc
a
a
c
bc bc
Figure 2: Total content of phytanic acid in milk fat of samples from five herds in May and
September. Error bars denote standard deviations and different letters above columns
indicate significant differences (p < 0.05).

Brita Ngum Che PhD Thesis 2012
174
Herd
DH1 DH2 DH3 DJ1 DJ2
RR
R p
hyta
nic
ac
id i
n t
ota
l p
hyta
nic
ac
id [
mg
/mg
]
0.0
0.1
0.2
0.3
0.4
0.5
0.6
May
September
a
b
f g
c
d
h
e
f f
Figure 3: Share of RRR phytanic acid in total phytanic acid in milk fat of samples from
five herds in May and September. Error bars denote standard deviations and different
letters above columns indicate significant differneces (p < 0.05).

Brita Ngum Che PhD Thesis 2012
175
Share of grazed legumes in dry matter intake
0.00 0.05 0.10 0.15 0.20 0.25
Sh
are
of
RR
R i
so
me
r in
to
tal
ph
yta
nic
ac
id
0.15
0.20
0.25
0.30
0.35
0.40
0.45
0.50
0.55
Figure 4: Relations between share of grazed legumes in dry matter intake and share of
RRR phytanic acid in total phytanic acid in milk fat.
Related Documents











