GLUCAGON-LIKE PEPTIDE 1 (GLP-1) OG MÆTHEDSFREMMENDE INGREDIENSER GLUCAGON-LIKE PEPTIDE 1 (GLP-1) AND SATIETY PROMOTING INGREDIENTS Ida Krog Nielsen, 201407852 Agrobiologi, Fødevarevidenskab Bachelorprojekt, 15 ECTS Vejleder: Mette Skou Hedemann Aflevering: Juni 2017

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GLUCAGON-LIKEPEPTIDE1(GLP-1)OGMÆTHEDSFREMMENDEINGREDIENSER
GLUCAGON-LIKEPEPTIDE1(GLP-1)
ANDSATIETYPROMOTINGINGREDIENTS
IdaKrogNielsen,201407852Agrobiologi,FødevarevidenskabBachelorprojekt,15ECTSVejleder:MetteSkouHedemannAflevering:Juni2017

2
SammendragFedmeer igennemdesenereårtierblevetetstadigstigendeproblempåverdensplan.Dette
skyldesadskilligefaktorer,ogdeteretproblem,somerkomplekstatløse.Dererkommetet
størrefokuspåmæthedsregulerendehormonersbetydning,ogdissekanmuligvisbliveendel
afdefaktorer,somblivervæsentligeiforebyggelseogbehandlingaffedmeogovervægt.Denne
opgave omhandler mæthedshormonet glucagon-like peptide 1 (GLP-1), herunder hvordan
GLP-1 syntetiseres, frigives ognedbrydes.DerudoverbeskrivesGLP-1’s adskillige effekter i
kroppen,herunderisærdensindflydelsepåblodsukker-ogmæthedsregulering.Ibehandling
aftype2-diabetesspillerGLP-1ogsåenrolle,hvorforskelligemedicinerbeskrives.
Idesenereårerderkommetstørrefokuspåforskelligeingredienser,somkanværepotentielt
mæthedsfremmende, og dermed have betydning i forbindelse med behandling af fedme. I
denneopgaveundersøges ikke-næringsstoffetgrøn teekstrakt, sommuligkandidat iatøge
sekretionafGLP-1,ogdermedhavemættendeeffekt.Grønteekstraktbestårhovedsageligtaf
polyphenoler,ogharvistathavepositiveffektpåfedme.Tilsidstbeskriveshvordanstofferne
testes i cellemodeller, hvor der fokuseres på STC-1 og HuTu-80 celler. Cellemodeller skal
anvendesmedomhu,ogderdiskutereshvornårdekanbrugesogikkebruges.

3
Indholdsfortegnelse
SAMMENDRAG........................................................................................................................................2
INTRODUKTION......................................................................................................................................4
FORMÅL..........................................................................................................................................................4
GLUCAGON-LIKEPEPTIDE1...................................................................................................................5
SYNTESE.........................................................................................................................................................5
SEKRETION.....................................................................................................................................................6
NEDBRYDNING..............................................................................................................................................10
EFFEKTER.....................................................................................................................................................10
MÆTHEDSREGULERING..................................................................................................................................12
BEHANDLINGAFTYPE2DIABETESMEDGLP-1...............................................................................................13
POTENTIELTMÆTHEDSFREMMENDESTOFFER.................................................................................14
GRØNTEEKSTRAKT.......................................................................................................................................15
CELLEMODELLER.................................................................................................................................18
STC-1CELLER..............................................................................................................................................19
HUTU-80CELLER.........................................................................................................................................20
DISKUSSION..........................................................................................................................................21
GRØNTEEKSTRAKT.......................................................................................................................................21
ANVENDELSEAFCELLEMODELLER..................................................................................................................21
KONKLUSION........................................................................................................................................22
PERSPEKTIVERING...............................................................................................................................22
REFERENCER........................................................................................................................................23

4
IntroduktionOvervægt og fedme er steget markant de sidste år, og betragtes som et alvorligt
sundhedsproblem.Mavetarmkanaleneransvarligforatsendeenrækkemæthedssignaler,som
erudløstviabådekemiskeogmekaniskereceptorerogmedieretafbådesensoriskenerverog
dels af en række appetitregulerende hormoner. Disse hormoner er frigivet fra
mavetarmkanalen og pankreas, og kommer via cirkulation direkte i kontakt med
appetitregulerendecentreihjernen.Derudoverpåvirkerdehjernecentreneviavagusnerven.I
denneopgave fokuserespåmæthedshormonetglucagon-likepeptide1(GLP-1),der frigives
somresponspåetmåltid.EnrækkeforsøgharvistatGLP-1kanøgemæthedenognedsætte
fødeindtag,ogerderforihøjgradinteressantibekæmpelseaf fedme.GLP-1bliverarbejdet
med somkandidat tilmedicin til behandling af fedme.Derudover undersøges faktorer, der
styrerdennaturligeudskillelseafGLP-1.Dissefaktorerkanværebestemteingredienser,som
kanøgemængdenafGLP-1sekretion,ogdervedværepotentieltmæthedsfremmende.
Formål
Opgaven har til formål at beskrive GLP-1’s rolle i mæthedsreguleringen. Der beskrives
forskelligepotentieltmæthedsfremmendestoffer,somøgersekretionenafhormonetGLP-1og
hvorledesdissestoffertestesicellemodeller(bachelorkontrakt).

5
Glucagon-likepeptide1
Syntese
Glucagon-likepeptide1(GLP-1)eret tarmhormon,somsyntetiseres iL-celler i tarmene.L-
cellernefindeshovedsageligtiileumogtyktarmen,ogeråben-typeepitelceller,someridirekte
kontaktmednæringsstofferneitarmlumen.GLP-1eretkløvningsproduktafproglukagon,som
erudtrykt iL-cellerne.Derudovererproglukagonudtrykt ia-celler ipankreasogneuroner
lokaliseretihjernestammenoghypothalamus.GLP-1eret30aminosyrerpeptidhormon,som
dannesvedposttranslationelbearbejdningafproglukagoniL-cellerne(Holst2007).Derfindes
to aktive former af GLP-1, som kaldes GLP-1 (7-36) amid og GLP-1 (7-37), hvor den
førstnævnteerdominant. I figur1 sesdenposttransitionellebearbejdningafproglukagon i
forskelligevæv,hvorproglukagonbliverkløvetafprohormonconvertase1/3,somerudtrykti
L-cellerne. Her frigives GLP-1, GLP-2 og det glukagon-indeholdende peptid glicentin og
oxyntomodulin.Ia-cellerbliverproglukagonprimærtkløvetafprohormonconvertase2,som
leder til dannelse af glukagon, glicentin-relateret pankreastisk peptid (GRPP) og ’det
overordnede proglukagon fragment’ (Lim & Brubaker 2006). Derudover menes det at
prohormonconvertase1/3ia-cellerkløverproglukagontilGLP-1,dogimindreskalaendiL-
cellerne. Denne pankreas-producerede GLP-1 har muligvis parakrin virkning på de
omkringliggendeb-cellerogfremmergendannelseogformeringafb-cellerne(Whalleyetal.,
2011).
Figur 1: Vævsspecifik posttranslationel bearbejdning af proglukagon danner forskellige proglucagon-afledte peptider.Prohormonconvertase2(PC2)ia-cellerfrigiverglicentin-relateretpankreatiskpeptid(GRPP),glukagon,interventingpeptid1(IP1), og ’major proglucagon fragment’. I L-cellerne kløves proglukagon vha. prohormon convertase 1/3 og dette giverglicentin,oxyntomodulin,GLP-1,IP2,ogGLP-2(Lim&Brubaker2006).

6
Sekretion
GLP-1 udskilles fra L-cellerne i respons på næringsstoffer i et to-fase mønster, hvor den
tidligereresponsnårsithøjeste15minutterefteretmåltid,ogvareriomkring30minutter,
hvordetsene,mindre,responspeakerefter90-120minutter(Rasketal.2001).Ivoksneerden
gennemsnitlige plasmakoncentration af amideret GLP-1 omkring 5-10 pmol/L i fastende
personer,og40-50pmol/L90minutterefterindtagelseafetmåltid(Padidelaetal.2009).Idet
hovedandelenafL-cellerfindesidensidstedelaftyndtarmen,erdetusandsynligtatdenførste
faseafGLP-1udskillelsenskyldesdirektekontaktafnæringsstoffermedL-cellerne.Adskillige
studierharvistatdetautonomenervesystem,specieltvagusnerven,spillerenvigtigrolleiat
medieredentidligeudskillelseafGLP-1.Vagusnervensrolle,somenvigtigmediator,erblevet
vistpåstudierafrotter.Detblevpåvistatvagotomi,hvorvagusnervenerskåretover,blokerer
fedtinduceretGLP-1-sekretion,hvorimoddirekteelektriskstimuleringafvagusnervensceliac-
grene,somindvirkerpåjejunum,ileumogtyktarm,øgersekretionafGLP-1(Rocca&Brubaker
1999).Derudoverbidragerneurotransmitternegastrin-releasingpeptid(GRP)ogacetylcholin
(ACh),oghormonetgastriskinhibitoriskpeptid(GIP)tildenhurtigefrigivelseafGLP-1efter
indtagelseafetmåltid.UdskillelseafGLP-1herskyldesikkedirektekontaktafnæringsstoffer
medL-cellerne,menskerderimodindirektegennemenneuro-endokrinpathway,somkanses
påfigur2.
Figur2:Oversigtoverdenproximale-distaleneuendrokrinepathway,somharindflydelsepåGLP-1sekretionfrailealeL-cellerfrarotter(LimandBrubaker,2006).

7
TilatpåvisedettidligereresponsafGLP-1,erderlavetenmodelpårotter,hvornæringsstoffer
varforhindretatnåtildendistaletarm,ogdervedundgåkontaktmedL-cellerne.Glukoseeller
fedtblevplaceretiduodenum,hvilketinduceredeenomgåendeoglangvarigstimuleringafL-
celler, som kunne sammenlignes med GLP-1 sekretion niveauer, når næringsstoffer var
placeret direkte i ileum (Roberge & Brubaker 1993). Derudover viste forsøget, at når
næringsstoffervarplaceretiduodenum,blevGIPfrigivetfraK-celler.GIPaktivererafferente
vagusnerver, som stimulerer L-cellerne indirekte. Stimuleringen sker igennem efferente
vagusnerverogenteriskeneuroner,somfrigiverAChogGRP.AChharstimulerendeeffekter,
som L-cellerne er meget følsomme overfor. GRP er et neuropeptid, frigivet fra GRPergic
neuroner i det enteriske nervesystem. Disse stimulatorer forårsager i sidste ende GLP-1
frigivelse.GIPkanogsåvedhøjereniveauerstimulereL-cellernedirekte.(Rocca&Brubaker
1999).
Der findes flere mekanismer for GLP-1 sekretion, hvor sekretionen sker som respons på
fødeindtagafkulhydrater,proteinerogfedt.GlukosestimulererGLP-1sekretionvednatrium
(Na+)kobletoptagviasodium-glucosetransporter1(SGLT1),somtransportereretglukose
molekyle sammen med to natrium-ioner, hvilket forårsager cellemembrandepolarisering.
Depolariseringenaktiverervoltage-gatedecalciumkanaler,ogderstrømmerekstracellulært
calciumind,hvilketaktivererexocytoseafGLP-1.Derudoverskerderogsåoptagafglukosevia
GLUT2,somerelektroneutral.VedoptagafglukosebådegennemGLUT2ogSGLT1skerder
intracellulærmetabolismetilATP,hvilketforårsagerlukningafATP-følsommekalium(KATP)
kanaler.Dette forårsagerogså celledepolarisering,og sammemekanismesomovenfor sker.
FruktosestimulererogsåGLP-1sekretionviaGLUT-5,hvilketskerelektro-neutraltogkunATP-
afhængigt.
FedtsyrerstimulererGLP-1sekretionvedaktiveringafGaq-subunitpå forskelligeG-protein
kobledereceptorer(GPCR).SCFA(short-chainfattyacids)aktivererreceptorerneGPR41og
GPR43, hvor LCFA (long-chain fatty acids) aktiverer receptorerne GPR40 og GPR120. Her
stimuleres sekretionen ved aktivering af phospholipase C (PLC), der omdanner
phosphatidylinositol 4,5-biphosphate (PIP2) til inositol triphosphat (IP3) og diacylglycerol
(DAG). IP3 forårsager mobilisering af calcium fra intracellulære lagre, og DAG aktiverer
proteinkinaseC(PKC).BeggemekanismerforårsagerexocytosisafGLP-1.
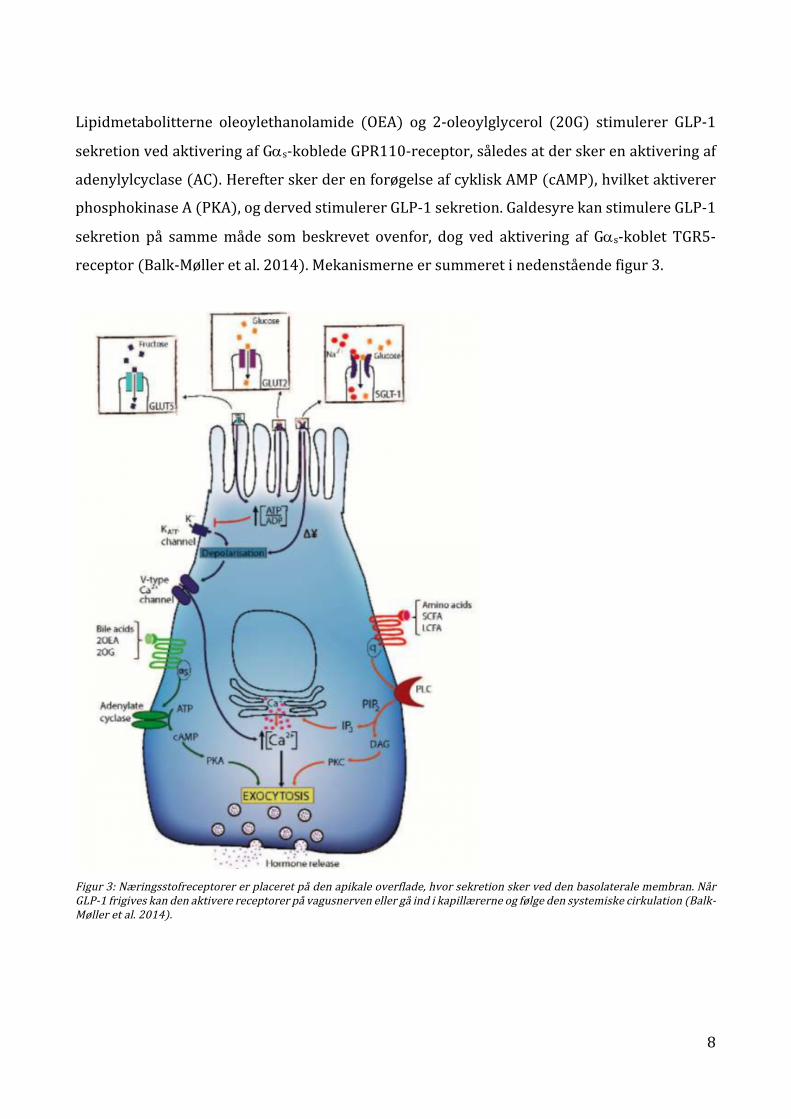
8
Lipidmetabolitterne oleoylethanolamide (OEA) og 2-oleoylglycerol (20G) stimulerer GLP-1
sekretionvedaktiveringafGas-kobledeGPR110-receptor,såledesatderskerenaktiveringaf
adenylylcyclase(AC).HerefterskerderenforøgelseafcykliskAMP(cAMP),hvilketaktiverer
phosphokinaseA(PKA),ogdervedstimulererGLP-1sekretion.GaldesyrekanstimulereGLP-1
sekretion på sammemåde som beskrevet ovenfor, dog ved aktivering af Gas-koblet TGR5-
receptor(Balk-Mølleretal.2014).Mekanismerneersummeretinedenståendefigur3.
Figur3:Næringsstofreceptorererplaceretpådenapikaleoverflade,hvorsekretionskerveddenbasolateralemembran.NårGLP-1frigiveskandenaktiverereceptorerpåvagusnervenellergåindikapillærerneogfølgedensystemiskecirkulation(Balk-Mølleretal.2014).

9
Proteiner kan lede til GLP-1 frigivelse gennem Ca2+-receptoren (CaSR), hvor bestemte
aminosyrer forårsagerøget intracellulærtCa2+ (Mace et al. 2012).Derudover findespeptid
transporteren, PEPT1, som er en brush-border cotransporter. PEPT1 transporterer di- og
tripeptiderfratarmlumenogindicellensammenmedH+.Detteledertilcelledepolariseringog
efterfølgendeåbningafvoltage-gatedecalciumkanaler,somtilsidstledertilGLP-1frigivelse
(Diakogiannakietal.2013).
Forskellige smage opfanges af smagsreceptorceller (TCR’er), som findes i smagsløgene på
tungen, og derudover udtrykkes smagsreceptorerne i L-cellerne i tarmene. Når TCR’er på
tungenaktiveresvedsmage,senderdesignalervideretilhjerneområder,somerinvolvereti
smagsopfattelse. Ved aktivering af smagsreceptorerne i tarmen frigives GLP-1. TCR’er
klassificeresifiretyper,hvortype-1celleroverførersmagensalt.Type-2cellerudtrykkerG-
protein koblede receptorer (GPCR’er), som opfatter sød, umami og bitter. Type-3 celler
udtrykker receptorer som aktiveres af sure smage, og disse receptorer er cykliske kanaler.
Type-4cellermenesatværestamceller.DerfindestoklasserindenforsmagsGPCR’er;’taste1
receptor family’ (TAS1R)og ’taste2receptor family’ (TAS2R).Sødogumami fornemmesaf
TAS1R, som er heterodimerisk og består af en familie på tre receptorer; TAS1R1, TAS1R2,
TAS1R3.TAS1R1ogTAS1R3heterodimerereceptorerbliveraktiveretafumamismag,hvor
receptorerafTAS1R2ogTAS1R3aktiveresafsødesmage(Chaudhari&Roper2010).TAS2R
receptorfamilien harmere end 25 undertyper imennesker, og reagerer på bitre fødevarer
(Meyerhofetal.2010).a-gusducin,somera-underenhedenafG-proteinet,dererkoblet til
receptorerne,spillerenrolleisignalvidereførelsenårdetdrejersigomsød,umamiogbitter
smag.DeG-proteinkobledereceptorerstimulerera-gusducin,somaktivererphospholipaseCb2
(PLCb2).PLCb2kløverPIP2tilIP3ogDAG.DannelseafIP3ledertilforhøjetcytoplasmiskCa2+,
ogaktiveringaf’transientreceptorpotentielcation-channelsubfamilyMmember5’(TRPM5),
som leder til membrandepolarisering. Kombinationen af forhøjede Ca2+-niveauer og
membrandepolariseringresultereriåbningafstoreporer(gapjunctions),somfrigiverATP.
FrigivelseafATPaktivererpurinoreceptorerpånervefibre,derledertilsmagsopfattelse.Denne
mekanismeforegårpåsmagsreceptorcellerneismagsløgenepåtungen.(Chaudhari&Roper
2010).Den sammemekanisme sker i smagsreceptorerne i L-cellerne, hvorderdog frigives
GLP-1istedetforATP(Calvo&Egan2015).

10
Nedbrydning
Når GLP-1 udskilles fra L-cellerne i tarmene nedbrydes det hurtigt af enzymet dipeptidyl
peptidase4(DPP-4),hvilketresultererienmegetkortlevetidforGLP-1.Halveringstidenfor
intaktGLP-1iplasmaer1-2minutter.DDP-4frakløveretdipeptidfraN-terminalen,hvilket
resultereriGLP-1(9-36)amidellerGLP-1(9-37),somerinaktivellervirkersomeninhibitor
vedGLP-1receptoren.Detodannedemetabolitterbliverogsånedbrudthurtigt,hovedsageligt
inyrerne,ogharenhalveringstidpåomkring4-5minutter(Holst2007).DDP-4erudtryktpå
overfladenaftarmepitelceller.DetteresultereriatmegetafGLP-1alleredenedbrydestilden
inaktivemetabolititarmene.Kunomkring25%afnyligtfrigivetGLP-1forladertarmeneien
intakt,aktivform(Hansenetal.2017).Enlignendenedbrydningpåomkring40-50%skeri
leveren, og det kan derfor beregnes at kun 10-15% af nyligt frigivet GLP-1 når ud i den
systemetiskecirkulationiintaktform(Holst,2007).
Effekter
Nårmanspiser,ogmadenkommerneditarmenedannesGLP-1,ogeffektenafGLP-1skerbåde
direktefratarmentilhjerne(vianerver)oggennemblodet.Frablodbanenfordeleshormonet
tilhelekroppen,ogvirkerpåadskilligeområderikroppen.Påfigur4seseffekterneafGLP-1.
Figur4:IllustrationafGLP-1’sfunktionerikroppen(Lundetal.2017).

11
Ibegyndelsenaf1900’erneblevdetvistatvissefaktorer,somproduceresaftarmslimhinden,
som reaktion på næringsindtagelse, kan stimulere frigivelse af stoffer fra den endokrine
pankreas,ogdervedreducereblodsukkerniveauet.Senereblevbegrebetinkretinbrugttilat
betegne disse glukose-sænkende tarmafledte faktorer. Udviklingen af radioimmunanalyse
hjalpmedatbekræftekommunikationenmellemtarmogdenendokrinepankreas,dadetblev
vist, at oral glukoseadministration giver en langt større stigning i plasma insulinniveauer
sammenlignet med den samme mængde glukose givet intravenøst. Fænomenet at insulin
responsen er større ved oral glukoseindtag, i forhold til intravenøs glukose infusion kaldes
inkretin-effekten.Detanslåsatinkretin-effekteneransvarligforomkring50-70%afdettotale
insulin udskilt efter oral glukoseadministration. (Baggio & Drucker 2007). Inkretiner er
således hormoner, der udskilles fra mave-tarmkanalen til cirkulationen, som reaktion på
næringsindtagelse, som forbedrer glukosestimuleret insulinsekretion. Hormoner der har
denneeffekt erhovedsageligtblevetbestemt til hormonerneGIPogGLP-1 (Nauck&Meier
2016).
GLP-1 virker på pankreas, som består af de langerhanske øer, der indeholder bl.a.
insulinproducerende b-celler og glukagonproducerende a-celler. Disse øer ligger spredt i
pankreas,somenslagskapslerpåblodårer,sådekanmåleblodsukkerniveauetiblodet.GLP-1
hjælper til glukosehomeostase ved at stimulere insulinsekretion (inkretin-effekt), og
derudovervedsinepankreatiskeogekstra-pankreatiskefunktioner.GLP-1erpåvistatvære
medtilatstimulereGIPudskillelse,vækstafb-celler,øgeinsulin-gentransskriptionogmRNA
stabilitet, og inhibere glukagon sekretion via direkte interaktion med GLP-1 receptoren i
pankreatiskea-celler,ogindirekteviastimuleringafsomatostatin-oginsulinsekretion.Dens
ekstra-pankreatiskeeffekter inkluderer inhiberingafmavesyreudskillelse,mavetømningog
bevægelighed, og modificering af raten af næringsstoffer, som kommer ind i
cirkulationssystemet.DermedspillerGLP-1enstor rolle i reguleringafblodsukkerniveauer
(Kim&Egan2008).DerudovererGLP-1blevetkarakteriseretsomenvigtigregulatorafappetit
og fødeindtag ved direkte stimulering anorektiske veje i hypothalamus og hjernestammen
igennemsignaleringafvagusnerven.Detteuddybesyderligereisenereafsnit.

12
Mæthedsregulering
Hypothalamus og hjernestammen er de vigtigste centralnervesystemregioner, som blandt
andet er ansvarlige for mæthedsreguleringen. Derudover spiller vagusnerven en betydelig
rolle.Figur5visersignalvejeneforGLP-1.
Figur5:Tarm-hjerneakse:reguleringaffødeindtagelse.NæringsstofferaktivererG-proteinkobledereceptorerpådenluminalesideafenteroendokrineceller f.eks.L-celler.Dennestimulering frigiver tarmhormonerheriblandtGLP-1, somkanpåvirkefødeindtagelsepåtrestadier:Vagusnerven,hjernestammenoghypothalamus.Forkortelser:GLP-1(glucagon-likepeptide1),ARC (nukleus arkuatus), AgRP (agouti-relateret peptid), NPY (neuropeptid Y), POMC (propiomelanocortin), PVN(paraventrikulærkerne),PYY(peptidYY)(Sametal.2012).
Nukleusarkuatus (ARC) ihypothalamusanses forat spilleenvigtig rolle iden fysiologiske
appetitregulering. ARC indeholder to populationer af neuroner, som ermodsatvirkende på
fødeindtagelse. De orexgeniske neuroner, som stimulerer appetit, udtrykker neuropeptid Y
(NPY)ogagouti-relateretprotein(AgRP).Derudoverfindesdeanorexigeniskeneuroner,der
hæmmer appetit og udtrykker pro-opiomelanocortin (POMC). ARC støder op til median
eminence,ogblodkarreneidenydrezoneafmedianeminenceerfenestrerede,hvilketgørat
cirkulerendehormonersomf.eks.GLP-1kankommeindivævetogpåvirkeaktivitetenafARC’s
neuronerdirekte (Chaudhri et al.2008).Tarmhormoner,heriblandtGLP-1, som frigives fra
mave-tarmkanalen, signalerer kortsigtet næringsstoftilgængelighed til ARC. Andre
cirkulerende faktorer, som insulin og leptin (et cirkulerende peptid frigivet fra fedtvæv)
videresender information vedrørende langsigtede energilagrer (Porte et al. 2002). Foruden

13
ARCmodtagerhjernestammensnucleustractussolitarus(NTS)ogareapostrema(AP)signaler
fraafferentevagusnerverogenrækkecirkulerendefaktorer.Dissecentrersamarbejdermed
hypothalamus.Mæthedssignaler,somdannesnårmanindtagerfødeogmavenudspiles,ogder
sker en aktivering af mekanoreceptorer og kemoreceptorer, sendes via vagusnerven til
centralnervesystemet om fylde i maven. Disse signaler samles i NTS, som derefter sender
signalervidere tilhypothalamus.GLP-1ogandre tarmhormonerkanpåvirkeNTS-neuroner
gennemAP,somstøderoptilNTS(Baraboietal.2010).
Behandlingaftype2diabetesmedGLP-1
Type 2 diabetes er karakteriseret ved insulinresistens, og relativ dysfunktion af b-celler i
pankreas,somførertilinsulinmangel.Sygdommenskyldes,atb-cellerneikkekanproducere
nok insulin i forbindelse med insulinresistensen. Insulinresistens vil sige, at cellerne ikke
reagerernokpåetnormaltinsulinniveau.Dettemedfører,atleverensendersukkerudiblodet,
somisidsteendeforårsageretkroniskforhøjetblodsukker(Ferrannini1998).Derudoverer
GLP-1 sekretion lavere end normalt ved type 2 diabetikere (Lugari et al. 2000). Disse
observationergørdetinteressantatbehandletype2diabetesmedGLP-1receptoragonister
ellerGLP-1analoger,pågrundafdensinkretin-effekt,ogatdenerlavtudtryktvedpatienter
med type 2 diabetes. Den hurtige nedbrydning af GLP-1 ved enzymet DPP-4 har ledt til
udvikling af nedbrydningsresistente GLP-1 receptor agonister og DPP-4 inhibitorer til
behandlingaftype2diabetes.
Indenfor nedbrydningsresistente GLP-1 receptor agonister findes Exendin-4 og Liraglutid.
Exendin-4eretnaturligtforekommendeGLP-1efterligning,someret39aminosyrerpeptid.
Deteroprindeligtisoleretfragiftetafgilaøglen,Helodermasuspectum,ogdenudviseromkring
53%aminosyreidentitetmeddetoprindeligeGLP-1,ogerenstærkagonisttilGLP-1receptoren
ipattedyr.Exendin-4indeholderenglyciniposition2,ogerderforikkesubstratforDPP-4,
hvilket forårsager en længere halveringstid end det oprindelige GLP-1 efter subkutan
administration.Exendin-4sænkerbloksukkersignifikantiprækliniskestudier,ogserudtilat
være10til100gangestærkereenddetoprindeligeGLP-1invivo,pga.afdenmegethurtigere
nedbrydningafdetoprindeligeGLP-1peptid.Exendin-4måinjicerestogangedagligt.(Baggio
&Drucker2007).Opdagelsenafexendin-4harledttiludviklingafensyntetiskversionkaldt

14
exentatid.Exentatidharenmeget længere invivohalveringstidendGLP-1ogerden første
inkretinefterligninggodkendttilbehandlingaftype2diabetes(Drucker&Nauck2006).Den
ermedtileffektivtatstimulere insulinsekretion,hæmmeglukagonogsænkeblodsukkeret.
Kliniskeforsøgharvist,atexentatiderbrugbartilbehandlingaftype2diabetes.Derudoverhar
exentatidvist,atværemedtilatreducerekropsvægtibehandledediabetikereifaseIIIkliniske
forsøg(Buseetal.2004;DeFronzoetal.2005;Kendalletal.2005).Detsessomenstorfordel,
atexentatidermedtilatgivevægttab,idetmangebehandlingeraftype2diabetesoftegiver
vægtforøgelse.Derudovererkvalmeofteenafsideeffekternevedbehandling(Murphy&Bloom
2006). Liraglutid er en human GLP-1 receptor analog, som er DPP-4 resistent og
langtidsvirkende. I kontrast til exendin-4harLiraglutid97%aminosyrerhomologimeddet
oprindeligeGLP-1.DenhartoaminosyreerstatningerogtilføjelseafenC16fedtsyrekæde,som
gørdetmuligtatbindenon-kovalenttilalbumin,ogdervedforlængedensfarmakokinetiske
profil.Liraglutidinjicereskunéngangdagligt,ogharsammeeffektersomdetoprindeligeGLP-
1(Degnetal.2004).DerudoverfindesogsåGLP-1receptoragonisten,Albiglutide,somkunskal
tageséngangugeligt(Rosenstocketal.2009).InhibitorerafGLP-1nedbrydningsenzymetDPP-
4kanværeSitagliptinellerlignendemedicin.Detkanindtagesmedføden,hvilketermedtilat
øgeplasmaniveauerneafGLP-1ogandrepeptider,somnedbrydesafDPP-4(Wagetetal.2011).
PotentieltmæthedsfremmendestofferDeterinteressant,atundersøgeomnæringsstofferellerikke-næringsstofferkanhjælpemed
atbehandlefedmevedmålrettetatstimulerenæringsstof-receptorerpåendokrineceller/L-
celler,ogdervedøgefrigivelseafGLP-1såkroppentror,atdenharindtagetetmåltid.Derer
undersøgt mange typer stoffer, som kan være potentielt mæthedsfremmende. Glukose,
proteinerogfedternæringsstoffer,somermegetundersøgtefordereseffekterpåmadindtag.
Derudover findes ikke-næringsstoffer,somogsåkanværepotentieltmæthedsfremmende. I
undersøgelsen af potentielt mæthedsfremmende stoffer tages der udgangspunkt i ikke-
næringsstoffet,grønteekstrakt,somhovedsageligtbestårafpolyphenoler.

15
GrønteekstraktBrugenafplanteekstraktertilvægttaberethurtigvoksendeterapeutiskområde,somerunder
stor undersøgelse. Grøn te brygges af tørrede blade fra planten Camellia senesis. Grøn te
indeholderstoremængderpolyphenoler,somrepræsenterer30%aftørvægtenaffriskeblade.
60-80% af disse polyphenoler er flavan-3-oler, kendt som catechiner. Hovedgrupperne af
catechinerer;(-)-epigallocatechingallate(EGCG),(-)-epigallocatechin(EGC),(-)-epicatechin
(EC)og(-)-epicathechingallat(ECG).GrønteindeholdermestafcatechinenEGCG,svarendetil
50-80% af det totale catechinindhold. Denne catechin er også kendt som den mest
sundhedsfremmende(Rainsetal.2011).
EGCGharvistathaveenreducerendeeffektpåmadindtag,ogdermedforebyggefedme(Kaoet
al.2000).EnundersøgelseafSongetal.(2015)viser,atEGCGstimulererfrigivelseafGLP-1.
Undersøgelsen havde til formål at undersøge, om polyphenoler kan stimulere sekretion af
mæthedsfremmendetarmhormoner.HertilblevderanvendtCaco-2celler,somstammerfra
humane tyktarms-cancer celler. Caco-2 cellerne blev behandlet direkte med polyphenoler,
efterfulgt af måling af frigivet hormoner i en buffer. De anvendte koncentrationer af
polyphenolerblevbestemtudfraderesopløselighedoggiftighedoverforCaco-2celler.Blandt
detestedepolyphenolerstimuleredeEGCGsignifikantGLP-1frigivelsefraCaco-2celler(p=
0,0001,figur6).
Figur6:FrigivelseafGLP-1fraCaco-2cellerstimuleretafpolyphenoler.Frigivelseblevmåltefter2timersinkubationmedpolyphenolerne. Koncentrationerne af polyphenolerne var; quercetin, apigenin, hesperidin og genistein, 100 µM; EGCG,cyanidinogresveratrol,300µM,chlorogenicacidogferulicacid,1mM,emodinogcurcumin,50µM.Kontrolerenprøve,hvorderkunertilsatassaybuffer.StatistisksignifikansblevbestemtvedDunnet’stestvedp<0,05(n=3)(Songetal.2015).

16
GLP-1 sekretionen viste sig at være dosisafhængigt. Caco-2 cellerne blev stimuleret af
forskellige doser af EGCG fra 100mM til 500mM, hvor GLP-1 sekretion viste sig at blive
induceretaf300mMsomminimumkoncentration,hvilketkansespåfigur7.
Figur7:EndosisafhængigresponsafGLP-1sekretionfraCaco-2cellerstimuleretafEGCG.StatistisksignifikansblevbestemtvedDunnet’stestvedp<0,05(n=3)(Songetal.2015).
Derudover blev det undersøgt, om EGCG stimulerede frigivelse af tarmhormoner på
vævsniveau.Detteblevudførtvedetexvivoassay,hvorduodenum,ileumogtyktarmframus
blevbrugt.Musetarmeneblevbehandleti45min.med1mMEGCG.Resultatetvistetydeligtat
GLP-1blevudskiltfraileum(p=0,006,figur8).Disseresultaterermedtilatvise,atEGCGeri
standtilatstimuleresekretionafGLP-1fradyr(Songetal.2015).
Figur8:SekretionafGLP-1framusetarmesomresponspåEGCG.Målingerblevtaget45minefterbehandlingmed(mørksøjle)elleruden(hvidsøjle)1mMEGCG.StatistisksignifikansblevbestemtvedStudentst-testvedp<0,05(n=3)(Songetal.2015).
Undersøgelsenviseraltialt,atEGCGstimulerersekretionafGLP-1fraCaco-2celler(invitro)
ogframusetarme(exvivo).DermedviserresultaterneatEGCGkanhaveenpotentielrollefor
appetitregulering og kontrol af energibalance. Dog skal resultaterne opnået fra Caco-2
cellelinjenfortolkesmedforsigtighed.Caco-2cellerstammeroprindeligt fraenkræftsvulst i

17
tyktarmen (human), men funktionelt ligner de tyndtarmsceller. Når cellerne dyrkes under
specifikke betingelserændrer de sig, så de ligner enterocytter i tyndtarmen (Hidalgo et al.
1989). Derfor kan det være en usikker cellelinje, da dyrkningen af cellerne kan påvirke,
hvordancellerneudartersig,ogdederfor ikke lignerceller franormaltvæv.Forsøgenebør
derfor formentligt gentages på andre cellelinjer, således at det er mere sandsynligt, at
resultaternekangenskabespånaturligtvæv.
Det er blevet vist at nogen ikke-næringsstoffer f.eks. polyphenoler, virker på
bittersmagsreceptorer, som hører under TAS2R (Meyerhof et al. 2010). Her er det blevet
demonstreret,atEGCGaktivererreceptorernehTAS2R14oghTAS2R39(Yamazakietal.2013;
Narukawa et al. 2011). Da disse bittersmagsreceptorer er blevet vist at inducere GLP-1
sekretion, stemmer det overens med, at EGCG kan stimulere GLP-1 sekretion, og dermed
muligvishaveindvirkningpåmæthedsreguleringogmadindtag.
IenundersøgelseafLiuetal.(2014),bleveffektenafgrønteekstrakt(GTE)testetpåpatienter
med type 2 diabetes og lipid-abnormaliteter i et randomiseret, dobbeltblindet, og placebo-
kontrolleret klinisk forsøg. Forsøgsresultater er baseret på 77 deltagere, somblev delt i to
grupper.Denenegruppe(39deltagere)indtog500mgGTE,tregangeomdagen,imensden
andengruppe(38deltagere)indtogcellulose(placebo)isammedoseroghyppighed.GTE’en
brugt til studiet var koffeinfri, og over 50% af indholdet var af catechinen EGCG. Forsøget
varedei16uger,ogniveauetafGLP-1blevmåltommorgenenefter8-9timersfaste.GLP-1
niveauetiEGCG-gruppenstegfra1.4±1.2til2.6±1.6medstatistisksignifikans(p=0.001).Der
blevikkevistnogensignifikantforskeliGLP-1niveauiplacebo-gruppen;1.9±2.3til2.2±1.8,(p
=0,60).Derudoverblevderikkeopnåetsignifikantforskelmellemgrupperne.Udoveratmåle
GLP-1niveauer,blevderbl.a.ogsåmåltpå insulinniveauerog insulinresistens. Insulin faldt
markantfra15.6±10.4til9.3±4.2(p=0.00)iEGCG-gruppen,ogfra17.0±14.8til12.3±7.5(p=
0.039) i placebo-gruppen, men der var ingen signifikant forskel mellem grupperne.
Insulinresistens-niveauer blevmålt af ’homeostatis model assessment of insulin resistance
index”(HOMA-IR).HOMA-IRindeksfaldtfra5.4±3.9til3.5±2.0medstatistisksignifikans(p=
0.004) i gruppen der indtog EGCG, hvor indekset også faldt i placebo-gruppen, men ikke
signifikant. Studiet viste altså øget GLP-1, lavere insulin og nedsat insulinresistens i EGCG-

18
gruppen.Disseresultatervardogkunsignifikanteigrupper,somhavdeværetsygeimindre
end5år.Derforkræverdetyderligeundersøgelse,hvorderbehøvesetstørreantaldeltagere,
foratundersøgedekliniskekaraktererafpatientermedbedreGLP-1responstilGTE.
Josic et al. (2010) har undersøgt om grøn te har mættende effekt i raske personer.
Undersøgelsenerbaseretpå14sundeograskedeltagereietcrossoverdesign,hvordeltagerne
tilfældigtentenfik300ml.grønteellervand.Detblevindtagetsammenmedenmorgenmad
beståendeafhvidtbrødogkalkun.Tilmålingafmæthedsfornemmelsehosdeltagernebleven
valideretvisuelanalogscore(VAS)brugt,somerbaseretpåetscorings-system.Derudoverblev
der brugt et omfattende spørgeskema, som vurderede forskellige følelser af mæthed.
Resultaterne fra undersøgelsen viste, at deltagerne oplevede signifikant højere
mæthedsfornemmelse, mindre trang til at spise deres yndlingsmad, og fandt det mindre
behagelig at spise endnu en mundfuld af den samme mad efter at have drukket grøn te
sammenlignetmedvand.Forskellige faktorerkanhave indflydelseog forklare resultaterne.
Smagsoplevelsenafgrøntekanhavehaftbetydningfordenøgedemæthedsfølelse.Deltagerne
føltehellerikkemeremisbehagvedindtagelseafgrønteendreferencemåltidet(vand),ogde
føltesighellerikkemeresygeigennemforsøgetmedgrønte.Derforkandenhøjerefølelseaf
mæthedikkeforklaresvedubehagafindtagelsenafgrønte.Desudenoplevededeltagerneen
stærkerefølelseaflysttilatindtagederesyndlingsmadellerendnuenmundfuldafdensamme
madefterreferencemåltidet.Ændringerihormonniveauerf.eks.GLP-1,kanværeårsagtilgrøn
te’s mæthedsfremmende effekt, men hormonniveauerne blev ikke målt i dette forsøg.
Undersøgelsen har alt i alt nogle begrænsninger, og resultaterne bør behandles med
forsigtighed. Forsøget var ikke blindet, og den øgede mæthedsfornemmelse kan være
forudindtaget. Et større antal deltagere, og derudover at inkludere overvægtige deltagere
kunnehaveindflydelsepåresultaterne.
CellemodellerIvoressøgenefter fødevarekomponenter, somkanstimuleresekretionafGLP-1,ogderved
ændre fødeindtag ved at give større mæthedsfornemmelse, bruges cellemodeller.
Enteroendokrine cellelinjer fra pattedyr bruges som udgangspunkt til at måle
tarmhormonresponspånæringsstofferogikke-næringsstoffer.Detervanskeligtatudviklein

19
vitromodeller af enteroendokrine cellelinjer, hovedsageligt fordi enteroendokrine celler er
sparsomt fordelt overmave-tarmkanalen, og lokaliseret sammenmed rigeligemængder af
enterocytter. Det er dog lykkedes for adskillige at succesfuldt danne in vitro screenings
bioassays (Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A.,
Requena, T., Swiatecka, D.,Wichers 2015). Blandt de enteroendokrine cellemodeller findes
STC-1ogHUTU-80,somderfokuserespåividereafsnit.
STC-1celler
TilatstudereGLP-1sekretionersekretiontumorcellelinjen(STC-1)megetbrugt.Denbesidder
mangeegenskabersomoprindeligeenteroendokrineceller,ogderforerdenmegetbrugttilat
identificereingredienser,derstimulerersekretionaftarmhormonerinvitro.Denerudviklet
fraproksimaletarmtumorerimus.STC-1cellelinjenerenheterogenenteroendokrincellelinje,
somudtrykkeradskilligemætheds-oginkretinhormoner,heriblandtCCK,GIP,PYY,pankreas
polypeptid, neurotensin og proglukagon-afledte peptider: GLP-1, GLP-2 og oxyntomodulin.
Egenskaben,atkunnefrigiveenstorvarietetaftarmhormoner,erogsåmedtilatgøreSTC-1
cellemodellenmegetbrugtogpopulær(Rindietal.1990).
Nårstofferneskaltestesfor,omdestimulererGLP-1sekretion,børfølgendeprocedureudføres.
Devalgtetestkomponenteropløsesiopvarmetbuffer.Bufferenskaltestesinden,foratsikreat
denpassersammenmeddetbestemteimmunoassay.TestopløsningenspHskaltilpassestil7.0-
7.4.Tilmåling af hormonsekretion, skal STC-1 cellernepodes i 6brøndsplader i suppleret
DMEM-medier(indeholderaminosyrerogvitaminer).Herstårcellerneietbestemtantaltimer
vedenbestemttemperatur,hvoreftermedierneaspireresogcellemonolagetvaskesmedden
valgtebuffer.Cellerneskaltilpassesdetnyeiklimaibuffereninogettid,oghereftertilsættes
testopløsningtilbrøndene.Cellerinkuberesi1-4timer,hvorerfaringerviser,atenperiodepå
3-4timereroptimaltforhormonsekretion.Derbøraltidinkluderespositivekontrollerkendt
for at stimulere tarmhormonsekretion og negative kontroller (buffer alene) i hver
forsøgsenhed.Efterinkubationtilsættesproteaseogphosphataseinhibitorforatbeskyttemod
hormonnedbrydning.Medietovercellerneopsamles,overførestilrørogcentrifugeresforat
fjernecellerester.Supernatanterneopsamlesogopbevares indenydereligeanalyse.Derbør
udføresmindst trebiologiskereplikater. Intracellulæreniveaueraf sekundæremessengere
(cAMP,IP3ogCa2+)brugesoftesomhurtigeindirekteindikatorerforhormonsekretion.Der

20
findesadskilligemetodertilmålingafdisse.TilpåvisningaftarmhormonerneCCK,PYYogGLP-
1findesenrækkefølsommeognøjagtigeimmunoassays(ELISAogRIA)(Verhoeckx,K.,Cotter,
P., López-Expósito, I., Kleiveland, C., Lea, T.,Mackie, A., Requena, T., Swiatecka, D.,Wichers
2015).
HuTu-80celler
HuTu-80erenhumancellelinje,somkommerfraen53årigkaukasiskmand.Cellernestammer
fraduodenum,ogudtrykkerreceptorerforbombesin(G-proteinkobledereceptorer).HuTu-
80cellerdyrkesisuppleretEMEMmedie(Eagle’sMinimumEssentialMedium)(CLScelllines
service,7/6-17).HuTu-80cellelinjener ikkeenoftebrugtcellelinje,hvilkenkanskyldes,at
erfaringerviser,atcellerneersværereathåndtereendforeksempelSTC-1celler.Dekanvære
sværeatdyrke,ogpludseligoplevesatværedøde(personligsamtale).
HuTu-80cellelinjener ikkenærsåbrugtsomSTC-1cellelinjen,ogderforfindesderikkeen
generel fremgangsmåde på, hvordan forskellige stoffer testes i denne cellemodel. HuTu-80
cellelinjenblevbrugtietforsøgafOhtsuetal.(2014).Sød-smagsreceptorerregulererGLP-1
sekretioniL-celler,hvorsignalsystemetaktiveretafdennereceptorblevundersøgtvha.HuTu-
80celler.Datestkomponenterneskulletestes,blevHuTu-80cellernepodetibrøndspladerog
inkuberetførmålingafGLP-1sekretion.Herefterblevcellernevaskettogangemedenbuffer,
og herefter inkuberet i bufferen. Efter blev de inkuberet i samme buffer, og derudover
forskelligekoncentrationeraftestkomponenterne.Supernatantenblevtilsidstiinkubationen
opsamletogcentrifugeretforatfjernecellerester.GLP-1koncentrationenisupernatantenblev
måltvedradioimmunoassay(RIA).Hverprøveblevanalyseretitoeksemplarer.
HuTu-80cellelinjenharderudovereksempelvisværebrugtietforsøg,hvorderblevundersøgt
omsteroidglycosidH.g.-12kunnestimulerehormonsekretion(CCK)fraenteroendokrineceller
(LeNevé et al. 2010). Cellelinjen har også været brugt i et forsøg, hvor de undersøgte om
liganderforTAS2R38receptor,kanstimulereGLP-1sekretion(Phametal.2016).

21
Diskussion
Grønteekstrakt
Omgrønteekstraktermæthedsfremmendeellerej,kanikkesvarespåendegyldigt. I første
undersøgelsevisteEGCGatstimulereGLP-1sekretionfraL-celler,hvilketblevpåvistpåCaco-
2cellerogimusetarme(Songetal.2015).IdetderervisusikkerhedvedCaco-2cellelinjen,bør
detogsåtestespåenandencellemodel.DerudoverblevEGCGtestetpåpatientermedtype2
diabetesoglipidabnormaliteter,hvorGLP-1sekretionblevøgetsignifikant(Liuetal.2014).I
densidsteundersøgelseblevdetmåltomEGCGvarmæthedsfremmende,ogdetblevtestetpå
raskemennesker.UndersøgelsenvisteatEGCGvarmedtilatøgemæthedsfornemmelsen(Josic
etal.2010).Iundersøgelsenvarderkun14deltagere,såienandenundersøgelsekrævesen
størreantaldeltagere, ogderudoverbørovervægtigedeltagere inkluderes.Pådenene side
tyder det på at grøn te ekstrakt kan øge GLP-1 sekretion og være potentielt
mæthedsfremmende,menpådenandensidekrævesderyderligerestudierogundersøgelser.
Anvendelseafcellemodeller
Cellemodeller bliver i høj grad anvendt i undersøgelsen af potentielt mæthedsfremmende
ingredienser, og spørgsmålet om, hvornår de kan bruges eller ikke bruges, er komplekst.
Forsøgsresultaterfracellemodellerskalanalyseresmedforsigtighed,idetatdetnetopforegår
på celleniveau. Resultaterne viser ikke resultater for en hel organisme. Resultater fra en
cellemodel,erresultatervistpåspecifiktdenenecelle,ogdettekangodtændresigogmåske
sletikkevirke,nårdetkommeroverpåenhelorganisme.Deteroftecellemodellerfradyrsom
bliveranvendt,menhumanecellemodellerbliverogsåihøjgradanvendt.Oftetestermanførst
på cellelinjer fra dyr, og derefter på humane cellelinjer. Dette er med til at gøre
forsøgsresultatermerebrugbare,idetdetnetopkommertætterepåenhelorganisme.Manskal
dogaltidvælgesincellelinjemedomhu,ogkiggepåhvorvalideretdener.
Udover cellemodeller bliver testkomponenterne også testet på dyr. Anvendelsen af
cellemodellerermedtilatsænkeanvendelsenafdyreforsøg,hvilketsessomenpositivting.
Grundentilatcellemodellerbegynderatkunneerstattedyreforsøg,skyldesblandtandetat
manharfåetbedremetoder,blevetbedretilatstandardisereforsøgene,ogfårmerevidenud
afdeenkelteforsøg.Pådenandensidenskalmanhuske,atcelleraltidkommerfraendonor,

22
hvilket har begrænsninger. Anvendelsen af dyr er med til at teste og fuldt ud forstå
behandlinger,somkanhaveindvirkningpåmenneskerogdyrssundhedpåenheltandenmåde,
endhvisderkunblevbrugt cellemodeller.Cellemodellervil altsåaldrigheltkunneerstatte
anvendelseafdyrtilforsøgafkropssystemer.
KonklusionOpgaven havde til formål at beskrive GLP-1’s rolle i mæthedsreguleringen, og derudover
potentieltmæthedsfremmende stoffer, som øger sekretionen af GLP-1, og til sidst hvordan
disse stoffer testes i cellemodeller. GLP-1 frigives fra L-celler i tarmen, og påvirker
appetitregulerendecentreihjernenviablodetogvagusnerven.Bådenæringsstofferogikke-
næringsstoffer er med til at stimulere sekretionen af GLP-1, og dermed virke potentielt
mæthedsfremmende. Grøn te ekstrakt, som fokuseres på i opgaven, viste sig at øge GLP-1
sekretionogøgemæthedsfornemmelsenienrækkeundersøgelser.Detkræverdogyderligere
undersøgelser af grøn te ekstrakt, for at kunne sige mere endegyldigt, om det er
mæthedsfremmende eller ej. Potentielt mæthedsfremmende stoffer testes i cellemodeller,
hvilketerenmegetbrugtmetode,sombådeharsinefordeleogulemper.
PerspektiveringFedmeogovervægteretstigendeproblemiverdenen,ogderarbejdesihøjgradpåatfinde
løsningertildette.Ændringerilivsstil,somatdyrkemeremotionogspisesundterenselvfølge,
menyderligeretiltagkanværenødvendige.Derfindesmegetmedicintilbehandlingaffedme,
mendetvilleværeenstorfordel,hvisnaturligesupplementerkunneanvendesibehandlingen
mod fedme, og på denne måde undgå anden form for medicinering. I opgaven er grøn te
ekstraktundersøgtfordenspotentieltmæthedsfremmendeeffekt.Grønteerbådeetsimpelt
ognaturligtsupplement,ogharpotentieltmæthedsfremmendeeffekter,hvordetkanværemed
tilatsnydekroppentilattroatdenermæt.Fremtidigtvilledetværeenstorfordel,hvisenpille
ellerkapselmedgrøn teekstraktkunnevirkesomfedmebehandling,ogmandervedkunne
undgåandenformformedicin.Udoveratkunnebrugestilbehandlingatfedme,kunnegrønte
ekstraktogsåværeenmuligkandidattilbehandlingaftype2diabetes.

23
ReferencerBaggio,L.L.&Drucker,D.J.,2007.BiologyofIncretins:GLP-1andGIP.Gastroenterology,
132(6),pp.2131–2157.
Balk-Møller,E.,Holst,J.J.&EhrenreichKuhre,R.,2014.Incretinsecretion:directmechanisms.
Baraboi,E.D.etal.,2010.Effectsofalbumin-conjugatedPYYonfoodintake:Therespective
rolesofthecircumventricularorgansandvagusnerve.EuropeanJournalof
Neuroscience,32(5),pp.826–839.
Buse,J.B.etal.,2004.Effectsofexenatide(exendin-4)onglycemiccontrolover30weeksin
sulfonylurea-treatedpatientswithtype2diabetes.DiabetesCare,27(11),pp.2628–2635.
Calvo,S.S.-C.&Egan,J.M.,2015.Theendocrinologyoftastereceptors.NatureReviews
Endocrinology,11(4),pp.213–227.Availableat:
http://www.nature.com/doifinder/10.1038/nrendo.2015.7.
Chaudhari,N.&Roper,S.D.,2010.Reviewseries:Thecellbiologyoftaste.TheJournalofCell
Biology,190(3),pp.285–296.Availableat:
http://www.jcb.org/cgi/doi/10.1083/jcb.201003144.
Chaudhri,O.B.,Field,B.C.T.&Bloom,S.R.,2008.Gastrointestinalsatietysignals.International
journalofobesity(2005),32Suppl7,pp.S28–S31.
CLScelllinesservice:http://www.clsgmbh.de(7/6-17)
DeFronzo,R.A.etal.,2005.Effectsofexenatide(exendin-4)onglycemiccontrolandweight
over30weeksinmetformin-treatedpatientswithtype2.DiabetesCare,28(5),pp.1092–
1100.
Degn,K.B.etal.,2004.OneWeek’sTreatmentWiththeLong-ActingGlucoseReleasein
PatientswithType2Diabetes.Diabetes,53(10),pp.1187–1194.
Diakogiannaki,E.etal.,2013.Oligopeptidesstimulateglucagon-likepeptide-1secretionin
micethroughproton-coupleduptakeandthecalcium-sensingreceptor.Diabetologia,
56(12),pp.2688–2696.
Drucker,D.J.&Nauck,M.A.,2006.Theincretinsystem:glucagon-likepeptide-1receptor
agonistsanddipeptidylpeptidase-4inhibitorsintype2diabetes.Lancet(London,
England),368(9548),pp.1696–1705.
Ferrannini,E.,1998.InsulinResistanceversusInsulinDeficiencyinNon-Insulin-Dependent
DiabetesMellitus:ProblemsandProspects.EndocrRev,19(4),pp.477–490.Availableat:

24
http://edrv.endojournals.org/cgi/content/abstract/19/4/477.
Hansen,L.etal.,2017.PeptidaseIVintheCapillariesSupplyingtheLCellsofthePorcine
Intestine*.,140(January),pp.5356–5363.
Hidalgo,I.J.,Raub,T.J.&Borchardt,R.T.,1989.Characterizationofthehumancoloncarcinoma
cellline(Caco-2)asamodelsystemforintestinalepithelialpermeability.
Gastroenterology,96(3),pp.736–49.Availableat:
http://www.ncbi.nlm.nih.gov/pubmed/2914637.
Holst,J.J.,2007.ThePhysiologyofGlucagon-likePeptide1.PhysiologicalReview,(225),
pp.1409–1439.
Josic,J.etal.,2010.Doesgreenteaaffectpostprandialglucose,insulinandsatietyinhealthy
subjects:arandomizedcontrolledtrial.NutritionJournal,9(1),p.63.Availableat:
http://nutritionj.biomedcentral.com/articles/10.1186/1475-2891-9-63.
Kao,Y.-H.,Hiipakka,R.A.&Liao,S.,2000.ModulationofEndocrineSystemsandFoodIntake
byGreenTeaEpigallactachinGallate.Endocrinology,141(3),pp.980–987.
Kendall,D.M.etal.,2005.Effectsofexenatide(exendin-4)onglycemiccontrolover30weeks
inpatientswithtype2diabetestreatedwithmetforminandasulfonylurea.Diabetes
Care,28(5),pp.1083–1091.
Kim,W.&Egan,J.M.,2008.Theroleofincretinsinglucosehomeostasisanddiabetes
treatment.Pharmacologicalreviews,60(4),pp.470–512.Availableat:
http://www.ncbi.nlm.nih.gov/pubmed/19074620%5Cnhttp://www.pubmedcentral.nih.
gov/articlerender.fcgi?artid=PMC2696340.
Lim,G.E.&Brubaker,P.L.,2006.Glucagon-LikePeptide1SecretionbytheL-Cell:TheView
FromWithin.Diabetes,55(Supplement2),pp.S70–S77.Availableat:
http://diabetes.diabetesjournals.org/cgi/doi/10.2337/db06-S020.
Liu,C.-Y.etal.,2014.EffectsofGreenTeaExtractonInsulinResistanceandGlucagon-Like
Peptide1inPatientswithType2DiabetesandLipidAbnormalities:ARandomized,
Double-Blinded,andPlacebo-ControlledTrial.PLoSONE,9(3),p.e91163.Availableat:
http://dx.plos.org/10.1371/journal.pone.0091163.
Lugari,R.etal.,2000.Effectofnutrientingestiononglucagon-likepeptide1(7-36amide)
secretioninhumantype1andtype2diabetes.Hormoneandmetabolicresearch=
Hormon-undStoffwechselforschung=Hormonesetmétabolisme,32,pp.424–8.
Availableat:http://www.ncbi.nlm.nih.gov/pubmed/11069208.

25
Lund,A.,K.Knob,F.&Vilsbøll,T.,2017.GLP-1-receptoragonistertilbehandlingaftype2-
diabetes–kliniskeforskelleogligheder.,4,p.4.
Mace,O.J.,Schindler,M.&Patel,S.,2012.TheregulationofK-andL-cellactivitybyGLUT2and
thecalcium-sensingreceptorCasRinratsmallintestine.TheJournalofphysiology,
590(Pt12),pp.2917–36.Availableat:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3448156&tool=pmcentrez
&rendertype=abstract.
Meyerhof,W.etal.,2010.TheMolecularReceptiveRangesofHumanTAS2RBitterTaste
Receptors.ChemicalSenses,35(2),pp.157–170.Availableat:
https://academic.oup.com/chemse/article-lookup/doi/10.1093/chemse/bjp092.
Murphy,K.G.&Bloom,S.R.,2006.Guthormonesandtheregulationofenergyhomeostasis.
Nature,444(7121),pp.854–859.Availableat:
http://www.ncbi.nlm.nih.gov/pubmed/17167473.
Narukawa,M.etal.,2011.Evaluationofthebitternessofgreenteacatechinsbyacell-based
assaywiththehumanbittertastereceptorhTAS2R39.BiochemicalandBiophysical
ResearchCommunications,405(4),pp.620–625.
Nauck,M.A.&Meier,J.J.,2016.Theincretineffectinhealthyindividualsandthosewithtype2
diabetes:physiology,pathophysiology,andresponsetotherapeuticinterventions.The
LancetDiabetes&Endocrinology,8587(15),pp.1–12.Availableat:
http://dx.doi.org/10.1016/S2213-8587(15)00482-9.
LeNevé,B.etal.,2010.ThesteroidglycosideH.g.-12fromHoodiagordoniiactivatesthe
humanbitterreceptorTAS2R14andinducesCCKreleasefromHuTu-80cells.American
journalofphysiology.Gastrointestinalandliverphysiology,299(6),pp.G1368-75.
Availableat:http://www.ncbi.nlm.nih.gov/pubmed/20930049.
Ohtsu,Y.etal.,2014.Diversesignalingsystemsactivatedbythesweettastereceptorin
humanGLP-1-secretingcells.MolecularandCellularEndocrinology,394(1–2),pp.70–79.
Availableat:http://dx.doi.org/10.1016/j.mce.2014.07.004.
Padidela,R.etal.,2009.Elevatedbasalandpost-feedglucagon-likepeptide1(GLP-1)
concentrationsintheneonatalperiod.EuropeanJournalofEndocrinology,160(1),
pp.53–58.
Pham,H.etal.,2016.Abitterpillfortype2diabetes?Theactivationofbittertastereceptor
TAS2R38canstimulateGLP-1releasefromenteroendocrineL-cells.Biochemicaland

26
BiophysicalResearchCommunications,475(3),pp.295–300.
Porte,D.,Baskin,D.G.&Schwartz,M.W.,2002.Leptinandinsulinactioninthecentralnervous
system.Nutritionreviews,60(10Pt2),pp.S20-9-84,85–7.Availableat:
http://www.ncbi.nlm.nih.gov/pubmed/12403080.
Rains,T.M.,Agarwal,S.&Maki,K.C.,2011.Antiobesityeffectsofgreenteacatechins:A
mechanisticreview.JournalofNutritionalBiochemistry,22(1),pp.1–7.Availableat:
http://dx.doi.org/10.1016/j.jnutbio.2010.06.006.
Rask,E.etal.,2001.Impairedincretinresponseafteramixedmealisassociatedwithinsulin
resistanceinnondiabeticmen.DiabetesCare,24(9),pp.1640–1645.
Rindi,G.etal.,1990.Developmentofneuroendocrinetumorsinthegastrointestinaltractof
transgenicmice.Heterogeneityofhormoneexpression.TheAmericanjournalof
pathology,136(6),pp.1349–63.Availableat:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1877573&tool=pmcentrez
&rendertype=abstract.
Roberge,J.N.&Brubaker,P.L.,1993.Regulationofintestinalproglucagon-derivedpeptide
secretionbyglucose-dependentinsulinotropicpeptideinanovelenteroendocrineloop.
Endocrinology,133(1),pp.233–240.
Rocca,A.S.&Brubaker,P.L.,1999.Roleofthevagusnerveinmediatingproximalnutrient-
inducedglucagon-likepeptide-1secretion.Endocrinology,140(4),pp.1687–1694.
RosenstockJ,ReuschJ,BushM,YangF,S.M.,2009.PotentialofAlbiglutide,aLong-ActingGLP-
1ReceptorAgonist,inType2Diabetes.DiabetesCare,32(10)(10),pp.1880–6.
Sam,A.H.etal.,2012.Theroleofthegut/brainaxisinmodulatingfoodintake.
Neuropharmacology,63(1),pp.46–56.Availableat:
http://www.sciencedirect.com/science/article/pii/S0028390811004631[AccessedMay
6,2017].
Song,W.-Y.etal.,2015.(-)-Epigallocatechin-3-gallateinducessecretionofanorexigenicgut
hormones.,57,pp.164–169.
Verhoeckx,K.,Cotter,P.,López-Expósito,I.,Kleiveland,C.,Lea,T.,Mackie,A.,Requena,T.,
Swiatecka,D.,Wichers,H.,2015.TheImpactofFoodBioactivesonHealthK.Verhoeckxet
al.,eds.,Cham:SpringerInternationalPublishing.Availableat:
http://www.scopus.com/inward/record.url?eid=2-s2.0-
84945157886&partnerID=tZOtx3y1.

27
Waget,A.etal.,2011.PhysiologicalandpharmacologicalmechanismsthroughwhichtheDPP-
4inhibitorsitagliptinregulatesglycemiainmice.Endocrinology,152(8),pp.3018–3029.
Whalley,N.M.etal.,2011.ProcessingofproglucagontoGLP-1inpancreaticalfa-cells:Isthisa
paracrinemechanismenablingGLP-1toactonbeta-cells?JournalofEndocrinology,
211(1),pp.99–106.
Yamazaki,T.etal.,2013.ActivationofthehTAS2R14humanbitter-tastereceptorby(-)-
epigallocatechingallateand(-)-epicatechingallate.Bioscience,Biotechnology,and
Biochemistry,77(9),pp.1981–1983.Availableat:
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Cit
ation&list_uids=24018685.
Related Documents










![IN2040: Funksjonell Programmering [3ex] Strømmer og utsatt ... · IN2040: Funksjonell Programmering Strømmer og utsatt evaluering StephanOepen Universitetet i Oslo 23. oktober2019](https://static.cupdf.com/doc/110x72/5e5212a67f5ada68f968a78e/in2040-funksjonell-programmering-3ex-strmmer-og-utsatt-in2040-funksjonell.jpg)
