Original article Haeme oxygenase activity protects the host against excessive cardiac inflammation during experimental Trypanosoma cruzi infection Fredy R.S. Gutierrez a, * ,1 , Wander R. Pavanelli e,b,1 , Tiago S. Medina b , Grace K. Silva b , Fla ´via S. Mariano b , Paulo M.M. Guedes f,b , Tiago W.P. Mineo g,b , Marcos A. Rossi c , Fernando Q. Cunha d , Joa ˜o S. Silva b a Biomedical Sciences Research Group, School of Medicine, Universidad Antonio Narin˜o, Bogota´, Colombia b Department of BiochemistryeImmunology, Ribeira˜o Preto School of Medicine, University of Sa˜o Paulo, Brazil c Department of Pathology, Ribeira˜o Preto School of Medicine, University of Sa˜o Paulo, Brazil d Department of Pharmacology, Ribeira˜o Preto School of Medicine, University of Sa˜o Paulo, Brazil e Centro de Cieˆncias Biolo´gicas, Departamento de Cieˆncias Patolo´gicas, Universidade Estadual de Londrina, Brazil f Departamento de Microbiologia e Parasitologia, Universidade Federal do Rio Grande do Norte, Natal, Brazil g Instituto de Cieˆncias Biome´dicas, Universidade Federal de Uberlaˆndia, Brazil Received 5 September 2012; accepted 4 October 2013 Abstract The infection with Trypanosoma cruzi induces a robust cardiac inflammation that plays a pathogenic role in the development of Chagas heart disease. In this study, we aimed at investigating the effects of Haem Oxygenase (HO) during experimental infection by T. cruzi in BALB/c and C57BL/6 mice. HO has recently emerged as a key factor modulating the immune response in diverse models of inflammatory diseases. In mice with two different genetic backgrounds, the pharmacologic inhibition of HO activity with zinc-protoporphyrin IX (ZnPPIX) induced enhanced myocarditis and reduced parasitaemia, which was accompanied by an amplified production of nitric oxide and increased influx of CD4 þ , CD8 þ and IFN-g þ cells to the myocardium in comparison with the control group. Conversely, treatment with haemin (an activator of HO) lead to a decreased number of intracardiac CD4 þ (but not CD8 þ ) cells compared to the control group. The mechanism involved in these observations is a modulation of the induction of regulatory T cells, because the stimulation or inhibition of HO was parallelled by an enhanced or reduced frequency of regulatory T cells, respectively. Hence, HO may be involved in the regulation of heart tissue inflammation and could be a potential target in conceiving future therapeutic approaches for Chagas disease. Ó 2013 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. Keywords: Trypanosoma cruzi; Haeme oxygenase; Regulatory T cells 1. Introduction Chagas disease is caused by the infection with the protozoan parasite Trypanosoma cruzi. It affects about 11 million people in Latin America and is emerging as an important public health problem in countries considered not endemic, mostly due to human migration and transmission through transplants or congenital infection. Among infected people, around 30% eventually develop a progressive form of myocardiopathy characterised by presence of heart fibrosis, cardiac hypertrophy, thromboembolism and conduction system abnormalities [1,2]. A well-established model for the study of immunopa- thology of Chagas disease is the experimental infection of mice, which leads to an intense myocarditis composed pri- marily of mononuclear cells. This inflammatory infiltration of the host cardiac tissues is critical for elimination of the * Corresponding author. Universidad Antonio Narin ˜o, Facultad de Medicina, Carrera 3 este No. 47 A e 15, Bogota ´, Colombia. Tel.: þ57 01 3204045. E-mail address: [email protected] (F.R.S. Gutierrez). 1 These two authors contributed equally to this work and they must be considered as co-first authors. Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase activity protects the host against excessive cardiac inflammation during experi- mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.doi.org/10.1016/j.micinf.2013.10.007 Microbes and Infection xx (2013) 1e12 www.elsevier.com/locate/micinf + MODEL 1286-4579/$ - see front matter Ó 2013 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. http://dx.doi.org/10.1016/j.micinf.2013.10.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

+ MODEL
Microbes and Infection xx (2013) 1e12www.elsevier.com/locate/micinf
Original article
Haeme oxygenase activity protects the host against excessive cardiacinflammation during experimental Trypanosoma cruzi infection
Fredy R.S. Gutierrez a,*,1, Wander R. Pavanelli e,b,1, Tiago S. Medina b, Grace K. Silva b,Flavia S. Mariano b, Paulo M.M. Guedes f,b, Tiago W.P. Mineo g,b, Marcos A. Rossi c,
Fernando Q. Cunha d, Joao S. Silva b
aBiomedical Sciences Research Group, School of Medicine, Universidad Antonio Narino, Bogota, ColombiabDepartment of BiochemistryeImmunology, Ribeirao Preto School of Medicine, University of Sao Paulo, Brazil
cDepartment of Pathology, Ribeirao Preto School of Medicine, University of Sao Paulo, BrazildDepartment of Pharmacology, Ribeirao Preto School of Medicine, University of Sao Paulo, Brazil
eCentro de Ciencias Biologicas, Departamento de Ciencias Patologicas, Universidade Estadual de Londrina, BrazilfDepartamento de Microbiologia e Parasitologia, Universidade Federal do Rio Grande do Norte, Natal, Brazil
g Instituto de Ciencias Biomedicas, Universidade Federal de Uberlandia, Brazil
Received 5 September 2012; accepted 4 October 2013
Abstract
The infection with Trypanosoma cruzi induces a robust cardiac inflammation that plays a pathogenic role in the development of Chagas heartdisease. In this study, we aimed at investigating the effects of Haem Oxygenase (HO) during experimental infection by T. cruzi in BALB/c andC57BL/6 mice. HO has recently emerged as a key factor modulating the immune response in diverse models of inflammatory diseases. In micewith two different genetic backgrounds, the pharmacologic inhibition of HO activity with zinc-protoporphyrin IX (ZnPPIX) induced enhancedmyocarditis and reduced parasitaemia, which was accompanied by an amplified production of nitric oxide and increased influx of CD4þ, CD8þ
and IFN-gþ cells to the myocardium in comparison with the control group. Conversely, treatment with haemin (an activator of HO) lead to adecreased number of intracardiac CD4þ (but not CD8þ) cells compared to the control group. The mechanism involved in these observations is amodulation of the induction of regulatory T cells, because the stimulation or inhibition of HO was parallelled by an enhanced or reducedfrequency of regulatory T cells, respectively. Hence, HO may be involved in the regulation of heart tissue inflammation and could be a potentialtarget in conceiving future therapeutic approaches for Chagas disease.� 2013 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved.
Keywords: Trypanosoma cruzi; Haeme oxygenase; Regulatory T cells
1. Introduction
Chagas disease is caused by the infection with the protozoanparasite Trypanosoma cruzi. It affects about 11million people inLatin America and is emerging as an important public health
* Corresponding author. Universidad Antonio Narino, Facultad de Medicina,
Carrera 3 este No. 47 A e 15, Bogota, Colombia. Tel.: þ57 01 3204045.
E-mail address: [email protected] (F.R.S. Gutierrez).1 These two authors contributed equally to this work and they must be
considered as co-first authors.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
1286-4579/$ - see front matter � 2013 Institut Pasteur. Published by Elsevier Ma
http://dx.doi.org/10.1016/j.micinf.2013.10.007
problem in countries considered not endemic, mostly due tohuman migration and transmission through transplants orcongenital infection. Among infected people, around 30%eventually develop a progressive form of myocardiopathycharacterised by presence of heart fibrosis, cardiac hypertrophy,thromboembolism and conduction system abnormalities [1,2].
A well-established model for the study of immunopa-thology of Chagas disease is the experimental infection ofmice, which leads to an intense myocarditis composed pri-marily of mononuclear cells. This inflammatory infiltration ofthe host cardiac tissues is critical for elimination of the
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007
sson SAS. All rights reserved.

2 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
parasite and is modulated by the orchestrated expression ofchemokine receptors on the surface of activated leukocytes inresponse to pro-inflammatory cytokines, primarily IFN-g andTNF-a produced by cells of the innate immune system and bycardiomyocytes [3,4]. The continued heart inflammation in-duces excessive collateral tissue damage that is caused by type1 effector T cells [3,5,6].
Although this inflammatory response is originally triggeredby the presence of the parasite, the pathogen is scarcely found,even in intensely inflamed heart tissues, leading some re-searchers to postulate that the exacerbated immune response isdue to loss of tolerance or autoimmune phenomena [7,8].Thus, understanding the mechanisms involved in the control ofthe cardiac inflammatory immune response elicited by T. cruzimay be crucial to define strategies aimed at ameliorating theresulting heart disease.
Haeme Oxygenase (HO) catalyse the degradation of hae-moglobin by cleaving the Haeme ring at the alpha methenebridge, producing biliverdin, iron, and carbon monoxide (CO),which are important endogenous anti-inflammatory mediators[9,10]. Three isoforms of HO have been identified. While HO-2 and HO-3 are constitutively expressed [11,12], the HO-1isoform is inducible [13] and is expressed under conditionsof oxidative stress, hypoxia, and intoxication by heavy metals,in response to cytokines [14,15], and in the presence of itssubstrate [16].
The activity of HO-1 has been linked to arrested macro-phage activation, reduced leukocyte migration and inflamma-tory cytokine production [17,18]. These anti-inflammatory,anti-apoptotic, and anti-lymphocyte proliferative effects ofHO-1 have been attributed to byproduct of the breakdown ofhaeme group [18e20]. In models of cardiac allograft andxenograft rejection, HO-1 protects the host from collateraltissue damage in a manner that is dependent on the presence ofCD4þCD25þ T cells [21,22]. Indeed, the pharmacologicalinduction of HO has anti-inflammatory properties, while itsinhibition leads to exacerbate inflammation [23].
One interesting mechanism by which HO-1 may regulateinflammation is through the induction of regulatory T cell(Treg) activity [24e27]. The presence of HO-1 increasesserum IL-10 and CD4þCD25þ Tregs in peripheral blood.Moreover, the transcription factor Foxp3 induces HO-1expression, suggesting that HO-1 may be one importantmechanism of Treg-mediated immune modulation [24].
In the present study, the effects of HO on the immuneresponse during experimental infection with T. cruzi weretested using pharmacological agonists and antagonists of HOactivity and it was found that this enzyme plays an importantrole in the modulation of inflammatory response at the heart.
2. Materials and methods
2.1. Animals
All the experimental protocols were reviewed and approvedby the committee on Ethics for Animal Care at the School ofMedicine in Ribeirao Preto, Universidade de Sao Paulo.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
Female BALB/c mice aged 6e8 weeks were bred and main-tained in the animal housing facility of the Department ofBiochemistry and Immunology. For survival and parasitaemiastudies, two independent groups of five animals were treatedintraperitoneally with either zinc-protoporphyrin IX(ZnPPIX), haemin or vehicle solution were followed for 40days after infection.
2.2. Treatments
Animals were treated daily with 5 mg/kg of ZnPPIX (HO-1inhibitor), 5 mg/kg of haemin (HO Inductor), or saline (controlgroup). The medications were purchased from PorphyrinProducts, Logan, UT, and diluted in saline. Treatment started1 h after infection with T. cruzi and went until 20 days p.i.
2.3. Parasites, experimental infection, and cardiacparasitism
Mice were infected via intraperitoneal injection (i.p.) with103 bloodstream forms of T. cruzi (Y strain) obtained from anintermediary strain-matched infected mouse. Parasitaemia wasevaluated in 5 ml of blood drawn from the tail vein [28].Before infection of intermediary mice, parasites were grown inand purified from the LLC-MK2 (ATCC) cell line. The cardiacparasitism of BAL/c and C57BL/6 mice was assessed by real-time PCR as previously described [29].
2.4. Measurement of bilirubin (activity of HO)
Bilirubin was measured in 50 ml of serum with a colouri-metric reaction, according to the manufacturer’s instructions(Labtest, Brazil). Reactions were read at 540 nm, and the re-sults were expressed as mg of bilirubin per ml of serum.
2.5. Measurement of liver enzymes
Blood samples were collected and immediately centrifugedat 4 �C. Enzyme activities of serum ALT and AST weredetermined by standard clinical tests according to the manu-facturer’s instructions (Roche, Switzerland), and the resultswere expressed in international units per ml.
2.6. Histologic analysis
Groups of five mice were euthanised after 10, 15, and 20days of infection. The heart, liver, and skeletal muscle werefixed in 10% formaldehyde in PBS, embedded in paraffin,sectioned, stained with haematoxylin eosin, and examined bylight microscopy.
2.7. Drug cytotoxicity and intracellular parasitism
Cultures of BMM were infected with trypomastigotes at amultiplicity of infection (MOI) of 5:1 for 2 h and treated withHaemin or ZnPPIX for 8 h. Drug cytotoxicity was assessedwith the CytoTox assay (Promega, USA) in infected or not
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

3F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
infected bone marrow-derived macrophages (BMM) [30], aswell as in T. cruzi trypomastigotes.
The cells were then stained with Giemsa, and the numbersof intracellular amastigotes were counted in 100 infected cellsin the optical microscope at a final magnification of 1000�(oil immersion), as previously described [31].
2.8. Cytokine assays
Cytokine concentrations in tissue homogenates of the heart,liver, and spleen were measured by ELISA. TGF-b, IFN-g, IL-10 (OpTEIA, BD Biosciences, USA), IL-4, and TNF-a(Duoset, R&D Systems, USA) were assayed following themanufacturer’s instructions. The reaction was visualised withperoxidase-conjugated streptavidin (Vector Laboratories,Burlingame, CA, USA) followed by a substrate containingTMB (Promega, USA) as a chromogen. The optical densities(O.D.) of the samples were then read at 450 nm, and thecytokine concentrations were calculated by extrapolating theO.D. obtained from a standard curve of each recombinantcytokine.
2.9. Nitric oxide quantification
Blood was obtained from the retro-orbital plexus of treatedor control mice at 10, 15, and 20 days after infection. Theserum was then separated and the samples treated with nitratereductase, following by nitrite concentration measurement bythe Griess method as previously described [32]. Briefly, 100 mlof reductase-treated serum was mixed with 100 ml of Griessreagent in a multi-well plate, and the absorbance at 550 nmwas read 10 min later. The NO2 concentration was determinedby reference to a standard curve of NaNO2 (1e200 mM).
2.10. Immunohistochemistry
Hearts removed from euthanised mice were embedded inthe OTC medium (Tissue-Tek, USA) and stored in liquid N2.Immunohistochemistry was performed in different frozen tis-sue sections with each of the following antibodies: rabbit IgGanti-mouse NOs2, rat anti-mouse CD8, rat anti-mouse CD4,goat anti-mouse IFN-g, and rabbit polyclonal serum anti-T.cruzi. All antibodies were diluted in phosphate-buffered salinecontaining 3% (w/v) non-fat dry milk. The staining wasvisualised using the avidin-peroxidase complex, 3, 30-dia-minobenzidine (Vector, Canada), and counterstaining withMayer’s haematoxylin.
A semi-quantitative analysis was then performed in T. cruziimmunostained tissues as follows: For CD4, CD8, and IFN-g,the number of stained cells in a fixed area of 50 mm2 wascounted; to quantify tissue parasitism, the number of parasitenests (considered as clusters of positive staining) observed ineach microscopic field at a magnification of 20� was used. Ineach animal, 200 different microscopic fields were counted,using three different nonconsecutive slides of tissues.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
2.11. Flow cytometry analysis
Cell phenotyping was performed on spleen cells from fourBALB/c mice belonging to each experimental group at daysten, fifteen, and twenty p.i. The following conjugated anti-bodies were used: anti-CD4-PerCP, anti-CD25-FITC, anti-PD-1-PE, anti-CTLA-4-PE, anti-Foxp-3-PE (BD-Pharmingen),and isotype-matched control mAbs. Cell viability wasconfirmed by negative staining with 0.2% trypan blue, and200,000 events were collected in a viable lymphocyte-enriched gate defined according to forward and side scatterparameters. Data acquisition was performed using a FACScanflow cytometer with Cell-Quest software (BectoneDickinsonUSA). The appropriate isotype-matched unrelated rat IgGmAbs were used as negative controls. Data analysis was per-formed with Cell-Quest software.
2.12. Foxp3 and IL-10 mRNA expression
Hearts from BALB/c or C57BL/6 mice were collected inTRIzol reagent (Invitrogen) 15 days after T. cruzi infection.The mRNA was extracted using the SV Total RNA IsolationSystem (Promega) according to the manufacturer’s in-structions, and cDNA was synthesised using 1 mg of RNAthrough a High-capacity cDNA Reverse Transcription Kit(Applied Biosystems, USA). The standard PCR conditionswere 50 �C (2 min) and 95 �C (10 min), followed by 40 cyclesof 94 �C (15 s), 58 �C (30 s) and 72 �C (30 s), with a standarddenaturation curve. The mean Ct values of duplicate cDNAsamples were used to determine the expression of the targetgene based on an internal control (GAPDH), which was usedto normalise the reaction.
2.13. Statistical analysis
The results are expressed as the means � SEM of threeindependent experiments. Two-way ANOVA followed by theBonferroni post-test was used to compare the three experi-mental groups at the two time points analysed. The log-rank(ManteleCox) test followed by the log-rank test for trendwas used to compare survival curves among the three exper-imental groups. In the case of Treg phenotyping analyses, one-way ANOVA was used, followed by Dunnett’s multiplecomparison tests. The statistical significance represented in thefigures was standardised as follows: n.s. for not significant, or* for p < 0.05.
3. Results
3.1. Treatment with ZnPPIX or haemin modulates theactivity of HO-1
We first aimed at validating the treatments with ZnPPIX orhaemin as an inhibitor and inducer of HO-1 activity, respec-tively, during T. cruzi infection. Hence, serum bilirubin wasmeasured at different time points after infection in animalsreceiving each treatment. As shown in Fig. 1A, the infection
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007
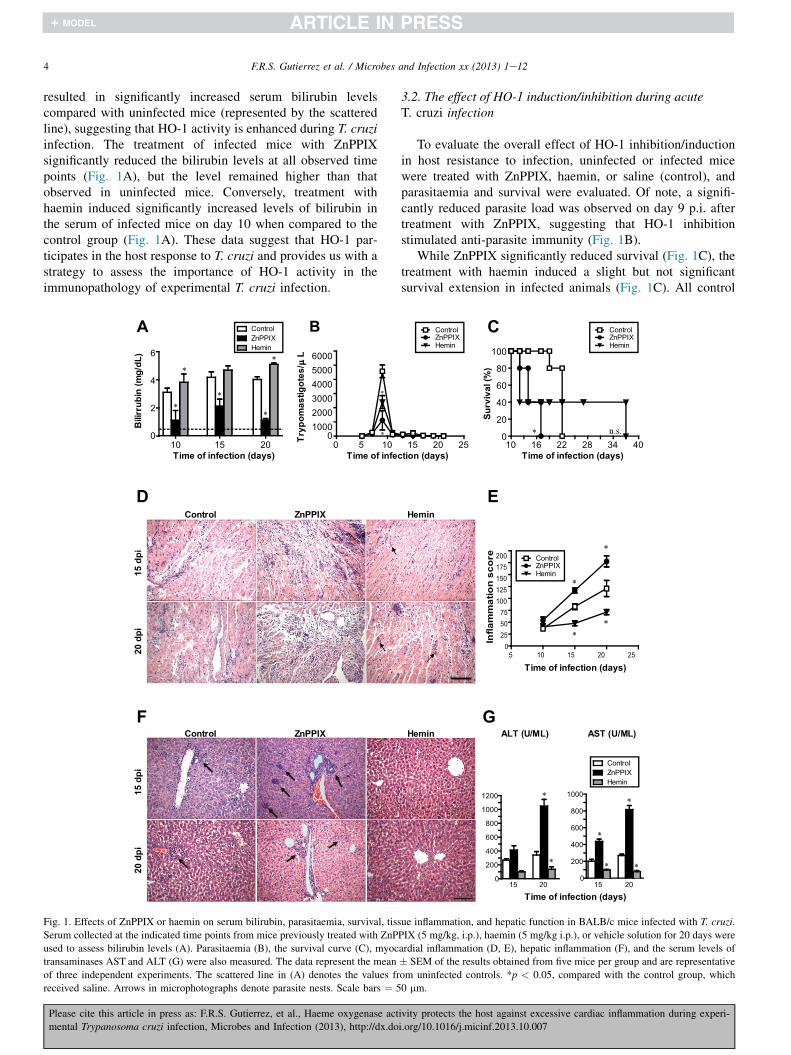
4 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
resulted in significantly increased serum bilirubin levelscompared with uninfected mice (represented by the scatteredline), suggesting that HO-1 activity is enhanced during T. cruziinfection. The treatment of infected mice with ZnPPIXsignificantly reduced the bilirubin levels at all observed timepoints (Fig. 1A), but the level remained higher than thatobserved in uninfected mice. Conversely, treatment withhaemin induced significantly increased levels of bilirubin inthe serum of infected mice on day 10 when compared to thecontrol group (Fig. 1A). These data suggest that HO-1 par-ticipates in the host response to T. cruzi and provides us with astrategy to assess the importance of HO-1 activity in theimmunopathology of experimental T. cruzi infection.
Fig. 1. Effects of ZnPPIX or haemin on serum bilirubin, parasitaemia, survival, tiss
Serum collected at the indicated time points from mice previously treated with ZnP
used to assess bilirubin levels (A). Parasitaemia (B), the survival curve (C), myoc
transaminases AST and ALT (G) were also measured. The data represent the mean
of three independent experiments. The scattered line in (A) denotes the values fr
received saline. Arrows in microphotographs denote parasite nests. Scale bars ¼ 5
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
3.2. The effect of HO-1 induction/inhibition during acuteT. cruzi infection
To evaluate the overall effect of HO-1 inhibition/inductionin host resistance to infection, uninfected or infected micewere treated with ZnPPIX, haemin, or saline (control), andparasitaemia and survival were evaluated. Of note, a signifi-cantly reduced parasite load was observed on day 9 p.i. aftertreatment with ZnPPIX, suggesting that HO-1 inhibitionstimulated anti-parasite immunity (Fig. 1B).
While ZnPPIX significantly reduced survival (Fig. 1C), thetreatment with haemin induced a slight but not significantsurvival extension in infected animals (Fig. 1C). All control
ue inflammation, and hepatic function in BALB/c mice infected with T. cruzi.
PIX (5 mg/kg, i.p.), haemin (5 mg/kg i.p.), or vehicle solution for 20 days were
ardial inflammation (D, E), hepatic inflammation (F), and the serum levels of
� SEM of the results obtained from five mice per group and are representative
om uninfected controls. *p < 0.05, compared with the control group, which
0 mm.
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

5F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
infected mice died on day 22 p.i. These data strongly suggestthat HO-1 activity modulates host survival during the acuteinfection. As HO-1 activity is known to regulate the inflam-matory response primarily by inducing the production of anti-inflammatory cytokines such as IL-10 [33,34], we next eval-uated the inflammatory response in the heart, skeletal muscle,and liver tissues of T. cruzi-infected mice. All infected animalsshowed an inflammatory response in the myocardium andliver, characterised by the diffuse infiltration of mononuclearcells and the sporadic presence of parasite nests. Interestingly,at 15 and 20 days p.i., the inflammatory reaction was signifi-cantly increased in the hearts of mice receiving ZnPPIX andreduced in the hearts of mice receiving haemin whencompared to the group receiving saline (Fig. 1D, E). Similarresults were observed for liver inflammatory damage (Fig. 1F),which reflected the levels of aminotransferases in plasma, asshown in Fig. 1G for ALT and AST, demonstrating that HO-1inhibition favours the establishment of a robust inflammatoryresponse in the liver and the myocardium during the acutephase of T. cruzi infection.
Pertinently, the treatment with ZnPPIX or Haemin did notinduce myocarditis in the uninfected mice. In addition, treat-ment of uninfected mice with haemin or ZnPPIX did not causedeath or alter weight body, even at 40 days after treatment (datanot shown). Moreover, ZnPPIX and haemin had no cytotoxiceffect on host cells, as verified by LDH release by macrophages(using Triton X-100 as a positive control), and these drugs didnot impair macrophage invasion of the cardiac tissues (Fig. S4).
Additional immunohistochemical examinations of heartsections revealed a significantly increased number of CD4(Fig. S1) and CD8 cells (Fig. S2) in the ZnPPIX groupcompared to the haemin or control groups, whereas signifi-cantly reduced numbers of CD4 (but not CD8) cells wereobserved after treatment with haemin (Fig. S1). By contrast,the production of IFN-g was significantly increased only in theZnPPIX group on day 20 p.i. (Fig. S3), suggesting that theHO-1 induction/blockade has a significant effect on the in-flammatory response during acute T. cruzi infection.
Fig. 2. Heart tissue parasitism of BALB/c mice treated with ZnPPIX or haemin. A, R
T. cruzi antigens on days 15 and 20 p.i. in heart tissues from BALB/c mice treated
nests. The data represent the mean � SEM of five mice per group and three indepen
saline. Scale bars ¼ 50 mm. C, Quantification of T. cruzi DNA by real-time PCR of
five mice per group and three independent experiments. *p < 0.05, compared wit
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
3.3. The inhibition of HO-1 leads to increase NOproduction and controls the cardiac parasitism in thehearts of T. cruzi-infected mice
To evaluate the impact of HO-1 activity on tissue para-sitism, immunohistochemical analyses were performed, andthe parasitism was quantified in the cardiac tissue of T. cruzi-infected mice in each experimental group. As expected fromour above results and previous reports [35], the infected micetreated with haemin e but not those receiving ZNPPIX,exhibited significantly increased numbers of parasite nests andDNAwhen compared to the control group (Fig. 2). To addresswhether haemin or ZnPPIX may have any cytotoxic effect onT. cruzi, we treated parasite trypomastigotes in vitro withZnPPIX or Haemin, and found that these drugs did not modifythe parasites motility (Fig. S4).
Mice treated with ZnPPIX showed increased levels of NOin the serum on days 15 and 20 p.i. compared to the controlgroup (control vs. ZnPPIX: 92.29 vs. 148.8 mM and 51.67 vs.82.99 mM, p < 0.05, on 15 and 20 days p.i., respectively,Fig. 5). Moreover, the treatment with haemin significantlyreduced the levels of NO in the serum of mice on day 15 p.i.when compared to the control group ( p < 0.05, Fig. 5). Theseresults demonstrate that NO is involved in the modulation ofthe inflammatory response by HO-1 during acute T. cruziinfection.
3.4. HO-1 activity defines the host cytokine profile andtissue parasitism during T. cruzi infection
The inhibition of HO-1 by ZnPPIX resulted in significantlyincreased production of IFN-g and TNF-a in the heart andspleen of infected mice on day 20 p.i. (Fig. 3). In the livers ofZnPPIX-treated mice, TNF-a was significantly increased onday 15 p.i. Surprisingly, reduced levels of IFN-g were detectedin the livers of animals receiving either ZnPPIX or haemin ondays 15 and 20 p.i. This may be a result of either the increasedliver parasitism for ZnPPIX or the reduced liver damage
epresentative photomicrography of immunohistochemistry for the detection of
with ZnPPIX, haemin, or vehicle. B, Quantification of the number of parasite
dent experiments. *p < 0.05, compared with the control group, which received
heart tissues. The data represent the mean � SEM of the results obtained from
h the control group, which received saline. Scale bars ¼ 50 mm.
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

IFN-γγ
0
50
100
150
200
250
Liv
er
0
50
100
150
200
250
ρg/m
L
10 15 200
100
200
300
400
TNF- α
0
500
1000
1500 ZnPPIXControl
Hemin
0
500
1000
1500
2000
2500
10 15 200
500
1000
1500
Time (days)
IL-10
0
1000
2000
3000
4000
5000
0
2000
4000
6000
8000
Sp
lee
n
10 15 200
2000
4000
6000
8000
TGF-β
0
2000
4000
6000
8000
10000
0
1000
2000
3000
4000
5000
6000
7000
10 15 200
1000
2000
3000
4000
5000
6000
He
art
*
*
*
*
*
*
*
**
*
*
*
n.s.
*
* *
*
*
n.s.
n.s.
*
*
*
*
*
**
* *
Fig. 3. Production of cytokines in tissues from T. cruzi-infected BALB/c mice treated with ZnPPIX or haemin. The concentrations of IFN-g, TNF-a, IL-10, and
TGF-b were evaluated by ELISA in tissue homogenates from BALB/c mice treated with ZnPPIX, haemin or vehicle solutions and euthanised at different time
points after infection with T. cruzi. The data represent the mean � SEM of five mice per group and three independent experiments. *p < 0.05, compared with the
control group, which received saline.
6 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
observed in liver after the treatment with haemin (Fig. 1Fand G).
In the spleen, ZnPPIX enhanced the levels of IFN-g andTNF-a on days 15 and 20 p.i., while haemin significantlyreduced the levels of IFN-g on day 10 p.i. and increased thelevels of TNF-a on day 15 p.i. (Fig. 3).
Moreover, the treatment with ZnPPIX significantly reducedthe levels of IL-10 in the hearts of infected mice on days 15and 20 p.i., while TGF-b was reduced only on day 20 p.i.Similar results were observed in the spleen for IL-10 and TGF-b. In the liver, ZnPPIX treatment was also effective inreducing the levels of IL-10 and TGF-b, although IL-10 wasnot significantly reduced on day 15 p.i.
Interestingly, animals that received haemin exhibitedsignificantly increased production of IL10 but not TGF-b inheart tissues on day 20 p.i. Again, a similar cytokine profilewas observed in the spleens of these mice. In the liver, nosignificant differences were found in the level of either cyto-kine in the tissues from haemin-treated animals compared tocontrols, except for a significant reduction of IL-10 on day15 p.i. (Fig. 3). These reduced levels of IL-10 in liver, againmay be reflecting a reduced inflammation in mice treated withhaemin, as shown in Fig. 1F. Taken together, These results
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
confirm our previous findings and provide strong evidence ofan immunoregulatory role of HO-1 during T. cruzi infection.
3.5. The presence of Tregs in cardiac tissue after T. cruziinfection is reduced by the blockade of HO-1
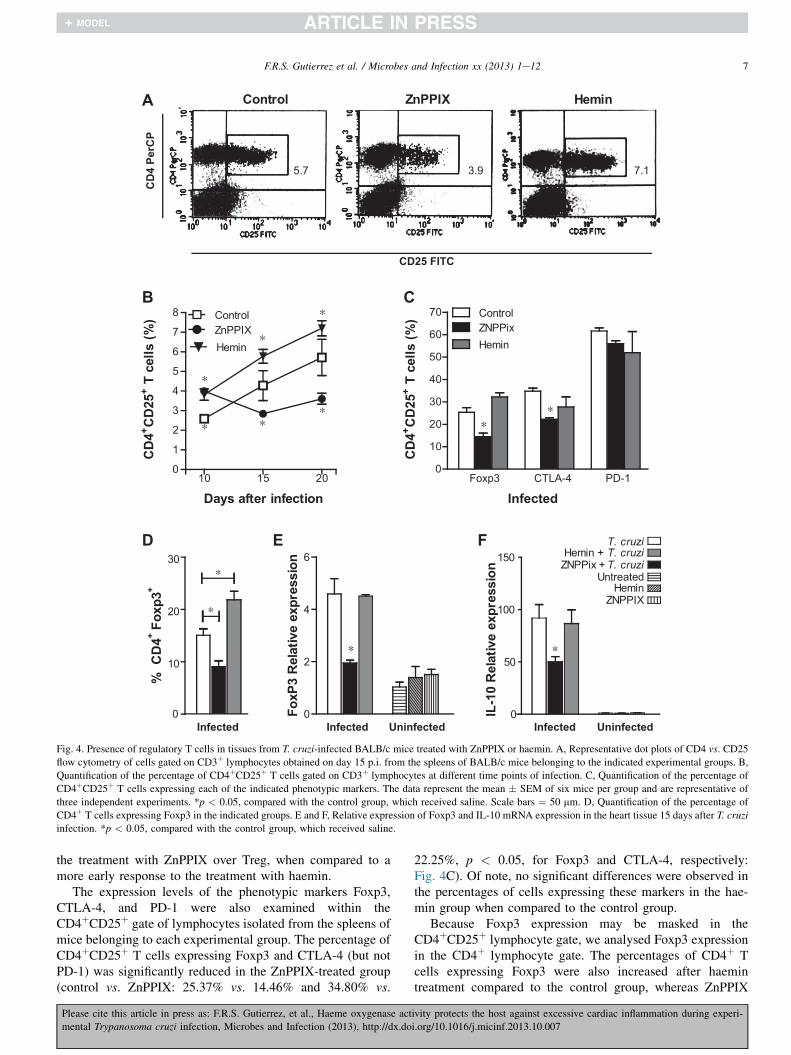
The effect of HO-1 inhibition/induction on the percentageof regulatory T cells (Tregs) from mice infected with T. cruziwas examined in lymphocytes extracted from the spleens of T.cruzi-infected BALB/c mice were analysed with a focus on theTreg subpopulation. The percentage of CD4þCD25þ cells wassignificantly reduced in mice treated with ZnPPIX comparedto the control group (control vs. ZnPPIX: 4.281% vs. 2.842%and 5.712% vs. 3.607%, p < 0.05, on days 15 and 20 p.i.,respectively, Fig. 4A and B). As expected, the percentages ofCD4þCD25þ cells were increased after treatment with haeminat all observed time points (control vs. haemin: 2.59% vs.3.818%; 4.28% vs. 5.772%, and 5.712% vs. 7.202%,p < 0.001, on days 10, 15 and 20 p.i., respectively, Fig. 4B).Of note, the percentages of Treg cells were increased on day10 p.i. in the group receiving ZnPPIX compared to the controlgroup (control vs. ZnPPIX: 2.593% vs. 3.998%, 95%,p < 0.001, Fig. 4B). This may reflect a delay in the effect of
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007
10 15 200
1
2
3
4
5
6
7
8ZnPPIXControl
Hemin
Days after infection
CD
4+C
D25
+ T
cells (%
)C
D4
P
erC
P
CD25 FITC
ZnPPIXControl Hemin
5.7 3.9 7.1
*
B
Foxp3 CTLA-4 PD-10
10
20
30
40
50
60
70 ControlZNPPixHemin
Infected
**
CD
4+C
D25
+ T
cells (%
)
C
A
***
*
*
0
2
4
6
UntreatedHemin
ZNPPIX
Hemin + T. cruzi
ZNPPix + T. cruzi
T. cruzi
*
Fo
xP
3 R
ela
tiv
e exp
ressio
n
E F
0
50
100
150
*
IL
-10 R
ela
tiv
e exp
ressio
n
0
10
20
30*
*
% C
D4
+
F
oxp
3 +
D
Infected InfectedInfected UninfectedUninfected
Fig. 4. Presence of regulatory T cells in tissues from T. cruzi-infected BALB/c mice treated with ZnPPIX or haemin. A, Representative dot plots of CD4 vs. CD25
flow cytometry of cells gated on CD3þ lymphocytes obtained on day 15 p.i. from the spleens of BALB/c mice belonging to the indicated experimental groups. B,
Quantification of the percentage of CD4þCD25þ T cells gated on CD3þ lymphocytes at different time points of infection. C, Quantification of the percentage of
CD4þCD25þ T cells expressing each of the indicated phenotypic markers. The data represent the mean � SEM of six mice per group and are representative of
three independent experiments. *p < 0.05, compared with the control group, which received saline. Scale bars ¼ 50 mm. D, Quantification of the percentage of
CD4þ T cells expressing Foxp3 in the indicated groups. E and F, Relative expression of Foxp3 and IL-10 mRNA expression in the heart tissue 15 days after T. cruzi
infection. *p < 0.05, compared with the control group, which received saline.
7F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
the treatment with ZnPPIX over Treg, when compared to amore early response to the treatment with haemin.
The expression levels of the phenotypic markers Foxp3,CTLA-4, and PD-1 were also examined within theCD4þCD25þ gate of lymphocytes isolated from the spleens ofmice belonging to each experimental group. The percentage ofCD4þCD25þ T cells expressing Foxp3 and CTLA-4 (but notPD-1) was significantly reduced in the ZnPPIX-treated group(control vs. ZnPPIX: 25.37% vs. 14.46% and 34.80% vs.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
22.25%, p < 0.05, for Foxp3 and CTLA-4, respectively:Fig. 4C). Of note, no significant differences were observed inthe percentages of cells expressing these markers in the hae-min group when compared to the control group.
Because Foxp3 expression may be masked in theCD4þCD25þ lymphocyte gate, we analysed Foxp3 expressionin the CD4þ lymphocyte gate. The percentages of CD4þ Tcells expressing Foxp3 were also increased after haemintreatment compared to the control group, whereas ZnPPIX
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

0
25
50
75
100
125
150
175
Days after infection
Seru
n n
itrit
e/n
itrate (μ
M) Control
ZNPPixHemin
10 2015
*
n.s.
*
*
Fig. 5. Production of nitric oxide in the sera of BALB/c mice treated with
ZnPPIX or haemin. BALB/c mice were treated with ZnPPIX, haemin, or saline
and euthanised on the indicated time points after infection with T. cruzi (10,
15, and 20 p.i.). The data represent the mean � SEM of the results obtained
from five mice per group and three independent experiments. *p < 0.05,
compared with the control group, which received saline. n.s.: Not significant.
8 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
treatment significantly reduced Foxp3 expression (Fig. 4D). Toconfirm these results, we analysed the expression of mRNAfor this transcription factor in the heart tissues 15 days after T.cruzi infection (Fig. 4E). Indeed, tissue Foxp3 mRNAexpression was reduced in the ZnPPIX-treated group but un-expectedly, remained unchanged in the mice treated withhaemin. This may suggest that the Tr1 type of Tregs (that do
6 8 10 12 140
2000
4000
6000
8000
Days after infection
**
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5 *
0
1
2
3 *IL-10 Foxp3
A
D E
Parasites/μL
Relative exp
ressio
n
ControlZnPPIXHemin
Control
HeminZnPPIX
Heart
Fig. 6. Effects of the HO-1 inhibition on T. cruzi-induced myocarditis in C57BL/6. C
treated with ZnPPix, haemin, or a vehicle solution during 12 d.p.i. The parasitaemia
to analyse the heart (B) and liver inflammation (E) using image J software. Slide
expression of Foxp3 and IL-10 mRNA transcripts by real-time PCR (D). *p < 0.
received saline.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
not express CD25 or foxp3) [36], may be participating in theregulation of the immune response in mice treated with hae-min, whereas Th3 type of Treg are directly modulated by thetreatment with ZnPPIX. Additionally, IL-10 mRNA expressionwas only decreased after ZnPPIX treatment, while haemintreatment did not alter IL-10 expression compared to thecontrol group (Fig. 4F), thus reinforcing that HO-1 modulatesTregs during T. cruzi infection.
3.6. The inhibition of HO-1 leads to enhancedmyocarditis in T. cruzi-infected C57BL/6 mice through areduction of Tregs
The previous findings HO-1 inhibition in BALB/c micewere also confirmed by using T. cruzi-infected C57BL/6 micethat were treated with haemin and ZnPPIX, and examined onday 14 p.i. Confirming our data from BALB/c mice, bothtreatments were able to reduce the parasitaemia during thecourse of the infection (Fig. 6A). Consistently, the increasedheart inflammatory index observed in ZnPPIX-treated mice(Fig. 6B) was critical to the control of cardiac parasitism onday 14 p.i. (Fig. 6C). In contrast, the treatment with haemin orZnPPIX did not promote liver inflammation in T. cruzi-infected C57BL/6 mice (Fig. 6E).
To understand whether the inhibition of HO-1 was able toreduce the expression of regulatory markers in the cardiactissue of C57BL/6 mice, the relative expression of Foxp3 and
0
5000
10000
15000 *
0
2000
4000
6000
0
2000
4000
6000
8000 *
B C
Nu
cleu
s co
un
ts /136 m
m2
T. cru
zi D
NA
(n
g)
Heart
Control ZnPPIX Hemin
Nu
cleu
s co
un
ts /136 m
m2
Liver
ControlZnPPIXHemin
57BL/6 mice (5 animals per group) were infected with Y strain of T. cruzi and
was evaluated until day 13 d.p.i. (A). The animals were euthanised (14 d.p.i.)
s of cardiac tissues were used to quantify the parasitism (C) and the relative
05 indicates a significant difference compared with the control group, which
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

9F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
IL-10 genes were analysed after T. cruzi infection. We found astriking reduction in the Foxp3 and IL-10 mRNA expressionafter ZnPPIX treatment 14 days p.i (Fig. 6D). These results arein agreement with that from BALB/c mice and are in supportof our hypothesis that the inhibition of HO-1 promotesmyocarditis by reducing the presence of Tregs in the hearttissues during experimental T. cruzi infection.
4. Discussion
The present study evaluated the effect of the specific inhi-bition or induction of HO-1 (with ZnPPIX or haemin,respectively) on the inflammatory response induced by T. cruziinfection. Experimental evidence is provided demonstratingthat HO-1 activity has an important immunomodulatory effectand is in part responsible for host protection from thedisproportionate inflammatory response during T. cruziinfection.
HO-1 represents an intrinsic anti-inflammatory systemdownstream of the cytokine IL-10, with the ability to regulateleukocyte function and restrain the expression of cytokines,primarily as a result of the production of carbon monoxide,one of the products of HO-1-mediated Haeme degradation[37]. We demonstrated that infection with T. cruzi promotesthe activation of this enzyme in vivo by significantlyincreasing the production of its main metabolite, bilirubin.
Biliverdin, which arises from free haeme catabolism byHO-1, is an anti-inflammatory molecule that is converted bybiliverdin reductase into bilirubin. Bilirubin is known as astrong anti-inflammatory, anti-oxidant molecule that protectscells against apoptosis during an oxidative stress response.Biliverdin and bilirubin are likely to protect against the in-flammatory response during several disease processes,including ischaemia and reperfusion injury, severe sepsis,vascular lesions, and the rejection of transplanted organs [52].
The reduced inflammatory response observed after haemintreatment can be related to the reduced activity of NOS2because nitric oxide (NO) is involved in anti-parasitic re-sponses during this infection [38]. In connection with this,mice treated with haemin showed reduced levels of serum NO.These findings are also in agreement with a previous studyusing experimental models of the inflammatory response eli-cited by zymosan where reduced NOS2 activity was observedafter HO-1 induction with haemin [39].
Through interaction with several mediators, NO can controlphysiologic responses directly or indirectly involved ininflammation [40]. This includes crosstalk between NO andHO-1-derived carbon monoxide, which has been previouslydocumented [41,42]. Hence, the immunomodulatory effectsobserved after HO-1 induction/inhibition in the present studymay be related to the following mechanisms: 1. Increasedlevels of CO are expected after HO-1 induction and have beenshown to be a potent anti-inflammatory factor [43]; 2. COdirectly binds to NOS2, inactivating it. 3. NOS2 (the majorsource of NO during T. cruzi infection) is a substrate of HO-1;therefore, increased HO-1 activity can accelerate NOS2degradation. In fact, our results showed an increased
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
production of NO in the sera of mice treated with ZNPPIX andreduced levels of this mediator in mice receiving haemin. 4.Because the active site of NOS2 requires two haeme mole-cules, an increase in the activity of HO accelerates thedegradation of haeme, reducing NOS2 activity [42]. 5. Thegeneration of free iron by haeme decomposition may inhibitthe production of NOS2 by suppressing its nucleartranscription.
The most important finding of the present study is that theeffects of inhibition/induction of HO-1 are coupled to theexpression of Foxp3 in heart tissues in the experimental modelof T. cruzi-induced myocarditis. We propose that the immu-nomodulatory effects herein observed after the inhibition/in-duction of HO-1 can be explained by changes in the balancebetween NOS2/NO and HO-1/CO, which has been largelydocumented. The biochemical aspects of the interactions be-tween NO, CO, hydrogen sulfide, and metabolites from theparasite deserve further investigation.
In addition, although not measured in the present work, it ispossible that haemin treatment induces increased arginaseactivity because NO production is deviated by the expressionof arginase-1. Indeed, a recent publication showed that thatHO-1 induced by haemin led to increased arginase activity andphagocytic ability and decreased NOS2 activity via the p38MAPK pathway in macrophages [44]. Increased arginase ac-tivity in macrophages has also been associated with a higherparasite burden [45,46].
HO-1 activity is responsible for the metabolism of Haemefrom haemoglobin. In this process, free Fe, CO2, and bili-verdin are released. Free iron is a nutritional factor for T. cruziand other microbes [47]. Indeed, haemin, which is an inducerof HO-1 activity and oxidative stress, provides nutritionalsupport for T. cruzi proliferation [48]. However, if ferritin isreduced, free iron ions produced by HO activity are also ableto react with H2O2 to produce hydroxyl free radicals,increasing host’s tissue damages from oxidative stress.
Recently, Paiva et al. reported that the treatment of culturedHmox1�/� macrophages with CoPP (a different HO-1inducer), lead to reduced parasite outgrowth via upregulationof the transcriptional factor NRF-2 [49]. This suggest thatCoPP effects may be not exclusively due to HO1-1 induction.In fact, CoPP may be inducing increased phosphorylation ofErk1/2, conferring a remarkable resistance of cells toapoptosis, as has been demonstrated in cardiac stem cells [50].
Conversely, our current study uses haemin that unlikeCoPP, induces HO-1 without sequestering iron via the NLR-2pathway. Using hosts of two different genetic backgrounds, wefound an increased tissue parasitism after HO-1 induction,which is consistent with the increased presence of regulatory Tcells in tissues and reduced inflammation observed in haemintreated mice, providing experimental evidence that supportsthe idea that HO-1 plays a crucial immunomodulatory roleduring T. cruzi infection.
Surprisingly, Paiva et al. observed that CoPP-induced HO-1did not influence adaptive immunity during T. cruzi infection[49]. By contrast, we observed reduced numbers of CD4 andCD8 cells in addition to decreased IFN-g and TNF-a level in
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

10 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
heart tissue after HO-1 activation. These differences are mostlikely due to the use of a different HO-1 inducer, the selec-tiveness of its effect on HO-1, a different treatment schedule,and different mice or parasite strains.
Because we have detected parasites by immunohisto-chemistry and real-time PCR, that are more sensitive assaythan the direct visualisation of parasites in Giemsa stainedmacrophages by optical microscopy, used by Paiva et al., theseapparently contradictory results may also reflect differences inthe methods used. Additionally, the distinct microbiota ofmice from different animal facilities has been suggested to bea factor influencing the immune response [51,52]. However,the HO-1 inhibitor (SnPP) used by Paiva et al. reduced theparasitaemia during acute infection, a finding that was alsoobserved in our study with the HO-1 inhibitor ZnPPIX.
Finally, oxidative stress (in which HO-1 induction plays acentral role) is able to impair the host’s immunity to intra-cellular parasites. In fact, the selective induction of Tregs hasbeen demonstrated in human diseases characterised byexcessive levels of oxidative stress [53]. Thus, increased tissueparasitism after HO-1 induction may occur as a consequenceof a combination of effects of HO-1 activity on differenteffector mechanisms of immunity (here we provide evidenceof the effect on the induction of Treg), leading to a parasitegrowth.
We evaluated the effects of the HO-1 induction/blockade onthe inflammatory response in the heart in vivo and observedthat the inhibition of HO-1 significantly increased the in-flammatory response, while the induction of HO-1 consider-ably reduced myocarditis in T. cruzi-infected mice,irrespective of the host’s genetic background. Moreover, theinduction of HO-1 noticeably attenuated the hepatic inflam-matory response induced by the infection in BALB/c mice, asevidenced by the histological analysis and the levels oftransaminases, and these results are corroborated by a previousreport showing that the induction of HO-1 protects the liverfrom inflammatory injury in experimental models of the sys-temic inflammatory response [54].
Furthermore, we observed that mice treated with ZnPPIXwere more susceptible to the infection induced by T. cruzi,although they had increased recruitment of cells to the heartand, consequently, a reduced parasitic load in the blood.Conversely, animals treated with haemin were more resistantto the infection. Altogether, these data suggest that the in-duction of HO-1 is critical for the protection of host’s tissuesagainst the intense inflammatory response induced by theparasite and that HO-1 activity improves the host survival.Indeed, a previous study showed that HO-1 induction throughgene transfer exerts a protective effect against inflammatorytissue injury in experimental models of hypoxia in the lung byattenuating the inflammatory response [55].
The treatment with haemin reduced the production of thecytokine TNF-a and increased the levels of IL-10 in the heart,although the effects on other organs were variable. We did notobserve a significant effect of haemin on the production ofTGF-b, although ZnPPIX induced a dramatic reduction in thelevels of this cytokine in all studied organs. These findings are
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
in agreement with those from previous reports, whichdemonstrated that treatment with haemin increased IL-10 andthe frequency and activity of CD4þCD25þ cells, while havinga marginal effect on the levels of TGF-b in a model ofexperimental airway allergy [25]. In fact, the effect of haeminwas entirely dependent on the production of IL-10 because IL-10�/� mice did not show an increased frequency or activity ofCD4þCD25þ cells [25].
HO-1 is expressed constitutively in CD4þCD25þ T cellsand can be induced in CD4þCD25� T cells through stimula-tion with anti-CD28 antibodies [27,56]. In agreement withthese data, we observed that the induction of HO-1 by haeminpromoted a significant increase in the number of CD4þCD25þ
T cells in the heart. The reciprocal effect was observed inanimals that received the HO-1 inhibitor ZnPPIX. Addition-ally, HO-1 induction significantly increased the expression ofthe transcription factor Foxp3, in accordance with previousreports [24]. While haemin did not have a significant effect onthe percentage of CD4þCD25þ cells expressing the negativeco-stimulatory molecules CTLA-4 and PD-1, the inhibition ofHO-1 by ZnPPIX strikingly reduced the percentage ofCD4þCD25þ cells expressing both Foxp3 and CTLA-4.Although not assessed in the present study, HO-1 modula-tion is expected to have a direct effect on the suppressiveactivity of Tregs, as observed in previous studies [24,25].
The expression of HO-1 does not appear to be restricted toTregs. It can also be expressed on CD8 T cells [26] and APCs[26]. Thus, in our study, it is possible that a combination ofeffects of induced HO-1 expression (with haemin) or itsblockade (with ZnPPIX) in both APCs and T cells accountedfor the final phenotype observed after each treatment.
In addition to inducing oxidative stress due to its de-rivatives, NO plays an essential role in the control of parasitereplication [4] and the immune response [57]. We found thatthe induction of HO-1 led to a significant reduction of NO.Thus, it is possible that the severe myocarditis associated withthe cytotoxic effects resulting from the excessive production ofNO in the infected animals treated with ZnPPIX increasestissue destruction, resulting in the death of the mice.
An increased presence of amastigote nests was observed inthe hearts of infected animals receiving haemin, along withlow levels of NO and reduced cytokine production (IFN-g andTNF-a), which are crucial for the control of parasite prolif-eration in tissues [5]. In addition, these mice exhibitedincreased production of the cytokines IL-10 and TGF-b, thatare important for the induction and maintenance of Tregs [58]and may also contribute to the persistence of the parasite in thehost tissues [29].
The homeostatic control of the immune response is crucialfor host survival after intense inflammatory diseases. Parasiteshave evolved mechanisms to exploit the immune system,allowing them to survive within the host and in this process,they may induce an altered immune homeostasis, leading toextensive inflammatory damage to host tissues.
In conclusion, our results clearly showed that HO activationby its substrate haemin resulted in the amelioration of theinflammatory response in the heart and liver, protecting
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

11F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
infected mice from tissue damage, and significantly promotingtheir survival. This study provides experimental evidenceencouraging further research on the therapeutic potential ofHO-1 as a regulator of the immune response.
Acknowledgements
Financial support. Fundacao de Amparo a Pesquisa doEstado de Sao Paulo (FAPESP), The Millennium Institute forVaccine Development and Technology, and Conselho Nacio-nal de Desenvolvimento Cientıfico e Tecnologico (CNPQ).
Appendix A. Supplementary data
Supplementary data related to this article can be found athttp://dx.doi.org/10.1016/j.micinf.2013.10.007.
References
[1] E.C. Sabino, A.L. Ribeiro, V.M. Salemi, C. Di Lorenzo Oliveira,
A.P. Antunes, M.M. Menezes, B.M. Ianni, L. Nastari, F. Fernandes,
G.M. Patavino, V. Sachdev, L. Capuani, C. de Almeida-Neto,
D.M. Carrick, D. Wright, K. Kavounis, T.T. Goncalez, A.B. Carneiro-
Proietti, B. Custer, M.P. Busch, E.L. Murphy, Ten-year incidence of
Chagas cardiomyopathy among asymptomatic Trypanosoma cruzi-sero-
positive former blood donors, Circulation 127 (2013) 1105e1115.
[2] A. Rassi Jr., A. Rassi, J.A. Marin-Neto, Chagas disease, Lancet 375
(2010) 1388e1402.
[3] M.M. Teixeira, R.T. Gazzinelli, J.S. Silva, Chemokines, inflammation
and Trypanosoma cruzi infection, Trends Parasitol. 18 (2002) 262e265.[4] F.S. Machado, G.A. Martins, J.C. Aliberti, F.L. Mestriner, F.Q. Cunha,
J.S. Silva, Trypanosoma cruzi-infected cardiomyocytes produce chemo-
kines and cytokines that trigger potent nitric oxide-dependent trypano-
cidal activity, Circulation 102 (2000) 3003e3008.[5] D.F. Hoft, C.S. Eickhoff, Type 1 immunity provides optimal protection
against both mucosal and systemic Trypanosoma cruzi challenges, Infect
Immun. 70 (2002) 6715e6725.[6] E. Cunha-Neto, J. Kalil, Heart-infiltrating and peripheral T cells in the
pathogenesis of human Chagas’ disease cardiomyopathy, Autoimmunity
34 (2001) 187e192.
[7] E. Cunha-Neto, A.M. Bilate, K.V. Hyland, S.G. Fonseca, J. Kalil,
D.M. Engman, Induction of cardiac autoimmunity in Chagas heart dis-
ease: a case for molecular mimicry, Autoimmunity 39 (2006) 41e54.
[8] F.R. Gutierrez, P.M. Guedes, R.T. Gazzinelli, J.S. Silva, The role of
parasite persistence in pathogenesis of Chagas heart disease, Parasite
Immunol. 31 (2009) 673e685.
[9] K. Sato, J. Balla, L. Otterbein, R.N. Smith, S. Brouard, Y. Lin,
E. Csizmadia, J. Sevigny, S.C. Robson, G. Vercellotti, A.M. Choi,
F.H. Bach, M.P. Soares, Carbon monoxide generated by heme
oxygenase-1 suppresses the rejection of mouse-to-rat cardiac transplants,
J. Immunol. 166 (2001) 4185e4194.
[10] K.A. Kirkby, C.A. Adin, Products of heme oxygenase and their potential
therapeutic applications, Am. J. Physiol. Renal Physiol. 290 (2006)
F563eF571.
[11] F. Seta, L. Bellner, R. Rezzani, R.F. Regan, M.W. Dunn, N.G. Abraham,
K. Gronert, M. Laniado-Schwartzman, Heme oxygenase-2 is a critical
determinant for execution of an acute inflammatory and reparative
response, Am. J. Pathol. 169 (2006) 1612e1623.
[12] W.K. McCoubrey Jr., T.J. Huang, M.D. Maines, Isolation and charac-
terization of a cDNA from the rat brain that encodes hemoprotein heme
oxygenase-3, Eur. J. Biochem. 247 (1997) 725e732.
[13] A.M. Choi, J. Alam, Heme oxygenase-1: function, regulation, and
implication of a novel stress-inducible protein in oxidant-induced lung
injury, Am. J. Respir. Cell Mol. Biol. 15 (1996) 9e19.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
[14] M. Rizzardini, M. Terao, F. Falciani, L. Cantoni, Cytokine induction of
haem oxygenase mRNA in mouse liver. Interleukin 1 transcriptionally
activates the haem oxygenase gene, Biochem. J. 290 (Pt 2) (1993)
343e347.
[15] K. Mitani, H. Fujita, A. Kappas, S. Sassa, Heme oxygenase is a positive
acute-phase reactant in human Hep3B hepatoma cells, Blood 79 (1992)
1255e1259.
[16] R. Tenhunen, H.S. Marver, R. Schmid, The enzymatic conversion of
heme to bilirubin by microsomal heme oxygenase, Proc. Natl. Acad. Sci.
U S A 61 (1968) 748e755.
[17] L.E. Otterbein, F.H. Bach, J. Alam, M. Soares, H. Tao Lu, M. Wysk,
R.J. Davis, R.A. Flavell, A.M. Choi, Carbon monoxide has anti-
inflammatory effects involving the mitogen-activated protein kinase
pathway, Nat. Med. 6 (2000) 422e428.
[18] M.J. Alcaraz, P. Fernandez, M.I. Guillen, Anti-inflammatory actions of
the heme oxygenase-1 pathway, Curr. Pharm. Des. 9 (2003) 2541e2551.[19] L.E. Otterbein, B.S. Zuckerbraun, M. Haga, F. Liu, R. Song, A. Usheva,
C. Stachulak, N. Bodyak, R.N. Smith, E. Csizmadia, S. Tyagi,
Y. Akamatsu, R.J. Flavell, T.R. Billiar, E. Tzeng, F.H. Bach, A.M. Choi,
M.P. Soares, Carbon monoxide suppresses arteriosclerotic lesions asso-
ciated with chronic graft rejection and with balloon injury, Nat. Med. 9
(2003) 183e190.
[20] C. Wunder, R.F. Potter, The heme oxygenase system: its role in liver
inflammation, Curr. Drug Targets Cardiovasc. Haematol. Disord. 3
(2003) 199e208.
[21] M.P. Soares, Y. Lin, J. Anrather, E. Csizmadia, K. Takigami, K. Sato,
S.T. Grey, R.B. Colvin, A.M. Choi, K.D. Poss, F.H. Bach, Expression of
heme oxygenase-1 can determine cardiac xenograft survival, Nat. Med. 4
(1998) 1073e1077.
[22] C. Chauveau, D. Bouchet, J.C. Roussel, P. Mathieu, C. Braudeau,
K. Renaudin, L. Tesson, J.P. Soulillou, S. Iyer, R. Buelow, I. Anegon,
Gene transfer of heme oxygenase-1 and carbon monoxide delivery
inhibit chronic rejection, Am. J. Transpl. 2 (2002) 581e592.
[23] D. Willis, A.R. Moore, R. Frederick, D.A. Willoughby, Heme oxygenase:
a novel target for the modulation of the inflammatory response, Nat.
Med. 2 (1996) 87e90.
[24] B.M. Choi, H.O. Pae, Y.R. Jeong, Y.M. Kim, H.T. Chung, Critical role of
heme oxygenase-1 in Foxp3-mediated immune suppression, Biochem.
Biophys. Res. Commun. 327 (2005) 1066e1071.
[25] Z.W. Xia, W.W. Zhong, L.Q. Xu, J.L. Sun, Q.X. Shen, J.G. Wang,
J. Shao, Y.Z. Li, S.C. Yu, Heme oxygenase-1-mediated CD4þCD25 high
regulatory T cells suppress allergic airway inflammation, J. Immunol.
177 (2006) 5936e5945.
[26] J.F. George, A. Braun, T.M. Brusko, R. Joseph, S. Bolisetty,
C.H. Wasserfall, M.A. Atkinson, A. Agarwal, M.H. Kapturczak, Sup-
pression by CD4þCD25þ regulatory T cells is dependent on expression
of heme oxygenase-1 in antigen-presenting cells, Am. J. Pathol. 173
(2008) 154e160.
[27] H.O. Pae, G.S. Oh, B.M. Choi, S.C. Chae, H.T. Chung, Differential
expressions of heme oxygenase-1 gene in CD25� and CD25þ subsets of
human CD4þ T cells, Biochem. Biophys. Res. Commun. 306 (2003)
701e705.
[28] Z. Brener, Biology of Trypanosoma cruzi, Ann. Rev. Microb. 27 (1973)
347e382.
[29] S. Kumar, R.L. Tarleton, Antigen-specific Th1 but not Th2 cells provide
protection from lethal Trypanosoma cruzi infection in mice, J. Immunol.
166 (2001) 4596e4603.
[30] F.M. Marim, T.N. Silveira, D.S. Lima Jr., D.S. Zamboni, A method for
generation of bone marrow-derived macrophages from cryopreserved
mouse bone marrow cells, PloS One 5 (2010) e15263.
[31] G.K. Silva, F.R. Gutierrez, P.M. Guedes, C.V. Horta, L.D. Cunha,
T.W. Mineo, J. Santiago-Silva, K.S. Kobayashi, R.A. Flavell, J.S. Silva,
D.S. Zamboni, Cutting edge: nucleotide-binding oligomerization domain
1-dependent responses account for murine resistance against Trypano-
soma cruzi infection, J. Immunol. 184 (2010) 1148e1152.
[32] W.R. Pavanelli, F.R. Gutierrez, F.S. Mariano, C.M. Prado, B.R. Ferreira,
M.M. Teixeira, C. Canetti, M.A. Rossi, F.Q. Cunha, J.S. Silva, 5-Lip-
oxygenase is a key determinant of acute myocardial inflammation and
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007

12 F.R.S. Gutierrez et al. / Microbes and Infection xx (2013) 1e12
mortality during Trypanosoma cruzi infection, Microbes Infect. 12
(2010) 587e597.
[33] F. Tamion, V. Richard, S. Renet, C. Thuillez, Protective effects of heme-
oxygenase expression against endotoxic shock: inhibition of tumor ne-
crosis factor-alpha and augmentation of interleukin-10, J. Trauma 61
(2006) 1078e1084.
[34] A.M. Vicente, M.I. Guillen, M.J. Alcaraz, Heme oxygenase-1 induction
and regulation in unstimulated mouse peritoneal macrophages, Biochem.
Pharmacol. 65 (2003) 905e909.
[35] C.F. Souza, A.B. Carneiro, A.B. Silveira, G.A. Laranja, M.A. Silva-Neto,
S.C. Costa, M.C. Paes, Heme-induced Trypanosoma cruzi proliferation is
mediated by CaM kinase II, Biochem. Biophys. Res. Commun. 390
(2009) 541e546.
[36] N. Ohkura, Y. Kitagawa, S. Sakaguchi, Development and maintenance of
regulatory T cells, Immunity 38 (2013) 414e423.
[37] T.S. Lee, L.Y. Chau, Heme oxygenase-1 mediates the anti-inflammatory
effect of interleukin-10 in mice, Nat. Med. 8 (2002) 240e246.
[38] J.S. Silva, F.S. Machado, G.A. Martins, The role of nitric oxide in the
pathogenesis of Chagas disease, Front Biosci. 8 (2003) s314e325.[39] A.M. Vicente, M.I. Guillin, M.J. Alcaraz, Participation of heme
oxygenase-1 in a model of acute inflammation, Exp. Biol. Med. (May-
wood) 228 (2003) 514e516.
[40] R. Korhonen, A. Lahti, H. Kankaanranta, E. Moilanen, Nitric oxide
production and signaling in inflammation, Curr. Drug Targets Inflamm.
Allergy 4 (2005) 471e479.
[41] C.L. Hartsfield, Cross talk between carbon monoxide and nitric oxide,
Antioxid. Redox Signal. 4 (2002) 301e307.
[42] J. Tejero, A. Biswas, Z.Q. Wang, R.C. Page, M.M. Haque, C. Hemann,
J.L. Zweier, S. Misra, D.J. Stuehr, Stabilization and characterization of a
heme-oxy reaction intermediate in inducible nitric-oxide synthase, J.
Biol. Chem. 283 (2008) 33498e33507.
[43] P. Fagone, K. Mangano, M. Coco, V. Perciavalle, G. Garotta,
C.C. Romao, F. Nicoletti, Therapeutic potential of carbon monoxide in
multiple sclerosis, Clin. Exp. Immunol. 167 (2012) 179e187.[44] C. Hualin, X. Wenli, L. Dapeng, L. Xijing, P. Xiuhua, P. Qingfeng, The
anti-inflammatory mechanism of heme Oxygenase-1 induced by hae-
min in Primary rat Alveolar macrophages, Inflammation 35 (3) (2012
Jun) 1087e1093, http://dx.doi.org/10.1007/s10753-011-9415-4. PMID:
22160840.
[45] C. Stempin, L. Giordanengo, S. Gea, F. Cerban, Alternative activation
and increase of Trypanosoma cruzi survival in murine macrophages
stimulated by cruzipain, a parasite antigen, J. Leukoc. Biol. 72 (2002)
727e734.
[46] H. Cuervo, M.A. Pineda, M.P. Aoki, S. Gea, M. Fresno, N. Girones,
Inducible nitric oxide synthase and arginase expression in heart tissue
during acute Trypanosoma cruzi infection in mice: arginase I is expressed
in infiltrating CD68þmacrophages, J. Infect Dis. 197 (2008) 1772e1782.
Please cite this article in press as: F.R.S. Gutierrez, et al., Haeme oxygenase act
mental Trypanosoma cruzi infection, Microbes and Infection (2013), http://dx.do
[47] M.C. Taylor, J.M. Kelly, Iron metabolism in trypanosomatids, and its
crucial role in infection, Parasitology 137 (2010) 899e917.
[48] A. Ciccarelli, L. Araujo, A. Batlle, E. Lombardo, Effect of haemin on
growth, protein content and the antioxidant defence system in Trypa-
nosoma cruzi, Parasitology 134 (2007) 959e965.[49] C.N. Paiva, D.F. Feijo, F.F. Dutra, V.C. Carneiro, G.B. Freitas, L.S. Alves,
J. Mesquita, G.B. Fortes, R.T. Figueiredo, H.S. Souza, M.R. Fantappie,
J. Lannes-Vieira, M.T. Bozza, Oxidative stress fuels Trypanosoma cruzi
infection in mice, J. Clin. Invest. 122 (2012) 2531e2542.
[50] C. Cai, L. Teng, D. Vu, J.Q. He, Y. Guo, Q. Li, X.L. Tang,
G. Rokosh, A. Bhatnagar, R. Bolli, The heme oxygenase 1 inducer
(CoPP) protects human cardiac stem cells against apoptosis through
activation of the extracellular signal-regulated kinase (ERK)/NRF2
signaling pathway and cytokine release, J. Biol. Chem. 287 (2012)
33720e33732.
[51] I. Yang, D. Eibach, F. Kops, B. Brenneke, S. Woltemate, J. Schulze,
A. Bleich, A.D. Gruber, S. Muthupalani, J.G. Fox, C. Josenhans,
S. Suerbaum, Intestinal microbiota composition of Interleukin-10 defi-
cient C57BL/6J mice and susceptibility to helicobacter hepaticus-
induced colitis, PloS One 8 (2013) e70783.
[52] R. Duarte, A.M. Silva, L.Q. Vieira, L.C. Afonso, J.R. Nicoli, Trypano-
soma cruzi: influence of predominant bacteria from indigenous digestive
microbiota on experimental infection in mice, Exp. Parasitol. 111 (2005)
87e96.
[53] D. Mougiakakos, C.C. Johansson, R. Kiessling, Naturally occurring
regulatory T cells show reduced sensitivity toward oxidative stress-
induced cell death, Blood 113 (2009) 3542e3545.[54] T. Wen, Z.M. Wu, Y. Liu, Y.F. Tan, F. Ren, H. Wu, Upregulation of heme
oxygenase-1 with haemin prevents D-galactosamine and
lipopolysaccharide-induced acute hepatic injury in rats, Toxicology 237
(2007) 184e193.
[55] F. Amersi, R. Buelow, H. Kato, B. Ke, A.J. Coito, X.D. Shen, D. Zhao,
J. Zaky, J. Melinek, C.R. Lassman, J.K. Kolls, J. Alam, T. Ritter, H.D. Volk,
D.G. Farmer, R.M. Ghobrial, R.W. Busuttil, J.W. Kupiec-Weglinski,
Upregulation of heme oxygenase-1 protects genetically fat Zucker rat livers
from ischemia/reperfusion injury, J. Clin. Invest. 104 (1999) 1631e1639.
[56] F.S. Benghiat, L. Graca, M.Y. Braun, S. Detienne, F. Moore,
S. Buonocore, V. Flamand, H. Waldmann, M. Goldman, A. Le Moine,
Critical influence of natural regulatory CD25þ T cells on the fate of
allografts in the absence of immunosuppression, Transplantation 79
(2005) 648e654.
[57] C. Nathan, M.U. Shiloh, Reactive oxygen and nitrogen intermediates in
the relationship between mammalian hosts and microbial pathogens,
Proc. Natl. Acad. Sci. U S A 97 (2000) 8841e8848.
[58] Y. Belkaid, R.B. Blank, I. Suffia, Natural regulatory T cells and parasites:
a common quest for host homeostasis, Immunol. Rev. 212 (2006)
287e300.
ivity protects the host against excessive cardiac inflammation during experi-
i.org/10.1016/j.micinf.2013.10.007
Related Documents











