GLUTAMATE RECEPTORS IN THE OLFACTORY BULB SYNAPTIC CIRCUITRY: HETEROGENEITY AND SYNAPTIC LOCALIZATION OF N-METHYL--ASPARTATE RECEPTOR SUBUNIT 1 AND AMPA RECEPTOR SUBUNIT 1 M. GIUSTETTO,* P. BOVOLIN,² A. FASOLO,² M. BONINO,² D. CANTINO* and M. SASSOE v -POGNETTO*‡ *Department of Human Anatomy and Physiology, Corso Massimo d’Azeglio 52, I-10126, Turin, Italy ²Department of Animal Biology, Via Accademia Albertina 17, I-10124, Turin, Italy Abstract––In this study, we analysed the molecular heterogeneity and synaptic localization of the N-methyl--aspartate receptor subunit 1 and the Æ-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptor subunit 1 in the olfactory bulb glomerular synaptic circuitry. Semiquantitative reverse transcriptase polymerase chain reaction showed that approximately 40% of the N-methyl--aspartate receptor subunit 1 messenger RNA splice variants contain the N1 exon, which conveys specific functional properties on the channel. In other forebrain and hindbrain regions that we examined, the ratio of the N1-containing (receptor subunit 1 1XX ) to N1-lacking (receptor subunit 1 0XX ) N-methyl--aspartate receptor subunit 1 messenger RNAs varied considerably. The cellular and subcellular distribution of N-methyl--aspartate receptor subunit 1 and AMPA receptor subunit 1 was investigated with antibodies generated against the C-terminal domain of the individual subunits [Petralia R. S. et al. (1994) J. Neurosci. 14, 667–696; Wenthold R. J. et al. (1992) J. biol. Chem. 267, 501–507]. Both N-methyl--aspartate receptor subunit 1 and AMPA receptor subunit 1 were localized to the postsynaptic density of asymmetric synapses established by olfactory receptor neuron terminals with the dendrites of mitral and tufted cells. Not all of these synapses, however, were labelled. These results are consistent with the notion that glutamate is the neurotransmitter at the olfactory nerve to mitral and tufted cell synapses, and suggest a high heterogeneity in the expression of the postsynaptic glutamate receptors. Copyright ? 1996 IBRO. Published by Elsevier Science Ltd. Key words: NMDA receptors, AMPA receptors, reverse transcriptase polymerase chain reaction, electron-microscopic immunocytochemistry, olfactory receptor neurons, rat. Olfactory receptor neurons (ORNs), located in the nasal mucosa, transduce odorant signals into im- pulses which are transmitted via the olfactory nerve to the glomerular layer of the main olfactory bulb. 40 At this level, the axon terminals of ORNs synapse on the apical dendrites of the primary output neurons, mitral and tufted (M/T) cells, and a class of intrinsic neurons, the periglomerular cells (reviewed by Greer 11 ). Recent studies indicate that glutamate may mediate neurotransmission between olfactory nerve fibres and postsynaptic neurons. Glutamate is enriched in ORN terminals 5,18,35 and application of glutamate results in a rapid and sustained depolarization of M/T cells. 2 The depolar- izing potential consists of two components, which can be blocked by non-N-methyl--aspartate (NMDA) receptor and NMDA receptor antagon- ists, respectively. 2,8 The postsynaptic responses of neurons to gluta- mate are mediated through both ionotropic recep- tors, which are ligand-gated cationic channels (reviewed by Monaghan et al. 23 ), and metabotropic receptors, which are coupled to G-proteins (reviewed by Nakanishi 27 ). Ionotropic receptors have been grouped into three major classes, defined by selective interactions with their specific agonists NMDA, Æ-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) and kainate. 23 Recently, it has become clear that ionotropic glutamate receptors are oligomeric complexes composed of different subunits. Molecular cloning and expression studies have shown that the subunits co-assemble in different combinations to form hetero-oligomeric receptors, which differ in pharmacological and biophysical properties. 13,26,38 The functional diversity of ionotropic glutamate receptors is also generated by differential RNA splic- ing. Each of the cloned AMPA receptor subunits (GluR1–4) occurs in two spliced isoforms, designated flip and flop, which are differentially expressed ‡To whom correspondence should be addressed. Abbreviations: AMPA, Æ-amino-3-hydroxy-5-methyl-4- isoxazolepropionate; GluR, AMPA receptor subunit; M/T, mitral/tufted; NMDA, N-methyl--aspartate; NR, NMDA receptor subunit; ORN, olfactory receptor neuron; PCR, polymerase chain reaction; RT, reverse transcriptase. Pergamon Neuroscience Vol. 76, No. 3, pp. 787–798, 1997 Copyright ? 1996 IBRO. Published by Elsevier Science Ltd Printed in Great Britain 0306–4522/97 $17.00+0.00 PII: S0306-4522(96)00285-0 787

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GLUTAMATE RECEPTORS IN THE OLFACTORYBULB SYNAPTIC CIRCUITRY: HETEROGENEITY AND
SYNAPTIC LOCALIZATION OF N-METHYL--ASPARTATERECEPTOR SUBUNIT 1 AND AMPA RECEPTOR SUBUNIT 1
M. GIUSTETTO,* P. BOVOLIN,† A. FASOLO,† M. BONINO,† D. CANTINO* andM. SASSOEv-POGNETTO*‡
*Department of Human Anatomy and Physiology, Corso Massimo d’Azeglio 52, I-10126, Turin, Italy
†Department of Animal Biology, Via Accademia Albertina 17, I-10124, Turin, Italy
Abstract––In this study, we analysed the molecular heterogeneity and synaptic localization of theN-methyl--aspartate receptor subunit 1 and the á-amino-3-hydroxy-5-methyl-4-isoxazolepropionate(AMPA) receptor subunit 1 in the olfactory bulb glomerular synaptic circuitry. Semiquantitative reversetranscriptase polymerase chain reaction showed that approximately 40% of the N-methyl--aspartatereceptor subunit 1 messenger RNA splice variants contain the N1 exon, which conveys specific functionalproperties on the channel. In other forebrain and hindbrain regions that we examined, the ratio of theN1-containing (receptor subunit 11XX) to N1-lacking (receptor subunit 10XX) N-methyl--aspartatereceptor subunit 1 messenger RNAs varied considerably. The cellular and subcellular distribution ofN-methyl--aspartate receptor subunit 1 and AMPA receptor subunit 1 was investigated with antibodiesgenerated against the C-terminal domain of the individual subunits [Petralia R. S. et al. (1994) J. Neurosci.14, 667–696; Wenthold R. J. et al. (1992) J. biol. Chem. 267, 501–507]. Both N-methyl--aspartate receptorsubunit 1 and AMPA receptor subunit 1 were localized to the postsynaptic density of asymmetric synapsesestablished by olfactory receptor neuron terminals with the dendrites of mitral and tufted cells. Not all ofthese synapses, however, were labelled.These results are consistent with the notion that glutamate is the neurotransmitter at the olfactory nerve
to mitral and tufted cell synapses, and suggest a high heterogeneity in the expression of the postsynapticglutamate receptors. Copyright ? 1996 IBRO. Published by Elsevier Science Ltd.
Key words: NMDA receptors, AMPA receptors, reverse transcriptase polymerase chain reaction,electron-microscopic immunocytochemistry, olfactory receptor neurons, rat.
Olfactory receptor neurons (ORNs), located in thenasal mucosa, transduce odorant signals into im-pulses which are transmitted via the olfactory nerveto the glomerular layer of the main olfactorybulb.40 At this level, the axon terminals of ORNssynapse on the apical dendrites of the primaryoutput neurons, mitral and tufted (M/T) cells, and aclass of intrinsic neurons, the periglomerular cells(reviewed by Greer11). Recent studies indicate thatglutamate may mediate neurotransmission betweenolfactory nerve fibres and postsynaptic neurons.Glutamate is enriched in ORN terminals5,18,35 andapplication of glutamate results in a rapid andsustained depolarization of M/T cells.2 The depolar-izing potential consists of two components, whichcan be blocked by non-N-methyl--aspartate
(NMDA) receptor and NMDA receptor antagon-ists, respectively.2,8
The postsynaptic responses of neurons to gluta-mate are mediated through both ionotropic recep-tors, which are ligand-gated cationic channels(reviewed by Monaghan et al.23), and metabotropicreceptors, which are coupled to G-proteins (reviewedby Nakanishi27). Ionotropic receptors have beengrouped into three major classes, defined by selectiveinteractions with their specific agonists NMDA,á-amino-3-hydroxy-5-methyl-4-isoxazolepropionate(AMPA) and kainate.23 Recently, it has become clearthat ionotropic glutamate receptors are oligomericcomplexes composed of different subunits. Molecularcloning and expression studies have shown that thesubunits co-assemble in different combinations toform hetero-oligomeric receptors, which differ inpharmacological and biophysical properties.13,26,38
The functional diversity of ionotropic glutamatereceptors is also generated by differential RNA splic-ing. Each of the cloned AMPA receptor subunits(GluR1–4) occurs in two spliced isoforms, designatedflip and flop, which are differentially expressed
‡To whom correspondence should be addressed.Abbreviations: AMPA, á-amino-3-hydroxy-5-methyl-4-isoxazolepropionate; GluR, AMPA receptor subunit;M/T, mitral/tufted; NMDA, N-methyl--aspartate; NR,NMDA receptor subunit; ORN, olfactory receptorneuron; PCR, polymerase chain reaction; RT, reversetranscriptase.
Pergamon
Neuroscience Vol. 76, No. 3, pp. 787–798, 1997Copyright ? 1996 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain0306–4522/97 $17.00+0.00PII: S0306-4522(96)00285-0
787

throughout the brain.37 In situ analyses have shownthat the olfactory bulb predominantly expresses theflop version of GluR mRNAs.37 Among the NMDAreceptors, the NR1 subunit arises from a single genein which three exons (N1, C1, C2) may be alterna-tively spliced, giving rise to a total of eight isoforms,seven of which have been identified in cDNA libraries(reviewed by Zukin and Bennett48). The NR1 splicevariants have been called NR1000-111, where the ‘‘0’’and the ‘‘1’’ subscripts indicate the absence or pres-ence of each of the three exons48 (alternative nameshave been employed by Durand et al.,6,7 Hollmann etal.,12 Nakanishi et al.28 and Sugihara et al.39). It hasbeen shown that differential RNA splicing of theNR1 subunit alters the structural, physiological andpharmacological properties of homomeric as well asheteromeric NMDA receptors.48 In particular, ex-pression of the N1 exon conveys specific functionalproperties on the channel, including a greatly en-hanced responsiveness to protein kinase C and largercurrent amplitudes.48
There is evidence that different types of glutamatereceptors are present at various locations in theolfactory bulb synaptic circuitry (reviewed byTrombley and Shepherd40). In situ hybridizationand immunocytochemical studies have identifiedseveral glutamate receptor subunits which havespecific cellular and laminar distribution pat-terns.10,15,16,21,22,24,25,30–32,43 However, the preciselocalization of these subunits within the synapticcircuits of the main olfactory bulb is not known. Arecent study42 has examined the expression of twometabotropic glutamate receptors (mGluR1á andmGluR5) in the rat olfactory bulb. Here we focus onionotropic receptors. The aim of the present study istwo-fold. First, we analysed by reverse transcriptasepolymerase chain reaction (RT-PCR) the expressionand relative abundance of N1-containing (NR11XX)and N1-lacking (NR10XX) NR1 mRNAs in the olfac-tory bulb, as well as in other brain regions. Second,we investigated, using immunocytochemistry, thelocalization of the NR1 subunit of the NMDA recep-tor and the GluR1 subunit of the AMPA receptor inthe olfactory bulb, in particular in the glomerularsynaptic circuitry.
EXPERIMENTAL PROCEDURES
Reverse transcriptase polymerase chain reaction
Adult Wistar rats (10–12 weeks old) were killed underanaesthesia and different brain regions were dissected outand frozen on dry ice. Total RNA was isolated from thefrozen tissue by the method of Chomczymsky and Sacchi.4
The yield of total RNA was determined by measuring theabsorbance of an aliquot of the ethanol-precipitated stockat 260/280 nm. Oligonucleotide primers for PCR were syn-thesized using an Applied Biosystem Inc. model 391 DNAsynthesizer. Primers were designed from the publishedsequence of the rat NR1 cDNA25 in order to yield PCRproducts spanning the insertion site of the N1 alternativelyspliced exon.48 To minimize variability in ampli-fication reactions, primers were designed so that they hadequal length (24 bp) and the same GC content (50%).
Primer sequences were 5*-GTATCCACCTGAGTTTCCTTCGCA-3* (nucleotides 395–419) and 5*-CTCGCTTGCAGAAAGGATGATGAC-3* (nucleotides 653–675). One microgram of total RNA was reverse-transcribedwith 200 units of M-MLV reverse transcriptase (Life Tech-nologies) in 50 mM Tris–HCl, pH 8.3, 75 mM KCl, 3 mMMgCl2, 1 mM deoxynucleotide triphosphates (dNTPs,Pharmacia) and 2.5 mM random hexamers (Pharmacia) in afinal volume of 20 µl. The reaction mixture was incubated at37) for 1 h, heat denatured at 95)C for 5 min and diluted 1:4in sterile water. Single cDNA aliquots or series of aliquotscontaining increasing amounts of the reverse transcribedmaterial were amplified with Taq Polymerase (Promega) inan MJ Programmable Thermal Controller (PT-10060). Theamplification mixture contained cDNA, 1 µM of specificNR1 subunit primers, 200 µM dNTPs, 1.5 mM MgCl2,10 mM Tris–HCl, pH 9.0, 50 mM KCl, 0.1% Triton X-100and 1.5 units of Taq Polymerase in 50 µl volume. Traceamounts of [32P]dCTP (Amersham), 1–2 µCi/sample, wereadded to the reaction when subsequent quantitation wasnecessary. The mixtures were amplified for 27 or 30 cycles,where each amplification cycle consisted of a denaturationstep (94)C, 45 s), an annealing step (60)C, 1 min) and anelongation step (72)C, 1 min) with a final 15 min elongation.Amplification products were separated by agarose gel elec-trophoresis, as described elsewhere.3 The DNA bands werevisualized by ethidium bromide staining of the agarose gels.In semiquantitative experiments, individual bands wereexcised and counted by Cerenkov counting of the incorpor-ated radioactivity. The ratios of counts incorporated intothe NR10XX amplification product to counts incorporatedinto the amplified NR11XX product were evaluated. Resultsare expressed as means& S.E.M. of values from fourseparate determinations.
Antibodies
The antibodies used in this study were raised in rabbitsagainst synthetic peptides corresponding to the C-terminalsequence of NR1 and GluR1, respectively.31,32 The anti-bodies were purchased from Chemicon and were used atdilutions of 1:25 (anti-NR1) and 1:12.5 (anti-GluR1). Thespecificity of the antibodies has been determined by immu-noblot analyses of brain tissue or transfected cell mem-branes.31,32,44 The antibody against GluR1 recognizes boththe flip and flop versions of this subunit. The antibodyagainst NR1 recognizes four of seven known splice variants(NR1011, NR1111, NR1001, NR1101) of this receptor sub-unit.32,48 The mRNAs encoding for these splice variants areall abundantly expressed in the olfactory bulb.17
Immunocytochemistry
Adult Wistar rats were deeply anaesthetized with Keta-mine (100 mg/kg) and Xylazine (5 mg/kg) and transcardiallyperfused with 0.01 M phosphate-buffered saline at pH 7.4,followed by the fixative solution (4% paraformaldehydewith or without 0.1% glutaraldehyde in phosphate buffer).The olfactory bulbs were removed, postfixed for 1 h in thesame fixative and washed in phosphate buffer. They werethen embedded in agar–agar and sectioned transversallywith a Vibratome at 70 µm. The sections were cryoprotectedin 30% sucrose and repeatedly frozen and thawed to en-hance antibody penetration. They were then collected incold phosphate-buffered saline and processed free-floatingaccording to the avidin–biotin–peroxidase method, as de-scribed elsewhere.36 Following incubation in the primaryand secondary antibodies, the sections were reacted in3,3*-diaminobenzidine, and then the diaminobenzidine reac-tion product was silver intensified and gold toned.36 Somesections were mounted on glass slides, coverslipped andobserved and photographed with a Zeiss photomicroscope(Axiophot). The other sections were postfixed with 1%osmium tetroxide, dehydrated in acetone and flat-embeddedin Epon 812. Serial ultrathin sections were taken
788 M. Giustetto et al.

perpendicular to the plane of the Vibratome sections. Theywere stained with uranyl acetate and lead citrate andexamined at 80 kV with a Philips EM 410 electronmicroscope.To assess non-specific staining due to the immunoproce-
dure, in every experiment some control sections were incu-bated in buffer without primary antibodies. This procedurealways resulted in a complete lack of immunoreactivity.
RESULTS
NR10XX versus NR11XX messenger RNA in olfactorybulb and other brain regionsIn order to study the splicing heterogeneity of the
N1 exon (=63 bp) in the olfactory bulb by RT-PCRwe chose a pair of primers flanking the splicingjunctions (Fig. 1A). Amplification of cDNA gener-ated two PCR products migrating on agarose gels astwo bands of the expected size: a lower band of280 bp (NR10XX) and an upper band of 343 bp(NR11XX), each corresponding to four of the eightdifferent splicing products of the NR1 gene.12,48 In
preliminary experiments performed on olfactory bulbtotal RNA, we observed that NR10XX and NR11XXmRNAs were both expressed (Fig. 1B). However, theNR10XX to NR11XX ratio appeared to be lowercompared to the 5:1 ratio suggested previously forthe adult rat forebrain.17,28,39 This prompted us toextend the RT-PCR analysis to other forebrain re-gions (hippocampus, cerebral cortex, striatum) andto hindbrain regions (cerebellum, brainstem). Figure2A shows that 27 cycles, with the PCR cyclingconditions described in Experimental Procedures, arenecessary for mRNA detection in all tissues exceptthe brainstem, whose optimal detection needs 30cycles. As estimated by both ethidium bromide stain-ing (Fig. 2A) and radioactive incorporation (Fig. 2B),the highest amounts of NR10XX mRNA are expressedin the hippocampus and striatum, while thehighest expression of the NR11XX mRNA is presentin the cerebellum. Forebrain tissues show a gen-eral prevalence of NR10XX over NR11XX mRNA,
Fig. 1. Detection of NR10XX and NR11XX splicing variants by RT-PCR. (A) Schematic diagram of NR1cDNA with the region selected for PCR amplification and corresponding PCR products. Cross-hatchedboxes represent the first three transmembrane domains predicted from hydropathy plots. Black boxescorrespond to untranslated regions. The box above the sequence bars represents the N1 63-bp insert. (B)Separation of RT-PCR products by agarose gel electophoresis. Total RNA was extracted from ratolfactory bulbs (OB), reverse transcribed and amplified in the presence of the NR1-specific primers asdescribed in Experimental Procedures. Two PCR products, corresponding to the alternatively spliced
products schematized in A, are visible. The size marker (M) is a 1-kb DNA ladder.
Glutamate receptors in rat olfactory bulb 789

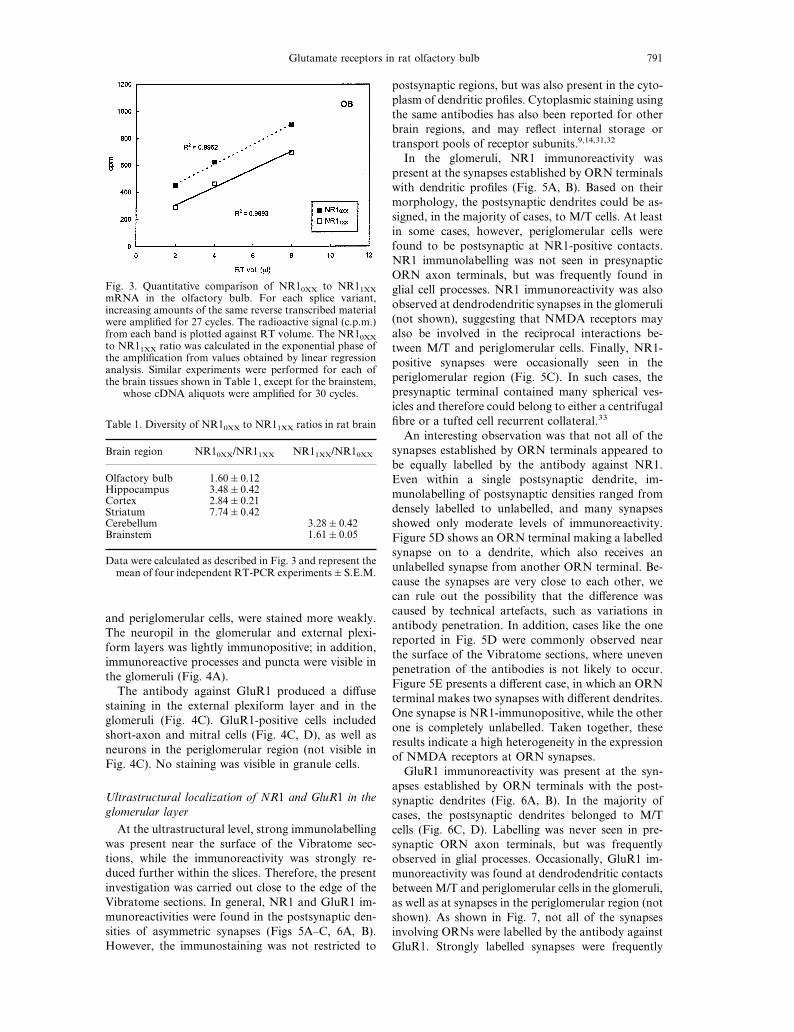
although at different ratios. In order to be able tomake more precise quantitative statements, we per-formed semiquantitative PCR on increasing concen-trations of each cDNA sample. The NR10XX/NR11XXratio was measured during the exponential phase ofthe amplification reaction, and Fig. 3 shows the resultsplotted from a representative experiment performedon the olfactory bulb. A similar quantitative analysiswas done for all the other tissues and the results aresummarized in Table 1. Different forebrain regionsshow quite dissimilar NR10XX to NR11XX ratios. Theolfactory bulb shows the lowest ratio (1.6:1), suggest-ing an almost equivalent expression of NR1 subunitswith or without the N1 insert. In the striatum, expres-sion of the N1 exon is low (0XX/1XX ~ 8:1); con-versely, the cerebellum, which shows the highest
amount of N1 expression, has an NR10XX/NR11XXratio of 1:3. In conclusion, the RT-PCR experimentsshow that the NR10XX and NR11XX splice variants aredifferentially expressed in different brain regions, andthat the olfactory bulb contains high contents of bothNR10XX and NR11XX mRNAs.
Light-microscopic immunocytochemistry
The results obtained with light-microscopic im-munocytochemistry are in good agreement with thosereported in previous studies.21,22,31,32 The antibodyagainst NR1 stained all major cell types in the mainolfactory bulb (Fig. 4A, B). Dense staining wasevident in the cell bodies and major dendrites of M/Tcells, while most interneurons, including granular
Fig. 2. Representative RT-PCR experiment showing NR10XX and NR11XX mRNA expression in variousbrain regions. (A) Gel electrophoresis of RT-PCR products, each derived from distinct RNA pools.One-tenth of the reverse transcribed material was amplified for 27 (upper gel) or 30 (lower gel) PCR cycles.(B) NR10XX and NR11XX mRNA contents as determined after 27 cycles. Radiolabelled PCR productswere excised from the gel and counted. OB, olfactory bulb; CX*, whole cerebral cortex; CX**,
frontoparietal cortex; HP, hippocampus; ST, striatum; CB, cerebellum; BS, brainstem.
790 M. Giustetto et al.
and periglomerular cells, were stained more weakly.The neuropil in the glomerular and external plexi-form layers was lightly immunopositive; in addition,immunoreactive processes and puncta were visible inthe glomeruli (Fig. 4A).The antibody against GluR1 produced a diffuse
staining in the external plexiform layer and in theglomeruli (Fig. 4C). GluR1-positive cells includedshort-axon and mitral cells (Fig. 4C, D), as well asneurons in the periglomerular region (not visible inFig. 4C). No staining was visible in granule cells.
Ultrastructural localization of NR1 and GluR1 in theglomerular layer
At the ultrastructural level, strong immunolabellingwas present near the surface of the Vibratome sec-tions, while the immunoreactivity was strongly re-duced further within the slices. Therefore, the presentinvestigation was carried out close to the edge of theVibratome sections. In general, NR1 and GluR1 im-munoreactivities were found in the postsynaptic den-sities of asymmetric synapses (Figs 5A–C, 6A, B).However, the immunostaining was not restricted to
postsynaptic regions, but was also present in the cyto-plasm of dendritic profiles. Cytoplasmic staining usingthe same antibodies has also been reported for otherbrain regions, and may reflect internal storage ortransport pools of receptor subunits.9,14,31,32
In the glomeruli, NR1 immunoreactivity waspresent at the synapses established by ORN terminalswith dendritic profiles (Fig. 5A, B). Based on theirmorphology, the postsynaptic dendrites could be as-signed, in the majority of cases, to M/T cells. At leastin some cases, however, periglomerular cells werefound to be postsynaptic at NR1-positive contacts.NR1 immunolabelling was not seen in presynapticORN axon terminals, but was frequently found inglial cell processes. NR1 immunoreactivity was alsoobserved at dendrodendritic synapses in the glomeruli(not shown), suggesting that NMDA receptors mayalso be involved in the reciprocal interactions be-tween M/T and periglomerular cells. Finally, NR1-positive synapses were occasionally seen in theperiglomerular region (Fig. 5C). In such cases, thepresynaptic terminal contained many spherical ves-icles and therefore could belong to either a centrifugalfibre or a tufted cell recurrent collateral.33
An interesting observation was that not all of thesynapses established by ORN terminals appeared tobe equally labelled by the antibody against NR1.Even within a single postsynaptic dendrite, im-munolabelling of postsynaptic densities ranged fromdensely labelled to unlabelled, and many synapsesshowed only moderate levels of immunoreactivity.Figure 5D shows an ORN terminal making a labelledsynapse on to a dendrite, which also receives anunlabelled synapse from another ORN terminal. Be-cause the synapses are very close to each other, wecan rule out the possibility that the difference wascaused by technical artefacts, such as variations inantibody penetration. In addition, cases like the onereported in Fig. 5D were commonly observed nearthe surface of the Vibratome sections, where unevenpenetration of the antibodies is not likely to occur.Figure 5E presents a different case, in which an ORNterminal makes two synapses with different dendrites.One synapse is NR1-immunopositive, while the otherone is completely unlabelled. Taken together, theseresults indicate a high heterogeneity in the expressionof NMDA receptors at ORN synapses.GluR1 immunoreactivity was present at the syn-
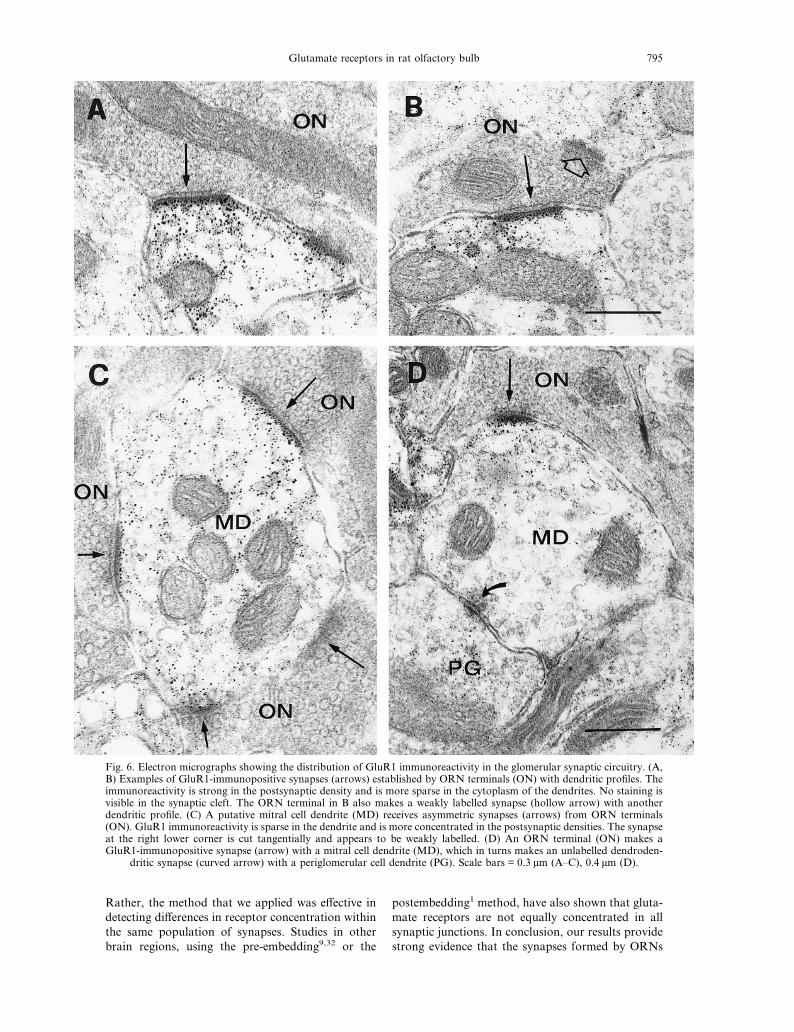
apses established by ORN terminals with the post-synaptic dendrites (Fig. 6A, B). In the majority ofcases, the postsynaptic dendrites belonged to M/Tcells (Fig. 6C, D). Labelling was never seen in pre-synaptic ORN axon terminals, but was frequentlyobserved in glial processes. Occasionally, GluR1 im-munoreactivity was found at dendrodendritic contactsbetween M/T and periglomerular cells in the glomeruli,as well as at synapses in the periglomerular region (notshown). As shown in Fig. 7, not all of the synapsesinvolving ORNs were labelled by the antibody againstGluR1. Strongly labelled synapses were frequently
Fig. 3. Quantitative comparison of NR10XX to NR11XXmRNA in the olfactory bulb. For each splice variant,increasing amounts of the same reverse transcribed materialwere amplified for 27 cycles. The radioactive signal (c.p.m.)from each band is plotted against RT volume. The NR10XXto NR11XX ratio was calculated in the exponential phase ofthe amplification from values obtained by linear regressionanalysis. Similar experiments were performed for each ofthe brain tissues shown in Table 1, except for the brainstem,
whose cDNA aliquots were amplified for 30 cycles.
Table 1. Diversity of NR10XX to NR11XX ratios in rat brain
Brain region NR10XX/NR11XX NR11XX/NR10XX
Olfactory bulb 1.60&0.12Hippocampus 3.48&0.42Cortex 2.84&0.21Striatum 7.74&0.42Cerebellum 3.28&0.42Brainstem 1.61&0.05
Data were calculated as described in Fig. 3 and represent themean of four independent RT-PCR experiments&S.E.M.
Glutamate receptors in rat olfactory bulb 791

Fig. 4. Micrographs showing the distribution of NR1 (A, B) and GluR1 (C, D) immunoreactivities on Vibratome sectionsof the rat olfactory bulb. (A, B) Strong NR1 immunoreactivity is visible in mitral (MC) and tufted (TC) cells.Periglomerular (PG) and granule (GC) cells are stained more weakly. The glomerulus (GL) in A contains NR1-positiveprocesses and puncta. (C, D) GluR1 immunoreactivity is present in the glomerular (GL), external plexiform (EPL) andmitral cell (MC) layers. Short-axon cells (SA) in the granule cell layer (GCL) are also GluR1-positive. The granule cells are
not labelled. Scale bars = 20 µm (A, B, D), 33 µm (C).

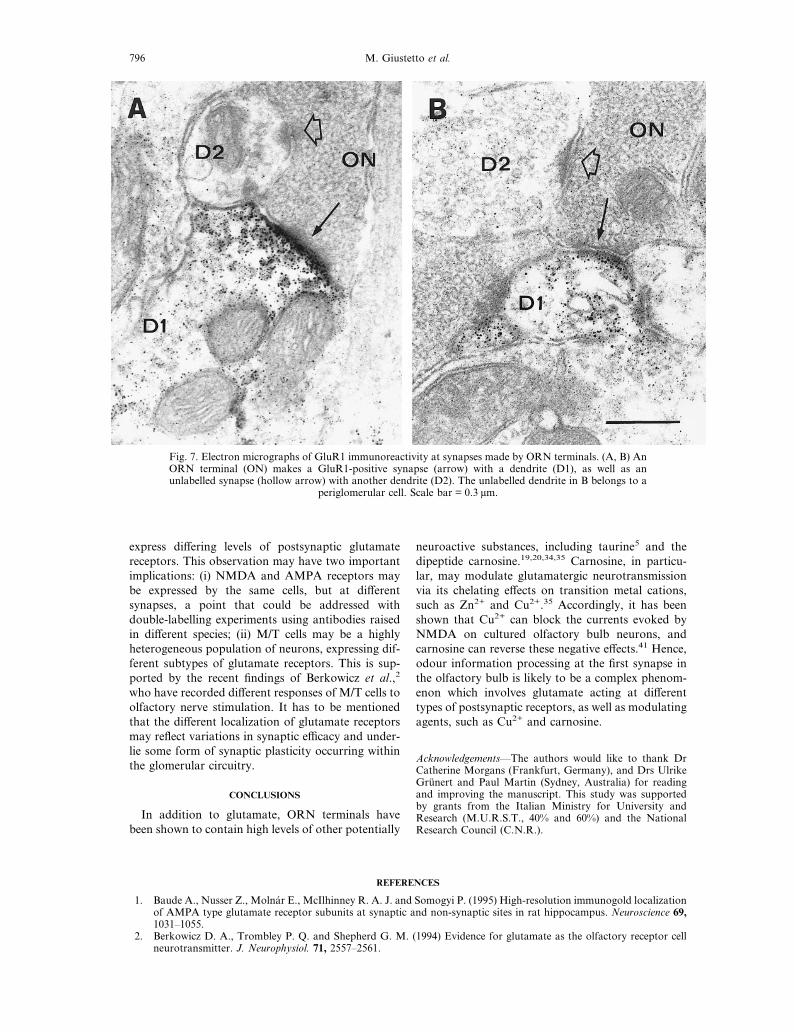
observed among other synapses which were notlabelled, even within the same postsynaptic dendrites.Figure 7A and B shows two cases in which an ORNterminal makes a GluR1-immunopositive synapse witha dendritic profile, as well as a GluR1-negative synapsewith another dendritic profile. The unlabelled dendritein Fig. 7B can be identified as a periglomerular celldendrite. These results indicate that there is great vari-ation in the expression of GluR1 at ORN synapses.
DISCUSSION
In this study we have investigated the synapticlocalization of two glutamate receptor subunits in theolfactory bulb glomeruli. The results showed thatboth the NMDA receptor subunit NR1 and theAMPA receptor subunit GluR1 are present at thesynapses established by ORN terminals with post-synaptic dendrites. A recent study42 has shown that ametabotropic glutamate receptor (mGluR1á) is alsoexpressed at the olfactory nerve to M/T cell synapses.Taken together, these observations are consistent withprevious data indicating that glutamate is the neuro-transmitter released by olfactory nerve fibres.2,35
It is only recently that glutamate has been regardedas the ORN transmitter. It has been shown that ORNterminals contain high levels of glutamate immuno-reactivity,35 and that exogenous application of gluta-mate mimics the response of M/T cells to olfactorynerve stimulation.2 This response consists of a rapiddepolarization of short duration, which can beblocked by the non-NMDA receptor antagonist 6,7-dinitroquinoxaline-2,3-dione, and a depolarization ofslower onset but longer duration which is sensitiveto the NMDA receptor antagonist -2-amino-5-phosphonovalerate. 2,8
Our present results confirm and extend, at a mo-lecular level, the data obtained from electrophysi-ological studies. The fast component of the M/T cellresponse may be explained in terms of an AMPAreceptor channel type, containing the GluR1 subunit.In addition, other AMPA-binding subunits of theglutamate receptor have been shown to be present inthe olfactory bulb,15,21,31 and may be co-expressedwith GluR1 to form heteromeric receptors.15,29,37
The prolonged depolarization induced in M/T cellsby the application of glutamate results from theactivation of NMDA receptors. NMDA receptorsare heteromeric complexes containing NR1 com-bined with one or more NR2 subunits.13,26 The NR2subunits are important in determining the pharmaco-logical and electrophysiological properties of theNMDA receptor channels.13,16,24,26 The main olfac-tory bulb has been shown to express all of the clonedNR2 subunits (NR2A–D), which have specific cellu-lar distribution patterns.43 Therefore, the olfactorybulb may contain different subtypes of NMDAreceptors, each with different response properties.This complexity is further increased by the exist-
ence of at least seven isoforms of the NR1 subunit,
arising from the alternative splicing of three exons.Exon 5 encodes an insert of 21 amino acids (termedN1) that is present in the predicted amino-terminusdomain. Recent studies6,7,12,45,46 have demonstratedthat the presence of the N1 insert reduces affinity foragonist, increases current amplitude, favours poten-tiation by protein kinase C activators and eliminatespolyamine potentiation at saturating concentrationsof glycine. On the other hand, splicing out the N1exon produces marked potentiation by Zn2+. Assuggested previously,17,47 alternative splicing of theN1 exon is differentially regulated in various brainregions. Our quantitative RT-PCR data indicatethat, in the olfactory bulb, N1 is present in approxi-mately 40% of the total NR1 transcripts, as opposedto a lower N1 percentage in other forebrain regions,such as the hippocampus (22%), cerebral cortex(26%) and striatum (11%). Therefore, NMDA recep-tors of the olfactory bulb may constitute a highlyheterogeneous population, generated by combiningthe NR1 spliced isoforms with the four differentNR2 subunits. This also means that our immuno-cytochemical results may reflect the presence offunctionally different forms of NMDA receptors.Clearly, more anatomical and electrophysiologicalstudies are needed to determine the exact subunitcomposition of the glutamate receptor types that arepresent within the glomerular synaptic circuitry, aswell as their involvement in odour informationprocessing.Our results indicate that NMDA and AMPA re-
ceptor subunits are not equally concentrated in allsynapses established by ORN terminals. Figures 5and 7 show examples of strongly labelled synapsesamong other synapses which contain scarce immuno-reactivity or no immunoreactivity at all. Even withinindividual dendrites, not all of the synapses estab-lished by ORN terminals were labelled by the anti-bodies against NR1 or GluR1. The possibility thatthis result is an artefact of the pre-embedding immu-noperoxidase method must be taken into consider-ation. Recently, Baude et al.1 have compared theefficacy of the immunoperoxidase and the postem-bedding immunogold methods for the ultrastructurallocalization of AMPA-type glutamate receptors.While the pre-embedding method is highly sensitive,it can suffer from technical limitations, such as pooraccessibility of the antibodies due to penetrationproblems and diffusion of the immunoperoxidasereaction product, which may result in secondarydeposition of label. The present analysis was carriedout near the surface of the Vibratome sections, whereheavy labelling was present in most dendritic profiles,reflecting an even penetration of the antibodies. Inaddition, variations in antibody penetration cannotexplain the presence of both labelled and unlabelledsynapses within the same dendrite (Fig. 5D). Ourresults also clearly indicate that the immunoperoxi-dase reaction product did not diffuse aspecifically toall synapses within the same postsynaptic element.
Glutamate receptors in rat olfactory bulb 793
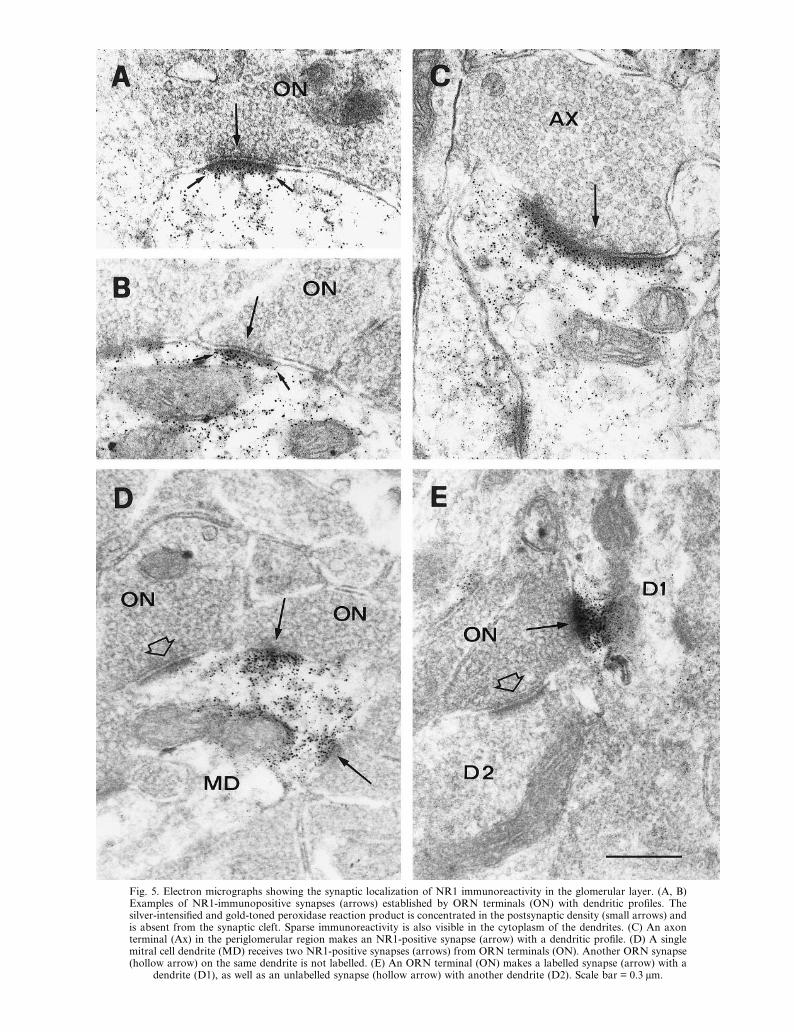
Fig. 5. Electron micrographs showing the synaptic localization of NR1 immunoreactivity in the glomerular layer. (A, B)Examples of NR1-immunopositive synapses (arrows) established by ORN terminals (ON) with dendritic profiles. Thesilver-intensified and gold-toned peroxidase reaction product is concentrated in the postsynaptic density (small arrows) andis absent from the synaptic cleft. Sparse immunoreactivity is also visible in the cytoplasm of the dendrites. (C) An axonterminal (Ax) in the periglomerular region makes an NR1-positive synapse (arrow) with a dendritic profile. (D) A singlemitral cell dendrite (MD) receives two NR1-positive synapses (arrows) from ORN terminals (ON). Another ORN synapse(hollow arrow) on the same dendrite is not labelled. (E) An ORN terminal (ON) makes a labelled synapse (arrow) with a
dendrite (D1), as well as an unlabelled synapse (hollow arrow) with another dendrite (D2). Scale bar = 0.3 µm.
Rather, the method that we applied was effective indetecting differences in receptor concentration withinthe same population of synapses. Studies in otherbrain regions, using the pre-embedding9,32 or the
postembedding1 method, have also shown that gluta-mate receptors are not equally concentrated in allsynaptic junctions. In conclusion, our results providestrong evidence that the synapses formed by ORNs
Fig. 6. Electron micrographs showing the distribution of GluR1 immunoreactivity in the glomerular synaptic circuitry. (A,B) Examples of GluR1-immunopositive synapses (arrows) established by ORN terminals (ON) with dendritic profiles. Theimmunoreactivity is strong in the postsynaptic density and is more sparse in the cytoplasm of the dendrites. No staining isvisible in the synaptic cleft. The ORN terminal in B also makes a weakly labelled synapse (hollow arrow) with anotherdendritic profile. (C) A putative mitral cell dendrite (MD) receives asymmetric synapses (arrows) from ORN terminals(ON). GluR1 immunoreactivity is sparse in the dendrite and is more concentrated in the postsynaptic densities. The synapseat the right lower corner is cut tangentially and appears to be weakly labelled. (D) An ORN terminal (ON) makes aGluR1-immunopositive synapse (arrow) with a mitral cell dendrite (MD), which in turns makes an unlabelled dendroden-
dritic synapse (curved arrow) with a periglomerular cell dendrite (PG). Scale bars = 0.3 µm (A–C), 0.4 µm (D).
Glutamate receptors in rat olfactory bulb 795
express differing levels of postsynaptic glutamatereceptors. This observation may have two importantimplications: (i) NMDA and AMPA receptors maybe expressed by the same cells, but at differentsynapses, a point that could be addressed withdouble-labelling experiments using antibodies raisedin different species; (ii) M/T cells may be a highlyheterogeneous population of neurons, expressing dif-ferent subtypes of glutamate receptors. This is sup-ported by the recent findings of Berkowicz et al.,2
who have recorded different responses of M/T cells toolfactory nerve stimulation. It has to be mentionedthat the different localization of glutamate receptorsmay reflect variations in synaptic efficacy and under-lie some form of synaptic plasticity occurring withinthe glomerular circuitry.
CONCLUSIONS
In addition to glutamate, ORN terminals havebeen shown to contain high levels of other potentially
neuroactive substances, including taurine5 and thedipeptide carnosine.19,20,34,35 Carnosine, in particu-lar, may modulate glutamatergic neurotransmissionvia its chelating effects on transition metal cations,such as Zn2+ and Cu2+.35 Accordingly, it has beenshown that Cu2+ can block the currents evoked byNMDA on cultured olfactory bulb neurons, andcarnosine can reverse these negative effects.41 Hence,odour information processing at the first synapse inthe olfactory bulb is likely to be a complex phenom-enon which involves glutamate acting at differenttypes of postsynaptic receptors, as well as modulatingagents, such as Cu2+ and carnosine.
Acknowledgements—The authors would like to thank DrCatherine Morgans (Frankfurt, Germany), and Drs UlrikeGrunert and Paul Martin (Sydney, Australia) for readingand improving the manuscript. This study was supportedby grants from the Italian Ministry for University andResearch (M.U.R.S.T., 40% and 60%) and the NationalResearch Council (C.N.R.).
REFERENCES
1. Baude A., Nusser Z., Molnar E., McIlhinney R. A. J. and Somogyi P. (1995) High-resolution immunogold localizationof AMPA type glutamate receptor subunits at synaptic and non-synaptic sites in rat hippocampus. Neuroscience 69,1031–1055.
2. Berkowicz D. A., Trombley P. Q. and Shepherd G. M. (1994) Evidence for glutamate as the olfactory receptor cellneurotransmitter. J. Neurophysiol. 71, 2557–2561.
Fig. 7. Electron micrographs of GluR1 immunoreactivity at synapses made by ORN terminals. (A, B) AnORN terminal (ON) makes a GluR1-positive synapse (arrow) with a dendrite (D1), as well as anunlabelled synapse (hollow arrow) with another dendrite (D2). The unlabelled dendrite in B belongs to a
periglomerular cell. Scale bar = 0.3 µm.
796 M. Giustetto et al.

3. Bovolin P., Santi M.-R., Memo M., Costa E. and Grayson D. R. (1992) Distinct developmental patterns of expressionof rat á1, á5, ã2 and ã2 ã-aminobutyric acidA receptor subunit mRNAs in vivo and in vitro. J. Neurochem. 59,62–72.
4. Chomczymski P. and Sacchi N. (1987) Single-step method of RNA isolation by acid guanidium thiocyanate–phenol–chloroform extraction. Analyt. Biochem. 162, 156–159.
5. Didier A., Ottersen O. P. and Storm-Mathisen J. (1994) Differential subcellular distribution of glutamate and taurinein primary olfactory neurones. NeuroReport 6, 145–148.
6. Durand G. M., Bennet M. V. L. and Zukin R. S. (1993) Splice variants of the N-methyl--aspartate receptor NR1identify domains involved in regulation by polyamines and protein kinase C. Proc. natn. Acad. Sci. U.S.A. 90,6731–6735.
7. Durand G. M., Gregor P., Zheng X., Bennet M. V. L., Uhl G. R. and Zukin R. S. (1992) Cloning of an apparent splicevariant of the rat N-methyl--aspartate receptor NMDAR1 with an altered sensitivity to polyamines and activators ofprotein kinase C. Proc. natn. Acad. Sci. U.S.A. 89, 9359–9363.
8. Ennis M., Jiang M., Zimmer L. A. and Shipley M. T. (1994) Glutamate or aspartate is a major transmitter in themammalian olfactory nerve (Abstract). Soc. Neurosci. Abstr. 20, 275.
9. Farb C. R., Aoki C. and Ledoux J. E. (1995) Differential localization of NMDA and AMPA receptor subunits in thelateral and basal nuclei of the amygdala: a light and electron microscopic study. J. comp. Neurol. 362, 86–108.
10. Gall C., Sumikawa K. and Lynch G. (1990) Levels of mRNA for a putative kainate receptor are affected by seizures.Proc. natn. Acad. Sci. U.S.A. 87, 7643–7647.
11. Greer C. A. (1991) Structural organization of the olfactory system. In Smell and Taste in Health and Disease (edsGetchell T. V. et al.), pp. 65–81. Raven Press, New York.
12. Hollmann M., Boulter J., Maron C., Beasley L., Sullivan J., Pecht G. and Heinemann S. (1993) Zinc potentiatesagonist-induced currents at certain splice variants of the NMDA receptor. Neuron 10, 943–954.
13. Hollmann M. and Heinemann S. (1994) Cloned glutamate receptors. A. Rev. Neurosci. 17, 31–108.14. Jaarsma D., Wenthold R. J. and Mugnaini E. (1995) Glutamate receptor subunits at mossy fiber–unipolar brush cell
synapses: light and electron microscopic immunocytochemical study in cerebellar cortex of rat and cat. J. comp.Neurol. 357, 145–160.
15. Keinanen K., Wisden W., Sommer B., Werner P., Herb A., Verdoorn Todd A., Sakmann B. and Seeburg P. H. (1990)A family of AMPA-selective glutamate receptors. Science 249, 556–560.
16. Kutsuwada T., Kashiwabuchi N., Mori H., Sakimura K., Kushiya E., Araki K., Meguro H., Masaki H., KumanishiT., Arakawa M. and Mishina M. (1992) Molecular diversity of the NMDA receptor channel. Nature 358, 36–41.
17. Laurie D. J. and Seeburg P. H. (1994) Regional and developmental heterogeneity in splicing of the rat brainNMDAR1 mRNA. J. Neurosci. 145, 3180–3194.
18. Liu C.-J., Grandes P., Matute C., Cuenod M. and Streit P. (1989) Glutamate-like immunoreactivity revealed in ratolfactory bulb, hippocampus and cerebellum by monoclonal antibody and sensitive staining method. Histochemistry90, 427–445.
19. Margolis F. L. (1974) Carnosine in the primary olfactory pathway. Science 184, 909–911.20. Margolis F. L. (1978) Carnosine. Trends Neurosci. 1, 42–44.21. Martin L. J., Blackstone C. D., Levey A. I., Huganir L. R. and Price D. L. (1993) AMPA glutamate receptor subunits
are differentially distributed in rat brain. Neuroscience 53, 327–358.22. Molnar E., Baude A., Richmond S. A., Patel P. B., Somogyi P. and McIlhinney R. A. J. (1993) Biochemical and
immunocytochemical characterization of antipeptide antibodies to a cloned GluR1 glutamate receptor subunit:cellular and subcellular distribution in the rat forebrain. Neuroscience 53, 307–326.
23. Monaghan D. T., Bridges R. J. and Cotman C. W. (1989) The excitatory amino acid receptors: their classes,pharmacology and distinct properties in the function of the central nervous system. A. Rev. Pharmac. Toxic. 29,365–402.
24. Monyer H., Sprengel R., Schoepfer R., Herb H., Higuchi M., Lomeli H., Burnashev N., Sakmann B. and SeeburgP. H. (1992) Heteromeric NMDA receptors: molecular and functional distinction of subtypes. Science 256, 1217–1221.
25. Moriyoshi K., Masu M., Ishii T., Shigemoto R., Mizuno N. and Nakanishi S. (1991) Molecular cloning andcharacterization of the rat NMDA receptor. Nature 354, 31–37.
26. Nakanishi S. (1992) Molecular diversity of glutamate receptors and implications for brain function. Nature 258,597–603.
27. Nakanishi S. (1994) Metabotropic glutamate receptors: synaptic transmission, modulation and plasticity. Neuron 13,1031–1037.
28. Nakanishi N., Axel R. and Shneider N. A. (1992) Alternative splicing generates functionally distinct N-methyl--aspartate receptors. Proc. natn. Acad. Sci. U.S.A. 89, 8552–8556.
29. Nakanishi N., Shneider N. A. and Axel R. (1990) A family of glutamate receptor genes: evidence for the formation ofheteromultimeric receptors with distinct channel properties. Neuron 5, 569–581.
30. Petralia R. S., Wang Y.-X. and Wenthold R. J. (1994) The NMDA receptor subunits NR2A and NR2B showhistological and ultrastructural localization patterns similar to those of NR1. J. Neurosci. 14, 6102–6120.
31. Petralia R. S. and Wenthold R. J. (1992) Light and electron immunocytochemical localization of AMPA-selectiveglutamate receptors in rat brain. J. comp. Neurol. 318, 329–354.
32. Petralia R. S., Yokotani N. and Wenthold R. J. (1994) Light and electron microscope distribution of the NMDAreceptor subunit NMDAR1 in the rat nervous system using a selective anti-peptide antibody. J. Neurosci. 14,667–696.
33. Pinching A. J. and Powell T. P. S. (1971) The neuropil of the periglomerular region of the olfactory bulb. J. Cell Sci.9, 379–409.
34. Sakai M., Kenji K., Karasawa N., Yoshida M. and Nagatsu I. (1988) Carnosine-like immunoreactivity in the olfactorybulb of the rat: an electron microscopic study. Brain Res. 438, 335–338.
35. Sassoe-Pognetto M., Cantino D., Panzanelli P., Verdun di Cantogno L., Giustetto M., Margolis F. L., De Biasi S. andFasolo A. (1993) Presynaptic co-localization of carnosine and glutamate in olfactory neurones. NeuroReport 5, 7–10.
36. Sassoe-Pognetto M., Wassle H. and Grunert U. (1994) Glycinergic synapses in the rod pathway of the rat retina: conebipolar cells express the á1 subunit of the glycine receptor. J. Neurosci. 14, 5131–5146.
Glutamate receptors in rat olfactory bulb 797

37. Sommer B., Keinanen K., Verdoorn T. A., Wisden W., Burnashev N., Herb A., Kohler M., Takagi T., Sakmann B.and Seeburg P. H. (1990) Flip and flop: a cell-specific functional switch in glutamate-operated channels of the CNS.Science 249, 1580–1585.
38. Sommer B. and Seeburg P. H. (1992) Glutamate receptor channels: novel properties and new clones. Trends pharmac.Sci. 13, 291–296.
39. Sugihara H., Moriyoshi K., Ishii T., Masu M. and Nakanishi S. (1992) Structures and properties of seven isoforms ofthe NMDA receptor generated by alternative splicing. Biochem. biophys. Res. Commun. 1853, 826–832.
40. Trombley P. Q. and Shepherd G. M. (1993) Synaptic transmission and modulation in the olfactory bulb. Curr. Opin.Neurobiol. 3, 540–547.
41. Trombley P. Q. and Shepherd G. M. (1994) Zinc, copper and carnosine modulation of amino acid receptors in theolfactory bulb (Abstract). Soc. Neurosci. Abstr. 20, 736.
42. van den Pol A. N. (1995) Presynaptic metabotropic glutamate receptors in adult and developing neurons:autoexcitation in the olfactory bulb. J. comp. Neurol. 359, 253–271.
43. Watanabe M., Inoue Y., Sakimura K. and Mishina M. (1993) Distinct distribution of five N-methyl--aspartatereceptor channel subunit mRNAs in the forebrain. J. comp. Neurol. 338, 377–390.
44. Wenthold R. J., Yokotani N., Doi K. and Wada K. (1992) Immunochemical characterization of the non-NMDAglutamate receptor using subunit specific antibodies: evidence for a hetero-oligomeric structure in rat brain. J. biol.Chem. 267, 501–507.
45. Zhang L., Zheng X., Paupard M. C., Wang A. P., Santchi L., Friedman L. K., Zukin R. S. and Bennet M. V. L. (1994)Spermine potentiation of recombinant N-methyl--aspartate receptors is affected by subunit composition. Proc. natn.Acad. Sci. U.S.A. 91, 10883–10887.
46. Zheng X., Zhang L., Durand G. M., Bennet M. V. L. and Zukin R. S. (1994) Mutagenesis rescues spermine and Zn2+
potentiation of recombinant NMDA receptors. Neuron 12, 811–818.47. Zhong J., Carrozza D. P., Williams K., Pritchett D. B. and Molinoff P. B. (1995) Expression of mRNAs encoding
subunits of the NMDA receptor in developing rat brain. J. Neurochem. 64, 531–539.48. Zukin R. S. and Bennett M. V. L. (1995) Alternatively spliced isoforms of the NMDAR1 receptor subunit. Trends
Neurosci. 18, 306–313.
(Accepted 7 May 1996)
798 M. Giustetto et al.
Related Documents











