Rend. Fz}. Acc. Lincei s. 9, v. 8:t39-149 (1997) Patologia. -- Glial Ceil Line-Derzved Neurotrophic Factor (GDNF) stimulates Ret activity. Nota di FRANCESCA CARLOMAGNO, ROSA MARINA MELILLO, GIULIANA SALVATORE, ROBERTA VISCONTI~ GABRIELLA DE VITA, GELSY LUPOLI, MICHELE MARINACCIO, ALFREDO FUSCO, MASSIMO SANTORO e GIANCARLO VECCHIO, presentata(*) dal Corrisp. G. Vecchio. Aus'Iv,aCT. -- Ret is a receptor-like tyrosine kinase involved in several neoplastic and developmental dis- eases affecting thyroid gland and tissues of neuroectodermal origin. Rearrangements of the ret gene are ob- served in a sizeable fraction of human thyroid papillary carcinomas. <<Gain of function~) of ret has been shown to be caused by mutations associated with multiple endocrine neoplasia type 2A (MEN2A) or 2B (MEN2B) syndromes. Conversely, congenital megacolon or Hirschsprung's disease (HSCR) is associated with <<loss of function)~ of ret. Here, we show that glial cell line-derived neurotrophic factor (GDNF), a member of the TGF]3 super-family, is able to stimulate in vivo tyrosine kinase activity of Ret. These effects were specific to GDNF, since neither other members of the TGF[5 family nor other neurotrophins and un- related growth factors exerted the same effects. Consistently, ectopic expression of Pet in NIH 3T3 cells rendered them responsive to the mitogenic action of GDNF. These results allow the conclusion that GDNF is a functional ligand for Ret and suggest that Ret-GDNF interaction plays an important role in the pathogenesis of diseases associated with ret mutations. Kt~v WORDS: Ligand; Transformation; Tyrosine kinase; Neurotrophin. Rb\.SSUNTO. - - I1 Fattore Neurotrofico derivato dalla Gha (GDNF) stt)nola l'atts di Ret. Ret ~un recet- tore con attivit.a tirosino chinasica coinvolto in alcune malattie neoplastiche e dello sviluppo riguardanti [a ghiandola tiroidea ed i tessuti di origine neuroectodermica. Riarrangiamenti del gene ret sono stati ossem, ati in un consistente numero di carcinomi papi]liferi tiroidei umani. Inoltre, l'attivazione del potenziale trasfor- mante diret 6 causata da mutazioni associate alle sindromi da neoplasia endocrina multipla tipo ~\ (MEN2A) o tipo 2B (MEN2B). A1 contrario, il megacolon congenito o malattia di Hirschsprung (HSCR) associato a mutazioni causanti una perdita di funzione di ret. In questo lavoro, dimostriamo che il fattore neurotrofico derivato da cellule gliali (GDNF), membro della super famiglia del TGFIB, ~ in grado di stirno- lare in vivo l'attivita tirosino chinasica di Ret. Questi effetti sono specifici per il GDNF, dal momento che nessuno degli altri componenti della famiglia del TGF]3 o nessuna altra neurotrofina o fattore di crescita appartenente ad altre famiglie esercita lo stesso effetto. Questi risultati ci permettono di concludere che il GDNF 8 il ligando funzionate di Ret e di suggerire che l'interazione Ret-GDNF gioca un ruolo molto ina- portante nella patogenesi delle malattie associate con le mutazioni di ret. LIST OF ABBI(EVIATIONS USED: NGF: nerve growth factor; GDNF: glial cell line derived neurotrophic fac- tor; BDNF: brain derived neurotrophic factor; FGF: fibroblast growth factor; CNTF: ciliary, neurotrophic factor; EGF: epidermal growth factor; PDGF: platelet derived growth factor; NT3: neurotrophic factor 3; NT4: neurotrophic factor 4; TGF c~ and [3: transforming growth factors 0~ and ~; HSCR: Hirschsprung's disease. INTRODUCTION The ret gene encodes a tyrosine-kinase receptor, Ret, (Takahashi et al., 1988) with potential transforming activity, as shown by its oncogenic activation, by somatic rear- (*) Nella seduta del 18 aprile 1997.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Rend. Fz}. Acc. Lincei s. 9, v. 8:t39-149 (1997)
P a t o l o g i a . - - G l i a l C e i l L i n e - D e r z v e d N e u r o t r o p h i c Factor ( G D N F ) s t i m u l a t e s R e t
activity. N o t a di FRANCESCA CARLOMAGNO, ROSA MARINA MELILLO, GIULIANA SALVATORE,
ROBERTA VISCONTI~ GABRIELLA D E VITA, GELSY LUPOLI, MICHELE MARINACCIO, ALFREDO
FUSCO, MASSIMO SANTORO e GIANCARLO VECCHIO, presentata(*) dal Corrisp. G .
V e c c h i o .
Aus'Iv, aCT. - - Ret is a receptor-like tyrosine kinase involved in several neoplastic and developmental dis- eases affecting thyroid gland and tissues of neuroectodermal origin. Rearrangements of the ret gene are ob- served in a sizeable fraction of human thyroid papillary carcinomas. <<Gain of function~) of ret has been shown to be caused by mutations associated with multiple endocrine neoplasia type 2A (MEN2A) or 2B (MEN2B) syndromes. Conversely, congenital megacolon or Hirschsprung's disease (HSCR) is associated with <<loss of function)~ of ret. Here, we show that glial cell line-derived neurotrophic factor (GDNF), a member of the TGF]3 super-family, is able to stimulate in vivo tyrosine kinase activity of Ret. These effects were specific to GDNF, since neither other members of the TGF[5 family nor other neurotrophins and un- related growth factors exerted the same effects. Consistently, ectopic expression of Pet in NIH 3T3 cells rendered them responsive to the mitogenic action of GDNF. These results allow the conclusion that GDNF is a functional ligand for Ret and suggest that Ret-GDNF interaction plays an important role in the pathogenesis of diseases associated with ret mutations.
Kt~v WORDS: Ligand; Transformation; Tyrosine kinase; Neurotrophin.
Rb\.SSUNTO. - - I1 Fattore Neurotrofico derivato dalla Gha (GDNF) stt)nola l'atts di Ret. Ret ~ un recet- tore con attivit.a tirosino chinasica coinvolto in alcune malattie neoplastiche e dello sviluppo riguardanti [a ghiandola tiroidea ed i tessuti di origine neuroectodermica. Riarrangiamenti del gene ret sono stati ossem, ati in un consistente numero di carcinomi papi]liferi tiroidei umani. Inoltre, l'attivazione del potenziale trasfor- mante d i r e t 6 causata da mutazioni associate alle sindromi da neoplasia endocrina multipla tipo ~ \ (MEN2A) o tipo 2B (MEN2B). A1 contrario, il megacolon congenito o malattia di Hirschsprung (HSCR) associato a mutazioni causanti una perdita di funzione di ret. In questo lavoro, dimostriamo che il fattore neurotrofico derivato da cellule gliali (GDNF), membro della super famiglia del TGFIB, ~ in grado di stirno- lare in vivo l'attivita tirosino chinasica di Ret. Questi effetti sono specifici per il GDNF, dal momento che nessuno degli altri componenti della famiglia del TGF]3 o nessuna altra neurotrofina o fattore di crescita appartenente ad altre famiglie esercita lo stesso effetto. Questi risultati ci permettono di concludere che il GDNF 8 il ligando funzionate di Ret e di suggerire che l'interazione Ret-GDNF gioca un ruolo molto ina- portante nella patogenesi delle malattie associate con le mutazioni di ret.
LIST OF ABBI(EVIATIONS USED: NGF: nerve growth factor; GDNF: glial cell line derived neurotrophic fac- tor; BDNF: brain derived neurotrophic factor; FGF: fibroblast growth factor; CNTF: ciliary, neurotrophic factor; EGF: epidermal growth factor; PDGF: platelet derived growth factor; NT3: neurotrophic factor 3; NT4: neurotrophic factor 4; TGF c~ and [3: transforming growth factors 0~ and ~; HSCR: Hirschsprung's disease.
INTRODUCTION
T h e ret g e n e e n c o d e s a t y r o s i n e - k i n a s e r e c e p t o r , Re t , ( T a k a h a s h i et al . , 1 9 8 8 ) w i t h
p o t e n t i a l t r a n s f o r m i n g act iv i ty , as s h o w n b y its o n c o g e n i c a c t i v a t i o n , b y s o m a t i c r ea r -
(*) Nella seduta del 18 aprile 1997.

140 F. CARLOMAGNO ET AL.
rangements, in papillary thyroid carcinomas (Grieco et al., 1990). Several observations suggest that ret may serve a role in the migration, committment, survival and differenti- ation of cell lineages of the nervous system. Re t expression is found in migrating neural crest ceils and in the developing peripheral and central nervous systems in several species (Pachnis et al., 1993; Schuchardt et al., 1994). Consistently, mutations of ret are associated with several neurochristopathies (diseases affecting tissues derived from neural crest), including multiple endocrine neoplasia type 2A and 2B (MEN2A and MEN2B), familial medullary thyroid carcinoma (FMTC), and Hirschsprung's disease. Substitutions of extracellular cysteine residues or of methionine 918 of the cytoplasmic domain of ret are associated to MEN2A/FMTC (Mulligan et al., 1993; Donis-Keller et al., 1993) or to MEN2B (Carlson et al., 1994; Hofstra et al., 1994) syndromes, respect- ively. Hirschsprung's (HSCR) disease is regarded as the consequence of premature ar- rest of the craniocaudal migration of neural crest-derived enteric neurons toward the anal end of the rectum, between the 5th and the 12th week of gestation. This causes the absence of autonomic ganglion cells within intestinal parasympathetic Meissner's and Auerbach's plexuses and, as a consequence, a functional obstruction resulting in mega- colon. Heterogeneous ret mutations characterize some cases of HSCR (Romeo et al.,
1994; Edery et al., 1994). Consistently, targeted disruption o f f e r causes a lack of enter- ic ganglion cells in homozygous mice (Schuchardt et ell., 1994). Ret involvement in neu- ral differentiation is also indicated by the observation that activated ret isoforms are able to induce differentiation of rat pheochromocytoma (Califano et al., 1996) and hu- man neuroblastoma cells (D'Alessio et al., 1995). On the basis of these considerations Ret was predicted to function as the receptor for a factor endowed with neurotrophic activity.
Here we show that a recently discovered neurotrophic factor, glial cell line derived neurotrophic factor (GDNF) (Linet al., 1993) acts as a functional ligand for Ret. In- deed, it stimulates Ret enzymatic function and elicits mitogenic and/or differentiative effects in Ret expressing cells.
M A T E R I A L S ANI ) MI:Tr l IODS
Cells.
NIH 3T3 cells were grown in Dulbecco's modified Eagle's medium (DMEM) sup- plemented with 10% foetal calf serum. NIH 3T3 cells transfected with an LTR- based ret expression vector have been described helsewere (Santoro et al., 1995). NIH 3T3 cells expressing the epidermal growth factor receptor /re t chimera (Santoro et al., 1994), mutated ret MEN2A, ret MEN2B, (Santoro et al., 1995), ret HSCR972 and ret HSCR32 (Carlomagno et al., 1996) have also been described. PC12 cells were grown in RPMI 1640 (Gibco, BRL) supplemented with 10% horse serum and 5% foetal calf serum. SK-N-BE cells were grown in RPMI 1640 (Gibco, BRL) supplemented with 10% foetal calf serum. [3H]thymidine incorporation assays were performed as de- scribed previously (Santoro et al., 1994). Briefly, NIH 3T3 transfectants, grown to con-

GLLAL CELL LINE.DERIVED NEUROTROPHIC FACTOR (GDNF) STLMUIaX.TES RET ACTIVH'Y 141
fluence in 24-well plates (Costar), were serum-starved for 24 h and then treated with GDNF for 24 h in the presence of 4 [tCi of [3H]thymidine per ml. Data are expressed as a mitogenic index, calculated as the fraction of stimulation obtained in the presence of GDNF with respect to the stimulation obtained with 1% calf serum.
Pet phosphorylation assays.
NGF, GDNF, NT-3, NT-4, BDNF and CNTF were purchased from Alomone labs., Israel. EGF, PDGF, TGFfSs and FGFs were from Upstate Biothechnology (Lake Placid, NY). Immunoprecipitation and immunoblotting experiments were performed as described by Santoro et al. (1994). Briefly, cells were lysed in a buffer containing 50 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES; pH 7.5), 1% (vol/vol) Triton X-100, 50 mM NaC1, 5 mM ethylene glycol-bis([5-amino-ethyl ether) N, N, N', N' tetraacetic acid (EGTA), 50 mM NaF, 20 mM sodium pyrophosphate, 1 mM sodium vanadate, 2 roam phenylmethylsulphonyl fluoride, 0.2 ~tg each of apro- tinin and leupeptin per ml. Lysates were clarified by centrifugation at 10,000 • g for 15 min and the supernatant processed for immunoblotting or for immunoprecipitation. Protein concentration was estimated by the Bio-Rad kit (Bio Rad Laboratories, Her- cules, Ca). To analyse tyrosine-phosphorylation of Ret products, comparable amounts of Ret were immunoprecipitated and assayed for phosphotyrosine (pTyr) content. An- ti-Ret is a polyclonal antibody directed against the tyrosine-kinase domain of Ret (San- toro et al., 1994) and the anti-phosphotyrosine was a monoclonal antibody (4G10) pur- chased from Upstate Biotechnology, Lake Placid, NY. Immunoblots were stained with appropriate secondau antibodies and revealed with the Amersham ECL system (Amersham Life Sciences, Buckinghamshire, UK).
R E S U L T S
GDNF stimulates Ret tyrosine phoaTahorylation.
Based on the physiological and pathological role of Ret in the nervous system, we tested the ability of several neurotrophins to stimulate Ret tyrosine phosphorylation in vivo. As recipients we employed NIH 3T3 cells, which are naive for Ret expression, re- combinantly engineered to express Ret (NIH-ret) (Santoro et aI., 1995), and two cell lines which naturally express ret: PC12 rat pheochromocytoma, and SK-N-BE human neuroblastoma cell lines. We initially analysed the effects of the glial cell line-derived neurotrophic factor (GDNF), in comparison to growth factors of the NGF family (NGF, BDNF, NT-3 and NT-4) and to CNTF, another ligand with known neurotroph- ic properties. Only GDNF was able to induce detectable tyrosine phosphorylation of Pet in all three cell lines tested (fig. 1A)I Several other unrelated growth factors includ- ing basic FGF, acid FGF, EGF, and PDGF did not show any effect on Ret (fig. 1B and data not shown). GDNF belongs to the TGF[3 superfamily (Lin et al., 1993); thus we tested whether other members of this family could affect Ret phosphorylated status. Three members of the TGFf3 superfamily (TGF]31, TGF[32, TGF(33) were not capable
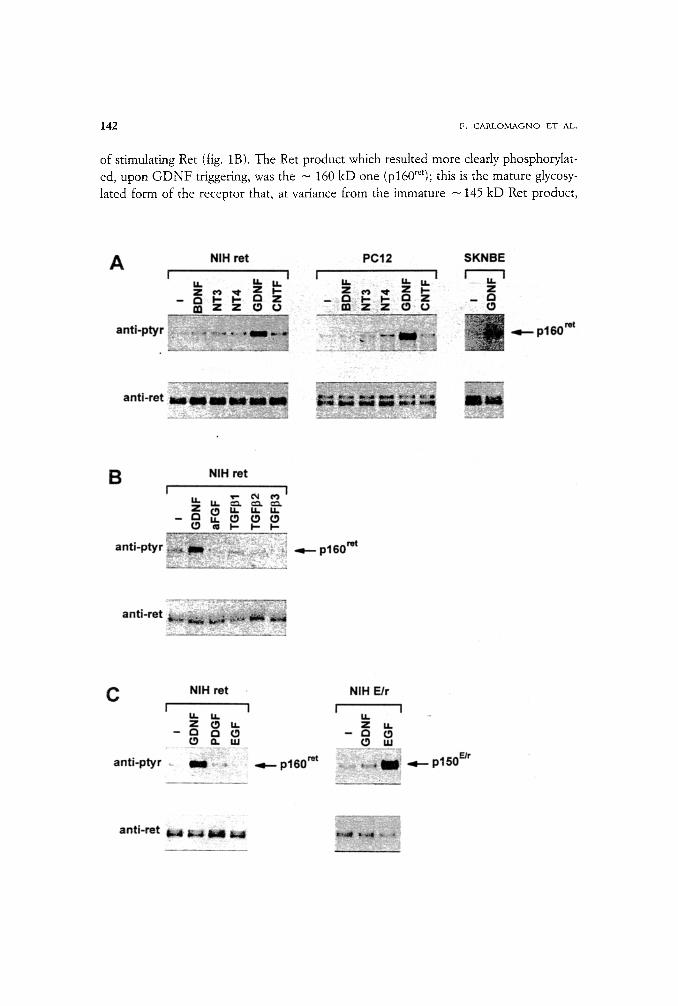
142 F. C A R L O M A G N O lET AL.
of stimulating Ret (fig. 1B). The Ret product which resulted more clearly phosphorylat- ed, upon G D N F triggering, was the - 160 kD one (p160~et); this is the mature glycosy- lated form of the receptor that, at variance from the immature - 145 kD Ret product,

GLb\L CELL LINE-DERIVED NEUROTROPHIC FACTOR /GDNF) STIMULATES RET ACTI'vTIW 143
is present on the cell surface (Asai et al., 1995). An additional evidence of the specificity of the observed effect derived from the observation that Ret tyrosine phosphorylation, following G D N F stimulation, was dependent on the presence of the extracellular do- main of Ret. Accordingly, the phosphorylation of an EGFR-Ret chimera, in which Ret
extracellular domain was replaced with that of the EGFR (Santoro et al., 1994), was in- duced by E G F but not by G D N F (fig. 1C).
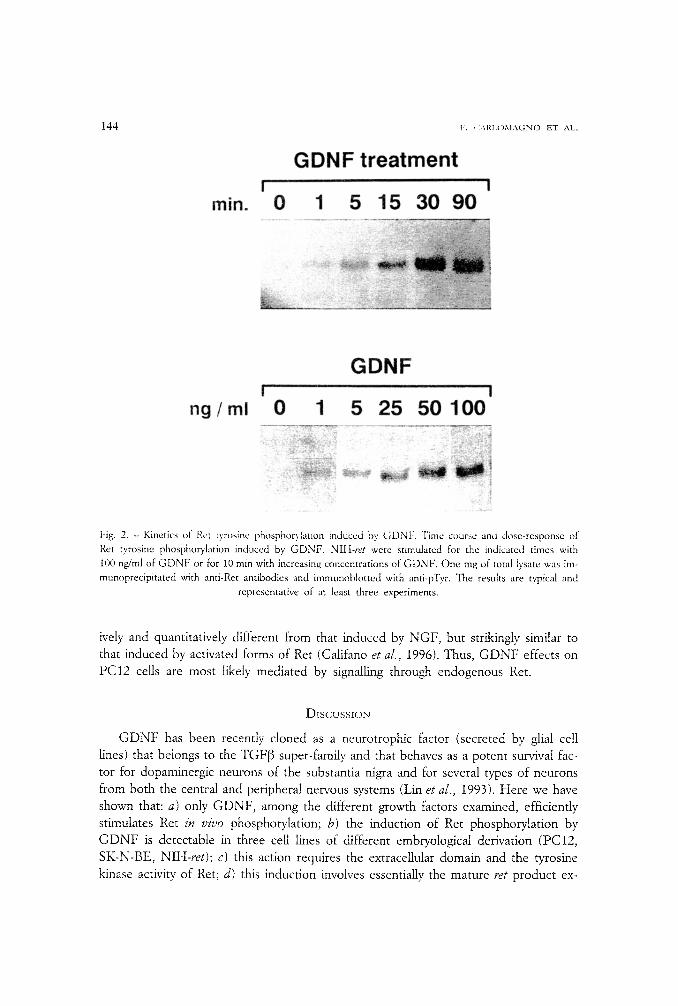
Time-course and dose*response experiments were performed to evaluate the kinet- ics of Ret phosphorylation induced by G D N F . In NIH-ret cells G D N F was able to in- duce phosphorylation of Ret rapidly, as quickly as 1 min after its addition, and at con-
centrations as low as 1 ng/ml (fig. 2). Maximal responses were obtained at 30 rain of stimulation and with a G D N F concentration around 50 ng/ml. Moreover, the stimula- tion of Ret activity by G D N F was confirmed when in vitro kinase assays were per- formed (data not shown).
Biological effects o f Ret-tn'ggerz)~g by G D N F .
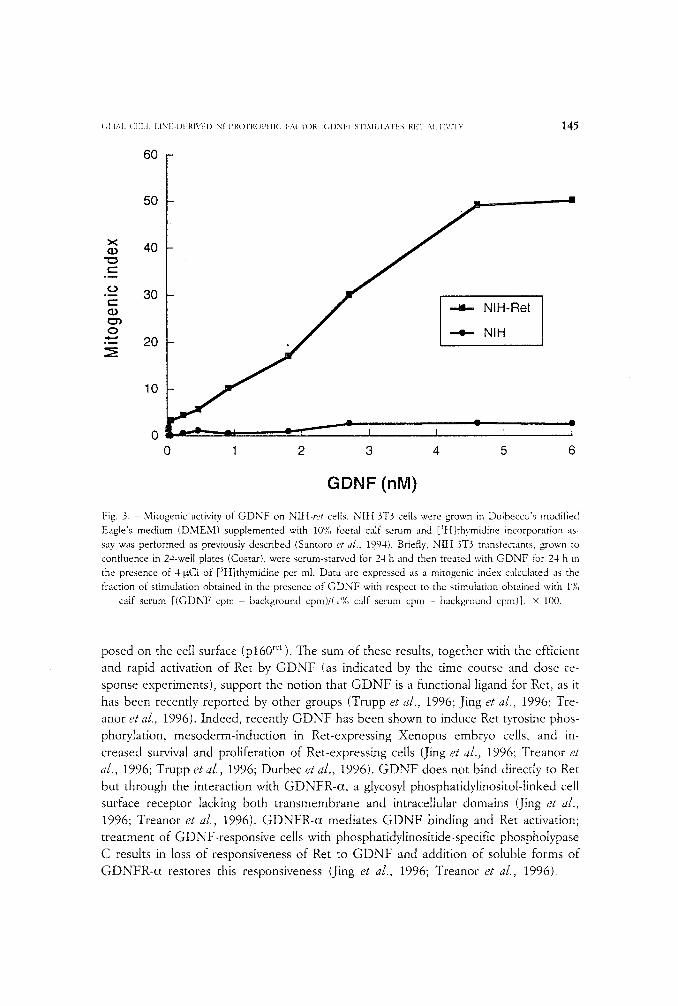
Induction of Ret phosphorylation upon G D N F triggering corresponded to an acti- vation of its biological activity. Ret activation should result into proliferation of NIt-t-ret cells, as it has been reported for various forms of activated ret (Grieco et al., 1990; San- toro et al., 1994, 1995; Asai et al., 1995). Indeed, when we evaluated, by a thymidine in- corporation assay, whether Ret stimulation by G D N F was endowed with mitogenic ef- fects, we found that G D N F was able to induce DNA synthesis, in a dose-dependent way, in NIH-ret , but not in parental N I H 3T3 cells (fig. 3).
The availability of a molecule able to activate Ret, i.e. G D N F , allowed us to study the effects of wild type Ret in a nearly <<physiological setting~. We decided to use PC12 cells which have been widely employed as a model of neuronal differentiation (Greene and Tishler, 1976). Nerve growth factor (NGF) treatment of PC12 induces differentia- tion to a sympathetic neuronal phenotype, which is characterized by long neurite exten- sion (fig. 4). We have reported, recently, that various activated forms of ret induced a pattern of differentiation, characterized by induction of immediate and delayed genes, and a peculiar flat morphology, which is different from that induced by N G F (Califano et al., 1996). GDNF-t rea ted PC12 cells acquired a flat morphology, with short neuritic processes (fig. 4). O f note, the GDNF- induced pattern of differentiation was qualitat-
4 Fig. t. - Tyrosine phosphorylation of Ret induced by GDNF: gowth factor specificity of Ret activation. NIH 3T3 transfected with LTR-ret (NLH-ret), PC12, SK-N-BE, or NIH 3T3 transfected with an EGFR-ret chimeric receptor (NIH-E/r) ceils were treated for 10 min at 37 ~ with 100 ng/ml of the indicated growth factors. Two mg (NIH-ret and NIH-E/r) or four mg (PC12 and SK-N-BE) of protein lysates were immu- noprecipitated with a potyclonat antibody to Ret; one half of the immunoprecipitate was immunobtotted with anti-Ret (anti-Ret) and one half with anti-phosphotyrosine (anti-pTyr). Pet proteins are detected as
145 kD and - 160 kD proteins; the - 160 kD species represents the mature glycosylated protein pre- sent on the cell surface, whereas the - 145 kD form is an immature precursor; the molecular weight diffe- rence between the two forms is even more evident in PCt2 cells, presumably as a result of differences in glycosylation in different cell lines. The molecular weight of the ~ 160-kD protein (p160 ~' ) is indicated.
144 F. ~7\RI,~3MAGNO ET AL,
Fig. 2. - Kinetics of Ret t.vrosinc phosphorylation induced by GDNF. Time course and dose-response of Ret tyrosine phosphorylation induced by GDNF. NItt-ret were stimulated for the indicated times with 100 ng/ml of GDNF or for 10 min with increasing concentrations of GDNF. One mg of total lysate was im-
munoprecipitated with anti-Ret antibodies and immunoblotted with anti-pTyr. The results are typical and representative of at least three experiments.
ively and quantitatively different from that induced by NGF, but strikingly similar to that induced by activated forms of Ret (Califano et al., 1996). Thus, GDNF effects on PC12 cells are most likely mediated by signalling through endogenous Ret.
D I s c u s s I O N
GDNF has been recently cloned as a neurotrophic factor (secreted by gliat cell Lines) that belongs to the TGF[3 super-family and that behaves as a potent survival fac- tor for dopaminergic neurons of the substantia nigra and for several types of neurons from both the central and peripheral nervous systems (Lin et al., 1993). Here we have shown that: a) only GDNF, among the different growth factors examined, efficiently stimulates Ret in vivo phosphorylation; b) the induction of Ret phosphorylation by GDNF is detectable in three cell lines of different embryological derivation (PC12, SK-N-BE, NIH-ret ) ; c) this action requires the extracellular domain and the tyrosine kinase activity of Ret; d) this induction involves essentially the mature ret product ex-
( , I [~,l. (~EI.I . I . [NI~-I31"~I{I\q 'D N E I ! R O T R t ) P I [ I C F A C T O R I ( ; I ) N I : I S T I M U I , A T E S l{E'r A(Trr~,rri~ '
6O
145
5O
4 0
-~ 30
g, ~ 20
10
0 0 - - I
0 6
H-Ret
A
- I ~ r I I
1 2 3 4 5
GDNF (nM)
Fig. 3. - Mitogenic activity of GDNF on NIII-ret cells. NIt{ 3T3 cells were grown in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10'}(, foetal calf serum and [)H]thymidine incorporation as-
sav was performed as previously described (Santoro et al., 19.94). Briefly, NIH 3T3 transfectants, grown to
confluence in 24-well plates (Costar), were serum-starved for 24 h and then treated with GDNF for 24 h in
the presence of 4 ~tCi of [~t{]thymidine per ml. Data are expressed as a mitogenic index calculated as the
fraction of stimulation obtained in the presence of GDNF with respect to the stimulation obtained with 1%
calf serum [(GDNF cpm - background cpm)/(l"/o calf serum cpm - background cpm)]. • 100.
posed on the cell surface (p160 r~t ). The sum of these results, together with the efficient and rapid activation of Ret by GDNF (as indicated by the time course and dose re- sponse experiments), support the notion that GDNF is a functional ligand for Ret , as it has been recently reported by other groups (Trupp et al., 1996; Jing et aJ., 1996; Tre- anor et al., 1996). Indeed, recently GDNF has been shown to induce Ret tyrosine phos- phorylation, mesoderm-induction in Ret-expressing Xenopus embuo cells, and in- creased survival and proliferation of Ret-expressing cells (Jing et al., 1996; Treanor et aL, 1996; Trupp et aJ., 1996; Durbec et al., 1996). GDNF does not bind directly to Ret but through the interaction with GDNFR-c*, a glycosyl phosphatidylinositol-tinked cell surface receptor lacking both transmembrane and intracellular domains (Jing et al., 1996; Treanor et al., 1996). GDNFR-c* mediates GDNF binding and Ret activation; treatment of GDNF-responsive cells with phosphatidylinositide-specific phospholypase C results in loss of responsiveness of Ret to GDNF and addition of soluble forms of GDNFR-ct restores this responsiveness (Jing et al., 1996; Treanor et a/., 1996).

146 F, C A R L O M A G N O E T AI..
Fig. 4. - Morphological effects of GDNF on PCI2 cells. PC12 were grown for 7 days in the presence or in the absence of 100 ng/ml of NGF or GDNF as recommended by Greene and Tishler (t976). Representati-
ve microphotographs are shown.
The results presented here indicate that in NIH 3T3, PC12 and SK N BE cells, in the absence of exogenously transfected GDNFR-ct, GDNF is able to induce readily detectable kinase activity of Ret and that, in NIH-ret cells, this stimulation results in mitogenesis and in PC12 in a neuronal phenotype. A plausible explanation for the dis- crepancy between the results obtained by us and those mentioned above is that low lev- els of GDNFR-(~ are endogenously expressed in the cell lines tested in the present work. Indeed, an RT-PCR analysis demonstrated the presence of low levels of GDNFR-ct transcripts in our NIH 3T3 cells (R. M. Melitlo and M. Santoro, unpub- lished results).
The demonstration that GDNF is an important component in the Ret stimulation machinery has been recently confirmed by the observation that, in analogy with target- ed disruption of ret (Schuchardt et aL, 1994), mice carrying targeted null mutations in the GDNF gene were reported to exhibit the absence of kidneys and the complete loss of enteric neurons in the digestive tract (Sanchez et al., 1996).
However, the possibility, that additional mechanisms of induction of Ret activity may exist have also to be considered. For instance, our preliminary results indicate that retinoic acid treatment of SK-N-BE cells causes overexpression and phosphorylation of Ret to a stoichiometry which is even higher than that induced by GDNF (F. Carlomag- no, V. de Franciscis and M. Santoro, unpublished results).
In conclusion, these findings have important implications for the role of the GDNF-Ret interaction in physiological and pathological conditions. In some settings, paracrine and/or autocrine interactions could be envisaged. For example, Ret products expressed in motoneurons (Pachnis et al., 1993) could be stimulated by GDNF elabo- rated by glial or muscle cells (Linet al., 1993). To this regard, it is worth noting that the upregulation of GDNF mRNA in the transected rat sciatic nerve corresponds to the

GLDkL CELL LINE-DERIVED NEUROTROPHIC FACTOR (GDNF) SThMULATES RET ACTIVITY 147
upregulation of Ret upon transection of the rat hypoglossal nerve (Colucci-D'Amato et al., 1996). These observations suggest that GDNF-Ret interaction may be involved in nerve repair. Similarly, Ret is expressed in cerebellar Purkinje cells and in the nigro- striatal region (Trupp et a[., 1996; Colucci-D'Amato et al., 1996). The neurotrophic ac- tion of G D N F on Purkinje cells and dopaminergic neurons of the substantia nigra (Lin et al., 1993) suggests a role for GDNF-Ret interaction in the growth and/or differentia- tion of these structures. Finally, Ret-GDNF interaction is also likely to have functions in districts outside the nervous system. Ret is expressed in thyroid (C-cells), adrenal medulla, kidney, salivary glands as well as testis (Pachnis et al., 1993; G. Viglietto and M. Santoro, unpublished results), and G D N F has been reported to be expressed in the same tissues (Sanchez et al., 1996).
Finally, the isolation of a ligand for Ret will allow a further definition of the mecha- nisms by which activated ret alleles, associated with familial tumoral syndromes, deter- mine neoplastic transformation and thus, it will help clarifying the pathogenesis of these human disorders.
ACI,.D, IOWLEDGEMENTS
We are grateful to Prof. G. Salvatore for his continuous encouragement and enthousiasm in supporting this work. We also thank P. P. Di Fiore for his suggestions, A. M. Cirafici and M. Pascotto for their contri- bution to this work and V. de Franciscis for the PC12 cells. This study was supported by the Associazione Italiana per la Ricerca sul Cancro (AIRC) and by the Progetto Finalizzato ACRO, Sottoprogetto 2, Biologia Molecolare. Part of the work of G. Vecchio was accomplished while he was a Fogarty Scholar-inRcsidcnce of the National Institutes of Health, Md., USA.
REFEILENCES
As,u N., IWAS~ u'rA T., NL\'rsuv,~'~a M., T,XKAI IASi u M, 1995. Mechani~vn ofactivatz?m of the retproto-onco~qene by. multiple endocrine neoplasia 2A mutanons. Mol. Cell. Biol., 15: 1613-1619.
C..vLnV,,:,~{,'. D., D'AlmssI{.) A., CoLu(:c1.D'A~t,wo G. L., DE Vm\ G., MoN,v.:o C., SANTELLI G., Dl Floe, l.: P. P., VECCI IIO G., Fus(o A., SANTORO M., DE FI~\NCISCIS V., 1996. MEN2 ret mutants dz~sociatc cell differentia- tion ~om inhibinon of prohferation and abrogate nerve growth factor responsiveness in the PC I2 cells. Proc. Natl. Acad. Sci USA, 93: 7933-7937.
C,U~LO,*L'\(~NO F., DE VITA G., BERLINGIERI M.T., I)E FRANCISClS V., MELILLO R. M., COGXNTUONI V., KI~kOS M. H., DI FI{oP, E P. P., Fusco A., SANTOIr IX/I,, 1996. Molecular heterogenei{'y of RET loss offimction in Hz~sch~rung's dz}ease. EMBO J., 15: 2717-2725.
CARLSON K. M., Dou S., C~ ~l D., SCaVARDa N., Tosl u,',IA K., JacKsoN C. E., WFLLS S. A., GOODFELLOW P., Do- NIs-KELLEr~ H., 1994. Single missense mutation in the tyrosine-kinase catalytk" domain of the PET proto- oncogene is associated wtth multiple endocrine neoplasia type 2B. Proc. Natl. Acad. Sci. USA, 91: 1579-1583.
COLUCCI-D'A,~,L-\TO G. L., D'ALEsslo A., FILLDXTREAU G., Fnomo T., DI GI?~'4BEI<ARDINO L., CIllAPPETFA G., VECCHIO G.~ F u s c o A., SANTORO ~X/L 7 DE FIL.\NCISCIS V., 1996. Presence ofphyszblogically stimulated RET in adult rat brain: induction of ret expression during nerve regeneration. Cell Growth Diff., 7: 1081-1086.
D'ALEssIo A., DE VITA G., C,vLi G., NITSCH L., Fusco A., V>:ccruo G., SANTELLI G., SANTORO M., DE FRAN- cmcIs V., 1995. Expression of the RET oncogene induces the SK-N-BE neuroblastoma cells to differen- tiate. Cell Growth Diff., 6: 1387-1394.

148 F. CARLONIAGNO ET A_L.
DONIS-KELLER H., Dou S., Clu D., CARLSON K. M., TOSIIIMA K., LA~MORE T. C., HOWE J. R., MOLEY J. F., GOODFELL()W P., WELLS S. A., 1993. Mutations in the RET" proto-oncogene are assoczated with MEN2A and FMTC. Hum. M01. Genet., 2: 851-856.
DuPamc P., hL~cos-Gu~EtU~EZ C. V., Km~NNY C., GvaGomu M., WAR'nOW~VXRA K., SUVANTO P., SMrrH D., PONDER B., Cos'rANTINt F., SA~'a~tA M., SamoLa H., PACUNm V., 1996. GDNFszgnallbzg through the Ret receptor tyrosine kinase. Nature, 381: 789-793.
EDERY P., LYONNET S., MULL*CAN L. M., PELET A., Dow E., ABEL L., HOLDEI~ S., NIHOuL.F~r,A-T~2 C., PONDER B. A. J., MUNN~CIt A., 1994. Mutations of the RETproto-oncogene in Hirschsprung's disease. Nature, 367: 378-380.
GVmENE L. A., TtSttLER A. S., 1976. Establishment of a noradrenergic clonal cell line o/rat pheochromocytoma cells which respond to nerve growth/actor. Proc. Natl. Acad. Sci. USA, 73: 2424-2428.
GvaECO M., SANTORO M., BERUNGmRt M. T., MEULLO R. M., DONGHI R., BONGaV, ZONE I., PmRo~'I M. A., DELDX PORTA G., Fus(:o A., VECCHtO G., 1990. PTC as a novel rearranged form o/the ret proto-oncogene and is frequently detected in vivo in human thyroid papillary carcinomas. Cell, 60: 557-563.
HoEsTmx R. M. W., LANDSVATEF. R. M., CEcct tEPdNI I., STULP R. P., STELWAGEN T., Luo Y., PAS1NI B., HOPPEN- El~J. W. M., WXN AMS'rEL H. C. P., ROMEO G., Ln's C. J. M., BuYs C. H. C. M., 1994. A mutation in the PET proto-oncogene assoctated with multiple endocrine neoplasia type 2B and sporadic medtdlau thyrot~ carcinoma. Nature, 367: 375-376.
J~NG S., Wt~N D., Yu Y., H o l s P. L., Luo Y., RaNG M., T.v, an~ R., AN'IONIO L., t Iu Z., CuPPI.ES R., Louts J. c. , Hu S., AL'raocK B. W., Fox G. M., 1996. GDNF-induced activation of the Ret protein tyrosine kinase is mediated by GDNFR-alpha, a novel receptor for GDNF. Cell, 85: 1113-1124.
LIN L. F., DollEirrY D. H., Line J. D., BI.:K'rEsll S., Cou.INs F., 1993. GDNF: a gkal cell line-derived neu- rotrophic fzLzor for midhrain dopaminergic neurons. Science, 260:113%1132.
MULIa{,AN L. M., KwoK J. B. J., HDu.I~',- C. S., EI.SDON M. J., ENG C., GP, RDNt::I', E., Low~ D. R., Moi.E S. E., Moo~u~ J. K., P..u,I L., PONDH~ M. A., TELENIUS H., TIINNACt.WFE A., PONDEI~ B. A. J., 1993. Germ-line mutations of the RET proto-oncz)gene in multep[e endocrine neoplasia type 2A. Nature, 363: 458- 460.
PA(:~N~S V., M,.XNKOO B., COST, XNT~NI F., 1993. E.xpression of the c-RETproto-oncogene during mouse emb~,oge- nests'. Development, 119: 1005-1017.
ROMEO G., Roac~ ir.."rro P., Luo Y., BAI<)NI" V., Sl-l.:I M., CE{:C~ H~IN~ L, PAs~N~ B., Boc{zt,\m)t R., LEI.ZONI< hl., K,'.;,m.XI~EN tt., Mal~TU(:{:ml.l.o G., 1994. Point mutations affecting the" tyrosine kinase dontain o/the RET proto-oncogene bz Hirschspn~ng ;" dtXease. Nature, 367: 377-378.
SAN(:~IrZ M., SmosSvxT~A(;o L, FRISEN J., HE B., Ln~ S. A., B,xm~acn) M., 1996. Renal agents,.} and the ab- sence of enteric neurons in mice lacking GDNF. Nature, 382: 70-73.
SaNTO~O M., WONt: T. W., A,<o{'a P., SANTOS E., NLvr{)s~ovA B., GluECO M., F~s{:o A., D~ F~ORE P. P., 1994. An epidermal growth jae2tor receptor/ret chintq'ra generates m#ogem'c and transforming signals evidence/or a ret-spectfic signaling pathway. M01. Cell. Bi01., 14: 663-675.
SaaTOi~O M., Car~LOM,~;aO F., ROMANO A., B,.)'vrAt~.o D. P., Da'rH,x~ N. A., GRa-X:o M., Fusco A., VEcc~ uo G., hL~TOS~OWX B., Ka~AUS M. H., D~ F~o~E P. P , 1995. Germ-line mutations of MEN2A and MEN2B acti- vate RET as a dominant transforming gene by_ different ntoleeular mechanisms. Science, 267: 381- 383.
S(:HUC~UU~T A., D'AGaT~ V., L,\RSSON-BLOMBERG L., Cos'rAr~nr~ F., P,\(:HN~S V., 1994. Defects in the kidne? and enters? nervous system of mice lacking the tyrosine kinase receptor Ret. Nature, 367: 380-383.
T,.\KAHAStU M., BUMA Y., Iw:vxlo'ro T., INAGUNL'\ V., IKEDA H., H~u H., 1988. C/ont)ag and expresston o/the ret protooncogene encoding a tyrosine kinase with two potentzal transmembrane domains. Onc0gene, 3: 571-578.
T~E,\NORJ. J. S., GOOD*~DXN L., DE SAUVA6E F., STONE D. M., POULSEN K T., BECK C. D., GmxY C., Am, aANtNi M. P., POLLOCK R. A., HEwri F., PItILLIPS H. 8., GODDAm) A., MOORE M. W., BuJ-BELLO A., D~wEs A. M., Asa~ N., TAmX~L, XSH~ M., VANDLEN R., HENDERSON C. E., ROSEN'nUW A., 1996. Characterization of a multicomponent receptor/or GDNF. Nature, 382: 80-83.

GLIAL CELL LINE-DEIU~D NEUROTROP[IIC FACTOR IGDNF~ STIMU1J\TES RET ACTIVITY 149
Taupp M.. Am'NAS E., FMNZn.BI-~ M., NmssoN A. S., SLEImR B. A., Gai~;o~lu M., KILKENNY C., SAL.~z,~-Gz~t1~: so E., P.\cH~ls V., MiUMaE U., SaluOLA H., SAARMA M., h~]\NEZ C. F., 1996. Functional receptor~or GDNF encoded by the c-Ret proto-oncogene. Nature, 381: 785-789.
F. Carlomagno, R. M. Melillo, G. Salvatore, R. Visconti, G. De Vita, G. Lupoli, M. Marinaccio, M. Santoro, G. Vecchio:
Centro di Endocrinologia ed Oncologia Sperimentale del CNR Dipartimento di Biologia e Patologia Cellutare e Molecolare
Facolt,a di Medicina e Chirurgia Universk'a degli Studi di Napoli <<Federico 1I~>
Via S. Pansini, 5 - 80131 Napou
A. Fusco: Dipartimento di Medicina Sperimentale e Clinica
Facolt'~ di Medicina e Chirurgia di Catanzaro Universita degli Studi di Reggio Calabria
Via T. Campanella, 5 - 88100 CA'rANXM~O
Related Documents











