Pergamon 0098-8472(94)E0011-R EnvironmentalandExpenmentalBotany, Vol. 35, No. 1, pp. 83-92, 1995 Copyright © 1995 ElsevierScience Ltd Printed in Great Britain, All rights reserved 0098-8479/95 $9.50+0.00 GROWTH, PROLINE ACCUMULATION AND WATER RELATIONS OF NaC1-SELECTED AND NON-SELECTED CALLUS LINES OF DACTYLIS GLOMERA TA L. S. DUTTA GUPTA,* R. M. AUGE,-[" P. D. DENCHEV. • and B. V. CONGEI~§ • Department of Agricultural Engineering, Indian Institute of Technology, Kharagpur-721302, India; I"Department of Ornamental Horticulture and Landscape Design, University of Tennessee, Knoxville, TN 37901-1071, U.S.A.; and ~ Department of Plant and Soil Science, University of Tennessee, Knoxville, TN 37901-1071, U.S.A. (Received 28 July 1993; acceptedin revised form 11 February 1994) Dutta Gupta S., Aug6 R. M., Denchev P. D. and Conger B. V. Growth, proline accumulation and water relations in NaCl-selected and non-selected callus lines of Dactylis glomerata L. Environmental and Experimental Botany 35, 83-92, 1995.--Sodium chloride-tolerant calfi were selected from leaf- derived embryogenic calli ofDac~lis glomerata L. on agar solidified medium supplemented with 200 mM NaC1, a concentration lethal to non-selected calli. Growth characteristics, water relations and proline accumulation pattern were compared in selected and non-selected lines. The objective was to gain an understanding of the mechanism(s) of tolerance in the NaCl-tolerant line. Growth in the selected fine, as expressed in terms of tolerance index (ratio of fresh wt. on NaC1 medium:fresh wt. on NaC1 free medium x 100), was greater than that of the non-selected line at all levels of NaC1 between 50 and 300 mM. There was no significant difference in proline accumulation in the selected and non-selected lines. Maintenance of turgor by osmotic adjustment was observed in the non- selected line despite decreased growth. In contrast, the selected line lost either the need or the ability to adjust osmotically. There was little or no increase in symplastic osmolality in the selected line when exposed to NaC1. Presumably, selection was made for a salt-excluding tissue that has lost the ability to accumulate solutes and adjust turgor with NaC1 stress. I@ words: NaCl-tolerance, tissue culture, osmotic adjustment, orchardgrass, Dactylis glomerata L. INTRODUCTION There have been many reports on in vitro selection of NaCl-tolerant cell/callus lines.(12' 10,19,30,351 The iso- lation of NaCl-tolerant cultures may offer potential for the retrieval of salt-tolerant plants. Such cultures also represent an ideal system to assess the physio- logical effects of salt and/or water stress at the cellu- lar level.(2a/Salt stress results from a lowering of the water potential of the plant environment and from § Author to whom all correspondence should be addressed. Abbreviations'. WC = water content; W = water potential; ROWC = relative osmotic water potential. 83 Wp = turgor potential; W, = osmotic potential;

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pergamon
0098-8472(94)E0011-R
EnvironmentalandExpenmentalBotany, Vol. 35, No. 1, pp. 83-92, 1995 Copyright © 1995 Elsevier Science Ltd
Printed in Great Britain, All rights reserved 0098-8479/95 $9.50+0.00
GROWTH, PROLINE ACCUMULATION AND WATER RELATIONS OF NaC1-SELECTED AND NON-SELECTED CALLUS LINES OF
DACTYLIS GLOMERA TA L.
S. DUTTA GUPTA,* R. M. AUGE,-[" P. D. DENCHEV. • and B. V. CONGEI~§
• Department of Agricultural Engineering, Indian Institute of Technology, Kharagpur-721302, India; I" Department of Ornamental Horticulture and Landscape Design, University of Tennessee, Knoxville,
TN 37901-1071, U.S.A.; and ~ Department of Plant and Soil Science, University of Tennessee, Knoxville, TN 37901-1071, U.S.A.
(Received 28 July 1993; accepted in revised form 11 February 1994)
Dutta Gupta S., Aug6 R. M., Denchev P. D. and Conger B. V. Growth, proline accumulation and water relations in NaCl-selected and non-selected callus lines of Dactylis glomerata L. Environmental and Experimental Botany 35, 83-92, 1995.--Sodium chloride-tolerant calfi were selected from leaf- derived embryogenic calli ofDac~lis glomerata L. on agar solidified medium supplemented with 200 mM NaC1, a concentration lethal to non-selected calli. Growth characteristics, water relations and proline accumulation pattern were compared in selected and non-selected lines. The objective was to gain an understanding of the mechanism(s) of tolerance in the NaCl-tolerant line. Growth in the selected fine, as expressed in terms of tolerance index (ratio of fresh wt. on NaC1 medium:fresh wt. on NaC1 free medium x 100), was greater than that of the non-selected line at all levels of NaC1 between 50 and 300 mM. There was no significant difference in proline accumulation in the selected and non-selected lines. Maintenance of turgor by osmotic adjustment was observed in the non- selected line despite decreased growth. In contrast, the selected line lost either the need or the ability to adjust osmotically. There was little or no increase in symplastic osmolality in the selected line when exposed to NaC1. Presumably, selection was made for a salt-excluding tissue that has lost the ability to accumulate solutes and adjust turgor with NaC1 stress.
I@ words: NaCl-tolerance, tissue culture, osmotic adjustment, orchardgrass, Dactylis glomerata L.
INTRODUCTION
There have been many reports on in vitro selection of NaCl-tolerant cell/callus lines.(12' 10,19,30,351 The iso- lation of NaCl-tolerant cultures may offer potential
for the retrieval of salt-tolerant plants. Such cultures also represent an ideal system to assess the physio- logical effects of salt and/or water stress at the cellu- lar level. (2a/Salt stress results from a lowering of the water potential of the plant environment and from
§ Author to whom all correspondence should be addressed. Abbreviations'. WC = water content; W = water potential;
ROWC = relative osmotic water potential.
83
Wp = turgor potential; W, = osmotic potential;

84 S. DUTTA GUPTA et al.
the toxic effect oi" ions. (25) Osmotic adjustment is frequently cited as one mechanism enabling stressed plants to maintain turgor and is necessary for growth of cells under saline conditions. A large number of in vitro studies has been conducted with tobacco, (4'5'2°'22'24'4°) Citrus sp. (2'3'26) and Brassica sp. 0°'n'33) to elucidate the tolerance mechanism(s). These reports have unequivocally indicated turgor restoration by osmotic adjustment, but in some instances with reduced growth. (4)
It has been suggested that growth reduction in adapted cells is not due to an inability of the cells to adjust osmotically. An alteration in the yield threshold and/or extensibility of the cell walls to turgor must have occurred during NaCI adaptation. (8) That is, complete restoration ofturgor may not be required since growth could be restored by changes either in the yield threshold or cell wall extensibility. Unfortunately, studies on the water relations of cultured cells of gramineous species under NaC1 stress have received little attention, particularly whether they are capable of main- taining turgor and/or restoring their growth rate. °3'16) Comparison of salt-tolerant and non-tol- erant cultures with an emphasis on growth and turgor maintenance enables measurement of the range of cellular tolerance and may also reveal the turgor/growth relations of cells.
The present study was conducted to compare growth, proline accumulation and osmotic adjust- ment in NaC1 selected and non-selected callus lines oforchardgrass (Dac~lisglomerata L.) with the objec- tive of assessing these relationships to gain a better understanding of the tolerance mechanism(s). The genotype, 'Embryogen-P '(l~ used in these studies has a high capacity for regeneration through somatic embryogenesis and we have used it pre- viously in pressure-volume analyses of in vitro cultures. °)
MATERIALS AND METHODS
Initiation and maintenance of callus cultures In vitro cultures were initiated from young leaf
segments and maintained on Schenk and Hilde- brandt medium (stl containing 30 gM 3,6-dichloro- 2-methoxybenzoic acid (dicamba) and 0.8% agar (SH-30). All cultures were subcukured (transferred to fresh medium) at 4-week intervals and kept in the dark at 27 -t- I°C.
Selection procedure For selection of a NaCl-tolerant callus fine,
actively growing embryogenic calli were exposed to increased concentrations of NaCI (0, 50, 100, 150, 200, 250 or 300 mM) to determine the con- centration at which growth was completely inhibited. Cell survival was non-existent at 250 mM. Therefore, a concentration of 200 mM was used for selection. At this concentration most of the calli turned brown within 4 weeks. However, a portion which remained healtlay in appearance was sub- cultured every 4 weeks onto fresh medium con- taining 200 mM NaCI. After 7 months, a callus was identified which exhibited continued growth without discoloration. This callus line will be referred to as the NaC1- selected line to distinguish it from the non-selected (control) line.
Characteristics of the selected line For quantification of NaCl-tolerance, embryo-
genic callus pieces from non-selected and selected cultures initiated on the same date, were transferred to SH-30 medium containing 0, 50, 100, 150, 200, 250 or 300 mM NaCI. There were 20 replications (dishes) of five or six calli (200 ___ 25 mg) for each concentration. The cultures were incubated in the dark at 25°C and transferred to fresh medium three times at 14-day intervals. Fresh wt. per replicate was measured after 56 days and expressed as a tolerance index (TI):
Fresh wt. on NaC1 medium T I = x 100.
Fresh wt. on NaC1 free medium
The fresh wt. to dry wt. ratio (Fw:Dw) was also measured after drying the tissue at 80°C for 24 hr. Relative growth rates (g g- 1 week- 1) were based on the increases in fresh and dry wts over the 56-day period using the formula for exponential growth: GR = [In (final wt . ) - ln (initial wt.)]/time. (m)
Stabili~ test Calli from the selected line were allowed to grow
on salt free medium for 84 days. They were then transferred back to SH-30 medium containing 100, 150 or 200 mM NaC1. The effect of NaC1 was studied as in the selection experiment. The line which retained its NaCl-tolerance on 200 mM NaC1 was referred to as the stable selected line.

GROWTH, PROLINE ACCUMULATION AND WATER 85
Estimation offiee proline Free proline was estimated according to the
methods of Singh et alJ sT~ Three replicate samples were measured for each treatment and proline con- centration is expressed on a dry wt. basis.
Measurement of water relations Callus water relations were measured using pres-
sure-volume analysis I39) as described previously. Ill Two SC-10 thermocouple psychrometers with sam- ple changers connected to NT-3 nanovoltmeter thermometers (Decagon Devices, Pullman, WA, U.S.A.) were used to derive temperature and gV readings for conversion into water potential (W) values. Psychrometers were calibrated with a graded series of NaC1 solutions. Standard psy- chrometer operating procedures and cautions were observed./9) Calli were removed from plates, weighed and placed in sample cups in the psy- chrometer chamber. All manipulations were per- formed in an enclosed, humid chamber to minimize evaporative loss from tissue. Samples were removed from the psychrometer and allowed to dehydrate on the bench between successive W determinations. Estimations of W were obtained at several levels of callus relative water content (RWC) between hydration at harvest (100% RWC) and 60% of harvest RWC. Turgor pressure (Wp) was computed as the difference between W and osmotic potential (W~). The inverse of - W (y axis) was plotted against the cumulative volume lost (~ x axis) and a linear regression fitted to the linear segment of the curve which developed when bulk turgor had been lost; 139) correlation coefficients for regressions were above 0.990 (most were above 0.995) for plots of all callus samples in this study. The y intercept of this line (_ W~ 1) gave the inverse of the harvest W~ and the x intercept gave the total volume of osmotic (sym- plastic) water in the sample (V~ef). Apoplastic water percentage was computed as total water in the sample --V~er. The bulk tissue elastic modulus (e) was computed as A~p/AROWC = (~p~est_ ~IJprgOr loss)/(ROWCharvest - ROWCtU,gor loss). R O W C , the relative osmotic water content, was computed as 100(V=f-- V)/Vr~r. The initial point in the linear segment of the PV curve was regarded as the ~P (and ~P,) at the turgor loss point, and gave RWC at turgor loss.
RESULTS
Growth characteristics Culture weights and relative growth rates of non-
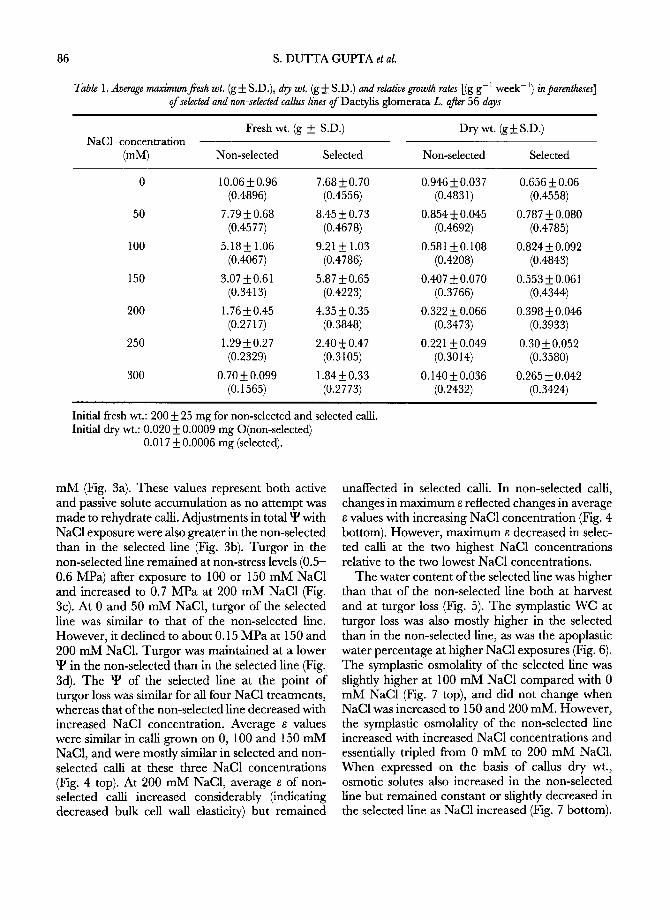
selected and selected callus lines as a function of increased NaC1 concentration are presented in Table 1. Under non-stress (0 mM NaC1), the selec- ted line did not grow as well over the 56-day period as the non-selected line. However, with the addition of NaC1, the growth rate of the selected line was greater than that of the non-selected line at each concentration. The final fresh wts of the selected line after 56 days was approximately 1.8-2.6 times greater than that of the non-selected line at con- centrations of 100 mM and above. Growth of the two lines, as expressed by TI, revealed a similar trend (Fig. 1). The selected line maintained on 50 and 100 mM NaC1 showed a TI of more than 100%. Further, the TI of the selected line was more than double that of the non-selected line at NaC1 concentrations of >/100 mM. The 100 mM NaC1 was optimal for the growth of the selected line. The Fw:Dw ratios of the selected line remained about the same up to 200 mM NaC1 and then declined (Fig. 1). The Fw:Dw ratio steadily declined in non- selected cultures between 0 and 200 mM NaCI after which it remained constant.
Stability test Tolerance of the selected line to 200 mM NaCI
was stable through six successive transfers under non-stress conditions. This was tested by plating it onto a medium containing 100, 150 or 200 mM NaC1. The stable selected line showed a reduced level of tolerance but still had significantly better growth than the non-selected line (Fig. 1).
Free proline content A linear increase in proline concentration was
observed in both fines (Fig. 2). However, except for 100 mM NaC1, there were no significant differences between the lines.
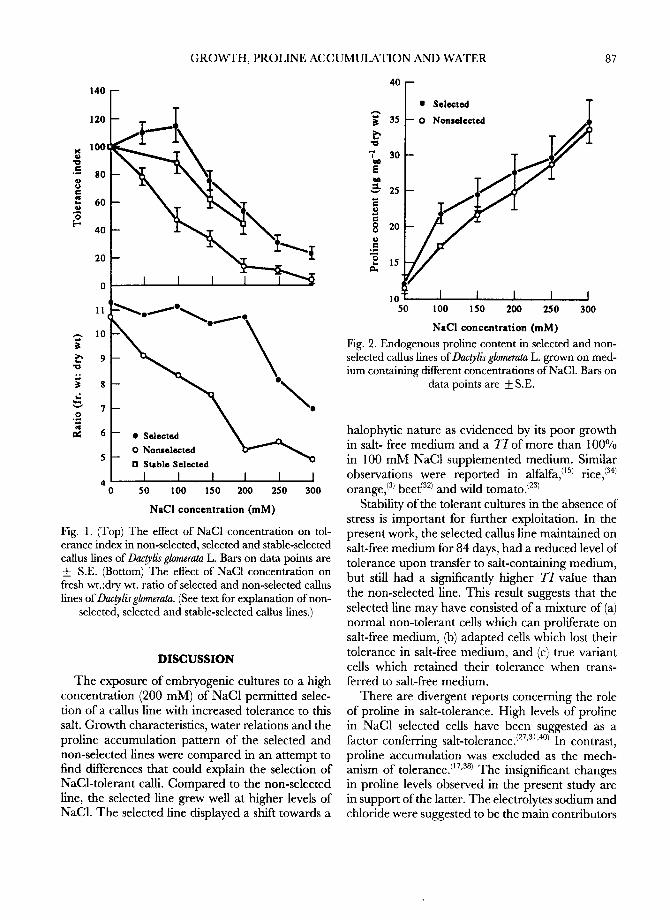
Water relations The osmotic potential of non-selected calli at 200
mM NaC1 was 1.25 MPa lower than that of non- selected calli at 0 mM NaCI, whereas the osmotic potential of selected calli at 200 mM NaC1 was only 0.26 MPa lower than that of the selected calli at 0
86 S. D U T r A GUPTA et al.
Table 1. Average maximum flesh wt. (g _+ S.D.), d~y wt. (g _+ S.D.) and relative growth rates [(g g- l week- 1) in parentheses] of selected and non-selected callus lines 0f Dactylis glomerata L. after 56 days
Fresh wt. (g _+ S.D.) NaC1 concentration
(mM) Non-selected Selected
Dry wt. (g-+ S.D.)
Non-selected Selected
0 10.06_+0.96 7 . 6 8 _ _ _ 0 . 7 0 0 .946___0.037 0.656_+0.06 (0.4896) (0.4556) (0.4831) (0.4558)
50 7.79-I-0.68 8 . 4 5 _ _ _ 0 . 7 3 0.854-t-0.045 0.787+0.080 (0.4577) (0.4678) (0.4692) (0.4785)
100 5.18-1- 1.06 9.21 _ 1.03 0.581 -t-0.108 0.824___0.092 (0.4067) (0.4786) (0.4208) (0.4843)
150 3.07+0.61 5.87_+0.65 0.407-t-0.070 0.553___0.061 (0.3413) (0.4223) (0.3766) (0.4344)
200 1.76-t-0.45 4.35-t-0.35 0.322__.0.066 0.398___0.046 (0.2717) (0.3848) (0.3473) (0.3933)
250 1.29-t-0.27 2.40-t-0.47 0.221 ___0.049 0.30-t-0.052 (0.2329) (0.3105) (0.3014) (0.3580)
300 0.70_+0.099 1.84_+0.33 0.140_+0.036 0.265_+0.042 (0.1565) (0.2773) (0.2432) (0.3424)
Initial fresh wt.: 200-I-25 mg for non-selected and selected caUi. Initial dry wt.: 0.020_ 0.0009 mg O(non-selected)
0.017 ___ 0.0006 mg (selected).
m M (Fig. 3a). These values represent both active and passive solute accumulation as no attempt was made to rehydrate calli. Adjustments in total • with NaCI exposure were also greater in the non-selected than in the selected line (Fig. 3b). Turgor in the non-selected line remained at non-stress levels (0.5- 0.6 MPa) after exposure to 100 or 150 m M NaC1 and increased to 0.7 MPa at 200 m M NaC1 (Fig. 3c). At 0 and 50 m M NaC1, turgor of the selected line was similar to that of the non-selected line. However, it declined to about 0.15 MPa at 150 and 200 m M NaC1. Turgor was maintained at a lower
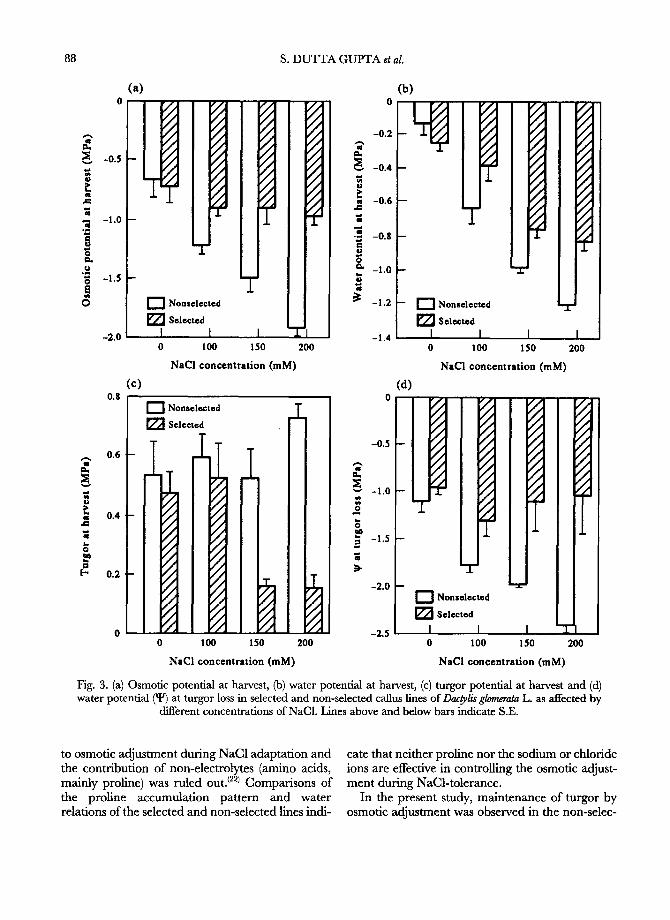
in the non-selected than in the selected line (Fig. 3d). The • of the selected line at the point of turgor loss was similar for all four NaC1 treatments, whereas that of the non-selected line decreased with increased NaC1 concentration. Average e values were similar in calli grown on 0, 100 and 150 m M NaC1, and were mostly similar in selected and non- selected calli at these three NaC1 concentrations (Fig. 4 top). At 200 m M NaCI, average ~ of non- selected calli increased considerably (indicating decreased bulk cell wall elasticity) but remained
unaffected in selected calfi. In non-selected calli, changes in max imum e reflected changes in average
values with increasing NaC1 concentration (Fig. 4 bottom). However, maximum ~ decreased in selec- ted calfi at the two highest NaC1 concentrations relative to the two lowest NaC1 concentrations.
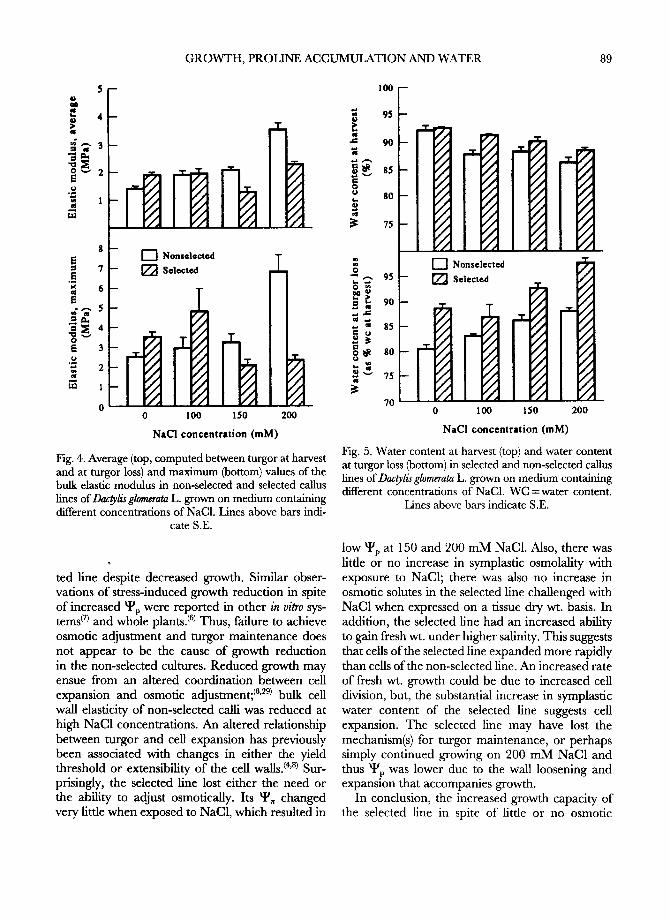
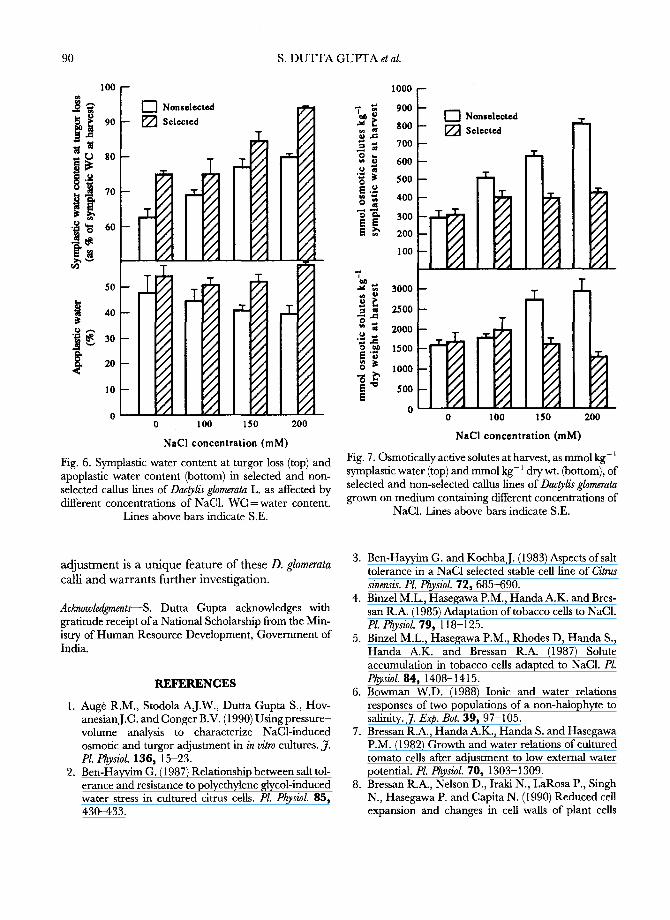
The water content of the selected line was higher than that of the non-selected line both at harvest and at turgor loss (Fig. 5). The symplastic WC at turgor loss was also mostly higher in the selected than in the non-selected fine, as was the apoplastic water percentage at higher NaC1 exposures (Fig. 6). The symplastic osmolality of the selected line was slightly higher at 100 m M NaC1 compared with 0 m M NaC1 (Fig. 7 top), and did not change when NaC1 was increased to 150 and 200 mM. However, the symplastic osmolality of the non-selected line increased with increased NaC1 concentrations and essentially tripled from 0 m M to 200 m M NaC1. When expressed on the basis of callus dry wt., osmotic solutes also increased in the non-selected line but remained constant or slightly decreased in the selected line as NaC1 increased (Fig. 7 bottom).
GROWTH, PROLINE ACCUMULATION AND WATER 87
140 - -
1oo~
8o
~, 6 o -
2O
o I I I I I ' ~
II - ' ~ ' - o ~ * ~ . ~ .
" " 7 ID 0
¢~ 6 - - *t Selected ~
5 - - 0 Nonselected U ' - -
n Stable Selected
4 I I I I I I 0 50 100 150 200 250 300
N a C i c o n c e n t r a t i o n ( m M )
Fig. 1. (Top) The effect of NaC1 concentration on tol- erance index in non-selected, selected and stable-selected callus lines ofDac~lis glomerata L. Bars on data points are _ S.E. (Bottom) The effect of NaC1 concentration on fresh wt.:dry wt. ratio of selected and non-selected callus lines of Dac~lisglomerata. (See text for explanation of non-
selected, selected and stable-selected callus lines.)
40
~" 35
"7 3O
E em
~ 25
8 2o
• Selected T
- 0 Nonselected A
D I S C U S S I O N
The exposure of embryogenic cultures to a high concentration (200 mM) of NaC1 permitted selec- tion of a callus line with increased tolerance to this salt. Growth characteristics, water relations and the proline accumulation pattern of the selected and non-selected lines were compared in an attempt to find differences that could explain the selection of NaCl-tolerant calli. Compared to the non-selected line, the selected line grew well at higher levels of NaCI. The selected line displayed a shift towards a
10 I I I [ I 50 100 150 200 250 300
N a C I c o n c e n t r a t i o n ( r a M )
Fig. 2. Endogenous proline content in selected and non- selected callus lines ofDactylisglomerata L. grown on med- ium containing different concentrations of NaC1. Bars on
data points are + S.E.
halophytic nature as evidenced by its poor growth in salt- free medium and a T I of more than 100% in 100 m M NaC1 supplemented medium. Similar observations were reported in alfalfa, (~5/ r i c e , (34)
orange,/3/beet/3~/and wild tomato./23/ Stability of the tolerant cultures in the absence of
stress is important for further exploitation. In the present work, the selected callus line maintained on salt-free medium for 84 days, had a reduced level of tolerance upon transfer to salt-containing medium, but still had a significantly higher T I value than the non-selected line. This result suggests that the selected line may have consisted of a mixture of (a) normal non-tolerant cells which can proliferate on salt-free medium, (b) adapted cells which lost their tolerance in salt-free medium, and (c) true variant cells which retained their tolerance when trans- ferred to salt-free medium.
There are divergent reports concerning the role of proline in sah-tolerance. High levels of proline in NaC1 selected cells have been suggested as a factor conferring salt-tolerance./27'3~'4°/In contrast, proline accumulation was excluded as the mech- anism of tolerance./17'38/ The insignificant changes in proline levels observed in the present study are in support of the latter. The electrolytes sodium and chloride were suggested to be the main contributors
88 S. DUTTA GUPTA et al.
(a) (b) .¢ / f j / A ~/~ : / ~" J f1' ,gA ¢ J ,-11/ /I -0.2 -- /CAC'A ~ ///1
-0.5 -
• ~ -O.It - -
~a ~ -I.0 -- "X"
D Nonselected -1.2 -- r~ Nonselected ~-=
-5.o s ° l ° ° " t I _ , . , I ) 0 100 150 200 0 100 150 200
NaCI concentration (raM) NaCI concentration (raM)
(c) (d) 0 . 8 ~ ~ 0
I--I ~o,,=1~=d ,..T., -0.5 i
0 . 6
~ 0 . 4 - -
0.2 ~"
-2.0 - r-I No,,=l=c~d ]
0 _2.5 ~ Selected ' I b 0 100 150 200 0 100 150 200
NaCI concentration (raM) NaCI concentration (mM)
Fig. 3. (a) Osmotic potential at harvest, (b) water potential at harvest, (c) turgor potential at harvest and (d) water potential (W) at turgor loss in selected and non-selected callus lines ofDac~lisglomerata L. as affected by
different concentrations of NaC1. Lines above and below bars indicate S.E.
to osmotic adjustment during NaC1 adaptation and the contribution of non-electrolytes (amino acids, mainly proline) was ruled out. (221 Comparisons of the proline accumulation pattern and water relations of the selected and non-selected lines indi-
cate that neither proline nor the sodium or chloride ions are effective in controlling the osmotic adjust- ment during NaCl-tolerance.
In the present study, maintenance of turgor by osmotic adjustment was observed in the non-selec-
GROWTH, PROLINE ACCUMULATION AND WATER 89
5 m u e~
e~
a
[ ]
7
-Mft - - [ ] N o n s e l e c t e d
- - ~ ' ~ S e l e c t e d
"r
0 100 150 2 0 0
N a C I c o n c e n t r a t i o n ( r a M )
Fig. 4. Average (top, computed between turgor at harvest and at turgor loss) and maximum (bottom) values of the bulk elastic modulus in non-selected and selected callus lines ofDac~lisglomerata L. grown on medium containing different concentrations of NaC1. Lines above bars indi-
cate S.E.
ted line despite decreased growth. Similar obser- vations of stress-induced growth reduction in spite of increased Wp were reported in other in vitro sys- tems (7) and whole plants. (6) Thus, failure to achieve osmotic adjustment and turgor maintenance does not appear to be the cause of growth reduction in the non-selected cultures. Reduced growth may ensue from an altered coordination between cell expansion and osmotic adjustment; (a'29) bulk cell wall elasticity of non-selected calli was reduced at high NaC1 concentrations. An altered relationship between turgor and cell expansion has previously been associated with changes in either the yield threshold or extensibility of the cell walls. I4'8) Sur- prisingly, the selected line lost either the need or the ability to adjust osmotically. Its W, changed very little when exposed to NaC1, which resulted in
100 -
9 5 - U
t "~ 9 0 -
0 o 80 - -
~ ~ 5 -
0 - - . - - 95 - - g~
.~ ,1 85
e t S0
70
[ ] N o n s c l c c t e d
[ ] S e l e c t e d
o
1 I00 150
o
l / , , ¢ / / / / /
f /
f / , ¢ j
/ / / / / / / J
/ J / /
! f / / /
2 0 0
NaCI concentration (raM)
Fig. 5. Water content at harvest (top) and water content at turgor loss (bottom) in selected and non-selected callus lines ofDac~lisglomerata L. grown on medium containing different concentrations of NaC1. WC = water content.
Lines above bars indicate S.E.
low ~p at 150 and 200 mM NaC1. Also, there was little or no increase in symplastic osmolafity with exposure to NaC1; there was also no increase in osmotic solutes in the selected line challenged with NaC1 when expressed on a tissue dry wt. basis. In addition, the selected line had an increased ability to gain fresh wt. under higher salinity. This suggests that cells of the selected line expanded more rapidly than cells of the non-selected line. An increased rate of fresh wt. growth could be due to increased cell division, but, the substantial increase in symplastic water content of the selected line suggests cell expansion. The selected line may have lost the mechanism(s) for turgor maintenance, or perhaps simply continued growing on 200 m M NaC1 and thus ~v was lower due to the wall loosening and expansion that accompanies growth.
In conclusion, the increased growth capacity of the selected line in spite of little or no osmotic
90 S. DUTFA GUPTA et al.
100
9 0 - -
~1 80 --
50
~ 30
2o
l0
[ ] Nonselccted [ ] Selected
T " - / / L / / / / / / / / , / / / /
/ / / / / / / /
/ / / / / / / I
l / / / / /
/ / / /
/ / / /
/ / l /
l / l / / / / /
/ / / /
lO0
! 1 / /
T / / / I / / / /
/ I / / / / / /
l / / / / /
l /
/ / / / / / / / / / / / / ] / / / / / / / / / / / / / /
/ /
150
NaCI concentration (mM)
,.T--
.L
200
Fig. 6. Symplastic water content at turgor loss (top) and apoplastic water content (bottom) in selected and non- selected callus fines of Dac~lis glomerata L. as affected by different concentrations of NaC1. WC =water content.
Lines above bars indicate S.E.
1000 --
"7 ~ 900
soo
= ~ 7oo O t . .o 600 t~
50o
O a
E 200
100
"7 ~ 3ooo
'~ ~ 2500
2000 <'7: 'i ,,oo
1000
5o.
o
m
[] Nonselected
__ -- [ ] Selected 1
-
.T-
0 100
22
150 200
NaC! concentration (raM)
Fig. 7. Osmotically active solutes at harvest, as mmol kg- l symplastic water (top) and mmol kg- l dry wt. (bottom), of selected and non-selected callus fines of Dac~lis glomerata grown on medium containing different concentrations of
NaC1. Lines above bars indicate S.E.
adjustment is a unique feature of these D. glomerata calli and warrants further investigation.
Acknowledgments~S. Dutta Gupta acknowledges with gratitude receipt of a National Scholarship from the Min- istry of Human Resource Development, Government of India.
REFERENCES
1. Aug6 R.M., Stodola AJ.W., Dutta Gupta S., Hov- anesianJ.C, and C onger B.V. (1990) Using pressure- volume analysis to characterize NaCl-induced osmotic and turgor adjustment in in vitro cultures, aT. Pl. Physiol. 136, 15 23.
2. Ben-Hayyim G. (1987) Relationship between salt tol- erance and resistance to polyethylene glycol-induced water stress in cultured citrus cells. Pl. PhysioL 85, 430-433.
3. Ben-Hayyim G. and KochbaJ. (1983) Aspects of salt tolerance in a NaC1 selected stable cell line of Citrus sinensis. Pl. Physiol. 72, 685-690.
4. Binzel M.L., Hasegawa P.M., Handa A.K. and Bres- san R.A. (1985) Adaptation of tobacco cells to NaC1. Pl. Physiol. 79, 118-125.
5. Binzel M.L., Hasegawa P.M., Rhodes D, Handa S., Handa A.K. and Bressan R.A. (1987) Solute accumulation in tobacco cells adapted to NaC1. Pl. Physiol. 84, 1408-1415.
6. Bowman W.D. (1988) Ionic and water relations responses of two populations of a non-halophyte to salinity. 07. Exp. Bot. 39, 97-105.
7. Bressan R.A., Handa A.K., Handa S. and Hasegawa P.M. (1982) Growth and water relations of cultured tomato cells after adjustment to low external water potential. Pl. Physiol. 70, 1303-1309.
8. Bressan R.A., Nelson D., Iraki N., LaRosa P., Singh N., Hasegawa P. and Capita N. (1990) Reduced cell expansion and changes in cell walls of plant cells

GROWTH, PROLINE ACCUMULATION AND WATER 91
adapted to NaC1. Pages 137-171 in F. Katterman, ed. Environmental injury to plants. Academic Press, Inc., San Diego.
9. Briscoe R.D. (1984) Thermocouple psychrometers for water potential measurements. Pages 1-14 in Proc. NA TO Advanced Study Institute on Advanced Agricultural Instrumentation. II Ciocco (Pisa), Italy.
10. Chandler S.F. and Thorpe T.A. (1987) Charac- terization of growth, water relations, and proline accumulation in sodium sulfate tolerant callus of Brassica napus L. cv. Westar (Canola). Pl. Physiol. 84, 106-111.
11. Chandler S.F., Mandal B.B. and Thorpe T.A. (1986) Effect of sodium sulphate on tissue cultures of Brassica napus cv. Westar and Brassica campestris, cv. Tobin. J. Pl. Physiol. 126, 105 117.
12. Chandler S.F. and Thorpe T.A. (1986) Variation from plant tissue cultures: Biotechnological appli- cation to improving salinity tolerance. Biotechnol. Adv. 4, 117-135.
13. Chandler S. and Vasil I.K. (1984) Selection and characterization of NaC1 tolerant cells from embryo- genic cultures ofPennisetumpurpureum Schum. (Napier Grass). Pl. Sci. Lett. 37, 157-164.
14. Conger B.V. and Hanning G.E. (1991) Registration of Embryogen-P orchardgrass germplasm with a high capacity for somatic embryogenesis from in vitro cultures. Crop Sci. 31,855.
15. Croughan T.P., Stavarek SJ. and Rains D.W. (1978) Selection of a NaCI tolerant line of cultured alfalfa cells. Crop Sci. 18, 959-963.
16. Danies RJ. and Gould A.R. (1985) The cellular basis of salt tolerance studied with tissue cultures of the halophytic grass lhstichlis spicata. J. Pl. Physiol. 119, 269 280.
17. Dix PJ. and Pearce R.S. (1981) Proline accumulation in NaCl-resistant and sensitive cell lines of Aqcotiana sylvestris. Z. Pflanzenphysiol. 102, 243-248.
18. Dracup M. (1991) Increasing salt tolerance of plants through cell culture requires greater understanding of tolerance mechanisms. Aust. J. Pl. Physiol. 18, 1 15.
19. Epstein E. and Rains D.W. (1987) Advances in salt tolerance. Pages 113 125 in W.H. Gabelman and B.C. Loughman, eds. Genetic aspects of plant mineral nutrition. Martinus Nijhof Publs. Dordrecht, Boston, Lancaster.
20. Gibbs J., Dracup M., Greenway H. and McComb J.A. (1989) Effects of high NaC1 on growth, turgor and internal solutes of tobacco callus. J. Pl. Physiol. 134, 61-69.
21. Glenn E.P. and O'LearyJ.W. (1984) Relationship between salt accumulation and water content of dicotyledonous halophytes. Pl. Cell Envir. 7, 253 261.
22. Hasegawa P.M., Bressan R.A. and Handa A.K. (1986) Cellular mechanisms of salinity tolerance. Hort Sci. 21, 1317-1324.
23. Hassan N.S. and Wilkins D.A. (1988) In vitro selection for salt tolerant fines in Lycopersiconperuvianum. Pl. Cell Rep. 7, 463-466.
24. Heyser J.W. and Nabors M.W. (1981) Osmotic adjustment of cultured tobacco cells (Nicotiana tabacum var. Samsum) grown on sodium chloride. Pl. Physiol. 67, 720-727.
25. Jennings D.H. (1976) The effect of sodium chloride on higher plants. Biol. Rev. 51,453-486.
26. KochbaJ., Ben-Hayymin G., Speigel-Roy P., Saad S. and Neumann H. (1982) Selection of stable salt- tolerant callus cell lines and embryos in Citrus sinensis and C. aurantium. Z. Pflanzenphysiol. 106, 111 118.
27. Kumar V. and Sharma D.R. (1989) Isolation and characterization of sodium chloride-resistant callus culture of Vigna radiata (L.) Wilczek var. radiata. J. Exp. Bot. 40, 143-147.
28. Lerner H.R. (1985)Adaptation to salinityat theplant cell level. Pl. Soil 89, 3-14.
29. Meyer R.F. and BoyerJ.S. (1981) Osmoregulation, solute distribution, and growth in soybean seedlings having low water potentials. Planta 151,482-489.
30. Nabors M.W. (1990)Environmental stress resistance. Pages 167-186 in PJ. Dix, ed. Plant cell line selection. VCH Verlagsgesellschaft mbH, Weinheim, Germany.
31. Pandey R. and Ganapathy P.S. (1985) The proline enigma: NaCl-tolerant and NaCl-sensitive callus lines of Cicer aTietinum. Pl. Sci. 40, 13-17.
32. Pau E.-C. and Thorpe T.A. (1986) Differential response of non-selected and Na2SO4- selected cal- lus cultures of Beta vulgaris L. to salt stress. J. Pl. Physiol. 123, 241-248.
33. Peak K.Y., Chandler S.F. and Thorpe T.A. (1988) Physiological effects of Na2SO4 and NaC1 on callus cultures of Brassica campestris (Chinese cabbage). Physiol. Pl. 72, 160 166.
34. Rains D.W., Croughan T.P. and Stavarek SJ. (1980) Selection of salt-tolerant plants using tissue culture. Pages 279 292 in D.W. Rains, R.C. Valentine and A. Hollaender, eds. Genetic engineering of osmoregulation. Plenum Press, New York,
35. Rains D.W., Croughan S.S. and Croughan T.P. (1986) Isolation and characterization of mutant cell lines and plants: Salt tolerance. Pages 537-547 in I.K. Vasil, ed. Cell culture and somatic cell genetics of plants, Vol. 3. Plant regeneration and genetic variability. Academic Press, New York.
36. Schenk R.U. and Hildebrandt A.C. (1972) Medium and techniques for induction and growth of mono- cotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 50, 199-204.

92 S. DUTTA GUPTA et al.
37. Singh T.N., Paleg L.C. and Aspinall D. (1973) Stress metabolism. I. Nitrogen metabolism and growth in the barley plant during water stress. Aust. 07. Biol. Sci. 269 45-56.
38. Tal M., Katz A., Heikin M. and Dehan K. (1979) Salt tolerance in the wild relatives of the cultivated tomato: proline accumulation in Lycopersicon escu- lentum Mill., L. peruvianum Mill. and Solanum pennelli Cor. treated with NaC1 and polyethylene glycol. New Phytol. 82, 349-355.
39. Tyree M.T. andJarvis P.G. (1982) Water in tissues and cells. Pages 35-77 in O.L. Lange, P.S. Nobel, C.B. Osmond, H. Ziegler eds. Encyclopedia of plant physiolog~ 12B, physiological plant ecolog~ II. Springer- Verlag, New York.
40. Watad A.A., Reinhold L. and Lerner H.R. (1983) Comparison between a stable NaCl-selectedNicotiana cell line and the wild type. K +, Na +, and proline pools as a function of salinity. Pl. Physiol. 75~ 624- 629.
Related Documents











