Habitat, occurrence and conservation of Saharo-Arabian-Turanian element Forsskaolea tenacissima L. in the Iberian Peninsula Javier Cabello*, Domingo Alcaraz, Francisco Go ´ mez-Mercado, Juan F. Mota, Javier Navarro, Julio Pen ˜as & Esther Gime ´nez *Departamento de Biologı´a Vegetal y Ecologı´a. Universidad de Almerı´a, E-04120 Almerı´a,Spain (Received 30 January 2002, accepted 11 June 2002) The aim of this study is to assess the Iberian populations of Forsskaolea tenacissima L. according to its biogeographical interest, habitat, geographical range and conservation status. Results point out that they are restricted to gravel wadis of Tabernas Desert (SE Spain), are scarcely included in protected areas and represent historically isolated populations with relict behaviour. We also describe a new association, Senecioni-Forsskaoleetum tenacissimae. Conservation status of species is cause for concern and two conservation actions must be carried out. Firstly, protected areas should house Forsskaolea populations and secondly, phytosociological characteriza- tion of a community allows inventorying its habitat and directing conserva- tion efforts to community level. # 2002 Elsevier Science Ltd. Keywords: biodiversity; flora; phytosociology; protected areas; reduced geographical range; relict populations; semi-arid ecosystems; Tabernas Desert Introduction The origin of the steppe areas in the Iberian Peninsula along the Pleistocene is an old classic controversial issue. The so-called ‘Steppe Theory’ (Del Villar, 1915) considered the vegetation of these ecosystems as the result of the destruction of primitive sclerophilous forests. For this reason the flora and vegetation of these ecosystems have been undervalued for conservation priorities. However, the most recent trend is to consider the possibility of persistence of non-forested areas across the Pleistocene in the Iberian Peninsula (e.g. Terradas, 1986; Gonza ´lez-Berna ´ldez, 1989; Sua ´rez Cardona et al., 1992; Blondel & Vigne, 1993; Suc et al., 1995; Willis, 1996; Ribera & Blasco-Zumeta, 1998). This debate is important because as Go ´mez-Campo (1985) pointed out, the origin of Mediterranean flora has deep implications with regard to conservation attitudes. In SE Spain, steppe areas are semi-arid ecosystems that support a notable richness in *Corresponding author. Fax: +34-950-015-069. E-mail: [email protected] 0140-1963/03/040491 + 10 $30.00/0 # 2002 Elsevier Science Ltd. Journal of Arid Environments (2003) 53: 491–500 doi:10.1006/jare.2002.1062

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

0
Journal of Arid Environments (2003) 53: 491–500doi:10.1006/jare.2002.1062
Habitat, occurrence and conservation ofSaharo-Arabian-Turanian element Forsskaolea
tenacissima L. in the Iberian Peninsula
Javier Cabello*, Domingo Alcaraz, Francisco Gomez-Mercado,Juan F. Mota, Javier Navarro, Julio Penas & Esther Gimenez
*Departamento de Biologıa Vegetal y Ecologıa. Universidad de Almerıa,E-04120 Almerıa, Spain
(Received 30 January 2002, accepted 11 June 2002)
The aim of this study is to assess the Iberian populations of Forsskaoleatenacissima L. according to its biogeographical interest, habitat, geographicalrange and conservation status. Results point out that they are restricted togravel wadis of Tabernas Desert (SE Spain), are scarcely included inprotected areas and represent historically isolated populations with relictbehaviour. We also describe a new association, Senecioni-Forsskaoleetumtenacissimae. Conservation status of species is cause for concern and twoconservation actions must be carried out. Firstly, protected areas shouldhouse Forsskaolea populations and secondly, phytosociological characteriza-tion of a community allows inventorying its habitat and directing conserva-tion efforts to community level.
# 2002 Elsevier Science Ltd.
Keywords: biodiversity; flora; phytosociology; protected areas; reducedgeographical range; relict populations; semi-arid ecosystems; Tabernas Desert
Introduction
The origin of the steppe areas in the Iberian Peninsula along the Pleistocene is an oldclassic controversial issue. The so-called ‘Steppe Theory’ (Del Villar, 1915)considered the vegetation of these ecosystems as the result of the destruction ofprimitive sclerophilous forests. For this reason the flora and vegetation of theseecosystems have been undervalued for conservation priorities. However, the mostrecent trend is to consider the possibility of persistence of non-forested areas acrossthe Pleistocene in the Iberian Peninsula (e.g. Terradas, 1986; Gonzalez-Bernaldez,1989; Suarez Cardona et al., 1992; Blondel & Vigne, 1993; Suc et al., 1995; Willis,1996; Ribera & Blasco-Zumeta, 1998).
This debate is important because as Gomez-Campo (1985) pointed out, the originof Mediterranean flora has deep implications with regard to conservation attitudes. InSE Spain, steppe areas are semi-arid ecosystems that support a notable richness in
*Corresponding author. Fax: +34-950-015-069. E-mail: [email protected]
140-1963/03/040491 + 10 $30.00/0 # 2002 Elsevier Science Ltd.

492 J. CABELLO ET AL.
species, with a large number of endemisms and Mesogean elements (Peinado et al.,1992; Cabello, 1997). One of the latest is the Saharo-Arabian–Turanian elementForsskaolea tenacissima L. (Danin & Orshan, 1999) extending into the Mediterraneanregion (Fig. 1). This plant has a disperse geographical distribution along Palestine(Zohary, 1966; Evenari et al., 1971), Egypt (Ayyard & Ghabbour, 1986), North Africaand Sahara Desert (Ozenda, 1983; Le Houerou, 1986, 1995) and SE Spain (Sagredo,1987; Paiva, 1993).
Literature (Sagredo, 1987; Paiva, 1993; Lorite et al., 1998), suggests that thegeographical range in Europe of this species is restricted to a few localities in SEIberian Peninsula, where the regional climate is notably semi-arid both due to therainfall shadow effect of the main Betic ranges and its proximity to northern Africa(Geiger, 1973).
Regarding the biogeographical and ecological interest of this species in a Europeanbiodiversity scenario, the aim of this study is to assess the populations of F. tenacissimaL. in the Iberian Peninsula according to its habitat, geographical range andconservation status. Taking into account that conservation becomes more logicalwhen directed to syntaxa or to ecosystems than to a single species (Gomez-Campo,1985), we studied the habitat of the species to enhance its role as a conservationtarget.
Material and methods
The study was based on collections, literature and fieldwork carried out during1999–2000. To assess the habitat of the species we studied biotope, communityand bioclimatic traits. At the community level we studied the phytosociologicalbehaviour of the species according to the Zurich–Montpellier school methods(Braun-Blanquet, 1979; Gehu & Rivas-Martınez, 1981). Taxa nomenclature firstfollows Castroviejo et al. (1986–2001) and Tutin et al. (1964–1980) for thosetaxa non-cited in the former reference. The syntaxonomical scheme followsRivas-Martınez et al. (2001). For the description of this new association weused International Code of Phytosociological Nomenclature (Weber et al., 2000).For bioclimatic characterization of the populations, we considered bio-climatic It index proposed by Rivas-Martınez & Loidi (1999). Conservationstatus was studied to assess the occurrence of populations in protected areas(Fig. 2).
Results
Geographical range
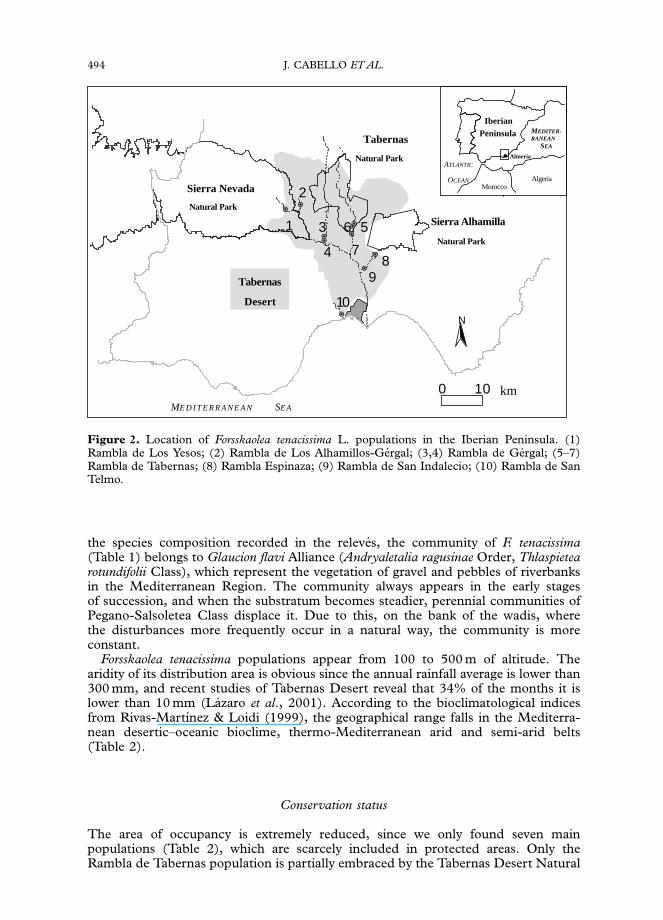
Iberian populations are restricted to gravel banks of some wadis of Tabernas Desert(Fig. 2): Rambla de Espinaza (Losa & Rivas Goday, 1968), Rambla de San Indalecio,Rambla de Gergal (Kunkel, 1987), Rambla de Tabernas (Kunkel, 1987; Sagredo,1987), Rambla de los Yesos (Lorite et al., 1998). As a result of fieldwork we confirmedall citations, and we added new localities: Rambla de San Telmo (Herbario de laUniversidad de Almerıa, 1317) y Rambla de Los Alhamillos-Gergal (Herbario de laUniversidad de Almerıa, 1318).
Habitat
Iberian populations inhabit on gravel banks. This biotope emerges naturally inwadis, and in man-made terraces in flood plains of Tabernas Desert. Considering
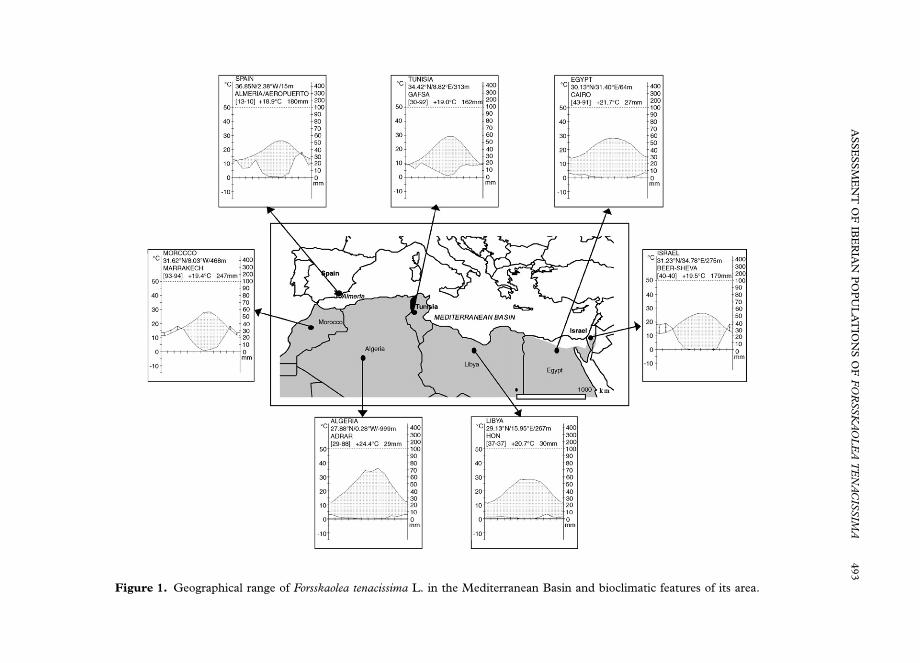
Figure 1. Geographical range of Forsskaolea tenacissima L. in the Mediterranean Basin and bioclimatic features of its area.
AS
SE
SS
ME
NT
OF
IBE
RIA
NP
OP
UL
AT
ION
SO
FF
OR
SS
KA
OL
EA
TE
NA
CIS
SIM
A493
km
89
1 6
7
53
2
4
10
0 10
N
Sierra Nevada
Natural Park
Sierra Alhamilla
Natural Park
Tabernas
Natural Park
ME D I T E R R A N E A N SE A
Tabernas
Desert
Almería
Iberian Peninsula MEDITER-
RANEANSEA
ATLANTIC
OCEANMorocco
Algeria
Figure 2. Location of Forsskaolea tenacissima L. populations in the Iberian Peninsula. (1)Rambla de Los Yesos; (2) Rambla de Los Alhamillos-Gergal; (3,4) Rambla de Gergal; (5–7)Rambla de Tabernas; (8) Rambla Espinaza; (9) Rambla de San Indalecio; (10) Rambla de SanTelmo.
494 J. CABELLO ET AL.
the species composition recorded in the releves, the community of F. tenacissima(Table 1) belongs to Glaucion flavi Alliance (Andryaletalia ragusinae Order, Thlaspietearotundifolii Class), which represent the vegetation of gravel and pebbles of riverbanksin the Mediterranean Region. The community always appears in the early stagesof succession, and when the substratum becomes steadier, perennial communities ofPegano-Salsoletea Class displace it. Due to this, on the bank of the wadis, wherethe disturbances more frequently occur in a natural way, the community is moreconstant.
Forsskaolea tenacissima populations appear from 100 to 500 m of altitude. Thearidity of its distribution area is obvious since the annual rainfall average is lower than300 mm, and recent studies of Tabernas Desert reveal that 34% of the months it islower than 10 mm (Lazaro et al., 2001). According to the bioclimatological indicesfrom Rivas-Martınez & Loidi (1999), the geographical range falls in the Mediterra-nean desertic–oceanic bioclime, thermo-Mediterranean arid and semi-arid belts(Table 2).
Conservation status
The area of occupancy is extremely reduced, since we only found seven mainpopulations (Table 2), which are scarcely included in protected areas. Only theRambla de Tabernas population is partially embraced by the Tabernas Desert Natural
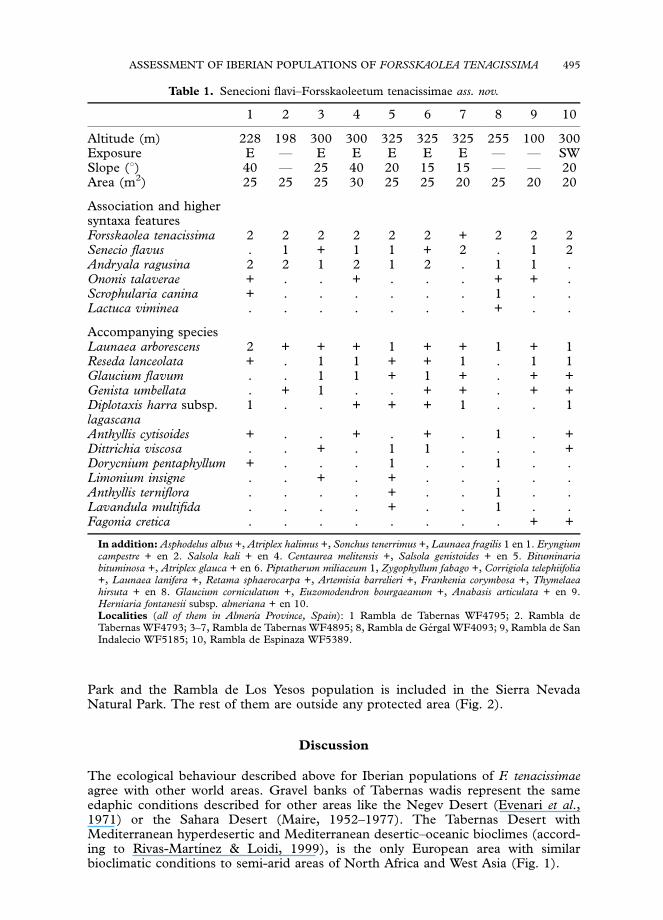
Table 1. Senecioni flavi–Forsskaoleetum tenacissimae ass. nov.
1 2 3 4 5 6 7 8 9 10
Altitude (m) 228 198 300 300 325 325 325 255 100 300Exposure E F E E E E E F F SWSlope (1) 40 F 25 40 20 15 15 F F 20Area (m2) 25 25 25 30 25 25 20 25 20 20
Association and highersyntaxa featuresForsskaolea tenacissima 2 2 2 2 2 2 + 2 2 2Senecio flavus . 1 + 1 1 + 2 . 1 2Andryala ragusina 2 2 1 2 1 2 . 1 1 .Ononis talaverae + . . + . . . + + .Scrophularia canina + . . . . . . 1 . .Lactuca viminea . . . . . . . + . .
Accompanying speciesLaunaea arborescens 2 + + + 1 + + 1 + 1Reseda lanceolata + . 1 1 + + 1 . 1 1Glaucium flavum . . 1 1 + 1 + . + +Genista umbellata . + 1 . . + + . + +Diplotaxis harra subsp.lagascana
1 . . + + + 1 . . 1
Anthyllis cytisoides + . . + . + . 1 . +Dittrichia viscosa . . + . 1 1 . . . +Dorycnium pentaphyllum + . . . 1 . . 1 . .Limonium insigne . . + . + . . . . .Anthyllis terniflora . . . . + . . 1 . .Lavandula multifida . . . . + . . 1 . .Fagonia cretica . . . . . . . . + +
In addition: Asphodelus albus +, Atriplex halimus +, Sonchus tenerrimus +, Launaea fragilis 1 en 1. Eryngiumcampestre + en 2. Salsola kali + en 4. Centaurea melitensis +, Salsola genistoides + en 5. Bituminariabituminosa +, Atriplex glauca + en 6. Piptatherum miliaceum 1, Zygophyllum fabago +, Corrigiola telephiifolia+, Launaea lanifera +, Retama sphaerocarpa +, Artemisia barrelieri +, Frankenia corymbosa +, Thymelaeahirsuta + en 8. Glaucium corniculatum +, Euzomodendron bourgaeanum +, Anabasis articulata + en 9.Herniaria fontanesii subsp. almeriana + en 10.Localities (all of them in Almerıa Province, Spain): 1 Rambla de Tabernas WF4795; 2. Rambla deTabernas WF4793; 3–7, Rambla de Tabernas WF4895; 8, Rambla de Gergal WF4093; 9, Rambla de SanIndalecio WF5185; 10, Rambla de Espinaza WF5389.
ASSESSMENT OF IBERIAN POPULATIONS OF FORSSKAOLEA TENACISSIMA 495
Park and the Rambla de Los Yesos population is included in the Sierra NevadaNatural Park. The rest of them are outside any protected area (Fig. 2).
Discussion
The ecological behaviour described above for Iberian populations of F. tenacissimaeagree with other world areas. Gravel banks of Tabernas wadis represent the sameedaphic conditions described for other areas like the Negev Desert (Evenari et al.,1971) or the Sahara Desert (Maire, 1952–1977). The Tabernas Desert withMediterranean hyperdesertic and Mediterranean desertic–oceanic bioclimes (accord-ing to Rivas-Martınez & Loidi, 1999), is the only European area with similarbioclimatic conditions to semi-arid areas of North Africa and West Asia (Fig. 1).
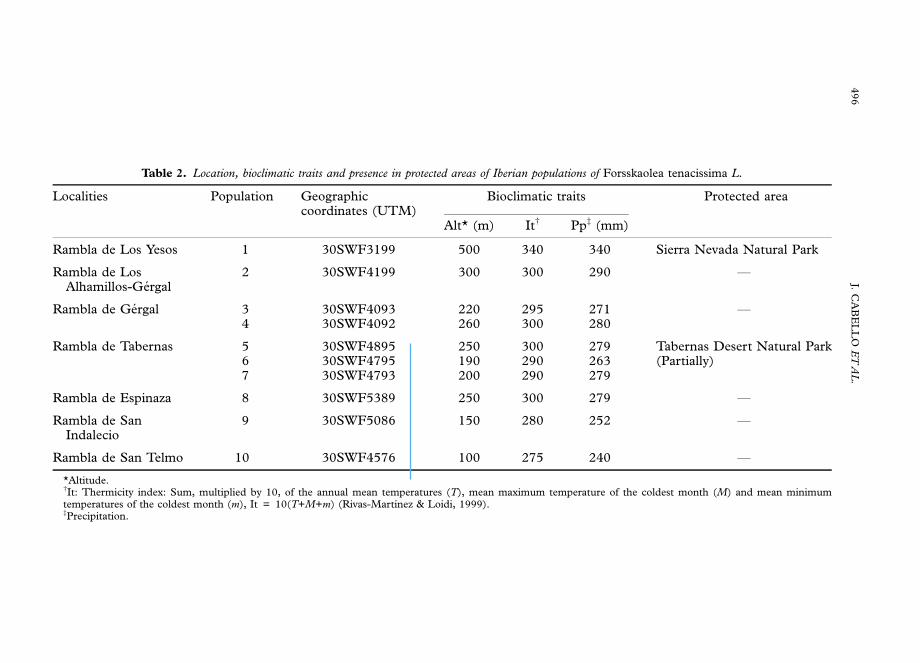
Table 2. Location, bioclimatic traits and presence in protected areas of Iberian populations of Forsskaolea tenacissima L.
Localities Population Geographiccoordinates (UTM)
Bioclimatic traits Protected area
Alt* (m) Itw Ppz (mm)
Rambla de Los Yesos 1 30SWF3199 500 340 340 Sierra Nevada Natural Park
Rambla de LosAlhamillos-Gergal
2 30SWF4199 300 300 290 F
Rambla de Gergal 34
30SWF409330SWF4092
220260
295300
271280
F
Rambla de Tabernas 567
30SWF489530SWF479530SWF4793
250190200
300290290
279263279
Tabernas Desert Natural Park(Partially)
Rambla de Espinaza 8 30SWF5389 250 300 279 F
Rambla de SanIndalecio
9 30SWF5086 150 280 252 F
Rambla de San Telmo 10 30SWF4576 100 275 240 F
*Altitude.wIt: Thermicity index: Sum, multiplied by 10, of the annual mean temperatures (T), mean maximum temperature of the coldest month (M) and mean minimumtemperatures of the coldest month (m), It = 10(T+M+m) (Rivas-Martınez & Loidi, 1999).zPrecipitation.
496
J.C
AB
EL
LO
ET
AL
.

ASSESSMENT OF IBERIAN POPULATIONS OF FORSSKAOLEA TENACISSIMA 497
Iberian F. tenacissima communities present biogeographical and floristic traits that allowus to propose a new association. Thermo- and meso-Mediterranenan shrublands of stonyareas and dry rivers distributed throughout the East Iberian Peninsula and BalearicIslands belongs to Andryaletum ragusinae association (G. flavi Alliance), led by Andryalaragusina L. and Mercurialis tomentosa L. (Bolos, 1967). However, the community of theTabernas Desert wadis is led by both F. tenacissima and Senecio flavus (Decne.) Sch. Bip.The first one is a Saharo-Arabian–Turanian element and the second one is a WestMediterranean element with restricted distribution in the Iberian Peninsula. From abiogeographical standpoint and at regional scale, both elements are differential taxa fromthe Occidental-Almerian District with regard to the rest of the Almerian SectorFMurcian–Almerian Province (Peinado et al., 1992; Mota et al., 1997).
Because of this floristic feature and the frequent presence in the releves ofOnonis talaverae, another restricted Iberian–North-African element (Devesa, 2000),we described a new endemic association for the Occidental-Almerian District(Almerian Sector, Murcian–Almerian Province) (Table 1, syntypus rel. 4). Thepresence in the releves of Salsolo-Peganion elements as accompanying species is due tothe pioneer perennial vegetation which is the spatial and successional contiguousvegetation.
Preservation of isolated populations and capturing the geographic range of species iswidely demanded in Conservation Biology (Rojas, 1992; Sutherland, 2000; Scottet al., 2001). Reasons for this are the possible effects on biodiversity elements causedby global and regional climate changes (Peters & Lovejoy, 1992), the need to maintaingenetic diversity (Jones et al., 2001), and the fact that stochastic events may threatenresources restricted to just one area (Pressey et al., 1993, 1994). Moreover, Lesica &Allendorf (1995) suggest that populations on the periphery of their ranges may be themost genetically variable and thus evolutionarily valuable if they are confronted withcontracting ranges, and so may be less vulnerable to anthropogenic or naturalchanges.
The main objective of this work was to assess F. tenacissima as a conservationflora priority in the European context. Taking into account that the last connectionbetween Africa and Europe was 3 millions years ago (Sanz de Galdeano, 1997),the Tabernas Desert populations represent historically (at geological-evolut-ionary time scale) isolated populations from the rest of African, Arabian and Turanianpopulations, and hence, as we showed above, they are interesting from biodiversitypoint of view. In addition, its occurrence in the SE Iberian Peninsula has agreat importance from a biogeographical approach. This is due to two facts: onthe one hand, it reflects both the floristic interchange between SE Iberian Peninsulaand northern Africa and western Asia during the Messinian desiccation ofthe Mediterranean Sea (Suarez et al., 1992), and, on the other one, it stressesthe hypothesis of considering the presence of steppe habitats along the Pleistocene inthe Iberian peninsula. Some authors suggest that those areas, less affected by glaciers,constituted refuges for xerophytic elements during the Quaternary Period (Bolos,1951; Blanca, 1991; Cabello, 1997).
Whereas in other areas such as Israel, it is a common taxon (The Hebrew UniversityHerbarium, 2000) in the Iberian area it is rare. Predictive distribution based onbioclimatic and ecological conditions suggest that the species should appear in most ofthe Murcian–Almerian Province. However, as a result of the fieldwork, even within theTabernas Desert, we have observed just a few populations. Two reasons for such arestricted area of occupancy might be either a low dispersal capacity of F. tenacissimaor the fact that Tabernas basin materials are mainly Tortonian marine marls, andhence many wadis have high salinity conditions. In addition, peripheral populations ofa given species tend to be smaller and more isolated than core populations. In anycase, the relict behaviour of Iberian populations is clearly stressed by their reducedgeographical range.

498 J. CABELLO ET AL.
According to the last Red List of Spanish Vascular Flora (VV.AA., 2000), F.tenacissimae, with the IUCN threatened category VU D2, is facing a high riskof extinction in the wild in the medium-term future. We believe that it is a causefor concern that Iberian populations are scarcely included in protected areas.Categorization of species according to their vulnerability to extinction in red listsdoes not represent an effective measure to preserve biodiversity. Two conserva-tion actions could be carried out. At a local scale, present protected areas inSE Iberian Peninsula must be reviewed, embracing Forsskaolea populations asconservation targets. At a regional scale, the phytosociological characterization ofthis new described community allows the inventory and incorporation of this biotopein the European Communities CORINE information and Mapping System (EUR12587, 1991), to be subsequently included in the EU Habitats Directive 92/43/EEC.
Syntaxonomic scheme
Class Thlaspietea rotundifolii Br.-Bl. 1948 + Order Andryaletalia ragusinae Rivas Godayex Rivas Goday & Esteve 1972. Alliance Glaucion flavi Br.-Bl. Ex Tchou 1948
References
Ayyard, M.A. & Ghabbour, S.I. (1986). Hot deserts of Egypt and the Sudan, Chapter 5.In: Evenari, M., Noy-Meier, I. & Goodall, D.W. (Eds), Ecosystems of the World, Vol. 12B,Chapter 5, pp. 149–202. Amsterdam: Elsevier. 421 pp.
Blanca, G. (1991). Diversidad y singularidad florıstica de las estepas almerienses. Boletın delInstituto de Estudios Almerienses, 9–10: 7–28.
Blondel, J. & Vigne, J.D. (1993). Space time and man as determinants of diversity of birds andmammals in the Mediterranean region. In: Ricklefs, R.E. & Schulter, D. (Eds), SpeciesDiversity in Ecological Communities, pp. 135–146. Chicago: Chicago University Press. 416 pp.
Bolos, O. (1951). Algunas consideraciones sobre las especies esteparias en la Penınsula Iberica.Anales del Instituto Botanico A.J. Cavanilles, 10: 445–453.
Bolos, O. (1967). Comunidades vegetales de las comarcas proximas al litoral situados entre losrıos Llobregat y Segura. Memorias de la Real Academia de Ciencias de Barcelona, 38: 3–281.
Braun Blanquet, J. (1979). Fitosociologıa. Bases para el estudio de las comunidades vegetales. Madrid:Blume Ediciones. 820 pp.
Cabello, J. (1997). Factores ambientales, estructura y diversidad en comunidades de matorralmediterraneo semiarido (Tabernas-Sierra Alhamilla-Nıjar, SE Iberico). Tesis doctoral (ined.).Universidad de Almerıa. 612 pp.
Castroviejo, S. (Eds) (1986–2001). Flora Iberica. Plantas Vasculares de la Penınsula Iberica e IslasBaleares, Vols. I–VIII. Madrid: Real Jardın Botanico, Consejo Superior de InvestigacionesCientıficas.
Danin, A. & Orshan, G. (1999). Vegetation of Israel. I. Desert and Coastal Vegetation. Leiden:Backhuys Publ. 346 pp.
Del Villar, H. (1915). Avance geobotanico sobre la pretendida estepa central espanola. Iberica,23: 328–333.
Devesa, J.A. (2000). Ononis L. In: Talavera, S., Adel, C., Castroviejo, S., Herrero, A., RomeroZarco, C., Salgueiro & Velayos (Eds.) Flora Iberica, Vol. VII(II), pp. 590–646. Madrid: RealJardın Botanico, Consejo Superior de Investigaciones Cientıficas. 1119 pp.
EUR 12587 (1991). CORINE Biotopes Manual F A Method to Identify and Describe ConsistentlySites of Major Importance for Nature Conservation. Data Specifications, Vol. 3. Luxembourg:Office for Official Publications of the European Communities. 300 pp.
Evenari, M., Shanan, L. & Tadmor, N. (1971). The Negev. The Challenge of a Desert. Cambridge,MA: Harvard University Press. 437 pp.
Gehu, J.M. & Rivas Martınez, S. (1981). Notions fondamentales de Phytosociologie. In:Dierschcke, H. (Ed.), Syntaxonomie. Berichte der Internationalen Symposien der InternationalenVereinigung fur Vegetationskunde, pp. 5–33. Vaduz: J. Cramer. 217 pp.

ASSESSMENT OF IBERIAN POPULATIONS OF FORSSKAOLEA TENACISSIMA 499
Geiger, F. (1973). El Sureste espanol y los problemas de la aridez. Revista de Geografıa, 7:166–209.
Gomez Campo, C. (1985). The conservation of Mediterranean plants: Principles and problems.In: Gomez Campo (Ed.), Plant Conservation in the Mediterranean Area, Chapter 1, pp. 3–8.Dordrecht: DR W. Junk Publ. 269 pp.
Gonzalez Bernaldez, F. (1989). Ecosistemas aridos y endorreicos espanoles. Seminario sobre ZonasAridas en Espana, pp. 223–238. Madrid: Real Academia de Ciencias Exactas, Fısicas yNaturales. 308 pp.
Jones, B., Gliddon, C. & Good, J.E.G. (2001). The conservation of variation in geographicallyperipheral populations: Lloydia serotina (Liliaceae) in Britain. Biological Conservation, 101:147–156.
Kunkel, G. (1987). Florula del desierto almeriense. Instituto de Estudios Almerienses, ColeccionInvestigacion, 5: 1–252.
Lazaro, R., Rodrigo, F.S., Gutierrez, L., Domingo, F. & Puigdefabregas, J. (2001). Analysis of a30-year rainfall record (1967–1997) in semi-arid SE Spain for implications on vegetation.Journal of Arid Environments, 48: 373–395.
Le Houerou, H.N. (1986). The desert and arid zones of Northern Africa. In: Evenari, M.,Noy-Meier, I. & Goodall, D.W. (Eds), Ecosystems of the world, Vol. 12B, Chapter 4,pp. 101–148. Amsterdam: Elsevier. 421 pp.
Le Houerou, H.N. (1995). Bioclimatologie et biogeographie des steppes arides du Nord de l’Afrique.Diversite biologique, developpement durable et desertisation. Montpellier: CIHEAM. 397 pp.
Lesica, P. & Allendorf, F.W. (1995). When are peripheral populations valuable for conservation.Conservation Biology, 9: 753–760.
Lorite, J., Navarro, F.B. & Valle, F. (1998). Aportaciones a la flora vascular de SierraNevada (Almerıa): el elemento Murciano-Almeriense. Acta Botanica Malacitana, 23:256–259.
Losa Espana, M. & Rivas Goday, S. (1968). Estudio florıstico y geobotanico de la provincia deAlmerıa, I. Archivos del Instituto de Aclimatacion, 3. Almerıa: Consejo Superior deInvestigaciones Cientıficas, Patronato ‘‘Alonso de Herrera’’. 111 pp.
Maire, R. (1952–1977). Flore de l’Afrique du Nord, 14 Vols. Parıs: Paul Lechevalier.Mota, J., Cabello, J., Cueto, M., Gomez Mercado, F., Gimenez, E. & Penas, J. (1997). Datos
sobre la vegetacion del sureste de Almerıa. (Desierto de Tabernas, Karst en Yesos de Sorbas y Cabo deGata). Almerıa: Servicio de Publicaciones de la Universidad de Almerıa. 130 pp.
Ozenda, P. (1983). Flore du Sahara. Paris: Centre National de la Recherche Scientifique. 625 pp.Paiva, J. (1993). Forsskaolea L. In: Castroviejo, S. et al. (Eds), Flora Iberica, Vol. III, pp. 272–275.
Madrid: Real Jardın Botanico, Consejo Superior de Investigaciones Cientıficas. 730 pp.Peinado, M., Alcaraz, F. & Martınez Parras, J.M. (1992). Vegetation of Southeastern Spain. Flora
et Vegetatio Mundi, X. Berlin-Stuttgart: J. Cramer. 487 pp.Peters, R.L. & Lovejoy, T.E. (1992). Global Warming and Biological Diversity. New Haven, CT:
Yale University Press. pp.Pressey, R.L., Humphries, C.J., Margules, C.R., Vanewright, R.I. & Williams, P.H. (1993).
Beyond opportunism: key principles for systematic reserve selection. Trends in Ecology andEvolution, 8: 124–128.
Pressey, R.L., Johnson, I.R. & Wilson, P.D. (1994). Shapes of irremplaceability: towardsa measure of the contribution of sites to a conservation goal. Biological Conservation, 3:242–262.
Ribera, I. & Blasco-Zumeta, J. (1998). Biogeographical links between steppe insects in theMonegros region (Aragon, NE Spain) the eastern Mediterranean, and central Asia. Journal ofBiogeography, 25: 969–986.
Rivas Martınez, S. & Loidi, J. (1999). Bioclimatology of the Iberian Peninsula. In: Iter IbericumA.D. MIM. (Excursus geobotanicus per Hispanian et Lusitaniam, ante XLII Symposium SocietatisInternationalis Scientiae Vegetationis Bilbao mense Iulio celebrandum dicti Anni). ItineraGeobotanica, 13: 41–47.
Rivas Martınez, S., Fernandez Gonzalez, F., Loidi, J. & Lousa, M. (2001). Syntaxonomicalchecklist of vascular plant communities of Spain and Portugal to association level. ItineraGeobotanica, 14: 5–341.
Rojas, M. (1992). The species problem and conservation: what are we protecting? BiologicalConservation, 3: 206–208.

500 J. CABELLO ET AL.
Sagredo, R. (1987). Flora de Almerıa. Plantas vasculares de la provincia. Almerıa: Instituto deEstudios Almerienses. 552 pp.
Sanz de Galdeano, C. (1997). La zona interna betico-rifena. Monografica Tierras del Sur.Granada: Servicio de Publicaciones de la Universidad de Granada. 316 pp.
Scott, J.M., Murray, M., Wright, R.G., Csuti, B., Morgan, P. & Pressey, R.L. (2001).Representation of natural vegetation in protected areas: capturing the geographic range.Biodiversity and Conservation, 10: 1297–1301.
Suarez Cardona, F., Sainz Ollero, H., Santos Martınez, T. & Gonzalez Bernaldez, F. (1992). Lasestepas ibericas. Madrid: Ministerio de Obras Publicas y Transportes. 160 pp.
Suc, J.P., Bertinin, A., Combourieu-Nebout, N., Diniz, F., Leroy, S., Russo-Ermolli, E., Zheng,Z., Bessais, E. & Ferrier, J. (1995). Structure of West Mediterranean vegetation and climatesince 5,3 m.a. Acta Zoologica Cracoviensia, 38: 3–16.
Sutherland, W.J. (2000). The Conservation Handbook. Research, Management and Policy. Oxford:Blackwell Science Ltd. 278 pp.
Terradas, J. (1986). El paisatge vegetal dels Monegros: assaig d’interpretacio. Orsis 2: 71–95.The Hebrew University Herbarium (2000). Vascular plants F HUJ Herbarium. BioGIS
(database). http://www.biogis.huji.ac.ilTutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M. &
Webb, D.A. (1964–1980). Flora Europaea, Vols. 1–5. Cambridge: Cambridge University Press.Weber, H.E., Moravec, J. & Theurillat, J.P. (2000). International Code of Phytosociological
Nomenclature (3rd Ed). Journal of Vegetation Science, 11: 739–768.Willis, K.J. (1996). Where did all the flowers go? The fate of temperate European flora during
glacial periods. Endeavour, 20: 110–114.VV.AA. (2000). Lista Roja de la Flora Vascular Espanola (valoracion segun categorıas UICN).
Conservacion Vegetal, 6: 11–38.Zohary, M. (1966). Flora Palestina. Part one. Text. Jerusalem: Jerusalem Academic Press. 367 pp.
Related Documents











