Probiotic characteristics of Lactobacillus acidophilus and Lactobacillus paracasei and their effects on immune response and gene expression in mice By Gunaranjan Paturi A thesis submitted in fulfilment of the requirements for the degree of Doctor of Philosophy Centre for Plant and Food Science University of Western Sydney Australia March 2007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Probiotic characteristics of Lactobacillus acidophilus and
Lactobacillus paracasei and their effects on immune response and
gene expression in mice
By
Gunaranjan Paturi
A thesis submitted in fulfilment of the requirements for the degree of
Doctor of Philosophy
Centre for Plant and Food Science
University of Western Sydney
Australia
March 2007

i
Statement of Authentication
The work presented in this thesis is, to the best of my knowledge and belief, original
except as acknowledged in the text. I hereby declare that I have not submitted this
material, either in full or in part, for a degree at this or any other institution.
Gunaranjan Paturi

ii
Table of contents
Statement of Authentication ...................................................................................... i
Acknowledgements.................................................................................................... vi
List of Publications...................................................................................................vii
List of Tables ............................................................................................................. ix
List of Figures ............................................................................................................ xi
List of Abbreviations ..............................................................................................xiii
ABSTRACT ................................................................................................................ 1
CHAPTER 1. Introduction ....................................................................................... 3
1.1. GUT MICROFLORA ........................................................................................... 4
1.2. GUT IMMUNE SYSTEM .................................................................................. 11
1.3. PROBIOTICS ..................................................................................................... 14
1.4. IMMUNE RESPONSES OF LACTIC ACID BACTERIA ............................... 17
1.5. THERAPEUTIC EFFECTS OF LACTIC ACID BACTERIA .......................... 20
1.5.1. Diarrhoeal diseases.................................................................................. 20
1.5.2. Inflammatory bowel disease ................................................................... 21
1.5.3. Irritable bowel syndrome ........................................................................ 21
1.5.4. Helicobacter pylori infections................................................................. 22
1.6. FUNCTIONAL GENOMICS AND LACTIC ACID BACTERIA .................... 26
1.7. AIMS................................................................................................................... 28
CHAPTER 2. Selection and characterisation of Lactobacillus and
Bifidobacterium species for potential probiotic strains......................................... 29
2.1. ABSTRACT........................................................................................................ 30
2.2. INTRODUCTION .............................................................................................. 31
2.3. MATERIALS AND METHODS........................................................................ 35
2.3.1. Microorganisms and growth conditions.................................................. 35
2.3.2. Acid tolerance ......................................................................................... 35
2.3.3. Bile tolerance .......................................................................................... 36
2.3.4. Hydrophobicity assay.............................................................................. 37
2.3.5. Autoaggregation and coaggregation assays ............................................ 38

iii
2.3.6. Statistical analysis ................................................................................... 39
2.4. RESULTS ........................................................................................................... 40
2.4.1. Tolerance to simulated gastric juice........................................................ 40
2.4.2. Tolerance to bile salts.............................................................................. 41
2.4.3. Surface hydrophobicity of Lactobacillus and Bifidobacterium strains... 46
2.4.4. Autoaggregation and coaggregation of Lactobacillus and
Bifidobacterium strains ..................................................................................... 46
2.5. DISCUSSION ..................................................................................................... 51
2.6. CONCLUSIONS................................................................................................. 56
CHAPTER 3. Effect of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26
on gut and systemic immunity and bacterial translocation in mice .................... 57
3.1. ABSTRACT........................................................................................................ 58
3.2. INTRODUCTION .............................................................................................. 59
3.3. MATERIALS AND METHODS........................................................................ 63
3.3.1. Animals ................................................................................................... 63
3.3.2. Microorganisms ...................................................................................... 63
3.3.3. Feeding procedure................................................................................... 63
3.3.4. Histological samples ............................................................................... 64
3.3.5. Immunofluorescent detection of IgA cells .............................................. 64
3.3.6. Detection of cytokine producing cells in small intestine ........................ 65
3.3.7. Splenocytes ............................................................................................. 65
3.3.8. Lymphocyte proliferation assay.............................................................. 66
3.3.9. Estimation of cytokines in blood and splenocytes .................................. 66
3.3.10. Phagocytosis.......................................................................................... 67
3.3.11. Bacterial translocation assay................................................................. 67
3.3.12. Statistical analysis ................................................................................. 68
3.4. RESULTS ........................................................................................................... 69
3.4.1. Determination of IgA producing cells..................................................... 69
3.4.2. Effect of LAB on cytokines in gut .......................................................... 69
3.4.3. Lymphocyte proliferative responses ....................................................... 70
3.4.4. Effect of LAB on cytokines in blood serum and splenocytes................. 70
3.4.5. Phagocytic activity .................................................................................. 77
3.4.6. Bacterial translocation............................................................................. 77

iv
3.5. DISCUSSION ..................................................................................................... 83
3.6. CONCLUSIONS................................................................................................. 89
CHAPTER 4. Immunostimulatory responses of L. acidophilus LAFTI L10 and
L. paracasei LAFTI L26 in cholera toxin mice ...................................................... 90
4.1. ABSTRACT........................................................................................................ 91
4.2. INTRODUCTION .............................................................................................. 92
4.3. MATERIALS AND METHODS........................................................................ 96
4.3.1. Mice ........................................................................................................ 96
4.3.2. Lactobacillus strains and feeding procedure........................................... 96
4.3.3. Cholera toxin immunisation.................................................................... 97
4.3.4. Intestinal fluid and serum preparation..................................................... 97
4.3.5. Determination of nitric oxide.................................................................. 97
4.3.6. Estimation of Immunoglobulin A ........................................................... 98
4.3.7. Histological samples ............................................................................... 98
4.3.8. Immunofluorescent detection of cytokine producing cells ..................... 99
4.3.9. Statistical analysis ................................................................................... 99
4.4. RESULTS ......................................................................................................... 100
4.4.1. Nitric oxide production in intestinal fluid and serum ........................... 100
4.4.2. Immunoglobulin A antibodies in intestinal fluid and serum................. 100
4.4.3. Detection of cytokine producing cells in small intestine ...................... 103
4.5. DISCUSSION ................................................................................................... 107
4.6. CONCLUSIONS............................................................................................... 111
CHAPTER 5. Microarray analysis of L. acidophilus LAFTI L10 induced gene
expression in the small intestine of mice .............................................................. 112
5.1. ABSTRACT...................................................................................................... 113
5.2. INTRODUCTION ............................................................................................ 114
5.3. MATERIALS AND METHODS...................................................................... 117
5.3.1. Animals ................................................................................................. 117
5.3.2. Lactic acid bacteria ............................................................................... 117
5.3.3. Experimental design.............................................................................. 117
5.3.4. RNA isolation ....................................................................................... 118
5.3.5. Microarray analysis............................................................................... 119

v
5.3.6. Synthesis of cRNA probes .................................................................... 119
5.3.7. Hybridisation of probes to oligonucleotide microarray ........................ 119
5.3.8. Data Analysis ........................................................................................ 120
5.4. RESULTS ......................................................................................................... 121
5.5. DISCUSSION ................................................................................................... 137
5.6. CONCLUSIONS............................................................................................... 145
CHAPTER 6. Concluding summary .................................................................... 146
REFERENCES ....................................................................................................... 154

vi
Acknowledgements
I am sincerely thankful to A/Prof. Kasipathy Kailasapathy for his supervision,
guidance and enthusiastic encouragement throughout this study. I am also thankful
for sharing his knowledge and time.
My deepest gratitude to Dr. Michael Phillips for his guidance and constructive
comments. I am especially thankful for sharing his knowledge in microbiology
during the progression of this study.
I am thankful to A/Prof. Shelley Burgin for the friendship and support I received
during this study. Many thanks to Dr. Mark Jones for his help during the animal
experiments.
I am grateful to Prof. Gabriela Perdigon for sharing her expertise in immunology
during this study at University of Western Sydney, Australia and also at Centro de
Referencias para Lactobacilos, Tucuman, Argentina. My sincere thanks to Prof.
Gerald Tannock for giving me the opportunity to visit his laboratory at University of
Otago, Dunedin, New Zealand to enhance my skills in gut microbiology.
I wish to thank all my colleagues for their friendship, advice and support during this
study at University of Western Sydney, Australia. I am grateful to all the technical
and administrative staff of Centre for Plant and Food Science and School of Natural
Sciences for their assistance. I am also thankful to Commonwealth of Australia and
University of Western Sydney for providing me the scholarship and resources to
pursue this study.
Last but not the least, I am thankful to my wife, parents, family members and friends
for their continued support, love and encouragement.

vii
List of Publications
Journal Papers
1. Paturi G., Phillips, M., Jones, M., Kailasapathy, K., 2007. Immune enhancing
effects of Lactobacillus acidophilus LAFTI L10 and Lactobacillus paracasei
LAFTI L26 in mice. International Journal of Food Microbiology 115, 115-
118.
2. Paturi G., Phillips, M., Kailasapathy, K. Effect of probiotic strains
Lactobacillus acidophilus LAFTI L10 and Lactobacillus paracasei LAFTI
L26 on systemic immune functions and bacterial translocation in mice.
Journal of Food Protection (Manuscript accepted).
3. Paturi G., Phillips, M., Kailasapathy, K. Oral administration of Lactobacillus
acidophilus LAFTI L10 modulate immune responses and gene expression in
the small intestine of cholera toxin mice model. (Manuscript submitted).
4. Paturi G., Phillips, M., Kailasapathy, K. Microarray analysis of Lactobacillus
acidophilus LAFTI L10 induced gene expression in the small bowel of mice.
(Manuscript submitted).
Conference Presentations
1. Paturi G., Kailasapathy, K., Phillips, M., Jones, M., 2006. Unravelling the
effects of Lactobacillus acidophilus in murine cholera toxin model.
Proceedings of the Annual Australian Society for Microbiology Conference,
Gold Coast, Australia. (Oral Presentation).
2. Paturi G., Kailasapathy, K., Phillips, M., Jones, M., 2006. Microarray
analysis of gene expression in murine small intestine modulated by
Lactobacillus acidophilus. The International Conference on Nutrigenomics
and Gut Health, Auckland, New Zealand. (Oral Presentation).

viii
3. Paturi G., Kailasapathy, K., Phillips, M., Jones, M., 2005. Immunological
effects of probiotic bacteria with reference to Lactobacillus acidophilus and
Lactobacillus paracasei. Proceedings of the Annual Australian Society for
Microbiology Conference, Canberra, Australia. (Poster Presentation).
4. Paturi G., Kailasapathy, K., Phillips, M., Jones, M., Perdigon, G., 2005.
Studies of probiotic bacteria on immunostimulating properties in gut immune
system. 38th
Annual Australian Institute of Food Science and Technology
Convention, Sydney, Australia. (Poster Presentation).
5. Paturi G., Kailasapathy, K., Phillips, M., Perdigon, G., Jones, M., 2005.
Potential immunostimulating properties of Lactobacillus acidophilus (LAFTI
L10) and Lactobacillus paracasei (LAFTI L26) in mice. Beneficial Microbes,
American Society for Microbiology Conference, Nevada, USA. (Poster
Presentation).

ix
List of Tables
Table 1.1. Microorganisms that are commonly considered as probiotics.
16
Table 1.2. Therapeutic properties of lactic acid bacteria. (Adapted from Saxelin et al., 2005; Reid et al., 2006).
24, 25
Table 2.1. Survival of Lactobacillus and Bifidobacterium strains in simulated gastric conditions for 3 h.
43,44
Table 2.2. Survival of Lactobacillus and Bifidobacterium strains in milk-yeast medium with oxgall.
45
Table 3.1. Proliferative responses of spleen cells stimulated with ConA and LPS in mice fed with either L.
acidophilus L10 or L. paracasei L26.
74
Table 3.2. Translocation of Lactobacillus spp. to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed with either L.
acidophilus L10 or L. paracasei L26 for 14-days.
80
Table 3.3. Translocation of total anaerobes to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed with either L.
acidophilus L10 or L. paracasei L26 for 14-days.
81
Table 3.4. Translocation of enteric bacteria to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed with either L.
acidophilus L10 or L. paracasei L26 for 14-days.
82
Table 5.1. Effect of L. acidophilus L10 on gene expression (Up- and down-regulated) in the small intestine of normal mice.
123,124,
125
Table 5.2. Functional analysis of differentially expressed genes (Up-regulated) after oral administration of L. acidophilus L10 in normal mice.
126
Table 5.3. Functional analysis of differentially expressed genes (Down-regulated) after oral administration of L. acidophilus L10 in normal mice.
127
Table 5.4. Pathway analysis of differentially expressed genes (Up- and down-regulated) after oral administration of L. acidophilus L10 in normal mice.
128
Table 5.5. Effect of L. acidophilus L10 on gene expression (Up- and down-regulated) in the small intestine of cholera toxin mice model.
131, 132,
133
Table 5.6. Functional analysis of differentially expressed genes (Up-regulated) after oral administration of L. acidophilus L10 in cholera toxin mice model.
134
Table 5.7. Functional analysis of differentially expressed genes (Down-regulated) after oral administration of L. acidophilus L10 in cholera toxin mice model.
135

x
Table 5.8. Pathway analysis of differentially expressed genes (Up- and down-regulated) after oral administration of L. acidophilus L10 in cholera toxin mice model.
136

xi
List of Figures
Figure 1.1. Development of human and murine gastrointestinal ecosystems. Abbreviations in the figure: IELs - intraepithelial lymphocytes; LP - lamina propria; sIgM - surface immunoglobulin M; sIgA - surface immunoglobulin A; TCR - T-cell receptor. (Adapted from McCracken and Lorenz, 2001).
6
Figure 1.2. Overview of gastrointestinal (GI) tract. (A) Microbial distribution (Anaerobic and aerobic genera) in various compartments of the GI tract. (B) Functional properties of intestinal microflora influencing intestinal mucosa. (Adapted from O’Hara and Shanahan, 2006).
10
Figure 1.3. Schematic representation of gut immune system. (Adapted from McCracken and Lorenz, 2001).
13
Figure. 2.1. Surface hydrophobicity of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B. infantis ASCC 1912 and L. acidophilus ASCC 2400.
48
Figure 2.2. Autoaggregation abilities of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B. infantis ASCC 1912 and L. acidophilus ASCC 2400.
49
Figure 2.3. Coaggregation abilities of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B. infantis ASCC 1912 and L. acidophilus ASCC 2400.
50
Figure 3.1. Effect of A) L. acidophilus L10 and B) L. paracasei L26 on immunoglobulin (Ig)-A producing cells in the small intestine of mice.
72
Figure 3.2. Effect of orally administered L. acidophilus L10 or L. paracasei L26 on A) interleukin (IL)-10 and B) interferon (IFN)-� cytokine producing cells in the small intestine of mice.
73
Figure 3.3. Estimation of cytokines A) interleukin (IL)-10 and B) interferon (IFN)-� in serum of mice fed with L. acidophilus L10 or L. paracasei L26.
75
Figure 3.4. Production of A) interleukin (IL)-10 and B) interferon (IFN)-� by spleen cells from mice orally fed with either L. acidophilus L10 or L.
paracasei L26.
76
Figure 3.5. Effect of A) L. acidophilus L10 and B) L.
paracasei L26 on phagocytic activity of the peritoneal macrophages.
79
Figure 4.1. Nitric oxide production in A) intestinal fluid and B) serum of mice fed with L. acidophilus L10 or L. paracasei L26.
101

xii
Figure 4.2. Immunoglobulin (Ig)-A antibody response to cholera toxin in A) intestinal fluid and B) serum of mice fed with L. acidophilus L10 or L.
paracasei L26.
102
Figure 4.3. Effect of L. acidophilus L10 and L. paracasei L26 on (A) interferon (IFN)-� and tumor necrosis factor (TNF)-�; (B) interleukin (IL)-4, IL-6 and IL-10 in the small intestine of mice.
104,105
Figure 4.4. Histological sections of the small intestine in mice fed with L. acidophilus L10 showing A) interleukin (IL)-10 and B) interferon (IFN)-� producing cells labelled with rabbit anti-mouse IL-10 and IFN-� respectively and secondary antibody as goat anti-rabbit conjugated with FITC, which fluoresces. Magnification 400X.
106

xiii
List of Abbreviations
CD Cluster of differentiation
CFU Colony Forming Units
ConA Concanavalin A
CT Cholera toxin
FITC Fluorescein isothiocyanate
GALT Gut-associated lymphoid tissue
GI Gastrointestinal
GRAS Generally Regarded As Safe
IBD Inflammatory Bowel Disease
IBS Irritable Bowel Syndrome
IFN Interferon
Ig Immunoglobulin
IL Interleukin
LAB Lactic acid bacteria
LPS Lipopolysaccharide
MLN Mesenteric lymph node
MRS de Man, Rogosa and Sharpe agar or broth
NGYC Non-fat skim milk, glucose, yeast extract and cysteine medium
NK Natural Killer (Cell)
NO Nitric oxide
PBS Phosphate buffered saline
TNF Tumor necrosis factor
WC Wilkins-Chalgren agar

1
ABSTRACT
Probiotic bacteria such as Lactobacillus and Bifidobacterium species are normal
inhabitants of healthy gastrointestinal (GI) tract, which may promote beneficial
effects on host through limiting the growth of undesirable microorganisms and
modulating the immune system. In the present study, Lactobacillus and
Bifidobacterium strains were screened for their in vitro acid and bile tolerance,
autoaggregation, coaggregation and hydrophobic abilities to identify potential
probiotic bacteria. Lactobacillus acidophilus LAFTI L10 and Lactobacillus
paracasei LAFTI L26 were selected based on their overall tolerance to in vitro acidic
conditions to further investigate their influence on various immune functions and
gene expression in mice.
Immunofluorescent analysis of small intestine in mice fed with L. acidophilus or L.
paracasei demonstrated an increase of immunoglobulin (Ig)-A, interleukin (IL)-10
and interferon (IFN)-� producing cells compared to control mice. In systemic
immune response, proliferative responses of splenocytes stimulated with mitogens,
concanavalin A (ConA) and lipopolysaccharide (LPS) showed differences between
L. acidophilus and L. paracasei strains. In comparison to control mice, IL-10 and
IFN-� levels in blood serum and splenocytes stimulated with ConA were enhanced in
mice fed with either L. acidophilus or L. paracasei. Increased phagocytic activity of
peritoneal macrophages against Candida albicans was determined in mice fed with
L. acidophilus or L. paracasei. Translocation of Lactobacillus spp. and total
anaerobes to Peyer’s patches as well as mesenteric lymph nodes were modulated in
L. acidophilus or L. paracasei-fed mice. Furthermore, there was no bacterial

2
translocation to spleen, liver or blood in mice fed with L. acidophilus or L.
paracasei.
In cholera toxin (CT) mice model, L. acidophilus-fed mice increased the nitric oxide
(NO) levels in serum, whereas L. paracasei-fed mice enhanced the NO levels in
serum and intestinal fluid. Mice fed with L. acidophilus or L. paracasei showed an
increased IgA response to CT in intestinal fluid and serum compared to control mice.
The analysis of cytokine producing cells in small intestine of L. acidophilus or L.
paracasei-fed mice showed the regulation of Th1 and Th2 cytokines such as IFN-�,
tumor necrosis factor (TNF)-�, IL-4, IL-6 and IL-10.
Microarray analysis of L. acidophilus induced gene expression in small intestine of
normal mice demonstrated an up- and down-regulation of 27 and 35 genes, which are
involved in cell-cell signalling, cell growth and proliferation, cell death and various
metabolic functions. In CT mice model, L. acidophilus influenced an up- and down-
regulation of 28 and 32 genes in small intestine that are involved in cell functions,
drug metabolism and immune responses.
In summary, L. acidophilus and L. paracasei showed tolerance to various gastric
conditions and bile salts. Lactobacillus acidophilus and L. paracasei enhanced gut
and systemic immune functions, particularly non-specific and specific immune
responses in normal and CT mice. Moreover, L. acidophilus regulated the genes
involved in various biological functions in small bowel of normal and CT mice,
which provided a basis in understanding the pathways through which these bacteria
are beneficial to the host.

3
CHAPTER 1
Introduction

4
1.1. GUT MICROFLORA
The gastrointestinal (GI) tract of human consists of diversified microflora ranging
between 300 - 500 bacterial species. The microflora of GI tract is about 10 times
higher than eukaryotic cells in the human body (Simon and Gorbach, 1986;
Bengmark, 1998). In a healthy host, enteric bacteria that were acquired from birth
colonise the alimentary tract and their composition remains mostly constant
thereafter. Aerobic and facultative microorganisms such as coliforms, lactobacilli
and streptococci are first to colonise the GI tract, which are considered to be
important in reducing oxidation process in the intestine that lead to the colonisation
of anaerobic bacteria such as Bacteroides and Bifidobacterium species.
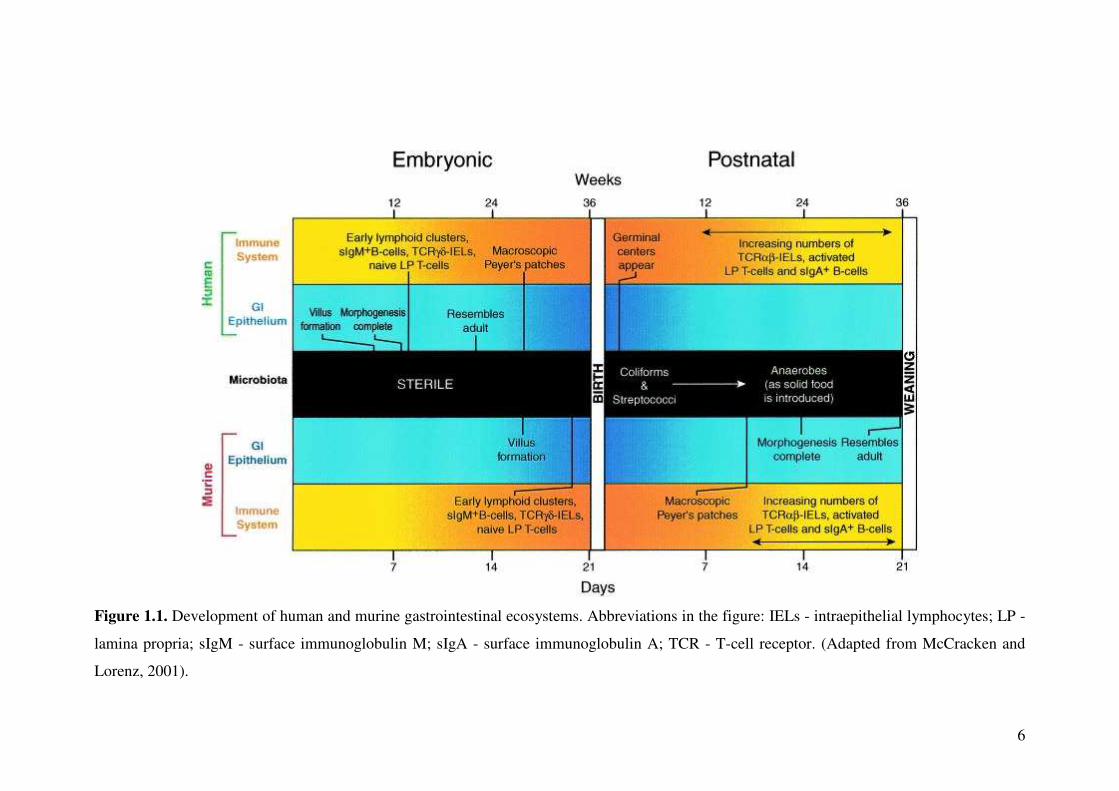
The consumption of solid food gradually transforms the intestinal microflora into
stable microbial communities in adults, where anaerobic microbes prevail (Fig. 1.1).
The microflora in adult GI tract comprises of 99.9% obligate anaerobes, where
gastric secretions and rapid motility in the upper small intestine restrict them to 103
organisms per millilitre of luminal fluids at this site and between 1011 - 1012
organisms per gram or millilitre of colon contents (Xu and Gordon, 2003; Backhed et
al., 2005; Fig. 1.2A).
Bacterial composition in GI tract differ based on anatomical properties of the
interacting sites and other conditions such as pH, oxygen availability and transit time
of food contents. Bacterial species belonging to genera Bacteroides, Bifidobacterium,
Fusobacterium, Butyrivibrio, Clostridium, Eubacterium and Lactobacillus are some
of the resident microflora in GI tract. Predominantly, bifidobacteria and lactobacilli

5
represents 90% of the bacterial population, whereas less than 0.01% consists of
diversified bacterial population such as pathogenic microorganisms (Tournut, 1993).
6
Figure 1.1. Development of human and murine gastrointestinal ecosystems. Abbreviations in the figure: IELs - intraepithelial lymphocytes; LP -
lamina propria; sIgM - surface immunoglobulin M; sIgA - surface immunoglobulin A; TCR - T-cell receptor. (Adapted from McCracken and
Lorenz, 2001).

7
The intestinal microflora influence wide variety of host functions such as synthesis
of biotin, folate, vitamin K and conversion of unabsorbed dietary sugars into short-
chain fatty acids, which can be used as energy source by colonic mucosa (O’Hara
and Shanahan, 2006; Fig. 1.2B). The ability of commensal microflora differs in
promoting the development of gut associated lymphoid tissues (GALT). However,
commensal microflora play an important role in maintaining immune homeostasis in
developing and adult gut. Fermentation processes of colonic microflora enable to
salvage energy from food components that have not been digested in the upper GI
tract. Gibson et al. (1999) suggested that colonic bacterial fermentation provide
approximately 7% - 8% of total dietary energy that was required daily by the host.
Backhed et al. (2004) demonstrated that germ free mice require higher uptake of
calories to maintain normal body mass in comparison to normal mice.
Gut microflora also interacts with the host immune system in order to protect from
pathogenic bacteria and for the overall development and function of GI tract (Fig.
1.2B). Several gnotobiotic studies demonstrated the beneficial role of microflora in
influencing the physiological functions of gut. In comparison to normal animals,
reduction of digestive enzyme activity and local cytokine production were detected
in germ-free animals. Earlier studies suggested that the absence of indigenous
microflora modified the intestinal epithelial cells, which lead to the changes in
morphological and functional properties of germ free animals (Alam et al., 1994;
Wostmann, 1996). In germ free animals, development of GALT, lamina propria
cellularity and mucosal vascularity was also less effective. However, introducing
intestinal microflora into germ free mice helped the restoration of specific mucosal
immune system (Umesaki et al., 1995). Furthermore, germ-free mice demonstrated

8
the inhibition of macrophage chemotaxis and phagocytic activity and less number of
spleen-derived macrophage precursors (Wostmann, 1996; Nicaise et al., 1998).
Earlier studies reported that commensal bacterial ligands are important in the
development and function of gut mucosal immune system (Rakoff-Nahoum et al.,
2004; Mazmanian et al., 2005). These commensal bacteria are also capable of
influencing the regulation of T-helper (Th) cell type-1 or type-2 cytokines (Cebra,
1999; Shanahan, 2002a). Cytokines such as interleukin (IL)-1�, IL-2, tumor necrosis
factor (TNF)-� and transforming growth factor (TGF)-� increased the proliferation of
intestinal stromal cells (Fritsch et al., 1999).
Intestinal mucosal surfaces are vulnerable to various harmful microorganisms from
outside environment that transit through the intestine, which may be harmful to the
host. In order to protect the host from these harmful microorganisms, active secretory
gut immune system has evolved. The release of secretory immunoglobulin (sIg)-A
into the intestinal fluid occurs in healthy host, which is important in preventing the
adherence of foreign antigens to epithelial cells. This process is known as immune
exclusion, which provides protection to mucosal surfaces without causing
inflammatory responses. The gut microflora also acts as a barrier in protecting the
host from pathogenic bacteria by producing regulatory factors such as short-chain
fatty acids and bacteriocins. Cebra, (1999) suggested the antagonistic activity of
intestinal microflora in inhibiting the colonisation of pathogenic bacteria in gut. The
innate immune defence mechanisms of intestinal epithelial cells include mucins and
antimicrobial compounds such as lysozyme and defensins, which are important in

9
preventing the microbial adherence to epithelial cell surfaces (Magnusson and
Stjernstorm, 1982; Ouellette, 1999).
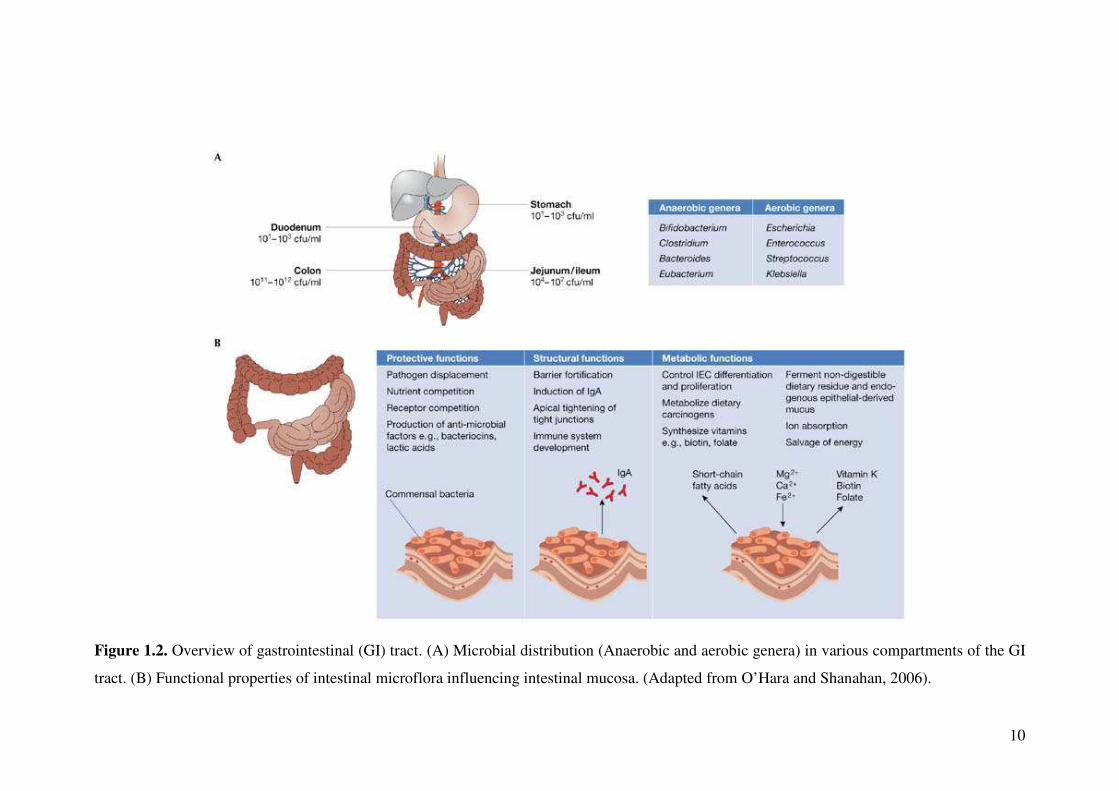
10
Figure 1.2. Overview of gastrointestinal (GI) tract. (A) Microbial distribution (Anaerobic and aerobic genera) in various compartments of the GI
tract. (B) Functional properties of intestinal microflora influencing intestinal mucosa. (Adapted from O’Hara and Shanahan, 2006).

11
1.2. GUT IMMUNE SYSTEM
In humans, gut is considered as a major immune organ. Gut mucosal surfaces are
constantly exposed to antigens derived from both diet and intestinal microflora. Gut
defence mechanisms have the capability of discriminating non-pathogenic
commensal bacteria and antigens from enteropathogens. Specifically, gut immune
responses towards antigens derived from commensal bacteria are non-harmful to the
organism, which further induce systemic tolerance against intestinal antigens. This
process can be expressed as immunological tolerance, which is important for gut
integrity. Earlier studies reported the mechanisms that are involved in immunological
tolerance (Schwartz, 1989; Garside et al., 1999). It was also suggested that the failure
of immunological tolerance towards luminal bacteria could lead to inflammatory
bowel disease (Duchmann et al., 1995).
The mucosal immune system is separated from gut lumen by a thin layer of intestinal
epithelial cells that lines the mucosal intestinal surface. These intestinal epithelial
cells are important in providing first line defence by preventing lumen antigens from
accessing the host immune system. However, antigens constantly interact with
immunosensory cells of the intestinal epithelium through different mechanisms (Fig.
1.3). The sampling of luminal antigens is performed through M-cell, which resides in
specialised epithelium that overlies the Peyer’s patches (Neutra et al., 1996). The
antigen presentation and affinity maturation of the transported luminal antigens occur
in Peyer’s patches (McGhee et al., 1999; Owen, 1999). In Peyer’s patches, activated
antigen-specific lymphocytes migrate to mesenteric lymph nodes and re-enter the
circulation through thoracic duct. Perdue, (1999) suggested that the transport of
antigen molecules can occur either through transcellular or paracellular pathways.

12
The other route of sampling antigens could be through the dendrites of intestinal
dendritic cells (Rescigno et al., 2001). The intestinal dendritic cells are also
important in inducing local immune responses towards commensal bacteria through
ingestion and retaining live commensal bacteria, which are further transported to
mesenteric lymph nodes (Macpherson and Uhr, 2004). This process suggests the role
of mesenteric lymph nodes as potential gatekeepers by preventing the access of
commensal bacteria to internal environment.
Lamina propria and intraepithelial spaces are considered as main effector sites in the
intestine. The lamina propria mainly consists of over 60% immune cells, which
include immunoglobulin (Ig)-A producing B-cells, T-cells and macrophages
(McGhee et al., 1999). In the lamina propria, B-cells undergo differentiation into
plasmacytes in order to secrete dimeric IgA. The intestinal epithelial cells are capable
of producing polymeric Ig receptors, which transport dimeric IgA into the intestinal
lumen (Underdown and Schiff, 1986). It is often described that plasma cells on the
mucosal surfaces as well as in the lamina propria are capable of secreting upto 40 mg
of IgA per day, which is an important characteristic for an active gut immune system.
In lamina propria, T-cells have the capability of interacting with B-cell
immunoglobulin isotype (Lebman et al., 1990; Shockett and Stavnezer, 1991). The
T-cells in lamina propria are mainly CD4+ T-helper lymphocytes, which express
CD40 ligands and release cytokines such as IL-4, IL-5 and IL-10 (Liu and Arpin,
1997). Furthermore, cytokines released from CD4+ T-cells and intestinal epithelial
cells modulate IgA producing B-cells, which are known to be important in mucosal
defence mechanism.
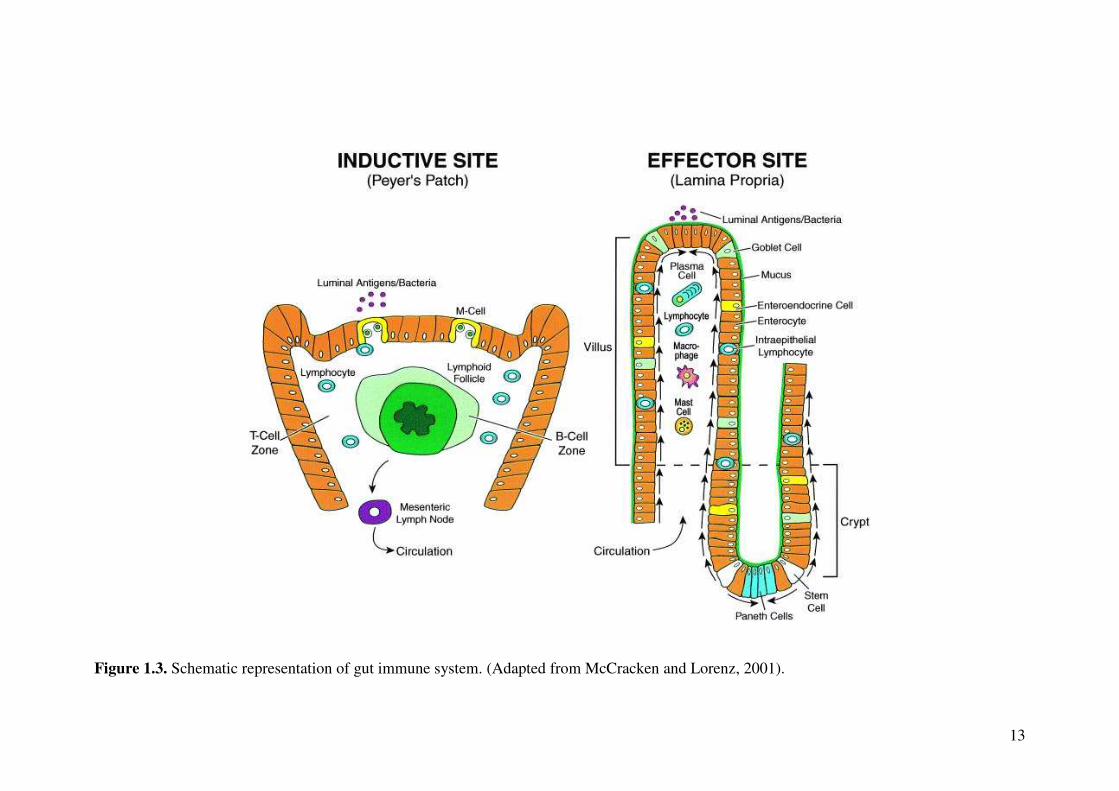
13
Figure 1.3. Schematic representation of gut immune system. (Adapted from McCracken and Lorenz, 2001).

14
1.3. PROBIOTICS
The term probiotic, meaning ‘for life’ was first coined in 1960s in order to describe
“substances secreted by one microorganism that stimulate the growth of another”
(Lilly and Stillwell, 1965). However, the role of these beneficial bacteria in
promoting positive health effects upon host has been fully recognised only in the last
two decades. Fuller, (1989) redefined the word probiotics as “viable microorganisms
that contribute to intestinal microbial balance and have the potential to improve the
health of their human host”. Recently, probiotics have been defined as “live
microorganisms which when administered in adequate amounts confer a health
benefit on the host” (Reid et al., 2003).
Probiotics are live microbial food supplements that have been used for many years in
animal feed industry. However, probiotics are now widely used in the manufacturing
of fermented dairy products. Of the numerous microbes that are beneficial
inhabitants of GI tract, microorganisms that have been considered as probiotics
include yeast (Guslandi et al., 2000), Escherichia coli (Tromm et al., 2004) and
enterococci (Lund and Edlund, 2001). The most commonly used have been
bifidobacteria and lactic acid bacteria (LAB) such as lactobacilli, lactococci and
streptococci (Madsen, 2001; Isolauri et al., 2002; Table 1.1). Lactobacilli are rod-
shaped bacteria, which are capable of producing short-chain fatty acids, acetate and
lactate through carbohydrate fermentation process. Several Lactobacillus species are
known to inhabit the intestinal tracts of humans and animals. In large intestine,
bifidobacteria constitute in large numbers among other beneficial microorganisms,
which are capable of synthesising vitamins and digestive enzymes (Gibson et al.,
1995). Generally, Lactobacillus and Bifidobacterium strains are used in the

15
development of probiotic products intended for human consumption due to the belief
that these bacteria are members of intestinal microflora and also considered as
“generally regarded as safe” (GRAS) (Berg, 1998; Klein et al., 1998).
The probiotic effects are strain-specific, therefore it is difficult to generalise the
beneficial effects of a probiotic strain even comparing to strains of the same species.
Recent Food and Agriculture Organisation (FAO) and World Health Organisation
(WHO) guidelines suggested that probiotic bacteria incorporated in food products
must demonstrate their tolerance to gastric juices and bile salts in order to survive
during their transit in GI tract (FAO/WHO, 2001; 2002). Additionally, probiotic
bacteria must be capable of colonising the GI tract and have the potential in
maintaining their efficacy during shelf-life of the product. Potential probiotic bacteria
possess some or all of the functional properties such as: (i) tolerance to acid, bile,
enzyme and oxygen; (ii) ability to adhere host epithelial cells; (iii) colonisation in the
GI tract; (iv) pathogen exclusion; (v) production of antimicrobial substances towards
pathogens; (vi) non-pathogenic and non-carcinogenic and (vii) favourable towards
well balanced microbial gut ecosystem. Several studies reported the influence of
probiotic bacteria on synthesis of various energy sources such as folic acid
production (Deeth and Tamime, 1981; Alm, 1982) and short-chain fatty acids
(Mallett et al., 1989). Furthermore, studies also showed that probiotic bacteria are
also capable of influencing various immune responses such as humoral, cellular and
non-specific immunity (Matsuzaki et al., 1998; Chiang et al., 2000; Cross et al.,
2002; Elahi et al., 2005; Takahashi et al., 2006).

16
Table 1.1. Microorganisms that are commonly considered as probiotics.
Lactobacillus spp.
Bifidobacterium spp.
Others
L. acidophilus B. adolescentis Enterococcus faecalis
L. brevis B. animalis Enterococcus faecium
L. casei B. breve Escherichia coli Nissle
L. crispatus B. bifidum Saccharomyces boulardii
L. curvatus B. infantis Streptococcus cremoris
L. delbrueckii subsp.
bulgaricus
B. lactis Streptococcus diacetylactis
L. fermentum B. longum Streptococcus intermedius
L. gasseri B. thermophilum Streptococcus salivarius
L. johnsonii Streptococcus thermophilus
L. lactis
L. paracasei
L. plantarum
L. reuteri
L. rhamnosus

17
1.4. IMMUNE RESPONSES OF LACTIC ACID BACTERIA
Certain strains of LAB have the potential in contributing towards host defence
mechanisms of GI tract (Deplancke and Gaskins, 2001) through interactions with the
immune system (Lu and Walker, 2001). Several factors such as survival, adherence,
colonisation and sites of interaction in GI tract influence the ability of immune
enhancing properties of LAB. Earlier studies reported that probiotic bacteria are
capable of influencing host cell functions such as immune responses and intestinal
barrier integrity (Rosenfeldt et al., 2004; Sartor, 2005). The capability of dendritic
cells in recognising bacterial antigens also helps in regulating T-cell responses. In
vitro studies demonstrated the differential cytokine expression of murine dendritic
cells influenced by LAB and probiotic cocktail VSL#3, which favoured Th1, Th2 or
Th3 immune responses (Christensen et al., 2002; Hart et al., 2004). Particularly,
Lactobacillus strains demonstrated the regulation of IL-10 producing T-cells through
interaction with dendritic cell-specific intracellular adhesion molecule 3-grabbing
nonintegrin (Smits et al., 2005).
Epithelial cells of the intestine and colon also play an important role in immune
mechanisms. Zhang et al. (2005) reported that Caco-2 cells cultured with L.
rhamnosus Goldin-Gorbach (GG) regulated NF-�B signalling pathway to reduce
TNF-� induced IL-8 production. Lactobacillus reuteri and VSL#3 also demonstrated
their influence on NF-�B signalling pathway (Ma et al., 2004; Petrof et al., 2004).
Probiotic bacteria are also capable of strengthening the intestinal barrier. For
example, probiotic E. coli Nissle 1917 influenced the regulation of NF-�B signalling
pathway by inducing the expression of antimicrobial peptide �-defensin-2 (Wehkamp

18
et al., 2004), which could be important in enhancing intestinal barrier functions such
as inhibiting the growth of pathogenic bacteria.
Several reports have suggested that early exposure of live microorganisms with
dietary antigens could be helpful in developing gut barrier functions (Helgeland et
al., 1996; Sudo et al., 1997). Moreau et al. (1978) reported the development of gut
barrier through increased duodenal IgA plasmocyte cells. Peyer’s patches are known
to be important in sampling luminal bacterial antigens where they interact with
antigen-presenting cells carrying major histocompatibility complex (MHC) class II
(Weiner et al., 1994). The antigens that are transported through Peyer’s patches are
considered important towards the overall development of local secretory immune
responses. Subsequently, immune responses initiated in GALT can influence
immune responses at other mucosal sites. In vitro cell culture models have been used
to demonstrate the immune responses induced by non-pathogenic bacteria. As an
example, Haller et al. (2000) reported the effects of non-pathogenic bacteria in
inducing cytokines such as IL-8, IL-10 and TNF-� by Caco-2 cells.
Many investigators reported the influence of LAB in inducing sIgA responses in
GALT. Orally administered L. acidophilus and L. casei increased the sIgA levels in
small intestine of mice (Perdigon et al., 1995). In mice, L. acidophilus and
Bifidobacterium species modulated the mucosal and systemic immunity through
increased IgA responses to cholera toxin, which was used as an oral immunogen
(Tejada-Simon et al., 1999a). Lactobacillus GG also showed the capability in
increasing IgA response in children with Crohn’s disease (Malin et al., 1996) and
acute rotavirus diarrhoea (Kaila et al., 1992). Moreover, several studies also reported

19
the capability of probiotic bacteria in modulating IgE production. For example,
decrease in the production of IgE in serum was detected in mice orally fed with L.
casei Shirota (Matsuzaki et al., 1998). Similar results related to the reduction of IgE
production was also observed in IL-10 deficient mice fed with L. plantarum 229v
(Schultz et al., 2002).
The regulation of both innate and acquired immune responses was mediated through
cytokines. Several studies reported the influence of LAB in regulating T-cell
functions. As an example, cell wall and cytoplasmic components of LAB
demonstrated the capability of stimulating cytokines such as IL-6 and TNF-� and
nitric oxide (NO) (Tejada-Simon and Pestka, 1999b). Maassen et al. (2000) also
reported an increase of TNF-�, IL-2 and IL-1� in gut villi of mice fed with L. reuteri
or L. brevi. Furthermore, Lactobacillus strains demonstrated the enhancement of
TNF-� and IFN-� producing cells in the lamina propria of gut (Perdigon et al., 2002).
The cytokine, IFN-� is capable of enhancing nitric oxide production, activating
macrophages and NK cells. The phagocytic activity of macrophages plays an
important role in preventing microbial infections, whereas NK cells are pivotal in
defence against viral infections and cancer. Schiffrin et al. (1995) demonstrated the
supplementation of L. johnsonii La1 or B. lactis Bb12 for 3 weeks, which increased
the phagocytic activity of peripheral blood leukocytes in humans. Interestingly, the
influence of probiotic bacteria differ in the modulation of phagocytic activity in
healthy and allergic subjects (Pelto et al., 1998). Furthermore, Donnet-Hughes et al.
(1999) also reported the potential of LAB on phagocytic activity of the peripheral
blood leukocytes in healthy adult volunteers.

20
In general, probiotic bacteria can be regarded as a dietary adjuvant, which
beneficially affect the host physiology through modulating mucosal and systemic
immune functions. Lactic acid bacteria have been considered as potential candidates
for oral vaccination purposes due to their various functional properties and long
history of usage as safe food grade organisms. Maassen et al. (2000) demonstrated
the differences between Lactobacillus strains in inducing cytokines and their
adjuvant properties in gut. Although several studies reported the adjuvant effects of
LAB with various antigens, adjuvant characteristics of Lactobacillus strains often
vary due to the differences in bacterial strains, experimental designs, animals and
antigens (Tejada-Simon et al., 1999a; Cano et al., 2002; Plant and Conway, 2002; He
et al., 2005; Kim et al., 2005).
1.5. THERAPEUTIC EFFECTS OF LACTIC ACID BACTERIA
Lactic acid bacteria have the potential to induce several beneficial effects on human
health, which are listed in Table 1.2. The insights into the therapeutic effects of LAB
in GI tract and their effects on host immune functions are discussed below.
1.5.1. Diarrhoeal diseases
Certain LAB are proven to be beneficial in the prevention or amelioration of various
diarrhoeal diseases. Several human intervention studies reported the probiotic effects
of L. rhamnosus GG against rotavirus diarrhoea in infants (Isolauri et al., 1991; Kaila
et al., 1992; Majamaa et al., 1995). Szajewska et al. (2001) demonstrated the
capability of L. rhamnosus GG in reducing the duration of diarrhoea in comparison
to placebo. Additionally, L. rhamnosus GG prevented the urease-producing bacterial
growth during rotavirus diarrhoea (Isolauri et al., 1994). Recently, Paton et al. (2005)

21
demonstrated the treatment and prevention of E. coli diarrhoea through a toxin
binding recombinant probiotic. Probiotic bacteria also showed the capability of
producing bacteriocins such as nisin in order to inhibit the growth of pathogenic
bacteria (Dodd and Gasson, 1994; Miraglia del Giudice and De Luca, 2004).
1.5.2. Inflammatory bowel disease
Inflammatory bowel disease (IBD) has a complex etiology, which was characterised
by chronic or recurrent intestinal inflammation. Ulcerative colitis and Crohn’s
disease are the best known forms of IBD. The mechanisms responsible for the cause
of IBD still remain unclear. Several studies suggested that certain probiotic bacteria
are capable of treating mild or moderately active IBD (Fedorak and Madsen, 2004;
Sartor, 2004). Particularly, Pathmakanthan et al. (1999) reported the decrease of
lactobacilli in the inflamed mucosal tissues of ulcerative colitis patients. Therefore,
restoration of microflora in GI tract through selected probiotic bacteria could be
considered as a therapeutic strategy for ameliorating intestinal inflammation.
1.5.3. Irritable bowel syndrome
Irritable bowel syndrome (IBS) is associated with manifestation of abdominal pain,
bloating, flatulence and diarrhoea. Several studies suggested the role of colonic
microflora in pathogenesis of IBS (Bradley et al., 1987; Madden et al., 2001).
However, the cause of IBS is still unknown. In double blind and placebo controlled
studies, orally administered L. plantarum reduced flatulence and abdominal pain
(Nobaek et al., 2000; Niedzielin et al., 2001). Brigidi et al. (2001) also reported the
beneficial effects of VSL#3 by improving the clinical picture and altering the faecal

22
microflora composition with an increase of lactobacilli, bifidobacteria and S.
thermophilus in patients having IBS. In another study, L. plantarum did not either
modify the colonic fermentation process or improve symptoms in patients with IBS
(Sen et al., 2002). Recently, O’Mahony et al. (2005) suggested the
immunomodulatory role of B. infantis by improving the symptoms in patients with
IBS. However, further studies are needed to define the mechanisms of probiotic
bacterial influence on IBS.
1.5.4. Helicobacter pylori infections
Helicobacter pylori are responsible for causing chronic gastritis and idiopathic peptic
ulcer disease. Previous studies showed the potential of probiotic bacteria in inhibiting
the growth of H. pylori (Midolo et al., 1995; Kabir et al., 1997). Aiba et al. (1998)
showed the capability of L. salivarius in inhibiting the growth of H. pylori by
producing high amounts of lactic acid. It was postulated that higher levels of lactic
acid production could be helpful in reducing the urease activity of H. pylori. It has
also been reported that the combination of probiotic cultures such as L. acidophilus
La5 and B. latics Bb12 showed significant decrease in the urease activity after 6
weeks of oral administration in humans with H. pylori (Wang et al., 2004). In
contrast, a probiotic combination of L. rhamnosus GG, L. rhamnosus LC705, B.
breve Bb99 and Propionibacterium freudenreichii subsp. shermanii JS were not
effective in improving intestinal inflammation symptoms in humans with H. pylori
(Myllyluoma et al., 2005). Further studies are needed to determine the etiology of the
disease and well defined experimental methodologies could possibly substantiate the
efficacy of probiotic bacteria.

23
The mechanisms of probiotic bacterial interaction with the host need to be
investigated to elucidate their potential role especially in promoting human health
and well-being. Earlier studies attempted to define the cellular targets of probiotic
bacteria, whereas the molecular basis of the probiotic functions remains unknown.
The emerging demand of probiotic bacteria in functional food applications require
fundamental understanding of the molecular structures that are involved in
microbial-host interactions. Moreover, genomic approaches are considered necessary
in identifying the key gene targets of probiotic functions.
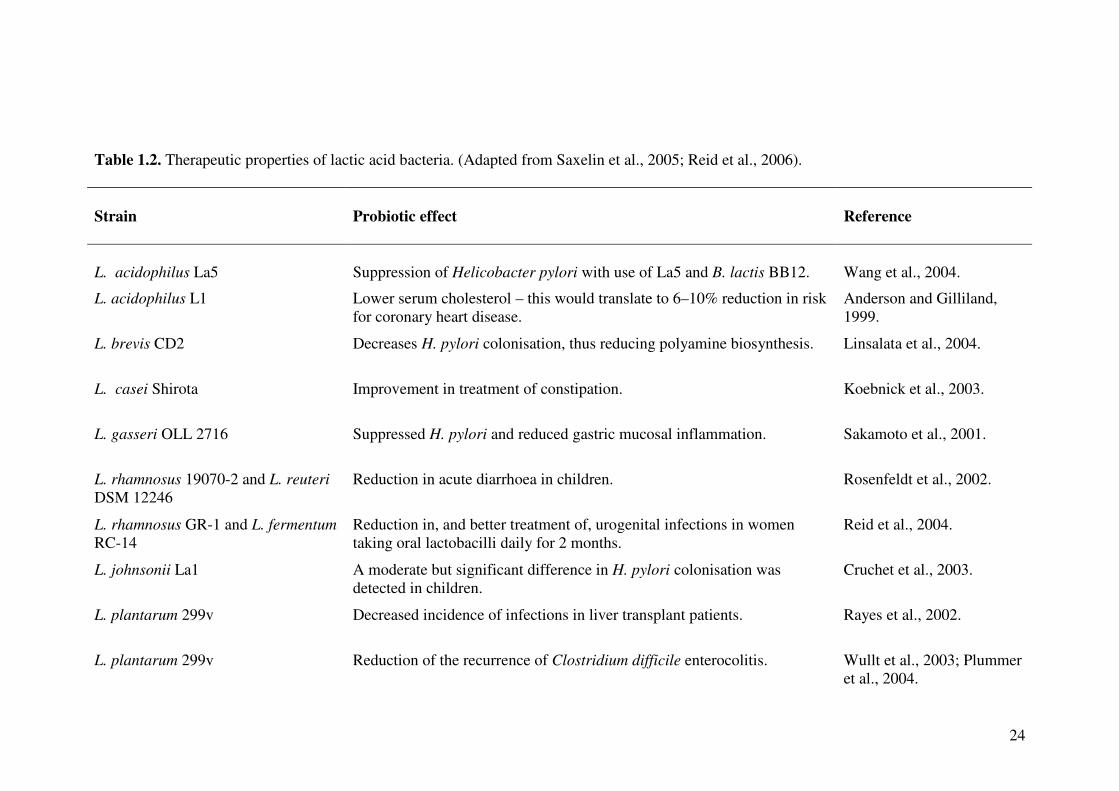
24
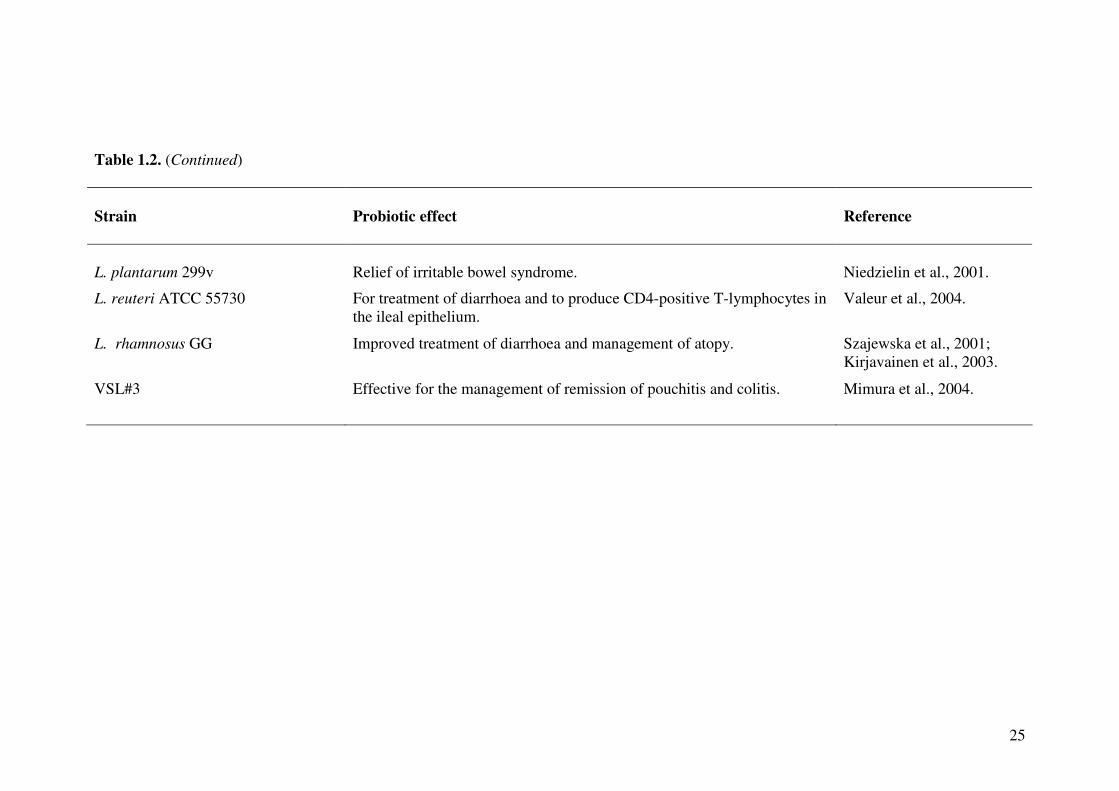
Table 1.2. Therapeutic properties of lactic acid bacteria. (Adapted from Saxelin et al., 2005; Reid et al., 2006).
Strain
Probiotic effect
Reference
L. acidophilus La5
Suppression of Helicobacter pylori with use of La5 and B. lactis BB12.
Wang et al., 2004.
L. acidophilus L1 Lower serum cholesterol – this would translate to 6–10% reduction in risk for coronary heart disease.
Anderson and Gilliland, 1999.
L. brevis CD2 Decreases H. pylori colonisation, thus reducing polyamine biosynthesis. Linsalata et al., 2004.
L. casei Shirota Improvement in treatment of constipation. Koebnick et al., 2003.
L. gasseri OLL 2716 Suppressed H. pylori and reduced gastric mucosal inflammation. Sakamoto et al., 2001.
L. rhamnosus 19070-2 and L. reuteri DSM 12246
Reduction in acute diarrhoea in children. Rosenfeldt et al., 2002.
L. rhamnosus GR-1 and L. fermentum RC-14
Reduction in, and better treatment of, urogenital infections in women taking oral lactobacilli daily for 2 months.
Reid et al., 2004.
L. johnsonii La1 A moderate but significant difference in H. pylori colonisation was detected in children.
Cruchet et al., 2003.
L. plantarum 299v Decreased incidence of infections in liver transplant patients. Rayes et al., 2002.
L. plantarum 299v Reduction of the recurrence of Clostridium difficile enterocolitis. Wullt et al., 2003; Plummer et al., 2004.
25
Table 1.2. (Continued)
Strain
Probiotic effect
Reference
L. plantarum 299v
Relief of irritable bowel syndrome.
Niedzielin et al., 2001.
L. reuteri ATCC 55730 For treatment of diarrhoea and to produce CD4-positive T-lymphocytes in the ileal epithelium.
Valeur et al., 2004.
L. rhamnosus GG Improved treatment of diarrhoea and management of atopy. Szajewska et al., 2001; Kirjavainen et al., 2003.
VSL#3 Effective for the management of remission of pouchitis and colitis. Mimura et al., 2004.

26
1.6. FUNCTIONAL GENOMICS AND LACTIC ACID BACTERIA
The word “functional genomics” constitute genomics, transcriptomics, proteomics,
metabolomics and bioinformatics, which can also be termed as systems biology. The
incorporation of functional genomics in understanding the complex intestinal ecosystem
is gaining momentum to elucidate the relationship between diet and human health.
However, understanding the role of nutrients in modulating gene and protein expression
and subsequently their influence on cellular and metabolic functions are necessary in
preventing diet-related diseases.
Molecular approaches have demonstrated that lactobacilli was able to survive through its
intestinal passage and altered the microbial population and diversity of Lactobacillus
species, but did not induce any adverse effects on dominant microflora (de Vos et al.,
2004; Zoetendal et al., 2004). Similarly, molecular techniques also showed that L. casei
and L. plantarum are active during the transit from stomach to colon in mice and played
an important role in protein synthesis (Bron et al., 2004; Oozeer et al., 2004).
Mechanisms such as autoaggregation and adhesion were considered as potential
properties of LAB in colonising the GI tract. Roos et al. (1999) reported that genes
coding for aggregation helicase and maltose-binding proteins are necessary in increasing
the autoaggregation ability of L. reuteri 1063. Earlier studies demonstrated that
lipoteichoic acid was involved in the adherence of L. johnsonii La1 to intestinal Caco-2
cells (Granato et al., 1999). The genome sequencing of L. johnsonii La1 uncovered the
genes that were involved in lipoteichoic acid synthesis and cell surface molecules
(Vaughan and Mollet, 1999). Furthermore, Mack et al. (1999) hypothesised the role of
L. plantarum 299v and L. rhamnosus GG in inhibiting the adherence of pathogenic E.

27
coli to HT-29 intestinal cells through increased expression of intestinal mucin genes
such as MUC2 and MUC3.
Several in vivo studies reported the influence of diet on gene expression. As an example,
Rao et al. (2001) showed that mice fed with low diet in selenium decreased the
expression of genes involved in detoxification and increased the expression of genes that
were involved in DNA damage processing, oxidative stress and cell-cycle control.
Similarly, magnesium deficiency up-regulated the genes involved in protection and
repair of oxidative stress in thymocytes of rat (Petrault et al., 2002). Blanchard et al.
(2001) demonstrated the potential of cDNA microarray analysis in revealing the
intestinal gene expression changes that occurred in early stages of zinc deficiency in
rats. Furthermore, Hooper et al. (2001) demonstrated the capability of commensal
bacteria in modulating gene expression that are involved in various intestinal functions
such as nutrient absorption, mucosal barrier fortification and xenobiotic metabolism.
The use of conventional methods restricted the understanding to a particular gene
function, whereas the DNA microarray technique was capable of unravelling the overall
gene expression patterns of the cell (Brown and Botstein, 1999; Khan et al., 1999).
Whole genome analysis of gene expression in GI tract could further elucidate the
microbial-host interactions. Earlier, several studies reported the beneficial effects of
LAB on various biological functions of host, however applications of novel molecular
approaches such as microarrays in combination with functional assays is necessary to
unravel new target’s that were influenced by the probiotic bacteria.

28
1.7. AIMS
The overall aim of this study was to characterise the in vitro probiotic properties of L.
acidophilus LAFTI L10 and L. paracasei LAFTI L26 and evaluate their influence on
mucosal and systemic immune responses and gene expression in mice.
The specific aims of this study were:
1. Characterisation of Lactobacillus and Bifidobacterium species for potential
probiotic strains (Chapter 2).
2. Examination of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26
influence on mucosal and systemic immunity and bacterial translocation in mice
(Chapter 3).
3. Determination of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 effects
on mucosal and systemic immune responses in cholera toxin mice model
(Chapter 4).
4. Investigation of L. acidophilus LAFTI L10 influence on gene expression in the
small bowel of normal and cholera toxin mice (Chapter 5).

29
CHAPTER 2
Selection and characterisation of Lactobacillus and Bifidobacterium species for
potential probiotic strains

30
2.1. ABSTRACT
The present study aimed to investigate the in vitro probiotic properties of Lactobacillus
and Bifidobacterium strains. A total of 12 strains were screened towards their tolerance
to various acidic conditions. Following an initial screening, 6 strains were selected to
further characterise their probiotic properties such as tolerance to bile salts,
hydrophobicity, autoaggregation and coaggregation. The overall reduction in cell
survivability of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 in different pH
conditions was less compared to other bacterial strains. Particularly, these strains
showed high tolerance to pH 2.0 with more than 105 CFU/ml in viability. Lactobacillus
acidophilus LAFTI L10 and L. paracasei LAFTI L26 demonstrated high survivability
especially in 1.0% bile salts when compared with other bacterial strains. A correlation
between hydrophobicity and autoaggregation ability was observed among the bacterial
strains. In contrast, B. lactis Bb12, demonstrated high degree of hydrophobicity, whereas
autoaggregation ability was lower compared to other bacterial strains. Furthermore,
coaggregation of L. acidophilus LAFTI L10 with other bacterial strains showed
significant differences in their aggregation capabilities compared to L. paracasei LAFTI
L26. Overall, L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 showed better
tolerance to acid and bile conditions and also their surface adherence characteristics
could be considered for their development as potential probiotic strains.

31
2.2. INTRODUCTION
Probiotic bacteria are commonly referred to as beneficial microbes, which are frequently
used as active ingredients in fermented milk products. The use of probiotic bacteria in
food products has been recently increased due to their potential attributes such as
contributing to host defence mechanisms and improving the microbial balance of the GI
tract. In order to exert beneficial effects on the host, probiotic bacteria are required to
survive in enough numbers while transiting through constantly changing gastric and bile
conditions of GI tract. The general acidity of stomach ranges from pH 2.5 to 3.5
(Holzapfel et al., 1998), however the pH of stomach could also be as low as pH 1.5
(Lankaputhra and Shah, 1995) or as high as pH 6.0 or higher after the intake of food
(Johnson, 1977). There were no specific pH conditions or in vitro methods set to screen
potential probiotic strains, however a wide range of pH conditions and in vitro methods
were used to screen the acid tolerance of Lactobacillus and Bifidobacterium strains
(Conway et al., 1987; Lankaputhra and Shah, 1995; Charteris et al., 1998; Chou and
Weimer, 1999; Chung et al., 1999; Zarate et al., 2000).
Several studies reported the screening of Lactobacillus and Bifidobacterium strains
tolerance to bile salts (Lankaputhra and Shah, 1995; Prasad et al., 1998; Chung et al.,
1999; Truelstrup Hansen et al., 2002). In the small intestine, transit of food may take
between 1 – 4 h (Smith, 1995), which can have a pH of around 8.0 (Keele and Neil,
1965). Bile concentrations ranging from 0.3% – 1.0% were used for in vitro screening to
identify potential probiotic strains that were resistant to bile salts (Prasad et al., 1998;
Jacobsen et al., 1999; Truelstrup Hansen et al., 2002). Goldin and Gorbach (1992)

32
suggested that bile salts of 0.15% – 0.3% concentrations were suitable for screening
Lactobacillus and Bifidobacterium strains for human consumption.
Microbial adhesion to intestinal cells is considered as a potential characteristic of
probiotic bacteria, which could enable them to colonise the GI tract and induce potential
health effects (Pedersen and Tannock, 1989; Alander et al., 1997). In order to
demonstrate a potential probiotic strain, adhesion ability is considered as one of the
screening criteria for selecting probiotic bacteria (Fogh et al., 1977). The in vivo
bacterial adhesion studies involve difficulties to characterise the interactions especially
in humans, which led to the development of several in vitro methods as model systems
for screening potential adherent probiotic bacteria (Mayra-Makinen et al., 1983; Conway
and Kjellberg, 1989; Ouwehand et al., 2001). Earlier studies showed the in vitro
adherence capability of probiotic bacteria to Caco-2 cells (Kimoto et al., 1999; Todoriki
et al., 2001; Fernandez et al., 2003; Lee et al., 2003). However, several studies suggested
indirect in vitro methods to characterise the adhering properties of probiotic bacteria that
demonstrated the correlation between cell adhesion ability, autoaggregation and cell
surface hydrophobicity (Wadstrom et al., 1987; Marin et al., 1997; Del Re et al., 1998;
Perez et al., 1998; Del Re et al., 2000; Kos et al., 2003; Pan et al., 2006).
The adhesion of bacterial cells is a multi-step process that involves physical contact with
cell surface, which could be influenced by structural composition of the cell wall
membrane and interacting surfaces. The overall adherence of microorganisms also
depends upon non-specific physical interactions between the two surfaces, which enable
them to interact specifically between adhesins and complementary receptors (Freter,

33
1992; Rojas and Conway, 1996; Perez et al., 1998). Previous studies suggested that
several factors contribute the adhesion of probiotic bacteria to host epithelial cells, such
as cell surface hydrophobicity and autoaggregation (Cesena et al., 2001; Ehrmann et al.,
2002; Ventura et al., 2002; Kos et al., 2003), lipoteichoic acids (Sherman and Savage,
1986; Granato et al., 1999) and cell wall proteins (Walter et al,., 2005). Autoaggregation
demonstrated its importance in adhesion to intestinal cell wall, whereas coaggregation
was crucial to form as a barrier to prevent the colonisation of pathogenic
microorganisms (Boris et al., 1997; Reid et al., 1988; Del Re et al., 2000).
Autoaggregation and coaggregation properties were considered to be important for
probiotic bacteria to persist in different locations of mucosal surfaces (Jankovic et al.,
2003). Castagliuolo et al. (2005) reported the importance of aggregation characteristics
of L. crispatus in exerting protective effects in colitis mice model.
It was reported previously, that cell surface properties such as hydrophobicity may affect
autoaggregation and adhesion ability of the bacteria (Wadstrom et al., 1987; Perez et al.,
1998; Del Re et al., 2000). It is well known that attractive forces between non-polar
surfaces in water solutes control the hydrophobic interactions. However, cell surface
structures of bacteria may have the potential to undergo physiochemical changes to alter
the hydrophobic ability in response to environmental conditions such as pH, bile salts or
ionic strength.
In the present study, tolerance to low pH and high bile conditions, hydrophobicity,
autoaggregation and coaggregation ability were considered as potential probiotic
characteristics (Prasad et al., 1998; Del Re et al., 2000; Kos et al., 2003; Castagliuolo et

34
al., 2005; Otero et al., 2006; Bujalance et al., 2007). Initially, 12 strains of Lactobacillus
and Bifidobacterium species were screened for their tolerance to simulated gastric juice
at various pH levels (pH 2.0, 3.0, 4.0 and 6.5). Based on acid tolerance, 6 strains were
selected to further investigate their probiotic properties such as tolerance to bile salts
(0.5% and 1.0%), hydrophobicity, autoaggregation and coaggregation.

35
2.3. MATERIALS AND METHODS
2.3.1. Microorganisms and growth conditions
The bacterial strains used in this study were obtained from various suppliers in
Australia. Lactobacillus acidophilus ASCC 2400, Lactobacillus casei subsp. casei
ASCC 2603, Lactobacillus paracasei subsp. paracasei ASCC 5437, Lactobacillus
rhamnosus ASCC 2625, Bifidobacterium breve ASCC 1900, Bifidobacterium bifidum
ASCC 1903, Bifidobacterium infantis ASCC 1912 and Bifidobacterium longum ASCC
5188 were obtained from Australian Starter Culture Research Centre (Werribee, VIC,
Australia). DSM Food Specialties (Moorebank, NSW, Australia) provided Lactobacillus
acidophilus LAFTI L10, Lactobacillus paracasei LAFTI L26 and Bifidobacterium
animalis subsp. lactis LAFTI B94. Bifidobacterium animalis subsp. lactis Bb12 was
supplied by Chr. Hansen, Bayswater, VIC, Australia.
All bacterial strains were grown anaerobically in gas jars using GasPak System (Oxoid,
Adelaide, Australia) for 24 h at 37°C in de Man, Rogosa and Sharpe (MRS) broth
(Oxoid, Adelaide, Australia). The bacterial cells were harvested at 4000 x g for 15 min
at 4°C and washed twice with sterile 0.01 M phosphate buffered saline (PBS) solution.
The cell viability of the bacterial cultures was determined by spread plate count on MRS
agar, which was incubated under anaerobic conditions at 37°C for 48 h.
2.3.2. Acid tolerance
The acid tolerance of Lactobacillus and Bifidobacterium strains was determined by
using milk-based medium (NGYC medium: 12% non-fat skim milk, 2% glucose, 1%

36
yeast extract and 0.05% L-cysteine) as described by Lankaputhra and Shah, (1995). The
NGYC medium was adjusted to pH 2.0, 3.0, 4.0 or 6.5 (Control) with 5 M HCL or 1 M
NaOH. The bacterial cell suspensions were added to the pH adjusted NGYC medium,
which were then incubated anaerobically for 3 h at 37°C. The bacterial survivability was
determined by spread plate count on MRS agar after incubated anaerobically at 37°C for
48 h. The average and standard error were estimated from 6 independent experiments.
The overall reduction in the viability of bacterial strains in different pH conditions was
calculated using the formula:
Overall reduction of viability = (pH 6.5 – pH 4.0) + (pH 6.5 – pH
3.0) + (pH 6.5 – pH 2.0)
2.3.3. Bile tolerance
The bile tolerance of Lactobacillus and Bifidobacterium strains was determined by using
milk-yeast extract medium as described by Truelstrup Hansen et al. (2002). The milk-
yeast extract medium (pH 6.9) consists of 10% non-fat skim milk powder, 0.5% yeast
extract, 0.05% L-cysteine and 0% (Control), 0.5% or 1.0% (w/v) bile salts (Oxgall,
Sigma, Australia). The bacterial cell suspensions were inoculated into the milk-yeast
extract medium and incubated anaerobically for 6 h at 37°C.
The survival of the bacterial cells was determined by spread plate count on MRS agar
after anaerobically incubated at 37°C for 48 h. The average and standard error were

37
estimated from 6 independent experiments. The overall reduction in the viability of the
bacterial strains in different bile concentrations was calculated using the formula:
Overall reduction of viability = (0% - 0.5%) + (0% - 1.0%)
2.3.4. Hydrophobicity assay
The adhesion of bacterial strains to hydrocarbons was determined according to Perez et
al. (1998). Briefly, bacterial cultures were grown anaerobically for 24 h at 37°C in MRS
broth and harvested at 4000 x g for 15 min at 4°C followed by washing twice with
sterile 0.01 M PBS solution. Bacterial cells were resuspended in 0.01 M PBS and
absorbance (A0) was read at 600 nm. A volume of 0.6 ml of n-hexadecane (Merck,
Melbourne, Australia) was added to 3-ml aliquots of bacterial cell suspension. The
samples were mixed thoroughly by vortexing for 2 min. The 2-phase separation was
observed by incubating the samples at 37°C for 1 h. Aqueous phase was removed and
absorbance (A) was measured at 600 nm. Hydrophobicity assay was repeated 6 times to
estimate the average and standard error. The percentage of cell surface hydrophobicity is
expressed as:
[ ] 100x/)(%)(cityHydrophobi OO AAAH −=
Where A0 and A represents the absorbance of aqueous phase before and after contact
with n-hexadecane.

38
2.3.5. Autoaggregation and coaggregation assays
The bacterial strains were grown anaerobically for 24 h at 37°C in MRS broth and
harvested at 4000 x g for 15 min at 4°C followed by washing twice with sterile 0.01 M
PBS solution. The bacterial cells were resuspended in 0.01 M PBS, which were used to
perform autoaggregation and coaggregation assays.
Autoaggregation assay was performed according to Kos et al. (2003). The bacterial cell
suspensions of 4-ml aliquots were mixed thoroughly for 10 s and incubated at 37°C for 6
h. The absorbance (A) of the upper suspension (0.1 ml) was measured at 600 nm.
Autoaggregation assay was repeated 6 times to estimate the average and standard error.
The autoaggregation percentage is expressed as:
100x)/(1(%)ationAutoaggreg ot AA−=
Where At represents the absorbance at 6 h and A0 represents the absorbance at 0 h.
Coaggregation assay was performed according to Handley et al. (1987). Briefly, equal
volumes (2-ml aliquots) of different bacterial cell suspensions were mixed together by
vortexing for 10 s, while the control tubes consist of 4-ml aliquots of individual bacterial
cell suspensions. The mixed bacterial cell suspensions and the control tubes were
incubated at 37°C for 6 h and absorbance (A) of the upper suspensions (0.1 ml) was
measured at 600 nm. Coaggregation assay was repeated 6 times to estimate the average
and standard error.

39
The coaggregation percentage is expressed as:
( )( )
100x2/yx
)yx()2/)yx((%ionCoaggregat
AA
AAA
+
+−+=
Where Ax and Ay represent individual bacterial strains in the control tubes and A(x + y)
represents the mixture of bacterial strains.
2.3.6. Statistical analysis
The results were expressed as mean ± standard error of mean (SEM). The data were
analysed using the one-way analysis of variance procedure of SPSS (Version 12.0.1).
Significant differences between groups were identified by Duncan’s multiple range test
(SPSS, Version 12.0.1).

40
2.4. RESULTS
2.4.1. Tolerance to simulated gastric juice
The effect of various gastric conditions on Lactobacillus and Bifidobacterium strains is
shown in Table 2.1. All the bacterial strains showed differences in cell viability in
various pH conditions for 3 h. Lactobacillus acidophilus LAFTI L10, L. paracasei
LAFTI L26 and B. lactis LAFTI B94 strains were tolerant to acid at pH 2.0 with a
reduction of 2.95 to 3.89 log colony-forming units (CFU)/ml. Particularly, L.
acidophilus LAFTI L10 and L. paracasei LAFTI L26 were tolerant to pH 2.0 with high
survival at 105 CFU/ml, whereas B. lactis LAFTI B94, B. infantis ASCC 1912, B. lactis
Bb12 and L. acidophilus ASCC 2400 showed the capability of surviving at 104 CFU/ml
after 3 h of incubation. At pH 2.0, L. rhamnosus ASCC 2625, B. bifidum ASCC 1903, B.
longum ASCC 5188, L. paracasei ASCC 5437 and B. breve ASCC 1900 showed less
survival ranging from 101 to 103 CFU/ml. Furthermore, there was no survival of L. casei
ASCC 2603 after incubating for 3 h at pH 2.0.
The ability of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, L. lactic LAFTI
B94, B. infantis ASCC 1912, B. lactis Bb12 and L. acidophilus ASCC 2400 to survive at
pH 3.0 was more than 106 CFU/ml after incubation for 3 h. All other Lactobacillus and
Bifidobacterium strains also showed tolerance to pH 3.0 with survival ranging from 103
to 105 CFU/ml. Particularly at pH 3.0, L. acidophilus LAFTI L10 demonstrated greater
tolerance with less reduction in cell viability by 1.95 CFU/ml, which was followed by L.
paracasei LAFTI L26 and B. infantis ASCC 1912 by a decrease of 2.5 CFU/ml. In pH
4.0, L. acidophilus LAFTI L10 showed higher tolerance with a decrease of 1.25

41
CFU/ml. Similarly, B. infantis ASCC 1912, B. lactis Bb12 and L. acidophilus ASCC
2400 also showed high tolerance to pH 4.0 with more than 107 CFU/ml, whereas L.
paracasei LAFTI L26, L. lactic LAFTI B94 and L. casei ASCC 2603 survived at 106
CFU/ml. The other Lactobacillus and Bifidobacterium strains showed survival of 105
CFU/ml at pH 4.0 (Table 2.1). All the Lactobacillus and Bifidobacterium strains that
were screened for their tolerance to pH 2.0, pH 3.0 and pH 4.0 showed significant
variations (P < 0.01) in their cell viability when compared with pH 6.5 (Control).
Based on the survival of Lactobacillus and Bifidobacterium strains at various pH
conditions after incubating for 3 h, overall reduction of cell viability was calculated to
determine the potential probiotic strains. Lactobacillus acidophilus LAFTI L10, L.
paracasei LAFTI L26, L. lactic LAFTI B94, B. infantis ASCC 1912, B. lactis Bb12 and
L. acidophilus ASCC 2400 exhibited greater tolerance to various pH conditions,
particularly at pH 2.0 with survival more than 104 CFU/ml. As shown in Table 2.1, top 6
bacterial strains were selected to further investigate their probiotic properties.
2.4.2. Tolerance to bile salts
The effect of bile salts on the growth of Lactobacillus and Bifidobacterium strains are
shown in Table 2.2. The exposure of bile salts at 0.5% and 1.0% for 6 h was less
detrimental to L. paracasei LAFTI L26, which demonstrated less reduction in overall
cell viability. All Lactobacillus and Bifidobacterium strains in 1.0% bile salts showed
less than 1.0 CFU/ml reduction in survival with exception to L. acidophilus ASCC 2400,
which exhibited a decrease of 1.15 CFU/ml in cell viability. The resistance of

42
Lactobacillus and Bifidobacterium strains towards 0.5% bile salts also showed reduction
in cell survival, which was not higher than 1.0 CFU/ml. The overall reduction of cell
viability in different bile salts demonstrated that L. paracasei LAFTI L26 was a
potential probiotic strain with high tolerance to bile salts, whereas B. lactis LAFTI B94
and L. acidophilus LAFTI L10 showed similar overall reduction of survival at different
bile salts concentrations. However, L. acidophilus LAFTI L10 showed better tolerance
to 1.0% bile salts compared to B. lactis LAFTI B94.
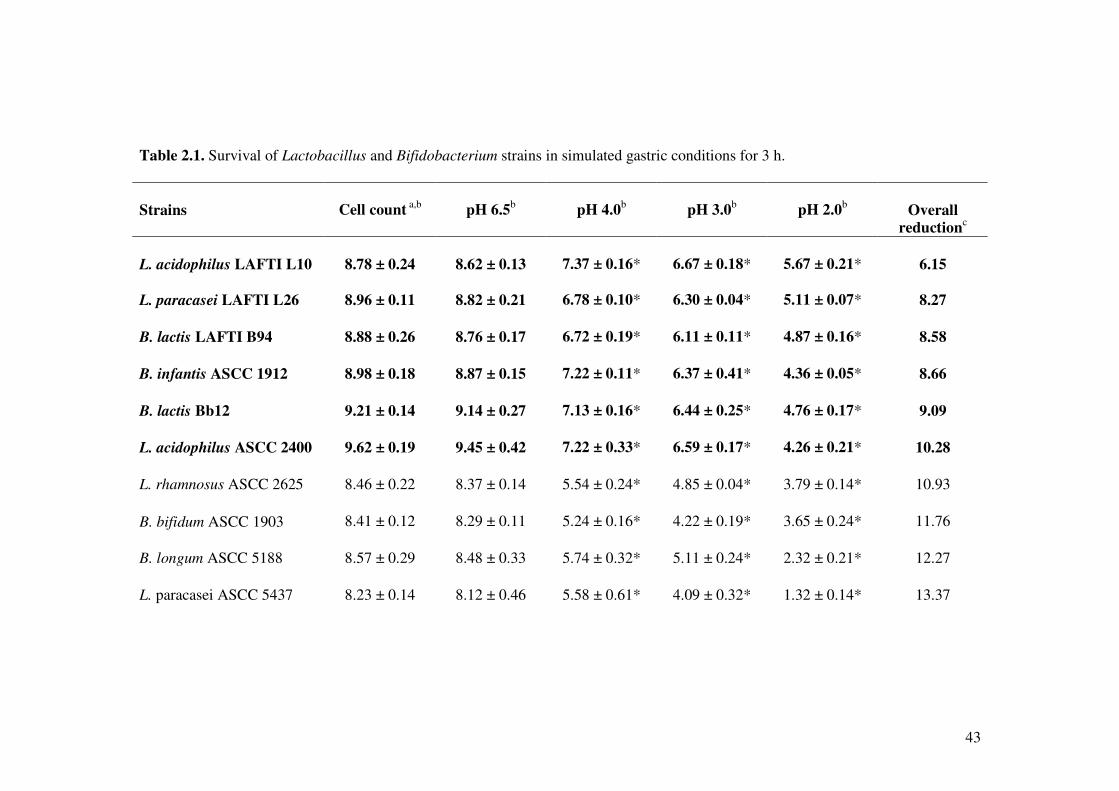
43
Table 2.1. Survival of Lactobacillus and Bifidobacterium strains in simulated gastric conditions for 3 h.
Strains
Cell count a,b
pH 6.5b
pH 4.0b
pH 3.0b
pH 2.0b
Overall
reductionc
L. acidophilus LAFTI L10
8.78 ± 0.24
8.62 ± 0.13
7.37 ± 0.16*
6.67 ± 0.18*
5.67 ± 0.21*
6.15
L. paracasei LAFTI L26
8.96 ± 0.11 8.82 ± 0.21 6.78 ± 0.10* 6.30 ± 0.04* 5.11 ± 0.07* 8.27
B. lactis LAFTI B94 8.88 ± 0.26 8.76 ± 0.17 6.72 ± 0.19* 6.11 ± 0.11* 4.87 ± 0.16* 8.58
B. infantis ASCC 1912
8.98 ± 0.18 8.87 ± 0.15 7.22 ± 0.11* 6.37 ± 0.41* 4.36 ± 0.05* 8.66
B. lactis Bb12
9.21 ± 0.14 9.14 ± 0.27 7.13 ± 0.16* 6.44 ± 0.25* 4.76 ± 0.17* 9.09
L. acidophilus ASCC 2400
9.62 ± 0.19 9.45 ± 0.42 7.22 ± 0.33* 6.59 ± 0.17* 4.26 ± 0.21* 10.28
L. rhamnosus ASCC 2625
8.46 ± 0.22 8.37 ± 0.14 5.54 ± 0.24* 4.85 ± 0.04* 3.79 ± 0.14* 10.93
B. bifidum ASCC 1903
8.41 ± 0.12 8.29 ± 0.11 5.24 ± 0.16* 4.22 ± 0.19* 3.65 ± 0.24* 11.76
B. longum ASCC 5188 8.57 ± 0.29 8.48 ± 0.33 5.74 ± 0.32* 5.11 ± 0.24* 2.32 ± 0.21* 12.27
L. paracasei ASCC 5437
8.23 ± 0.14 8.12 ± 0.46 5.58 ± 0.61* 4.09 ± 0.32* 1.32 ± 0.14* 13.37
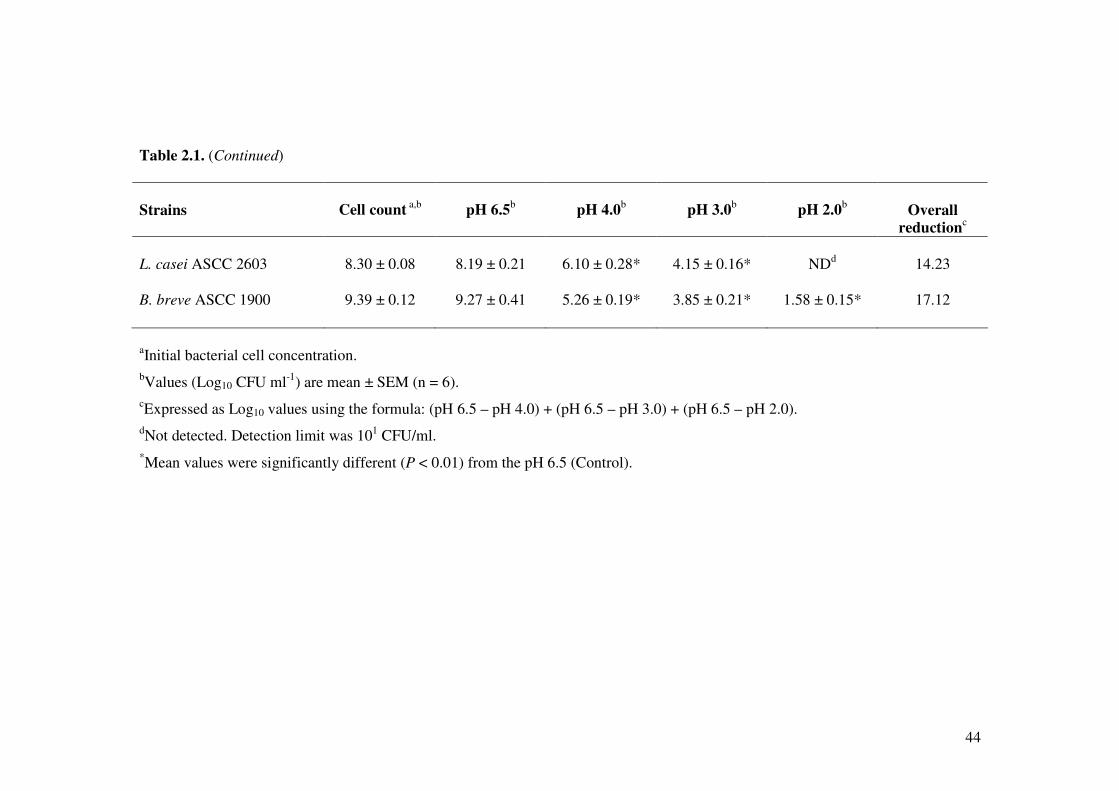
44
Table 2.1. (Continued)
Strains
Cell count a,b
pH 6.5b
pH 4.0b
pH 3.0b
pH 2.0b
Overall
reductionc
L. casei ASCC 2603
8.30 ± 0.08
8.19 ± 0.21
6.10 ± 0.28*
4.15 ± 0.16*
NDd
14.23
B. breve ASCC 1900 9.39 ± 0.12 9.27 ± 0.41 5.26 ± 0.19* 3.85 ± 0.21* 1.58 ± 0.15* 17.12
aInitial bacterial cell concentration.
bValues (Log10 CFU ml-1) are mean ± SEM (n = 6).
cExpressed as Log10 values using the formula: (pH 6.5 – pH 4.0) + (pH 6.5 – pH 3.0) + (pH 6.5 – pH 2.0).
dNot detected. Detection limit was 101 CFU/ml.
*Mean values were significantly different (P < 0.01) from the pH 6.5 (Control).
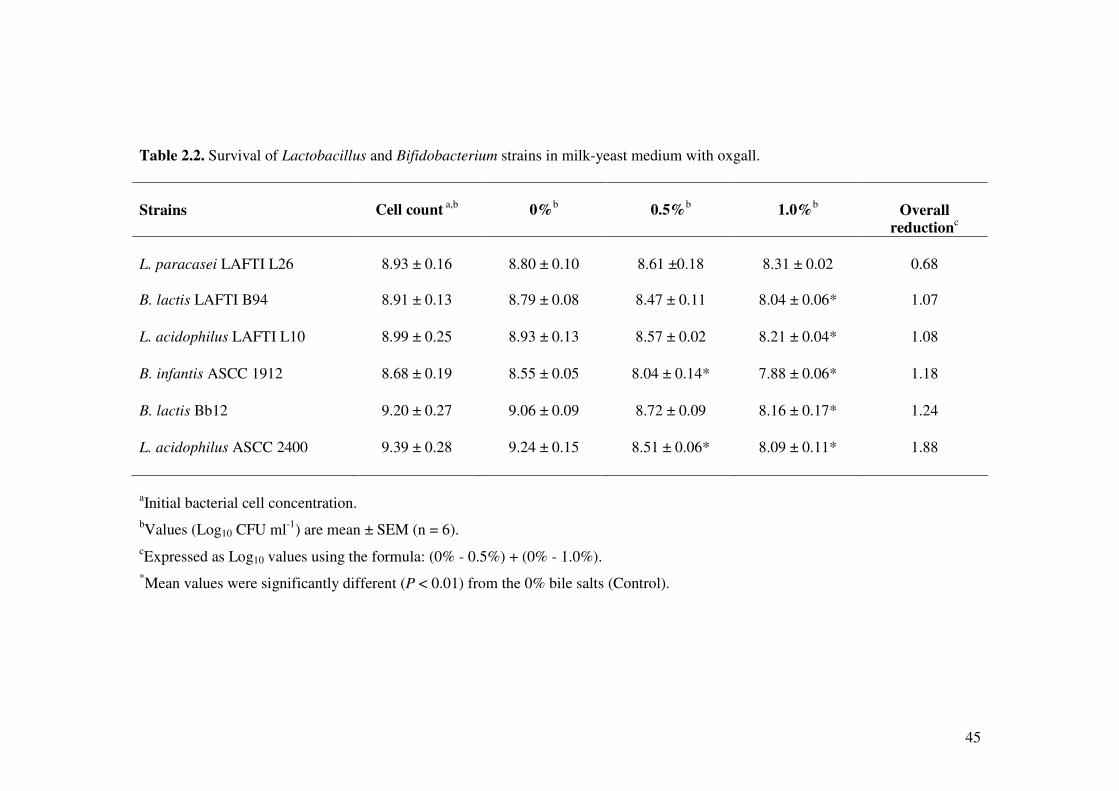
45
Table 2.2. Survival of Lactobacillus and Bifidobacterium strains in milk-yeast medium with oxgall.
Strains
Cell count a,b
0%b
0.5%b
1.0%b
Overall
reductionc
L. paracasei LAFTI L26
8.93 ± 0.16
8.80 ± 0.10
8.61 ±0.18
8.31 ± 0.02
0.68
B. lactis LAFTI B94
8.91 ± 0.13 8.79 ± 0.08 8.47 ± 0.11 8.04 ± 0.06* 1.07
L. acidophilus LAFTI L10
8.99 ± 0.25 8.93 ± 0.13 8.57 ± 0.02 8.21 ± 0.04* 1.08
B. infantis ASCC 1912
8.68 ± 0.19 8.55 ± 0.05 8.04 ± 0.14* 7.88 ± 0.06* 1.18
B. lactis Bb12
9.20 ± 0.27 9.06 ± 0.09 8.72 ± 0.09 8.16 ± 0.17* 1.24
L. acidophilus ASCC 2400
9.39 ± 0.28 9.24 ± 0.15 8.51 ± 0.06* 8.09 ± 0.11* 1.88
aInitial bacterial cell concentration.
bValues (Log10 CFU ml-1) are mean ± SEM (n = 6).
cExpressed as Log10 values using the formula: (0% - 0.5%) + (0% - 1.0%).
*Mean values were significantly different (P < 0.01) from the 0% bile salts (Control).

46
2.4.3. Surface hydrophobicity of Lactobacillus and Bifidobacterium strains
The cell surface hydrophobicity of Lactobacillus and Bifidobacterium strains to n-
hexadecane was investigated as shown in Figure 2.1. The results indicate that B. lactis
Bb12 (25%) showed higher surface hydrophobicity compared to less hydrophobic B.
infantis ASCC 1912 (10%). Specifically, LAFTI strains, such as L. acidophilus L10, L.
paracasei L26 and B. lactis B94 demonstrated similar cell surface hydrophobicity
ranging between 14% – 16%. Furthermore, L. acidophilus ASCC 2400 also exhibited
similar hydrophobic levels by 14%.
2.4.4. Autoaggregation and coaggregation of Lactobacillus and Bifidobacterium
strains
The autoaggregation of bacterial strains was investigated on the basis of their
sedimentation properties after 6 h as shown in Figure 2.2. All Lactobacillus and
Bifidobacterium strains demonstrated the differences in their autoaggregation abilities.
Particularly, L. paracasei LAFTI L26, B. lactis LAFTI B94 and L. acidophilus ASCC
2400 showed higher autoaggregation ability by 33%, whereas L. acidophilus LAFTI L10
and B. infantis ASCC 1912 indicated similar autoaggregation profiles by 30%.
Particularly, B. lactis Bb12 exhibited autoaggregation ability by 29%, which was less in
comparison to other Lactobacillus and Bifidobacterium strains that were screened in this
study.

47
Coaggregation of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 and
individually with other potential probiotic strains such as, B. lactis LAFTI B94, B. lactis
Bb12, B. infantis ASCC 1912 and L. acidophilus ASCC 2400 was examined (Fig. 2.3).
The coaggregation ability was expressed as a percentage reduction in the absorbance of
mixed bacterial suspension compared to individual microbial suspension. Lactobacillus
acidophilus LAFTI L10 and L. paracasei LAFTI L26 indicated higher coaggregation
ability by 75% compared to their individual coaggregation ability with other potential
probiotic strains. The coaggregation ability of L. acidophilus LAFTI L10 and B. lactis
LAFTI B94 was increased by 54%, which was significantly different (P < 0.01)
compared to L. paracasei LAFTI L26 coaggregation with B. lactis LAFTI B94 (24%).
Simultaneously, L. acidophilus LAFTI L10 demonstrated increased (P < 0.01)
coaggregation by 53% with B. lactis Bb12 compared to L. paracasei LAFTI L26 with B.
lactis Bb12 (18%). The coaggregation of L. acidophilus LAFTI L10 or L. paracasei
LAFTI L26 with B. infantis ASCC 1912 showed no differences (P > 0.05) in their
aggregation capabilities. However, coaggregation of L. acidophilus LAFTI L10 or L.
paracasei LAFTI L26 with L. acidophilus ASCC 2400 demonstrated significant
differences (P < 0.01).
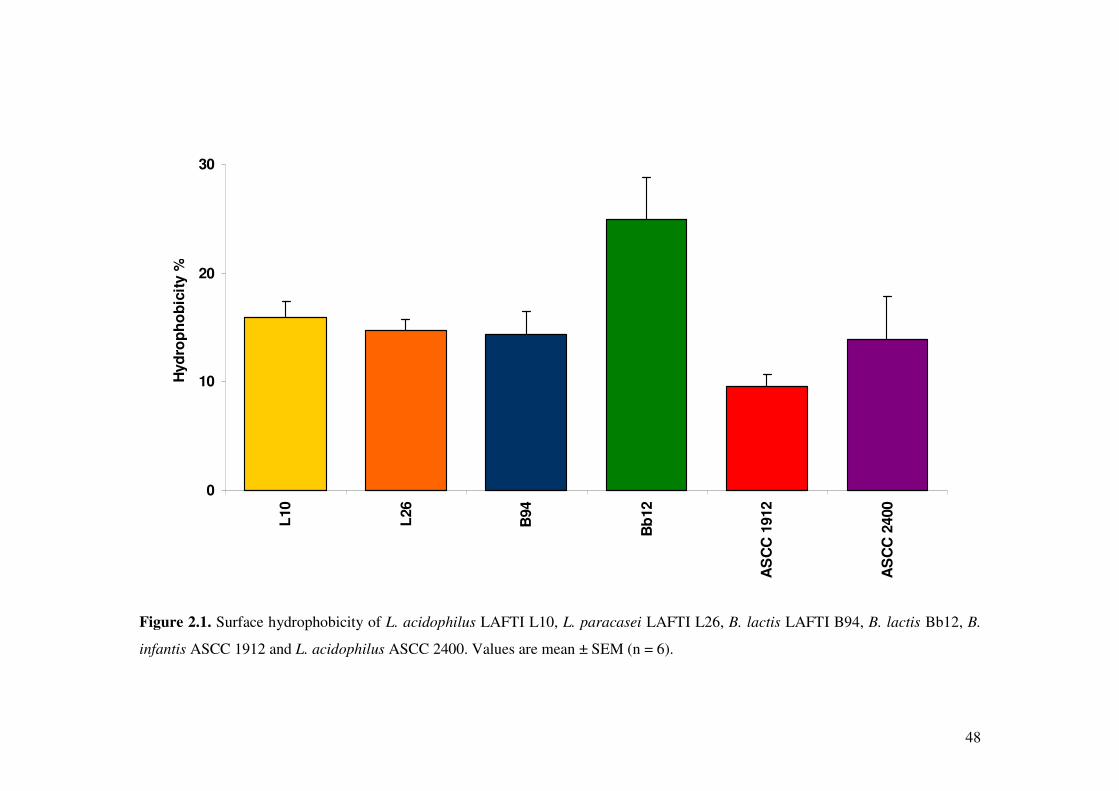
48
0
10
20
30
L10
L26
B94
Bb
12
AS
CC
1912
AS
CC
2400
Hyd
rop
ho
bic
ity %
Figure 2.1. Surface hydrophobicity of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B.
infantis ASCC 1912 and L. acidophilus ASCC 2400. Values are mean ± SEM (n = 6).
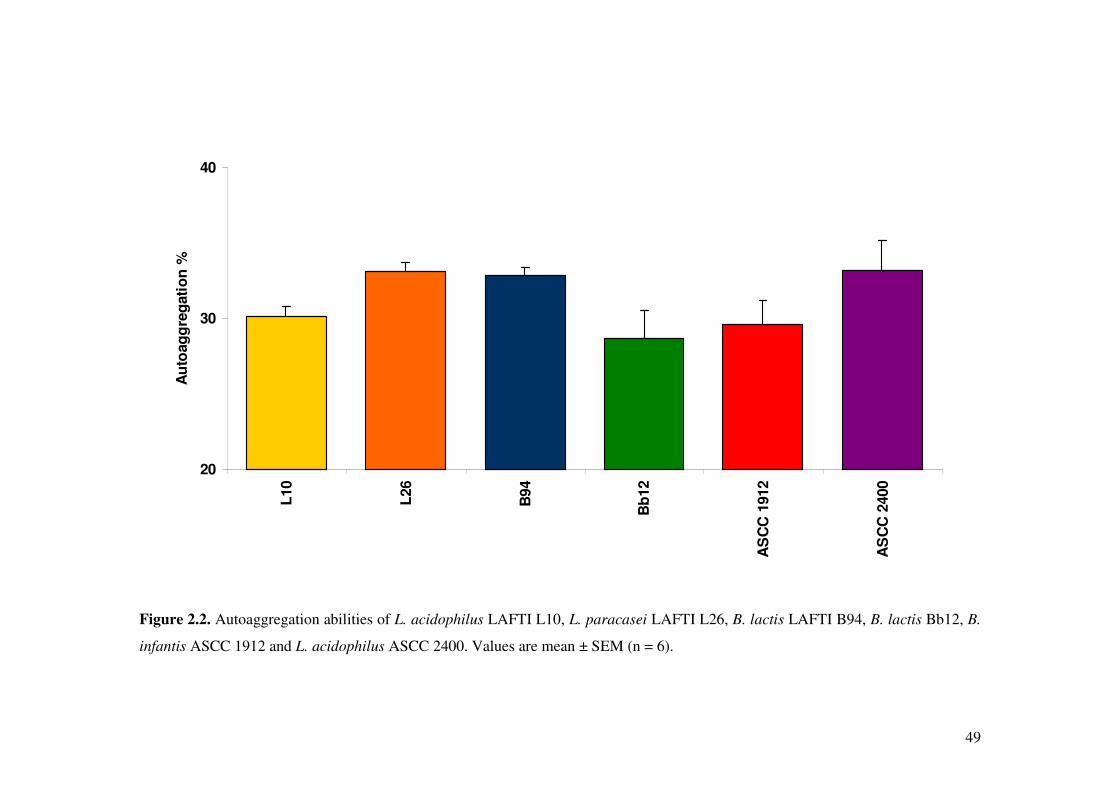
49
20
30
40
L10
L26
B94
Bb
12
AS
CC
1912
AS
CC
2400
Au
toag
gre
gati
on
%
Figure 2.2. Autoaggregation abilities of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B.
infantis ASCC 1912 and L. acidophilus ASCC 2400. Values are mean ± SEM (n = 6).
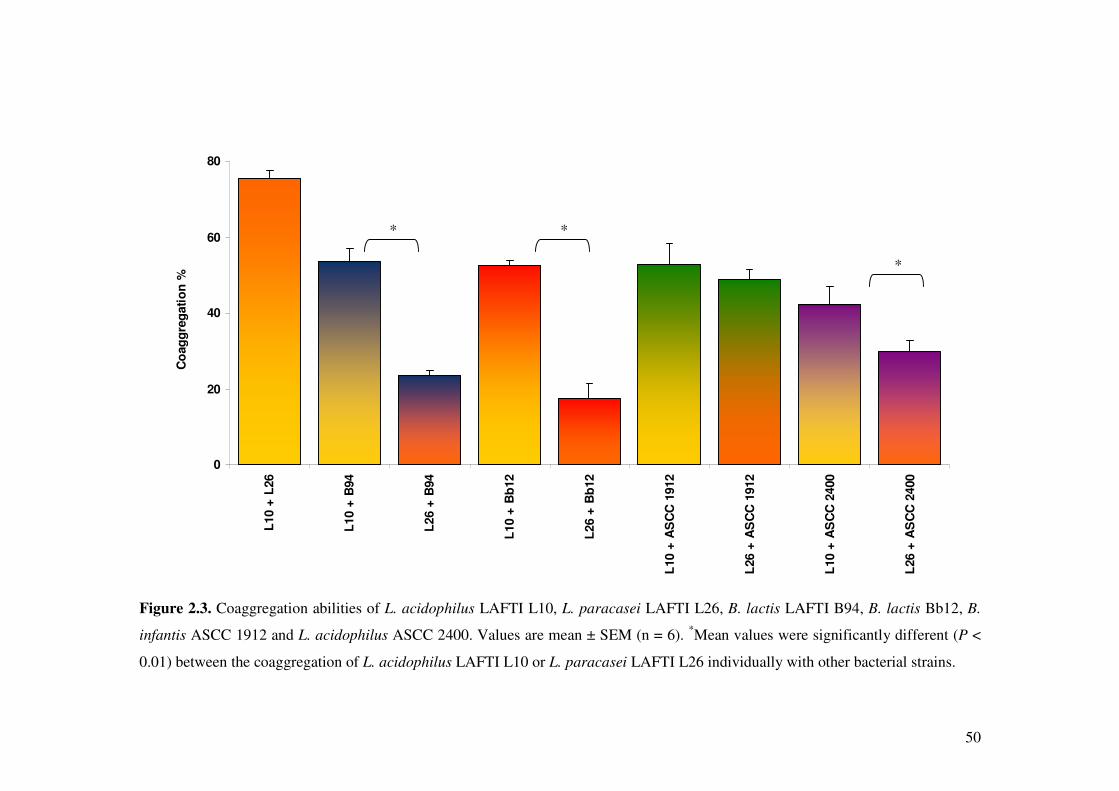
50
0
20
40
60
80
L10 +
L26
L10 +
B94
L26 +
B94
L10 +
Bb
12
L26 +
Bb
12
L10 +
AS
CC
1912
L26 +
AS
CC
1912
L10 +
AS
CC
2400
L26 +
AS
CC
2400
Co
ag
gre
gati
on
%
Figure 2.3. Coaggregation abilities of L. acidophilus LAFTI L10, L. paracasei LAFTI L26, B. lactis LAFTI B94, B. lactis Bb12, B.
infantis ASCC 1912 and L. acidophilus ASCC 2400. Values are mean ± SEM (n = 6). *Mean values were significantly different (P <
0.01) between the coaggregation of L. acidophilus LAFTI L10 or L. paracasei LAFTI L26 individually with other bacterial strains.
* *
*

51
2.5. DISCUSSION
In order to exert beneficial effects, probiotic bacteria must be tolerant to harsh gastric
conditions and high bile salt concentrations in the GI tract. In humans, pH of normal
gastric juice can be below 3.0, which can significantly prevent all bacterial growth.
Although the pH of stomach could be as low as 1.0, several studies reported the
preferred pH as 3.0 for in vitro assays (Garriga et al., 1998; Suskovic et al., 1997).
Usman and Hosono, (1999) also suggested the survival of probiotic bacteria at pH
3.0 for 2 h as an optimal screening condition for characterising probiotic strains
towards acid tolerance.
In the present study, overall reduction in cell viability of bacterial strains at various
pH conditions was considered as selection criteria, which lead to the identification of
6 Lactobacillus and Bifidobacterium strains. Particularly, these 6 Lactobacillus and
Bifidobacterium strains showed better survival at pH 2.0 with cell viability higher
than 104 CFU/ml (Table 2.1). Lactobacillus acidophilus LAFTI L10 remained highly
tolerant to various pH conditions with less overall decrease in cell viability. In pH
2.0, L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 survived by more than
105 CFU/ml, whereas other bacterial strains that were selected remained at 104
CFU/ml. Strain differences at various pH conditions were observed, which were
consistent to the earlier studies that reported similar strain-specific variations to
simulated gastric conditions (Truelstrup Hansen et al., 2002; Mishra and Prasad,
2005).
Several studies reported the effects of various bile salt concentrations on the growth
of probiotic bacteria (Clark and Martin, 1994; Lankaputhra and Shah, 1995; Prasad

52
et al., 1998; Truelstrup Hansen et al., 2002). There were no reports suggesting the
precise concentration of bile salts that can be used as selection criteria towards
probiotic characterisation. The bile concentration varies accordingly in the human GI
tract; however it was believed to be 0.3% w/v (Sjovall, 1959, Gilliland et al., 1984).
In the current study, bile salt concentrations of 0.5% and 1.0% were used for
screening bacterial strains towards bile tolerance.
Lactobacillus and Bifidobacterium strains used in this study survived by more than
108 CFU/ml in 1.0% bile salts with exception to B. infantis ASCC 1912, which
survived at 107 CFU/ml. Lactobacillus paracasei LAFTI L26 showed an overall
higher tolerance to various bile salt concentrations, whereas L acidophilus LAFTI
L10 and B. lactis LAFTI B94 showed similar reductions in the overall cell
survivability. In contrast, L. acidophilus LAFTI L10 demonstrated higher tolerance
to 1.0% bile salts compared to B. lactis LAFTI B94. Interestingly, LAFTI strains
such as L. acidophilus L10, L. paracasei L26 and B. lactis B94 demonstrated high
tolerance to simulated gastric conditions and bile salt concentrations. These
characteristics demonstrate that LAFTI strains are likely to survive in varying GI
tract conditions, where they could exert positive health effects on the host.
Adhesion of bacterial strains to GI tract is necessary to colonise and induce
beneficial effects on the host. Several direct and indirect in vitro methods were used
to study the adherence ability of probiotic bacteria, but still there was no consensus
about defining an optimal method that could predict the in vivo microbial adhesion.
In the current study, microbial adhesion to n-hexadecane was used to predict the
adherence ability of Lactobacillus and Bifidobacterium strains. Bifidobacterium

53
lactis Bb12 showed higher hydrophobicity when compared with other bacterial
strains. The LAFTI strains, L. acidophilus L10, L. paracasei L26 and B. lactis B94
demonstrated similar hydrophobic abilities ranging between 14% – 16%.
Particularly, B. infantis ASCC 1912 showed less hydrophobicity compared to other
bacterial strains screened in the current study. The differences in hydrophobic ability
of bacterial strains observed in this study could be due to variations in the chemical
composition of cell surface components (Pelletier et al., 1997), particularly
lipoteichoic acid-protein complex (Sherman and Savage, 1986) or presence of cell
surface hydrophobic protein moiety (Ronner et al., 1990). Furthermore, Gomez
Zavaglia et al. (2002) demonstrated the influence of bile salts, which decreased the
autoaggregation ability of Bifidobacterium strains.
Earlier, several studies reported the composition, structure and forces of interaction
that are related to bacterial adherence with intestinal epithelial cells (Greene and
Klaenhammer, 1994; Pelletier et al., 1997; Perez et al., 1998). Moreover, the ability
of aggregation is related to cell surface characteristics of bacterial strains
(Vandevoorde et al., 1992; Del Re et al., 2000). Microbial aggregation is considered
as a desirable characteristic of probiotic bacteria (Drago et al., 1997; Mastromarino
et al., 2002; Castagliuolo et al., 2005). In the present study, autoaggregation ability of
bacterial strains was examined, which could be considered as an indication of
adherence characteristics. All the strains used in this study showed differences in
their autoaggregation abilities. Interestingly, LAFTI strains, L. acidophilus L10, L.
paracasei L26 and B. lactis B94 showed similarities in their autoaggregation and
hydrophobic abilities. These results were consistent with the earlier reports, which
suggested similarities in bacterial autoaggregation and hydrophobicity (Gomez

54
Zavaglia et al., 2002; Aslim et al., 2007). However, B. lactis Bb12 demonstrated
higher hydrophobicity and lower autoaggregation ability compared to other bacterial
strains used in the current study. Similar differences in autoaggregation and
hydrophobicity of bacterial strains was also reported earlier (Del Re et al., 2000).
The coaggregation ability of bacterial strains isolated from human dental plaque was
the first study to suggest the process of adhesion between genetically distinct
bacterial strains. However, coaggregation is now being investigated among the
bacteria that were isolated from human oral cavity, mammalian GI tracts and human
urogenital tract (Rickard et al., 2003). Drago et al. (1997) demonstrated the
coaggregation ability of Lactobacillus strains with enterotoxigenic E. coli,
Salmonella enteritidis and Vibrio cholerae. Earlier studies suggested the
phenomenon of coaggregation between bacterial strains isolated from dental plaque
of healthy individuals (Kolenbrander, 2000) and beneficial effects of cell aggregation
in promoting the colonisation of lactobacilli in GI and vaginal tracts (Vandevoorde et
al., 1992; Kmet and Lucchini, 1997; Cesena et al., 2001; Jankovic et al., 2003).
In the present study, coaggregation of L. acidophilus LAFTI L10 with B. lactis
LAFTI B94, B. lactis Bb12 and L. acidophilus ASCC 2400 showed significant
differences compared to L. paracasei LAFTI L26. Vizoso Pinto et al. (2007)
demonstrated higher degree of coaggregation ability of L. johnsonii compared to L.
casei Shirota and L. rhamnosus GG with human pathogens such as E. coli, L.
monocytogenes, Staphylococcus aureus and Salmonella Typhimurium. In the current
study, coaggregation of L. acidophilus LAFTI L10 with L. paracasei LAFTI L26
demonstrated higher coaggregating ability compared to their individual

55
coaggregation abilities with other bacterial strains. Especially, coaggregation abilities
of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 with pathogenic bacteria
need to be investigated, which may constitute as a protective mechanism against
infection (Reid et al., 1988; Schachtsiek et al., 2004).

56
2.6. CONCLUSIONS
Lactobacillus acidophilus LAFTI L10 and L. paracasei LAFTI L26 were selected
for subsequent in vivo studies based on their overall tolerance to various pH
conditions, especially due to their higher survival at pH 2.0. These strains also
showed better tolerance to bile conditions particularly to 1.0% bile salts. All bacterial
strains demonstrated the correlation of adherence abilities such as hydrophobicity
and autoaggregation with exception to B. lactis Bb12. Lactobacillus acidophilus
LAFTI L10 showed higher coaggregation abilities with L. paracasei LAFTI L26 and
also with other probiotic bacterial strains.

57
CHAPTER 3
Effect of L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 on gut and
systemic immunity and bacterial translocation in mice

58
3.1. ABSTRACT
The immune enhancing properties of Lactobacillus acidophilus LAFTI L10 and
Lactobacillus paracasei LAFTI L26 in mice were investigated. Each mouse
(BALB/c) was orally fed with cultures of either L. acidophilus or L. paracasei at 108
CFU/50µl per day for 14-days. Immunofluorescence assay of the small intestine in
mice demonstrated the capability of both Lactobacillus strains in enhancing
immunoglobulin (Ig)-A, interleukin (IL)-10 and interferon (IFN)-� producing cells.
In systemic immune response, proliferative responses of splenocytes to concanavalin
A (ConA; T-cell mitogen) and lipopolysaccharide (LPS; B-cell mitogen) were
significantly higher in mice fed with L. acidophilus. ConA-induced splenocyte
proliferative responses were increased significantly in mice fed with L. paracasei,
however there was no significant difference of splenocyte proliferative responses to
LPS compared to control mice. In mice fed with either L. acidophilus or L.
paracasei, IL-10 and IFN-� levels in blood serum and splenocytes stimulated with
ConA were increased compared to the control group. Both Lactobacillus strains
showed similar patterns in activating the phagocytic activity of peritoneal
macrophages. Lactobacillus acidophilus or L. paracasei-fed mice modulated the
translocation of Lactobacillus spp. and total anaerobes to Peyer’s patches and
mesenteric lymph nodes when compared to control mice. Furthermore, there was no
indication of disruption of intestinal mucosal integrity and thus no bacterial
translocation to spleen, liver or blood in mice fed with L. acidophilus or L.
paracasei. The results of this study indicate that L. acidophilus and L. paracasei are
potential enhancers of gut and systemic immunity and are non-pathogenic, as
suggested by their bacterial translocation profiles in healthy mice.

59
3.2. INTRODUCTION
The mucous covering of gastrointestinal (GI) tract has various important functions
such as providing a well balanced habitat for beneficial intestinal microflora and
protecting the host from pathogenic bacterial infection (Corfield et al., 2001). Lactic
acid bacteria (LAB) are known to stimulate immune responses of gut associated
lymphoid tissues (GALT) through lymphoid cells of GI tract (Herias et al., 1999;
Perdigon et al., 1994; Perdigon et al., 1999). Evidence is accumulating to
demonstrate that certain LAB promote health benefits by stimulating immune
responses through phagocytosis (Perdigon et al., 1986), enhanced release of
cytokines (Marin et al., 1998; Miettinen et al., 1998; Kato et al., 1999; Tejada-Simon
et al., 1999) and increased mucosal and secretory antibody levels in response to
Escherichia coli 0157:H7 infection (Shu and Gill, 2002) and Shigella sonnei (Nader
de Macias et al., 1992). Several reports have shown that LAB influences the balance
of Th1/Th2 cytokines (Borruel et al., 2003; Morita et al., 2002; Wallace et al., 2003).
Stimulation of cells to produce cytokines and elimination of toxic compounds are
related to the surface adherence properties of LAB (Gomez Zavaglia et al., 2002).
The effect of probiotic bacteria on immune responses is often evaluated through in
vitro and ex vivo measurements of cytokine or immunoglobulin production, T- or B-
cell proliferation. Gill et al. (2000) demonstrated that orally administered LAB
enhances proliferative responses of splenocytes to concanavalin A (ConA; T-cell
mitogen) and lipopolysaccharide (LPS; B-cell mitogen). However, Kirjavainen et al.
(1999) reported that oral administration of LAB inhibits lymphocyte proliferation.
Several scientific studies reported the influence of LAB in inducing cytokines such
as interferon (IFN)-�, tumor necrosis factor (TNF)-�, interleukin (IL)-6, IL-10, IL-12

60
and IL-18 in human blood mononuclear cells (Miettinen et al., 1996, Solis-Pereyra et
al., 1997; Miettinen et al., 1998). Furthermore, LAB influenced the enhancement of
IFN-� levels in ConA stimulated human peripheral blood lymphocytes (De Simone et
al., 1986).
Macrophages are regulatory cells that play an important role in inflammation and
host defence-related functions such as phagocytosis, production of cytokines and
mediators, antigen presentation, antimicrobial and tumoricidal activity. Phagocytosis
is a critical function of mononuclear phagocytes and neutrophils. The ability of
lactobacilli to induce cytokine production by macrophages (Marin et al., 1998;
Hessle et al., 1999) and stimulation of macrophages through NF-kappa B and STAT
transcription factor has been demonstrated (Mestecky et al., 1999). Human
volunteers fed with either L. acidophilus La1 or B. bifidum Bb12 enhanced the
phagocytosis of peripheral blood leukocytes, which persisted for 6 weeks after oral
ingestion of these strains (Schiffrin et al., 1997).
Intestinal bacterial translocation is the passage of viable indigenous bacteria from
gastrointestinal lumen to lymphatic organs (Peyer’s patches and mesenteric lymph
nodes), spleen, liver, peritoneal cavity and blood stream (Berg, 1992). Translocation
of microorganisms includes E. coli and certain members of Enterobacteriaceae and
rarely anaerobes (Wells et al., 1988). Bacterial translocation in healthy mice occurs
continuously at a low rate, which is a highly regulated biological process (Ma et al.,
1990). The disruption of intestinal barrier or incompetent host immune system that is
unable to confine an infection could lead the pathogenic bacteria to bloodstream and
cause septicemia (Berg, 1999). However, translocation of LAB from gut to GALT is

61
considered as a normal and beneficial physiological process associated with the
stimulation of immune responses (Bengmark and Jeppsson, 1995). Furthermore,
scientific studies also reported the potential role of LAB in protecting gut barrier
functions in animal models (Luyer et al., 2005; Daniel et al., 2006; Zareie et al.,
2006).
Earlier reports showed the influence of probiotic bacteria on host immune functions
in a strain dependent manner and therefore it is inappropriate to generalise the
probiotic beneficial effects that are specific to genus or species (Perdigon et al.,
2001; Schiffrin and Blum, 2002). As shown in Chapter 2, Lactobacillus and
Bifidobacterium strains were screened to select bacterial strains that are tolerant to
simulated gastric juices and bile salts. After subsequent screening of the bacterial
strains, L. acidophilus LAFTI L10 and L. paracasei LAFTI L26 were identified as
potential probiotic strains based on their tolerance to low pH and high bile salts.
Lactobacillus acidophilus L10 and L. paracasei L26 also demonstrated better
adhesive abilities such as autoaggregation and coaggregation, which are known to be
important in bacterial adhesion to intestinal mucosa in order to influence host
immune functions. The commercial probiotic strains may lose their potential
probiotic properties and also change their physiological processes due to rigorous
industrial processing conditions and repeated subculturing. Earlier in vitro studies
reported that L. acidophilus L10 and L. paracasei L26 inhibited the growth of
Listeria monocytogenes and E. coli 0111 (Pidcock et al., 2002). Further studies
conducted using L. acidophilus L10 in mice showed the ability to inhibit the growth
of L. monocytogenes in the GI tract (Mahoney and Henriksson, 2003). Elahi et al.
(2005) demonstrated that mice fed with L. acidophilus L10 enhanced the clearance of

62
Candida albicans from oral cavities, which are correlated with increased levels of
IFN-� and nitric oxide in saliva.
There were no scientific reports that suggested the ability of L. acidophilus L10 or L.
paracasei L26 in stimulating in vivo gut immune responses. However, little was
known about the capability of L. acidophilus L10 or L. paracasei L26 in stimulating
distant immune responses at systemic levels (Cayzer et al., 2001; Clancy et al., 2006)
and furthermore no scientific studies reported the influence of these strains on
bacterial translocation. Therefore, this study investigated the effects of L. acidophilus
L10 and L. paracasei L26 on various indices of immune functions in gut and
systemic immunity and their influence on bacterial translocation in healthy mice.

63
3.3. MATERIALS AND METHODS
3.3.1. Animals
Male 6 - 8 wks old BALB/c mice were obtained from Biological Resources Centre,
The University of New South Wales, Sydney, Australia. Control and experimental
groups consisted of six mice each, which were caged separately at 22 ± 1°C and
housed under 12 h light-dark photoperiod. All mice were fed with standard mouse
chow (Gordon's Specialty Stock Feeds, Sydney, Australia) and provided ad libitum
access to water. The University of Western Sydney (Sydney, Australia) Animal Care
and Ethics Committee approved all protocols for the animal experiment.
3.3.2. Microorganisms
Lactobacillus acidophilus LAFTI L10 and L. paracasei LAFTI L26 strains were
obtained from DSM culture collection (DSM Food Specialties Ltd., Sydney,
Australia). Lactobacillus acidophilus and L. paracasei were grown anaerobically in
gas jars using GasPak System (Oxoid, Adelaide, Australia) for 24 h at 37°C in de
Man, Rogosa and Sharpe (MRS) broth (Oxoid, Adelaide, Australia). The bacterial
cells were harvested at 4000 x g for 15 min at 4°C and washed twice with sterile 0.01
M phosphate buffered saline (PBS) solution.
3.3.3. Feeding procedure
After acclimatisation for 1 week, mice were fed daily by oral gavage with either L.
acidophilus or L. paracasei of 108 colony-forming units (CFU) in 50µl of 10% (w/v)
skimmed milk powder (SMP) for 14-days. Control group mice received 50µl of SMP
without lactic acid bacteria. The viability of bacteria was determined by spread plate

64
count of bacterial serial dilutions on MRS agar (Oxoid, Adelaide, Australia), which
were incubated anaerobically in gas jars using GasPak System at 37°C for 48 h.
3.3.4. Histological samples
At the end of 14-day feeding trail, mice were euthanized by carbon dioxide
inhalation and small intestines were removed, dissected into segments and stored in
95% ethanol (v/v). The tissues were then embedded in paraffin blocks and the
sections were processed for immunofluorescence assays.
3.3.5. Immunofluorescent detection of IgA cells
The number of immunoglobulin (Ig)-A secreting cells was determined in histological
sections of the mice small intestine by direct immunofluorescence assay.
Histological sections (4-µm) were deparaffinised by immersion in xylene and
rehydrated in ethanol, which were incubated with 1% blocking solution of bovine
serum albumin (Sigma-Aldrich, Sydney, Australia) for 30 min at room temperature.
The sections were then incubated with 1/100 dilution of anti-IgA mono-specific
antibody (�–chain specific) conjugated with fluorescein isothiocyanate (FITC)
(Sigma-Aldrich, Sydney, Australia) in 0.01 M PBS solution for 30 min at 37°C. The
sections were washed three times with 0.01 M PBS solution and the immune cells
were observed by using a fluorescence light microscope. Results were expressed as
number of positive fluorescent cells in 10 fields of vision at 1000x magnification.

65
3.3.6. Detection of cytokine producing cells in small intestine
Interleukin–10 and IFN-� producing cells were detected in histological sections of
the mice small intestine by indirect immunofluorescence assay. After
deparaffinization and rehydration in ethanol, histological sections (4-µm) were
incubated with 1/100 dilution of rabbit anti-mouse IL-10 and IFN-� polyclonal
antibodies (Peprotech Inc., Rocky Hill, USA) for 60 min at room temperature. The
incubation was followed by washing twice with 0.01 M PBS solution. The sections
were then incubated with 2/100 dilution of goat anti-rabbit antibody conjugated with
FITC (Jackson ImmunoResearch Laboratories Inc., West Grove, USA) for 45 min at
room temperature. The number of fluorescent cells was counted using a fluorescence
light microscope and expressed as positive fluorescent cells in 10 fields of vision at
1000x magnification.
3.3.7. Splenocytes
Spleens from mice were removed aseptically and placed individually in complete
RPMI-1640 medium containing 10% fetal calf serum (FCS), 100 U/ml penicillin and
100 µg/ml streptomycin. Splenocytes were isolated by chopping the spleen tissue
into small pieces with sterile scissors and forcing the spleen cell suspension through
a 5 ml syringe. The resulting spleen cell suspension was then transferred into a tube
containing 5 ml of complete RPMI-1640 medium and centrifuged at 300 x g for 10
min. To lyse erythrocytes, cells were resuspended in ACK lysis buffer (Tris-NH4Cl)
and incubated at room temperature for 5 min followed by washing the cells twice in
complete RPMI-1640 medium. The splenic lymphocyte suspensions were adjusted to
a final concentration of 2 x 106 cells/ml in complete RPMI-1640 medium. Trypan
blue exclusion method was used to determine the cell viability.

66
3.3.8. Lymphocyte proliferation assay
A 50 µl of complete RPMI-1640 medium with 2 x 106 cells ml-1 were added in
triplicates to the wells of a 96-well tissue culture plate and cultured in the presence or
absence of T- and B-cell mitogens. A 50 µl/well of pre-optimised concentrations of
mitogens, concanavalin A (ConA, 2.5 µg/ml; Sigma, Sydney, Australia) and
lipopolysaccharide (LPS, 5 µg/ml, derived from E. coli; Sigma, Sydney, Australia)
were added to the wells. Control wells received 50 µl of complete RPMI-1640
medium. The cells were cultured for 72 h at 37°C in a humidified incubator with CO2
and air (5: 95, v/v). The cell proliferation over the final 4 h of culture was determined
by using CellTiter 96 AQueous One Solution Cell Proliferation Assay (Promega,
Madison, USA). The absorbance of each well was read at 492nm using a microtitre
plate reader (Multiskan Titertek, Huntsville, USA).
3.3.9. Estimation of cytokines in blood and splenocytes
Blood was collected upon euthanizing the mice after 14-day feeding trial and left to
coagulate overnight at 4°C. Serum was then processed from blood by centrifugation
at 2000 x g for 10 min and stored at -20°C until cytokines were assayed. Serum was
used to estimate the presence of IL-10 and IFN-� using mouse IL-10 and mouse IFN-
� sandwich ELISA kits (Chemicon International Inc., Temecula, USA).
Spleen cell suspensions (2 x 106 cells ml-1) were added in triplicates to the wells of a
96-well tissue culture plate and cultured in the presence of ConA (2.5 µg/ml) for 48 h
at 37°C in a humidified incubator with CO2 and air (5: 95, v/v). The supernatant
fractions were harvested from cell suspensions and stored at -80°C until cytokine

67
analysis. Estimation of IL-10 and IFN-� in the supernatants was determined using
mouse IL-10 (Chemicon International Inc., Temecula, USA) and mouse IFN-�
(Assay Designs Inc., Ann Arbor, USA) sandwich ELISA kits.
3.3.10. Phagocytosis
Peritoneal macrophages were isolated from the peritoneal cavity of each mouse by
flushing with 5 ml of RPMI-1640 medium. Peritoneal macrophages were washed
twice and adjusted to a final concentration of 106 cells/ml in RPMI-1640 medium.
Heat killed (100°C, 15 min) Candida albicans ATCC 10231 suspension (106 cells
ml-1) of 0.2 ml opsonised with 10% mouse autologous serum for 15 min at 37°C was
added to 0.2 ml of each macrophage suspension at a concentration of 106 cells/ml
and incubated for 30 min at 37°C. The results were expressed as percentage of
phagocytosis that was determined by counting over 100 macrophages using a Nikon
optical microscope.
3.3.11. Bacterial translocation assay
Bacterial translocation to Peyer’s patches, mesenteric lymph nodes (MLN), spleen,
liver and blood was determined in control and experimental group mice. The Peyer’s
patches, MLN, spleen and a sample of liver tissue were aseptically removed from
each animal and washed twice with 0.01 M PBS solution, which were transferred to
pre-weighed tubes containing 0.01 M PBS. The organs were weighed, homogenised
and serially diluted in 0.01 M PBS. Blood collected through cardiac puncture was
also serially diluted in 0.01 M PBS. Tissue homogenates and blood suspensions from
each mouse were plated separately in triplicates on MRS agar for detection of

68
Lactobacillus spp., Wilkins-Chalgren (WC) Anaerobe agar (Oxoid, Adelaide,
Australia) containing 5% horse blood (Oxoid, Adelaide, Australia) for detection of
total anaerobes and MacConkey agar (Oxoid, Adelaide, Australia) for detection of
enteric bacteria. Plates were incubated under anaerobic (MRS and WC agar plates)
and aerobic (MacConkey agar plates) conditions at 37°C for 48 h. Colonies were
enumerated on appropriate agar plates and expressed as either colony forming units
per g (CFU g-1) or per ml (CFU ml-1).
3.3.12. Statistical analysis
The results were expressed as mean ± standard error of mean (SEM). The data were
analysed using the one-way analysis of variance procedure of SPSS (Version 12.0.1).
Significant differences between groups were identified by Duncan’s multiple range
test (SPSS, Version 12.0.1).

69
3.4. RESULTS
3.4.1. Determination of IgA producing cells
The effect of orally administered LAB on IgA producing cells in lamina propria of
the small intestine was determined by direct immunofluorescence assay (Fig. 3.1A,
B). In comparison to control mice, mice orally fed with L. acidophilus or L.
paracasei significantly enhanced (P < 0.001) the IgA producing cells in the small
intestine. Lactobacillus acidophilus-fed mice showed an increase of 68% in IgA cells
compared to control mice, whereas L. paracasei-fed mice demonstrated 20%
increase in IgA cells. The differences between L. acidophilus and L. paracasei
experimental groups were significant (P < 0.001) in enhancing the number of IgA
cells.
3.4.2. Effect of LAB on cytokines in gut
The effect of oral administration of LAB in enhancing cytokine producing cells in
lamina propria of the small intestine was determined by indirect immunofluorescence
assay (Fig. 3.2A, B). Significant increase (P < 0.001) of IL-10 and IFN-� producing
cells were detected in the small intestine of mice fed with L. acidophilus or L.
paracasei when compared to the control mice. Specifically, L. acidophilus-fed mice
showed an increase of 63% of IL-10 producing cells and more than 200% of IFN-�
producing cells compared to control group. Simultaneously, L. paracasei
demonstrated an increase of 18% of IL-10 and 95% of IFN-� producing cells,
however the increase was lesser compared to mice fed with L. acidophilus.

70
3.4.3. Lymphocyte proliferative responses
The proliferative responses of spleen cells stimulated with ConA and LPS were
estimated to determine the effect of orally administered LAB on T- and B-cell
functions in each experimental group (Table 3.1). The proliferative responses of
splenocytes stimulated with ConA were significantly higher in mice fed with L.
acidophilus (P < 0.01) or L. paracasei (P < 0.05) compared to control mice. In
comparison to control mice, LPS induced proliferative responses were increased (P <
0.05) in mice fed with L. acidophilus. The splenocyte proliferative responses induced
by LPS were higher in mice fed with L. paracasei but were not significantly different
(P > 0.05) compared to control mice. In contrast, L. acidophilus-fed mice
demonstrated higher (P < 0.01) proliferative responses when stimulated with ConA
and LPS compared to mice fed with L. paracasei.
3.4.4. Effect of LAB on cytokines in blood serum and splenocytes
The effects of L. acidophilus and L. paracasei in systemic immunity were
determined by estimating the serum cytokines. Lactobacillus acidophilus or L.
paracasei demonstrated a significant increase (P < 0.001) of IL-10 and IFN-� levels
in serum compared to control mice (Fig. 3.3A, B). The increase of IL-10 and IFN-�
levels detected in serum of mice fed with L. acidophilus were higher (P < 0.001)
compared to mice fed with L. paracasei.
The effects of ConA induced cytokine production in spleen cells from mice fed with
L. acidophilus or L. paracasei were estimated (Fig. 3.4A, B). In mice fed with L.
acidophilus, production of IL-10 (P < 0.01) and IFN-� (P < 0.05) by splenocytes
were enhanced when compared to control mice. Splenocytes from mice fed with L.

71
paracasei also demonstrated significantly higher levels of IL-10 and IFN-� (P <
0.05) compared to control mice. The differences between mice fed with L.
acidophilus and L. paracasei in enhancing IL-10 production was significant (P <
0.01). However, there were no differences (P > 0.05) between L. acidophilus and L.
paracasei in inducing IFN-� levels.
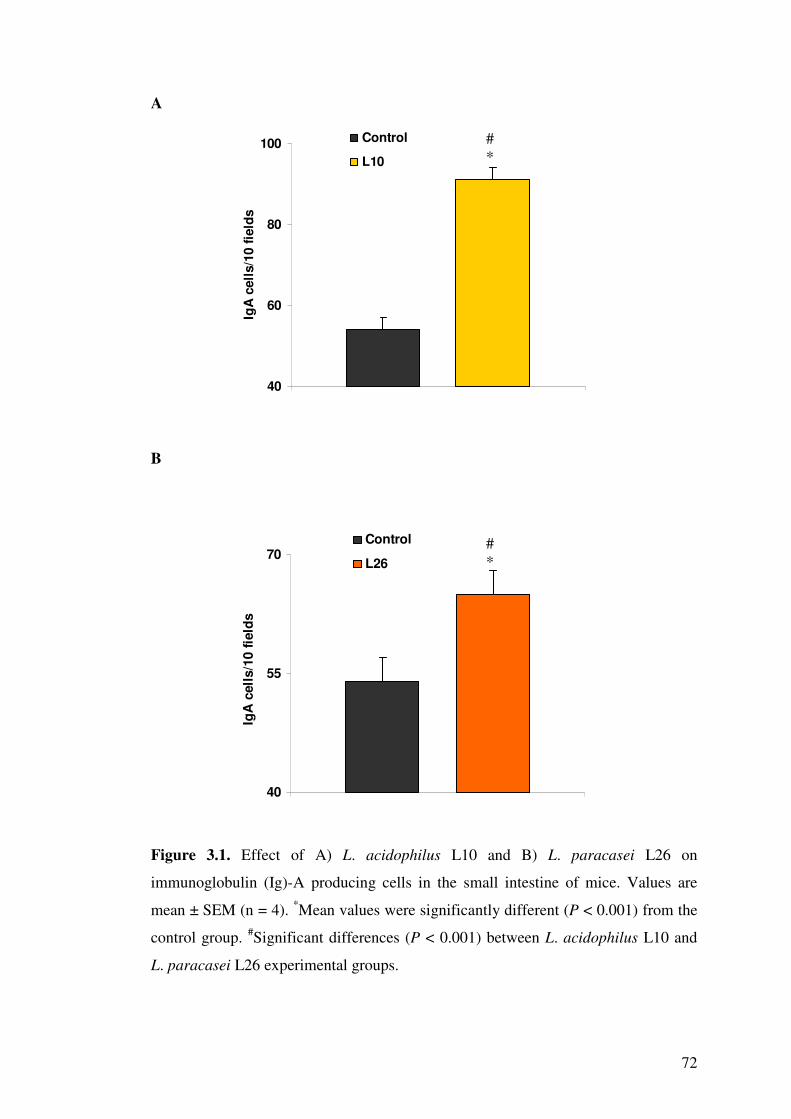
72
A
40
60
80
100
IgA
cell
s/1
0 f
ield
s
Control
L10
B
40
55
70
IgA
cell
s/1
0 f
ield
s
Control
L26
Figure 3.1. Effect of A) L. acidophilus L10 and B) L. paracasei L26 on
immunoglobulin (Ig)-A producing cells in the small intestine of mice. Values are
mean ± SEM (n = 4). *Mean values were significantly different (P < 0.001) from the
control group. #Significant differences (P < 0.001) between L. acidophilus L10 and
L. paracasei L26 experimental groups.
# *
# *
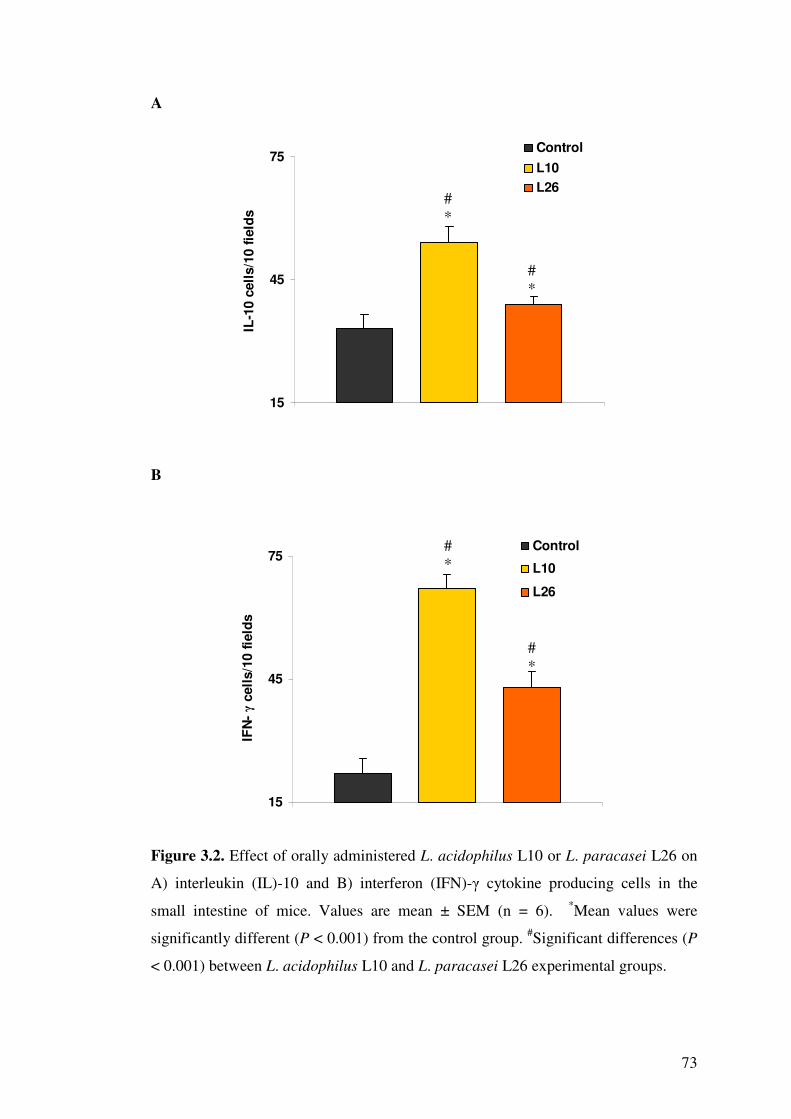
73
A
15
45
75
IL-1
0 c
ell
s/1
0 f
ield
s
Control
L10
L26
B
15
45
75
IFN
- � c
ell
s/1
0 f
ield
s
Control
L10
L26
Figure 3.2. Effect of orally administered L. acidophilus L10 or L. paracasei L26 on
A) interleukin (IL)-10 and B) interferon (IFN)-� cytokine producing cells in the
small intestine of mice. Values are mean ± SEM (n = 6). *Mean values were
significantly different (P < 0.001) from the control group. #Significant differences (P
< 0.001) between L. acidophilus L10 and L. paracasei L26 experimental groups.
# *
# *
# *
# *

74
Table 3.1. Proliferative responses of spleen cells stimulated with ConA and LPS in
mice fed with either L. acidophilus L10 or L. paracasei L26.
Mitogen stimulated splenocytes (Absorbance – OD 492 nm)
Group ConA LPS
Control
3.28 ± 0.19 2.19 ± 0.07
L. acidophilus L10
5.04 ± 0.29 **, # 4.31 ± 0.10 *, #
L. paracasei L26
4.25 ± 0.32 *, # 2.71 ± 0.08 #
Values are mean ± SEM (n = 6).
Significantly different from the control mice *P < 0.05; **P < 0.01.
Significant differences between L. acidophilus L10 and L. paracasei L26
experimental groups #P < 0.01.
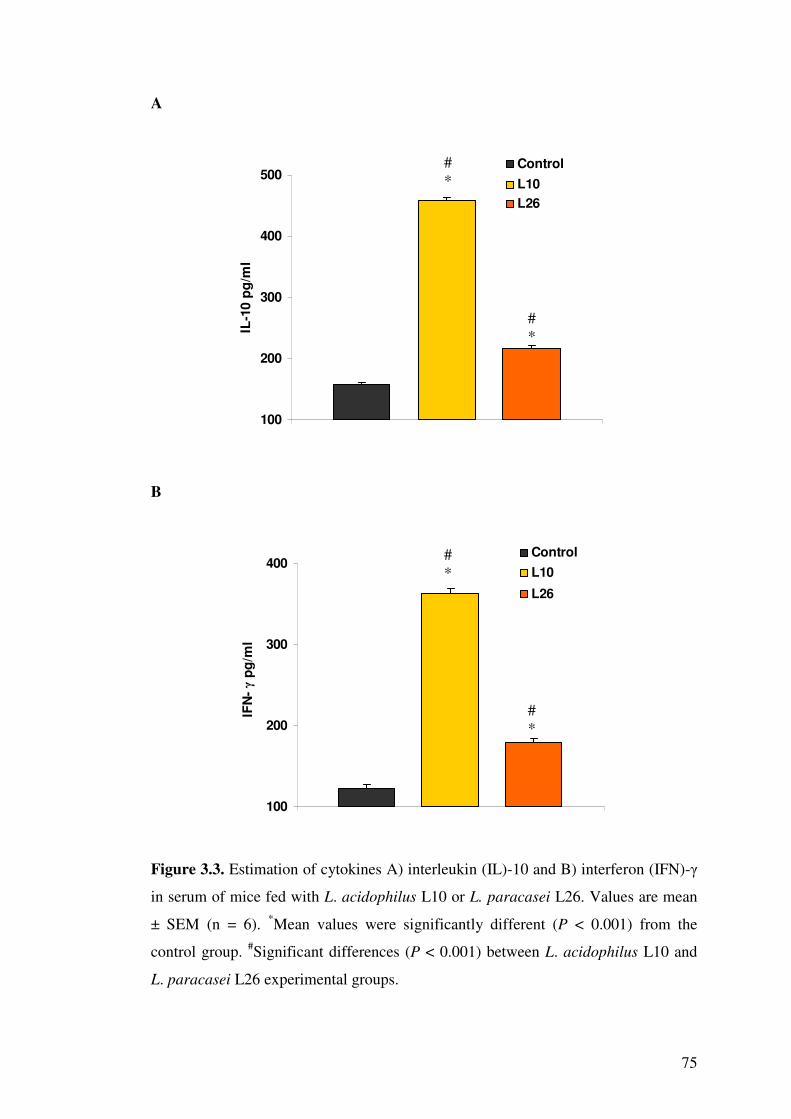
75
A
100
200
300
400
500
IL-1
0 p
g/m
l
Control
L10
L26
B
100
200
300
400
IFN
- � p
g/m
l
Control
L10
L26
Figure 3.3. Estimation of cytokines A) interleukin (IL)-10 and B) interferon (IFN)-�
in serum of mice fed with L. acidophilus L10 or L. paracasei L26. Values are mean
± SEM (n = 6). *Mean values were significantly different (P < 0.001) from the
control group. #Significant differences (P < 0.001) between L. acidophilus L10 and
L. paracasei L26 experimental groups.
# *
# *
# *
# *
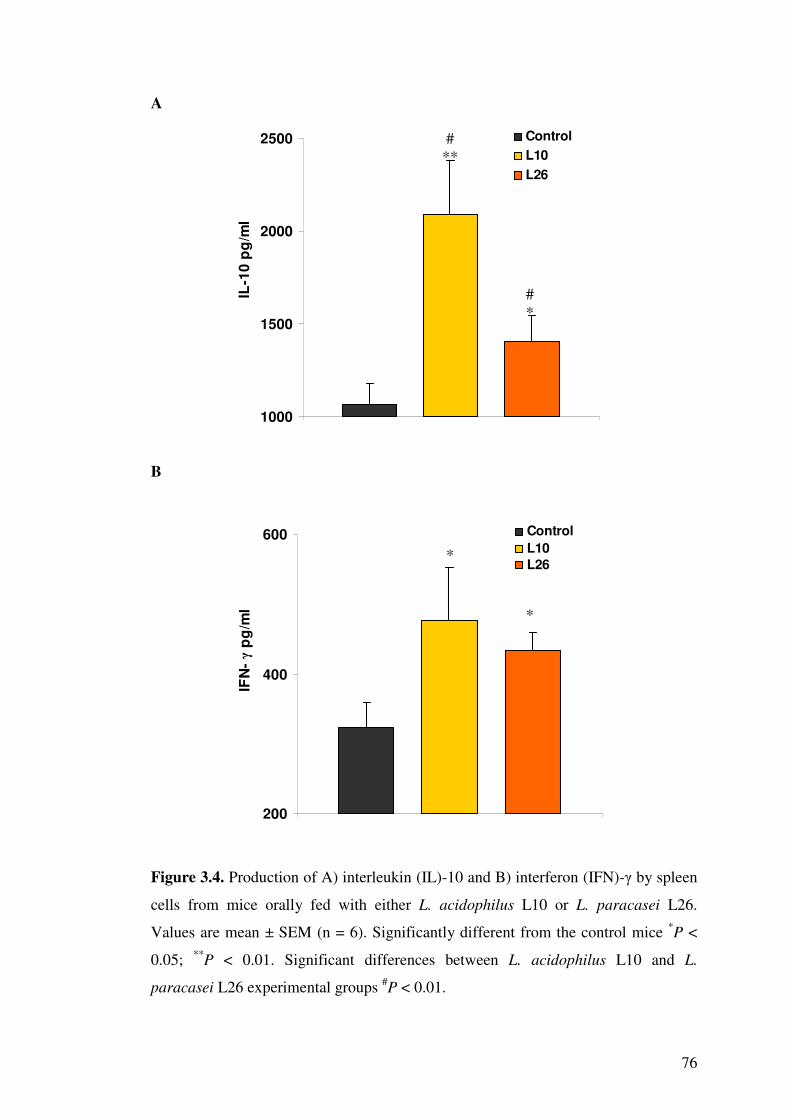
76
A
1000
1500
2000
2500
IL-1
0 p
g/m
l
Control
L10
L26
B
200
400
600
IFN
- � p
g/m
l
Control
L10
L26
Figure 3.4. Production of A) interleukin (IL)-10 and B) interferon (IFN)-� by spleen
cells from mice orally fed with either L. acidophilus L10 or L. paracasei L26.
Values are mean ± SEM (n = 6). Significantly different from the control mice *P <
0.05; **P < 0.01. Significant differences between L. acidophilus L10 and L.
paracasei L26 experimental groups #P < 0.01.
*
*
# *
# **

77
3.4.5. Phagocytic activity
The phagocytic activity of peritoneal macrophages in mice fed with either L.
acidophilus or L. paracasei was determined (Fig. 3.5A, B). Macrophages isolated
from the peritoneal cavity of mice fed with L. acidophilus or L. paracasei showed
significantly increased (P < 0.01) phagocytic activity against Candida albicans when
compared with the control mice. In contrast, L. acidophilus and L. paracasei strains
showed no differences (P > 0.05) in inducing phagocytic activity of the peritoneal
macrophages.
3.4.6. Bacterial translocation
Bacterial translocation to Peyer’s patches, MLN, spleen, liver and blood was
determined in mice fed with L. acidophilus or L. paracasei to evaluate possible
adverse effects that could lead to systemic dysfunction. Translocation of
Lactobacillus spp. to Peyer’s patches was significantly different in mice fed with L.
acidophilus (P < 0.01) or L. paracasei (P < 0.05) compared to control mice (Table
3.2). In mice fed with L. acidophilus or L. paracasei, Lactobacillus spp. translocated
to MLN was also increased (P < 0.01) compared to control mice. There were no
differences (P > 0.05) between mice fed with L. acidophilus and L. paracasei in
relation to the increased translocation of Lactobacillus spp. to Peyer’s patches and
MLN. Furthermore, there was no incidence of translocation of Lactobacillus spp. to
spleen, liver and blood in mice fed with either L. acidophilus or L. paracasei and
control group (Table 3.2).
Total anaerobes translocated to Peyer’s patches in L. acidophilus-fed mice was
higher (P < 0.01) compared to control mice (Table 3.3). Translocation of total

78
anaerobes to Peyer’s patches was also increased in mice fed with L. paracasei but
not significantly different (P > 0.05) when compared with control mice. Mice fed
with either L. acidophilus or L. paracasei showed increased (P < 0.01) translocation
of total anaerobes to MLN compared to control mice (Table 3.3). There were no
significant differences (P > 0.05) between mice fed with L. acidophilus and L.
paracasei in relation to increased translocation of total anaerobes to Peyer’s patches.
However, L. acidophilus and L. paracasei strains demonstrated the differences in
translocation of total anaerobes to MLN. There was no evidence of translocation of
total anaerobes to spleen, liver and blood in mice fed with either L. acidophilus or L.
paracasei and control group (Table 3.3).
In mice fed with L. acidophilus or L. paracasei and control group, translocation of
enteric bacteria to Peyer’s patches, MLN, spleen, liver and blood was not detected
(Table 3.4).
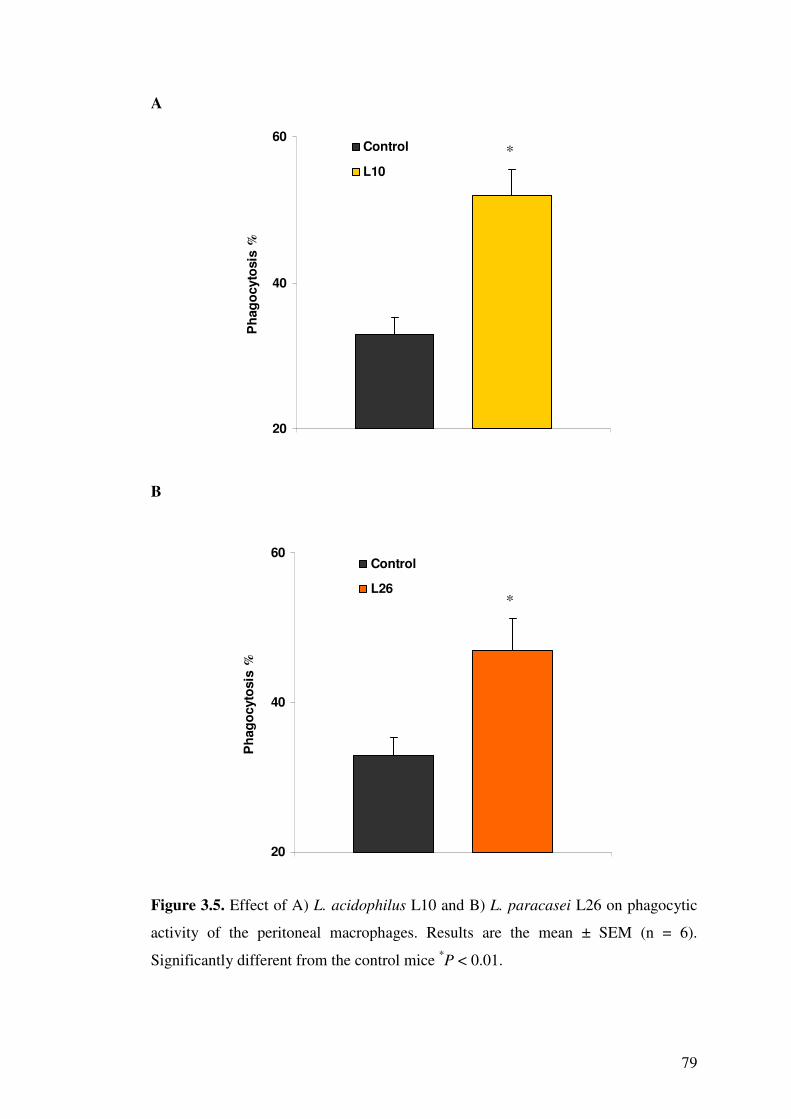
79
A
20
40
60
Ph
ag
ocyto
sis
%
Control
L10
B
20
40
60
Ph
ag
ocyto
sis
%
Control
L26
Figure 3.5. Effect of A) L. acidophilus L10 and B) L. paracasei L26 on phagocytic
activity of the peritoneal macrophages. Results are the mean ± SEM (n = 6).
Significantly different from the control mice *P < 0.01.
*
*
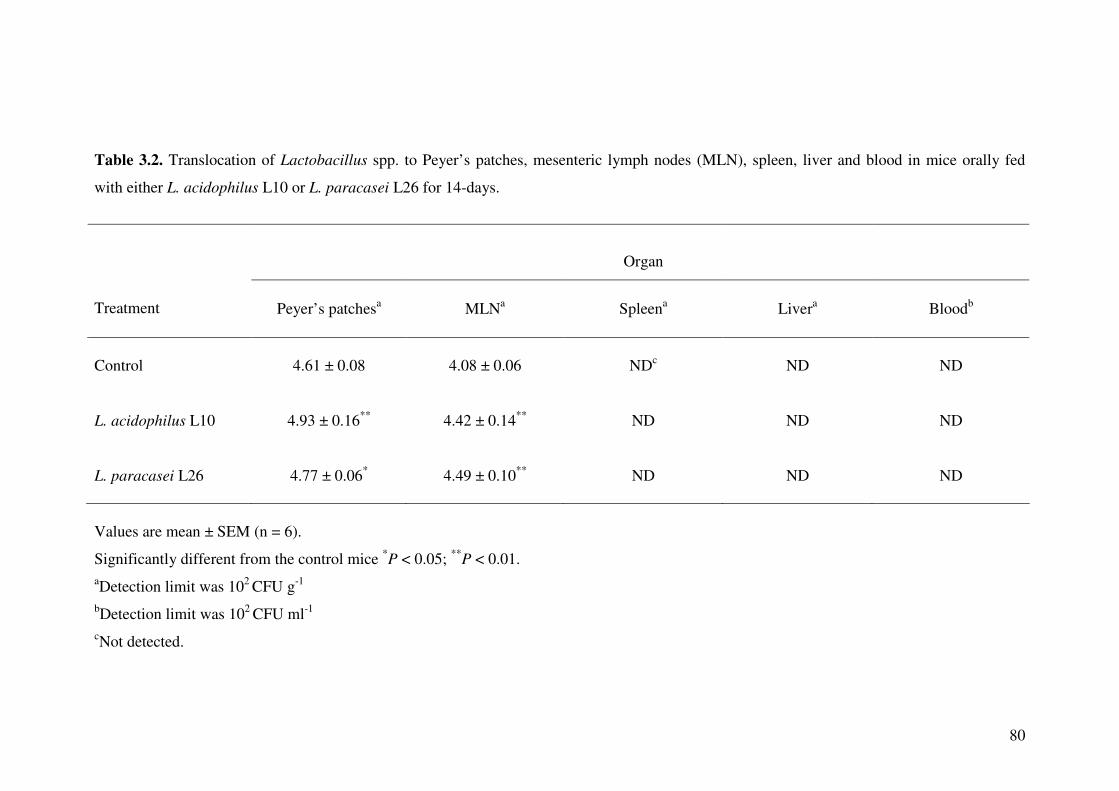
80
Table 3.2. Translocation of Lactobacillus spp. to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed
with either L. acidophilus L10 or L. paracasei L26 for 14-days.
Organ
Treatment Peyer’s patchesa MLNa Spleena Livera
Bloodb
Control
4.61 ± 0.08 4.08 ± 0.06 NDc ND
ND
L. acidophilus L10
4.93 ± 0.16** 4.42 ± 0.14** ND ND
ND
L. paracasei L26
4.77 ± 0.06* 4.49 ± 0.10** ND ND
ND
Values are mean ± SEM (n = 6).
Significantly different from the control mice *P < 0.05; **P < 0.01.
aDetection limit was 102 CFU g-1
bDetection limit was 102 CFU ml-1
cNot detected.
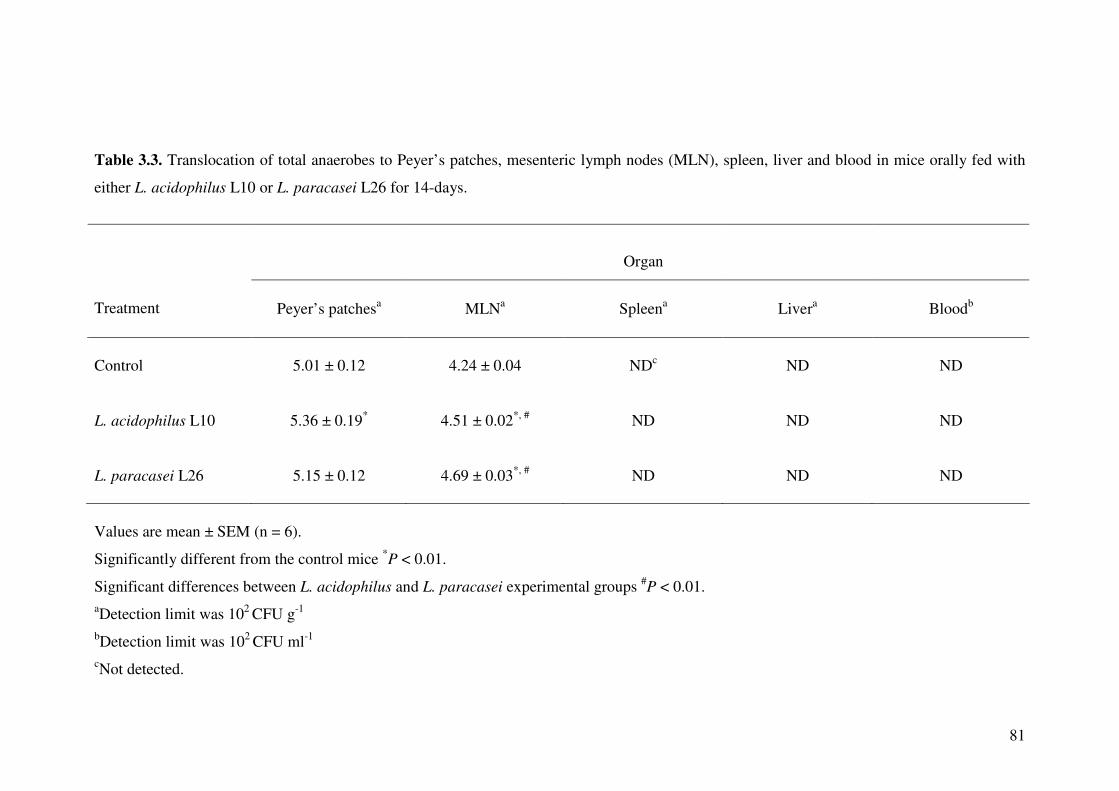
81
Table 3.3. Translocation of total anaerobes to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed with
either L. acidophilus L10 or L. paracasei L26 for 14-days.
Organ
Treatment Peyer’s patchesa MLNa Spleena Livera
Bloodb
Control
5.01 ± 0.12 4.24 ± 0.04 NDc ND
ND
L. acidophilus L10
5.36 ± 0.19* 4.51 ± 0.02*, # ND ND
ND
L. paracasei L26
5.15 ± 0.12 4.69 ± 0.03*, # ND ND
ND
Values are mean ± SEM (n = 6).
Significantly different from the control mice *P < 0.01.
Significant differences between L. acidophilus and L. paracasei experimental groups #P < 0.01.
aDetection limit was 102 CFU g-1
bDetection limit was 102 CFU ml-1
cNot detected.
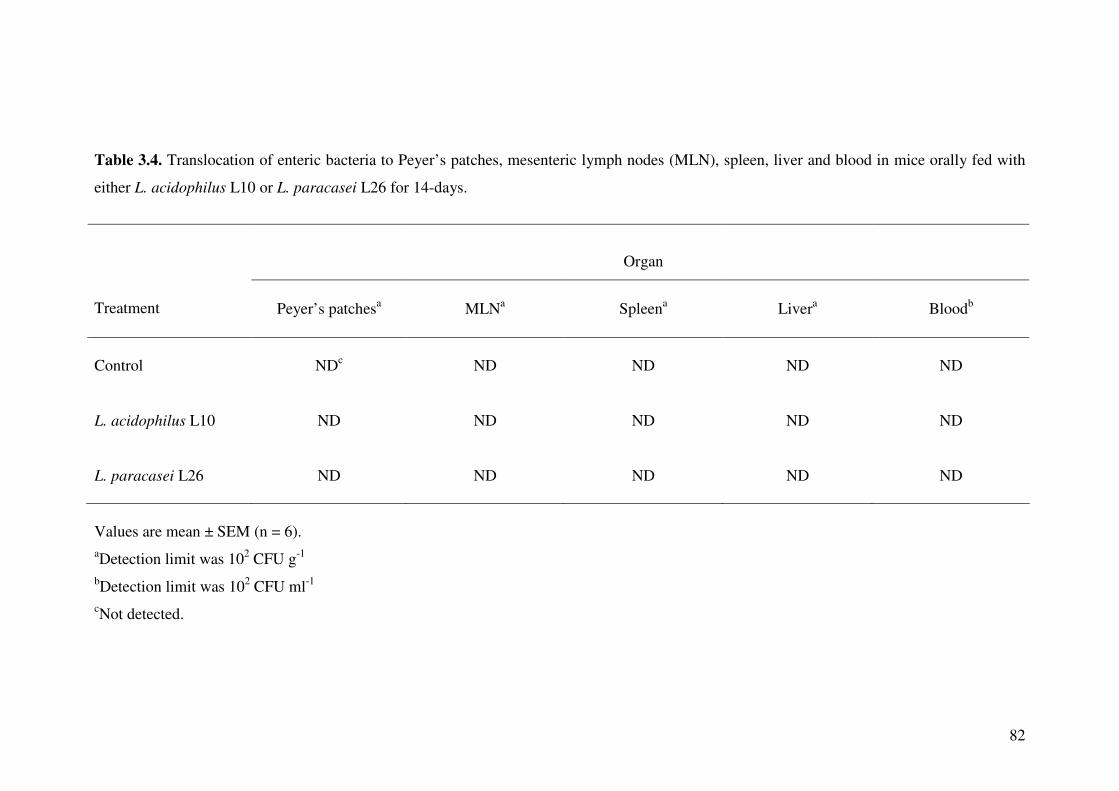
82
Table 3.4. Translocation of enteric bacteria to Peyer’s patches, mesenteric lymph nodes (MLN), spleen, liver and blood in mice orally fed with
either L. acidophilus L10 or L. paracasei L26 for 14-days.
Organ
Treatment Peyer’s patchesa MLNa Spleena Livera
Bloodb
Control
NDc ND ND ND
ND
L. acidophilus L10
ND ND ND ND
ND
L. paracasei L26
ND ND ND ND
ND
Values are mean ± SEM (n = 6).
aDetection limit was 102 CFU g-1
bDetection limit was 102 CFU ml-1
cNot detected.

83
3.5. DISCUSSION
The external secretions of IgA in GI tract play an active role in protecting the
surfaces of mucus membrane from pathogenic infections and carcinogens (Mestecky
and McGhee, 1987). The formation of immune complexes in intestinal lumen to bind
antigens was the protective effect of antigen-specific secretory IgA, which interferes
with the adherence of pathogenic microorganisms to intestinal epithelial cells and
also helps in neutralising toxins and viruses (Mestecky et al., 1999). In the current
study, L. acidophilus and L. paracasei strains enhanced the number of IgA producing
cells in the small intestine of mice (Fig. 3.1A, B), which suggest that these strains
could protect the GI tract from pathogenic microorganisms through increased IgA
response. In comparison to L. paracasei, L. acidophilus showed an increased number
of IgA producing cells in the lamina propria that could be beneficial to the host in
conferring greater protection against pathogenic microorganisms. Earlier, it was
reported that peptides derived from L. helveticus fermented milk enhanced the IgA
secretion in gut and systemic immunity in mice challenged with E. coli O157:H7
(Leblanc et al., 2004).
The Th1 cells produce important factors such as IL-2, IFN-� and tumor necrosis
factor for cell-mediated immunity. Cytokines like IL-4, IL-5 and IL-10 associated
with humoral immunity and allergic responses are produced by Th2 cells (Kidd,
2003). The Th1 cytokine, IFN-� is a multifunctional pro-inflammatory cytokine that
triggers innate immune responses such as phagocytosis and antimicrobial activity,
whereas Th2 cytokine, IL-10 regulates a number of events including IFN-� and
antibody production (Kidd, 2003). Both IL-10 and IFN- � were considered as pivotal
cytokines in inducing Th1 and Th2 responses (Perdigon et al., 2002). In this study, L.

84
acidophilus and L. paracasei strains were effective in activating mucosal and
systemic immune responses through the release of IL-10 and IFN-� cytokines (Fig.
3.2; 3.3; 3.4). Perdigon et al. (1990) reported that mice fed with fermented milk
consisting of LAB were protected against Salmonella Typhimurium infection.
Furthermore, Gould and Sonnenfeld, (1987) demonstrated that mice pre-treated with
IFN-� were also protected against Salmonella Typhimurium strain LT-2. In the
present study, increase of IFN-� producing cells in the intestinal mucosa of mice fed
with L. acidophilus or L. paracasei could therefore contribute to the protection of
mucosal surfaces against harmful pathogenic microorganisms.
Lymphocytes play an important role in the regulation of immune responses. T-
lymphocytes are the mediators of cellular immunity, whereas B-lymphocytes
develop into antibody producing cells in response to foreign antigens. In the current
study, mitogen-stimulated lymphocyte proliferative responses were used to asses the
T- and B-cell functions. Mice fed with L. acidophilus or L. paracasei exhibited the
enhancement of splenocyte proliferative responses to T-cell mitogen (ConA) and B-
cell mitogen (LPS). Although, LPS stimulated B-cell proliferative responses in L.
paracasei-fed mice was higher, but not significantly different from the control mice
(Table 3.1). The present study demonstrated the differences in mitogen-stimulated
proliferative responses of splenocytes, which could be due to strain-specific
variations. Kirjavainen et al. (1999) demonstrated similar strain-specific effects of
mitogen-stimulated splenocyte proliferative responses of T- and B-lymphocytes.
It is widely known that the balance between Th1 and Th2 responses was important to
elevate immunological responses against various diseases. Lactic acid bacteria are

85
known to modulate the balance of Th1 and Th2 immune responses (Maassen et al.,
2000; Perdigon et al., 2002). Human volunteers fed with lyophilised lactobacilli
showed an increased production of IFN-� in serum (De Simone et al., 1993). Schultz
et al. (2003) reported that oral administration of L. rhamnosus GG to healthy human
volunteers stimulated an increased secretion of anti-inflammatory cytokines (IL-10
and IL-4) and decreased secretion of pro-inflammatory cytokines (IFN-�, TNF-� and
IL-6). In the present study, IL-10 was significantly increased in serum as well as in
splenocytes stimulated with ConA of mice fed with L. acidophilus or L. paracasei.
The magnitude in the increment of IL-10 was higher than Th1 cytokine IFN-�, which
may be necessary to control the increase of the pro-inflammatory cytokine IFN-�
(Fig. 3.3; 3.4).
In the current study, splenocytes from mice fed with L. acidophilus or L. paracasei
significantly increased the production of IL-10 and IFN-� cytokines (Fig. 3.4A, B).
Roller et al. (2004) reported similar enhancement of IL-10 and IFN-� production in
Peyer’s patches of rats fed with synbiotics (Probiotics and prebiotics). In another
study, CD4+ T-cells from Peyer’s patches of mice fed with dietary
fructooligosaccharides showed dose-dependent increased production of IL-10 and
IFN-� levels (Hosono et al., 2003). The current study demonstrates that L.
acidophilus and L. paracasei were able to influence the regulation of Th1 and Th2
cytokines in gut and systemic immunity through increased production of IL-10 and
IFN-� (Fig. 3.2; 3.3; 3.4). In contrast, L. acidophilus-fed mice stimulated higher
levels of IL-10 and IFN-� compared to the increase detected in mice fed with L.
paracasei.

86
The influence of LAB on phagocytosis could be strain-specific, dose dependent or
survivability in the GI tract (Gill et al., 2000; Gill and Rutherfurd, 2001; Medici et
al., 2005). The differences in cell surface structures of Lactobacillus strains lead to
strain-specific variations in modulating immune responses (Sato et al., 1988). The
ability of LAB in stimulating phagocytic cells also depends on its survival and
persistence in GI tract and their adherence to intestinal epithelial cells (Perdigon and
Alvarez, 1992; Schiffrin et al., 1997). In the present study, orally administered L.
acidophilus or L. paracasei strains enhanced the phagocytic activity of peritoneal
macrophages (Fig. 3.5A, B), which suggests that these strains are tolerant to gastric
conditions of GI tract and can persist through the GI passage to induce host
immunological responses. Mice fed with L. acidophilus showed an increase of 21%
activation of peritoneal macrophages compared to control mice, whereas L.
paracasei-fed mice showed only 6% activation. However, there were no significant
differences observed between L. acidophilus and L. paracasei in inducing
macrophage activity.
Gut mucosal integrity is crucial in host immune defence mechanisms, such as
protection against pathogenic bacteria through interfering their adherence to
intestinal epithelial cells (Mestecky et al., 1999). Translocation of indigenous
microflora to distant intestinal sites such as spleen, liver and blood could occur due
to intestinal inflammation that causes the disruption of intestinal mucosal surfaces.
Vinderola et al. (2004) reported the translocation of various doses of different LAB
to liver, which activated gut and systemic immune responses. In the present study,
there was no evidence of translocation of intestinal microflora to spleen, liver or
blood in both the experimental group mice (Table 3.2; 3.3), which suggests that oral

87
administration of L. acidophilus or L. paracasei at 108 CFU/day did not induce
adverse effects such as disruption of immune defence mechanisms that leads to
inflammation of intestinal mucosal layers and bacterial translocation to sterile
organs.
The absence of bacterial translocation to spleen, liver or blood also demonstrates the
active role of immune defence mechanisms that could have inhibited the
translocation beyond MLN. Additionally, enteric bacteria were not detected in
Peyer’s patches, MLN, spleen, liver and blood either in control or experimental
group mice (Table 3.4).
Systemic dissemination of bacteria in mice has been correlated with the adhesion of
probiotic bacteria to intestinal epithelial cells (Wagner et al., 1997). Earlier reports
suggested that adhesion of bacterial strains to gut epithelial cells is necessary to
modulate immune responses (Perdigon and Alvarez, 1992; Schiffrin et al., 1997). In
the present study, L. acidophilus-fed mice showed increased translocation of
Lactobacillus spp. to Peyer’s patches (7%) and MLN (8%) compared to control
mice, whereas L. paracasei-fed mice demonstrated the translocation of Lactobacillus
spp. with an increase of 3% to Peyer’s patches and 10% to MLN (Table 3.2).
However, increased translocation of Lactobacillus spp. in mice fed with L.
acidophilus or L. paracasei were statistically significant compared to control mice
but showed very small differences. Similar small differences were also observed in
the translocation of total anaerobes to Peyer’s patches and MLN in both the
experimental groups with respect to control mice (Table 3.3). The results of this
study suggest that L. acidophilus and L. paracasei have potential adhesive properties

88
that can lead to the translocation of Lactobacillus spp. to Peyer’s patches and MLN,
which may contribute to the enhancement of various systemic immune functions.

89
3.6. CONCLUSIONS
Orally administered L. acidophilus and L. paracasei demonstrated the enhancement
of gut mucosal immunity in relation to the increase of IgA and cytokines (IL-10 and
IFN-�). In systemic immunity, L. acidophilus and L. paracasei enhanced the
phagocytic activity of peritoneal macrophages and release of IL-10 and IFN-�
cytokines from serum and splenocytes. In contrast, L. acidophilus showed higher
stimulation of gut and systemic immune responses compared to L. paracasei.
Additionally, there was no evidence of bacterial translocation to spleen, liver or
blood in mice fed with either L. acidophilus or L. paracasei. Therefore, L.
acidophilus and L. paracasei strains are considered non-pathogenic, as indicated by
their effects on bacterial translocation in healthy mice.

90
CHAPTER 4
Immunostimulatory responses of L. acidophilus LAFTI L10 and L. paracasei
LAFTI L26 in cholera toxin mice

91
4.1. ABSTRACT
Mice were orally fed with either L. acidophilus or L. paracasei for 14-days, during
which they were immunized twice with 10 �g of cholera toxin (CT). Lactobacillus
acidophilus-fed mice showed an increase (P < 0.01) in serum nitric oxide (NO)
levels. The increment of NO levels in intestinal fluid and serum of L. paracasei-fed
mice was significantly different compared to control mice. Immunoglobulin (Ig)-A
response to CT in intestinal fluid and serum were significantly enhanced by L.
acidophilus or L. paracasei. Cytokine producing cells in small intestine of mice fed
with either L. acidophilus or L. paracasei demonstrated the regulation of Th1 and
Th2 cytokines such as interferon (IFN)-�, tumor necrosis factor (TNF)-�, interleukin
(IL)-4, IL-6 and IL-10. The results of this study demonstrate that L. acidophilus and
L. paracasei are capable of enhancing gut immune responses as well as serum NO
and IgA in systemic levels of CT immunized mice.

92
4.2. INTRODUCTION
Gastrointestinal (GI) tract is considered to be a complex ecosystem consisting of
various resident microorganisms such as Escherichia coli, Clostridium spp.,
Streptococcus spp., Lactobacillus spp., Bacteroides spp., and Bifidobacterium spp.,
which are involved in the development of gut microflora and maintains host health
by exerting specific gut functions (McCracken and Lorenz, 2001). The microbial
population in GI tract, specifically, Lactobacillus and Bifidobacterium spp. were
considered as probiotic due to their beneficial health effects such as inhibiting the
growth of Escherichia coli (Forestier et al., 2001), Helicobacter pylori (Wang et al.,
2004) and resistance to diseases (Shanahan, 2002; de Waard et al., 2003).
Microorganisms of GI tract have the potential to influence immunoglobulin (Ig)-A
production for the development and activation of gut immune system (Dogi and
Perdigon, 2006). Furthermore, commensal bacteria also help in promoting the
development of B-cell population in Peyer’s patches that were not developed in
germ-free animals. In comparison to germ-free animals, IgA producing cells were
higher in intestinal lamina propria of animals possessing normal intestinal microflora
(Crabbe et al., 1968; Jiang et al., 2004). Due to the significance of IgA in host
immune functions, it is important to screen the capability of probiotic bacteria in
enhancing antibody response in gut and systemic immunity.
Oral administration of appropriate microbial strains could potentially restore and
maintain oral tolerance and host immune functions (Sudo et al., 1997; Kalliomaki et
al., 2001). Korhonen et al. (2001) reported that L. rhamnosus GG induced the
production of nitric oxide (NO), which played a functional role in protective
mechanisms of intestinal mucosa and regulating immune functions. In rats, dietary

93
supplementation of food with lactobacilli and nitrate increased the NO levels in small
intestine and caecum (Sobko et al., 2006). The endogenous NO was known to have
potential role in regulating various host functions including water and electrolyte
transport (Mourad et al., 1999) and motility (Mashimo et al., 2000). Scientific reports
speculated that some of the beneficial effects exerted by probiotic bacteria were
mediated through NO release in intestinal lumen (Xu and Verstraete, 2001; Lamine
et al., 2004; Sobko et al., 2005). Sobko et al. (2005) suggested the relationship
between increased number of intestinal microflora with higher NO levels in the
intestine of healthy new born infants. The source of NO detected in the lumen of new
born infants however remains unclear.
Gastrointestinal epithelial cells act as physical and chemical barriers by protecting
the host from harmful microorganisms that could hijack the regulation of cellular
molecules and signalling pathways (Cossart, 1997; Finlay and Falkow, 1997).
Epithelial cells of GI tract were considered important in mucosal immune system due
to their interdependency on adjacent lymphoid cells (Shanahan, 1999). Cytokines
such as interleukin (IL)-6, IL-10 and tumor necrosis factor (TNF)-� were few of the
several cytokines secreted by intestinal epithelial cells (McCracken and Gaskins,
1999). Enhanced secretory antibodies and variations in mucous layers along the
intestine could be crucial in strengthening the intestinal barrier of GI tract (Yuan and
Walker, 2004). Earlier studies reported the capability of probiotic bacteria in
normalising and reinforcing gut barrier functions and protecting the host from
pathogenic bacteria (Terpend et al., 1999; Madsen et al., 2001).

94
Cholera toxin (CT), the enterotoxin of Vibrio cholerae, which affects epithelial cells
by inducing massive salt and water secretion results in severe diarrhoea. The toxin is
considered as a potent adjuvant and immunogen that can influence mucosal and
systemic immune functions (Elson and Ealding, 1984). Several scientific reports
demonstrated CT as a mucosal adjuvant by enhancing IgA and IgG immune
responses, increasing T-cell antigen priming and inhibiting oral tolerance to
unrelated antigens (Elson and Ealding, 1984; Lycke et al., 1985; Hornqvist et al.,
1991). Mucosal adjuvants are also known to activate immune responses by
stimulating cytokines that are involved in innate and acquired immunity (Elson and
Dertzbaugh, 1994). The degradation of orally administered antigens could occur in
GI tract due to constant changes such as low gastric pH and high bile salts, which
further may lead to either ineffective or minimal immune responses. Considerable
attention was given to lactic acid bacteria as an adjuvant due to their general use as
safe food grade microorganisms in the manufacturing of fermented dairy products.
Several studies have shown the differences between LAB strains in inducing
adjuvant effects when delivered orally with antigens (McCracken and Gaskins, 1999;
Tejada-Simon et al., 1999a; Plant and Conway, 2002).
As shown in chapter 3, L. acidophilus LAFTI L10 and L. paracasei LAFTI L26
demonstrated the stimulation of various immune functions in gut and systemic
immunity. Particularly, L. acidophilus L10 demonstrated higher stimulation of gut
and systemic immune responses with respect to L. paracasei L26. However, it is
unclear whether L. acidophilus L10 and L. paracasei L26 have the potential to alter
the regulation of B- and T-cell functions after oral immunisation with an antigen in
BALB/c mice. Therefore, present study aimed to use CT as an oral antigen to

95
determine the immune responses enhanced by oral administration of L. acidophilus
L10 and L. paracasei L26 in mice. The effects of L. acidophilus L10 and L.
paracasei L26 on various indices of immune functions such as NO, CT-specific IgA
response and Th1/Th2 cytokine producing cells in the small intestine and serum NO
and CT-specific IgA levels were evaluated.

96
4.3. MATERIALS AND METHODS
4.3.1. Mice
Male 6-week old BALB/c mice were purchased from Biological Resources Centre,
The University of New South Wales, Sydney, Australia. Mice were housed at 22 ±
1°C under 12 h light-dark photoperiod and fed with standard mouse chow (Gordon’s
Speciality Stock Feeds, Sydney, Australia) and provided ad libitum access to water.
Control and experimental groups consisted of six mice each, which were randomly
allocated. All experiments were performed with the approval from Animal Care and
Ethics Committee of University of Western Sydney, Sydney, Australia.
4.3.2. Lactobacillus strains and feeding procedure
Lactobacillus acidophilus LAFTI L10 and Lactobacillus paracasei LAFTI L26 used
in this study were obtained from DSM culture collection (DSM Food Specialties
Ltd., Sydney, Australia). The bacterial strains were grown under anaerobic
conditions in gas jars using GasPak System (Oxoid, Adelaide, Australia) at 37°C for
24 h in de Man, Rogosa and Sharpe (MRS) broth (Oxoid, Adelaide, Australia) and
washed twice with sterile 0.01 M phosphate buffered saline (PBS; Sigma-Aldrich,
Sydney, Australia) solution following centrifugation at 4000 x g for 15 min at 4°C.
The viability of bacteria was determined by spread plate count of bacterial serial
dilutions on MRS agar (Oxoid, Adelaide, Australia), which were incubated
anaerobically for 48 h at 37°C. After acclimatisation for 1 week, mice (six per group)
were fed daily by oral gavage with 108 colony-forming units (CFU) of either L.
acidophilus or L. paracasei in 50 �l of 10% (w/v) skimmed milk powder (SMP) for
14-days. Control group mice received 50 �l of SMP without lactic acid bacteria.

97
4.3.3. Cholera toxin immunisation
Control and experimental group mice were orally immunized with 10 �g of cholera
toxin (Sigma-Aldrich, Sydney, Australia) in 25 �l of 0.1 M sodium bicarbonate
(Sigma-Aldrich, Sydney, Australia) on day 0 and 7. Mice were sacrificed by carbon
dioxide inhalation after 14-day feeding trail and intestinal fluid, blood and small
intestines were collected.
4.3.4. Intestinal fluid and serum preparation
Intestinal fluid of small intestine in each mouse was collected by flushing the
intestinal contents with 2 ml of 0.01 M PBS and centrifuged at 10,000 x g for 15 min
at 4°C. The supernatant was collected and stored at -20°C until use.
Blood was collected upon euthanizing the mice and left to coagulate overnight at 4°C
and centrifuged at 2000 x g for 10 min (4°C). The serum was collected and stored at
-20°C until use.
4.3.5. Determination of nitric oxide
Nitric oxide was determined by measuring nitrite accumulation in intestinal fluid and
serum using Griess reagent system (Promega, Sydney, Australia). Briefly, 50 µl of
intestinal fluid supernatant or serum were added in triplicates to the wells, which
were incubated with 50 µl of sulfanilamide solution for 10 min at room temperature
while protected from light. After adding 50 µl of N-1-napthylethylenediamine
dihydrochloride solution to all the wells, absorbance was read at 540 nm using a
microtitre plate reader (Multiskan Titertek, Huntsville, USA).

98
4.3.6. Estimation of Immunoglobulin A
The CT-specific IgA antibodies in intestinal fluid and serum were detected by
ELISA. Briefly, each well of the 96-well microtitre plate was coated with 100 �l of
CT (10 �g/ml) in 0.05 M carbonate-bicarbonate buffer, pH 9.6. For standard, 100 �l
of goat anti-mouse IgA (Bethyl Laboratories Inc., Montgomery, USA) was added to
the wells in triplicates at 10 �g/ml in 0.05 M carbonate-bicarbonate buffer, pH 9.6.
The plates were incubated overnight at 4°C and washed three times with 0.01 M PBS
containing 0.05% Tween 20 (PBS-T; Sigma-Aldrich, Sydney, Australia). Then the
plates were incubated for 2 h at 37°C with either 100 �l of mouse IgA-kappa (Sigma-
Aldrich, Sydney, Australia) as a standard or 100 �l of intestinal fluid or serum, which
were added in triplicates. The plates were washed three times with PBS-T and
incubated with 100�l of goat anti-mouse IgA horseradish peroxidase conjugate
(Sigma-Aldrich, Sydney, Australia) for 90 min at 37°C. Incubation was followed by
washing the plates three times with PBS-T and 200�l of o-phenylenediamine
dihydrochloride peroxide substrate (Sigma-Aldrich, Sydney, Australia) was added to
the wells and incubated for 30 min at room temperature. The absorbance was read at
450 nm using a microtitre plate reader (Multiskan Titertek, Huntsville, USA).
4.3.7. Histological samples
Mice small intestines were removed, dissected into segments and stored in 95%
ethanol (v/v). The tissues were then embedded in paraffin blocks and the sections
were processed for immunofluorescence assays.

99
4.3.8. Immunofluorescent detection of cytokine producing cells
The number of cytokine secreting cells in small intestine of mice was determined by
indirect immunofluorescence assay. The paraffin sections (4-�m) were
deparaffinised in xylene and rehydrated in ethanol, which were then washed twice
with 0.01 M PBS solution and incubated with a blocking solution (0.01 M PBS
containing 1% bovine serum albumin) for 30 min at room temperature. The sections
were incubated with 0.2 �g/ml (diluted in 0.01 M PBS) of rabbit anti-mouse IL-4,
IL-6, IL-10, interferon (IFN)-� and TNF-� (Peprotech Inc., Rocky Hill, USA)
polyclonal antibodies for 75 min at 37°C. The incubation was followed by washing
the sections twice with 0.01 M PBS. The sections were then incubated with goat anti-
rabbit antibody conjugated with fluorescein isothiocyanate (FITC; Jackson Immuno
Research Labs Inc., West Grove, USA) for 45 min at room temperature and washed
twice with 0.01 M PBS. The number of fluorescent cells was counted by using
fluorescence light microscope (Olympus Optical Co., Tokyo, Japan) and the results
were expressed as number of positive fluorescent cells in 10 fields of vision.
4.3.9. Statistical analysis
The results were expressed as mean ± standard error of mean (SEM). The data were
analysed using the one-way analysis of variance procedure of SPSS (Version 12.0.1).
Significant differences between groups were identified by Duncan’s multiple range
test (SPSS, Version 12.0.1).

100
4.4. RESULTS
4.4.1. Nitric oxide production in intestinal fluid and serum
The effect of orally administered Lactobacillus strains on NO levels was determined
by Griess reagent system. As shown in Fig. 4.1A, L. paracasei showed significant
increase (P < 0.01) of NO levels in intestinal fluid compared to control mice. Nitric
oxide levels in intestinal fluid of mice fed with L. acidophilus were also increased
but not significantly different when compared to control mice. The serum NO levels
were increased significantly (P < 0.01) in mice fed with either L. acidophilus or L.
paracasei (Fig. 4.1B). The differences between L. acidophilus and L. paracasei were
significant (P < 0.01) in enhancing NO levels in intestinal fluid and serum.
4.4.2. Immunoglobulin A antibodies in intestinal fluid and serum
The IgA antibodies specific to CT in mice fed with L. acidophilus or L. paracasei
were estimated by ELISA (Fig. 4.2). In intestinal fluid, CT-specific IgA antibodies
were increased significantly (P < 0.01) in mice fed with L. acidophilus or L.
paracasei compared to control mice (Fig. 4.2A). In systemic response, strong serum
IgA response to CT was detected in mice fed with L. acidophilus or L. paracasei,
which differed significantly (P < 0.001) from control mice (Fig. 4.2B). There were
no significant differences (P > 0.05) between L. acidophilus and L. paracasei in
inducing CT-specific IgA response in intestinal fluid. In contrast, L. acidophilus and
L. paracasei demonstrated the differences (P < 0.01) in enhancing CT-specific IgA
antibodies in serum.
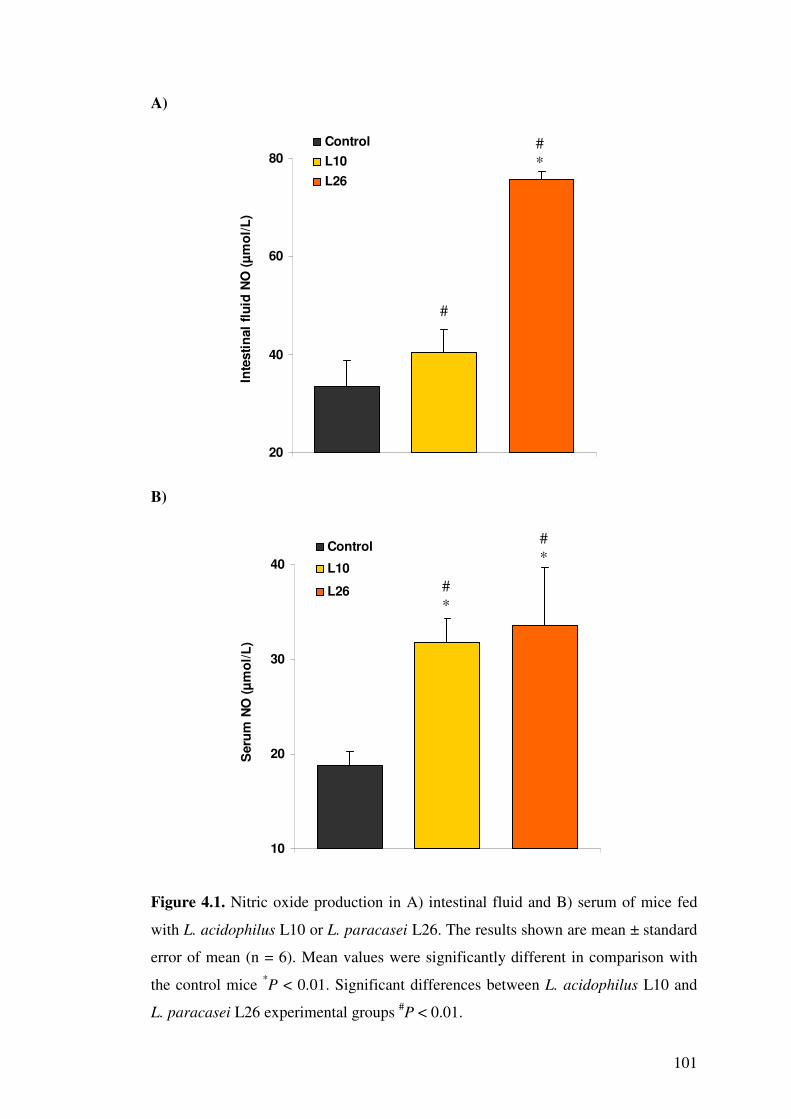
101
A)
20
40
60
80
Inte
sti
nal
flu
id N
O (
µm
ol/
L)
Control
L10
L26
B)
10
20
30
40
Seru
m N
O (
µm
ol/
L)
Control
L10
L26
Figure 4.1. Nitric oxide production in A) intestinal fluid and B) serum of mice fed
with L. acidophilus L10 or L. paracasei L26. The results shown are mean ± standard
error of mean (n = 6). Mean values were significantly different in comparison with
the control mice *P < 0.01. Significant differences between L. acidophilus L10 and
L. paracasei L26 experimental groups #P < 0.01.
#
*
#
#
*
#
*
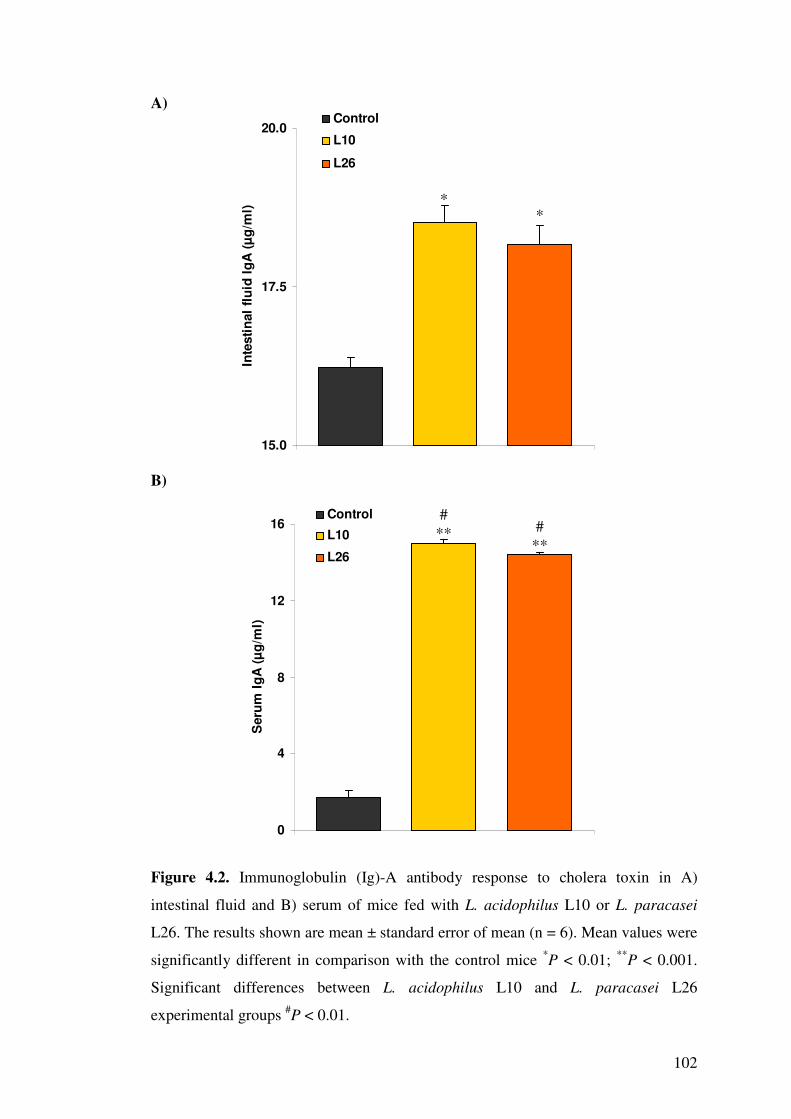
102
A)
15.0
17.5
20.0
Inte
sti
nal
flu
id I
gA
(µ
g/m
l)
Control
L10
L26
B)
0
4
8
12
16
Seru
m I
gA
(µ
g/m
l)
Control
L10
L26
Figure 4.2. Immunoglobulin (Ig)-A antibody response to cholera toxin in A)
intestinal fluid and B) serum of mice fed with L. acidophilus L10 or L. paracasei
L26. The results shown are mean ± standard error of mean (n = 6). Mean values were
significantly different in comparison with the control mice *P < 0.01; **
P < 0.001.
Significant differences between L. acidophilus L10 and L. paracasei L26
experimental groups #P < 0.01.
* *
#
** #
**

103
4.4.3. Detection of cytokine producing cells in small intestine
Cytokine producing cells in small intestine of mice fed with L. acidophilus or L.
paracasei were shown in Figures 4.3; 4.4. Th1 cytokine, IFN-� producing cells were
significantly increased in mice fed with L. acidophilus (P < 0.001) or L. paracasei (P
< 0.01) compared to control mice (Fig. 4.3A). The TNF-� was increased in mice fed
with L. acidophilus, but not significantly different from control mice. However,
TNF-� producing cells in L. paracasei-fed mice were significantly increased (P <
0.01) with respect to control mice (Fig. 4.3A).
In L. paracasei-fed mice, Th2 cytokine, IL-4 producing cells were enhanced (P <
0.01) when compared with control mice. Significant differences (P > 0.05) were not
detected in IL-4 producing cells when compared between mice fed with L.
acidophilus and control group (Fig. 4.3B). In comparison to control mice, IL-6
producing cells were increased in mice fed with either L. acidophilus (P < 0.05) or L.
paracasei (P < 0.001) (Fig. 4.3B). The IL-10 producing cells were also enhanced (P
< 0.01) in mice fed with L. acidophilus compared to control mice. The increase of
IL-10 producing cells of mice fed with L. paracasei did not show significant
differences (P > 0.05) when compared to control mice (Fig. 4.3B). There were no
significant differences (P > 0.05) between L. acidophilus and L. paracasei in
enhancing cytokines such as IFN-�, TNF-�, IL-4, IL-10 with exception to the
significant difference (P < 0.05) between L. acidophilus and L. paracasei in
increasing IL-6 cytokine producing cells in small intestine of mice.
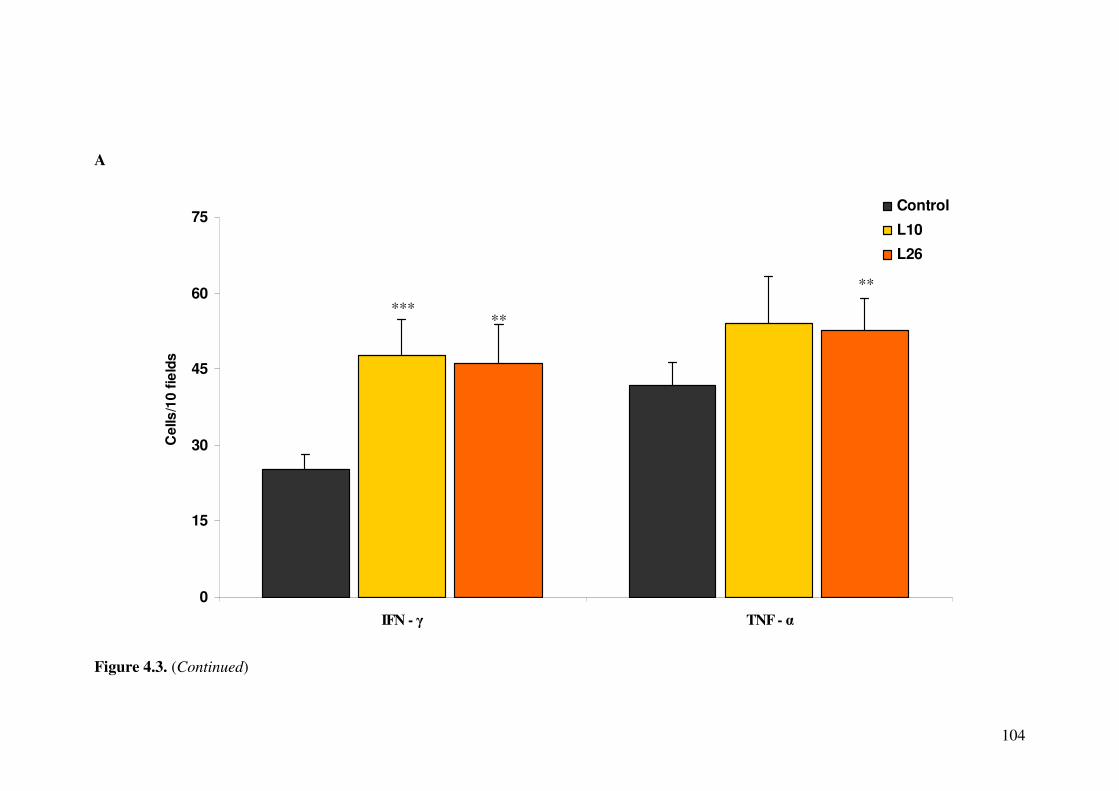
104
A
0
15
30
45
60
75
IFN - � TNF - �
Ce
lls
/10
fie
lds
Control
L10
L26
Figure 4.3. (Continued)
*** **
**
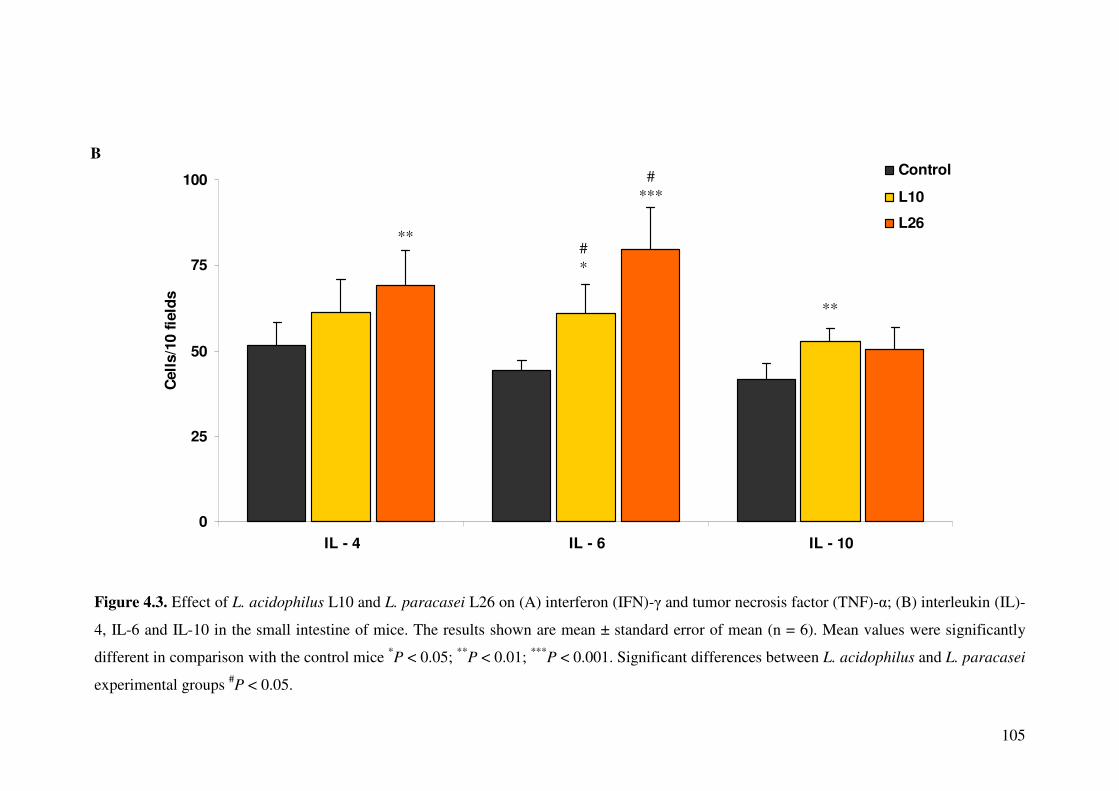
105
0
25
50
75
100
IL - 4 IL - 6 IL - 10
Cell
s/1
0 f
ield
sControl
L10
L26
Figure 4.3. Effect of L. acidophilus L10 and L. paracasei L26 on (A) interferon (IFN)-� and tumor necrosis factor (TNF)-�; (B) interleukin (IL)-
4, IL-6 and IL-10 in the small intestine of mice. The results shown are mean ± standard error of mean (n = 6). Mean values were significantly
different in comparison with the control mice *P < 0.05; **P < 0.01; ***
P < 0.001. Significant differences between L. acidophilus and L. paracasei
experimental groups #P < 0.05.
B
#
*
#
***
**
**
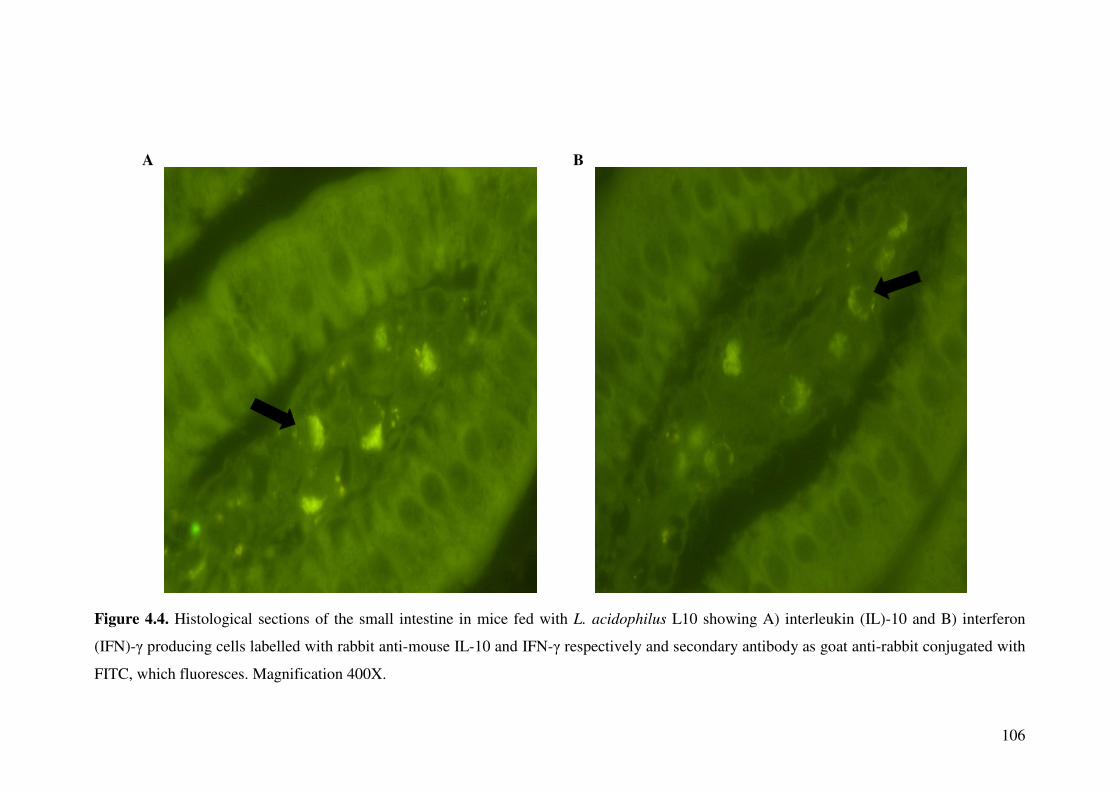
106
A B
Figure 4.4. Histological sections of the small intestine in mice fed with L. acidophilus L10 showing A) interleukin (IL)-10 and B) interferon
(IFN)-� producing cells labelled with rabbit anti-mouse IL-10 and IFN-� respectively and secondary antibody as goat anti-rabbit conjugated with
FITC, which fluoresces. Magnification 400X.

107
4.5. DISCUSSION
The NO in human body acts as a mediator in regulating various physiological and
pathophysiological processes (Moncada and Higgs, 1995). In physiological levels, NO
was essential in protecting intestinal mucosa through regulating mucus, bicarbonate
secretion and maintaining mucosal blood flow (Hutcheson et al., 1990; Alican and
Kubes, 1996; Lefer and Lefer, 1999). McCafferty et al. (1997) suggested that NO was
important in resolving gut inflammation and may also protect the GI tract in
inflammatory mechanisms. Several scientific reports indicated the involvement of NO in
protecting against intestinal mucosal permeability associated with reperfusion of
postischemic intestine (Payne and Kubes, 1993), preventing mucosal damage (Conforti
et al., 1993), and regulating GI tract protective mechanisms (Korhonen et al., 2001;
Lamine et al., 2004).
Elahi et al. (2005) correlated the increase of NO and IFN-� in saliva with enhanced
clearance of Candida albicans from oral cavities of L. acidophilus L10-fed mice.
Interestingly, results of the current study showed the capability of L. acidophilus and L.
paracasei in inducing NO levels in intestinal fluid and serum with increased IFN-�
producing cells in small intestine (Fig. 4.1; Fig. 4.3A). However, L. acidophilus-fed
mice showed no significant differences in increasing NO levels in intestinal fluid
compared to control mice. In relation to inducing NO levels in GI tract, L. paracasei
could be considered as a potential enhancer compared to L. acidophilus. Moreover, both
Lactobacillus strains showed significant enhancement of serum NO levels in systemic
immune response.

108
Gastrointestinal immune system consists of Peyer’s patches, mesenteric lymph nodes,
immunocompetent cells of lamina propria and mucosal epithelium. Lamina propria and
intraepithelial spaces are the main effector sites in the intestine. In lamina propria,
Peyer’s patch derived B-cells differentiate into dimeric IgA secreting plasma cells,
which were transported across epithelial cells and released as secretory IgA. The
secretory IgA is necessary in host defence mechanisms such as antigen binding and
preventing the adherence of pathogenic bacteria to intestinal epithelial cells (Hudault et
al., 1997; Gopal et al., 2001; Shu and Gill, 2002). In this study, L. acidophilus and L.
paracasei significantly enhanced CT-specific IgA response in intestinal fluid and serum
(Fig. 4.2). Intestinal lymphoid organ, Peyer’s patches were located in small intestine that
was considered as principal inductive sites of immune responses upon oral
administration of an antigen. In contrast to large intestine, small intestine is anatomically
connected to systemic immune system through lymphatic and blood circulation by
which the immune responses induced in small intestine could further lead to distant
mucosal sites in systemic immunity.
In the current study, increase of CT-specific IgA response was observed in intestinal
fluid of small intestine and blood serum of mice fed with L. acidophilus or L. paracasei,
which suggests that the release of CT-specific IgA antibodies in small intestine could
possibly lead into blood circulation. Therefore, blood serum could be regarded as a
potential marker to study the immunomodulation of orally administered antigens.
Furthermore, enhancement of CT-specific IgA response in intestinal fluid also suggests
the potential capability of L. acidophilus and L. paracasei strains, which could protect
the GI tract towards pathogenic microbial infections (Macpherson et al., 2001).

109
Goodrich and McGee, (1999) reported the ability of IL-6 in inducing terminal
differentiation of B-lymphocytes to IgA secreting cells. However, B-cells switching and
differentiation leading to plasmocyte secreting IgA also occurs in high IL-4, IL-5 and
transforming growth factor (TGF)-� environment (Blum et al., 1999). Despite the
capability of macrophages and T-cells to produce IL-6, intestinal epithelial cells are also
known to produce IL-6 (Vitini et al., 2000). In the present study, L. acidophilus and L.
paracasei increased IL-6 producing cells in small intestine (Fig. 4.3B). The
enhancement of IL-6 producing cells is considered to have a potential role in
augmenting the CT-specific IgA response in intestinal fluid and serum (Fig. 4.2). The
increase of IL-6 and IgA levels observed in this study is consistent with the earlier
studies that reported similar effects of lactic acid bacteria in enhancing IL-6 as well as
IgA responses (Miettinen et al., 1996; Galdeano and Perdigon, 2006).
Regulatory cytokines such as IL-4 and IL-10 are associated with humoral and allergic
immune responses. The IL-4 cytokine plays an important role in cell growth and
regulation of immune response particularly in inhibiting cytokines such as IL-1, IL-6,
IL-8 and TNF-� (Feghali and Wright, 1997). In the current study, differences in
stimulating IL-6 cytokine producing cells in mice fed with L. acidophilus or L.
paracasei were observed (Fig. 4.3B). The analysis of cytokine producing cells in small
intestine of mice showed the capability of L. acidophilus and L. paracasei strains in
modulating Th2 regulatory cytokines, such as IL-10 that plays an important role in
chronic inflammatory bowel disease (Braat et al., 2003) and amelioration of colitis (Di
Giacinto et al., 2005).

110
In the present study, pro-inflammatory cytokines such as IFN-� and TNF-� were
increased in mice fed with L. acidophilus or L. paracasei (Fig. 4.3A). In comparison to
control mice, L. acidophilus induced TNF-� cytokine producing cells were not
significantly different. Despite the role of IFN-� in inflammatory response, it can also
act as an effector molecule in immune responses against solid cancers (Numata et al.,
1991; Belardelli and Ferrantini, 2002). Macrophages and dendritic cells of lamina
propria are known to be crucial in producing various cytokines such as IL-6, IL-10, IL-
12, IFN-� and TNF-� (Husband et al., 1999). However, mast cells are also considered
necessary in releasing cytokines such as IL-4 and TNF-� that are important in initiating
immune and inflammatory responses (Feger et al., 2002). These various cell populations
from innate immune system might have been the source of cytokine producing cells in
lamina propria of small intestine that were determined in this study. Additionally,
current study also showed simultaneous stimulation of pro-inflammatory and regulatory
cytokines in mice fed with L. acidophilus or L. paracasei, which could be considered
necessary for maintaining a chronic and immunological balanced intestinal
inflammatory response termed as physiological inflammation (Cebra et al., 2005).

111
4.6. CONCLUSIONS
The current study demonstrates that L. acidophilus and L. paracasei were capable of
inducing NO levels in intestinal fluid and serum. Particularly, L. paracasei-fed mice
showed higher NO levels in intestinal fluid compared to mice fed with L. acidophilus.
Lactobacillus acidophilus and L. paracasei strains were demonstrated as potential
inducers of CT-specific IgA antibodies in gut and systemic immunity. Especially, L.
acidophilus induced higher CT-specific IgA levels in serum compared to L. paracasei-
fed mice. The analysis of cytokines in small intestine of mice reported the capability of
L. acidophilus and L. paracasei in regulating Th1 and Th2 cytokines. Particularly,
differences between L. acidophilus and L. paracasei were observed in inducing IL-6
cytokine producing cells in small intestine of mice.

112
CHAPTER 5
Microarray analysis of L. acidophilus LAFTI L10 induced gene expression in the
small intestine of mice

113
5.1. ABSTRACT
The microflora of gastrointestinal (GI) tract is known to be involved in promoting health
and physiological functions of the host. The influence of L. acidophilus in regulating
gene expression in the small intestine of normal and cholera toxin (CT) mice was
examined in this study. In the first experiment, mice were fed daily with L. acidophilus
at 108 CFU for 14-days, whereas in the second experiment, mice were fed with L.
acidophilus at 108 CFU/day for 14-days, during which they were immunized on day 0
and 7 with 10 �g of cholera toxin. In both the experiments, mice were euthanized after
14-day feeding trail and small intestines were excised to determine the gene expression
patterns using mouse oligonucleotide microarray. Microarray data was generated by
comparing the influence of L. acidophilus in regulating gene expression with the
respective control group mice. In the first experiment, normal mice fed with L.
acidophilus demonstrated the regulation of genes with an up-regulation of 27 and down-
regulation of 35 genes involved in various biological and molecular functions such as
cell-cell signalling, cell growth and proliferation, cell death and various metabolic
functions. In CT mice model, L. acidophilus influenced the up- and down-regulation of
28 and 32 genes that are involved in various cell functions, drug metabolism, immune
responses and metabolic functions. The present study demonstrated the influence of L.
acidophilus in regulating gene expression in the small bowel of normal and CT mice.
Furthermore, this study also provides a basis to identify the gene targets that may
represent a reference for further studies to define the pathways through which these
bacteria influence the host.

114
5.2. INTRODUCTION
Probiotic bacteria are considered as beneficial microbes due to their potential with
respect to different ways in which they can contribute to the positive health of GI tract.
Probiotic strains such as Lactobacillus and Bifidobacterium spp. are predominantly
incorporated into fermented dairy products. The interactions of probiotic bacteria in GI
tract are believed to be necessary in promoting host biological functions. Several reports
also suggested the probiotic effects on respiratory, subcutaneous and urogenital tract
functions (Tagg and Dierksen, 2003; Sheil et al., 2004; Reid, 2005). The microflora of
GI tract interacts with mucosal cell population, which includes epithelial cells (Hooper
et al., 2001). Commensal microbial population that colonises the GI tract acts as a
barrier in protecting the intestinal mucosa against pathogens through various
mechanisms, such as occupation of niches, antimicrobial production and competition for
essential nutrients.
The probiotic responses of microorganisms induced on host could be due to either
microbe-microbe or microbial-host interactions. Probiotic bacteria play an important
role in host defence mechanisms through activation of mucosal immune responses
(Shanahan, 2002a). Previous studies reported the capability of probiotic bacteria in
promoting gut barrier functions, balance of Th1/Th2 cytokines, enhancement of host
immune responses and interactions with gut associated lymphoid tissues (Erickson and
Hubbard, 2000; Isolauri et al., 2001). Furthermore, several studies also reported the
beneficial effects of probiotic bacteria in breast cancer model (de Moreno de LeBlanc et
al., 2005), respiratory infections (Racedo et al., 2006; Villena et al., 2006) and
inflammatory bowel diseases (Madsen et al., 2001; Schultz et al., 2002).

115
Probiotic bacteria especially Lactobacillus strains were characterised due to their
production of lactic acid, where most of them are indigenous inhabitants of human GI
tract and colonise in higher populations in the small intestine. Orally administered
probiotic bacteria are inducers of immune responses through potential inductive sites
such as M cells or follicle associated epithelial cells of Peyer’s patches on small
intestine. The immune responses induced in small intestine can effectively reach
systemic immune response due to its anatomical connection through lymphatic and
blood circulation, whereas immune responses induced in large intestine were mostly
limited to this environment. Vinderola et al. (2006) suggested the importance of small
intestine in understanding the immunomodulatory properties of orally administered kefir
microflora in mice.
The molecular interactions of gut microflora with host specifically in relation to
probiotic bacteria were not well understood. Microarray technology is a powerful tool,
which offers a comprehensive view of biological systems through monitoring the
expression of thousands of genes simultaneously and rapidly (Wu et al., 2001;
Sepulveda et al., 2002). Microarray analysis of the gene expression in host is necessary
to define these probiotic microorganisms as beneficial bacteria, which also provides
further insights into the interactions of these microbes with epithelial cells of GI tract.
In the current study, L. acidophilus L10 demonstrated its potential probiotic properties
such as tolerance to simulated gastric juices particularly at pH 2.0, better survival at
various bile salt concentrations and coaggregation abilities with other bacterial strains
(Chapter 2). Lactobacillus acidophilus L10 also showed its potential role in stimulating

116
various gut and systemic immune functions, especially non-specific and specific
immune responses in normal and cholera toxin (CT) mice (Chapters 3 & 4). Therefore,
L. acidophilus L10 was chosen to elucidate its influence on gene expression in the small
intestine of mice and also to provide further insights on genes that were involved in
regulating various biological and molecular functions of the host. This study was aimed
to investigate the effects of L. acidophilus L10 on gene expression patterns in the small
intestine of normal and CT mice.

117
5.3. MATERIALS AND METHODS
5.3.1. Animals
Male 6 - 8 wks old mice (BALB/c) were obtained from Biological Resources Centre,
The University of New South Wales, Sydney, Australia. Control and experimental
groups consisted of six mice each, which were caged separately at 22 ± 1°C and housed
under 12 h light-dark photoperiod. All mice were fed with standard mouse chow
(Gordon's Specialty Stock Feeds, Sydney, Australia) and provided ad libitum access to
water. All the protocols for the animal experiment were approved by the University of
Western Sydney Animal Care and Ethics Committee.
5.3.2. Lactic acid bacteria
Lactobacillus acidophilus LAFTI L10 used in this study was obtained from DSM
culture collection (DSM Food Specialties Ltd., Sydney, Australia). Lactobacillus
acidophilus was grown anaerobically in gas jars using GasPak System (Oxoid, Adelaide,
Australia) for 24 h at 37°C in de Man, Rogosa and Sharpe (MRS) broth (Oxoid,
Adelaide, Australia). The bacterial cells were harvested at 4000 x g for 15 min at 4°C
and washed twice with sterile 0.01 M phosphate buffered saline (PBS) solution.
5.3.3. Experimental design
Two experiments were conducted. In the first experiment, mice (n = 6) were fed daily by
oral gavage with 108 CFU of L. acidophilus in 50 �l of 10% (w/v) skimmed milk

118
powder (SMP) for 14-days. Control group mice received 50 �l of SMP without lactic
acid bacteria.
In the second experiment, mice (n = 6) were fed daily by oral gavage with 108 CFU of L.
acidophilus in 50 �l of 10% (w/v) SMP for 14-days. Control group mice received 50 �l
of SMP without lactic acid bacteria. Experimental and control group mice were orally
immunized with 10 �g of cholera toxin (Sigma-Aldrich, Sydney, Australia) in 25 �l of
0.1 M sodium bicarbonate (Sigma-Aldrich, Sydney, Australia) on day 0 and 7.
The viability of bacteria was determined by spread plate count of bacterial serial
dilutions on MRS agar (Oxoid, Adelaide, Australia), which were incubated
anaerobically in gas jars using GasPak System at 37°C for 48 h. In both the experiments,
mice were euthanized by carbon dioxide inhalation after 14-day feeding trail and small
intestines were removed for gene expression analysis.
5.3.4. RNA isolation
Total RNA was isolated from the small intestine of each mouse using TRIzol reagent
(Invitrogen, Melbourne, Australia) according to the manufacturer’s instructions. The
total RNA was further purified by RNeasy mini kit (Qiagen, Melbourne, Australia). The
purity and concentration of RNA was determined by spectrophotometer readings at 260
and 280 nm. The RNA integrity was confirmed by agarose gel electrophoresis. For
microarray analysis, equal amounts of RNA was pooled from all the mice (n = 6) of
experimental and control groups.

119
5.3.5. Microarray analysis
Microarray analysis was performed with RNA extracted from the experimental and
control groups using a compugen mouse oligonucleotide microarray consisting of
22,000 probes. A list of oligonucleotides immobilized on the array can be obtained from
the web-site: http://www.microarray.adelaide.edu.au/libraries.
5.3.6. Synthesis of cRNA probes
The total RNA pooled from individual experimental and control group mice was
amplified using a SuperScript Indirect RNA Amplification System (Invitrogen,
Melbourne, Australia) according to the manufacturer’s instructions. The purified cRNA
was dried under reduced pressure, dissolved in 9 µl of 0.1 M NaHCO3 (pH 9.0), mixed
with Cy3 or Cy5 and left in the dark to couple at room temperature for 60 min. The
labelled cRNA was mixed with 41 µl of Milli Q water and purified using a QIAquick
PCR purification kit (Qiagen, Melbourne, Australia). The purified fluorescent cRNA
samples were eluted into a clean tube with 90 µl of Milli Q water and dried under
reduced pressure.
5.3.7. Hybridisation of probes to oligonucleotide microarray
Prior to hybridization, each microarray slide was immersed in 50 ml of hot (60-95˚C)
Milli-Q water for 1 min, then each array was dried by centrifugation at 650 x g for 5
min. The labelled cRNA probes were then mixed with 0.64 µl of 25 mg/ml yeast tRNA,
4 µl of 2 mg/ml poly A and 20 µl of 1 mg/ml mouse Cot-1 DNA, dried under reduced

120
pressure, resuspended in 16 µl of formamide and 16 µl of 6.25 X SSC, denatured by
heating to 100˚C for 3 min and transferred directly to ice, followed by addition of 0.5 µl
of 10% SDS. The probes were then applied to the array and incubated at 42˚C overnight
in a humidified chamber. The arrays were washed in 0.5 X SSC containing 0.01% SDS
for 5 min, 0.5 X SSC for 3 min and rinsed in 0.2 X SSC for 3 min. The slides were dried
by centrifugation at 650 x g for 5 min and were then scanned using an Axon 4000B
microarray scanner. Data analysis was performed by using the single image tif files.
5.3.8. Data Analysis
The Cy5 and Cy3 fluorescent signal intensity of each gene on the array was extracted
using SPOT software (CSIRO, Australia). After the subtraction of morphological
background fluorescence, ratio of signal intensities (Cy5/Cy3) was calculated. Each
probe ratio was log 2 transformed and normalised using a LOWESS algorithm (Locally
weighted scatterplot smoothing) within each slide using LIMMA (Linear models for
microarray analysis). Between the arrays, normalisation was performed to have the log-
ratios with same median-absolute-deviation (MAD) across all arrays. Linear modelling
combined with Bayesian statistical analysis of the normalised data produced a ranked
list of genes that were likely to be consistently differentially expressed on all 4 arrays.

121
5.4. RESULTS
In both the experiments, microarray analysis of gene expression in the small bowel of
mice was analysed using oligonucleotide microarray consisting of 22,000 genes.
Analysis of data from arrays ranked the genes in order of likelihood of being
differentially expressed. In both the experiments, top 100 genes that were likely to be
differentially expressed were selected and their functional and pathway analysis were
determined through Ingenuity Pathways Analysis (Ingenuity systems,
www.ingenuity.com).
In the first experiment, 62 of the top 100 genes were mapped to Ingenuity Pathways
Knowledge Base associated with known biological functions. In 62 genes, 27 and 35
genes were up- and down-regulated (Table 5.1). Orally administered L. acidophilus in
normal mice altered the expression of genes that are involved in various biological and
molecular functions such as cell-cell signalling, cell growth, molecular transport, tissue
development, immune response and various metabolic functions (Table 5.2; 5.3). Genes
such as MAP4K1, H3 histone family 3A and HSPA1A, which are involved in p38
MAPK signalling, TGF-� signalling and ERF/MAPK signalling pathways were up-
regulated in L. acidophilus-fed mice (Table 5.4).
Genes related to cytochrome P450 family (CYP2B6, CYP2C8, CYP2C19) that are
involved in linoleic acid, fatty acid and arachidonic acid metabolic pathways were
down-regulated by L. acidophilus. Several other genes such as fibroblast growth factor
19 (FGF signalling), carboxyl ester lipase (Bile acid metabolism), AMY2A and ENPP3

122
(Starch and sucrose metabolism) and glutathione S-transferase A5 (Glutathione
metabolism) were also down-regulated in L. acidophilus-fed mice (Table 5.4).
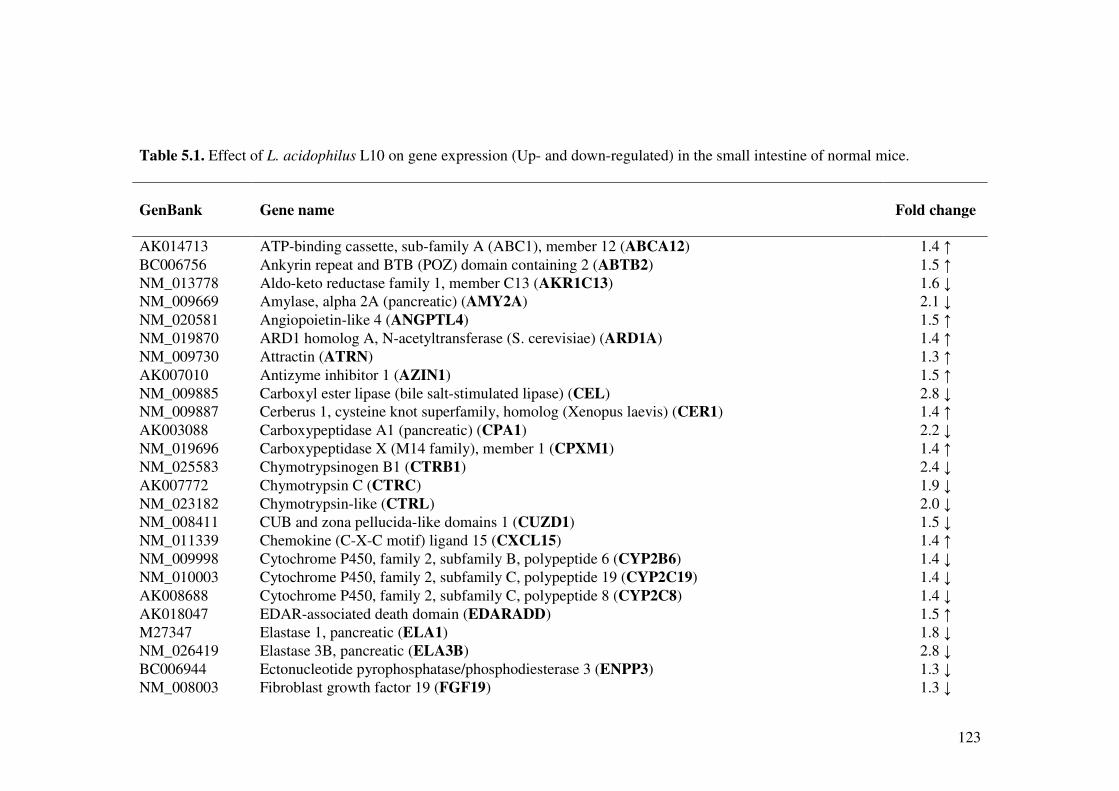
123
Table 5.1. Effect of L. acidophilus L10 on gene expression (Up- and down-regulated) in the small intestine of normal mice.
GenBank Gene name
Fold change
AK014713 ATP-binding cassette, sub-family A (ABC1), member 12 (ABCA12) 1.4 �
BC006756 Ankyrin repeat and BTB (POZ) domain containing 2 (ABTB2) 1.5 �
NM_013778 Aldo-keto reductase family 1, member C13 (AKR1C13) 1.6 �
NM_009669 Amylase, alpha 2A (pancreatic) (AMY2A) 2.1 �
NM_020581 Angiopoietin-like 4 (ANGPTL4) 1.5 �
NM_019870 ARD1 homolog A, N-acetyltransferase (S. cerevisiae) (ARD1A) 1.4 �
NM_009730 Attractin (ATRN) 1.3 �
AK007010 Antizyme inhibitor 1 (AZIN1) 1.5 �
NM_009885 Carboxyl ester lipase (bile salt-stimulated lipase) (CEL) 2.8 �
NM_009887 Cerberus 1, cysteine knot superfamily, homolog (Xenopus laevis) (CER1) 1.4 �
AK003088 Carboxypeptidase A1 (pancreatic) (CPA1) 2.2 �
NM_019696 Carboxypeptidase X (M14 family), member 1 (CPXM1) 1.4 �
NM_025583 Chymotrypsinogen B1 (CTRB1) 2.4 �
AK007772 Chymotrypsin C (CTRC) 1.9 �
NM_023182 Chymotrypsin-like (CTRL) 2.0 �
NM_008411 CUB and zona pellucida-like domains 1 (CUZD1) 1.5 �
NM_011339 Chemokine (C-X-C motif) ligand 15 (CXCL15) 1.4 �
NM_009998 Cytochrome P450, family 2, subfamily B, polypeptide 6 (CYP2B6) 1.4 �
NM_010003 Cytochrome P450, family 2, subfamily C, polypeptide 19 (CYP2C19) 1.4 �
AK008688 Cytochrome P450, family 2, subfamily C, polypeptide 8 (CYP2C8) 1.4 �
AK018047 EDAR-associated death domain (EDARADD) 1.5 �
M27347 Elastase 1, pancreatic (ELA1) 1.8 �
NM_026419 Elastase 3B, pancreatic (ELA3B) 2.8 �
BC006944 Ectonucleotide pyrophosphatase/phosphodiesterase 3 (ENPP3) 1.3 �
NM_008003 Fibroblast growth factor 19 (FGF19) 1.3 �
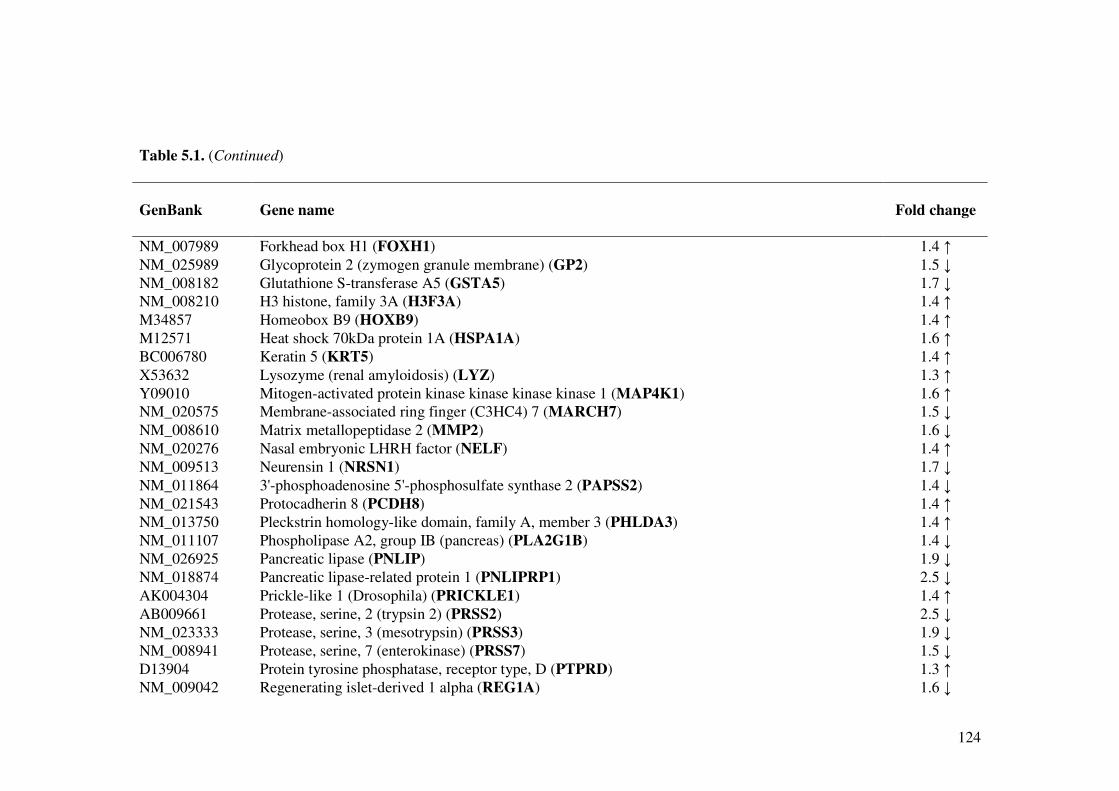
124
Table 5.1. (Continued)
GenBank Gene name
Fold change
NM_007989 Forkhead box H1 (FOXH1) 1.4 �
NM_025989 Glycoprotein 2 (zymogen granule membrane) (GP2) 1.5 �
NM_008182 Glutathione S-transferase A5 (GSTA5) 1.7 �
NM_008210 H3 histone, family 3A (H3F3A) 1.4 �
M34857 Homeobox B9 (HOXB9) 1.4 �
M12571 Heat shock 70kDa protein 1A (HSPA1A) 1.6 �
BC006780 Keratin 5 (KRT5) 1.4 �
X53632 Lysozyme (renal amyloidosis) (LYZ) 1.3 �
Y09010 Mitogen-activated protein kinase kinase kinase kinase 1 (MAP4K1) 1.6 �
NM_020575 Membrane-associated ring finger (C3HC4) 7 (MARCH7) 1.5 �
NM_008610 Matrix metallopeptidase 2 (MMP2) 1.6 �
NM_020276 Nasal embryonic LHRH factor (NELF) 1.4 �
NM_009513 Neurensin 1 (NRSN1) 1.7 �
NM_011864 3'-phosphoadenosine 5'-phosphosulfate synthase 2 (PAPSS2) 1.4 �
NM_021543 Protocadherin 8 (PCDH8) 1.4 �
NM_013750 Pleckstrin homology-like domain, family A, member 3 (PHLDA3) 1.4 �
NM_011107 Phospholipase A2, group IB (pancreas) (PLA2G1B) 1.4 �
NM_026925 Pancreatic lipase (PNLIP) 1.9 �
NM_018874 Pancreatic lipase-related protein 1 (PNLIPRP1) 2.5 �
AK004304 Prickle-like 1 (Drosophila) (PRICKLE1) 1.4 �
AB009661 Protease, serine, 2 (trypsin 2) (PRSS2) 2.5 �
NM_023333 Protease, serine, 3 (mesotrypsin) (PRSS3) 1.9 �
NM_008941 Protease, serine, 7 (enterokinase) (PRSS7) 1.5 �
D13904 Protein tyrosine phosphatase, receptor type, D (PTPRD) 1.3 �
NM_009042 Regenerating islet-derived 1 alpha (REG1A) 1.6 �
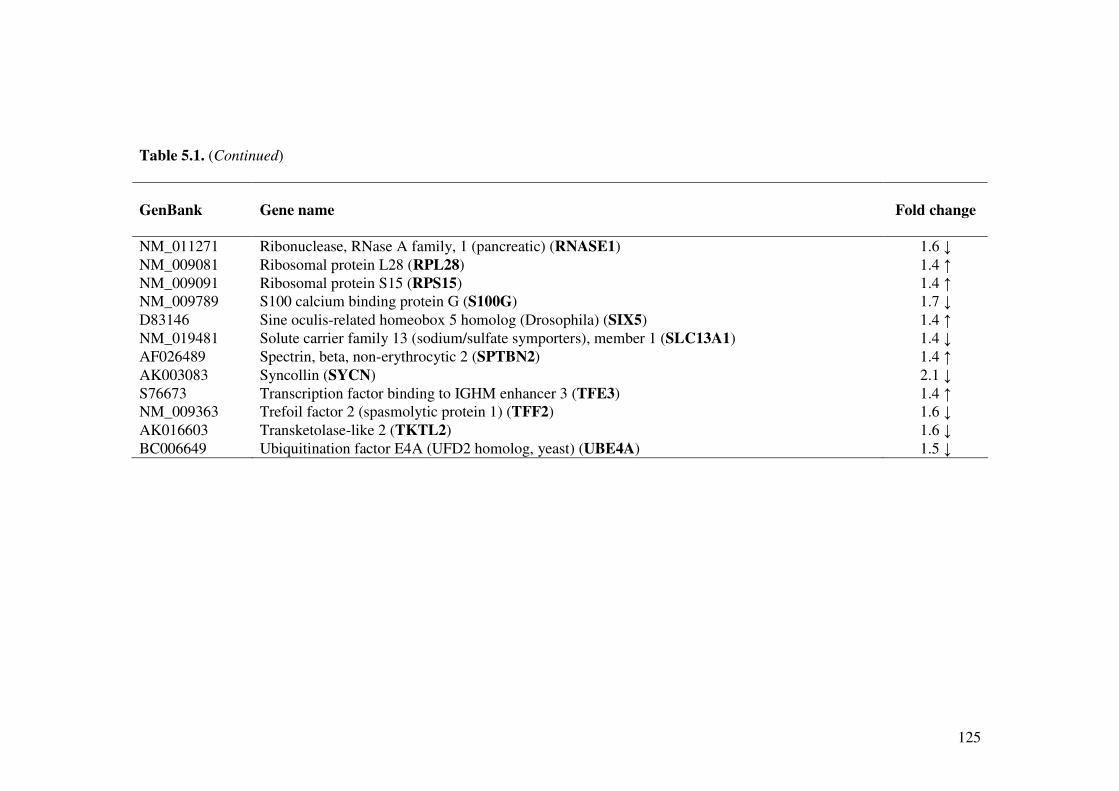
125
Table 5.1. (Continued)
GenBank Gene name
Fold change
NM_011271 Ribonuclease, RNase A family, 1 (pancreatic) (RNASE1) 1.6 �
NM_009081 Ribosomal protein L28 (RPL28) 1.4 �
NM_009091 Ribosomal protein S15 (RPS15) 1.4 �
NM_009789 S100 calcium binding protein G (S100G) 1.7 �
D83146 Sine oculis-related homeobox 5 homolog (Drosophila) (SIX5) 1.4 �
NM_019481 Solute carrier family 13 (sodium/sulfate symporters), member 1 (SLC13A1) 1.4 �
AF026489 Spectrin, beta, non-erythrocytic 2 (SPTBN2) 1.4 �
AK003083 Syncollin (SYCN) 2.1 �
S76673 Transcription factor binding to IGHM enhancer 3 (TFE3) 1.4 �
NM_009363 Trefoil factor 2 (spasmolytic protein 1) (TFF2) 1.6 �
AK016603 Transketolase-like 2 (TKTL2) 1.6 �
BC006649 Ubiquitination factor E4A (UFD2 homolog, yeast) (UBE4A) 1.5 �
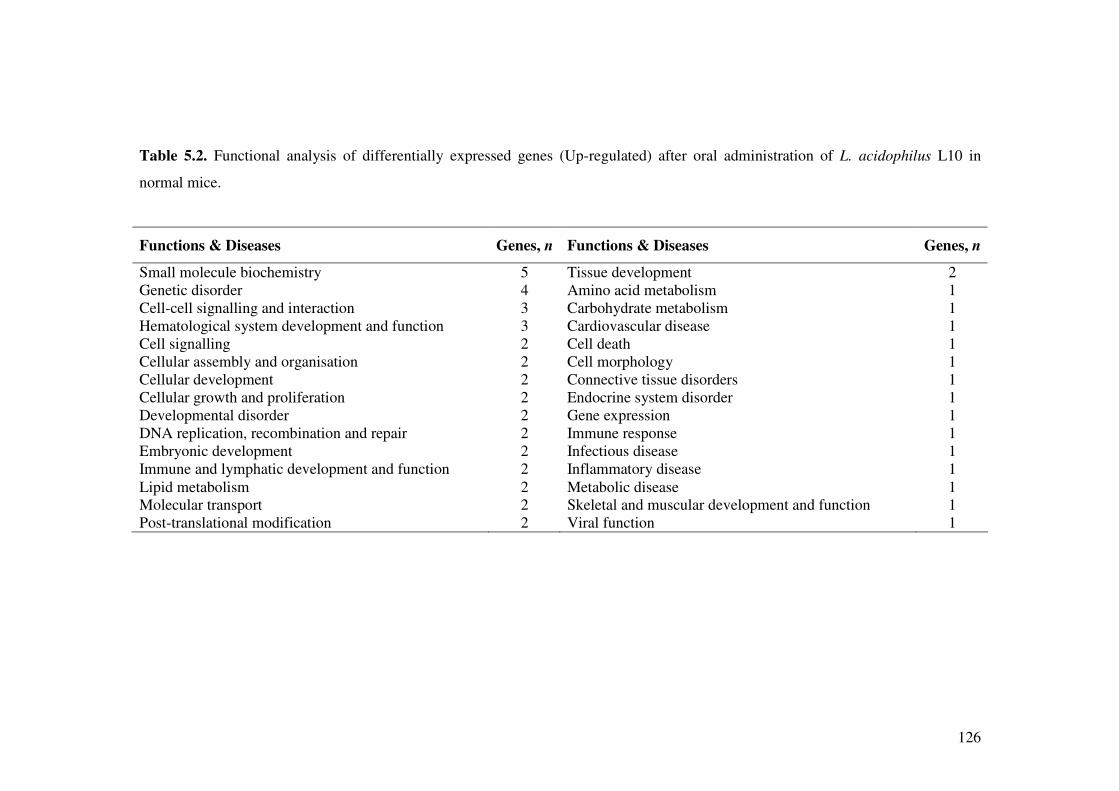
126
Table 5.2. Functional analysis of differentially expressed genes (Up-regulated) after oral administration of L. acidophilus L10 in
normal mice.
Functions & Diseases Genes, n Functions & Diseases Genes, n
Small molecule biochemistry 5 Tissue development 2 Genetic disorder 4 Amino acid metabolism 1 Cell-cell signalling and interaction 3 Carbohydrate metabolism 1 Hematological system development and function 3 Cardiovascular disease 1 Cell signalling 2 Cell death 1 Cellular assembly and organisation 2 Cell morphology 1 Cellular development 2 Connective tissue disorders 1 Cellular growth and proliferation 2 Endocrine system disorder 1 Developmental disorder 2 Gene expression 1 DNA replication, recombination and repair 2 Immune response 1 Embryonic development 2 Infectious disease 1 Immune and lymphatic development and function 2 Inflammatory disease 1
Lipid metabolism 2 Metabolic disease 1 Molecular transport 2 Skeletal and muscular development and function 1 Post-translational modification 2 Viral function 1
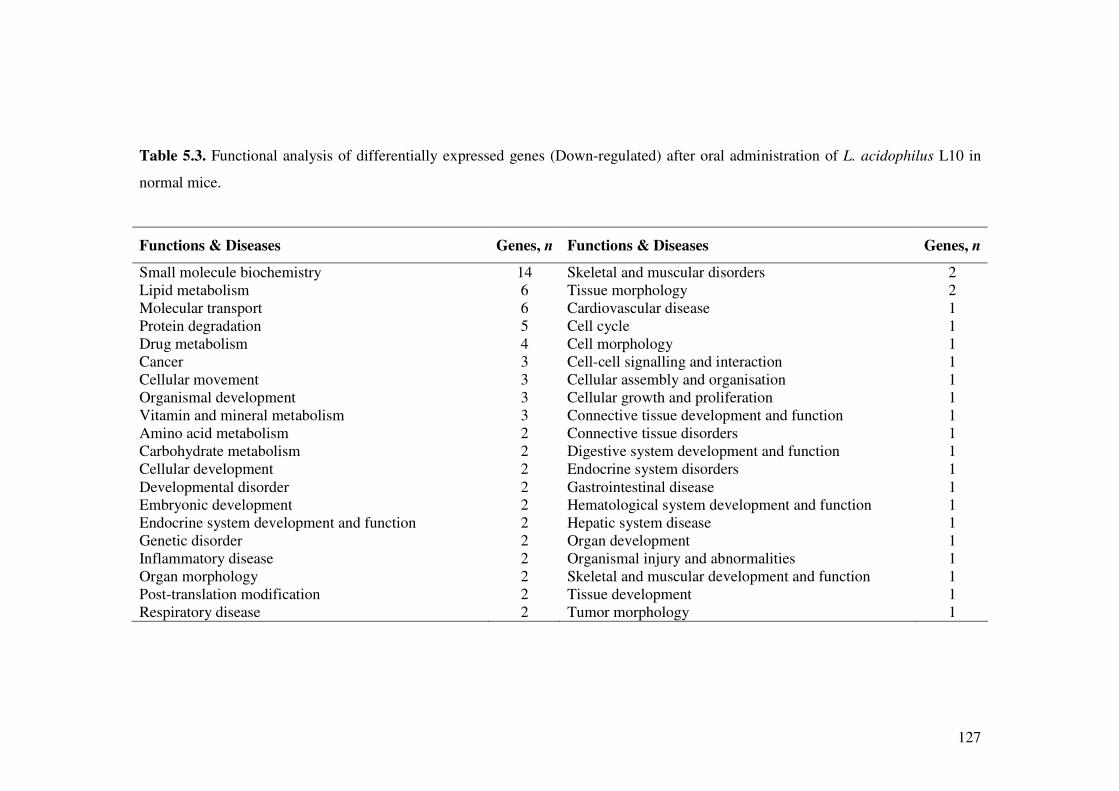
127
Table 5.3. Functional analysis of differentially expressed genes (Down-regulated) after oral administration of L. acidophilus L10 in
normal mice.
Functions & Diseases Genes, n Functions & Diseases Genes, n
Small molecule biochemistry 14 Skeletal and muscular disorders 2 Lipid metabolism 6 Tissue morphology 2 Molecular transport 6 Cardiovascular disease 1 Protein degradation 5 Cell cycle 1 Drug metabolism 4 Cell morphology 1 Cancer 3 Cell-cell signalling and interaction 1 Cellular movement 3 Cellular assembly and organisation 1 Organismal development 3 Cellular growth and proliferation 1 Vitamin and mineral metabolism 3 Connective tissue development and function 1 Amino acid metabolism 2 Connective tissue disorders 1 Carbohydrate metabolism 2 Digestive system development and function 1 Cellular development 2 Endocrine system disorders 1
Developmental disorder 2 Gastrointestinal disease 1 Embryonic development 2 Hematological system development and function 1 Endocrine system development and function 2 Hepatic system disease 1 Genetic disorder 2 Organ development 1 Inflammatory disease 2 Organismal injury and abnormalities 1 Organ morphology 2 Skeletal and muscular development and function 1 Post-translation modification 2 Tissue development 1 Respiratory disease 2 Tumor morphology 1
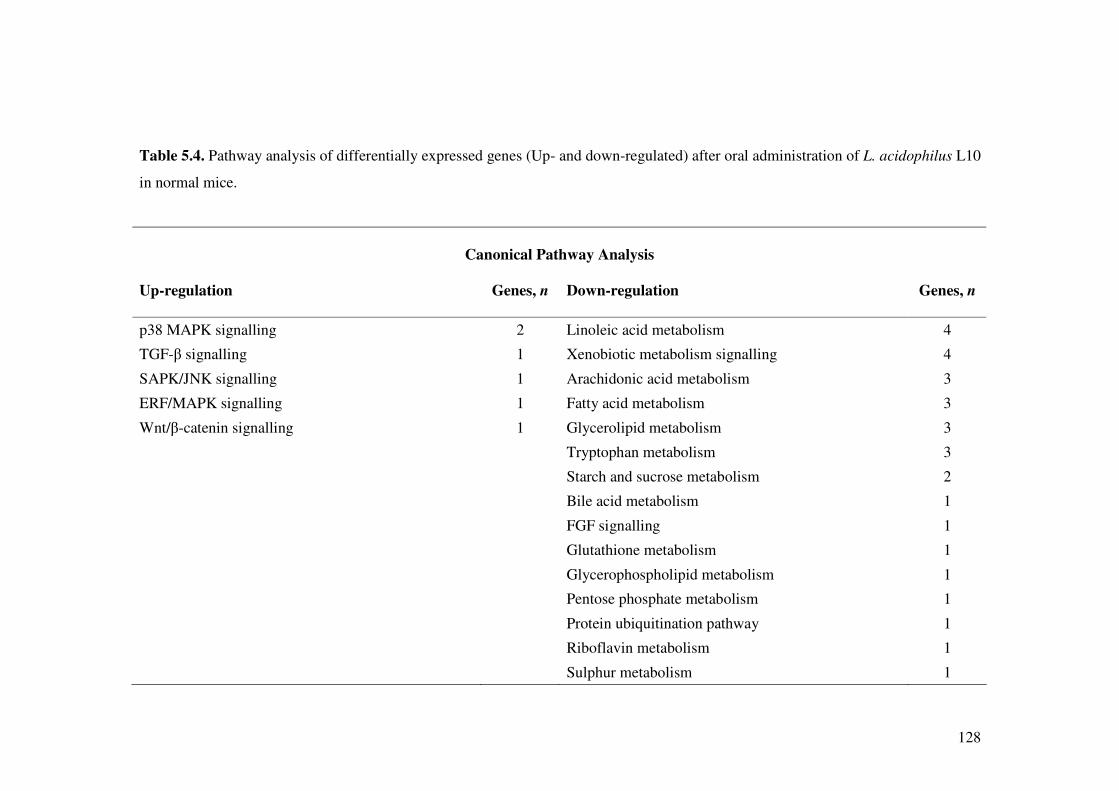
128
Table 5.4. Pathway analysis of differentially expressed genes (Up- and down-regulated) after oral administration of L. acidophilus L10
in normal mice.
Canonical Pathway Analysis
Up-regulation
Genes, n Down-regulation Genes, n
p38 MAPK signalling 2 Linoleic acid metabolism 4
TGF-� signalling 1 Xenobiotic metabolism signalling 4
SAPK/JNK signalling 1 Arachidonic acid metabolism 3
ERF/MAPK signalling 1 Fatty acid metabolism 3
Wnt/�-catenin signalling 1 Glycerolipid metabolism 3
Tryptophan metabolism 3
Starch and sucrose metabolism 2
Bile acid metabolism 1
FGF signalling 1
Glutathione metabolism 1
Glycerophospholipid metabolism 1
Pentose phosphate metabolism 1
Protein ubiquitination pathway 1
Riboflavin metabolism 1
Sulphur metabolism 1

129
In the second experiment, regulation of gene expression by L. acidophilus in the small
intestine of CT mice model was examined. Sixty of the top 100 genes were mapped to
the Ingenuity Pathways Knowledge Base associated with known biological functions. In
60 genes, 28 and 32 genes were up- and down-regulated (Table 5.5). Lactobacillus
acidophilus influenced the expression of genes that are involved in various cell functions
such as cell death, cellular movement, cell cycle, cellular growth and proliferation.
Several other genes regulated are also involved in functions related to molecular
transport, immune responses, immunological and inflammatory diseases, gastrointestinal
diseases and various metabolic functions (Table 5.6; 5.7).
Genes related to cytochrome P450 family (CYP2C9, CYP2C19, CYP2C38, CYP3A5)
that are involved in various metabolic pathways such as arachidonic acid metabolism,
fatty acid metabolism, linoleic acid metabolism and tryptophan metabolism were up-
regulated in the small bowel of L. acidophilus-fed mice (Table 5.8). Additionally, genes
involved in oxidative phosphorylation such as COX7C, NDUFC1, NDUFC2, PPA1 and
UCRC were up-regulated in mice fed with L. acidophilus. Fibronectin 1, which is
involved in integrin and actin cytoskeleton signalling pathways was also up-regulated in
mice fed with L. acidophilus.
Orally administered L. acidophilus down-regulated genes such as BIRC3, TNFRSF1A,
CD74 that are involved in apoptosis, death receptor signalling, antigen presenting
pathway, IL-6 and p38 MAPK signalling pathways. Furthermore, L. acidophilus also
down-regulated several genes that are involved in various functions related to eicosanoid
signalling (DPEP1), glutathione metabolism (ANPEP), N-Glycan biosynthesis

130
(MAN1B1), starch and sucrose metabolism (SI), cAMP-mediated signalling and G-
protein coupled receptor signalling pathways (ADORA1) (Table 5.8).
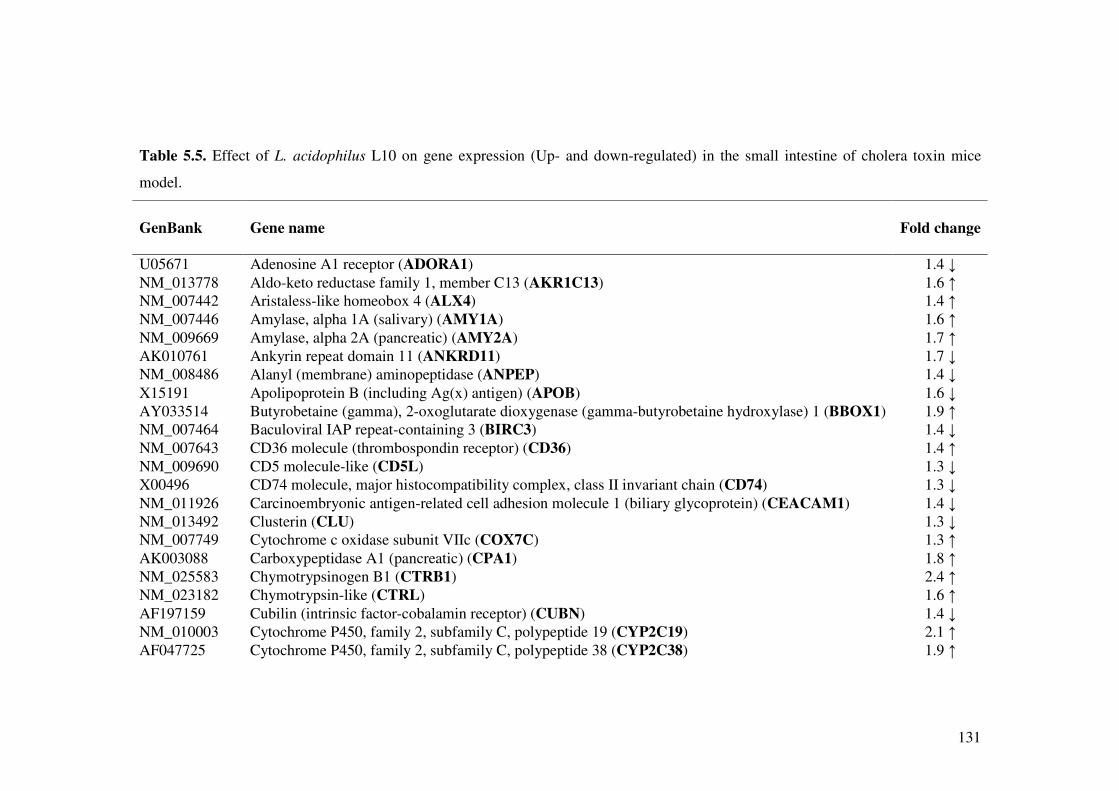
131
Table 5.5. Effect of L. acidophilus L10 on gene expression (Up- and down-regulated) in the small intestine of cholera toxin mice
model.
GenBank Gene name
Fold change
U05671 Adenosine A1 receptor (ADORA1) 1.4 �
NM_013778 Aldo-keto reductase family 1, member C13 (AKR1C13) 1.6 �
NM_007442 Aristaless-like homeobox 4 (ALX4) 1.4 �
NM_007446 Amylase, alpha 1A (salivary) (AMY1A) 1.6 �
NM_009669 Amylase, alpha 2A (pancreatic) (AMY2A) 1.7 �
AK010761 Ankyrin repeat domain 11 (ANKRD11) 1.7 �
NM_008486 Alanyl (membrane) aminopeptidase (ANPEP) 1.4 �
X15191 Apolipoprotein B (including Ag(x) antigen) (APOB) 1.6 �
AY033514 Butyrobetaine (gamma), 2-oxoglutarate dioxygenase (gamma-butyrobetaine hydroxylase) 1 (BBOX1) 1.9 �
NM_007464 Baculoviral IAP repeat-containing 3 (BIRC3) 1.4 �
NM_007643 CD36 molecule (thrombospondin receptor) (CD36) 1.4 �
NM_009690 CD5 molecule-like (CD5L) 1.3 �
X00496 CD74 molecule, major histocompatibility complex, class II invariant chain (CD74) 1.3 �
NM_011926 Carcinoembryonic antigen-related cell adhesion molecule 1 (biliary glycoprotein) (CEACAM1) 1.4 �
NM_013492 Clusterin (CLU) 1.3 �
NM_007749 Cytochrome c oxidase subunit VIIc (COX7C) 1.3 �
AK003088 Carboxypeptidase A1 (pancreatic) (CPA1) 1.8 �
NM_025583 Chymotrypsinogen B1 (CTRB1) 2.4 �
NM_023182 Chymotrypsin-like (CTRL) 1.6 �
AF197159 Cubilin (intrinsic factor-cobalamin receptor) (CUBN) 1.4 �
NM_010003 Cytochrome P450, family 2, subfamily C, polypeptide 19 (CYP2C19) 2.1 �
AF047725 Cytochrome P450, family 2, subfamily C, polypeptide 38 (CYP2C38) 1.9 �
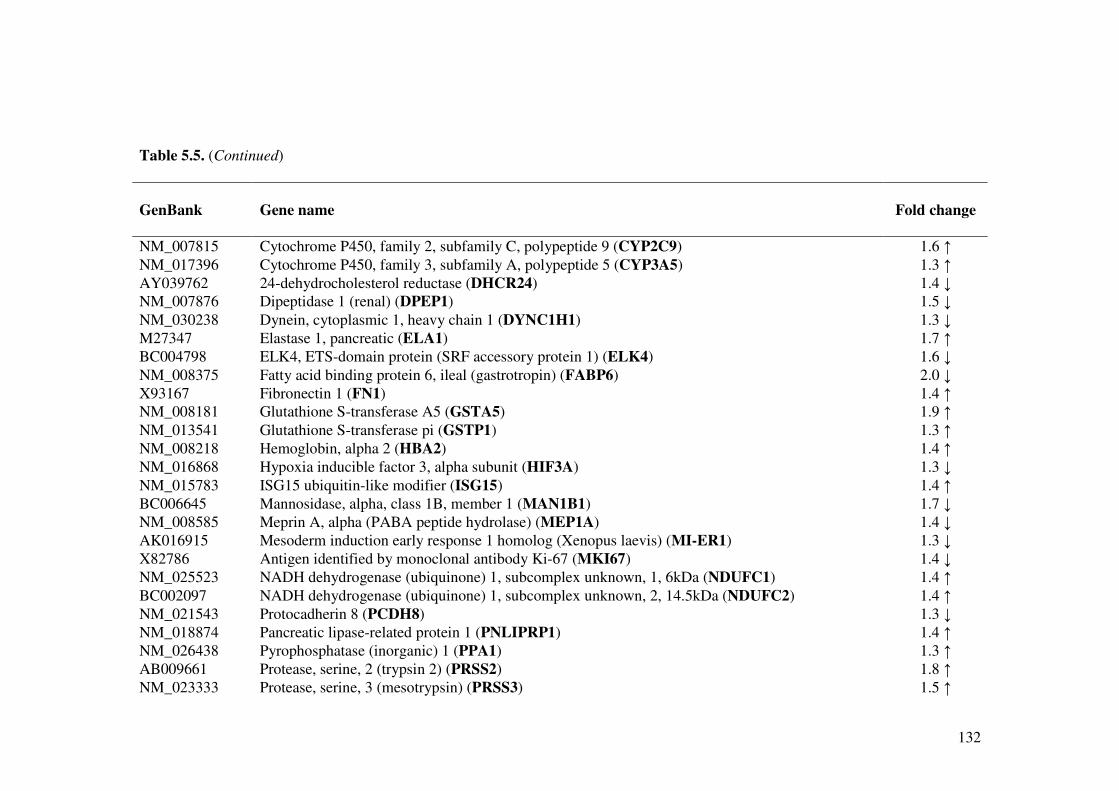
132
Table 5.5. (Continued)
GenBank Gene name
Fold change
NM_007815 Cytochrome P450, family 2, subfamily C, polypeptide 9 (CYP2C9) 1.6 �
NM_017396 Cytochrome P450, family 3, subfamily A, polypeptide 5 (CYP3A5) 1.3 �
AY039762 24-dehydrocholesterol reductase (DHCR24) 1.4 �
NM_007876 Dipeptidase 1 (renal) (DPEP1) 1.5 �
NM_030238 Dynein, cytoplasmic 1, heavy chain 1 (DYNC1H1) 1.3 �
M27347 Elastase 1, pancreatic (ELA1) 1.7 �
BC004798 ELK4, ETS-domain protein (SRF accessory protein 1) (ELK4) 1.6 �
NM_008375 Fatty acid binding protein 6, ileal (gastrotropin) (FABP6) 2.0 �
X93167 Fibronectin 1 (FN1) 1.4 �
NM_008181 Glutathione S-transferase A5 (GSTA5) 1.9 �
NM_013541 Glutathione S-transferase pi (GSTP1) 1.3 �
NM_008218 Hemoglobin, alpha 2 (HBA2) 1.4 �
NM_016868 Hypoxia inducible factor 3, alpha subunit (HIF3A) 1.3 �
NM_015783 ISG15 ubiquitin-like modifier (ISG15) 1.4 �
BC006645 Mannosidase, alpha, class 1B, member 1 (MAN1B1) 1.7 �
NM_008585 Meprin A, alpha (PABA peptide hydrolase) (MEP1A) 1.4 �
AK016915 Mesoderm induction early response 1 homolog (Xenopus laevis) (MI-ER1) 1.3 �
X82786 Antigen identified by monoclonal antibody Ki-67 (MKI67) 1.4 �
NM_025523 NADH dehydrogenase (ubiquinone) 1, subcomplex unknown, 1, 6kDa (NDUFC1) 1.4 �
BC002097 NADH dehydrogenase (ubiquinone) 1, subcomplex unknown, 2, 14.5kDa (NDUFC2) 1.4 �
NM_021543 Protocadherin 8 (PCDH8) 1.3 �
NM_018874 Pancreatic lipase-related protein 1 (PNLIPRP1) 1.4 �
NM_026438 Pyrophosphatase (inorganic) 1 (PPA1) 1.3 �
AB009661 Protease, serine, 2 (trypsin 2) (PRSS2) 1.8 �
NM_023333 Protease, serine, 3 (mesotrypsin) (PRSS3) 1.5 �
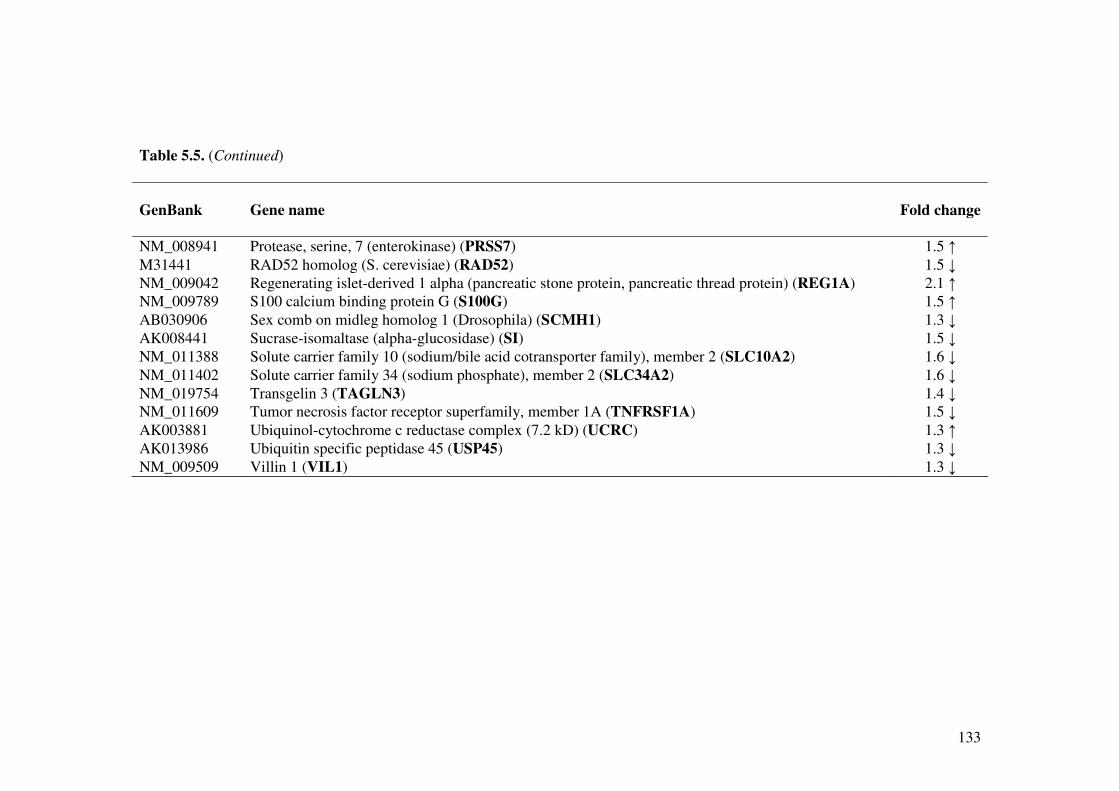
133
Table 5.5. (Continued)
GenBank Gene name
Fold change
NM_008941 Protease, serine, 7 (enterokinase) (PRSS7) 1.5 �
M31441 RAD52 homolog (S. cerevisiae) (RAD52) 1.5 �
NM_009042 Regenerating islet-derived 1 alpha (pancreatic stone protein, pancreatic thread protein) (REG1A) 2.1 �
NM_009789 S100 calcium binding protein G (S100G) 1.5 �
AB030906 Sex comb on midleg homolog 1 (Drosophila) (SCMH1) 1.3 �
AK008441 Sucrase-isomaltase (alpha-glucosidase) (SI) 1.5 �
NM_011388 Solute carrier family 10 (sodium/bile acid cotransporter family), member 2 (SLC10A2) 1.6 �
NM_011402 Solute carrier family 34 (sodium phosphate), member 2 (SLC34A2) 1.6 �
NM_019754 Transgelin 3 (TAGLN3) 1.4 �
NM_011609 Tumor necrosis factor receptor superfamily, member 1A (TNFRSF1A) 1.5 �
AK003881 Ubiquinol-cytochrome c reductase complex (7.2 kD) (UCRC) 1.3 �
AK013986 Ubiquitin specific peptidase 45 (USP45) 1.3 �
NM_009509 Villin 1 (VIL1) 1.3 �
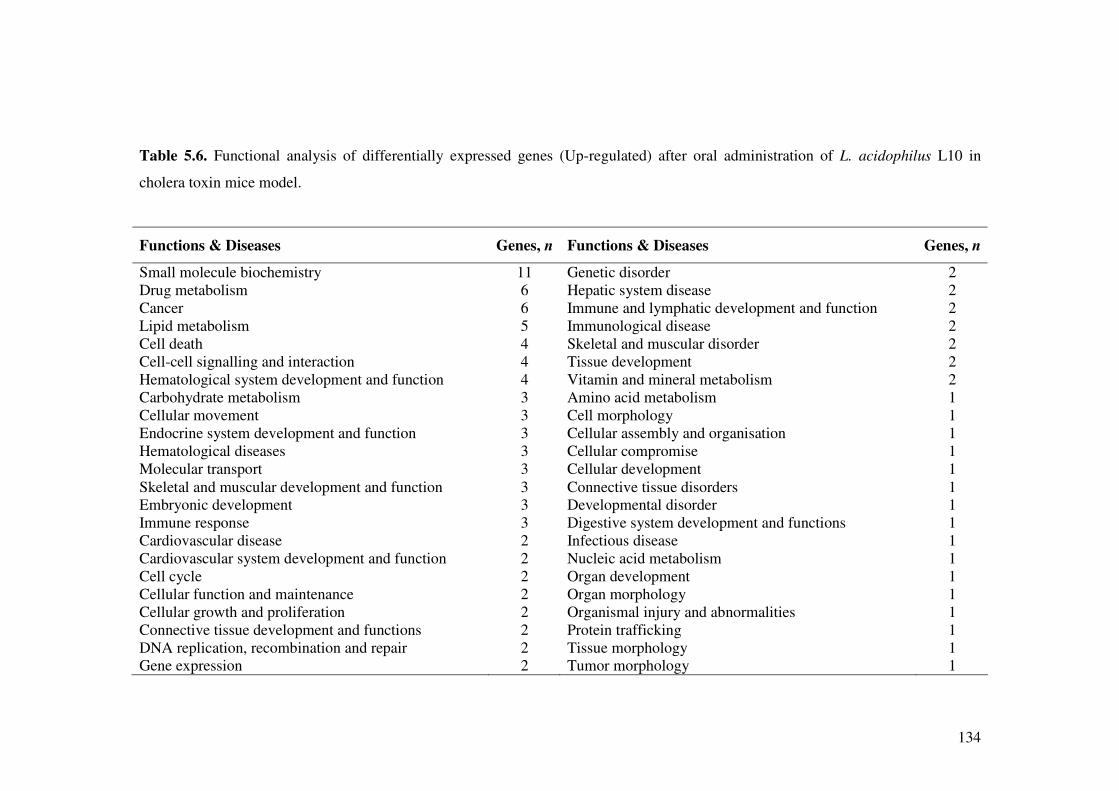
134
Table 5.6. Functional analysis of differentially expressed genes (Up-regulated) after oral administration of L. acidophilus L10 in
cholera toxin mice model.
Functions & Diseases Genes, n Functions & Diseases Genes, n
Small molecule biochemistry 11 Genetic disorder 2 Drug metabolism 6 Hepatic system disease 2 Cancer 6 Immune and lymphatic development and function 2 Lipid metabolism 5 Immunological disease 2 Cell death 4 Skeletal and muscular disorder 2 Cell-cell signalling and interaction 4 Tissue development 2 Hematological system development and function 4 Vitamin and mineral metabolism 2 Carbohydrate metabolism 3 Amino acid metabolism 1 Cellular movement 3 Cell morphology 1 Endocrine system development and function 3 Cellular assembly and organisation 1 Hematological diseases 3 Cellular compromise 1 Molecular transport 3 Cellular development 1
Skeletal and muscular development and function 3 Connective tissue disorders 1 Embryonic development 3 Developmental disorder 1 Immune response 3 Digestive system development and functions 1 Cardiovascular disease 2 Infectious disease 1 Cardiovascular system development and function 2 Nucleic acid metabolism 1 Cell cycle 2 Organ development 1 Cellular function and maintenance 2 Organ morphology 1 Cellular growth and proliferation 2 Organismal injury and abnormalities 1 Connective tissue development and functions 2 Protein trafficking 1 DNA replication, recombination and repair 2 Tissue morphology 1 Gene expression 2 Tumor morphology 1
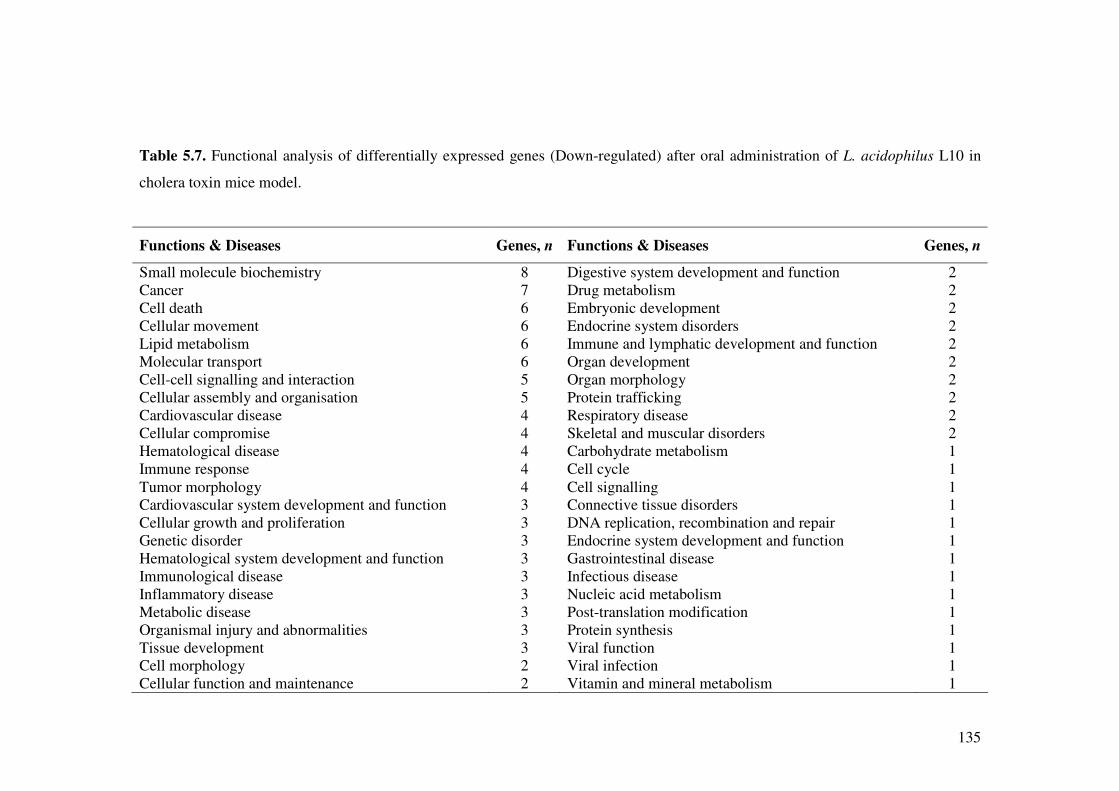
135
Table 5.7. Functional analysis of differentially expressed genes (Down-regulated) after oral administration of L. acidophilus L10 in
cholera toxin mice model.
Functions & Diseases Genes, n Functions & Diseases Genes, n
Small molecule biochemistry 8 Digestive system development and function 2 Cancer 7 Drug metabolism 2 Cell death 6 Embryonic development 2 Cellular movement 6 Endocrine system disorders 2 Lipid metabolism 6 Immune and lymphatic development and function 2 Molecular transport 6 Organ development 2 Cell-cell signalling and interaction 5 Organ morphology 2 Cellular assembly and organisation 5 Protein trafficking 2 Cardiovascular disease 4 Respiratory disease 2 Cellular compromise 4 Skeletal and muscular disorders 2 Hematological disease 4 Carbohydrate metabolism 1 Immune response 4 Cell cycle 1
Tumor morphology 4 Cell signalling 1 Cardiovascular system development and function 3 Connective tissue disorders 1 Cellular growth and proliferation 3 DNA replication, recombination and repair 1 Genetic disorder 3 Endocrine system development and function 1 Hematological system development and function 3 Gastrointestinal disease 1 Immunological disease 3 Infectious disease 1 Inflammatory disease 3 Nucleic acid metabolism 1 Metabolic disease 3 Post-translation modification 1 Organismal injury and abnormalities 3 Protein synthesis 1 Tissue development 3 Viral function 1 Cell morphology 2 Viral infection 1 Cellular function and maintenance 2 Vitamin and mineral metabolism 1
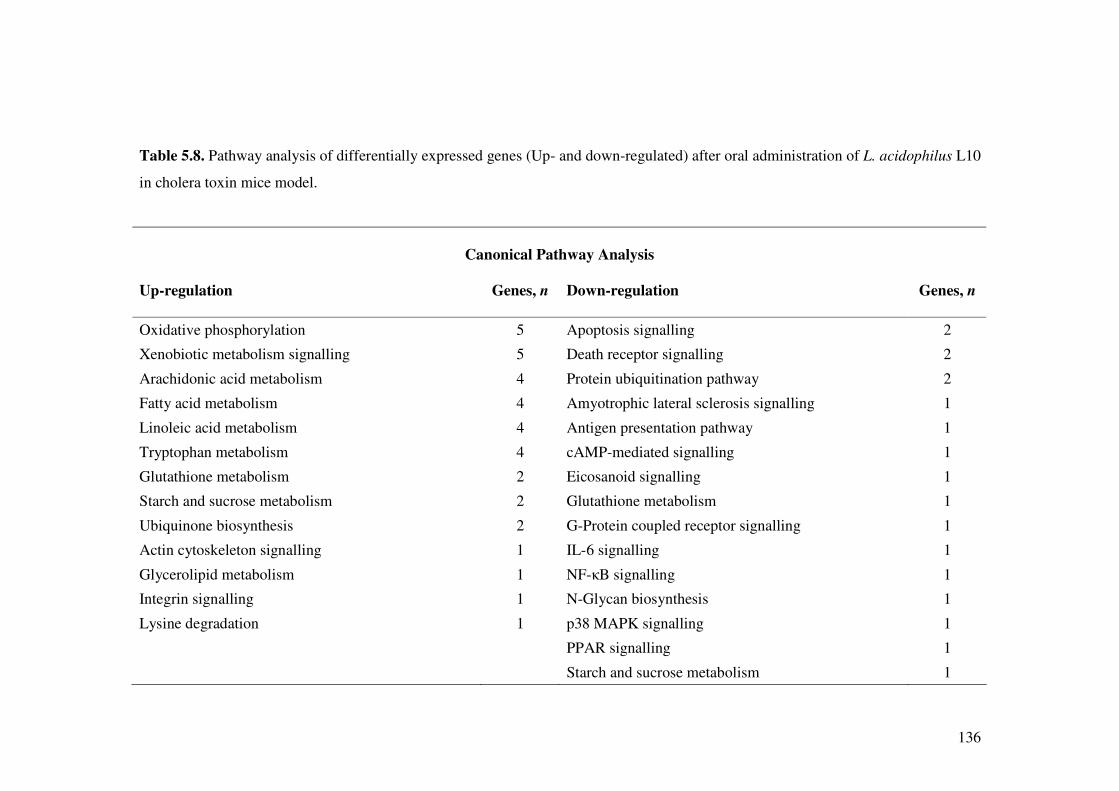
136
Table 5.8. Pathway analysis of differentially expressed genes (Up- and down-regulated) after oral administration of L. acidophilus L10
in cholera toxin mice model.
Canonical Pathway Analysis
Up-regulation
Genes, n Down-regulation Genes, n
Oxidative phosphorylation 5 Apoptosis signalling 2
Xenobiotic metabolism signalling 5 Death receptor signalling 2
Arachidonic acid metabolism 4 Protein ubiquitination pathway 2
Fatty acid metabolism 4 Amyotrophic lateral sclerosis signalling 1
Linoleic acid metabolism 4 Antigen presentation pathway 1
Tryptophan metabolism 4 cAMP-mediated signalling 1
Glutathione metabolism 2 Eicosanoid signalling 1
Starch and sucrose metabolism 2 Glutathione metabolism 1
Ubiquinone biosynthesis 2 G-Protein coupled receptor signalling 1
Actin cytoskeleton signalling 1 IL-6 signalling 1
Glycerolipid metabolism 1 NF-�B signalling 1
Integrin signalling 1 N-Glycan biosynthesis 1
Lysine degradation 1 p38 MAPK signalling 1
PPAR signalling 1
Starch and sucrose metabolism 1

137
5.5. DISCUSSION
The GI tract harbours a well balanced microflora, which is in close and continuous
contact with epithelial and immune cells that are essential for a functionally balanced
immune system. Earlier studies suggested that complex modifications of epithelial cells
could occur due to the interactions between probiotic bacteria and enterocytes (Lu and
Walker, 2001). The cross-talk between microorganisms and enterocytes is required to
influence the mucosal barrier functions, which could favour positive health or cause
disease (Kohler et al., 2003). Lactobacilli remain as a major component of human
intestinal microflora that are frequently considered as good candidates in the preparation
of functional foods. The ability of several LAB strains in modulating host innate and
acquired immune responses has been demonstrated in many in vivo animal models.
However, influence of LAB on gene expression of the intestinal epithelial cells remains
unclear.
The present study investigated the effect of orally administered L. acidophilus on gene
expression in the small intestine of mice. Compugen mouse oligonucleotide microarray
was used to identify genes that were influenced by L. acidophilus through comparing the
gene expression profiles in the small intestine of L. acidophilus-fed mice with the
control group. Orally administered L. acidophilus induced the expression of genes that
are involved in immune and lymphatic development such as attractin and chemokine (C-
X-C motif) ligand 15. Attractin is a normal serum glycoprotein of 175 kDa, which is
expressed on activated T-cells. Attractin gene encoded protein is involved in initial
immune cell clustering during inflammatory responses, which may also regulate
chemotactic activity of chemokines. Attractin protein is also crucial in mediating the

138
clustering of non-proliferative T-lymphocytes and spreading of monocytes (Duke-Cohan
et al., 1998). Furthermore, Nagle et al. (1999) reported the potential role of attractin in
decreasing the obesity that was increased by diet in mice. Chemokine receptors are
suggested to be involved in allergic diseases (Dulkys et al., 2004). Chemokine CXCL12
and its receptor are known to be important in the migration of intestinal epithelial cells,
development of enterocytes and promoting mucosal barrier integrity (Smith et al., 2005).
The expression of chemokine (C-X-C motif) ligand 15 (CXCL15) was enhanced by L.
acidophilus, which is also known to be regulated by the influence of CD14 gene in
immune cells (Benhnia et al., 2005).
Several genes involved in various cellular functions such as cell-cell signalling, cell
development, cell death and cell morphology were up-regulated by L. acidophilus. As an
example, Protocadherin 8 (PCDH8) gene involved in cell signalling was enhanced after
oral administration of L. acidophilus. Protocadherin 8 belongs to protocadherin gene
family, a subfamily of cadherin superfamily that are crucial in cell interactions and
morphogenesis of epithelial tissue (Yagi and Takeichi, 2000; Rhee et al., 2003). Heat
shock protein 1A (HSPA1A) gene involved in cell signalling mechanism was up-
regulated by L. acidophilus. The heat shock proteins are known to be capable of
increasing tumor necrosis factor (TNF)-� protein in macrophages (Zheng et al., 2004).
Additionally, genes involved in cell signalling mechanism such as protein tyrosine
phosphatase, receptor type D (PTPRD) and mitogen-activated protein kinase kinase
kinase kinase 1 (MAP4K1) were up-regulated by L. acidophilus. The PTPRD gene is a
member of protein-tyrosine phosphatase (PTP) family, which are signalling molecules

139
involved in variety of cellular processes including cell growth, differentiation, mitotic
cycle, and oncogenic transformation. The protein-tyrosine phosphatases are also
involved in dephosphorylation of amino acids (Pulido et al., 1995). The MAPK family is
a major signalling system, which is considered as an intracellular mediator of
inflammation and also known to play a functional role in c-Jun amino-terminal kinases
(JNKs)/stress-activated protein kinases (SAPKs) pathway (Hu et al., 1996; van den
Blink et al., 2002).
Other genes up-regulated by L. acidophilus are involved in lipid metabolism such as
angiopoietin-like 4 (ANGPTL4) and ATP-binding cassette, sub-family A, member 12
(ABCA12). The ANGPTL4 is a member of angiopoietin/angiopoietin-like gene family,
which is a blood-borne hormone directly involved in regulating glucose homeostasis,
lipid metabolism, and insulin sensitivity. Human clinical trials reported the decrease of
ANGPTL4 serum levels compared to healthy subjects, which suggested that lower
ANGPTL4 levels could be a causative factor for type 2 diabetes (Xu et al., 2005).
Furthermore, Kaneda et al. (2002) reported that aberrant methylation of this gene is
associated with human gastric cancers. The ABCA12 gene encoded protein is a member
of ATP-binding cassette transporters superfamily, which is involved in transporting
various molecules across extra- and intracellular membranes.
Another gene enhanced by oral administration of L. acidophilus is lysozyme, which is
generally regarded as an anti-microbial agent found in human milk, spleen, white blood
cells and plasma. Several studies reported the importance of lysozyme expression in

140
human B-lymphocytes and gastric adenocarcinoma tissues (Hogerkorp et al., 2003; Oien
et al., 2003).
Lactobacillus acidophilus down-regulated genes involved in lipid metabolism such as
pancreatic lipase (PNLIP), carboxyl ester lipase (CEL) and phospholipase A2, group IB
(PLA2G1B). Pancreatic lipase (PNLIP) is a member of lipase gene family, which
encodes a carboxyl esterase that is essential for efficient digestion of dietary fats.
Pancreatic triglyceride lipase deficiency was shown to have minimal affect on dietary fat
absorption, however dietary cholesterol absorption was decreased in mice (Huggins et
al., 2003). The CEL is a glycoprotein secreted into digestive tract from pancreas, which
is necessary in the catabolism of cholesterol (Ikeda et al., 2002) and also influence the
secretion and assembly of lipoprotein through ceramide hydrolysis in mice intestine
(Kirby et al., 2002). Moreover, PLA2G1B gene involved in the catalyses of releasing
fatty acids from glycero-3-phosphocholines was down-regulated by L. acidophilus.
Several forms of PLA2 are known to be present in spleen, macrophages, leukocytes and
erythrocytes. In vitro studies using HL-60 cells reported the influence of interferon
(IFN)-� protein in increasing PLA2G1B protein activity (Visnjic et al., 1997).
Lactobacillus acidophilus down-regulated trefoil factor 2 (TFF2) gene, which is a
member of trefoil family that are commonly known as stable secretory proteins
expressed in gastrointestinal mucosa. Trefoil family secreted proteins play an active role
in protecting the mucosa by stabilising mucus layer and healing epithelial cells. In vitro
studies using human bronchial epithelial cell line, BEAS-2B demonstrated the capability
of TFF-peptides in modulating the inflammatory responses by regulating TNF-� induced

141
secretion of interleukin (IL)-6 and IL-8 (Graness et al., 2002). Furthermore, matrix
metallopeptidase 2 (MMP2) gene was also down-regulated by L. acidophilus. The
proteins that belong to MMP family are involved in tissue remodelling, arthritis and
metastasis disease processes. Several studies reported the role of MMP activity in
apoptosis of epithelial cells (Wiseman et al., 2003) and cancer (Egeblad and Werb,
2002).
In the second experiment, L. acidophilus induced gene expression in the small intestine
was determined in cholera toxin mice using compugen mouse oligonucleotide
microarray. Lactobacillus acidophilus induced gene expression profiles in the small
intestine of mice were identified by comparing with the control group. Mice fed with L.
acidophilus demonstrated the up-regulation of CD36 gene, which is involved in cell-cell
signalling mechanism. The CD36 encoded protein acts as a receptor for thrombospondin
in various cell lines. The CD36 protein also involves in various adhesive processes due
to its functional role as cell adhesion molecule. Recent study reported the role of CD36
in proinflammatory responses, particularly as a recognition receptor that mediates
microglial and macrophage response to beta-amyloid (El Khoury et al., 2003).
Several genes involved in cell death such as glutathione S-transferase pi (GSTP1),
fibronectin 1 (FN1) and ISG15 ubiquitin-like modifier (ISG15) were up-regulated by L.
acidophilus. The GSTP1 is a polymorphic gene, which belongs to GST family of
enzymes that are crucial in catalysing the conjugation of many hydrophobic and
electrophilic compounds through reduced glutathione. The GSTP1 gene is known to be
functional in xenobiotic metabolism and is also involved in the regulation of cell

142
signalling pathways (Gate et al., 2004). The FN1 gene is a glycoprotein, which involves
in cell adhesion and migration processes including host defence functions. Earlier it was
reported that fibronectin type III A1A2 domain was capable of regulating T-lymphocyte
proliferation and cytokine production (Puente Navazo et al., 2001). Furthermore,
fibronectin protein also showed the influence in wound healing process (Rybarczyk et
al., 2003). Moreover, ISG15 gene was up-regulated, which is involved in cell death
mechanism. The ISG15 gene, which is a member of cytokine cascade mechanism, plays
an important role in several host immune functions such as innate immune response and
regulating interferon signalling. D'Cunha et al. (1996) also suggested the role of ISG15
gene in immunomodulatory effects of IFN-� or IFN-�.
Lactobacillus acidophilus down-regulated genes involved in cell growth and
proliferation such as carcinoembryonic antigen-related cell adhesion molecule 1
(CEACAM1), clusterin (CLU) and tumor necrosis factor receptor superfamily, member
1A (TNFRSF1A). The CEACAM1 gene belongs to carcinoembryonic antigen (CEA)
gene family, which is a cell surface molecule known to have regulatory effects on T-cell
functions. Earlier studies reported the involvement of CEACAM1 protein in decreasing
tumor development and oxazolone colitis (Fournes et al., 2001; Iijima et al., 2004).
Furthermore, CEACAM1 protein also acts as a signal-transducing receptor, which could
regulate early maturation and activation of dendritic cells (Kammerer et al., 2001). The
CLU is a glycoprotein of 75-80 kDa in size secreted in physiological fluids, which is
known to be involved in apoptosis and colorectal cancer (Scaltriti et al., 2004; Andersen
et al., 2007). Moreover, L. acidophilus also down-regulated TNFRSF1A gene, which is
a member of TNF-receptor superfamily that are potential regulators of apoptosis, a

143
process considered to be important in maintaining immune homeostasis. Particularly,
TNFRSF1A protein is known to have a functional role as a regulator in inflammatory
responses. Several studies also reported the involvement of TNF-receptor proteins in
decreasing obesity, liver inflammation and tumor formation (Chan et al., 2003; Francis
et al., 2003; Arnott et al., 2004).
Oral administration of L. acidophilus down-regulated villin 1 (VIL1), which is an actin-
binding protein localises to the brush borders of intestine. Villin 1 gene is a member of
gelsolin family of proteins, which acts as a regulator in hepatocyte growth factor-
induced epithelial cell motility (Athman et al., 2003). The CD74 gene was also down-
regulated by L. acidophilus, which is considered to be important in the activation of
ERK-1/2 MAP kinase cascade and cell proliferation (Leng et al., 2003). Furthermore, L.
acidophilus down-regulated apolipoprotein B (APOB), which occurs in two forms in
plasma known as APOB-48 and APOB-100. Interestingly, Farese et al. (1995) reported
the potential role of APOB gene in diet-induced hypercholesterolemia mice model.
The present study generated a list of genes that were influenced by L. acidophilus in
normal and cholera toxin mice. To gain further insights into the potential value of this
data, it is important to analyse the gene expression in various segments of GI tract as
well as in distant mucosal site such as spleen in animal models. This study evaluated the
effects of a specific probiotic strain in the small intestine of mice for a specific period.
Therefore, it is also reasonable to predict that various factors such as strain variations,
dosage or duration of the probiotic feeding may affect in modulating the gene

144
expression. Furthermore, human trials with different clinical settings are necessary to
understand the clinical value of the microarray data reported in this study.
Nevertheless, this study represents a basic methodology to understand the influence of L.
acidophilus in regulating gene expression that was involved in various biological
functions.

145
5.6. CONCLUSIONS
Oral administration of L. acidophilus influenced the regulation of genes involved in
various biological functions of the small bowel in normal and cholera toxin mice. The
current study generated a comprehensive list of genes that were affected by L.
acidophilus in small intestine of mice, which lay a basis for further understanding the
interactions between probiotic bacteria and the host.

146
CHAPTER 6
Concluding summary

147
The current study examined the probiotic properties of L. acidophilus LAFTI L10 and L.
paracasei LAFTI L26 using various in vitro methods. The influence of orally
administered L. acidophilus and L. paracasei on gut and systemic immune responses in
normal and cholera toxin (CT) mice were investigated. Furthermore, L. acidophilus
induced gene expression was examined in the small intestine of normal and CT mice.
The intestinal microflora play a significant role in influencing the structural and
functional development of gut immune system. Lactic acid bacteria are non-pathogenic
food grade microorganisms that are natural components of intestinal microflora. Lactic
acid bacteria are important in a well balanced intestinal microbial ecosystem, which are
also known for their health-promoting attributes. Earlier studies suggested that probiotic
bacteria must be tolerant to adverse conditions of GI tract, especially in stomach that has
lower pH and high bile salt concentrations (Prasad et al., 1998; Mishra and Prasad,
2005). Furthermore, adhesive ability of probiotic bacteria to intestinal mucosa is also
considered as a prerequisite for colonisation and modulation of host immune functions
(Castagliuolo et al., 2005; Aslim et al., 2007).
The initial work in this study was focused in characterising the probiotic properties of
Lactobacillus and Bifidobacterium species to identify potential probiotic strains for
further in vivo mice studies. Lactobacillus acidophilus LAFTI L10 and L. paracasei
LAFTI L26 showed higher tolerance to simulated gastric conditions and bile salts
(Chapter 2). Lactobacillus acidophilus and L. paracasei demonstrated the correlation
between in vitro adhesive properties such as hydrophobicity and autoaggregation.
Coaggregation abilities of L. acidophilus and L. paracasei with other bacterial strains

148
showed significant variations, which could be due to the differences in their cell surface
proteins. Particularly, L. acidophilus demonstrated higher level of coaggregation with L.
paracasei and other probiotic strains. The tolerance of L. acidophilus and L. paracasei
to gastric pH 2.0 and 1.0% bile salts demonstrated the potential of these strains to
survive and colonise especially in the small intestine to induce beneficial effects on the
host.
Orally administered L. acidophilus and L. paracasei at 108 CFU/day for 14-days in
BALB/c mice stimulated the host immune functions (Chapter 3). Immunofluorescent
examination of small intestine in mice demonstrated the capability of L. acidophilus and
L. paracasei in enhancing immunoglobulin (Ig)-A producing B-cells, which are
necessary in mucosal defence mechanisms. The T-cells in small intestine of mice fed
with either L. acidophilus or L. paracasei showed an increase of anti-inflammatory
cytokine, interleukin (IL)-10 and pro-inflammatory cytokine, interferon (IFN)-�.
Lactobacillus acidophilus and L. paracasei were capable of enhancing IL-10 cytokine,
which is known to have a beneficial role in preventing immune hypersensitivity/atopy
(Majamaa and Isolauri, 1997; Kalliomaki et al., 2001) and alleviating inflammatory
bowel disease (Madsen et al., 1999; Matsumoto et al., 2001). In systemic immunity,
proliferative responses of splenocytes stimulated with concanavalin A (ConA) and
lipopolysaccharide (LPS) were increased in mice fed with L. acidophilus or L.
paracasei. Particularly, LPS induced proliferative responses of splenocytes in mice fed
with L. paracasei were not significantly higher compared to control mice.

149
Mice fed with either L. acidophilus or L. paracasei regulated the systemic immune
response by increasing Th1 cytokine, IFN-� and Th2 cytokine, IL-10 in serum and
spleen cells (Chapter 3). In the current study, stimulation of systemic immune responses
by probiotic bacteria might have occurred through the bacterial antigen uptake by M-
cells of Peyer’s patches in the small intestine. In Peyer’s patches, probiotic bacterial
antigens interact with lymphocytes that lead to the activation of antigen-specific
lymphocytes, which could have migrated through mesenteric lymph nodes and then
entered into blood stream via thoracic duct to stimulate systemic immune functions.
Lactobacillus acidophilus and L. paracasei also showed the capability in stimulating
non-specific immune functions such as macrophage activity (Chapter 3). The peritoneal
macrophages of mice fed with L. acidophilus or L. paracasei demonstrated the
enhancement of phagocytic activity against C. albicans compared to control mice.
Vinderola et al. (2004) suggested that lack of influence of probiotic bacteria in activating
systemic immune functions such as macrophage activity could be due to the inhibitory
effect of IL-10. Interestingly, inhibitory effect of IL-10 on macrophage activation was
not observed in the current study, where L. acidophilus and L. paracasei enhanced the
Th2 cytokine, IL-10 in serum and spleen cells (Chapter 3). As shown in chapter 2,
tolerance of L. acidophilus and L. paracasei to simulated acid and bile conditions and
their better adherence capabilities may have favoured them to persist, colonise and
adhere in GI tract of mice to stimulate the macrophage activity (Perdigon and Alvarez,
1992; Schiffrin et al., 1997).

150
The differences in translocation of Lactobacillus spp. and total anaerobes to Peyer’s
patches and MLN were observed in mice fed with L. acidophilus or L. paracasei
(Chapter 3). Particularly, L. acidophilus-fed mice showed higher translocation of
Lactobacillus spp. and total anaerobes to Peyer’s patches compared to L. paracasei-fed
mice and control group. Mice fed with L. acidophilus or L. paracasei showed increased
bacterial translocation to Peyer’s patches and MLN, which could be due to their
adhesive properties (Wagner et al., 1997) that may have contributed to the enhancement
of various gut and systemic immune functions. Furthermore, oral administration of L.
acidophilus or L. paracasei at 108 CFU/day for 14-days in BALB/c mice did not cause
intestinal inflammation, as there was no indication of translocation of Lactobacillus spp.
and total anaerobes to sterile organs such as spleen, liver and blood.
The influence of orally administered L. acidophilus and L. paracasei on host immune
responses was investigated in CT mice (Chapter 4). Lactobacillus acidophilus and L.
paracasei showed the differences in inducing nitric oxide (NO) production in intestinal
fluid and serum. Particularly, L. paracasei demonstrated as a potential inducer of NO
levels in intestinal fluid and serum compared to L. acidophilus. The potential of L.
paracasei in inducing the release of NO in intestinal fluid may be necessary to play a
protective role in colitis (Wallace et al., 1999; Lamine et al., 2004). Both, L. acidophilus
and L. paracasei increased the IgA response to CT in intestinal fluid and serum, which
could be considered as a potential attribute necessary in host immune defence
mechanisms.

151
As shown in chapter 4, immunofluorescent studies demonstrated the potential of L.
acidophilus and L. paracasei in regulating Th1- and Th2-cytokines in the small intestine
of CT mice. Both L. acidophilus and L. paracasei enhanced the IL-6 producing cells in
small intestine, which may have contributed to the stimulation of IgA response to CT in
intestinal fluid and serum. Lactobacillus acidophilus and L. paracasei showed the
regulation of anti-inflammatory cytokine, IL-10 and pro-inflammatory cytokine, IFN-�
in the small intestine of CT mice. Similar effects of L. acidophilus and L. paracasei in
regulating IL-10 and IFN-� was also observed in the small intestine of normal mice
(Chapter 3). In the current study, stimulation of IFN-� producing cells in small intestine
of normal and CT mice fed with L. acidophilus or L. paracasei may have contributed to
the increase in uptake of bacterial antigens into Peyer’s patches (Sutas et al., 1997),
which lead to the stimulation of gut and systemic immune functions (Chapters 3 & 4).
Moreover, mice fed with L. acidophilus or L. paracasei also enhanced the stimulation of
tumor necrosis factor (TNF)-� and IL-4 cytokines in small intestine of CT mice. Further
studies on dose-dependent oral administration of L. acidophilus and L. paracasei are
needed to understand the cytokine profiles at different time intervals.
Considering the in vitro probiotic characteristics of L. acidophilus and its immune
enhancing properties in normal and CT mice, L. acidophilus induced gene expression in
the small intestine of normal and CT mice was investigated using mouse oligonucleotide
microarray (Chapter 5). In normal mice, L. acidophilus altered the gene expression in
the small intestine through up- and down-regulation of 27 and 35 genes. Orally
administered L. acidophilus affected the genes that are important in cell signalling such
as HSPA1A and MAP4K1. The up-regulation of HSPA1A by L. acidophilus showed its

152
potential in interacting with intestinal epithelial cells to maintain cytoskeletal integrity.
Particularly, L. acidophilus-fed mice up-regulated MAP4K1 gene, which is an
intracellular signalling molecule that may have influenced the expression of HSPA1A in
intestinal epithelial cells (Tao et al., 2006). Moreover, L. acidophilus-fed mice up-
regulated the genes involved in immune responses (attractin and CXCL15) and cell
signalling (PCDH8), whereas the down-regulated genes are functional in lipid
metabolism (CEL, PLA2G1B and PNLIP) and apoptosis (MMP2).
In CT mice model, L. acidophilus modulated the gene expression in small intestine
through an up- and down-regulation of 28 and 32 genes. Orally administered L.
acidophilus up-regulated CD36 gene, which is involved in cell-cell signalling and
known to have a significant role in pro-inflammatory responses (El Khoury et al., 2003).
Other genes up-regulated by L. acidophilus are involved in cell death such as FN1,
GSTP1 and ISG15. Particularly, genes that belong to cytochrome P450 family
(CYP2C9, CYP2C19, CYP2C38 and CYP3A5) were also up-regulated by L.
acidophilus. Interestingly, L. acidophilus down-regulated TNFRSF1A gene, which is
considered to have a regulatory role in inflammatory mechanisms (Chan et al., 2003).
Additionally, L. acidophilus also down-regulated genes such as VIL1, CD74 and APOB.
The present study investigated the influence of L. acidophilus on gene expression in the
small intestine of mice, since it is considered as an important site for probiotic bacterial
interactions with intestinal epithelial cells. The analysis of L. acidophilus induced gene
expression in distant mucosal site such as spleen could be useful in further elucidating
the mechanisms through which these probiotic bacteria are beneficial to the host.

153
Moreover, the current study generated the pool of genes in small intestine of normal and
CT mice that were modified by oral administration of L. acidophilus. Lactobacillus
acidophilus induced gene expression was analysed in normal and CT mice. Therefore, it
is reasonable to suggest that the analysis of gene expression modulated by L. acidophilus
was limited and could possibly differ in various animal models. Furthermore, validating
the influence of L. acidophilus on gene expression in human clinical trails is necessary
to unravel the beneficial role of probiotic bacteria.

154
REFERENCES

155
Aiba, Y., Suzuki, N., Kabir, A.M., Takagi, A., Koga, Y., 1998. Lactic acid-mediated
suppression of Helicobacter pylori by the oral administration of Lactobacillus
salivarius as a probiotic in a gnotobiotic murine model. American Journal of
Gastroenterology 93, 2097-2101.
Alam, M., Midtvedt, T., Uribe, A., 1994. Differential cell kinetics in the ileum and colon
of germfree rats. Scandinavian Journal of Gastroenterology 29, 445-451.
Alander, M., Korpela, R., Saxelin, M., Vilpponen-Salmela, T., Mattila-Sandholm, T.,
von Wright, A., 1997. Recovery of Lactobacillus rhamnosus GG from human
colonic biopsies. Letters in Applied Microbiology 24, 361-364.
Alican, I., Kubes, P., 1996. A critical role for nitric oxide in intestinal barrier function
and dysfunction. American Journal of Physiology 270, G225-G237.
Alm, L., 1982. Effect of fermentation on lactose, glucose, and galactose content in milk
and suitability of fermented milk products for lactose intolerant individuals.
Journal of Dairy Science 65, 346-352.
Andersen, C.L., Schepeler, T., Thorsen, K., Birkenkamp-Demtroder, K., Mansilla, F.,
Aaltonen, L.A., Laurberg, S., Orntoft, T.F., 2007. Clusterin expression in normal
mucosa and colorectal cancer. Molecular & Cellular Proteomics 6, 1039-1048.
Anderson, J.W., Gilliland, S.E., 1999. Effect of fermented milk (yoghurt) containing
Lactobacillus acidophilus L1 on serum cholesterol in hypercholesterolemic
humans. Journal of the American College of Nutrition 18, 43-50.
Arnott, C.H., Scott, K.A., Moore, R.J., Robinson, S.C., Thompson, R.G., Balkwill, F.R.,
2004. Expression of both TNF-alpha receptor subtypes is essential for optimal
skin tumour development. Oncogene 23, 1902-1910.

156
Aslim, B., Onal, D., Beyatli, Y., 2007. Factors influencing autoaggregation and
aggregation of Lactobacillus delbrueckii subsp. bulgaricus isolated from
handmade yogurt. Journal of Food Protection 70, 223-227.
Athman, R., Louvard, D., Robine, S., 2003. Villin Enhances Hepatocyte Growth Factor-
induced Actin Cytoskeleton Remodeling in Epithelial Cells. Molecular Biology
of the Cell 14, 4641-4653.
Backhed, F., Ding, H., Wang, T., Hooper, L.V., Koh, G.Y., Nagy, A., Semenkovich,
C.F., Gordon, J.I., 2004. The gut microbiota as an environmental factor that
regulates fat storage. Proceedings of the National Academy of Sciences USA
101, 15718-15723.
Backhed, F., Ley, R.E., Sonnenburg, J.L., Peterson, D.A., Gordon, J.I., 2005. Host-
bacterial mutualism in the human intestine. Science 307, 1915-1920.
Belardelli, F., Ferrantini, M., 2002. Cytokines as a link between innate and adaptive
antitumor immunity. Trends in Immunology 23, 201-208.
Bengmark, S., 1998. Ecological control of the gastrointestinal tract. The role of probiotic
flora. Gut 42, 2-7.
Bengmark, S., Jeppsson, B., 1995. Gastrointestinal surface protection and mucosa
reconditioning. Journal of Parenteral and Enteral Nutrition 19, 410-415.
Benhnia, M.R., Wroblewski, D., Akhtar, M.N., Patel, R.A., Lavezzi, W., Gangloff, S.C.,
Goyert, S.M., Caimano, M.J., Radolf, J.D., Sellati, T.J., 2005. Signaling through
CD14 Attenuates the Inflammatory Response to Borrelia burgdorferi, the Agent
of Lyme Disease. Journal of Immunology 174, 1539-1548.

157
Berg, R.D., 1992. Translocation of enteric bacteria in health and disease. Current Studies
in Hematology and Blood Transfusion 59, 44-65.
Berg, R.D., 1998. Probiotics, prebiotics or 'conbiotics'?. Trends in Microbiology 6, 89-
92.
Berg, R.D., 1999. Bacterial translocation from the gastrointestinal tract. Advances in
experimental medicine and biology 473, 11-30.
Blanchard, R.K., Moore, J.B., Green, C.L., Cousins, R.J., 2001. Modulation of intestinal
gene expression by dietary zinc status: effectiveness of cDNA arrays for
expression profiling of a single nutrient deficiency. Proceedings of the National
Academy of Sciences USA 98, 13507-13513.
Blum, S., Delneste, Y., Alvarez, S., Haller, D., Perez, P.F., Bode, Ch., Hammes, W.P.,
Pfeifer, A.M.A., Schiffrin, E.J., 1999. Interactions between commensal
bacteria and mucosal immunocompetent cells. International Dairy Journal 9, 63-
68.
Boris, S., Suarez, J.E., Barbes, C., 1997. Characterization of the aggregation promoting
factor from Lactobacillus gasseri, a vaginal isolate. Journal of Applied
Microbiology 83, 413-420.
Borruel, N., Casellas, F., Antolin, M., Llopis, M., Carol, M., Espiin, E., Naval, J.,
Guarner, F., Malagelada, J.R., 2003. Effects of nonpathogenic bacteria on
cytokine secretion by human intestinal mucosa. American Journal of
Gastroenterology 98, 865-870.
Braat, H., Peppelenbosch, M.P., Hommes, D.W., 2003. Interleukin-10-based therapy for
inflammatory bowel disease. Expert Opinion on Biological Therapy 3, 725-731.

158
Bradley, H.K., Wyatt, G.M., Bayliss, C.E., Hunter, J.O., 1987. Instability in the faecal
flora of a patient suffering from food-related irritable bowel syndrome. Journal
of Medical Microbiology 23, 29-32.
Brigidi, P., Vitali, B., Swennen, E., Bazzocchi, G., Matteuzzi, D., 2001. Effects of
probiotic administration upon the composition and enzymatic activity of human
fecal microbiota in patients with irritable bowel syndrome or functional diarrhea.
Research in Microbiology 152, 735-741.
Bron, P.A., Grangette, C., Mercenier, A., de Vos, W.M., Kleerebezem M., 2004.
Identification of Lactobacillus plantarum genes that are induced in the
gastrointestinal tract of mice. Journal of Bacteriology 186, 5721-5729.
Brown, P.O., Botstein, D., 1999. Exploring the new world of the genome with DNA
microarrays. Nature Genetics 21, 33-37.
Bujalance, C., Moreno, E., Jimenez-Valera, M., Ruiz-Bravo, A., 2007. A probiotic strain
of Lactobacillus plantarum stimulates lymphocyte responses in immunologically
intact and immunocompromised mice. International Journal of Food
Microbiology 113, 28-34.
Cano, P.G., Aguero, G., Perdigon, G., 2002. Immunological effects of yogurt addition to
a re-nutrition diet in a malnutrition experimental model. Journal of Dairy
Research 69, 303-316.
Castagliuolo, I., Galeazzi, F., Ferrari, S., Elli, M., Brun, P., Cavaggioni, A., Tormen, D.,
Sturniolo, G.C., Morelli, L., Palu, G., 2005. Beneficial effect of auto-aggregating
Lactobacillus crispatus on experimentally induced colitis in mice. FEMS
Immunology and Medical Microbiology 43, 197-204.

159
Cayzer, C., Patel, K., Wang, X., Florin, T., 2001. Effect of probiotics on Th1 and Th2
cytokines: Relevance to pathogenesis and treatment of ulcerative colitis. Journal
of Gastroenterology and Hepatology 16(S1), 36.
Cebra, J.J., 1999. Influences of microbiota on intestinal immune system development.
American Journal of Clinical Nutrition 69, 1046S-1051S.
Cebra, J.J., Jiang, H.Q., Boiko, N., Tlaskalova-Hogenova, H., 2005. The role of
mucosal microbiota in the development, maintenance, and pathologies of the
mucosal immune system. In: Mestecky, J., Lamm, M.E., Strober, W.,
Bienenstock, J., McGhee, J.R., Mayer, L., (eds.), Mucosal Immunology,
Academic Press, New York, pp. 335-368.
Cesena, C., Morelli, L., Alander, M., Siljander, T., Tuomola, E., Salminen, S., Mattila-
Sandholm, T., Vilpponen-Salmela, T., von Wright, A., 2001. Lactobacillus
crispatus and its nonaggregating mutant in human colonization trials. Journal of
Dairy Science 84, 1001-1010.
Chan, F.K., Shisler, J., Bixby, J.G., Felices, M., Zheng, L., Appel, M., Orenstein, J.,
Moss, B., Lenardo, M.J., 2003. A role for tumor necrosis factor receptor-2 and
receptor-interacting protein in programmed necrosis and antiviral responses.
Journal of Biological Chemistry 278, 51613-51621.
Charteris, W.P., Kelly, P.M., Morelli, L., Collins, J.K., 1998. Development and
application of an in vitro methodology to determine the transit tolerance of
potentially probiotic Lactobacillus and Bifidobacterium species in the upper
human gastrointestinal tract. Journal of Applied Microbiology 84, 759-768.
Chiang, B.L., Sheih, Y.H., Wang, L.H., Liao, C.K., Gill, H.S., 2000. Enhancing
immunity by dietary consumption of a probiotic lactic acid bacterium

160
(Bifidobacterium lactis HN019): optimization and definition of cellular immune
responses. European Journal of Clinical Nutrition 54, 849–855.
Chou, L.S., Weimer, B., 1999. Isolation and characterization of acid- and bile-tolerant
isolates from strains of Lactobacillus acidophilus. Journal of Dairy Science 82,
23-31.
Christensen, H.R., Frokiaer, H., Pestka, J.J., 2002. Lactobacilli differentially modulate
expression of cytokines and maturation surface markers in murine dendritic cells.
Journal of Immunology 168, 171-178.
Chung, H.S., Kim, Y.B., Chun, S.L., Ji, G.E., 1999. Screening and selection of acid and
bile resistant bifidobacteria. International Journal of Food Microbiology 47, 25-
32.
Clancy, R.L., Gleeson, M., Cox, A., Callister, R., Dorrington, M., D'Este, C., Pang, G.,
Pyne, D., Fricker, P., Henriksson, A., 2006. Reversal in fatigued athletes of a
defect in interferon gamma secretion after administration of Lactobacillus
acidophilus. British Journal of Sports Medicine 40, 351-354.
Clark, P.A., Martin, J.H., 1994. Selection of bifidobacteria for use as dietary adjuncts in
cultures dairy foods: III – Tolerance to simulated bile concentrations of human
small intestines. Cultured Dairy Products Journal 29, 18-21.
Conforti, A., Donini, M., Brocco, G., Del Soldato, P., Benoni, G., Cuzzolin, L., 1993.
Acute anti-inflammatory activity and gastrointestinal tolerability of diclofenac
and nitrofenac. Agents and Actions 40, 176-180.
Conway, P.L., Gorbach, S.L., Goldin, B.R., 1987. Survival of lactic acid bacteria in the
human stomach and adhesion to intestinal cells. Journal of Dairy Science 70, 1-
12.

161
Conway, P.L., Kjelleberg, S., 1989. Protein-mediated adhesion of Lactobacillus
fermentum strain 737 to mouse stomach squamous epithelium. Journal of
General Microbiology 135, 1175-1186.
Corfield, A.P., Carroll, D., Myerscough, N., Probert, C.S., 2001. Mucins in the
gastrointestinal tract in health and disease. Frontiers in Bioscience 6, D1321-
D1357.
Cossart, P., 1997. Host/pathogen interactions. Subversion of the mammalian cell
cytoskeleton by invasive bacteria. Journal of Clinical Investigation 99, 2307-
2311.
Crabbe, P.A., Bazin, H., Eyssen, H., Heremans, J.F., 1968. The normal microbial flora
as a major stimulus for proliferation of plasma cells synthesizing IgA in the gut.
The germ-free intestinal tract. International Archives of Allergy and Applied
Immunology 34, 362-375.
Cross, M.L., Mortensen, R.R., Kudsk, J., Gill, H.S., 2002. Dietary intake of
Lactobacillus rhamnosus HNOO1 enhances production of both Th1 and Th2
cytokines in antigen-primed mice. Medical Microbiology and Immunology 191,
49-53.
Cruchet, S., Obregon, M.C., Salazar, G., Diaz, E., Gotteland, M., 2003. Effect of the
ingestion of a dietary product containing Lactobacillus johnsonii La1 on
Helicobacter pylori colonization in children. Nutrition 19, 716– 721.
Daniel, C., Poiret, S., Goudercourt, D., Dennin, V., Leyer, G., Pot, B., 2006. Selecting
lactic acid bacteria for their safety and functionality by use of a mouse colitis
model. Applied and Environmental Microbiology 72, 5799-5805.

162
D'Cunha, J., Knight, E.Jr., Haas, A.L., Truitt, R.L., Borden, E.C., 1996.
Immunoregulatory properties of ISG15, an interferon-induced cytokine.
Proceedings of the National Academy of Sciences USA 93, 211-215.
de Moreno de LeBlanc, A., Matar, C., Leblanc, N., Perdigon, G., 2005. Effects of milk
fermented by Lactobacillus helveticus R389 on a murine breast cancer model.
Breast Cancer Research 7, R477-R486.
De Simone, C., Bianchi Salvadori, B., Negri, R., Ferrazzi, M., Baldinelli, L., Vesely, R.,
1986. The adjuvant effect of yogurt on production of gamma-interferon by
ConA-stimulated human peripheral blood lymphocytes. Nutrition Reports
International 33, 419–433.
De Simone, C., Vesely, R., Bianchi-Salvadori, B., Jirillo, E., 1993. The role of
probiotics in modulation of the immune system in man and animals. International
Journal of Immunotherapy 9, 23-28.
de Vos, W.M., Bron, P.A., Kleerebezem, M., 2004. Post-genomics of lactic acid bacteria
and other food-grade bacteria to discover gut functionality. Current Opinion in
Biotechnology 15, 86-93.
de Waard, R., Claassen, E., Bokken, G.C., Buiting, B., Garssen, J., Vos, J.G., 2003.
Enhanced immunological memory responses to Listeria monocytogenes in
rodents, as measured by delayed-type hypersensitivity (DTH), adoptive transfer
of DTH, and protective immunity, following Lactobacillus casei Shirota
ingestion. Clinical and Diagnostic Laboratory Immunology 10, 59-65.
Deeth, H.C., Tamime, A.Y., 1981. Yogurt, nutritive and therapeutic aspects. Journal of
Food Protection 44, 78-86.

163
Del Re, B., Busetto, A., Vignola, G., Sgorbati, B., Palenzona, D.L., 1998.
Autoaggregation and adhesion ability in a Bifidobacterium suis strain. Letters in
Applied Microbiology 27, 307-310.
Del Re, B., Sgorbati, B., Miglioli, M., Palenzona, D., 2000. Adhesion, autoaggregation
and hydrophobicity of 13 strains of Bifidobacterium longum. Letters in Applied
Microbiology 31, 438-442.
Deplancke, B., Gaskins, H.R., 2001. Microbial modulation of innate defense: goblet
cells and the intestinal mucus layer. American Journal of Clinical Nutrition 73,
1131S-1141S.
Di Giacinto, C., Marinaro, M., Sanchez, M., Strober, W., Boirivant, M., 2005. Probiotics
ameliorate recurrent Th1-mediated murine colitis by inducing IL-10 and IL-10-
dependent TGF-beta-bearing regulatory cells. Journal of Immunology 174, 3237-
3246.
Dodd, H.M., Gasson, M.J., 1994. Bacteriocins of Lactic acid bacteria. In: Gasson, M.J.,
de Vos, W.M., (eds.), Genetics and Biotechnology of Lactic acid bacteria,
Blackie Academic and Professional, Glasgow, pp. 211-251.
Dogi, C.A., Perdigon, G., 2006. Importance of the host specificity in the selection of
probiotic bacteria. Journal of Dairy Research 73, 357-366.
Donnet-Hughes, A., Rochat, F., Serrant, P., Aeschlimann, J.M., Schiffrin, E.J., 1999.
Modulation of nonspecific mechanisms of defense by lactic acid bacteria:
effective dose. Journal of Dairy Science 82, 863-869.
Drago, L., Gismondo, M.R., Lombardi, A., de Haen, C., Gozzini, L., 1997. Inhibition of
in vitro growth of enteropathogens by new Lactobacillus isolates of human
intestinal origin. FEMS Microbiology Letters 153, 455-463.

164
Duchmann, R., Kaiser, I., Hermann, E., Mayet, W., Ewe, K., Meyer zum Buschenfelde,
K.H., 1995. Tolerance exists towards resident intestinal flora but is broken in
active inflammatory bowel disease (IBD). Clinical and Experimental
Immunology 102, 448-455.
Duke-Cohan, J.S., Gu, J., McLaughlin, D.F., Xu, Y., Freeman, G.J., Schlossman, S.F.,
1998. Attractin (DPPT-L), a member of the CUB family of cell adhesion and
guidance proteins, is secreted by activated human T lymphocytes and modulates
immune cell interactions. Proceedings of the National Academy of Sciences
USA 95, 11336-11341.
Dulkys, Y., Buschermohle, T., Escher, S.E., Kapp, A., Elsner, J., 2004. T-helper 2
cytokines attenuate senescent eosinophil activation by the CXCR4 ligand
stromal-derived factor-1alpha (CXCL12). Clinical & Experimental Allergy 34,
1610-1620.
Egeblad, M., Werb, Z., 2002. New functions for the matrix metalloproteinases in cancer
progression. Nature reviews. Cancer 2, 161-174.
Ehrmann, M.A., Kurzak, P., Bauer, J., Vogel, R.F., 2002. Characterization of lactobacilli
towards their use as probiotic adjuncts in poultry. Journal of Applied
Microbiology 92, 966-975.
El Khoury, J.B., Moore, K.J., Means, T.K., Leung, J., Terada, K., Toft, M., Freeman,
M.W., Luster, A.D., 2003. CD36 mediates the innate host response to beta-
amyloid. Journal of Experimental Medicine 197, 1657-1666.
Elahi, S., Pang, G., Ashman, R., Clancy, R., 2005. Enhanced clearance of Candida
albicans from the oral cavities of mice following oral administration of
Lactobacillus acidophilus. Clinical and Experimental Immunology 141, 29-36.

165
Elson, C.O., Ealding, W., 1984. Generalized systemic and mucosal immunity in mice
after mucosal stimulation with cholera toxin. Journal of Immunology 132, 2736-
2741.
Elson, C.O., Dertzbaugh, M.T., 1994. Mucosal adjuvants. In: Ogra, P.L., Mestecky, J.,
Lamm, M.E., Strober, W., McGhee, R., Bienenstock, J., (eds.), Handbook of
Mucosal Immunology, Academic Press, San Diego, pp. 391-402.
Erickson, K.L., Hubbard, N.E., 2000. Probiotic immunomodulation in health and
disease. Journal of Nutrition 130, 403S-409S.
FAO/WHO., 2001. Evaluation of health and nutritional properties of powder milk and
live lactic acid bacteria. Cordoba, Argentina: Food and Agriculture Organization
of the United Nations and World Health Organization Expert Consultation
Report, pp. 1-34.
FAO/WHO., 2002. Guidelines for the evaluation of probiotics in food. London, Ontario:
Food and Agriculture Organization of the United Nations and World Health
Organization Working Group Report, pp. 1-11.
Farese, R.V.Jr., Ruland, S.L., Flynn, L.M., Stokowski, R.P., Young, S.G., 1995.
Knockout of the mouse apolipoprotein B gene results in embryonic lethality in
homozygotes and protection against diet-induced hypercholesterolemia in
heterozygotes. Proceedings of the National Academy of Sciences USA 92, 1774-
1778.
Fedorak, R.N., Madsen, K.L., 2004. Probiotics and the management of inflammatory
bowel disease. Inflammatory Bowel Diseases 10, 286-299.

166
Feger, F., Varadaradjalou, S., Gao, Z., Abraham, S.N., Arock, M., 2002. The role of
mast cells in host defense and their subversion by bacterial pathogens. Trends in
Immunology 23, 151–158.
Feghali, C.A., Wright, T.M., 1997. Cytokines in acute and chronic inflammation.
Frontiers in Bioscience 2, D12-D26.
Fernandez, M.F., Boris, S., Barbes, C., 2003. Probiotic properties of human lactobacilli
strains to be used in the gastrointestinal tract. Journal of Applied Microbiology
94, 449-455.
Finlay, B.B., Falkow, S., 1997. Common themes in microbial pathogenicity revisited.
Microbiology and Molecular Biology Reviews 61, 136-169.
Fogh, J., Fogh, J.M., Orfeo, T., 1977. One hundred and twenty seven cultured tumor cell
lines producing tumors in nude mice. Journal of National Cancer Institute 59,
221-226.
Forestier, C., De Champs, C., Vatoux, C., Joly, B., 2001. Probiotic activities of
Lactobacillus casei rhamnosus: in vitro adherence to intestinal cells and
antimicrobial properties. Research in Microbiology 152, 167-173.
Fournes, B., Sadekova, S., Turbide, C., Letourneau, S., Beauchemin, N., 2001. The
CEACAM1-L Ser503 residue is crucial for inhibition of colon cancer cell
tumorigenicity. Oncogene 20, 219-230.
Francis, G.A., Fayard, E., Picard, F., Auwerx, J., 2003. Nuclear receptors and the control
of metabolism. Annual Review of Physiology 65, 261-311.
Freter, R., 1992. Factors affecting the microecology of the gut. In: Fuller, R., (ed.),
Probiotics: The scientific basis, Chapman and Hall, London, pp. 111-144.

167
Fritsch, C., Orian-Rousseaul, V., Lefebvre, O., Simon-Assmann, P., Reimund, J.M.,
Duclos, B., Kedinger, M., 1999. Characterization of human intestinal stromal cell
lines: response to cytokines and interactions with epithelial cells. Experimental
Cell Research. 248, 391-406.
Fuller, R., 1989. Probiotics in man and animals. Journal of Applied Bacteriology 66,
365-378.
Galdeano, C.M, Perdigon, G., 2006. The probiotic bacterium Lactobacillus casei induces
activation of the gut mucosal immune system through innate immunity. Clinical
and Vaccine Immunology 13, 219-226.
Garriga, M., Pascual, M., Monfort, J.M., Hugas, M., 1998. Selection of lactobacilli for
chicken probiotic adjuncts. Journal of Applied Microbiology 84, 125-132.
Garside, P., Mowat, A.M., Khoruts, A., 1999. Oral tolerance in disease. Gut 44, 137-
142.
Gate, L., Majumdar, R.S., Lunk, A., Tew, K.D., 2004. Increased myeloproliferation in
glutathione S-transferase pi-deficient mice is associated with a deregulation of
JNK and Janus kinase/STAT pathways. Journal of Biological Chemistry 279,
8608-8616.
Gibson, G.R., Beatty, E.R., Wang, X., Cummings, J.H., 1995. Selective stimulation of
bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology
108, 975-982.
Gibson, G.R., Wynne, A., Bird, A., 1999. Microflora of the intestine: role and effects.
In: Sadler, M., Caballero, B., Strain, S., (eds.), Encyclopedia of human nutrition,
Academic press, London, pp. 1282-1289.

168
Gill, H.S., Rutherfurd, K.J., 2001. Viability and dose-response studies on the effects of
the immunoenhancing lactic acid bacterium Lactobacillus rhamnosus in mice.
British Journal of Nutrition 86, 285-289.
Gill, H.S., Rutherfurd, K.J., Prasad, J., Gopal, P.K., 2000. Enhancement of natural and
acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus
acidophilus (HN017) and Bifidobacterium lactis (HN019). British Journal of
Nutrition 83, 167-176.
Gilliland, S.E., Staley, T.E., Bush, L.J., 1984. Importance of bile tolerance of
Lactobacillus acidophilus used as a dietary adjunct. Journal of Dairy Science 67,
3045-3051.
Goldin, B.R., Gorbach, S.L., 1992. Probiotics for humans. In: Fuller, R., (ed.),
Probiotics: The scientific basis, Chapman and Hall, London, pp. 355-376.
Gomez Zavaglia, A., Kociubinski, G., Perez, P., Disalvo, E., De Antoni, G., 2002. Effect
of bile on the lipid composition and surface properties of bifidobacteria. Journal
of Applied Microbiology 93, 794-799.
Goodrich, M.E., McGee, D.W., 1999. Effect of intestinal epithelial cell cytokines on
mucosal B-cell IgA secretion: enhancing effect of epithelial-derived IL-6 but not
TGF-beta on IgA+ B cells. Immunology Letters 67, 11-14.
Gopal, P.K., Prasad, J., Smart, J., Gill, H.S., 2001. In vitro adherence properties of
Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and
their antagonistic activity against an enterotoxigenic Escherichia coli.
International Journal of Food Microbiology 67, 207-216.

169
Gould, C.L., Sonnenfeld, G., 1987. Effect of treatment with interferon-gamma and
concanavalin A on the course of infection of mice with Salmonella typhimurium
strain LT-2. Journal of Interferon Research 7, 255-260.
Granato, D., Perotti, F., Masserey, I., Rouvet, M., Golliard, M., Servin, A., Brassart, D.,
1999. Cell surface-associated lipoteichoic acid acts as an adhesion factor for
attachment of Lactobacillus johnsonii La1 to human enterocyte-like Caco-2 cells.
Applied and Environmental Microbiology 65, 1071-1077.
Graness, A., Chwieralski, C.E., Reinhold, D., Thim, L., Hoffmann, W., 2002. Protein
kinase C and ERK activation are required for TFF-peptide-stimulated bronchial
epithelial cell migration and tumor necrosis factor-alpha-induced interleukin-6
(IL-6) and IL-8 secretion. Journal of Biological Chemistry 277, 18440-18446.
Greene, J.D., Klaenhammer, T.R., 1994. Factors involved in adherence of lactobacilli to
human Caco-2 cells. Applied and Environmental Microbiology 60, 4487-4494.
Guslandi, M., Mezzi, G., Sorghi, M., Testoni, P.A., 2000. Saccharomyces boulardii in
maintenance treatment of Crohn's disease. Digestive Diseases and Sciences 45,
1462-1464.
Haller, D., Bode, C., Hammes, W.P., Pfeifer, A.M., Schiffrin, E.J., Blum, S., 2000. Non-
pathogenic bacteria elicit a differential cytokine response by intestinal epithelial
cell/leucocyte co-cultures. Gut 47, 79-87.
Handley, P.S., Harty, D.W., Wyatt, J.E., Brown, C.R., Doran, J.P., Gibbs, A.C., 1987. A
comparison of the adhesion, coaggregation and cell-surface hydrophobicity
properties of fibrillar and fimbriate strains of Streptococcus salivarius. Journal of
General Microbiology 133, 3207-3217.

170
Hart, A.L., Lammers, K., Brigidi, P., Vitali, B., Rizzello, F., Gionchetti, P., Campieri,
M., Kamm, M.A., Knight, S.C., Stagg, A.J., 2004. Modulation of human
dendritic cell phenotype and function by probiotic bacteria. Gut 53, 1602-1609.
He, F., Morita, H., Kubota, A., Ouwehand, A.C., Hosoda, M., Hiramatsu, M., Kurisaki,
J., 2005. Effect of orally administered non-viable Lactobacillus cells on murine
humoral immune responses. Microbiology and Immunology 49, 993-997.
Helgeland, L., Vaage, J.T., Rolstad, B., Midtvedt, T., Brandtzaeg, P., 1996. Microbial
colonization influences composition and T-cell receptor V beta repertoire of
intraepithelial lymphocytes in rat intestine. Immunology 89, 494-501.
Herias, M.V., Hessle, C., Telemo, E., Midtvedt, T., Hanson, L.A., Wold, A.E., 1999.
Immunomodulatory effects of Lactobacillus plantarum colonizing the intestine of
gnotobiotic rats. Clinical and Experimental Immunology 116, 283-290.
Hessle, C., Hanson, L.A., Wold, A.E., 1999. Lactobacilli from human gastrointestinal
mucosa are strong stimulators of IL-12 production. Clinical and Experimental
Immunology 116, 276-282.
Hogerkorp, C.M., Bilke, S., Breslin, T., Ingvarsson, S., Borrebaeck, C.A., 2003. CD44-
stimulated human B cells express transcripts specifically involved in
immunomodulation and inflammation as analyzed by DNA microarrays. Blood
101, 2307-2313.
Holzapfel, W.H., Haberer, P., Snel, J., Schillinger, U., Huis in't Veld, J.H., 1998.
Overview of gut flora and probiotics. International Journal of Food Microbiology
41, 85-101.

171
Hooper, L.V., Wong, M.H., Thelin, A., Hansson, L., Falk, P.G., Gordon, J.I., 2001.
Molecular analysis of commensal host-microbial relationships in the intestine.
Science 291, 881-884.
Hornqvist, E., Goldschmidt, T.J., Holmdahl, R., Lycke, N., 1991. Host defense against
cholera toxin is strongly CD4+ T cell dependent. Infection and Immunity 59,
3630-3638.
Hosono, A., Ozawa, A., Kato, R., Ohnishi, Y., Nakanishi, Y., Kimura, T., Nakamura, R.,
2003. Dietary fructooligosaccharides induce immunoregulation of intestinal IgA
secretion by murine Peyer's patch cells. Bioscience, Biotechnology, and
Biochemistry 67, 758-764.
Hu, M.C., Qiu, W.R., Wang, X., Meyer, C.F., Tan, T.H., 1996. Human HPK1, a novel
human hematopoietic progenitor kinase that activates the JNK/SAPK kinase
cascade. Genes & Development 10, 2251-2264.
Hudault, S., Lievin, V., Bernet-Camard, M.F., Servin, A.L., 1997. Antagonistic activity
exerted in vitro and in vivo by Lactobacillus casei (strain GG) against
Salmonella typhimurium C5 infection. Applied and Environmental Microbiology
63, 513-518.
Huggins, K.W., Camarota, L.M., Howles, P.N., Hui, D.Y., 2003. Pancreatic triglyceride
lipase deficiency minimally affects dietary fat absorption but dramatically
decreases dietary cholesterol absorption in mice. Journal of Biological Chemistry
278, 42899-42905.
Husband, A.J., Beagley, K.W., McGhee, J.R., 1999. Mucosal cytokines. In: Ogra, P.L.,
Mestecky, J., Lamm, M.E., Strober, W., Bienenstock, J., McGhee, J.R., (eds.),
Mucosal Immunology, Academic Press, New York, pp. 541-557.

172
Hutcheson, I.R., Whittle, B.J., Boughton-Smith, N.K., 1990. Role of nitric oxide in
maintaining vascular integrity in endotoxin-induced acute intestinal damage in
the rat. British Journal of Pharmacology 101, 815-820.
Iijima, H., Neurath, M.F., Nagaishi, T., Glickman, J.N., Nieuwenhuis, E.E., Nakajima,
A., Chen, D., Fuss, I.J., Utku, N., Lewicki, D.N., Becker, C., Gallagher, T.M.,
Holmes, K.V., Blumberg, R.S., 2004. Specific Regulation of T Helper Cell 1-
mediated Murine Colitis by CEACAM1. Journal of Experimental Medicine 199,
471-482.
Ikeda, I., Matsuoka, R., Hamada, T., Mitsui, K., Imabayashi, S., Uchino, A., Sato, M.,
Kuwano, E., Itamura, T., Yamada, K., Tanaka, K., Imaizumi, K., 2002.
Cholesterol esterase accelerates intestinal cholesterol absorption. Biochimica et
Biophysica Acta 1571, 34-44.
Isolauri, E., Juntunen, M., Rautanen, T., Sillanaukee, P., Koivula, T., 1991. A human
Lactobacillus strain (Lactobacillus casei sp strain GG) promotes recovery from
acute diarrhea in children. Pediatrics 88, 90-97.
Isolauri, E., Kaila, M., Mykkanen, H., Ling, W.H., Salminen, S., 1994. Oral
bacteriotherapy for viral gastroenteritis. Digestive Diseases and Sciences 39,
2595-2600.
Isolauri, E., Kirjavainen, P.V., Salminen, S., 2002. Probiotics: a role in the
treatment of intestinal infection and inflammation?. Gut 50(Suppl III), 54-59.
Isolauri, E., Sutas, Y., Kankaanpaa, P., Arvilommi, H., Salminen, S., 2001. Probiotics:
effects on immunity. American Journal of Clinical Nutrition 73, 444S-450S.
Jacobsen, C.N., Rosenfeldt Nielsen, V., Hayford, A.E., Moller, P.L., Michaelsen, K.F.,
Paerregaard, A., Sandstrom, B., Tvede, M., Jakobsen, M., 1999. Screening of

173
probiotic activities of forty-seven strains of Lactobacillus spp. by in vitro
techniques and evaluation of the colonization ability of five selected strains in
humans. Applied and Environmental Microbiology 65, 4949-4956.
Jankovic, I., Ventura, M., Meylan, V., Rouvet, M., Elli, M., Zink, R., 2003. Contribution
of aggregation-promoting factor to maintenance of cell shape in Lactobacillus
gasseri 4B2. Journal of Bacteriology 185, 3288-3296.
Jiang, H.Q., Thurnheer, M.C., Zuercher, A.W., Boiko, N.V., Bos, N.A., Cebra, J.J.,
2004. Interactions of commensal gut microbes with subsets of B- and T-cells in
the murine host. Vaccine 22, 805-811.
Johnson, L.R., 1977. Regulation of gastric secretion. In: Johnson, L.R., (ed.),
Gastrointestinal Physiology, CV Mosby., St. Louis, pp. 62-69.
Kabir, A.M., Aiba,Y., Takagi, A., Kamiya, S., Miwa, T., Koga, Y., 1997. Prevention of
Helicobacter pylori infection by lactobacilli in a gnotobiotic murine model. Gut
41, 49-55.
Kaila, M., Isolauri, E., Soppi, E., Virtanen, E., Laine, S., Arvilommi, H., 1992.
Enhancement of the circulating antibody secreting cell response in human
diarrhea by a human Lactobacillus strain. Pediatric Research 32, 141-144.
Kalliomaki, M., Salminen, S., Arvilommi, H., Kero, P., Koskinen, P., Isolauri, E., 2001.
Probiotics in primary prevention of atopic disease: a randomised placebo-
controlled trial. Lancet 357, 1076-1079.
Kammerer, R., Stober, D., Singer, B.B., Obrink, B., Reimann, J., 2001.
Carcinoembryonic antigen-related cell adhesion molecule 1 on murine dendritic
cells is a potent regulator of T cell stimulation. Journal of Immunology 166,
6537-6544.

174
Kaneda, A., Kaminishi, M., Yanagihara, K., Sugimura, T., Ushijima, T., 2002.
Identification of Silencing of Nine Genes in Human Gastric Cancers. Cancer
Research 62, 6645-6650.
Kato, I., Tanaka, K., Yokokura, T., 1999. Lactic acid bacterium potently induces the
production of interleukin-12 and IFN-gamma by mouse splenocytes.
International Journal of Immunopharmacology 21, 121-131.
Keele, C.A., Neil, E., 1965. Secretion of digestive juices. In. Wright, S., (ed.), Applied
Physiology, Oxford University Press, London, pp. 353-363.
Khan, J., Bittner, M.L., Chen, Y., Meltzer, P.S., Trent, J.M., 1999. DNA microarray
technology: the anticipated impact on the study of human disease. Biochimica et
Biophysica Acta 1423, M17-M28.
Kidd, P., 2003. Th1/Th2 balance: the hypothesis, its limitations, and implications for
health and disease. Alternative Medicine Review 8, 223-246.
Kim, H., Kwack, K., Kim, D.Y., Ji, G.E., 2005. Oral probiotic bacterial administration
suppressed allergic responses in an ovalbumin-induced allergy mouse model.
FEMS Immunology and Medical Microbiology 45, 259-267.
Kimoto, H., Kurisaki, J., Tsuji, N.M., Ohmomo, S., Okamoto, T., 1999. Lactococci as
probiotic strains: adhesion to human enterocyte-like Caco-2 cells and tolerance
to low pH and bile. Letters in Applied Microbiology 29, 313-316.
Kirby, R.J., Zheng, S., Tso, P., Howles, P.N., Hui, D.Y., 2002. Bile salt-stimulated
carboxyl ester lipase influences lipoprotein assembly and secretion in intestine: a
process mediated via ceramide hydrolysis. Journal of Biological Chemistry 277,
4104-4109.

175
Kirjavainen, P.V., El-Nezami, H.S., Salminen, S.J., Ahokas, J.T., Wright, P.F., 1999.
The effect of orally administered viable probiotic and dairy lactobacilli on mouse
lymphocyte proliferation. FEMS Immunology and Medical Microbiology 26,
131-135.
Kirjavainen, P.V., Salminen, S.J., Isolauri, E., 2003. Probiotic bacteria in the
management of atopic disease: underscoring the importance of viability. Journal
of Pediatric Gastroenterology and Nutrition 36, 223- 227.
Klein, G., Pack, A., Bonaparte, C., Reuter, G., 1998. Taxonomy and physiology of
probiotic lactic acid bacteria. International Journal of Food Microbiology 41,
103-125.
Kmet, V., Lucchini, F., 1997. Aggregation-promoting factor in human vaginal
Lactobacillus strains. FEMS Immunology and Medical Microbiology 19, 111-
114.
Koebnick, C., Wagner, I., Leitzmann, P., Stern, U., Zunft, H.J., 2003. Probiotic beverage
containing Lactobacillus casei Shirota improves gastrointestinal symptoms in
patients with chronic constipation. Canadian Journal of Gastroenterology 17,
655– 659.
Kohler, H., McCormick, B.A., Walker, W.A., 2003. Bacterial-enterocyte crosstalk:
cellular mechanisms in health and disease. Journal of Pediatric Gastroenterology
and Nutrition 36, 175-185.
Kolenbrander, P.E., 2000. Oral microbial communities: biofilms, interactions, and
genetic systems. Annual Review of Microbiology 54, 413-437.
Korhonen, R., Korpela, R., Saxelin, M., Maki, M., Kankaanranta, H., Moilanen, E.,
2001. Induction of nitric oxide synthesis by probiotic Lactobacillus rhamnosus

176
GG in J774 macrophages and human T84 intestinal epithelial cells. Inflammation
25, 223-232.
Kos, B., Suskovic, J., Vukovic, S., Simpraga, M., Frece, J., Matosic, S., 2003. Adhesion
and aggregation ability of probiotic strain Lactobacillus acidophilus M92.
Journal of Applied Microbiology 94, 981-987.
Lamine, F., Fioramonti, J., Bueno, L., Nepveu, F., Cauquil, E., Lobysheva, I., Eutamene,
H., Theodorou, V., 2004. Nitric oxide released by Lactobacillus farciminis
improves TNBS-induced colitis in rats. Scandinavian Journal of
Gastroenterology 39, 37-45.
Lankaputhra, W.E.V., Shah, N.P., 1995. Survival of Lactobacillus acidophilus and
Bifidobacterium spp in the presence of acid and bile salts. Cultured Dairy
Products Journal 30, 2-7.
Leblanc, J., Fliss, I., Matar, C., 2004. Induction of a humoral immune response
following an Escherichia coli O157:H7 infection with an immunomodulatory
peptidic fraction derived from Lactobacillus helveticus-fermented milk. Clinical
and Diagnostic Laboratory Immunology 11, 1171-1181.
Lebman, D.A., Lee, F.D., Coffman, R.L., 1990. Mechanism for transforming growth
factor beta and IL-2 enhancement of IgA expression in lipopolysaccharide-
stimulated B cell cultures. Journal of Immunology 144, 952-959.
Lee, Y.K., Puong, K.Y., Ouwehand, A.C., Salminen, S., 2003. Displacement of bacterial
pathogens from mucus and Caco-2 cell surface by lactobacilli. Journal of
Medical Microbiology 52, 925-930.
Lefer, A.M., Lefer, D.J., 1999. Nitric oxide. II. Nitric oxide protects in intestinal
inflammation. American Journal of Physiology 276, G572-G575.

177
Leng, L., Metz, C.N., Fang, Y., Xu, J., Donnelly, S., Baugh, J., Delohery, T., Chen, Y.,
Mitchell, R.A., Bucala, R., 2003. MIF signal transduction initiated by binding to
CD74. Journal of Experimental Medicine 197, 1467-1476.
Lilly, D.M., Stillwell, R.H., 1965. Probiotics: growth-promoting factors produced by
microorganisms. Science 147, 747-748.
Linsalata, M., Russo, F., Berloco, P., Caruso, M.L., Matteo, G.D., Cifone, M.G.,
Simone, C.D., Ierardi, E., Di Leo, A., 2004. The influence of Lactobacillus
brevis on ornithine decarboxylase activity and polyamine profiles in Helicobacter
pylori-infected gastric mucosa. Helicobacter 9, 165– 172.
Liu, Y.J., Arpin, C., 1997. Germinal center development. Immunological Reviews 156,
111-126.
Lu, L., Walker, W.A., 2001. Pathologic and physiologic interactions of bacteria with the
gastrointestinal epithelium. American Journal of Clinical Nutrition 73, 1124S-
1130S.
Lund, B., Edlund, C., 2001. Probiotic Enterococcus faecium strain is a possible recipient
of the vanA gene cluster. Clinical Infectious Diseases 32, 1384-1385.
Luyer, M.D., Buurman, W.A., Hadfoune, M., Speelmans, G., Knol, J., Jacobs, J.A.,
Dejong, C.H., Vriesema, A.J., Greve, J.W., 2005. Strain-specific effects of
probiotics on gut barrier integrity following hemorrhagic shock. Infection and
Immunity 73, 3686-3692.
Lycke, N., Lindholm, L., Holmgren, J., 1985. Cholera antibody production in vitro by
peripheral blood lymphocytes following oral immunization of humans and mice.
Clinical and Experimental Immunology 62, 39-47.

178
Ma, D., Forsythe, P., Bienenstock, J., 2004. Live Lactobacillus reuteri is essential for the
inhibitory effect on tumor necrosis factor alpha-induced interleukin-8 expression.
Infection and Immunity 72, 5308-5314.
Ma, L., Deitch, E., Specian, R., Steffen, E., Berg, R., 1990. Translocation of
Lactobacillus murinus from the gastrointestinal tract. Current Microbiology 20,
177-184.
Maassen, C.B., van Holten-Neelen, C., Balk, F., den Bak-Glashouwer, M.J., Leer, R.J.,
Laman, J.D., Boersma, W.J., Claassen, E., 2000. Strain-dependent induction of
cytokine profiles in the gut by orally administered Lactobacillus strains. Vaccine
18, 2613-2623.
Mack, D.R., Michail, S., Wei, S., McDougall, L., Hollingsworth, M.A., 1999. Probiotics
inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin
gene expression. American Journal of Physiology 276, G941-G950.
Macpherson, A.J., Hunziker, L., McCoy, K., Lamarre, A., 2001. IgA responses in the
intestinal mucosa against pathogenic and non-pathogenic microorganisms.
Microbes and Infection 3, 1021-1035.
Macpherson, A.J., Uhr, T., 2004. Induction of protective IgA by intestinal dendritic cells
carrying commensal bacteria. Science 303, 1662-1665.
Madden, J.A.J., Plummer, S., Sen, S., Dear, K., Tarry, S., Hunter, J.O., 2001.
Comparison of the caecal and faecal microflora of healthy subjects and patients
with irritable bowel syndrome (IBS). Gut 48(Suppl 1), A58.
Madsen, K., Cornish, A., Soper, P., McKaigney, C., Jijon, H., Yachimec, C., Doyle, J.,
Jewell, L., De Simone, C., 2001. Probiotic bacteria enhance murine and human
intestinal epithelial barrier function. Gastroenterology 121, 580-591.

179
Madsen, K.L., 2001. The use of probiotics in gastrointestinal disease. Canadian Journal
of Gastroenterology 15, 817-822.
Madsen, K.L., Doyle, J.S., Jewell, L.D., Tavernini, M.M., Fedorak, R.N., 1999.
Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice.
Gastroenterology 116, 1107-1114.
Magnusson, K.E., Stjernstrom, I., 1982. Mucosal barrier mechanisms. Interplay between
secretory IgA (SIgA), IgG and mucins on the surface properties and association
of salmonellae with intestine and granulocytes. Immunology 45, 239-248.
Mahoney, M., Henriksson, A., 2003. The effect of processed meat and meat starter
cultures on gastrointestinal colonization and virulence of Listeria monocytogenes
in mice. International Journal of Food Microbiology 84, 255-261.
Majamaa, H., Isolauri, E., 1997. Probiotics: a novel approach in the management of food
allergy. Journal of Allergy and Clinical Immunology 99, 179-185.
Majamaa, H., Isolauri, E., Saxelin, M., Vesikari, T., 1995. Lactic acid bacteria in the
treatment of acute rotavirus gastroenteritis. Journal of Pediatric Gastroenterology
and Nutrition 20, 333-338.
Malin, M., Suomalainen, H., Saxelin, M., Isolauri, E., 1996. Promotion of IgA immune
response in patients with Crohn's disease by oral bacteriotherapy with
Lactobacillus GG. Annals of Nutrition and Metabolism 40, 137-145.
Mallett, A.K., Bearne, C.A., Rowland, I.R., 1989. The influence of incubation pH on the
activity of rat and human gut flora enzymes. Journal of Applied Bacteriology 66,
433-437.

180
Marin, M.L., Benito, Y., Pin, C., Fernandez, M.F., Garcia, M.L., Selgas, M.D., Casas,
C., 1997. Lactic acid bacteria: hydrophobicity and strength of attachment to meat
surfaces. Letters in Applied Microbiology 24, 14-18.
Marin, M.L., Tejada-Simon, M.V., Lee, J.H., Murtha, J., Ustunol, Z., Pestka, J.J., 1998.
Stimulation of cytokine production in clonal macrophage and T-cell models by
Streptococcus thermophilus: comparison with Bifidobacterium sp. and
Lactobacillus bulgaricus. Journal of Food Protection 61, 859–864.
Mashimo, H., Kjellin, A., Goyal, R.K., 2000. Gastric stasis in neuronal nitric oxide
synthase-deficient knockout mice. Gastroenterology 119, 766-773.
Mastromarino, P., Brigidi, P., Macchia, S., Maggi, L., Pirovano, F., Trinchieri, V.,
Conte, U., Matteuzzi, D., 2002. Characterization and selection of vaginal
Lactobacillus strains for the preparation of vaginal tablets. Journal of Applied
Microbiology 93, 884-893.
Matsumoto, S., Watanabe, N., Imaoka, A., Okabe, Y., 2001. Preventive effects of
Bifidobacterium- and Lactobacillus-fermented milk on the development of
inflammatory bowel disease in senescence-accelerated mouse P1/Yit strain mice.
Digestion 64, 92-99.
Matsuzaki, T., Yamazaki, R., Hashimoto, S., Yokokura, T., 1998. The effect of oral
feeding of Lactobacillus casei strain Shirota on immunoglobulin E production in
mice. Journal of Dairy Science 81, 48–53.
Mayra-Makinen, A., Manninen, M., Gyllenberg, H., 1983. The adherence of lactic acid
bacteria to the columnar epithelial cells of pigs and calves. Journal of Applied
Bacteriology 55, 241-245.

181
Mazmanian, S.K., Liu, C.H., Tzianabos, A.O., Kasper, D.L., 2005. An
immunomodulatory molecule of symbiotic bacteria directs maturation of the host
immune system. Cell 122, 107-118.
McCafferty, D.M., Mudgett, J.S., Swain, M.G., Kubes, P., 1997. Inducible nitric oxide
synthase plays a critical role in resolving intestinal inflammation.
Gastroenterology 112, 1022-1027.
McCracken, V.J., Gaskins, H.R., 1999. Probiotics and the immune system. In: Tannock,
G.W., (ed.), Probiotics: A Critical Review, Horizon Scientific Press, England,
pp. 85-112.
McCracken, V.J., Lorenz, R.G., 2001. The gastrointestinal ecosystem: a precarious
alliance among epithelium, immunity and microbiota. Cellular Microbiology 3,
1-11.
McGhee, J.R., Lamm, M.E., Strober, W., 1999. Mucosal immune responses: An
overview. In: Ogra, P.L., Mestecky, J., Lamm, M.E., Strober, W., Bienenstock,
J., McGhee, J.R., (eds.), Mucosal Immunology, Academic Press, San Diego, pp.
485-506.
Medici, M., Vinderola, C.G., Weill, R., Perdigon, G., 2005. Effect of fermented milk
containing probiotic bacteria in the prevention of an enteroinvasive Escherichia
coli infection in mice. Journal of Dairy Research 72, 243-249.
Mestecky, J., McGhee, J.R., 1987. Immunoglobulin A (IgA): Molecular and cellular
interactions involved in IgA biosynthesis and immune response. Advances in
Immunology 40,153-245.
Mestecky, J., Russell, M.W., Elson, C.O., 1999. Intestinal IgA: novel views on its
function in the defence of the largest mucosal surface. Gut 44, 2-5.

182
Midolo, P.D., Lambert, J.R., Hull, R., Luo, F., Grayson, M.L., 1995. In vitro inhibition
of Helicobacter pylori NCTC 11637 by organic acids and lactic acid bacteria.
Journal of Applied Bacteriology 79, 475-479.
Miettinen, M., Matikainen, S., Vuopio-Varkila, J., Pirhonen, J., Varkila, K., Kurimoto,
M., Julkunen, I., 1998. Lactobacilli and streptococci induce interleukin-12 (IL-
12), IL-18 and gamma interferon production in human peripheral blood
mononuclear cells. Infection and Immunity 66, 6058–6062.
Miettinen, M., Vuopio-Varkila, J., Varkila, K., 1996. Production of human tumor
necrosis factor alpha, interleukin-6, and interleukin-10 is induced by lactic acid
bacteria. Infection and Immunity 64, 5403-5405.
Mimura, T., Rizzello, F., Helwig, U., Poggioli, G., Schreiber, S., Talbot, I.C., Nicholls,
R.J., Gionchetti, P., Campieri, M., Kamm, M.A., 2004. Once daily high dose
probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory
pouchitis. Gut 53, 108– 114.
Miraglia del Giudice, M., De Luca, M.G., 2004. The role of probiotics in the clinical
management of food allergy and atopic dermatitis. Journal of Clinical
Gastroenterology 38, S84-S85.
Mishra, V., Prasad, D.N., 2005. Application of in vitro methods for selection of
Lactobacillus casei strains as potential probiotics. International Journal of Food
Microbiology 103, 109-115.
Moncada, S., Higgs, E.A., 1995. Molecular mechanisms and therapeutic strategies
related to nitric oxide. FASEB Journal 9, 1319-1330.
Moreau, M.C., Ducluzeau, R., Guy-Grand, D., Muller, M.C., 1978. Increase in the
population of duodenal immunoglobulin A plasmocytes in axenic mice

183
associated with different living or dead bacterial strains of intestinal origin.
Infection and immunity 21, 532-539.
Morita, H., He, F., Fuse, T., Ouwehand, A.C., Hashimoto, H., Hosoda, M., Mizumachi,
K., Kurisaki, J., 2002. Adhesion of lactic acid bacteria to Caco-2 cells and their
effect on cytokine secretion. Microbiology and Immunology 46, 293-297.
Mourad, F.H., Turvill, J.L., Farthing, M.J., 1999. Role of nitric oxide in intestinal water
and electrolyte transport. Gut 44, 143-147.
Myllyluoma, E., Veijola, L., Ahlroos, T., Tynkkynen, S., Kankuri, E., Vapaatalo, H.,
Rautelin, H., Korpela, R., 2005. Probiotic supplementation improves tolerance to
Helicobacter pylori eradication therapy--a placebo-controlled, double-blind
randomized pilot study. Alimentary Pharmacology and Therapeutics 21, 1263-
1272.
Nader de Macias, M.E., Apella, M.C., Romero, N.C., Gonzalez, S.N., Oliver, G., 1992.
Inhibition of Shigella sonnei by Lactobacillus casei and Lactobacillus
acidophilus. Journal of Applied Bacteriology 73, 407-411.
Nagle, D.L., McGrail, S.H., Vitale, J., Woolf, E.A., Dussault, B.J.Jr., DiRocco, L.,
Holmgren, L., Montagno, J., Bork, P., Huszar, D., Fairchild-Huntress, V., Ge, P.,
Keilty, J., Ebeling, C., Baldini, L., Gilchrist, J., Burn, P., Carlson, G.A., Moore,
K.J., 1999. The mahogany protein is a receptor involved in suppression of
obesity. Nature 398, 148-152.
Neutra, M.R., Frey, A., Kraehenbuhl, J.P., 1996. Epithelial M cells: gateways for
mucosal infection and immunization. Cell 86, 345-348.

184
Nicaise, P., Gleizes, A., Sandre, C., Forestier, F., Kergot, R., Quero, A.M., Labarre, C.,
1998. Influence of intestinal microflora on murine bone marrow and spleen
macrophage precursors. Scandinavian Journal of Immunology 48, 585-591.
Niedzielin, K., Kordecki, H., Birkenfeld, B., 2001. A controlled, double-blind,
randomized study on the efficacy of Lactobacillus plantarum 299v in patients
with irritable bowel syndrome. European Journal of Gastroenterology and
Hepatology 13, 1143-1147.
Nobaek, S., Johansson, M.L., Molin, G., Ahrne, S., Jeppsson, B., 2000. Alteration of
intestinal microflora is associated with reduction in abdominal bloating and pain
in patients with irritable bowel syndrome. American Journal of Gastroenterology
95, 1231-1238.
Numata, A., Minagawa, T., Asano, M., Nakane, A., Katoh, H., Tanabe, T., 1991.
Functional evaluation of tumor-infiltrating mononuclear cells. Detection of
endogenous interferon-gamma and tumor necrosis factor-alpha in human
colorectal adenocarcinomas. Cancer 68, 1937-1943.
O'Hara, A.M., Shanahan, F., 2006. The gut flora as a forgotten organ. EMBO Reports 7,
688-693.
Oien, K.A., Vass, J.K., Downie, I., Fullarton, G., Keith, W.N., 2003. Profiling,
comparison and validation of gene expression in gastric carcinoma and normal
stomach. Oncogene 22, 4287-4300.
O'Mahony, L., McCarthy, J., Kelly, P., Hurley, G., Luo, F., Chen, K., O'Sullivan, G.C.,
Kiely, B., Collins, J.K., Shanahan, F., Quigley, E.M., 2005. Lactobacillus and
bifidobacterium in irritable bowel syndrome: symptom responses and
relationship to cytokine profiles. Gastroenterology 128, 541-551.

185
Oozeer, R., Mater, D.D., Goupil-Feuillerat, N., Corthier, G., 2004. Initiation of protein
synthesis by a labeled derivative of the Lactobacillus casei DN-114 001 strain
during transit from the stomach to the cecum in mice harboring human
microbiota. Applied and Environmental Microbiology 70, 6992-6997.
Otero, M.C., Morelli, L., Nader-Macías, M.E., 2006. Probiotic properties of vaginal
lactic acid bacteria to prevent metritis in cattle. Letters in Applied Microbiology
43, 91-97.
Ouellette, A.J., 1999. IV. Paneth cell antimicrobial peptides and the biology of the
mucosal barrier. American Journal of Physiology 277, G257-G261.
Ouwehand, A.C., Tuomola, E.M., Tolkko, S., Salminen, S., 2001. Assessment of
adhesion properties of novel probiotic strains to human intestinal mucus.
International Journal of Food Microbiology 64, 119-126.
Owen, R.L., 1999. Uptake and transport of intestinal macromolecules and
microorganisms by M cells in Peyer's patches - a personal and historical
perspective. Seminars in Immunology 11, 157-163.
Pan, W.H., Li, P.L., Liu, Z., 2006. The correlation between surface hydrophobicity and
adherence of Bifidobacterium strains from centenarians' faeces. Anaerobe 12,
148-152.
Pathmakanthan, S., Thornley, J.P., Hawkey, C.J., 1999. Mucosally associated bacterial
flora of the human colon: Quantitative and species specific differences between
normal and inflamed colonic biopsies. Microbial Ecology in Health and Disease
11, 169-174.

186
Paton, A.W., Jennings, M.P., Morona, R., Wang, H., Focareta, A., Roddam, L.F., Paton,
J.C., 2005. Recombinant probiotics for treatment and prevention of
enterotoxigenic Escherichia coli diarrhea. Gastroenterology 128, 1219-1228.
Payne, D., Kubes, P., 1993. Nitric oxide donors reduce the rise in reperfusion-induced
intestinal mucosal permeability. American Journal of Physiology 265, G189-
G195.
Pedersen, K., Tannock, G.W., 1989. Colonization of the porcine gastrointestinal tract by
lactobacilli. Applied and Environmental Microbiology 55, 279-283.
Pelletier, C., Bouley, C., Cayuela, C., Bouttier, S., Bourlioux, P., Bellon-Fontaine, M.N.,
1997. Cell surface characteristics of Lactobacillus casei subsp. casei,
Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains.
Applied and Environmental Microbiology 63, 1725-1731.
Pelto, L., Isolauri, E., Lilius, E.M., Nuutila, J., Salminen, S., 1998. Probiotic bacteria
down-regulate the milk-induced inflammatory response in milk-hypersensitive
subjects but have an immunostimulatory effect in healthy subjects. Clinical &
Experimental Allergy 28, 1474-1479.
Perdigon, G., Alvarez, S., 1992. Bacterial interactions in the gut. In: Fuller, R., (ed.),
Probiotics: The scientific basis, Chapman and Hall, London, pp. 146-180.
Perdigon, G., Alvarez, S., Rachid, M., Aguero, G., Gobbato, N., 1995. Immune system
stimulation by probiotics. Journal of Dairy Science 78, 1597-1606.
Perdigon, G., de Macias, M.E., Alvarez, S., Oliver, G., de Ruiz Holgado, A.A., 1986.
Effect of perorally administered lactobacilli on macrophage activation in mice,
Infection and Immunity 53, 404-410.

187
Perdigon, G., Fuller, R., Raya, R., 2001. Lactic acid bacteria and their effect on the
immune system. Current Issues in Intestinal Microbiology 2, 27-42.
Perdigon, G., Maldonado Galdeano, C., Valdez, J.C., Medici, M., 2002. Interaction of
lactic acid bacteria with the gut immune system. European Journal of Clinical
Nutrition 56, S21-S26.
Perdigon, G., Nader de Macias, M.E., Alvarez, S., Oliver, G., Pesce de Ruiz Holgado,
A.A., 1990. Prevention of gastrointestinal infection using immunobiological
methods with milk fermented with Lactobacillus casei and Lactobacillus
acidophilus. Journal of Dairy Research 57, 255-264.
Perdigon, G., Rachid, M., De Budeguer, M.V., Valdez, J.C., 1994. Effect of yogurt
feeding on the small and large intestine associated lymphoid cells in mice.
Journal of Dairy Research 61, 553-562.
Perdigon, G., Vintini, E., Alvarez, S., Medina, M., Medici, M., 1999. Study of the
possible mechanisms involved in the mucosal immune system activation by
lactic acid bacteria. Journal of Dairy Science 82, 1108-1114.
Perdue, M.H., 1999. Mucosal immunity and inflammation. III. The mucosal antigen
barrier: cross talk with mucosal cytokines. American Journal of Physiology 277,
G1-G5.
Perez, P.F., Minnaard, Y., Disalvo, E.A., De Antoni, G.L., 1998. Surface properties of
bifidobacterial strains of human origin. Applied and Environmental
Microbiology 64, 21-26.
Petrault, I., Zimowska, W., Mathieu, J., Bayle, D., Rock, E., Favier, A., Rayssiguier, Y.,
Mazur, A., 2002. Changes in gene expression in rat thymocytes identified by

188
cDNA array support the occurrence of oxidative stress in early magnesium
deficiency. Biochimica et Biophysica Acta 1586, 92-98.
Petrof, E.O., Kojima, K., Ropeleski, M.J., Musch, M.W., Tao, Y., De Simone, C.,
Chang, E.B., 2004. Probiotics inhibit nuclear factor-kappaB and induce heat
shock proteins in colonic epithelial cells through proteasome inhibition.
Gastroenterology 127, 1474-1487.
Pidcock, K., Heard, G.M., Henriksson, A., 2002. Application of nontraditional meat
starter cultures in production of Hungarian salami. International Journal of Food
Microbiology 76, 75-81.
Plant, L.J., Conway, P.L., 2002. Adjuvant properties and colonization potential of
adhering and non-adhering Lactobacillus spp following oral administration to
mice. FEMS Immunology and Medical Microbiology 34, 105-111.
Plummer, S., Weaver, M.A., Harris, J.C., Dee, P., Hunter, J., 2004. Clostridium difficile
pilot study: effects of probiotic supplementation on the incidence of C. difficile
diarrhoea. International Microbiology 7, 59-62.
Prasad, J., Gill, H., Smart, J., Gopal, P.K., 1998. Selection and Characterisation of
Lactobacillus and Bifidobacterium Strains for Use as Probiotics. International
Dairy Journal 8, 993-1002.
Puente Navazo, M.D., Valmori, D., Ruegg, C., 2001. The alternatively spliced domain
TnFnIII A1A2 of the extracellular matrix protein tenascin-C suppresses
activation-induced T lymphocyte proliferation and cytokine production. Journal
of Immunology 167, 6431-6440.
Pulido, R., Krueger, N.X., Serra-Pages, C., Saito, H., Streuli, M., 1995. Molecular
characterization of the human transmembrane protein-tyrosine phosphatase delta.

189
Evidence for tissue-specific expression of alternative human transmembrane
protein-tyrosine phosphatase delta isoforms. Journal of Biological Chemistry
270, 6722-6728.
Racedo, S., Villena, J., Medina, M., Aguero, G., Rodriguez, V., Alvarez, S., 2006.
Lactobacillus casei administration reduces lung injuries in a Streptococcus
pneumoniae infection in mice. Microbes and Infection 8, 2359-2366.
Rakoff-Nahoum, S., Paglino, J., Eslami-Varzaneh, F., Edberg, S., Medzhitov, R., 2004.
Recognition of commensal microflora by toll-like receptors is required for
intestinal homeostasis. Cell 118, 229-241.
Rao, L., Puschner, B., Prolla, T.A., 2001. Gene expression profiling of low selenium
status in the mouse intestine: transcriptional activation of genes linked to DNA
damage, cell cycle control and oxidative stress. Journal of Nutrition 131, 3175-
3181.
Rayes, N., Seehofer, D., Hansen, S., Boucsein, K., Muller, A.R., Serke, S., Bengmark,
S., Neuhaus, P., 2002. Early enteral supply of lactobacillus and fiber versus
selective bowel decontamination: a controlled trial in liver transplant recipients.
Transplantation 74, 123– 127.
Reid, G., 2005. Predictors of urinary tract infection after menopause: a prospective
study. American Journal of Medicine 118, 930-931.
Reid, G., Burton, J., Hammond, J.A., Bruce, A.W., 2004. Nucleic acid-based diagnosis
of bacterial vaginosis and improved management using probiotic lactobacilli.
Journal of Medicinal Food 7, 223-228.
Reid, G., Kim, S.O., Kohler, G.A., 2006. Selecting, testing and understanding probiotic
microorganisms. FEMS Immunology and Medical Microbiology 46, 149-157.

190
Reid, G., McGroarty, J.A., Angotti, R., Cook, R.L., 1988. Lactobacillus inhibitor
production against Escherichia coli and coaggregation ability with uropathogens.
Canadian Journal of Microbiology 34, 344-351.
Reid, G., Sanders, M.E., Gaskins, H.R., Gibson, G.R., Mercenier, A., Rastall, R.,
Roberfroid, M., Rowland, I., Cherbut, C., Klaenhammer, T.R., 2003. New
scientific paradigms for probiotics and prebiotics. Journal of Clinical
Gastroenterology 37, 105-118.
Rescigno, M., Urbano, M., Valzasina, B., Francolini, M., Rotta, G., Bonasio, R.,
Granucci, F., Kraehenbuhl, J.P., Ricciardi-Castagnoli, P., 2001. Dendritic cells
express tight junction proteins and penetrate gut epithelial monolayers to sample
bacteria. Nature Immunology 2, 361-367.
Rhee, J., Takahashi, Y., Saga, Y., Wilson-Rawls, J., Rawls, A., 2003. The protocadherin
papc is involved in the organization of the epithelium along the segmental border
during mouse somitogenesis. Developmental Biology 254, 248-261.
Rickard, A.H., Gilbert, P., High, N.J., Kolenbrander, P.E., Handley, P.S., 2003. Bacterial
coaggregation: an integral process in the development of multi-species biofilms.
Trends in Microbiology 11, 94-100.
Rojas, M., Conway, P.L., 1996. Colonization by lactobacilli of piglet small intestinal
mucus. Journal of Applied Bacteriology 81, 474-480.
Roller, M., Pietro Femia, A., Caderni, G., Rechkemmer, G., Watzl, B., 2004. Intestinal
immunity of rats with colon cancer is modulated by oligofructose-enriched inulin
combined with Lactobacillus rhamnosus and Bifidobacterium lactis. British
Journal of Nutrition 92, 931-938.

191
Ronner, U., Husmark, U., Henriksson, A., 1990. Adhesion of bacillus spores in
relation to hydrophobicity Journal of Applied Bacteriology 69, 550-556.
Roos, S., Lindgren, S., Jonsson, H., 1999. Autoaggregation of Lactobacillus reuteri is
mediated by a putative DEAD-box helicase. Molecular Microbiology 32, 427-
436.
Rosenfeldt, V., Benfeldt, E., Valerius, N.H., Paerregaard, A., Michaelsen, K.F., 2004.
Effect of probiotics on gastrointestinal symptoms and small intestinal
permeability in children with atopic dermatitis. Journal of Pediatrics 145, 612-
616.
Rosenfeldt, V., Michaelsen, K.F., Jakobsen, M, Larsen, C.N., Moller, P.L., Tvede, M.,
Weyrehter, H., Valerius, N.H., Paerregaard, A., 2002. Effect of probiotic
Lactobacillus strains on acute diarrhea in a cohort of nonhospitalized children
attending day-care centers. Pediatric Infectious Disease Journal 21, 417-419.
Rybarczyk, B.J., Lawrence, S.O., Simpson-Haidaris, P.J., 2003. Matrix-fibrinogen
enhances wound closure by increasing both cell proliferation and migration.
Blood 102, 4035-4043.
Sakamoto, I., Igarashi, M., Kimura, K., Takagi, A., Miwa, T., Koga, Y., 2001.
Suppressive effect of Lactobacillus gasseri OLL 2716 (LG21) on Helicobacter
pylori infection in humans. Journal of Antimicrobial Chemotherapy 47, 709-
710.
Sartor, R.B., 2004. Therapeutic manipulation of the enteric microflora in inflammatory
bowel diseases: antibiotics, probiotics, and prebiotics. Gastroenterology 126,
1620-1633.

192
Sartor, R.B., 2005. Probiotic therapy of intestinal inflammation and infections.
Current Opinion in Gastroenterology 21, 44-50.
Sato, K., Saito, H., Tomioka, H., Yokokura, T., 1988. Enhancement of host resistance
against Listeria infection by Lactobacillus casei: efficacy of cell wall preparation
of Lactobacillus casei. Microbiology and Immunology 32, 1189-1200.
Saxelin, M., Tynkkynen, S., Mattila-Sandholm, T., de Vos, W.M., 2005. Probiotic and
other functional microbes: from markets to mechanisms. Current Opinion in
Biotechnology 16, 204-211.
Scaltriti, M., Santamaria, A., Paciucci, R., Bettuzzi, S., 2004. Intracellular Clusterin
Induces G2-M Phase Arrest and Cell Death in PC-3 Prostate Cancer Cells1.
Cancer Research 64, 6174-6182.
Schachtsiek, M., Hammes, W.P., Hertel, C., 2004. Characterization of Lactobacillus
coryniformis DSM 20001T surface protein Cpf mediating coaggregation with
and aggregation among pathogens. Applied and Environmental Microbiology 70,
7078-7085.
Schiffrin, E.J., Blum, S., 2002. Interactions between the microbiota and the intestinal
mucosa. European Journal of Clinical Nutrition 56, S60-S64.
Schiffrin, E.J., Brassart, D., Servin, A.L., Rochat, F., Donnet-Hughes, A., 1997. Immune
modulation of blood leukocytes in humans by lactic acid bacteria: criteria for
strain selection. American Journal of Clinical Nutrition 66, 515S-520S.
Schiffrin, E.J., Rochat, F., Link-Amster, H., Aeschlimann, J.M., Donnet-Hughes, A.,
1995. Immunomodulation of human blood cells following the ingestion of lactic
acid bacteria. Journal of Dairy Science 78, 491-497.

193
Schultz, M., Linde, H.J., Lehn, N., Zimmermann, K., Grossmann, J., Falk, W.,
Scholmerich, J., 2003. Immunomodulatory consequences of oral administration
of Lactobacillus rhamnosus strain GG in healthy volunteers. Journal of Dairy
Research 70, 165-173.
Schultz, M., Veltkamp, C., Dieleman, L.A., Grenther, W.B., Wyrick, P.B., Tonkonogy,
S.L., Sartor, R.B., 2002. Lactobacillus plantarum 299v in the treatment and
prevention of spontaneous colitis in interleukin-10-deficient mice. Inflammatory
Bowel Diseases 8, 71-80.
Schwartz, R.H., 1989. Acquisition of immunologic self-tolerance. Cell 57, 1073-1081.
Sen, S., Mullan, M.M., Parker, T.J., Woolner, J.T., Tarry, S.A., Hunter, J.O., 2002.
Effect of Lactobacillus plantarum 299v on colonic fermentation and
symptoms of irritable bowel syndrome. Digestive Diseases and Sciences 47,
2615-2620.
Sepulveda, A.R., Tao, H., Carloni, E., Sepulveda, J., Graham, D.Y., Peterson, L.E.,
2002. Screening of gene expression profiles in gastric epithelial cells induced
by Helicobacter pylori using microarray analysis. Alimentary Pharmacology
& Therapeutics 16, 145-157.
Shanahan, F., 1999. Intestinal lymphoepithelial communication. Advances in
Experimental Medicine and Biology 473, 1-9.
Shanahan, F., 2002. Probiotics and inflammatory bowel disease: from fads and fantasy
to facts and future. British Journal of Nutrition 88, S5-S9.
Shanahan, F., 2002a. The host-microbe interface within the gut. Best Practice &
Research: Clinical Gastroenterology 16, 915-931.

194
Sheil, B., McCarthy, J., O'Mahony, L., Bennett, M.W., Ryan, P., Fitzgibbon, J.J., Kiely,
B., Collins, J.K., Shanahan, F., 2004. Is the mucosal route of administration
essential for probiotic function? Subcutaneous administration is associated with
attenuation of murine colitis and arthritis. Gut 53, 694-700.
Sherman, L.A., Savage, D.C., 1986. Lipoteichoic acids in Lactobacillus strains that
colonize the mouse gastric epithelium. Applied and Environmental Microbiology
52, 302-304.
Shockett, P., Stavnezer, J., 1991. Effect of cytokines on switching to IgA and alpha
germline transcripts in the B lymphoma I.29 mu. Transforming growth factor-
beta activates transcription of the unrearranged C alpha gene. Journal of
Immunology 147, 4374-4383.
Shu, Q., Gill, H.S., 2002. Immune protection mediated by the probiotic Lactobacillus
rhamnosus HN001 (DR20) against Escherichia coli O157:H7 infection in mice.
FEMS Immunology and Medical Microbiology 34, 59-64.
Simon, G.L., Gorbach, S.L., 1986. The human intestinal microflora. Digestive Diseases
and Sciences 31, 147S-162S.
Sjovall, J., 1959. On the concentration of bile acids in the human intestine during
absorption. Bile acids and sterioids 74. Acta Physiologica Scandinavica. 46, 339-
345.
Smith, J.M., Johanesen, P.A., Wendt, M.K., Binion, D.G., Dwinell, M.B., 2005.
CXCL12 activation of CXCR4 regulates mucosal host defense through
stimulation of epithelial cell migration and promotion of intestinal barrier
integrity. American Journal of Physiology - Gastrointestinal and Liver
Physiology 288, G316-G326.

195
Smith, T., 1995. The digestive system. In: Smith, T., (ed.), The human body, Ken Fin
Books, Collingwood, pp. 150-173.
Smits, H.H., Engering, A., van der Kleij, D., de Jong, E.C., Schipper, K., van Capel,
T.M., Zaat, B.A., Yazdanbakhsh, M., Wierenga, E.A., van Kooyk, Y.,
Kapsenberg, M.L., 2005. Selective probiotic bacteria induce IL-10-producing
regulatory T cells in vitro by modulating dendritic cell function through dendritic
cell-specific intercellular adhesion molecule 3-grabbing nonintegrin. Journal of
Allergy and Clinical Immunology 115, 1260-1267.
Sobko, T., Huang, L., Midtvedt, T., Norin, E., Gustafsson, L.E., Norman, M., Jansson,
E.A., Lundberg, J.O., 2006. Generation of NO by probiotic bacteria in the
gastrointestinal tract. Free Radical Biology & Medicine 41, 985-991.
Sobko, T., Norman, M., Noria, E., Gustafsson, L.E., Lundberg, J.O., 2005. Birth-related
increase in intracolonic hydrogen gas and nitric oxide as indicator of host-
microbial interactions. Allergy 60, 396-400.
Solis-Pereyra, B., Aattouri, N., Lemonnier, D., 1997. Role of food in the stimulation of
cytokine production. American Journal of Clinical Nutrition 66, 521S-525S.
Sudo, N., Sawamura, S., Tanaka, K., Aiba, Y., Kubo, C., Koga, Y., 1997. The
requirement of intestinal bacterial flora for the development of an IgE production
system fully susceptible to oral tolerance induction. Journal of Immunology 159,
1739-1745.
Suskovic, J., Brkic, B., Matosic, S., Maric, V., 1997. Lactobacillus acidophilus M92 as
potential probiotic strains. Milchwissenschaft 52, 430-435.
Sutas, Y., Autio, S., Rantala, I., Isolauri, E., 1997. IFN-gamma enhances
macromolecular transport across Peyer's patches in suckling rats: implications

196
for natural immune responses to dietary antigens early in life. Journal of
Pediatric Gastroenterology and Nutrition 24, 162-169.
Szajewska, H., Kotowska, M., Mrukowicz, J.Z., Armanska, M., Mikolajczyk, W., 2001.
Efficacy of Lactobacillus GG in prevention of nosocomial diarrhea in infants.
Journal of Pediatrics 138, 361-365.
Tagg, J.R., Dierksen, K.P., 2003. Bacterial replacement therapy: adapting 'germ warfare'
to infection prevention. Trends in Biotechnology 21, 217-223.
Takahashi, N., Kitazawa, H., Iwabuchi, N., Xiao, J.Z., Miyaji, K., Iwatsuki, K., Saito,
T., 2006. Immunostimulatory oligodeoxynucleotide from Bifidobacterium
longum suppresses Th2 immune responses in a murine model. Clinical and
Experimental Immunology 145, 130-138.
Tao, Y., Drabik, K.A., Waypa, T.S., Musch, M.W., Alverdy, J.C., Schneewind, O.,
Chang, E.B., Petrof, E.O., 2006. Soluble factors from Lactobacillus GG activate
MAPKs and induce cytoprotective heat shock proteins in intestinal epithelial
cells. American Journal of Physiology - Cell Physiology 290, C1018-C1030.
Tejada-Simon, M.V., Ustunol, Z., Pestka, J.J., 1999. Ex vivo effects of lactobacilli,
streptococci, and bifidobacteria ingestion on cytokine and nitric oxide production
in a murine model. Journal of Food Protection 62, 162-169.
Tejada-Simon, M.V., Lee, J.H., Ustunol, Z., Pestka, J.J., 1999a. Ingestion of yogurt
containing Lactobacillus acidophilus and Bifidobacterium to potentiate
immunoglobulin A responses to cholera toxin in mice. Journal of Dairy Science
82, 649-660.

197
Tejada-Simon, M.V., Pestka, J.J., 1999b. Proinflammatory cytokine and nitric oxide
induction in murine macrophages by cell wall and cytoplasmic extracts of lactic
acid bacteria. Journal of Food Protection 62, 1435-1444.
Terpend, K., Blaton, M.A., Candalh, C., Wal, J.M., Pochart, P., Heyman, M., 1999.
Intestinal barrier function and cow's milk sensitization in guinea pigs fed milk or
fermented milk. Journal of Pediatric Gastroenterology and Nutrition 28, 191-198.
Todoriki, K., Mukai, T., Sato, S., Toba, T., 2001. Inhibition of adhesion of food-borne
pathogens to Caco-2 cells by Lactobacillus strains. Journal of Applied
Microbiology 91, 154-159.
Tournut, J., 1993. The digestive flora of the pig and its variations. Recueil de Medecine
Veterinaire 169, 645-652.
Tromm, A., Niewerth, U., Khoury, M., Baestlein, E., Wilhelms, G., Schulze, J., Stolte,
M., 2004. The probiotic E. coli strain Nissle 1917 for the treatment of
collagenous colitis: first results of an open-label trial. Zeitschrift fur
Gastroenterologie 42, 365-369.
Truelstrup Hansen, L., Allan-Wojtas, P.M., Jin, Y.L., Paulson, A.T., 2002. Survival of
Ca-alginate microencapsulated Bifidobacterium spp. in milk and simulated
gastrointestinal conditions. Food Microbiology 19, 35-45.
Umesaki, Y., Okada, Y., Matsumoto, S., Imaoka, A., Setoyama, H., 1995. Segmented
filamentous bacteria are indigenous intestinal bacteria that activate intraepithelial
lymphocytes and induce MHC class II molecules and fucosyl asialo GM1
glycolipids on the small intestinal epithelial cells in the ex-germ-free mouse.
Microbiology and Immunology 39, 555-562.

198
Underdown, B.J., Schiff, J.M., 1986. Immunoglobulin A: strategic defense initiative at
the mucosal surface. Annual Review of Immunology 4, 389-417.
Usman, Hosono, A., 1999. Bile tolerance, taurocholate deconjugation, and binding of
cholesterol by Lactobacillus gasseri strains. Journal of Dairy Science 82, 243–
248.
Valeur, N., Engel, P., Carbajal, N., Connolly, E., Ladefoged, K., 2004. Colonization and
immunomodulation by Lactobacillus reuteri ATCC 55730 in the human
gastrointestinal tract. Applied and Environmental Microbiology 70, 1176– 1181.
van den Blink, B., Ten Hove, T., van den Brink, G.R., Peppelenbosch, M.P., van
Deventer, S.J., 2002. From extracellular to intracellular targets, inhibiting MAP
kinases in treatment of Crohn's disease. Annals of the New York Academy of
Sciences 973, 349-358.
Vandevoorde, L., Christiaens, H., Verstraete, W., 1992. Prevalence of coaggregation
reactions among chicken lactobacilli. Journal of Applied Bacteriology 72, 214-
219.
Vaughan, E.E., Mollet, B., 1999. Probiotics in the new millennium. Nahrung 43, 148-
153.
Ventura, M., Jankovic, I., Walker, D.C., Pridmore, R.D., Zink, R., 2002. Identification
and characterization of novel surface proteins in Lactobacillus johnsonii and
Lactobacillus gasseri. Applied and Environmental Microbiology 68, 6172-6181.
Villena, J., Racedo, S., Aguero, G., Alvarez, S., 2006. Yoghurt accelerates the recovery
of defence mechanisms against Streptococcus pneumoniae in protein-
malnourished mice. British Journal of Nutrition 95, 591-602.

199
Vinderola, C.G., Medici, M., Perdigon, G., 2004. Relationship between interaction sites
in the gut, hydrophobicity, mucosal immunomodulating capacities and cell wall
protein profiles in indigenous and exogenous bacteria. Journal of Applied
Microbiology 96, 230-243.
Vinderola, G., Perdigon, G., Duarte, J., Farnworth, E., Matar, C., 2006. Effects of the
oral administration of the products derived from milk fermentation by kefir
microflora on immune stimulation. Journal of Dairy Research 73, 472-479.
Visnjic, D., Batinic, D., Banfic, H., 1997. Arachidonic acid mediates interferon-gamma-
induced sphingomyelin hydrolysis and monocytic marker expression in HL-60
cell line. Blood 89, 81-91.
Vitini, E., Alvarez, S., Medina, M., Medici, M., de Budeguer, M.V., Perdigon, G., 2000.
Gut mucosal immunostimulation by lactic acid bacteria. Biocell 24, 223-232.
Vizoso Pinto, M.G., Schuster, T., Briviba, K., Watzl, B., Holzapfel, W.H., Franz, C.M.,
2007. Adhesive and chemokine stimulatory properties of potentially probiotic
Lactobacillus strains. Journal of Food Protection 70, 125-134.
Wadstrom, T., Andersson, K., Sydow, M., Axelsson, L., Lindgren, S., Gullmar, B.,
1987. Surface properties of lactobacilli isolated from the small intestine of pigs.
Journal of Applied Bacteriology 62, 513-520.
Wagner, R.D., Warner, T., Roberts, L., Farmer, J., Balish, E., 1997. Colonization of
congenitally immunodeficient mice with probiotic bacteria. Infection and
Immunity 65, 3345-3351.
Wallace, J.L., Vergnolle, N., Muscara, M.N., Asfaha, S., Chapman, K., McKnight, W.,
Del Soldato, P., Morelli, A., Fiorucci, S., 1999. Enhanced anti-inflammatory

200
effects of a nitric oxide-releasing derivative of mesalamine in rats.
Gastroenterology 117, 557-566.
Wallace, T.D., Bradley, S., Buckley, N.D., Green-Johnson, J.M., 2003. Interactions of
lactic acid bacteria with human intestinal epithelial cells: effects on cytokine
production. Journal of Food Protection 66, 466-472.
Walter, J., Chagnaud, P., Tannock, G.W., Loach, D.M., Dal Bello, F., Jenkinson, H.F.,
Hammes, W.P., Hertel, C., 2005. A high-molecular-mass surface protein (Lsp)
and methionine sulfoxide reductase B (MsrB) contribute to the ecological
performance of Lactobacillus reuteri in the murine gut. Applied and
Environmental Microbiology 71, 979-986.
Wang, K.Y., Li, S.N., Liu, C.S., Perng, D.S., Su, Y.C., Wu, D.C., Jan, C.M., Lai, C.H.,
Wang, T.N., Wang, W.M., 2004. Effects of ingesting Lactobacillus- and
Bifidobacterium-containing yogurt in subjects with colonized Helicobacter
pylori. American Journal of Clinical Nutrition 80, 737-741.
Wehkamp, J., Harder, J., Wehkamp, K., Wehkamp-von Meissner, B., Schlee, M.,
Enders, C., Sonnenborn, U., Nuding, S., Bengmark, S., Fellermann, K.,
Schroder, J.M., Stange, E.F., 2004. NF-kappaB- and AP-1-mediated induction of
human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle
1917: a novel effect of a probiotic bacterium. Infection and Immunity 72, 5750-
5758.
Weiner, H.L., Friedman, A., Miller, A., Khoury, S.J., al-Sabbagh, A., Santos, L.,
Sayegh, M., Nussenblatt, R.B., Trentham, D.E., Hafler, D.A., 1994. Oral
tolerance: immunologic mechanisms and treatment of animal and human organ-
specific autoimmune diseases by oral administration of autoantigens. Annual
Review of Immunology 12, 809-837.

201
Wells, C.L., Maddaus, M.A., Simmons, R.L., 1988. Proposed mechanisms for the
translocation of intestinal bacteria. Reviews of Infectious Diseases 10, 958-979.
Wiseman, B.S., Sternlicht, M.D., Lund, L.R., Alexander, C.M., Mott, J., Bissell, M.J.,
Soloway, P., Itohara, S., Werb, Z., 2003. Site-specific inductive and inhibitory
activities of MMP-2 and MMP-3 orchestrate mammary gland branching
morphogenesis. Journal of Cell Biology. 162, 1123-1133.
Wostmann, B.S., 1996. Immunology, including radiobiology and transplantation. In:
Wostmann, B.S., (ed.), Germfree and gnotobiotic animal models, CRC Press,
Boca Raton , pp. 101–125.
Wu, L., Thompson, D.K., Li, G., Hurt, R.A., Tiedje, J.M., Zhou, J., 2001. Development
and evaluation of functional gene arrays for detection of selected genes in the
environment. Applied and Environmental Microbiology 67, 5780-5790.
Wullt, M., Hagslatt, M.L., Odenholt, I., 2003. Lactobacillus plantarum 299v for the
treatment of recurrent Clostridium difficile-associated diarrhoea: a double-blind,
placebo-controlled trial. Scandinavian Journal of Infectious Diseases 35, 365-
367.
Xu, A., Lam, M.C., Chan, K.W., Wang, Y., Zhang, J., Hoo, R.L., Xu, J.Y., Chen, B.,
Chow, W.S., Tso, A.W., Lam, K.S., 2005. Angiopoietin-like protein 4 decreases
blood glucose and improves glucose tolerance but induces hyperlipidemia and
hepatic steatosis in mice. Proceedings of the National Academy of Sciences USA
102, 6086-6091.
Xu, J., Gordon, J.I., 2003. Inaugural Article: Honor thy symbionts. Proceedings of the
National Academy of Sciences USA 100, 10452-10459.

202
Xu, J., Verstraete, W., 2001. Evaluation of nitric oxide production by lactobacilli.
Applied Microbiology and Biotechnology 56, 504-507.
Yagi, T., Takeichi, M., 2000. Cadherin superfamily genes: functions, genomic
organization, and neurologic diversity. Genes & Development 14, 1169-1180.
Yuan, Q., Walker, W.A., 2004. Innate immunity of the gut: mucosal defense in health
and disease. Journal of Pediatric Gastroenterology and Nutrition 38, 463-473.
Zarate, G., Chaia, A.P., Gonzalez, S., Oliver, G., 2000. Viability and beta-galactosidase
activity of dairy propionibacteria subjected to digestion by artificial gastric and
intestinal fluids. Journal of Food Protection 63, 1214-1221.
Zareie, M., Johnson-Henry, K., Jury, J., Yang, P.C., Ngan, B.Y., McKay, D.M.,
Soderholm, J.D., Perdue, M.H., Sherman, P.M., 2006. Probiotics prevent
bacterial translocation and improve intestinal barrier function in rats following
chronic psychological stress. Gut 55, 1553-1560.
Zhang, L., Li, N., Caicedo, R., Neu, J., 2005. Alive and dead Lactobacillus rhamnosus
GG decrease tumor necrosis factor-alpha-induced interleukin-8 production in
Caco-2 cells. Journal of Nutrition 135, 1752-1756.
Zheng, L., He, M., Long, M., Blomgran, R., Stendahl, O., 2004. Pathogen-induced
apoptotic neutrophils express heat shock proteins and elicit activation of human
macrophages. Journal of Immunology 173, 6319-6326.
Zoetendal, E.G., Cheng, B., Koike, S., Mackie, R.I., 2004. Molecular microbial ecology
of the gastrointestinal tract: from phylogeny to function. Current Issues in
Intestinal Microbiology 5, 31-47.
Related Documents











