PHYSIOLOGICAL REVIEWS Vol. 78, No. 1, January 1998 Printed in U.S.A. Glial Calcium: Homeostasis and Signaling Function ALEXEJ VERKHRATSKY, RICHARD K. ORKAND, AND HELMUT KETTENMANN Department of Cellular Neurosciences, Max-Delbru ¨ ck Center for Molecular Medicine, Berlin-Buch, Germany; and Institute of Neurobiology, University of Puerto Rico, San Juan, Puerto Rico I. Introduction 100 II. Methodological Considerations 100 A. Types of glial cells 100 B. Experimental approaches to measure intracellular Ca 2/ 102 III. An Overview of Calcium Homeostasis in Glial Cells 102 A. Resting intracellular Ca 2/ in glial cells 102 B. Ca 2/ -permeable channels 103 C. Ca 2/ storage organelles, intracellular Ca 2/ release, and store-operated channels 103 D. Ca 2/ transporters 105 E. Intracellular Ca 2/ sensors and effectors 105 IV. Voltage-Gated Channels and Depolarization-Induced Calcium Signals 106 A. Schwann cells 106 B. Astrocytes 106 C. Oligodendrocytes 107 D. Mechanisms of glial cell depolarization 109 V. Neurotransmitter-Induced Calcium Signaling in Glial Cells 109 A. Glutamate 109 B. Purines and pyrimidines 115 C. Monoamines 117 D. g-Aminobutyric acid and glycine 118 E. Acetylcholine 119 F. Histamine 119 G. Substance P 120 H. Bradykinin 120 I. Endothelins 120 J. Other agonists linked to intracellular Ca 2/ regulation in glial cells 121 K. Heterogeneity of neurotransmitter receptor expression in glial cells 124 VI. Spatiotemporal Organization of Calcium Signals 125 A. Intracellular Ca 2/ oscillations 125 B. Intercellular Ca 2/ waves 127 VII. Glial Calcium Signaling and Neuron-Glial Interactions 128 VIII. Glial Calcium and Brain Pathology 129 IX. Calcium Signals and Glial Function 130 X. Concluding Remarks: Calcium Signals Are a Consequence of Glial Excitability 130 Verkhratsky, Alexej, Richard K. Orkand, and Helmut Kettenmann. Glial Calcium: Homeostasis and Signaling Function. Physiol. Rev. 78: 99 – 141, 1998. — Glial cells respond to various electrical, mechanical, and chemical stimuli, including neurotransmitters, neuromodulators, and hormones, with an increase in intracellular Ca 2/ concen- tration ([Ca 2/ ] i ). The increases exhibit a variety of temporal and spatial patterns. These [Ca 2/ ] i responses result from the coordinated activity of a number of molecular cascades responsible for Ca 2/ movement into or out of the cytoplasm either by way of the extracellular space or intracellular stores. Transplasmalemmal Ca 2/ movements may be controlled by several types of voltage- and ligand-gated Ca 2/ -permeable channels as well as Ca 2/ pumps and a Na / /Ca 2/ exchanger. In addition, glial cells express various metabotropic receptors coupled to intracellular Ca 2/ stores through the intracellular messenger inositol 1,4,5-trisphosphate. The interplay of different molecular cascades enables the development of agonist-specific patterns of Ca 2/ responses. Such agonist specificity may provide a means for intracellular and intercellular information coding. Calcium signals can traverse gap junctions between glial cells without decrement. These waves can serve as a substrate for integration of glial activity. By controlling gap junction conductance, Ca 2/ waves may define the limits of functional glial networks. Neuronal activity can trigger [Ca 2/ ] i signals in apposed glial cells, and moreover, there is some evidence that glial [Ca 2/ ] i waves can affect neurons. Glial Ca 2/ signaling can be regarded as a form of glial excitability. 99 0031-9333/98 $15.00 Copyright q 1998 the American Physiological Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PHYSIOLOGICAL REVIEWS
Vol. 78, No. 1, January 1998Printed in U.S.A.
Glial Calcium: Homeostasis and Signaling Function
ALEXEJ VERKHRATSKY, RICHARD K. ORKAND, AND HELMUT KETTENMANN
Department of Cellular Neurosciences, Max-Delbru¨ck Center for Molecular Medicine, Berlin-Buch, Germany; and
Institute of Neurobiology, University of Puerto Rico, San Juan, Puerto Rico
I. Introduction 100II. Methodological Considerations 100
A. Types of glial cells 100B. Experimental approaches to measure intracellular Ca2/ 102
III. An Overview of Calcium Homeostasis in Glial Cells 102A. Resting intracellular Ca2/ in glial cells 102B. Ca2/-permeable channels 103C. Ca2/ storage organelles, intracellular Ca2/ release, and store-operated channels 103D. Ca2/ transporters 105E. Intracellular Ca2/ sensors and effectors 105
IV. Voltage-Gated Channels and Depolarization-Induced Calcium Signals 106A. Schwann cells 106B. Astrocytes 106C. Oligodendrocytes 107D. Mechanisms of glial cell depolarization 109
V. Neurotransmitter-Induced Calcium Signaling in Glial Cells 109A. Glutamate 109B. Purines and pyrimidines 115C. Monoamines 117D. g-Aminobutyric acid and glycine 118E. Acetylcholine 119F. Histamine 119G. Substance P 120H. Bradykinin 120I. Endothelins 120J. Other agonists linked to intracellular Ca2/ regulation in glial cells 121K. Heterogeneity of neurotransmitter receptor expression in glial cells 124
VI. Spatiotemporal Organization of Calcium Signals 125A. Intracellular Ca2/ oscillations 125B. Intercellular Ca2/ waves 127
VII. Glial Calcium Signaling and Neuron-Glial Interactions 128VIII. Glial Calcium and Brain Pathology 129
IX. Calcium Signals and Glial Function 130X. Concluding Remarks: Calcium Signals Are a Consequence of Glial Excitability 130
Verkhratsky, Alexej, Richard K. Orkand, and Helmut Kettenmann. Glial Calcium: Homeostasis and SignalingFunction. Physiol. Rev. 78: 99–141, 1998.—Glial cells respond to various electrical, mechanical, and chemicalstimuli, including neurotransmitters, neuromodulators, and hormones, with an increase in intracellular Ca2/ concen-tration ([Ca2/]i). The increases exhibit a variety of temporal and spatial patterns. These [Ca2/]i responses resultfrom the coordinated activity of a number of molecular cascades responsible for Ca2/ movement into or out of thecytoplasm either by way of the extracellular space or intracellular stores. Transplasmalemmal Ca2/ movementsmay be controlled by several types of voltage- and ligand-gated Ca2/-permeable channels as well as Ca2/ pumpsand a Na//Ca2/ exchanger. In addition, glial cells express various metabotropic receptors coupled to intracellularCa2/ stores through the intracellular messenger inositol 1,4,5-trisphosphate. The interplay of different molecularcascades enables the development of agonist-specific patterns of Ca2/ responses. Such agonist specificity mayprovide a means for intracellular and intercellular information coding. Calcium signals can traverse gap junctionsbetween glial cells without decrement. These waves can serve as a substrate for integration of glial activity. Bycontrolling gap junction conductance, Ca2/ waves may define the limits of functional glial networks. Neuronalactivity can trigger [Ca2/]i signals in apposed glial cells, and moreover, there is some evidence that glial [Ca2/]i
waves can affect neurons. Glial Ca2/ signaling can be regarded as a form of glial excitability.
990031-9333/98 $15.00 Copyright q 1998 the American Physiological Society
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78100
I. INTRODUCTION transduction in response to neuronal activity. In addition,we consider what is known of changes in [Ca2/]i that
Glial intracellular calcium ([Ca2/]i), like that of other affect glial function and accompany pathological pro-eukaryotic cells, is highly regulated; the free [Ca2/]i is four cesses.to five orders of magnitude less than that in the narrowsystem of clefts that constitutes the functional extracellu-
II. METHODOLOGICAL CONSIDERATIONSlar environment of the nervous system. There is, there-fore, a steep electrochemical gradient favoring Ca2/ entry;transient cellular activation increasing Ca2/ permeability
A. Types of Glial Cellswill lead to a transient increase in [Ca2/]i . In addition,there are Ca2/ stores within the cell that may release Ca2/
in response to specific intracellular chemical messengers, Glial cells (Fig. 1) are found throughout the verte-brate central nervous system (CNS) (for overview on glialalso increasing [Ca2/]i . These transient rises of [Ca2/]i in
turn trigger or regulate various intracellular events, in- cell biology, see Ref. 223). The macroglial cells, astro-cytes, and oligodendrocytes are of ectodermal origin,cluding metabolic processes, gene expression, and ion
transport systems. Therefore, changes in [Ca2/]i act as an whereas the microglial cells are thought to stem fromthe mesoderm. Astrocytes are probably the most diverseeclectic second messenger system coordinating changes
in the external environment with intracellular processes. population of glial cells. One of their hallmarks is theexpression of intermediate filament proteins, glial fibril-These observations, in a wide variety of cells, have led to
a general appreciation of the specific role of [Ca2/]i in cell lary acidic protein (GFAP) or S100. There is a battery ofcommercially available antibodies that can be used assignaling (for general references, see Refs. 42, 73, 147,
245, 345). Cells have developed specialized machinery to markers to identify astrocytes. However, the expressionof GFAP can vary among astrocytes and can change dur-control the spatial and temporal characteristics of these
Ca2/ signals. These include transmembrane Ca2/ trans- ing development, particularly in pathological conditions.The astrocytic response to injury is marked by an increaseporters and Ca2/-permeable channels, cytoplasmic buff-
ers, and intracellular organelles that are able to accumu- in GFAP expression, and these cells are termed reactiveastrocytes. Astrocytes in culture probably represent reac-late, store, and release Ca2/.
In the nervous system, Ca2/ regulation has been ex- tive astrocytes, since they obviously sense the strangeculture environment and prominently express GFAP. Ex-tensively investigated and well characterized in a variety
of neurons (17, 134, 290, 387, 430). These investigations pression of GFAP is, however, not a marker for all astro-cytes: Muller cells in the retina do not express GFAP un-demonstrate that neuronal [Ca2/]i participates in the con-
trol of important neuronal functions, like electrical excit- der normal conditions, but only under pathological condi-tions. What then is the definition of an astrocyte? Theability, neurotransmitter release, and long-term changes
in synaptic efficacy. In parallel, but to a much lesser ex- answer may be that they characteristically have two con-tact sites, the neuronal membrane (synaptic regions in thetent, knowledge has accumulated on the homeostasis and
role of Ca2/ signaling in glial cells. The perception of the gray matter and axons in the white matter) and the borderof the CNS, either the blood system or the ventricularrole of glia in brain function has changed dramatically
over the last 10 years from that of a supporting glue walls. Astrocytes can be subdivided into three major pop-ulations: radial astrocytes, fibrous astrocytes, and proto-(Greek glia is glue) with mainly trophic functions to that
of a cell with dynamic interactions with neurons actively plasmic astrocytes with transition forms between thesepopulations. Bergmann glial cells of the cerebellum are aparticipating in nervous system function. This change oc-
curred after the development of new techniques, like prominent example of a radial (astrocytic glial cell). Fi-brous astrocytes send a large number of processes intopatch-clamp recording and Ca2/ imaging, that revealed
that glial cells express a wide variety of ion channels and all directions, whereas protoplasmic astrocytes, mainlylocated in the gray matter, have short ramified andneurotransmitter receptors that make them able to detect
and respond to neuronal activity (429, 432). Changes in crimped processes (for review of astrocyte morphology,see Ref. 354).glial [Ca2/]i have been measured under a variety of condi-
tions where glial cells are responding to electrical, me- Oligodendrocytes are the myelin-producing cells ofthe CNS (406). They produce myelin proteins such aschanical, and chemical stimuli. These fluctuations in
[Ca2/]i appear to be a consistent response of glial cells to myelin basic protein, proteolipid protein, myelin-associ-ated glycoprotein, and cyclic nucleotide phosphodiester-changes in the environment that lead to a change in glial
function; they are not passive responses and, therefore, ase. Antibodies against these proteins can be used as oli-godendrocyte markers. In white matter, their functioncan be considered a form of glial excitability mediated by
calcium. This review primarily includes recent insights seems to be well defined: they enwrap axons and form themyelin sheath. Oligodendrocytes are prominently found ininto the major mechanisms involved in the control of
[Ca2/]i and the role of changes in [Ca2/]i in glial signal white matter but can also be found in gray matter. There
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
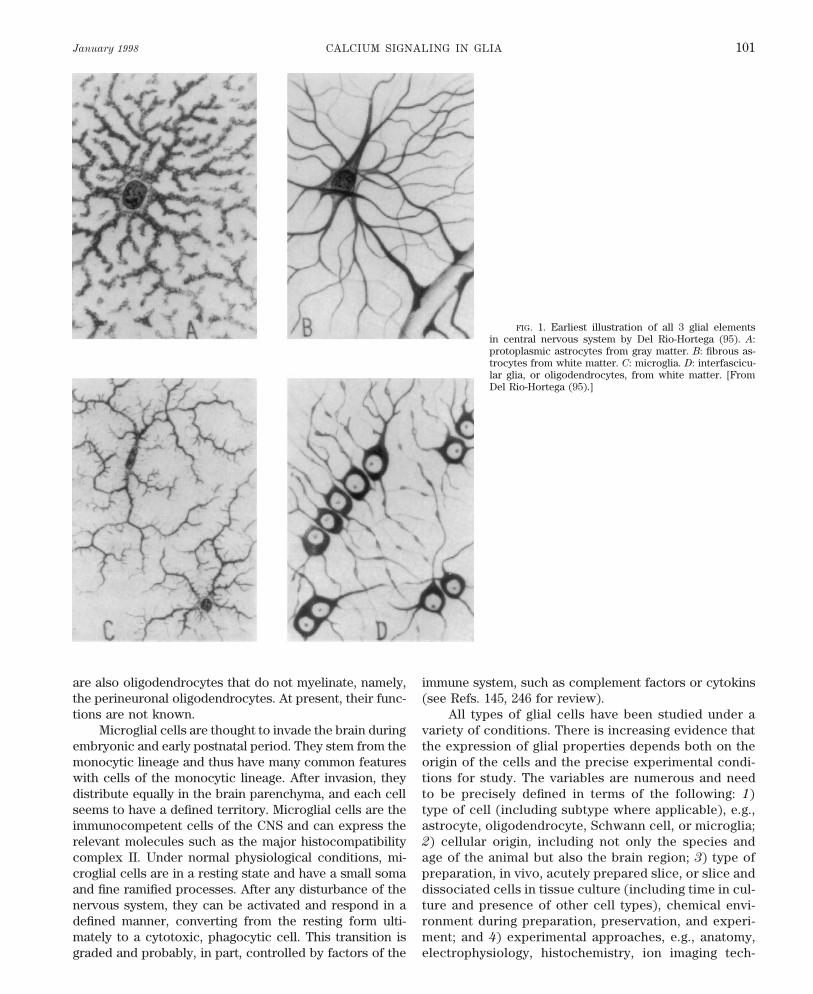
January 1998 CALCIUM SIGNALING IN GLIA 101
FIG. 1. Earliest illustration of all 3 glial elementsin central nervous system by Del Rio-Hortega (95). A:protoplasmic astrocytes from gray matter. B: fibrous as-trocytes from white matter. C: microglia. D: interfascicu-lar glia, or oligodendrocytes, from white matter. [FromDel Rio-Hortega (95).]
are also oligodendrocytes that do not myelinate, namely, immune system, such as complement factors or cytokins(see Refs. 145, 246 for review).the perineuronal oligodendrocytes. At present, their func-
tions are not known. All types of glial cells have been studied under avariety of conditions. There is increasing evidence thatMicroglial cells are thought to invade the brain during
embryonic and early postnatal period. They stem from the the expression of glial properties depends both on theorigin of the cells and the precise experimental condi-monocytic lineage and thus have many common features
with cells of the monocytic lineage. After invasion, they tions for study. The variables are numerous and needto be precisely defined in terms of the following: 1)distribute equally in the brain parenchyma, and each cell
seems to have a defined territory. Microglial cells are the type of cell (including subtype where applicable), e.g.,astrocyte, oligodendrocyte, Schwann cell, or microglia;immunocompetent cells of the CNS and can express the
relevant molecules such as the major histocompatibility 2) cellular origin, including not only the species andage of the animal but also the brain region; 3) type ofcomplex II. Under normal physiological conditions, mi-
croglial cells are in a resting state and have a small soma preparation, in vivo, acutely prepared slice, or slice anddissociated cells in tissue culture (including time in cul-and fine ramified processes. After any disturbance of the
nervous system, they can be activated and respond in a ture and presence of other cell types), chemical envi-ronment during preparation, preservation, and experi-defined manner, converting from the resting form ulti-
mately to a cytotoxic, phagocytic cell. This transition is ment; and 4) experimental approaches, e.g., anatomy,electrophysiology, histochemistry, ion imaging tech-graded and probably, in part, controlled by factors of the
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78102
niques (ion-sensitive dyes, ion probe microscopy), and from the cell of interest so that an actual backgroundfluorescence value can be determined (240).molecular biology.
Given the multitude of variables, it is hardly surpris-ing that there is little unanimity of opinion or even consis-tent results regarding the properties of neuroglia. The di-
III. AN OVERVIEW OF CALCIUMversity of results is tending to focus on a few central
HOMEOSTASIS IN GLIAL CELLSproblems. The control of [Ca2/]i and its variation duringglial responses is one of those problems.
The general mechanisms of [Ca2/]i homeostasis arecommon to all eukaryotic cells (see Refs. 73, 245, 345 forreview). Intracellular Ca2/ is determined by the interac-
B. Experimental Approaches to Measuretion of membrane Ca2/ transporters and cytoplasmic cal-
Intracellular Ca2/
cium buffers (Fig. 2). The Ca2/ transporters are repre-sented by several superfamilies of transmembrane Ca2/-
Attempts to measure [Ca2/]i in glial cells have paral- permeable channels, ATP-driven calcium pumps, and elec-leled those in other tissues and include the use of radioiso- trochemically driven Ca2/ exchangers. The resultingtope tracers, Ca2/ ion-selective electrodes, electron-probe Ca2/ fluxes may either deliver or remove Ca2/ from themicroscopy, and in more recent time Ca2/-sensitive fluo- cytoplasm. Upon entering the cytoplasm, most Ca2/ isrescent dyes (see Refs. 159, 385, 417 for review). Each of trapped by Ca2/-binding proteins; this determines thethe methods has serious limitations that dictate the choice Ca2/-buffering capacity of the cell. Calcium transportersof glial preparation for study. The initial measurements are localized in the cell membrane (providing Ca2/ ex-of [Ca2/]i were carried out in steadily growing cultures of change between the cell interior and exterior) and in theglial cell lines (47) and primary glial cultures (283). As membrane of intracellular organelles [e.g., endoplasmicindicated above, glial cells change during development. reticulum (ER), mitochondria, Golgi complex, and nu-Therefore, it was important to correlate the [Ca2/]i mea- cleus]. The latter forms the intracellular Ca2/ storage sys-surement with the developmental stage. Thus Ca2/ fluxes tem (352), which actively accumulates Ca2/. Accumulatedand [Ca2/]i recordings were carried out along with immu- Ca2/ is bound to intraluminal proteins, and it can be rap-nostaining with stage-specific antibodies (e.g., Refs. 182, idly released via intracellular Ca2/ channels. This general230, 436). scheme is applicable to all types of glial cells (429). A
Experiments on cultured cells raised the question of peculiar feature of glial cells is their high degree of hetero-whether the [Ca2/]i handling mechanisms remain unal- geneity with respect to the expression of various molecu-tered after the cells were removed from their natural envi- lar cascades involved in [Ca2/]i regulation.ronment and maintained under artificial conditions in theabsence of neurons. To solve this problem, [Ca2/]i re-cording techniques were applied first to freshly isolated
A. Resting Intracellular Ca2/ in Glial Cellscells (e.g. Refs. 103, 131) and then to cells in acutely pre-pared brain slices. Initially, the technique combiningpatch-clamp electrophysiological recordings with Ca2/- Free cytoplasmic Ca2/ is a minor part (õ0.001%) of
total calcium in glial cells. Most is associated with intra-sensitive fluorescent dyes was applied to neurons (13,265); later, it was used in glial cells (235, 237, 301). In cellular organelles (e.g., ER, mitochondria, and Golgi ap-
paratus). Resting [Ca2/]i in glial cells varies from 30–40these experiments, the patch-clamp whole cell configura-tion was employed to inject Ca2/-sensitive probes into to 200–400 nM (see Table 1). This variation is not only
among subtypes of glia, but also within the same popula-glial cells. This technique confines the [Ca2/]i recordingto a single, morphologically identified cell and allows si- tion of cells. It may reflect method-induced artifacts or
indicate the flexibility of [Ca2/]i homeostasis. Most mea-multaneous electrophysiological recording. However,prolonged intracellular dialysis can significantly disturb surements were made using membrane-permeable forms
of calcium indicators; thus all the problems associated[Ca2/]i regulation (235). As an alternative, Ca2/-sensitivedye can be injected via microelectrodes into the cell of with this method (uncertain calibration, dye Ca2/ buff-
ering, compartmentalization, and photobleaching) mayinterest (104), or the slices can be incubated with mem-brane-permeant forms of fluorescent Ca2/ probes (33, 239, contribute to the variability. Nevertheless, even in experi-
ments performed on Bergmann glial cells in cerebellar240, 349, 351). A major difficulty with this technique isthat the background fluorescence is unknown; this may slices (235) with careful intracellular calibration proce-
dures, the resting [Ca2/]i ranged from 30 to 200 nM. Thislead to a miscalculation of [Ca2/]i . This problem can beresolved by combining [Ca2/]i measurements from cells variability did not appear to reflect cell damage, because
in all cases the resting potential determined by whole cellloaded with a permeable form of the dye with subsequentintracellular dialysis. The latter helps to wash the dye recordings remained about normal (075 to 060 mV).
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 103
FIG. 2. General scheme of molecular cascades involved in intracellular calcium signaling (see discussion in text).VGCC, voltage-gated Ca2/ channels; SOCC, stores-operated Ca2/ channels; PMCA, plasmalemmal Ca2/-ATPase; Ca2/-BP, Ca2/ binding proteins; InsP3R, inositol 1,4,5-trisphosphate receptor/inositol 1,4,5-trisphosphate-gated Ca2/ channel;RyR, ryanodine receptors/Ca2/-gated Ca2/ channel; SERCA, sarco(endo)plasmic reticulum Ca2/-ATPase. IntracellularCa2/ sensors are as follows: CaM, calmodulin; CaM kinase, Ca2//calmodulin-dependent protein kinase; CaM AC, Ca2//calmodulin-dependent-adenylate cyclase; CaM-phosphatase, Ca2//calmodulin-dependent-protein phosphatase; Ras,p21ras guanine nucleotide-binding proteins; Raf, raf protein kinase; MEK, mitogen-activated/extracellular regulated ki-nase; MAPK, mitogen-activated protein kinase; IEG, immediate early genes.
B. Ca2/-Permeable Channels in almost all glial cell subtypes (397). Finally, certain typesof glial cells (e.g., retinal Muller cells or cultured astro-cytes) express nonspecific cation channels that may alsoThe initial electrophysiological surveys of glial cellspass Ca2/ (227, 356).of various origin (248, 249, 361; see Ref. 392 for review)
did not reveal voltage-sensitive channels. With improvedtechniques, e.g., voltage clamping and patch clamping, the
C. Ca2/ Storage Organelles, Intracellular Ca2/surprising finding was made that some glial cells exhibit a
Release, and Store-Operated Channelsvariety of voltage-gated ion channels that were previouslybelieved to be present only in electrically excitable cells(23, 37, 393). Several populations of both peripheral and Little is known about Ca2/ storage organelles in glial
cells. Many contain an elaborate ER (143, 354) that pre-central macroglia were shown to express voltage-gatedCa2/ channels similar to those found in neurons (392). sumably serves as a major substrate for rapidly exchang-
ing Ca2/ stores. Calcium accumulation by glial ER in-Later, it was found that Ca2/ influx through voltage-gatedchannels significantly increases [Ca2/]i in astro- and oligo- volves ER pumps that, like other cells, are inhibited by
thapsigargin (60, 235, 237) and cyclopiazonic acid (158).dendrocytes as well as in Schwann cells. However, notall glia express Ca2/ channels. For example, Bergmann Aplysia glial cells were found to have an unusual analog
of Ca2/ stores, so called ‘‘gliagrana’’ (217, 218), whichglial cells, microglia, and certain populations of astroglio-mas seem to lack voltage-dependent Ca2/ channels (192, may retain an enormously high (up to 50–100 mM) Ca2/
concentration. The density of these gliagrana varies with239, 429). Nevertheless, Ca2/ may enter glial cytoplasmvia ligand-gated channels that are abundantly expressed fluctuations in extracellular Ca2/ ([Ca2/]o). Increases or
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78104
TABLE 1. Resting [Ca2/]i in glial cells and caffeine and activated under physiological conditions,has been demonstrated only for periaxonal Schwann cells
Resting (258) and for freshly isolated Muller glial cells from sala-Method of [Ca2/]i [Ca2/]i , ReferencePreparation/Cell Type Recording nM No. mander retina (219). In astrocytes, data on CICR are con-
troversial; there is the one report that caffeine triggered aGlial cell lines
[Ca2/]i increase in cultured embryonic cortical astrocytesGlioma C6 Fura 2 75–200 263 (158). In contrast, several observations in cultured andGlioma C6BU-1 Quin 2 10–100 325
freshly isolated astrocytes failed to detect an obvious caf-feine-triggered [Ca2/]i effect (60, 103). However, ryano-Oligodendrocytes
dine and dantrolene, believed to be CICR antagonists,Culture/brain stem Fura 2 70 297modulated [Ca2/]i responses in astrocytes (60, 253). Fi-Culture/cerebrum Indo 1 5–120 107nally, in Bergmann glial cells, studied in cerebellar slices,
Astrocytes caffeine and ryanodine triggered a moderate [Ca2/]i eleva-tion and attenuated [Ca2/]i transients evoked by kainateCulture/rat/cortex Fura 2 34 { 4 203
Culture/rat/hippocampus Fura 2 150 324 (S. Kirischuk and A. Verkhratsky, unpublished observa-Acutely isolated/rat/ Indo 1
tions). In oligodendrocytes, caffeine and ryanodine didhippocampus 45–400 103Culture/rat/cortex Fura 2 85–220 156 not affect [Ca2/]i (238). In general (with several excep-Culture/mouse/cortex Fura 2, indo 1 272 tions), glial [Ca2/]i appears to be insensitive to caffeine,
1–2 wk 300–400reflecting either an absence of ryanodine receptors in glial4–5 wk 100–200
Culture/cortex Fura 2 50–160 140 cells or the specific expression of ‘‘brain’’ ryanodine re-Slice/mouse/cerebellar Fura 2 30–200 235 ceptor isoform (RyR3), which is not modulated by caf-Bergmann glial cells
feine (149, 395). This particular ryanodine receptor sub-[Ca2/]i , intracellular Ca2/ concentration. type could be activated by a newly discovered second
messenger, cyclic ADP ribose (cADPR) (141); the possibleeffect of cADPR on glial [Ca2/]i has not been investigated.
decreases in glial calcium depending on [Ca2/]o suggestThe amount of Ca2/ bound to internal stores may
the possible involvement of glia in the regulation ofalso regulate a distinct type of plasmalemmal Ca2/ perme-
[Ca2/]o . Similar stores have been described in frog epen-ability: it is widely recognized that the depletion of [Ca2/]idymal glia and human astrocytes (143).pools activates a capacitative Ca2/ influx. This influx isThe major mechanism for Ca2/ release from internalassociated with the activation of specific, store-operatedstores involves activation of inositol 1,4,5-trisphosphateplasmalemmal Ca2/ channels (75, 188). The existence of(InsP3)-gated Ca2/ release channels (InsP3 receptors, Refs.these channels in glial cells has not been clearly shown,34, 139). The production of InsP3, in turn, is achieved byalthough there are a number of suggestions that theythe activation of phospholipase C (PLC) coupled via Gmight be important for Ca2/ homeostasis in gliomas (175,proteins with numerous ‘‘metabotropic’’ plasmalemmal362), astrocytes (418), and microglial cells (292, 293), al-receptors. The nature of the InsP3 receptors subtypes inthough at least one group (347) reported that their at-different glial cells is not known in detail. In rat corticaltempts to find the capacitative Ca2/ entry in cultured as-astrocytes and cerebellar Bergmann glial cells, only typetrocytes failed. In microglial cells, the long-lasting activa-3 but not type 1 and 2 InsP3 receptors have been immuno-tion of capacitative Ca2/ entry after the maximal depletionlocalized (453). Oligodendrocytes were reported to tran-of intracellular Ca2/ stores has been described recentlysiently express type 1 InsP3 receptors in a short period(416). Once activated, the capacitative Ca2/ entry pathwayduring the onset of myelination (98). The direct activationin microglial cells remained operative for tens of minutes,of InsP3 receptors by photorelease of InsP3 from cagedcreating a steady-state [Ca2/]i elevation that dramaticallycompound was shown in cultured astrocytes (224, 382).outlasted the period of agonist action.Astrocytic InsP3 receptors appear to be substantially more
Mitochondria are another capacious Ca2/ storagesensitive to InsP3 than InsP3 receptors in Purkinje neu-site. However, their role in [Ca2/]i homeostasis in glialrons; the threshold InsP3 concentration for activation ofcells is little understood. The dissipation of the mitochon-the InsP3-gated channel in astrocytes was 0.2–0.5 mM,drial electrochemical gradient by protonophores [car-whereas in Purkinje neurons, it was 9 mM (224). Inositolbonyl cyanide m-chlorophenylhydrazone (CCCP) and car-1,4,5-trisphosphate-induced Ca2/ release is involved in thebonyl cyanide p-trifluoromethoxyphenylhydrazone] trig-majority of glial responses to neurotransmitters and neu-gered Ca2/ release in oligodendrocytes (236, 238).rohormones (see sect. V). The glial expression of anotherHowever, CCCP treatment did not influence the kinetictype of intracellular Ca2/ release channel, the Ca2/-gatedparameters of the depolarization-triggered [Ca2/]i tran-channel [or ryanodine receptor (RyR); Refs. 138, 287] issients (238). This suggests that mitochondrial Ca2/ accu-still debatable. Functional Ca2/-induced Ca2/ release
(CICR), sensitive to the classical modulators ryanodine mulation does not play an important role in calcium signal
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 105
termination under physiological conditions. Under patho- port in glial cells may be the target for nitric oxide. How-ever, the relative importance of Na//Ca2/ exchanger inlogical conditions, which may substantially disturb mito-
chondria, glial [Ca2/]i homeostasis might be markedly af- regulation of [Ca2/]i was questioned in several reportsthat showed only a minor effect of low [Na/]o on [Ca2/]ifected. Alternatively, mitochondria could play an active
role in Ca2/ signaling also under physiological conditions: (103, 203). Recently, it was demonstrated (156) that adecrease in [Na/]o , by itself, is not sufficient to increasein cultured oligodendrocytes, mitochondrial Ca2/ release/
accumulation actively shaped intracellular Ca2/ waves Ca2/ influx via the Na//Ca2/ exchanger in astrocytes. Nev-ertheless, under conditions of elevated intracellular Na/and [Ca2/]i oscillations originated from InsP3-sensitive
Ca2/ stores (388). In cultured astrocytes, histamine- concentration ([Na/]i), in ouabain-treated cells, lowering[Na/]o produced a dramatic increase in [Ca2/]i . Interest-evoked [Ca2/]i oscillations were accompanied by oscilla-
tions in intramitochondrial free Ca2/ (209), also sug- ingly, a ouabainlike compound has been proposed to actas a vertebrate adrenocortical hormone (163). Thus it isgesting that mitochondrial Ca2/ store may play an active
role in [Ca2/]i homeostasis. possible that the physiological effect of this compoundmight be to increase Ca2/ influx via an increase in [Na/]i .The intracellular distribution of active mitochondria
is different in oligodendrocyte progenitors and mature In Bergmann glial cells in situ, Na//Ca2/ exchangeralso seems to play a relatively minor role in regulatingoligodendrocytes. In the former, both rhodamine-123
staining and CCCP-induced Ca2/ release were confined resting [Ca2/]i . However, Ca2/ flux through the exchangerbecame significant under conditions of elevated [Na/]i .to the tips of cellular processes, suggesting that active
mitochondria were concentrated in these particular areas. Stimulation of Bergmann glial cells with kainate increased[Na/]i to ú30 mM, which turned the exchanger in theConversely, in mature oligodendrocytes, mitochondria
were evenly distributed (236). Presumably, the preferen- reverse mode, providing thus the additional pathway forCa2/ influx. This Ca2/ influx significantly altered both thetial localization of active mitochondria in the processes
of oligodendrocytic progenitors might be important to amplitude and kinetics of the kainate-triggered [Ca2/]i sig-nals (233).supply energy for protein synthesis during cellular
growth; it could also be important for [Ca2/]i handling in An astrocytic Na//Ca2/ exchanger may be an im-portant means for glia to regulate the ionic content in thethis subcellular compartment.interstitium. Neurons, when being electrically excited, candecrease both [Ca2/]o and [Na/]o in the intercellular clefts
D. Ca2/ Transporters (30). Under conditions of lowered [Na/]o , the Na//Ca2/
exchanger could reverse and supply the interstitium withA low [Ca2/]i and recovery from increases in [Ca2/]i Ca2/ by expelling it from adjacent astrocytes.
produced by receptors/channels activation is provided by Finally, Na//Ca2/ exchanger may be involved in medi-plasmalemmal Ca2/ pumps (57) and an electrochemically ating Ca2/ excitotoxicity under pathological conditions.driven Na//Ca2/ exchanger (40). There is little informa- In particular, reversal of Na//Ca2/ exchanger was foundtion on the properties of glial Ca2/ pumps. However, it to play an important role in the astrocytic injury due tohas been shown in oligodendrocytes that La3/-sensitive Ca2/ reperfusion after periods of Ca2/ depletion (a phe-Ca2/-ATPases are primarily responsible for the restora- nomenon similar to the‘‘Ca2/ paradox’’ well described intion of [Ca2/]i after a depolarization-triggered [Ca2/]i in- cardiac muscle). The reperfusion-induced Ca2/ excitotox-crease (238). In contrast, substantially more information icity was significantly decreased in astrocytes in whichis available on the expression of a Na//Ca2/ exchanger. expression of Na//Ca2/ exchanger was inhibited by treat-Initial evidences concerning the existence of functional ment with antisense oligodeoxynucleotides to the ex-Na//Ca2/ exchange in glial cells derived from radiotracer changer (282).experiments demonstrating that transmembrane fluxes of45Ca2/ in glial cells are controlled by extracellular Na/
E. Intracellular Ca2/ Sensors and Effectorsconcentration ([Na/]o) (254). In several astrocytic prepa-rations, reduction of the Na/ gradient, by lowering [Na/]o ,increased [Ca2/]i (96, 156, 158), reduced the Ca2/ efflux After entering the cytoplasm, Ca2/ binds to a number
of proteins that trigger various intracellular signal trans-rate (178, 410), and affected the kinetics of the stimulus-evoked [Ca2/]i (84, 203). Biochemical studies (both the duction pathways (Fig. 2, see Ref. 147 for review). Proba-
bly the best known cytoplasmic Ca2/ sensor is calmodulinpresence of protein and specific mRNA) revealed the ex-pression of a heart isoform of the Na//Ca2/ exchanger in (CaM), which regulates the functional activity of at least
three broad classes of enzymes, namely, CaM-dependentastroglial cells (156, 410). Astrocytic Na//Ca2/ exchangerwas inhibited by 30-min incubation with 0.1–1 mM protein kinases, protein phosphatases, and adenylate cy-
clases. The latter either interact with cytoplasmic en-ascorbic acid (409) and was stimulated by sodium nitro-prusside and 8-bromoguanosine 3*,5*-cyclic monophos- zymes or transfer the signal further down to the nucleus,
initiating other pathways responsible for gene expression.phate (11, 411), suggesting that Na/-dependent Ca2/ trans-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78106
An alternative way to connect cytoplasmic Ca2/ signals 010 mV, and was rapidly inactivated (complete decay ofcurrent tookÇ150–200 ms). The Ca2/ currents in culturedand gene expression is associated with Ras proteins
(small guanine nucleotide-binding proteins) which after Schwann cells were insensitive to L-type Ca2/ channelmodulators (nifedipine and BAY K 8644) but were blockedbeing activated by Ca2/ trigger a cascade of phosphoryla-
tion events that lead to a modulation of gene expression by 5 mM Co2/. In a minor cell subpopulation, a slowlydecaying, nifedipine-sensitive current component was ob-(127). Finally, cytoplasmic Ca2/ signals may propagate to
the nucleus, where they directly stimulate the synthesis served when using 89 mM Ba2/ as a charge carrier. Theexpression of voltage-gated Ca2/ channels should provideof immediate early genes as well as structural genes. Un-
fortunately, little is known of the expression and role of a means for generating [Ca2/]i transients upon Schwanncell depolarization. However, a direct attempt to measurethese systems in glial cells; their characterization in glia is
an important problem awaiting an experimental solution. [Ca2/]i elevation in Schwann cells in a similar DRG cocul-ture (267) failed to detect any measurable [Ca2/]i elevationin response to depolarization by 50 mM KCl.
IV. VOLTAGE-GATED CHANNELS AND Recently, voltage-gated Ca2/ channels were detectedDEPOLARIZATION-INDUCED in perisynaptic Schwann cells at the frog neuromuscularCALCIUM SIGNALS junction (368). Calcium channel expression in these cells
was visualized using either labeling with monoclonal anti-Voltage-gated Ca2/ channels form an important path- bodies against a2/d-subunit (monoclonal antibody 3007;
way for Ca2/ entry in excitable cells; the latter have been Ref. 424) or fluorescent phenylalkylamine (242). Bothfound to express a variety of Ca2/ channels, differing in markers clearly stained the Schwann cell membrane pri-their voltage dependence, kinetics, and pharmacological marily on the processes close to transmitter release sites.properties (177, 190). Calcium channels are integral mem- The morphological observations were substantiated bybrane proteins composed of five subunits, each playing a confocal video imaging of [Ca2/]i that demonstrated thatdistinct role in channel function. The Ca2/ channel sub- perisynaptic Schwann cells respond to high-KCl depolar-units are encoded by several gene families. The functional ization with [Ca2/]i transients sensitive to nimodipineheterogeneity of Ca2/ channels arises mainly from differ- (368). Thus it appears the perisynaptic peripheral glia ex-ences in a1-subunit proteins; at least six major subtypes press functional voltage-gated Ca2/ channels.of a1-subunit have been cloned and characterized (177).On the basis of physiological properties and pharmacolog-
B. Astrocytesical profile, Ca2/ channels are classified as low-voltage-activated or T-type channels and several types of high-
MacVicar (271) first demonstrated Ca2/ action poten-voltage-activated channels (code named as L, N, P, Q, andtials in cAMP-treated cultured cortical astrocytes whenR types). Molecular classification based on the diversitythe K/ conductance was blocked and 10 mM Ba2/ wasof a1-subunit distinguishes six types of Ca2/ channelsadded. Subsequently, similar Ca2/ action potentials were(CaCh1–6). Glial cells, although being inexcitable fromrecorded from Muller glial cells in freshly prepared retinalthe classical point of view (they are not able to generateslices (311), and voltage-clamp experiments on enzymati-action potentials) are capable of expressing voltage-gatedcally dissociated Muller cells revealed currents carried byCa2/ channels. These have been found in several popula-Ca2/ (311).tions of macroglial cells but so far not in microglia.
There are essentially two techniques to detect thepresence of voltage-gated Ca2/ channels, either by charac-
A. Schwann Cells terizing membrane currents using electrophysiologicaltechniques or by recording [Ca2/]i while activating Ca2/
channels with depolarization [commonly by elevating ex-Several early attempts to identify Ca2/ currents (ICa)in cultured Schwann cells (see Ref. 392 for review) were tracellular K/ concentration ([K/]o)]. Calcium currents
were characterized in detail in cultured cortical astrocytesunsuccessful. In 1991, Amedee et al. (5) discovered thatSchwann cells are able to express Ca2/ channels only (24, 71, 85, 273) and type 2 astrocytes from optic nerve
(23, 25). A special treatment of cortical astrocytic cultureswhen cocultured with neurons. Later, it was shown thatexpression of Ca2/ channels in Schwann cells could be was necessary to record Ca2/ currents, namely, the addi-
tion to the culture medium of agents that increase intracel-also induced by addition of a nonhydrolyzable analog ofadenosine 3*,5*-cyclic monophosphate (cAMP) to the cul- lular cAMP. These include treatment with dibutyryl cAMP
(71, 236, 273), forskolin, isoprotereneol, or certain typesture media (28). Whole cell patch-clamp studies ofSchwann cells in organotypic cultures of mouse dorsal of sera (24, 156). Coculturing with neurons had the same
effect (85). In untreated cortical astrocytes, Ca2/ currentsroot ganglia (DRG) revealed voltage-dependent Ca2/ cur-rents. At 10 mM Ca2/ outside, the ICa had an activation were usually undetectable. In contrast, in both freshly
isolated and cultured astrocytes from the optic nerve,threshold at 045 mV, current-voltage curve maximum at
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 107
Ca2/ currents could be recorded without such pretreat- natal (121) cortical astrocytes. In neonatal astrocytes,membrane depolarization with 25–100 mM KCl resultedment (23, 25). These results suggest that certain intracellu-in large increases in [Ca2/]i (up to 1 mM) that were inhib-lar metabolic processes (i.e., phosphorylation) are neces-ited by nimodipine and D-600 (121). In embryonic astro-sary to transfer Ca2/ channels between ‘‘silent’’ and func-cytes, maintained in confluent culture for 4–6 wk, applica-tional pools and furthermore that neurons may regulatetion of 50 mM KCl generated [Ca2/]i transients with anthe availability of Ca2/ channels in certain types of astro-amplitude of 300–400 nM (272). The KCl-triggered [Ca2/]icytes. This also indicates that astrocytes are heteroge-elevation was inhibited by nifedipine and significantly po-neous with respect to Ca2/ channel expression.tentiated by BAY K 8644, suggesting the influx was medi-The parameters of astrocytic Ca2/ currents and theated by L-type Ca2/ channels. In astrocytes kept in a cul-types of Ca2/ channels expressed vary in cells of differentture for only 1–2 wk, 50 mM KCl did not raise (but ratherorigin. Using a double-microelectrode voltage clamp inlowered) [Ca2/]i unless KCl was applied together withcultured cortical astrocytes, MacVicar and Tse (273) re-BAY K 8644. These results imply that astrocytes at earlycorded a Ca2/ current that closely resembled L-type cur-stages in culture have silent Ca2/ channels (272).rents described in neurons. This current inactivated
The variability of [Ca2/]i channels in cultured cellsslowly, had a typical voltage dependence (threshold atraised questions as to the presence and function of volt-020 mV and a maximum of the current-voltage curve atage-gated Ca2/ entry in vivo. Freshly isolated mature hip-/10 mV while using 10 mM Ba2/ as a charge carrier), waspocampal astrocytes (103) promptly respond to KCl appli-completely blocked by 1 mM nifedipine, and was potenti-cation with [Ca2/]i transients (400–800 nM in amplitudeated by b-adrenergic agonists via increased intracellularin response to 50 mM KCl). Potassium chloride-inducedcAMP. The amplitude of ICa in this preparation was quite[Ca2/]i transients were blocked by Co2/ and verapamil,substantial, reaching 4–6 nA at 5 mM extracellular Ba2/
but, in contrast to cultured astrocytes, they were resistantand 10 nA at 10 mM extracellular Ba2/. In contrast, cur-to dihydropyridines; depolarization-induced [Ca2/]i tran-rents recorded from optic nerve astrocytes were muchsients in freshly isolated astrocytes were not affected bysmaller, 200–400 pA, with 10 mM Ba2/ as a charge carrierdibutyryl cAMP. The importance of voltage-gated Ca2/
(23). Furthermore, two components of ICa were recordedchannels for [Ca2/]i regulation was further illustrated byfrom optic nerve astrocytes (23, 25): inactivating (whichrecording [Ca2/]i elevation evoked by depolarizing stepswas defined as a T-type ICa based on its kinetic and voltagein voltage-clamped fura 2-loaded astrocytes from hippo-dependence) and sustained, sharing properties of an L-campal slices (2). Thus the data available suggest thattype ICa (slow inactivation, sensitivity to low concentra-astrocytes in vivo express voltage-gated Ca2/ channels
tions of Cd2/, and voltage dependence).and that Ca2/ influx through these channels substantially
Similarly, small Ca2/ currents (õ250 pA at 5 mM Ca2/affects [Ca2/]i .outside) were recorded recently from immature astro-
cytes in acutely prepared hippocampal slices (Fig. 3, Ref.2). Hippocampal astrocytes in situ appear to express sev-
C. Oligodendrocyteseral types of Ca2/ channels as revealed by their sensitivityto various antagonists. The currents observed at voltagesbetween 050 and 020 mV had parameters typical for T- Oligodendrocytes are heterogeneous with respect totype ICa (fast inactivation and sensitivity to amiloride). the expression of voltage-gated Ca2/ channels. In culturesThe ICa recorded at higher potentials were partially sensi- from cortex, oligodendrocytes expressed both low-volt-tive to nimodipine, verapamil, and v-conotoxin, sug- age (T type) and high-voltage (presumably L type) Ca2/
gesting the coexpression of L- and N-type Ca2/ channels. currents (431, 436). The amplitudes of Ca2/ currents inFinally, with the employment of whole cell and perfo- mature oligodendrocytes were up to about 200 pA with
rated patch-clamp recordings, L-type Ca2/ currents were 20 mM Ca2/ as a charge carrier (Fig. 3). In contrast, volt-recorded in cultured human Muller cells (357). These cur- age-clamp analysis of membrane currents in cultured oli-rents were inhibited by dihydropyridines, but insensitive godendrocytes isolated from rat optic nerve did not revealto v-conotoxins. The reverse transcription (RT)-polymer- Ca2/ currents (23). In situ recordings from oligodendro-ase chain reaction (PCR) examination of total RNA de- cytes in a white matter preparation also failed to detectrived from cultured Muller cells revealed expression of voltage-gated Ca2/ currents (31). It could, however, notmRNAs specific for a1D-, a2-, and b3-channel subunits, be excluded that such channels are present in membranewhere a2-subunit was represented by a splice variant dis- patches remote from the soma such as in the paranodaltinct from skeletal muscle a2S- and brain a2B-isoforms. loops. It is unlikely that even large current injections into
An important question was to find out whether Ca2/ the soma will lead to significant membrane depolarizationinflux via voltage-gated channels alters [Ca2/]i . The initial at such distant regions. In support of such an unevenattempt to resolve this problem employed fura 2 and indo 1 distribution of voltage-gated channels is an observation
by Waxman and colleagues (393), who found that voltage-[Ca2/]i recordings from cultured embryonic (272) and neo-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
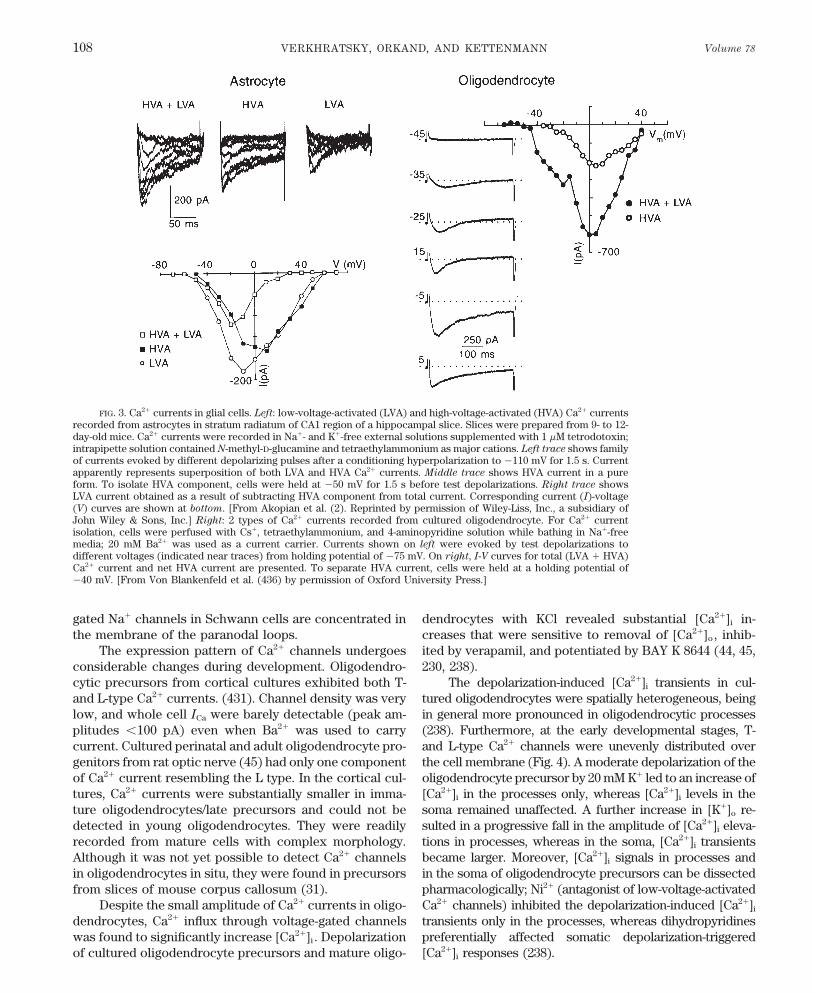
VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78108
FIG. 3. Ca2/ currents in glial cells. Left: low-voltage-activated (LVA) and high-voltage-activated (HVA) Ca2/ currentsrecorded from astrocytes in stratum radiatum of CA1 region of a hippocampal slice. Slices were prepared from 9- to 12-day-old mice. Ca2/ currents were recorded in Na/- and K/-free external solutions supplemented with 1 mM tetrodotoxin;intrapipette solution contained N-methyl-D-glucamine and tetraethylammonium as major cations. Left trace shows familyof currents evoked by different depolarizing pulses after a conditioning hyperpolarization to 0110 mV for 1.5 s. Currentapparently represents superposition of both LVA and HVA Ca2/ currents. Middle trace shows HVA current in a pureform. To isolate HVA component, cells were held at 050 mV for 1.5 s before test depolarizations. Right trace showsLVA current obtained as a result of subtracting HVA component from total current. Corresponding current (I)-voltage(V) curves are shown at bottom. [From Akopian et al. (2). Reprinted by permission of Wiley-Liss, Inc., a subsidiary ofJohn Wiley & Sons, Inc.] Right: 2 types of Ca2/ currents recorded from cultured oligodendrocyte. For Ca2/ currentisolation, cells were perfused with Cs/, tetraethylammonium, and 4-aminopyridine solution while bathing in Na/-freemedia; 20 mM Ba2/ was used as a current carrier. Currents shown on left were evoked by test depolarizations todifferent voltages (indicated near traces) from holding potential of 075 mV. On right, I-V curves for total (LVA / HVA)Ca2/ current and net HVA current are presented. To separate HVA current, cells were held at a holding potential of040 mV. [From Von Blankenfeld et al. (436) by permission of Oxford University Press.]
gated Na/ channels in Schwann cells are concentrated in dendrocytes with KCl revealed substantial [Ca2/]i in-creases that were sensitive to removal of [Ca2/]o , inhib-the membrane of the paranodal loops.
The expression pattern of Ca2/ channels undergoes ited by verapamil, and potentiated by BAY K 8644 (44, 45,230, 238).considerable changes during development. Oligodendro-
cytic precursors from cortical cultures exhibited both T- The depolarization-induced [Ca2/]i transients in cul-tured oligodendrocytes were spatially heterogeneous, beingand L-type Ca2/ currents. (431). Channel density was very
low, and whole cell ICa were barely detectable (peak am- in general more pronounced in oligodendrocytic processes(238). Furthermore, at the early developmental stages, T-plitudes õ100 pA) even when Ba2/ was used to carry
current. Cultured perinatal and adult oligodendrocyte pro- and L-type Ca2/ channels were unevenly distributed overthe cell membrane (Fig. 4). A moderate depolarization of thegenitors from rat optic nerve (45) had only one component
of Ca2/ current resembling the L type. In the cortical cul- oligodendrocyte precursor by 20 mM K/ led to an increase of[Ca2/]i in the processes only, whereas [Ca2/]i levels in thetures, Ca2/ currents were substantially smaller in imma-
ture oligodendrocytes/late precursors and could not be soma remained unaffected. A further increase in [K/]o re-sulted in a progressive fall in the amplitude of [Ca2/]i eleva-detected in young oligodendrocytes. They were readily
recorded from mature cells with complex morphology. tions in processes, whereas in the soma, [Ca2/]i transientsbecame larger. Moreover, [Ca2/]i signals in processes andAlthough it was not yet possible to detect Ca2/ channels
in oligodendrocytes in situ, they were found in precursors in the soma of oligodendrocyte precursors can be dissectedpharmacologically; Ni2/ (antagonist of low-voltage-activatedfrom slices of mouse corpus callosum (31).
Despite the small amplitude of Ca2/ currents in oligo- Ca2/ channels) inhibited the depolarization-induced [Ca2/]i
transients only in the processes, whereas dihydropyridinesdendrocytes, Ca2/ influx through voltage-gated channelswas found to significantly increase [Ca2/]i . Depolarization preferentially affected somatic depolarization-triggered
[Ca2/]i responses (238).of cultured oligodendrocyte precursors and mature oligo-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
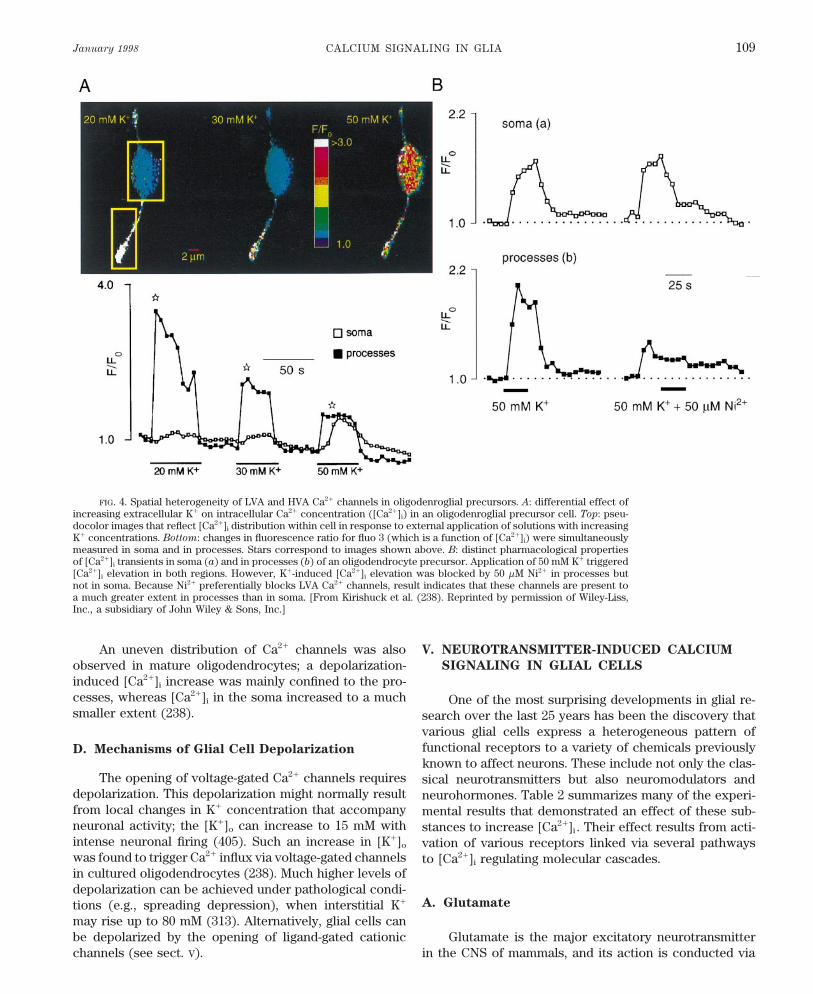
January 1998 CALCIUM SIGNALING IN GLIA 109
FIG. 4. Spatial heterogeneity of LVA and HVA Ca2/ channels in oligodenroglial precursors. A: differential effect ofincreasing extracellular K/ on intracellular Ca2/ concentration ([Ca2/]i) in an oligodenroglial precursor cell. Top: pseu-docolor images that reflect [Ca2/]i distribution within cell in response to external application of solutions with increasingK/ concentrations. Bottom: changes in fluorescence ratio for fluo 3 (which is a function of [Ca2/]i) were simultaneouslymeasured in soma and in processes. Stars correspond to images shown above. B: distinct pharmacological propertiesof [Ca2/]i transients in soma (a) and in processes (b) of an oligodendrocyte precursor. Application of 50 mM K/ triggered[Ca2/]i elevation in both regions. However, K/-induced [Ca2/]i elevation was blocked by 50 mM Ni2/ in processes butnot in soma. Because Ni2/ preferentially blocks LVA Ca2/ channels, result indicates that these channels are present toa much greater extent in processes than in soma. [From Kirishuck et al. (238). Reprinted by permission of Wiley-Liss,Inc., a subsidiary of John Wiley & Sons, Inc.]
An uneven distribution of Ca2/ channels was also V. NEUROTRANSMITTER-INDUCED CALCIUM
SIGNALING IN GLIAL CELLSobserved in mature oligodendrocytes; a depolarization-induced [Ca2/]i increase was mainly confined to the pro-cesses, whereas [Ca2/]i in the soma increased to a much One of the most surprising developments in glial re-smaller extent (238). search over the last 25 years has been the discovery that
various glial cells express a heterogeneous pattern offunctional receptors to a variety of chemicals previouslyD. Mechanisms of Glial Cell Depolarization
known to affect neurons. These include not only the clas-The opening of voltage-gated Ca2/ channels requires sical neurotransmitters but also neuromodulators and
depolarization. This depolarization might normally result neurohormones. Table 2 summarizes many of the experi-from local changes in K/ concentration that accompany mental results that demonstrated an effect of these sub-neuronal activity; the [K/]o can increase to 15 mM with stances to increase [Ca2/]i . Their effect results from acti-intense neuronal firing (405). Such an increase in [K/]o vation of various receptors linked via several pathwayswas found to trigger Ca2/ influx via voltage-gated channels to [Ca2/]i regulating molecular cascades.in cultured oligodendrocytes (238). Much higher levels ofdepolarization can be achieved under pathological condi-
A. Glutamatetions (e.g., spreading depression), when interstitial K/
may rise up to 80 mM (313). Alternatively, glial cells canbe depolarized by the opening of ligand-gated cationic Glutamate is the major excitatory neurotransmitter
in the CNS of mammals, and its action is conducted viachannels (see sect. V).
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78110
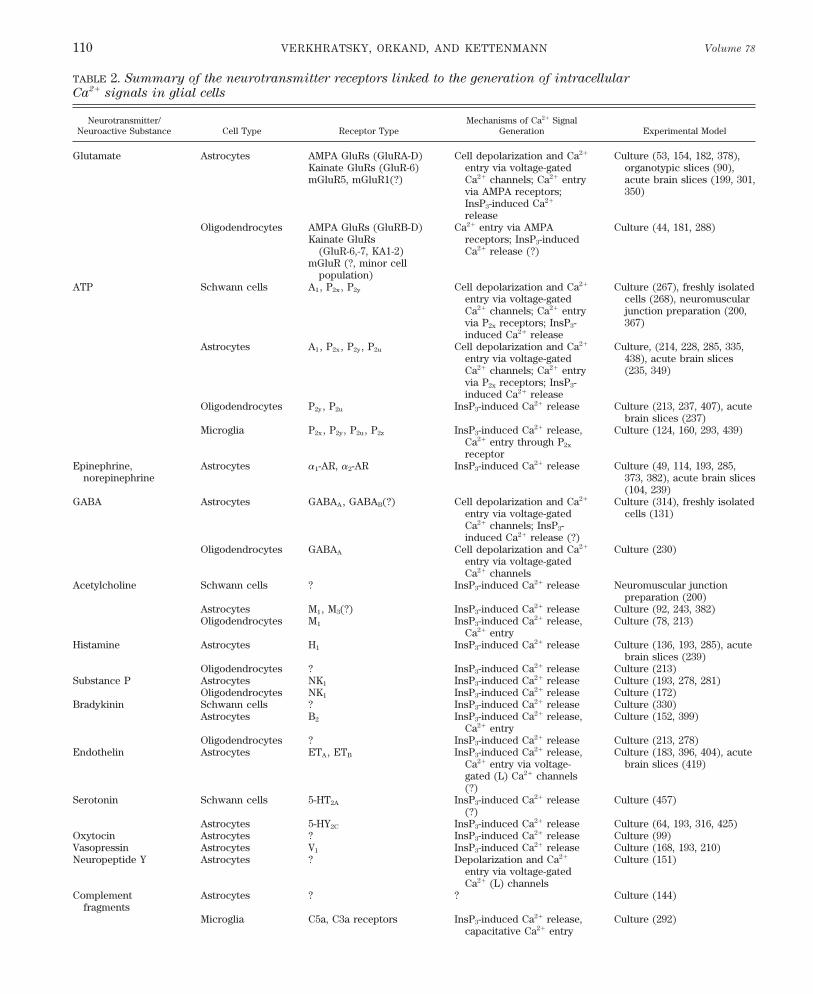
TABLE 2. Summary of the neurotransmitter receptors linked to the generation of intracellular
Ca2/ signals in glial cells
Neurotransmitter/ Mechanisms of Ca2/ SignalNeuroactive Substance Cell Type Receptor Type Generation Experimental Model
Glutamate Astrocytes AMPA GluRs (GluRA-D) Cell depolarization and Ca2/ Culture (53, 154, 182, 378),Kainate GluRs (GluR-6) entry via voltage-gated organotypic slices (90),mGluR5, mGluR1(?) Ca2/ channels; Ca2/ entry acute brain slices (199, 301,
via AMPA receptors; 350)InsP3-induced Ca2/
releaseOligodendrocytes AMPA GluRs (GluRB-D) Ca2/ entry via AMPA Culture (44, 181, 288)
Kainate GluRs receptors; InsP3-induced(GluR-6,-7, KA1-2) Ca2/ release (?)
mGluR (?, minor cellpopulation)
ATP Schwann cells A1, P2x , P2y Cell depolarization and Ca2/ Culture (267), freshly isolatedentry via voltage-gated cells (268), neuromuscularCa2/ channels; Ca2/ entry junction preparation (200,via P2x receptors; InsP3- 367)induced Ca2/ release
Astrocytes A1, P2x , P2y , P2u Cell depolarization and Ca2/ Culture, (214, 228, 285, 335,entry via voltage-gated 438), acute brain slicesCa2/ channels; Ca2/ entry (235, 349)via P2x receptors; InsP3-induced Ca2/ release
Oligodendrocytes P2y , P2u InsP3-induced Ca2/ release Culture (213, 237, 407), acutebrain slices (237)
Microglia P2x , P2y , P2u , P2z InsP3-induced Ca2/ release, Culture (124, 160, 293, 439)Ca2/ entry through P2x
receptorEpinephrine, Astrocytes a1-AR, a2-AR InsP3-induced Ca2/ release Culture (49, 114, 193, 285,
norepinephrine 373, 382), acute brain slices(104, 239)
GABA Astrocytes GABAA, GABAB(?) Cell depolarization and Ca2/ Culture (314), freshly isolatedentry via voltage-gated cells (131)Ca2/ channels; InsP3-induced Ca2/ release (?)
Oligodendrocytes GABAA Cell depolarization and Ca2/ Culture (230)entry via voltage-gatedCa2/ channels
Acetylcholine Schwann cells ? InsP3-induced Ca2/ release Neuromuscular junctionpreparation (200)
Astrocytes M1, M3(?) InsP3-induced Ca2/ release Culture (92, 243, 382)Oligodendrocytes M1 InsP3-induced Ca2/ release, Culture (78, 213)
Ca2/ entryHistamine Astrocytes H1 InsP3-induced Ca2/ release Culture (136, 193, 285), acute
brain slices (239)Oligodendrocytes ? InsP3-induced Ca2/ release Culture (213)
Substance P Astrocytes NK1 InsP3-induced Ca2/ release Culture (193, 278, 281)Oligodendrocytes NK1 InsP3-induced Ca2/ release Culture (172)
Bradykinin Schwann cells ? InsP3-induced Ca2/ release Culture (330)Astrocytes B2 InsP3-induced Ca2/ release, Culture (152, 399)
Ca2/ entryOligodendrocytes ? InsP3-induced Ca2/ release Culture (213, 278)
Endothelin Astrocytes ETA, ETB InsP3-induced Ca2/ release, Culture (183, 396, 404), acuteCa2/ entry via voltage- brain slices (419)gated (L) Ca2/ channels(?)
Serotonin Schwann cells 5-HT2A InsP3-induced Ca2/ release Culture (457)(?)
Astrocytes 5-HY2C InsP3-induced Ca2/ release Culture (64, 193, 316, 425)Oxytocin Astrocytes ? InsP3-induced Ca2/ release Culture (99)Vasopressin Astrocytes V1 InsP3-induced Ca2/ release Culture (168, 193, 210)Neuropeptide Y Astrocytes ? Depolarization and Ca2/ Culture (151)
entry via voltage-gatedCa2/ (L) channels
Complement Astrocytes ? ? Culture (144)fragments
Microglia C5a, C3a receptors InsP3-induced Ca2/ release, Culture (292)capacitative Ca2/ entry
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 111
TABLE 2—Continued
Neurotransmitter/ Mechanisms of Ca2/ SignalNeuroactive Substance Cell Type Receptor Type Generation Experimental Model
Platelet-activating Microglia PAFR InsP3-induced Ca2/ release, Culture (298, 365)factor capacitative Ca2/ entry
(?)Prostanoids Astrocytes FP-R InsP3-induced Ca2/ release Culture (198)Vasoactive intestinal Astrocytes ? InsP3-induced Ca2/ release Culture (119, 415)
polypeptidePDGF Oligodendrocytes ? InsP3-induced Ca2/ release Culture (120, 165)Angiotensin II Astrocytes AT1 InsP3-induced Ca2/ release, Culture (201, 441)
Ca2/ entry via voltage-gated (L) Ca2/ channels(?)
Trombin and albumin Astrocytes ? Intracellular release (?) Culture (87, 304, 321)Arachidonic acid Oligodendrocytes ? Ca2/ entry (?) Culture (391)Opioids Astrocytes m- (?) and k-opioid Ca2/ entry via voltage-gated Culture (116, 170, 401)
receptors (L) Ca2/ channels, InsP3-induced Ca2/ release (?)
Myelin Oligodendrocytes ? Intracellular Ca2/ release Culture (297)(?)
Benzodiazepine Astrocytes ? InsP3-induced Ca2/ release Culture (251)ligands
Reference numbers are given in parentheses. GABA, g-aminobutyric acid; AMPA, DL-a-amino-3-hydroxy-5-methylisoxazole-4-propionic acid;GluR, glutamate receptor; AR, adrenergic receptor; NK, neurokinin; ET, endothelin; 5-HT2, 5-hydroxytryptamine; InsP3, inositol 1,4,5-trisphosphate.
activation of highly diversified families of ionotropic and 1. Schwann cells
metabotropic receptors. The ionotropic glutamate recep-Initial suggestions that GluRs might be expressed bytors (GluRs) are ligand-gated cationic channels assembled
Schwann cells came from experiments on squid axons infrom five subunits. There are three groups of ionotropicwhich nerve stimulation appeared to trigger a hyperpolar-GluRs (according to their pharmacological properties),ization of periaxonal Schwann cells that could be
a-amino-3-hydroxy-5-methylisoxazole-4-propionic acidmimicked by glutamate (434) and blocked by a glutamate(AMPA), kainate, and N-methyl-D-aspartate (NMDA)antagonist (2-amino-4-phosphonobutyrate) (260, 261) or(180). The recent advances in recombinant DNA tech-internal administration of the Ca2/ chelator 1,2-bis(2-niques have given a precise characterization of the GluRsaminophenoxy)ethane-N,N,N*,N*-tetraacetic acid (262).subunit structure and revealed the molecular determi-The latter observation suggested the effect involved annants of GluRs permeability, gating mechanisms, and ago-increase in [Ca2/]i . At the same time, electrophysiologicalnist specificity (180, 207). The GluR subunits A, B, C, andexperiments appeared to reveal NMDA-mediated depolar-D (or 1–4) form AMPA-sensitive receptors, GluR5,izing responses in squid Schwann cells (117, 118). These-6, and -7, and subunits denoted as KA1 and KA2 assem-potentially interesting observations have not been con-
bled to form kainate-preferable GluRs. Finally, the NMDA-firmed in other molluscan species and require confirma-
sensitive GluR subfamily is formed by NMDA R1 andtion with more modern techniques.
NMDA R2A-D subunits (295). Various GluRs subunits canIn mammalian peripheral nerve, Schwann cells were
be differentially assembled forming homo- or heteromericintensively stained by specific antibodies against GluR B
channels that bear different functional properties. Theand D (97), suggesting the possible existence of functional
subunit structure of the GluRs determines their Ca2/ per-AMPA receptors. However, their link to changes in [Ca2/]imeability, with a crucial role for the GluR B subunit; chan-remains unknown. Only a minor fraction (õ10%) of
nels containing GluR B subunit are almost impermeable freshly dissociated Schwann cells from neonatal rat sci-to Ca2/, and those lacking this subunit in the channel atic nerves responded to glutamate with an increase inpentamer are highly Ca2/ permeable (52, 146). [Ca2/]i (267). Thus the involvement of glutamate receptors
Metabotropic glutamate receptors (mGluRs) also in [Ca2/]i control in Schwann cells is still unclear andcomprise a distinct gene family of at least eight members needs further examination.(mGluRs 1–8); the mGluRs belong to the so-called seven-membrane spanning domains receptors (307, 348). The
2. AstrocytesmGluR1 and -5 are coupled (via G proteins) with PLC,being thus the activators of the InsP3-mediated intracellu- The GluRs were probably the first neurotransmitterlar signaling pathway; other mGluRs are connected with receptors found in astroglia. In 1981, Orkand et al. (328)
found that glutamate depolarized glial cells in optic nerveadenylate cyclases.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78112
preparation of Necturus; later, in 1984, Kettenmann et 207). The latter findings stimulated the search for Ca2/-al. (221) and Bowman and Kimelberg (46) showed that permeable GluRs in glia. Initially, it was found that Co2/,excitatory amino acids (glutamate, aspartate, and kainate) which is thought to substitute for Ca2/ as a permeabledirectly depolarized cultured astrocytes. An alternative ion through AMPA/kainate GluRs, permeates and can bemechanism for a glutamate-dependent depolarization is stained within cerebellar type 2 astrocytes, suggesting thestimulation of the electrogenic Na/-dependent glutamate expression of Ca2/-permeable GluRs (355). High-Ca2/ per-transporter by an increase in external glutamate (12, 212). meable AMPA receptors were described in cultured Berg-However, the effectiveness of kainate, a specific agonist mann glial cells, and simultaneously, in situ hybridizationof kainate/AMPA receptors that is not transported by glu- indicated that these cells lack the GluR B subunit (53).tamate transporter, already implied the presence of gluta- These findings were consistent with the hypothesis thatmate receptors. This was substantiated by the observation the presence of GluR B in the channel heteromer inhibitsthat the glutamate effect is mediated by changes in intra- Ca2/ permeability (146). No GluR B subunit mRNA wascellular phosphoinositide turnover and transmembrane found to be associated with the expression of GluR Afluxes of 45Ca2/ (336). More recently, microfluorimetric (mainly) and GluR C subunits in glial cells (presumablytechniques revealed that glutamate induces complex astrocytes) from rat optic nerve (205). Northern blot anal-changes in [Ca2/]i characterized by distinct spatiotempo- ysis of mRNA for AMPA GluRs subtypes performed onral features often in the form of intracellular waves and primary cultured astrocytes revealed that cells isolatedoscillations. These glutamate-triggered Ca2/ responses from the brain stem express predominantly GluR D spe-were mediated by both transmembrane Ca2/ entry and cific mRNA (81).intracellular Ca2/ release, indicating the involvement of Cortical astrocytes and astrocyte progenitors ex-several types of GluRs (225). pressed GluR B mRNAs as well as mRNAs encoding GluRs
A) AMPA/KAINATE IONOTROPIC GLUTAMATE RECEPTORS. A, C, D and GluR6 subunits, as demonstrated by both1) Cultured cells. The initial observations of excitatory Northern blots (81) and RT-PCR technique (182). Despiteamino acid-induced depolarization of astrocytes were the apparent presence of GluR B subunit, these cells re-substantiated in voltage-clamp experiments that demon- spond to kainate with large [Ca2/]i transients (182). Thesestrated that glutamate, quisqualate, AMPA, and kainate, transients were not modified by Na/ removal from thebut not NMDA triggered Na//K/ currents in cultured as- bath and were not attenuated when intracellular Ca2/
trocytes from cerebrum and cerebellum (394, 449). These stores were blocked with thapsigargin. Furthermore, stim-currents were blocked by the specific antagonists of ulation of astrocytes by kainate, when external Ca2/ wasAMPA/kainate receptors, 6-cyano-7-nitroquinoxaline-2,3- replaced by Co2/ and [Na/]o was removed, caused fastdione (CNQX) and 6,7-dichloro-3-hydroxy-2-quinoxa- quenching of fura 2 signals, indicating that Co2/ enteredlinecarboxylic acid (394, 448). Single-channel recordings the cell via kainate-activated channels. These results sug-revealed that glutamate-activated currents had several gested that [Ca2/]i elevation in cortical astrocytes resultedconductance levels, and their kinetic properties were sim- mainly from Ca2/ entry via AMPA/kainate receptors (182).ilar to those for AMPA/kainate receptors in neurons (422, However, kainate-induced currents, measured under volt-448). Thus experimental evidence suggests that astrocytes age-clamp conditions in the same cells, were drasticallyare endowed with AMPA/kainate GluRs. decreased (Ç40 times) in the absence of Na/, suggesting
a low Ca2/ permeability of the receptor. Similarly, glialRecordings of [Ca2/]i with fura 2-based microfluori-metry demonstrated that kainate and AMPA (112, 154) cells (most likely immature astrocytes) acutely isolated
from the hippocampal CA1 stratum radiatum region, ex-raised [Ca2/]i in cultured cerebral, hippocampal, and cere-bellar astrocytes. This [Ca2/]i rise depended on [Ca2/]o hibited a low or intermediate Ca2/ permeability, as deter-
mined by potential-dependent characteristics of kainate-and was blocked by CNQX. Similar AMPA- and kainate-evoked [Ca2/]i transients were observed in retinal Muller induced ionic currents (378). Nevertheless, even these
small Ca2/ currents via low-Ca2/ permeability AMPA/kai-cells (437). In mixed cultures from neonatal rat brains, theCNQX-sensitive AMPA/kainate-triggered [Ca2/]i transients nate receptors are able to appreciably increase [Ca2/]i in
astrocytes. This may indicate a low Ca2/ buffer capacitywere mostly confined to type 1 astrocytes (204), sug-gesting differential expression of GluRs in astroglia. Thus in cortical astrocytes.activation of AMPA/kainate receptors in astrocytes pro- 2) In situ preparations. Initial evidence for the ex-motes Ca2/ influx that might result either from depolariza- pression of functional GluRs in glial cells in situ was re-tion-triggered activation of voltage-gated channels or from vealed by microelectrode recordings from astrocytes indirect Ca2/ influx through GluRs. rat hippocampal slices and amphibian optic nerve; appli-
cation of glutamate depolarized these astrocytes (414,Initial experiments on glutamate-induced Ca2/ signal-ing in glial cells coincided with the detection of Ca2/ per- 440). Later, patch-clamp recordings revealed AMPA-, kai-
nate-, and quisqualate-induced ionic currents sensitive tomeability of AMPA/kainate GluRs in neurons (179, 191)and the subsequent discovery of its molecular basis (52, CNQX in rabbit retinal astrocytes from in situ preparation
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
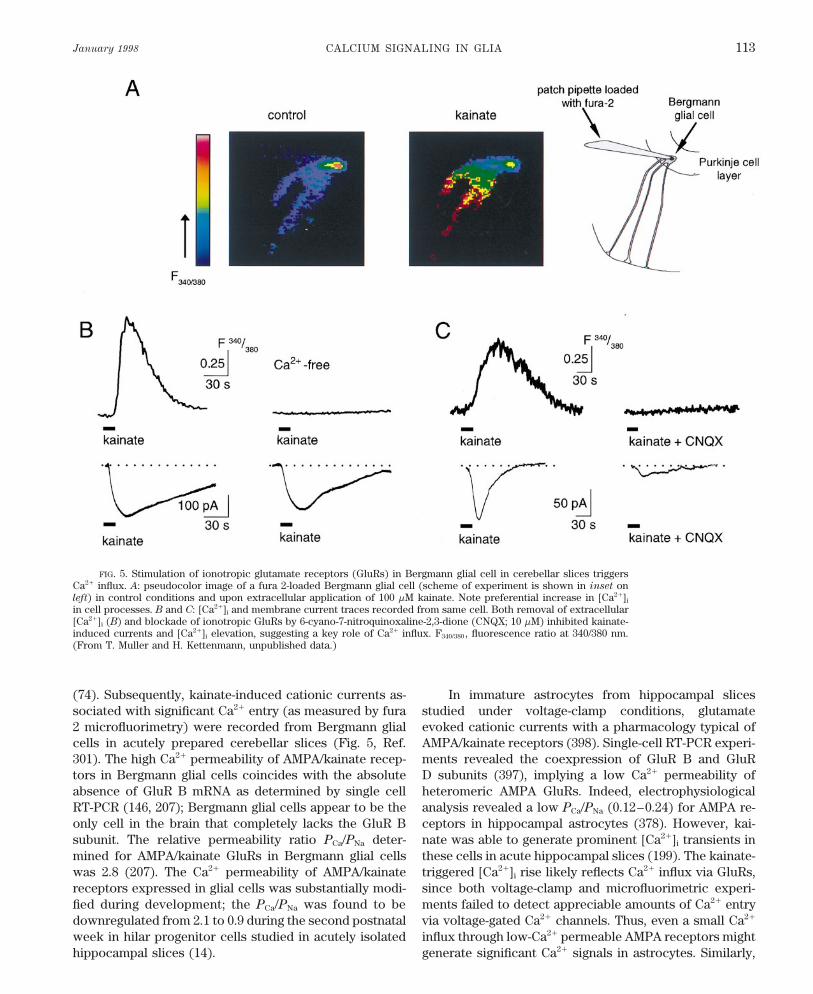
January 1998 CALCIUM SIGNALING IN GLIA 113
FIG. 5. Stimulation of ionotropic glutamate receptors (GluRs) in Bergmann glial cell in cerebellar slices triggersCa2/ influx. A: pseudocolor image of a fura 2-loaded Bergmann glial cell (scheme of experiment is shown in inset onleft) in control conditions and upon extracellular application of 100 mM kainate. Note preferential increase in [Ca2/]i
in cell processes. B and C: [Ca2/]i and membrane current traces recorded from same cell. Both removal of extracellular[Ca2/]i (B) and blockade of ionotropic GluRs by 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX; 10 mM) inhibited kainate-induced currents and [Ca2/]i elevation, suggesting a key role of Ca2/ influx. F340/380 , fluorescence ratio at 340/380 nm.(From T. Muller and H. Kettenmann, unpublished data.)
(74). Subsequently, kainate-induced cationic currents as- In immature astrocytes from hippocampal slicesstudied under voltage-clamp conditions, glutamatesociated with significant Ca2/ entry (as measured by fura
2 microfluorimetry) were recorded from Bergmann glial evoked cationic currents with a pharmacology typical ofAMPA/kainate receptors (398). Single-cell RT-PCR experi-cells in acutely prepared cerebellar slices (Fig. 5, Ref.
301). The high Ca2/ permeability of AMPA/kainate recep- ments revealed the coexpression of GluR B and GluRD subunits (397), implying a low Ca2/ permeability oftors in Bergmann glial cells coincides with the absolute
absence of GluR B mRNA as determined by single cell heteromeric AMPA GluRs. Indeed, electrophysiologicalanalysis revealed a low PCa/PNa (0.12–0.24) for AMPA re-RT-PCR (146, 207); Bergmann glial cells appear to be the
only cell in the brain that completely lacks the GluR B ceptors in hippocampal astrocytes (378). However, kai-nate was able to generate prominent [Ca2/]i transients insubunit. The relative permeability ratio PCa/PNa deter-
mined for AMPA/kainate GluRs in Bergmann glial cells these cells in acute hippocampal slices (199). The kainate-triggered [Ca2/]i rise likely reflects Ca2/ influx via GluRs,was 2.8 (207). The Ca2/ permeability of AMPA/kainate
receptors expressed in glial cells was substantially modi- since both voltage-clamp and microfluorimetric experi-ments failed to detect appreciable amounts of Ca2/ entryfied during development; the PCa/PNa was found to be
downregulated from 2.1 to 0.9 during the second postnatal via voltage-gated Ca2/ channels. Thus, even a small Ca2/
influx through low-Ca2/ permeable AMPA receptors mightweek in hilar progenitor cells studied in acutely isolatedhippocampal slices (14). generate significant Ca2/ signals in astrocytes. Similarly,
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78114
GFAP-positive mature rat hippocampal astrocytes re- Biochemical investigations clearly demonstrated an in-crease in intracellular InsP3 level in glutamate-treated as-sponded with a [Ca2/]i increase to kainate and AMPA;troglial cells (289, 336). The mGluR-mediated Ca2/ signal-these [Ca2/]i responses were blocked by CNQX (350). Fur-ing is widespread in astrocytes. A majority of [Ca2/]i re-thermore, neither verapamil nor nifedipine prevented kai-cordings from cultured astrocytes (1, 83, 129) suggest thatnate-induced [Ca2/]i transients, suggesting that Ca2/ influxa substantial part of the glutamate-induced [Ca2/]i eleva-occurred through GluRs.tion persists in Ca2/-free extracellular solutions, indicat-Finally, [Ca2/]i recordings using confocal microscopying that the Ca2/ comes from internal stores. These intra-demonstrated glutamate-evoked [Ca2/]i transients in peri-cellular Ca2/ responses were mimicked by a specific ago-axonal glial cells (presumably astrocytes) in optic nervesnist of mGluRs (1S,3R)-ACPD (129), pointing to the(247). The [Ca2/]i rise in periaxonal glial cells in the opticinvolvement of the GluR-InsP3 signal transduction chain.nerve was also elicited by 1-aminocyclopentane-1,3-dicar-The nature of astrocytic mGluRs is still unclear; the ex-boxylate [(1S,3R)-ACPD] and AMPA and was partially sen-pression of mGluR3 and mGluR5 only was found in gliasitive to the AMPA antagonist 6,7-dinitroquinoxaline-(370, 413). Strong mGluR5-dependent immunostaining2,3(1H,4H)-dione, suggesting the functional expression ofwas found in astrocytic processes in hypothalamus in situboth iono- and metabotropic GluRs in optic nerve glial(426); these processes surround complex synapses;cells.mGluRs may well be exposed to glutamate during synap-B) NMDA RECEPTORS. In one study, an NMDA-inducedtic activity. The importance of mGluRs in triggering Ca2/
[Ca2/]i rise was observed (1) in some cultured spinal cordresponses in astroglia was also confirmed by in situ exper-astrocytes. In most other studies of cultured astrocytes,iments. The [Ca2/]i transients mediated via mGluRs wereexamined with both electrophysiological techniques (394,found in both astrocytes in hippocampal slices (350) and422, 448, 449) and Ca2/-sensitive fluorescent dyes (182),Bergmann glial cells in cerebellar slices (232). In the latter,NMDA caused no effects on membrane permeability andthe expression of both mGluR1 and mGluR5 was deter-[Ca2/]i . A notable exception was experiments on radialmined by using single-cell RT-PCR (Kirischuk, Matiash, F.glia; studies of cultured retinal Muller cells found thatKirchhoff, H. Kettenmann, and A. Verkhratsky, unpub-glutamate promoted their proliferation via activation oflished observations). The relative expression of mGluR1/receptors with NMDA receptor (NMDAR) pharmacologymGluR5 receptors could be important for the shaping of(421). Subsequent electrophysiological observations de-glutamate-evoked [Ca2/]i transients. It has been demon-tected NMDA-evoked currents in Muller cells (358). Instrated recently that transfected cells that express exclu-
another type of radial glia, cerebellar Bergmann glial cellssively mGluR5 respond to glutamate with [Ca2/]i oscilla-
voltage-clamped in cerebellar slices, bath application oftions, whereas cells expressing mGluR1 had single-peak
1 mM NMDA evoked tiny (Ç30–60 pA) currents (300). In[Ca2/]i responses (216). The question of whether mGluR1/
situ hybridization revealed a significant level of expres-mGluR5 might be important for determining the kinetic
sion of NMDAR1 and -2B subunits mRNA in these cells characteristics of glial [Ca2/]i responses remains to be(266), although the exact composition of NMDA receptors clarified.assembled in the membrane remains unknown. TheNMDA-activated currents in Bergmann glial cells were
3. Oligodendrocytesnot associated with measurable changes in [Ca2/]i (300).Similarly, fura 2-based experiments failed to detect any Electrophysiological studies of cultured immature[Ca2/]i changes in retinal Muller cells challenged with oligodendrocytes and their progenitors (44, 142, 334) asNMDA (437). Finally, NMDA-activated currents have been well as oligodendrocyte progenitors in corpus callosumobserved in neocortical protoplasmic astrocytes (229) and slices (32) found glutamate-, AMPA-, and kainate-trig-in a small population of hippocampal astrocytes (398). gered ionic currents that were blocked by CNQX, sug-The question of whether NMDA can induce [Ca2/]i in- gesting AMPA/kainate ionotropic receptors were stimu-creases in astrocytes remains unclear. Using confocal lated. Indeed, Northern blots revealed the expression ofvideo imaging of hippocampal astrocytes, Porter and Mc- GluRs B, C, and D, GluR6 and -7, and KA1 and KA2 mRNAsCarthy (350) observed [Ca2/]i transients in response to in cells of the oligodendrocyte lineage (334). High expres-bath applications of NMDA; however, these Ca2/ re- sion of GluR B subunit implies a low Ca2/ permeabilitysponses could have been triggered indirectly by activation of oligodendrocyte AMPA/kainate channels.of neuronal terminals in the hippocampal slices with sub- Intracellular Ca2/ recordings from oligodendrocytesequent release of glutamate and activation of non- cultures also showed increases after application of gluta-NMDAR in glial cells. mate and its agonists. According to Borges et al. (44),
C) METABOTROPIC GLUTAMATE RECEPTORS. Another im- cytoplasmic Ca2/ increases after the activation of AMPA/portant route for generating Ca2/ signals in astroglia is kainate receptors in oligodendrocytic precursors resultedassociated with the activation of mGluRs and subsequent mainly from Ca2/ influx via voltage-gated channels. Other
authors (181, 182, 288) suggest that Ca2/ influx throughCa2/ release via InsP3-gated intracellular Ca2/ channels.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 115
GluRs can be also involved. Minor populations of cultured among glial cells. A majority of glial cells studied (includ-ing peripheral glia, macro- and microglia from the CNS)oligodendrocytes also exhibited mGluRs (181). The ex-
pression of AMPA/kainate GluRs in cells of the oligoden- appear to express various types of purinoreceptors. Inmany cases, activation of glial purinoreceptors leads todrocyte lineage was found to be developmentally down-
regulated. Mature oligodendrocytes lost the ability to re- increases in [Ca2/]i .spond to glutamate by activation of membrane currents
1. Schwann cells(44). In another study, the upregulation of GluR B subunit
An ATP-triggered increase in [Ca2/]i was first demon-abundance during transition of O2-A and CG-4 progenitorsstrated by Jahromi et al. (200) who monitored [Ca2/]i ininto oligo- or astrocytes was demonstrated (288). Like-perisynaptic Schwann cells at the frog neuromuscularwise, in optic nerves, quisqualate-stimulated Co2/ uptakejunction; they also suggested that Schwann cells respond(which is believed to reflect Co2/ entry through Ca2/-per-with a [Ca2/]i increase to synaptically released ATP aftermeable AMPA receptors) only in O-2A progenitor cellselectrical stimulation of the axon. The calcium transientsbut not in mature glia (137).recorded by Jahromi et al. (200) were confined to perisy-naptic Schwann cells only; the myelinating Schwann cellssituated periaxonally did not respond to ATP. IntracellularB. Purines and PyrimidinesCa2/ transients induced by ATP and mediated via activa-tion of P2y receptors were found in a majority of freshlyAdenosine 5*-triphosphate, adenosine, and relateddissociated Schwann cells from neonatal rat sciatic nervesubstances control a number of important physiological(267) as well as in periaxonal sciatic nerve Schwann cellsreactions and act as neurotransmitters in the peripheralstudied in situ (268). Interestingly, P2y-driven [Ca2/]i re-nervous system and CNS (54, 102, 133). In recent years,sponses in neonatal rat Schwann cells were significantlyclear evidence that ATP acts as an excitatory neurotrans-downregulated when cells were maintained in culture formitter in the CNS has been obtained (108, 109), and phar-several days. However, these responses were restored ei-macological and molecular characterization of ATP andther by the addition of membrane permeable cAMP ana-adenosine receptors (named purinoreceptors) waslogs (dibutyryl cAMP or 8-bromo-cAMP) to the cultureachieved. Purinoreceptors are represented by a broadmedia (267) or coculturing glial cells with DRG neuronsfamily of proteins classified into two major groups (88,(267, 268). In Schwann cell cultures obtained from adult133): 1) adenosine receptors (or P1 purinoreceptors code-rats and rabbits, the predominant receptor type was P2unamed also as A1-A4 receptors) coupled mainly with ade-(9), suggesting that the expression of particular purinore-nylate cyclases as well as with PLC (A1 receptors) and 2)ceptor subtypes might undergo developmental changes.receptors for ATP and related nucleotides known as P2Finally, an analysis of purinoreceptors at the frog neuro-purinoreceptors. On the basis of pharmacological proper-muscular junction perisynaptic Schwann cells revealedties, the P2 purinoreceptor family is subclassified into twometabotropic A1 adenosine and P2y purinoreceptors thatgroups: ionotropic receptors (P2x and P2z) and metabotro-both induce intracellular Ca2/ release and involve pertus-pic receptors (P2y , P2u , P2t and P2d). Advances in molecularsis toxin (PTX)-sensitive G protein transductioncloning extended this classification by showing that puri-pathways, as well as ionotropic P2x receptors that act vianoreceptors are encoded by two distinct gene familiesmembrane depolarization and Ca2/ entry through plas-(55). The family of P2x receptors comprise several sub-malemmal L-type Ca2/ channels (367). Voltage-clamp ex-types (labeled P2x1–7) of ligand-gated ionic channels withperiments on cultured mouse DRG Schwann cells alsoa unique two transmembrane domain topology; the mem-revealed the existence of an ATP-triggered cationic cur-
bers of P2x family differ in their ion selectivity and gatingrent, sensitive to P2 purinoreceptor antagonist suramin
properties. Cloned P2z receptors also belong to the P2x and carried to certain extent by Ca2/ (4). The unusuallygene family, and they are codenamed as P2x7 subtype;
high concentrations of ATP required to activate this cur-P2x7(z) receptors are large transmembrane pores that are
rent (dissociation constant Ç8.4 mM) suggests P2z recep-activated in fact by a tetraanionic form of ATP (ATP40)
tors are involved. All experimental data available indicateand may pass molecules with a molecular mass up to 1
that Schwann cells express purinoreceptors linked to thekDa. Cloned metabotropic receptors are classified as P2y regulation of [Ca2/]i , and this might be an important path-family, being represented by seven members (P2y1-P2y7). way for neuronal-glial interaction in the peripheral ner-They all are similar to other G protein-linked metabotro- vous system. This interaction is presumably accomplishedpic receptors by their seven-transmembrane domain via either synaptic or nonsynaptic release of purines afterstructure and are often associated with PLC and hence axonal electrical activity.InsP3 turnover. The P2y1 receptor pharmacologically
2. Astrocytesmatches P2y subtype, P2y2-P2u , P2y3 probably correspondsto P2t receptor; P2y4–7 are not yet assigned with known Numerous experiments on cultured astrocytes dem-subtypes. onstrate that they express both adenosine (P1) and P2
purinoreceptors involved in the regulation of [Ca2/]i .Purinoreceptors are unusually widely distributed
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78116
A) ADENOSINE (P1) PURINORECEPTORS. Apart from A1 have also been described in astrocyte-derived glioma cells(87). The coexpression of both P2y and P2u receptors wereand A2 receptors present in astroglia, which either inhibitdemonstrated for cultured cortical astrocytes from neona-(A1) or stimulate (A2) adenylate cyclase, recent experi-tal rat (228). That was shown by injecting the Xenopusments demonstrated that adenosine may also increaseoocytes with mRNA collected from confluent astrocytic[Ca2/]i in astrocytes (96, 335). This [Ca2/]i increase origi-cultures. The injected oocytes expressed both P2u andnates from intracellular Ca2/ stores. It was mimicked byP2y receptors as demonstrated by their pharmacologicalselective A1 receptor agonist N6-cyclopentyladenosine andprofile.inhibited by A1 antagonist 8-cylopentyl-1,3-dipropylxan-
Activation of P2y receptors leads to an increase intine (335). Additionally, stimulation of A1 receptors wasintracellular InsP3 in cultured cortical astrocytes (214,reported to potentiate the [Ca2/]i mobilizing activity of338, 340), resulting in InsP3-driven Ca2/ release from inter-other neuroactive substances, including histamine (335),nal stores. The involvement of PLC controlled InsP3 pro-substance P (96) and the mGluR agonist (1S,3R)-ACPDduction and activation of InsP3-gated ER Ca2/ channels(324). The ability of adenosine to induce Ca2/ mobilizationwas directly demonstrated in experiments on cultured as-from internal stores was further substantiated in video-trocytes from rat cerebral cortex and spinal cord. In bothimaging experiments on astrocytes in acutely isolated ratpreparations, an increase in intracellular InsP3 [by eitherhippocampal slices (349). The agonist/antagonist sensitiv-flash photolysis (382) or intracellular dialysis (375)] mim-ity of adenosine-induced [Ca2/]i transients in slice prepa-icked the effects of ATP. The sensitization of InsP3 recep-rations did not match the pharmacological profile for A1,tors by incubation of cultured astrocytes with timerosalA2 or A3 receptors; the possible involvement of A2b recep-dramatically increased the amplitude of ATP-inducedtors (452) was suggested. The [Ca2/]i mobilization medi-[Ca2/]i transients (347). Furthermore, an ATP-inducedated through adenosine receptors was reported to trigger[Ca2/]i increase was inhibited by heparin, an intracellularnitric oxide release from cultured astrocytes (202).antagonist of InsP3-gated Ca2/ channels, and prevented
B) P2 PURINORECEPTORS. Many studies of cultured as-by pharmacological inhibition of PLC (375).trocytes have shown that ATP increases [Ca2/]i . Both io-
The existence of functional P2y receptors in astroglialnotropic and metabotropic P2 purinoreceptors appear tocells was demonstrated in situ in Bergmann glial cellsparticipate in this response. The ATP-stimulated Ca2/ in-studied in acutely isolated cerebellar slices (Fig. 6). Aden-flux was found first in experiments measuring 45Ca2/ up-osine 5*-triphosphate and P2y agonists promptly raisedtake in astrocytes (309). This effect of ATP was inhibited[Ca2/]i (235). As shown in Figure 6, these Ca2/ responsesby LaCl3 , suggesting Ca2/ entered via plasmalemmal Ca2/were not affected by removal of external Ca2/ and were
channels. Subsequent current- and voltage-clamp studiesnot associated with measurable transmembrane currents.
found that ATP depolarized cultured cerebral astrocytesThus intracellular stores appear to be the exclusive source
by triggering an inward cationic current, most likely dueof Ca2/. This was confirmed in experiments in which the
to stimulation of P2x receptors (274, 438). This cationicATP-induced [Ca2/]i rise was blocked by blockade of ER
current may activate voltage-gated Ca2/ channels (438).pumps with thapsigargin, as well as intracellular perfusion
Alternatively, P2x receptors may have an appreciable Ca2/of these cells with heparin. Such experiments confirm that
permeability. Experiments with other cells suggest thatthe InsP3-induced Ca2/ release from ER Ca2/ stores is
several P2x receptor subtypes have a significant Ca2/ per- involved in the ATP response (235).meability (423). The Ca2/ current can be up to 6.5% of thetotal under physiological conditions (369). Furthermore,astrocytic calcium signaling could be mediated through 3. OligodendrocytesP2z ionotropic receptors as demonstrated on primary cul-tured rat cortical astrocytes (19). Cells of the oligodendrocyte lineage respond to ATP
Another important route for [Ca2/]i increases in as- by generating [Ca2/]i transients mediated by activation oftrocytes results from metabotropic P2 receptor stimula- metabotropic P2y and/or P2u purinoreceptors (213, 237,tion. Studies on purinoreceptor agonists suggest that the 407). The ATP-driven [Ca2/]i responses remain in Ca2/-predominant type of metabotropic ATP receptor ex- free solutions and are inhibited by thapsigargin and intra-pressed by astroglia is the P2y (214, 285, 374, 375), although cellular dialysis with heparin (237), indicating a role forthere are reports that primary cultured cortical (50, 338) the InsP3-mediated intracellular signal transduction chain.and dorsal spinal cord (176) astroglial cells express P2u Experiments in situ, in corpus callosum slices, also dem-receptors linked to InsP3 turnover and Ca2/ mobilization onstrated ATP-triggered [Ca2/]i transients in oligodendro-from internal stores. Likewise, P2u receptors linked to cytes (defined by their morphology and ionic current pat-[Ca2/]i increase were found in enteric glial cells (226), tern) (237). The ATP-induced Ca2/ signaling in oligoden-which are similar to astrocytes with respect to their mor- drocytes in brain slices was not accompanied by anyphology and GFAP expression. Intracellular Ca2/ in- changes in membrane permeability, further supporting the
suggestion of an intracellular origin.creases and [Ca2/]i oscillations mediated by P2u receptors
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
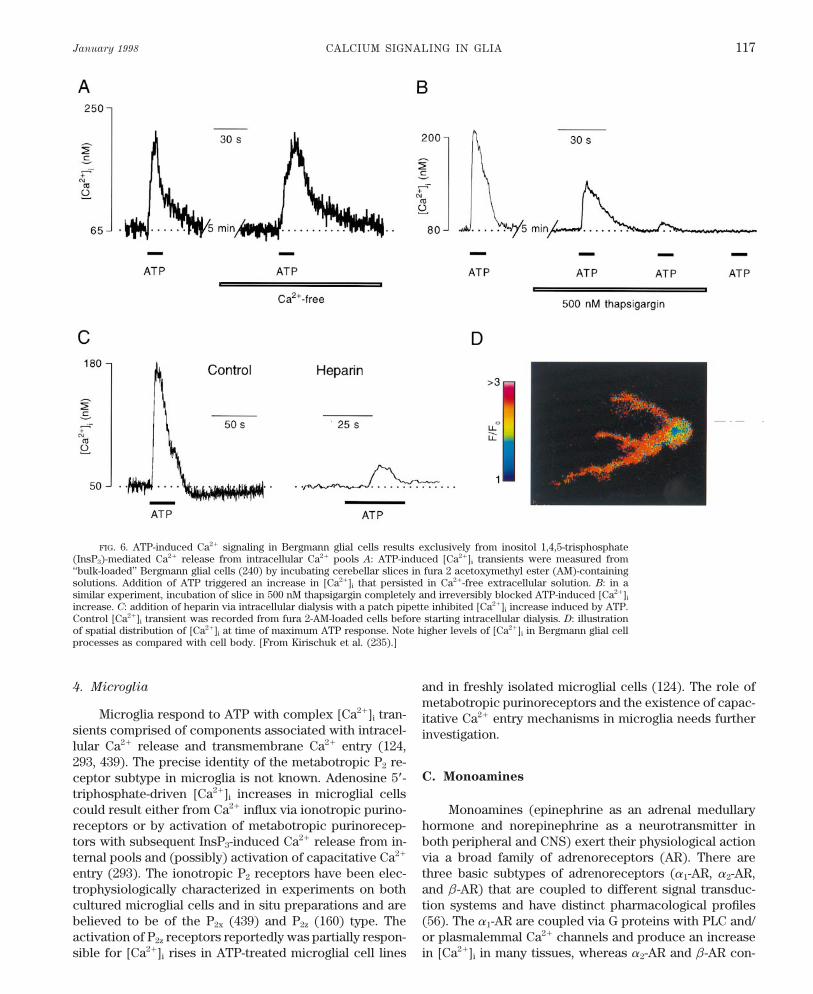
January 1998 CALCIUM SIGNALING IN GLIA 117
FIG. 6. ATP-induced Ca2/ signaling in Bergmann glial cells results exclusively from inositol 1,4,5-trisphosphate(InsP3)-mediated Ca2/ release from intracellular Ca2/ pools A: ATP-induced [Ca2/]i transients were measured from‘‘bulk-loaded’’ Bergmann glial cells (240) by incubating cerebellar slices in fura 2 acetoxymethyl ester (AM)-containingsolutions. Addition of ATP triggered an increase in [Ca2/]i that persisted in Ca2/-free extracellular solution. B: in asimilar experiment, incubation of slice in 500 nM thapsigargin completely and irreversibly blocked ATP-induced [Ca2/]i
increase. C: addition of heparin via intracellular dialysis with a patch pipette inhibited [Ca2/]i increase induced by ATP.Control [Ca2/]i transient was recorded from fura 2-AM-loaded cells before starting intracellular dialysis. D: illustrationof spatial distribution of [Ca2/]i at time of maximum ATP response. Note higher levels of [Ca2/]i in Bergmann glial cellprocesses as compared with cell body. [From Kirischuk et al. (235).]
4. Microglia and in freshly isolated microglial cells (124). The role ofmetabotropic purinoreceptors and the existence of capac-
Microglia respond to ATP with complex [Ca2/]i tran- itative Ca2/ entry mechanisms in microglia needs furthersients comprised of components associated with intracel- investigation.lular Ca2/ release and transmembrane Ca2/ entry (124,293, 439). The precise identity of the metabotropic P2 re-
C. Monoaminesceptor subtype in microglia is not known. Adenosine 5*-triphosphate-driven [Ca2/]i increases in microglial cellscould result either from Ca2/ influx via ionotropic purino- Monoamines (epinephrine as an adrenal medullary
hormone and norepinephrine as a neurotransmitter inreceptors or by activation of metabotropic purinorecep-tors with subsequent InsP3-induced Ca2/ release from in- both peripheral and CNS) exert their physiological action
via a broad family of adrenoreceptors (AR). There areternal pools and (possibly) activation of capacitative Ca2/
entry (293). The ionotropic P2 receptors have been elec- three basic subtypes of adrenoreceptors (a1-AR, a2-AR,and b-AR) that are coupled to different signal transduc-trophysiologically characterized in experiments on both
cultured microglial cells and in situ preparations and are tion systems and have distinct pharmacological profiles(56). The a1-AR are coupled via G proteins with PLC and/believed to be of the P2x (439) and P2z (160) type. The
activation of P2z receptors reportedly was partially respon- or plasmalemmal Ca2/ channels and produce an increasein [Ca2/]i in many tissues, whereas a2-AR and b-AR con-sible for [Ca2/]i rises in ATP-treated microglial cell lines
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78118
trol the activity of adenylate cyclase. In glial cells, the prepared hippocampal stratum radiatum slices (104) andin Bergmann glial cells studied in isolated cerebellar slicesexpression of adrenoreceptors and monoamine-triggered(239). In both studies, norepinephrine- and epinephrine-Ca2/ signaling has been found exclusively in astrocytes.evoked [Ca2/]i responses exhibited a1-AR pharmacologyAstrocytes express b-AR (184, 276) and a1-AR (256,(the effects were mimicked by a1-AR agonist PE, blocked383) both in culture and in situ (see also Ref. 284 forby a1-AR antagonist prazozin, and insensitive to a2-AR-review of AR in astrocytes). In cultured astrocytes, thespecific agents). The a1-AR-mediated [Ca2/]i responsesexpression of b-AR was only found in type 1-like cellswere characterized by a long-lasting plateau phase that(51); a1-AR are abundant in all types of astrocytes. Stimu-followed the initial [Ca2/]i rise. Removal of [Ca2/]o elimi-lation of a1-AR with norepinephrine increases intracellu-nated the plateau phase in Bergmann glia but not in hippo-lar levels of InsP3 (337, 339). Microfluorimetric [Ca2/]i re-campal astrocytes.cording techniques revealed that norepinephrine and
other monoamines increased cytoplasmic Ca2/ in culturedastrocytes (49, 193, 285, 317, 373). The pharmacological
D. g-Aminobutyric Acid and Glycineprofile of the monoamine-induced [Ca2/]i transients dem-onstrated a leading role for the a1-AR receptor, although
g-Aminobutyric acid (GABA) effects in the nervousseveral reports point out that activation of a2-AR can in-system are mediated by three distinct subtypes of recep-crease [Ca2/]i (285, 315, 461), presumably involving PLC-tors: GABAA and GABAC are intrinsic ligand-gated Cl0driven production of InsP3 (as revealed by the inhibitionchannels, whereas GABAB receptors are coupled to theirof a2-AR-stimulated [Ca2/]i elevation with the specific PLCeffectors (K/- or Ca2/-permeable plasmalemmal channels)blocker U-73122; Ref. 114). The density of a2-AR linkedvia G proteins (206, 308, 386). In contrast to neurons,to the generation of cytoplasmic Ca2/ signals was signifi-where GABA normally leads to a hyperpolarization, incantly upregulated by incubation of cortical astrocytic cul-macroglial cells GABA depolarizes the membrane (132,tures with dibutyryl cAMP (114). The a1-AR-mediated221, 222, 435). The difference is not due to the receptor[Ca2/]i transients are characterized by complex kinetics,but rather due to the different intracellular Cl0 levels in
often consisting of an initial peak followed by a long-the two cell types. Glial cells have a more positive Cl0
lasting plateau or oscillations (e.g., Refs. 373, 450). Theequilibrium potential (ECl) than neurons due to a higher
norepinephrine-induced [Ca2/]i increase in culturedintracellular Cl0 concentration. This is primarily due to
mouse astrocytes was substantially inhibited by chronic the activity of two inwardly directed Cl0 transporters, atreatment (7–14 days) with 1 mM lithium (63). The latter Na/-K/-2Cl0 cotransporter and a Cl0/HCO0
3 exchanger.is known to interfere with InsP3 signaling pathway and The Cl0 reversal potential in glial cells is about 035 mV,has therapeutic potential as a mood-stabilizing drug. Thus and opening Cl0 channels led to a depolarizing Cl0 effluxit is not excluded that the psychological effects of lithium (132, 435). The GABA-activated inward currents were re-are due to modulating astrocytic rather than neuronal re- corded in cells from astrocyte and oligodendrocyte lin-ceptor. eage employing both single cell (45, 131, 221) and brain
Shao and McCarthy (379, 382) determined the rela- slice models (32, 398).tion between the amplitude of the [Ca2/]i increase in corti- In many cases, GABA-induced depolarization of glialcal astrocytes and agonist concentration (379, 382). The cells exceeds the threshold for voltage-gated Ca2/ chan-threshold phenylephrine (PE) concentration was Ç1 mM, nels and thereby produces Ca2/ influx and measurablebut PE concentrations exceeding 5 mM triggered a maxi- [Ca2/]i transients. Such effects have been recorded frommal response. Moreover, using subsequent monitoring of cultured (314) and freshly isolated (131) astrocytes asa1-AR-mediated [Ca2/]i increase and the a1-AR receptors well as cultured cells of the oligodendrocyte lineage (230).density in the same cell, they (379) found that the ampli- Experiments with freshly isolated astrocytes and oligo-tude of [Ca2/]i response was independent of the density dendrocytes in culture demonstrated that GABA-inducedof a1-AR (despite that the latter varied between 10 and [Ca2/]i increases were exclusively due to Ca2/ entry via2,000 binding sites/1,000 mm2 membrane area). They sug- voltage-gated channels, since removal of [Ca2/]o or Ca2/
gested that a1-AR-mediated InsP3 formation triggers all-or- channel blockers inhibited the [Ca2/]i rise (131, 230). Innothing Ca2/ release from internal stores. The underlying experiments on cultured astrocytes, Nilson et al. (314)mechanism is unclear but may involve facilitation of suggested that in addition to plasmalemmal Ca2/ entry,InsP3-induced release by increased [Ca2/]i . In contrast, GABA triggered a release of Ca2/ from intracellular stores,experiments on cerebellar Bergmann glial cells (239) re- suggesting a possible involvement of GABAB receptors.vealed a flatter dependence of [Ca2/]i increase on agonist Another piece of evidence suggesting the expression ofconcentration, suggesting a gradual InsP3-triggered Ca2/ GABAB receptors coupled to Ca2/ entry pathway camerelease in these cells. from 45Ca2/ flux measurements (3), although this observa-
Recently, the expression of functional a1-AR linked tion has not been substantiated yet with physiologicaltechniques.to [Ca2/]i was found in situ in astrocytes from acutely
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 119
Some glial cells have been reported to express gly- lowed the activation of muscarinic receptors in oligoden-drocyte precursors (79).cine receptors that also increase Cl0 permeability (231).
Activation of these receptors causes depolarization of oli- Pharmacological assays indicate that astrocytes alsogodendrocytes and might also increase [Ca2/]i , but this have both M1 and M2 receptors (8, 302) and that M1 recep-has not yet been shown. tors are primarily responsible for InsP3 production (8). In
addition, in astrocytes from neonatal rat cerebral cortex,M3 receptors coupled to phosphoinisitide hydrolysis were
E. Acetylcholine found (243). The expression of functional MChRs recep-tors varies considerably between astrocytes isolated from
Acetylcholine reacts with two distinct families of cho- different brain regions; the carbachol-stimulated InsP3
linoreceptors, nicotinic cholinoreceptors (NChRs) that production mediated by M1 receptors was much higher incontain an integral cationic channel and metabotropic astrocytes derived from mesencephalon than in cells frommuscarinic cholinoreceptors (MChRs) that are coupled to the medulla-pons and cerebral hemispheres, being an-PLC and adenylate cyclase by means of G proteins (58, other example of functional heterogeneity among astro-122). The NChRs are assembled from a broad family of cytes (8).elementary subunits belonging to a distinct gene family;some NChR heteromeres have relatively high Ca2/ perme-ability (408, 433). There are five major subtypes of MChRs
F. Histamine(M1-M5) that have been identified and cloned (41). Theyhave the seven membrane spanning domain structure typi-cal for metabotropic receptors. Type 1, 3, and 5 MChRs
Histamine affects three types of receptors: H1 (cou-are coupled with PLC, thereby controlling InsP3 turnover,pled with PLC), H2 (positively connected with adenylateand MChRs types 2 and 4 decrease adenylate cyclase ac-cyclase), and H3 receptors that control histamine turnovertivity.and release (257). Histaminergic nerve terminals in theIn glial cells, NChRs have only been described inposterior hypothalamus release the transmitter nonsynap-invertebrates. Stimulation of NChRs in leech neuropiletically directly into the interstitial space (196). It is thusglial cells causes depolarization (18, 376) and increasedlikely that adjacent glial cells equipped with the proper[Ca2/]i due to both Ca2/ entry via NChRs and voltage-receptor are activated. Indeed, astrocytes as well as astro-activated plasmalemmal channels (376). In insects, therecytoma cell lines have both H1 and H2 receptors (187, 195,is evidence of NChRs from immunochemical studies196), and stimulation of H1 receptors increased intracellu-(255).lar InsP3 in these cells (10, 244, 305). Measurements ofMuscarinic receptors are widely distributed in mam-[Ca2/]i revealed intracellular Ca2/ transients in both astro-malian macroglial cells. Initially, MChRs were detected incytoma cell lines (286, 456) and primary cultured astro-both astrocytes (185, 366) and oligodendrocytes (366) bycytes (136, 193, 194, 285) mediated via H1 histamine recep-a variety of histochemical and immunocytochemicaltors. Experiments with cultured cortical astrocytes indi-methods. Stimulation of MChRs leads to accumulation ofcated that H1 receptors and histamine-evoked [Ca2/]iInsP3 in glial cell lines (250, 306) and macroglial cellsresponses were mainly confined to type 2 astrocytes and(astrocytes, Refs. 302, 337, 339; oligodendrocytes, Refs.a subpopulation of type 1 astrocytes (136, 244). The ex-78, 366). Thus InsP3-mediated [Ca2/]i increases were re-pression of H1 receptors coupled to [Ca2/]i was recentlycorded from many types of glial cells treated with MChRconfirmed in experiments in situ in cerebellar Bergmannagonists. This included astrocytoma cells (286, 320, 327),glial cells (239). In these cells, histamine induced [Ca2/]iperisynaptic Schwann cells at the frog neuromusculartransients in a dose-dependent manner (thresholdÇ1 mM;junction (200), primary cultured astrocytes (92, 382), andand maximal response at Ç100 mM). The histamine-in-cultured oligodendrocytes (78, 213). In the latter, PCRduced [Ca2/]i transients in Bergmann glial cells were sen-was used to demonstrate the expression of both M1 and M2
sitive to the H1 specific antagonist chlorpheniramine butMChRs. However, the carbachol-induced [Ca2/]i increasewere not mimicked by H2 and H3 agonists (dimarit and a-was preferentially sensitive to the M1 agonist pirenzipine,methylhistamine, respectively). Similarly, H2 and H3 antag-rather than the M2 antagonist methoctramin. This suggestsonists (ranitidine and thioperamide) had no effect. Pre-that InsP3 formation and Ca2/ mobilization in oligoden-treatment of Bergmann glial cells with thapsigargin abol-drocytes involves M1 muscarinic receptors (78). Interest-ished the histamine-induced [Ca2/]i response, suggestingingly, MChR-mediated [Ca2/]i responses in cells of thethat intracellular Ca2/ release was responsible for the H1oligodendrocyte lineage have two components, an initialreceptor-sensitive [Ca2/]i rise.rise associated with intracellular Ca2/ release and a long-
In cells of the oligodendrocyte lineage, histamine in-lasting plateau resulting from Ca2/ entry (78) via an un-creased [Ca2/]i in a subpopulation of cultured progenitorsknown mechanism. Both Ca2/ release and Ca2/ entry are
important for stimulation of c-fos expression, which fol- and mature oligodendrocytes (213).
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78120
G. Substance P population (267). The BK-triggered [Ca2/]i elevation in cul-tured astroglia was mediated mainly by B2 receptors asjudged by its pharmacological profile, although a minorSubstance P belongs to a family of neuropeptides,subpopulation of astrocytes may also contain B1 receptorsthe neurokinins, that affects neurons by stimulating neu-(152, 399). The relative importance of intracellular Ca2/rokinin receptors widely distributed in the brain (275).release/Ca2/ entry components in BK-induced [Ca2/]i ele-These receptors are represented by three subtypes (NK1-vation remains unclear. In one series of experiments onNK3) identified by different molecular structures and acultured astrocytes, BK-triggered [Ca2/]i transients con-distinct pharmacological profile (89, 384, 458). Amongsist of an initial Ca2/ release from ER stores followed bythem, NK1 receptors preferably bind substance P. All threelong-lasting Ca2/ entry component (152). Another studyNK receptors are typical G protein-coupled metabotropicwith the same type of cells found only the Ca2/ releasereceptors with a seven transmembrane domain structure.component (399). Apart from an [Ca2/]i rise, an inwardInteraction of substance P with NK1 receptors results incurrent was observed in cultured astrocytes upon BK ap-phosphoinositide hydrolysis (278, 279) in oligodendro-plication (152). The BK-dependent membrane current wascytes and type 2 astrocytes. Substance P and NK1 agonistsalso sensitive to B2 receptor antagonists. However, a clearincrease [Ca2/]i by releasing Ca2/ from thapsigargin-sensi-disparity in the concentration dependence betweentive stores in astrocytoma cell lines (43, 110, 353) in hu-changes in [Ca2/]i and membrane conductance was found.man glioma line (173), in primary cultured astrocytes (193,The 50% effective concentration (EC50) for [Ca2/]i rise was278, 281), and in cultured cells of oligodendrocyte lineageÇ10 nM BK, whereas EC50 for BK-induced currents was(172, 213). In astroglia, substance P-dependent InsP3 pro-Ç1 mM. These results suggest that distinct concentration-duction and subsequent intracellular Ca2/ release was re-dependent signaling pathways aimed at different targetsstricted to type 2 astrocytes (278). The NK1-mediatedare triggered by activation of B2 receptors on cultured[Ca2/]i responses, found in experiments with both glialastrocytes. Bradykinin-induced Ca2/ signaling was ob-cells lines (43) and primary cultured oligodendrocytesserved in Ç50–60% of cultured astrocytes and was a sub-(172), were relatively short and not affected by extracellu-ject of developmental regulation in oligodendroglia, i.e.,lar Ca2/ removal. This suggests that the Ca2/ is releasedBK triggered [Ca2/]i elevation inÇ50% of mature oligoden-from intracellular stores. Substance P-induced Ca2/ sig-drocytes compared with only 13% of oligodendrocyte pre-nals stimulate interleukin-1 production in cultured astro-cursors (213).cytes (281). The activation of NK1 receptors is accompa-
nied by astrocyte depolarization (444) due to the closureof K/ channels (15). What is the mechanism of K/ chan-
I. Endothelinsnels inhibition upon substance P stimulation remains un-clear; it could be mediated by an [Ca2/]i increase. This
Endothelins, represented by three major isoforms en-depolarization, in turn, could activate additional Ca2/ in-coded by distinct genes (ET-1, -2, -3; Ref. 197), were firstflux through voltage-gated channels.found to be potent vasoactive compounds (455). Later,expression of both endothelin mRNAs and a peptide
H. Bradykinin proper, as well as endothelin binding sites, were found tobe widely distributed in the brain (148, 208), and physio-logical responses were detected in neurons challengedTwo types of bradykinin (BK) receptors (B1 and B2)
have been described (161). Both subtypes belong to the with endothelins (70, 299). Endothelin-induced cellularreactions are mediated via three major receptor subtypesG protein-coupled metabotropic receptors linked to PLC
and InsP3 production. Bradykinin acts not only as an effec- (ETRs) bearing different sensitivity to endothelin iso-forms. The most abundant form in the brain is the nonse-tive vasodilatator but also as a neurotransmitter. Bradyki-
nin and its binding sites were found in the mammalian lective ETBR (which is equally sensitive to all 3 endothe-lins). The ETAR (preferably sensitive to ET-1 and ET-2,brain (135, 344), and various neurons express functional
BK receptors. Initial evidence for the presence of BK re- but not ET-3) is much less expressed. The ETCRs havebeen described only in nonmammalian tissues (26, 372).ceptors in glial cells came from biochemical experiments
that identified BK binding sites on cultured astrocytes (68) All endothelin receptors have an overall topology similarto G protein-coupled seven transmembrane domain me-and demonstrated stimulation of phopshoinisitide turn-
over in BK-treated cultured astrocytes (69, 278, 366) and tabotropic receptors, and their activation involves variousG proteins (either PTX sensitive or insensitive). The ETRsoligodendrocytes (278, 400). In physiological experiments,
using microfluorimetric [Ca2/]i recordings, BK was found are significantly involved in regulation in [Ca2/]i . Theytrigger the PLC-InsP3-Ca2/ release transduction chain asto increase [Ca2/]i in cultured Schwann cells (330) and in
cultured astrocytes (152, 399) and oligodendrocytes (213, well as stimulate transmembrane Ca2/ influx (372).Initially, endothelin receptors were found with auto-278). However, in freshly isolated Schwann cells, a BK-
induced [Ca2/]i rise was observed only in a minor cell radiography in various subpopulations of brain astrocytes
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 121
(186). Later, astrocytes were shown to express ETA and of endothelin-induced Ca2/ responses in Bergmann glialcells in situ (418). In the same cells, endothelin-triggeredETB receptors (111, 123, 162). Furthermore, in certain
brain regions, expression of endothelin-binding sites was [Ca2/]i transients exhibited a progressive rundown withrepeated endothelin applications (418). This could be thefound to be glial specific. The endothelin binding in the
cerebellum was quite similar in normal mice and in mu- result of the internalization of the ETRs (364).Endothelin-induced [Ca2/]i signaling might partici-tants lacking Purkinje neurons (pcd mutants), suggesting
that glial cells carry the majority of ETRs (270). Stimula- pate in the astroglial proliferative response. Increased lev-els of endothelins in brain tissue accompany various in-tion with endothelin led to an increase of InsP3 in astro-
cytes and astroglioma cells (70, 86, 263, 270, 459) and to sults in the CNS (22, 153). This observation suggests thehypothesis that ETRs are involved in the transduction ofincreases in [Ca2/]i in both glial cell lines (263, 456, 459)
and cultured astrocytes (155, 183, 280, 396, 404); recently, a mitogenic signal which, at least partially, underlies theactivation of astrocytes involved in brain pathology. Isch-ET-1-induced [Ca2/]i transients were also detected in mi-
croglial cells (293). The glial [Ca2/]i response to endothe- emic conditions are also accompanied by an increase inendothelin synthesis (22), and endothelin antagonists arelins was found to be insensitive to PTX (183, 396). The
endothelin-mediated Ca2/ mobilization in glioma cells was neuroprotective during brain ischemia (125). It has beendemonstrated that endothelin-induced [Ca2/]i increasesinhibited by opioid agonists presumably because they in-
hibit InsP3 production (20). The expression of ETRs linked play a pivotal role in stimulation of DNA synthesis andproliferative response of cultured type 1 astrocytes (396,to [Ca2/]i increases appears to be region specific, i.e., in
cortical type 1 astrocytes, endothelin effects on [Ca2/]i 404). In addition, endothelins were found to trigger imme-diate early genes expression; the level of Fos proteinswere mediated via ETAR (183), whereas cerebellar astro-
cytes predominantly expressed ETBR (396). The increases significantly in endothelin-challenged glial cellsfrom organotypically cultured cerebellar slices (403). Thisastrocytic expression of ETRs linked to [Ca2/]i was re-
cently demonstrated in experiments in situ on Bergmann suggests that ET receptors can play a role in developmentor during pathological events when glial cells undergoglial cells, in which endothelin produced an elevation in
[Ca2/]i . The Ca2/ responses in Bergmann glial cells were phases of proliferation. In this context, the glial endo-thelin system has been implicated in a number of patho-mediated by ETBR (418) as was revealed by singe-cell RT-
PCR analysis. physiological situations such as viral infections (269), Alz-heimer’s disease (460), and ischemia (454).The endothelin-induced [Ca2/]i transients in both cul-
tured and in situ astrocytes had heterogeneous kineticproperties varying from a simple peak response to a bi-
J. Other Agonists Linked to Intracellular Ca2/phasic [Ca2/]i rise, with a plateau lasting up to 20 minRegulation in Glial Cellsafter agonist washout (155, 183). Sometimes endothelin
triggered only a plateau without a defined peak (183). Asin the case of other agonist-triggered [Ca2/]i responses, 1. Serotoninthe initial peak resulted from internal Ca2/ release, while
Serotonin is a neurotransmitter in a number of brainthe plateau phase was entirely dependent on extracellularregions. It exerts its action via a broad spectrum of recep-Ca2/ (183, 263, 396, 404, 419). The nature of Ca2/ deliverytors that have been classified into four subfamilies (5-HT1pathway underlying the plateau phase of endothelin-in-to 5-HT4). Both 5-HT1 and 5-HT4 are coupled to adenylateduced responses remains controversial. Studying endo-cyclase, whereas 5-HT3 receptors are ligand-operated cat-thelin responses in cerebellar cultured glial cells, oneionic channels and 5-HT2 receptors are positively coupledstudy found it was completely (404) or partially (396)with PLC, thereby regulating InsP3 production (189, 462).inhibited by nifedipine, suggesting an important role forSerotonin activates phosphoinositide turnover in glial cellvoltage-gated Ca2/ channels, whereas in another study,lines (7) and raises [Ca2/]i in astrocytomas (325, 402) andnifedipine had only a marginal effect (183). The effects ofsubpopulations of primary cultured astrocytes (94, 193,ETR stimulation on Ca2/ channels might be mediated by316, 425). Type 1 astrocytes are more sensitive to seroto-diacylglycerol, since treating cultured astrocytes with anin (285) than type 2 astrocytes (92). Cultured astrocytesprotein kinase inhibitor causes a nifedipine-sensitive pla-challenged with serotonin display kinetically different re-teau (183). Diacylglycerol might induce phosphorylationsponses varying from single spikes to prolonged oscilla-of Ca2/ channels and shift their threshold toward moretions (316). In Schwann cells, results on the appearancenegative membrane potentials. The participation of Ca2/
of 5-HT receptors linked to [Ca2/]i are controversial. In achannels in the response may vary considerably from cellcomprehensive study of neuroactive substances, Lyons etto cell. A second possible mechanism for producing aal. (267) failed to detect an effect of serotonin on [Ca2/]iprominent plateau phase in endothelin-induced [Ca2/]i
in cultured Schwann cells. Another study of culturedtransients might be either endothelin-activated plas-Schwann cells (457) found a consistent increase in [Ca2/]imalemmal channels or capacitative Ca2/ entry. The capac-
itative Ca2/ entry pathway is likely to shape the plateau mediated by stimulation 5-HT2A receptors. In the latter
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78122
case, the Schwann cells were maintained in 4 mM and appeared to result entirely from Ca2/ influx via volt-age-gated channels, since either removal of Ca2/ from theforskolin; such treatment could promote the expression
of serotonin receptors. In cultured astrocytes, serotonin bath or the addition of Cd2/ (2 mM) prevented the [Ca2/]i
responses to NPY. The channels were activated by NPY-effects were mediated by 5-HT2 receptors, as detected byradioligand binding, Northern blots, and pharmacological triggered cell depolarization resulting from an inward cur-
rent in response to NPY application. This current resultedassays (94). Further studies revealed the pharmacologicalprofile of these receptors corresponds to the 5-HT2C sub- from the closing of K/ channels. The NPY-dependent mod-
ulation of K/ channels was PTX insensitive (151).type (64).
4. Complement fragments2. Oxytocin and vasopressin
Complement fragments (anaphylatoxins) are gener-Receptors for the closely related neurohormones ated during the activation of the complement system and
oxytocin and vasopressin have been found in several play an important role in various immunological reactionstypes of astroglial cells. Oxytocin caused an increase in (157). Recently, it was discovered that complement frag-InsP3 level in cultured astrocytes from cortex and cerebel- ments can be directly produced in the brain (21). Thelum but not from spinal cord (67). The oxytocin binding action of anaphylatoxins is achieved via activation of spe-sites were initially described in cultured hypothalamic as- cific receptors belonging to the metabotropic family andtrocytes (100, 101), and later, functional oxytocin recep- have a characteristic seven transmembrane structuretors linked to intracellular Ca2/ signaling were found in (363, 365). They are believed to regulate PLC-controlledthe same cells (99). Oxytocin-triggered [Ca2/]i transients InsP3 accumulation and initiate Ca2/ release from intracel-were observed in a majority of hypothalamic astrocytes. lular stores (294, 319). Initially, expression of complementThe responses usually had a monophasic peak with a fast fragments receptors was thought to be a perogative ofdecay to the initial [Ca2/]i level. A minority of cells (15%) leukocytes. However, functional anaphylatoxin receptorsunderwent [Ca2/]i oscillations. The transients remained were found in other cells including neuroglia. The recep-in Ca2/-deficient extracellular media, suggesting they orig- tors for complement fragment C5a were found in bothinated from intracellular Ca2/ release. They exhibited a primary cultured human fetal astrocytes and in a humanprominent rundown in response to successive oxytocin astrocyte cell line. Stimulation of these receptors raisedapplications apparently due to receptor desensitization [Ca2/]i in cultured astrocytes (144). Similarly, complementrather than depletion of stores (99). fragments C5a and C3a triggered a transient [Ca2/]i rise
The addition of vasopressin to cultured cortical and in both cultured microglial cells and microglial cells oncerebellar astrocytes also increased intracellular InsP3 the surface of acutely isolated corpus callosum slices(67). Subpopulations of cultured type 2 astrocytes (193), (292). The anaphylatoxin-induced [Ca2/]i transients re-astrocytes derived from rat neural lobes (168) and circum- corded in microglial cells result from Ca2/ release fromventricular organs (210), also responded to vasopressin intracellular stores followed by a capacitative Ca2/ influxwith an increase in [Ca2/]i . Naturally occurring fragments (292).of vasopressin increased [Ca2/]i in cultured astrocytesfrom the circumventricular organ (211). The effects of 5. Platelet-activating factorvasopressin on [Ca2/]i in astrocytes were mediated by V1
Platelet-activating factor (PAF) is known to substan-vasopressin receptors, as suggested by their pharmacolog-tially modulate neuronal functions by exerting a pleiotro-ical profile (210, 211). Vasopressin-triggered [Ca2/]i tran-pic effect and modulating synaptic transmission (27, 215).sients were maintained in Ca2/-free solutions, indicatingIts effect is mediated via stimulation of specific PAF re-that they resulted (as in the case of oxytocin) from aceptors (PAFR; Ref. 38), which control phosphoinositiderelease of Ca2/ from intracellular stores (210).turnover and induce Ca2/ release from internal stores invarious types of cells, including neurons (39, 445). The
3. Neuropeptide Y PAFRs have been also found in C6 glioma cells, primarycultured astrocytes, and oligodendrocytes (48). In cul-Neuropeptide Y (NPY), a member of the pancreatictured astrocytes, PAF was reported to increase cyto-polypeptide family (which includes also pancreatic poly-plasmic InsP3 (346). High expression of PAFR and PAF-peptide and peptide YY), was found to be expressed ininduced [Ca2/]i elevation (due to both internal Ca2/ re-the CNS where it exerts various regulatory effects (seelease and Ca2/ influx) has also been detected in immortal-Refs. 80, 106, 443 for review). In astrocytes, the specificized (365) and cultured (293, 298) microglial cells.binding of 125I-NPY was detected, suggesting the existence
of specific receptors (151). Applications of NPY, in micro-6. Prostanoidsmolar concentrations, to primary cultured cortical astro-
cytes caused an increase in [Ca2/]i (151). This increase Prostanoid receptors constitute a large family (442);these receptors are coupled to cAMP turnover (DP, EP2,was observed in 20–70% of cells in different preparations
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 123
and IP receptors) and to PLC and InsP3 production (EP1, Other blood-derived proteins, albumin and thrombin, havebeen reported to affect [Ca2/]i in several types of cells ofEP3; FP and TP receptors). The expression of FP and
TP receptors linked to phosphoinositide hydrolysis was glial origin. Albumin was reported to have a rather un-usual effect on [Ca2/]i in cultured astrocytes, decreasingdemonstrated in human astrocytoma cell line (241), and
FP receptor-mediated Ca2/ increases were found in pri- [Ca2/]i at low concentrations (õ2 mg/ml) and initiating[Ca2/]i spikes at high concentrations (303, 304). The de-mary cultured type 1 astrocytes (198).crease in [Ca2/]i in the presence of low albumin concen-trations was accompanied by a significant rise in the am-7. Vasoactive intestinal polypeptideplitude of thapsigargin- and glutamate-triggered [Ca2/]i
Vasoactive intestinal polypeptide (VIP) in concentra- transients. This suggested that albumin promotes Ca2/
tions of 0.1–1 nM was found to increase [Ca2/]i in a sub- accumulation into the ER Ca2/ stores (303). As albuminpopulation of cultured cortical type 1 astrocytes (119). concentrations rise in the brain after injury, this effectSimultaneous administration of VIP and subthreshold would potentiate neurotransmitter-induced Ca2/ signalsconcentrations of norepinephrine greatly increased the in astrocytes by increasing the releasable Ca2/ contentpercentage of responding astrocytes and enhanced the in intracellular stores. Such a potentiation might also beVIP [Ca2/]i response. The VIP-triggered [Ca2/]i increases relevant for astrocyte responses that are an importantpersist in Ca2/-free external solutions and are blocked by defense component during brain insults. Another bloodthapsigargin, suggesting they result from intracellular factor, thrombin, was reported to cause [Ca2/]i oscilla-Ca2/ release. Another endogenous ligand for VIP recep- tions in astrocytoma cells in nanomolar concentrations.tors, pituitary adenylate cyclase-activating polyleptide, re- Similar oscillations were produced by thrombin receptor-leases Ca2/ from internal stores in type 2 but not in type activating peptide. This suggests the involvement of a spe-1 astrocytes (415). cific plasmalemmal thrombin receptor (87).
8. Platelet-derived growth factor11. Arachidonic acid
Examples of platelet-derived growth factors (PDGFs)Arachidonic acid, which serves as a biologically ac-
are three proteins that are potent mitogenes (174). Thetive signaling molecule, was reported to directly raise
PDGF receptors (a and b) have been found in oligoden-[Ca2/]i in cultured rat spinal cord oligodendrocytes (391).
drocytes and Schwann cells, and PDGF was shown toThis [Ca2/]i rise appeared to result from stimulation of a
promote proliferation of various types of glial cells (360).Ca2/ influx pathway.
Intracellular Ca2/ transients as well as [Ca2/]i oscillationshave been found in oligodendrocyte cell line (120) and in
12. Opioidsoligodendrocyte precursors (165) in response to PDGF.In the latter, PDGF-induced Ca2/ signaling may be in- Several groups have reported that activation of opioidvolved in the initiation of cell differentiation. receptors trigger increase in [Ca2/]i in cultured astrocytes.
The precise nature of receptors involved remains unclear.9. Angiotensin II In astrocytes cultures from newborn mice cerebrum (401)
and rat cortex (116), morphine produced [Ca2/]i increasesAngiotensin II-induced [Ca2/]i increases have beenvia activation of k-opioid receptors (as [Ca2/]i responsesdemonstrated in human astrocytoma cell lines (412) andwere mimicked by selective k-receptor agonist U-69593).in several types of primary cultured astrocytes (201, 441).In contrast, another group (170) reported that morphine’sIn astroglial cultures derived from the hypothalamus andeffect on [Ca2/]i in cultured mice brain astrocytes wasbrain stem of adult rat, angiotensin II triggered [Ca2/]imediated through m-opioid receptors. In the latter prepa-transients with a clear biphasic (peak followed with pla-ration, morphine-triggered [Ca2/]i transients were simu-teau) time course. The initial peak resulted from Ca2/
lated by m-receptor agonist PL-017, and furthermore, ex-release from the internal stores (as judged by its insensi-pression of m-receptors was substantiated by immuno-tivity to [Ca2/]o and inhibition by thapsigargin and cyclopi-staining with specific antibodies. The intracellularazonic acid), whereas the plateau component reflectedmechanism for morphine-induced [Ca2/]i mobilization isCa2/ entry via nifedipine and cadmium-sensitive plas-also unclear. Eriksson et al. (116) demonstrated that stim-malemmal Ca2/ channels (441). The effects of angiotensinulation of k-opioid receptors results in a stimulation ofon [Ca2/]i in astrocytes were mediated via AT1 receptorsCa2/ influx through L-type voltage-gated channels. In con-(441).trast, Hauser et al. (170) believe that morphine triggersCa2/ release from thapsigargin- and dantrolene-sensitive
10. Serum albumin and thrombinintracellular depots (170). Stimulation of opioid receptorsin astrocytes also affects DNA synthesis and astrocyticDialyzed serum was reported to trigger [Ca2/]i in-
creases in primary cultured cerebellar astrocytes (321). growth. The role of [Ca2/]i in mediating these effects re-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78124
mains obscure, although it might be associated with intra- 381). Moreover, the same heterogeneity persists withincellular Ca2/ release (170). clones of cultured astrocytes, suggesting that the receptor
pattern is not inherited from the parent astrocyte (380).In a series of elegant experiments, Shao and McCarthy13. Myelin
(380, 381) demonstrated that the [Ca2/]i responsivenessIt is of interest that myelin, the main product of differ-
to various neuroligands can differ between two sister cellsentiated oligodendrocytes, produces [Ca2/]i transients in
immediately after astrocyte division. Thus astrocytes areboth oligodendrocytes (297) and brain stem neurons (296)
not born with a fixed set of functional receptors but canin vitro. This is accompanied by a block of oligodendro-
vary the expression of a variety of neurotransmitter recep-cytic motility and collapse of neuronal growth cones. The
tors depending on factors in their environment.mechanisms for myelin-triggered Ca2/ signaling substan-Similarly, neuroligand [Ca2/]i responsiveness of cul-tially differed between neurons and oligodendrocytes. In
tured oligodendrocytes is heavily controlled by cultureneurons, myelin promoted Ca2/ influx via voltage-gatedconditions. Even more interesting, the expression of neu-channels, whereas in oligodendrocytes, most of the Ca2/
roligand receptors linked to the Ca2/ signaling appearedwas released from intracellular stores. Thus, with the useto be controlled by oligodendrocyte-neuronal contactsof Ca2/ signaling, oligodendrocytes could recognize con-(171). Preventing glial-neuronal contacts by transectiontact with other oligodendrocytes, although the details ofof neurites in oligodenroglial-DRG cocultures significantlythe myelin-triggered changes in [Ca2/]i remain unclear.reduced the number of oligodendrocytes sensitive to aThe myelin basic protein in micromolar concentrationsvariety of neuroligands (ATP, carbachol, and histamine;also triggered a prominent [Ca2/]i increase associatedresponsiveness to BK was not affected; Ref. 171). Neu-with Ca2/ influx (420); this [Ca2/]i increase was fatal forronal control of receptors expression in neighboring gliaoligodendrocytes.appeared to be sensitive to tetrodotoxin, suggesting theimportant role of neuronal activity.14. Benzodiazepine receptors
An important question is whether this plasticity isApart of acting on GABAA receptors, benzodiazepines present not only when astrocytes are in culture but also
exert their action through a not yet precisely character- in vivo. Recently, this question has been investigated inized set of binding sites found in a variety of mammalian studies of neurotransmitter-induced Ca2/ signaling in glialcells and in glial cells in particular (447). Stimulation of cells in situ, in brain slices. Glial cells (both oligodendro-cultured rat astrocytes with endogenous benzodiazepine cytes and astrocytes) in corpus callosum slices demon-receptor ligand octadecaneuropeptide in nanomolar (0.1– strated prominent developmental changes in their neuro-10 nM) concentration triggered [Ca2/]i transients (251). transmitter receptor expression (33). In slices obtainedThis Ca2/ elevation was independent of extracellular Ca2/, from young (3–7 days old) mice, glial cells were ratherblocked by thapsigargin or by incubation with PTX for promiscuous, responding with [Ca2/]i increases to ATP,4 h. These data obviously suggest that benzodiazepine glutamate, histamine, GABA, norepinephrine, serotonin,receptors in astrocytes are coupled with the generation angiotensin II, BK, and substance P. In contrast, in olderof Ca2/ release from, presumably, InsP3-sensitive stores. (11–18 days old) animals, the expression of receptors was
limited to glutamate, ATP, and norepinephrine. Anothercomprehensive study of Ca2/ signaling-linked neurotrans-K. Heterogeneity of Neurotransmitter Receptormitter expression has been carried out in cerebellar Berg-Expression in Glial Cellsmann glial cells. These cells provide a unique model forstudying glial cells in slices because their characteristicThe data on the presence of neurotransmitter recep-morphology makes identification easy. The study foundtors presented above clearly demonstrate that glial cellsthat, contrary to cultured astrocytes, Bergmann glia al-can express functional receptors to almost all known neu-ways express a distinct set of receptors. These includerotransmitters, neuromodulators, and neurohormonesa1-adrenoreceptors, AMPA/kainate GluRs, mGluRs, H1(Table 2). The important question is whether all thesehistamine receptors, P2y purinoreceptors, and ETB endo-receptors are expressed in situ and which factors regulatethelin receptors (232, 235, 239, 419, 429). Other substancestheir expression. Most of the experiments on glial recep-known to induce Ca2/ signals in cultured macroglia weretors have been carried out in tissue-culture systems; ourineffective. Interestingly, the Purkinje cell layer, whereknowledge of the receptor pattern expressed by glial cellsBergmann glial cells are located, receives afferents (323)in vivo is far more limited. Glial cells in culture expressusing the following neurotransmitters: glutamate (parallela remarkable heterogeneity in their sensitivity to variousfibres), norepinephrine and ATP (terminals from locusneuroligands. Cultured astrocytes can respond at theceruleus), and histamine (histaminergic terminals fromsame time to a number of ligands with an increase intuberomammillary nucleus of posterior hypothalamus).[Ca2/]i . The pattern of receptor expression can differ sub-
stantially between cells in the same culture (92, 285, 380, Thus Bergmann glial cells express receptors that are ap-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
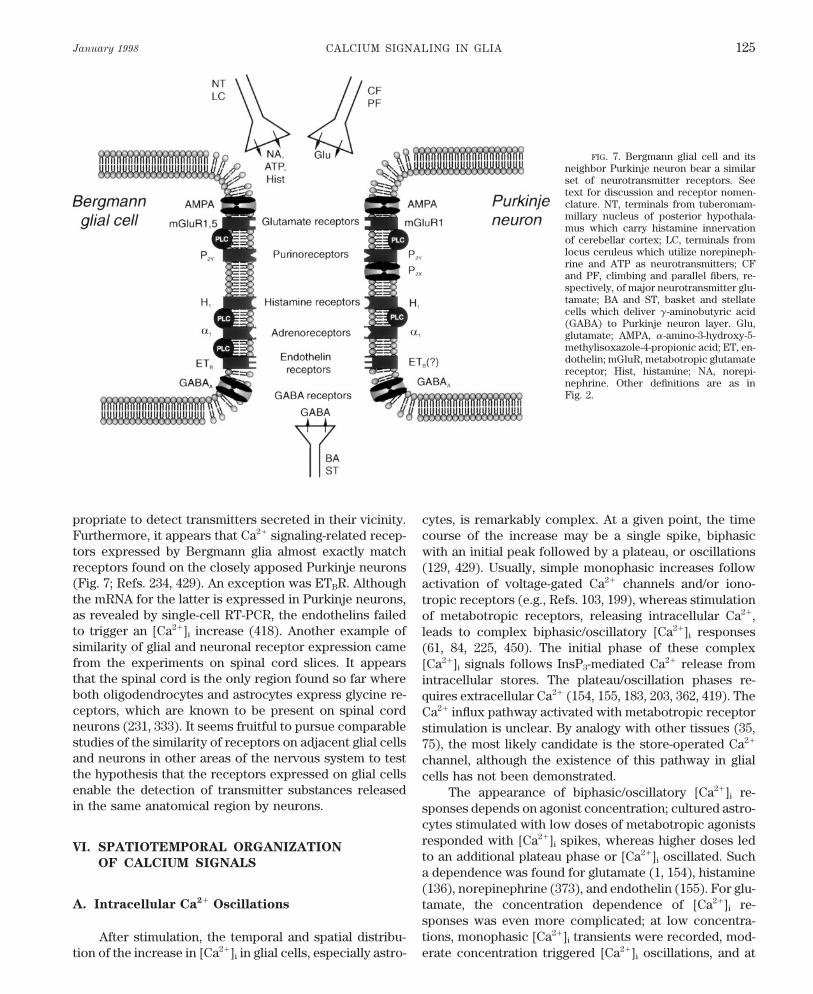
January 1998 CALCIUM SIGNALING IN GLIA 125
FIG. 7. Bergmann glial cell and itsneighbor Purkinje neuron bear a similarset of neurotransmitter receptors. Seetext for discussion and receptor nomen-clature. NT, terminals from tuberomam-millary nucleus of posterior hypothala-mus which carry histamine innervationof cerebellar cortex; LC, terminals fromlocus ceruleus which utilize norepineph-rine and ATP as neurotransmitters; CFand PF, climbing and parallel fibers, re-spectively, of major neurotransmitter glu-tamate; BA and ST, basket and stellatecells which deliver g-aminobutyric acid(GABA) to Purkinje neuron layer. Glu,glutamate; AMPA, a-amino-3-hydroxy-5-methylisoxazole-4-propionic acid; ET, en-dothelin; mGluR, metabotropic glutamatereceptor; Hist, histamine; NA, norepi-nephrine. Other definitions are as inFig. 2.
propriate to detect transmitters secreted in their vicinity. cytes, is remarkably complex. At a given point, the timeFurthermore, it appears that Ca2/ signaling-related recep- course of the increase may be a single spike, biphasictors expressed by Bergmann glia almost exactly match with an initial peak followed by a plateau, or oscillationsreceptors found on the closely apposed Purkinje neurons (129, 429). Usually, simple monophasic increases follow(Fig. 7; Refs. 234, 429). An exception was ETBR. Although activation of voltage-gated Ca2/ channels and/or iono-the mRNA for the latter is expressed in Purkinje neurons, tropic receptors (e.g., Refs. 103, 199), whereas stimulationas revealed by single-cell RT-PCR, the endothelins failed of metabotropic receptors, releasing intracellular Ca2/,to trigger an [Ca2/]i increase (418). Another example of leads to complex biphasic/oscillatory [Ca2/]i responsessimilarity of glial and neuronal receptor expression came (61, 84, 225, 450). The initial phase of these complexfrom the experiments on spinal cord slices. It appears [Ca2/]i signals follows InsP3-mediated Ca2/ release fromthat the spinal cord is the only region found so far where intracellular stores. The plateau/oscillation phases re-both oligodendrocytes and astrocytes express glycine re- quires extracellular Ca2/ (154, 155, 183, 203, 362, 419). Theceptors, which are known to be present on spinal cord Ca2/ influx pathway activated with metabotropic receptorneurons (231, 333). It seems fruitful to pursue comparable stimulation is unclear. By analogy with other tissues (35,studies of the similarity of receptors on adjacent glial cells 75), the most likely candidate is the store-operated Ca2/
and neurons in other areas of the nervous system to test channel, although the existence of this pathway in glialthe hypothesis that the receptors expressed on glial cells cells has not been demonstrated.enable the detection of transmitter substances released The appearance of biphasic/oscillatory [Ca2/]i re-in the same anatomical region by neurons. sponses depends on agonist concentration; cultured astro-
cytes stimulated with low doses of metabotropic agonistsresponded with [Ca2/]i spikes, whereas higher doses led
VI. SPATIOTEMPORAL ORGANIZATIONto an additional plateau phase or [Ca2/]i oscillated. Such
OF CALCIUM SIGNALSa dependence was found for glutamate (1, 154), histamine(136), norepinephrine (373), and endothelin (155). For glu-
A. Intracellular Ca2/ Oscillations tamate, the concentration dependence of [Ca2/]i re-sponses was even more complicated; at low concentra-tions, monophasic [Ca2/]i transients were recorded, mod-After stimulation, the temporal and spatial distribu-
tion of the increase in [Ca2/]i in glial cells, especially astro- erate concentration triggered [Ca2/]i oscillations, and at
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
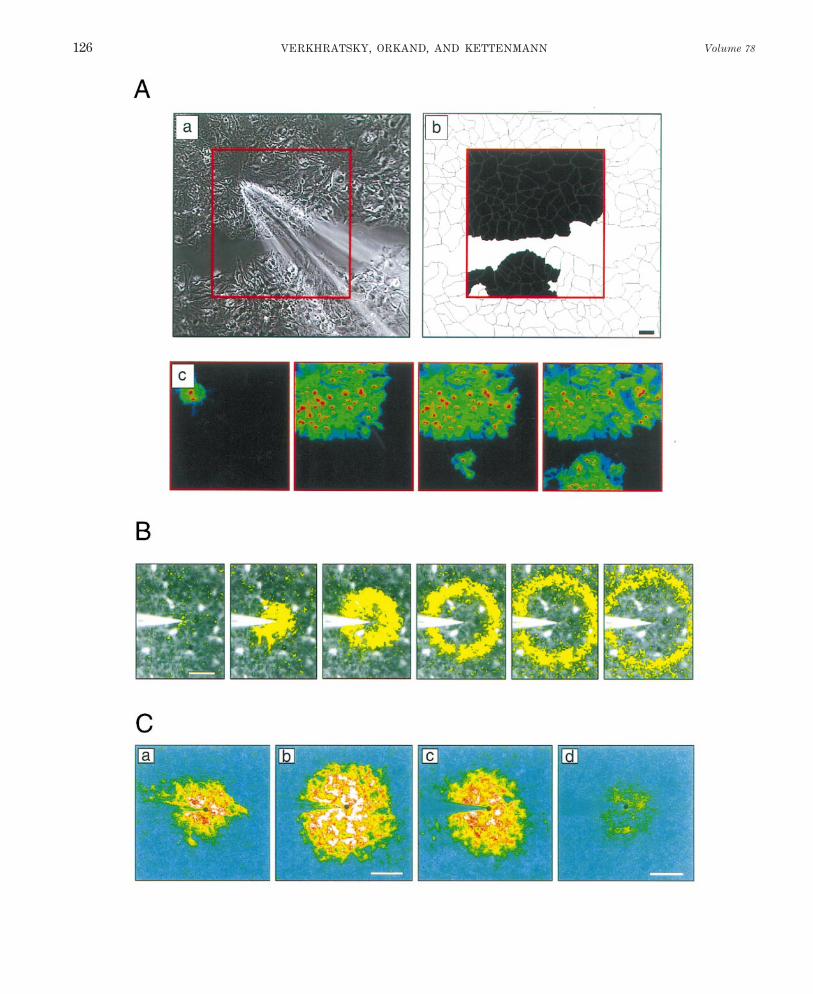
VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78126
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 127
high glutamate concentrations, the initial [Ca2/]i peak was the astrocytic network. The seminal observation wasfollowed by a sustained [Ca2/]i plateau (84). made by Cornell-Bell et al. (83), who found that in the
In addition to a complex temporal pattern, glial Ca2/ presence of glutamate, Ca2/ waves propagate through thesignaling exhibits spatial heterogeneity. In many glial cell astrocyte syncytium. The waves followed complex routes,populations, [Ca2/]i rises first in processes and then spreads without delays at cell borders, for hundreds of micronstoward the soma [e.g., in ATP or KCl-stimulated oligoden- at a velocity of Ç15–20 mm/s (83, 84, 428). The propaga-drocytes (237, 238), in histamine-stimulated astrocytes (194), tion of glutamate-triggered Ca2/ waves requires extracel-or in ATP-challenged Bergmann glial cells (235); cf. Figs. 4– lular Ca2/. An alternative way to induce intercellular Ca2/
6]. Such a pattern is expected from considerations of sur- waves is with focal mechanical stimulation. These wavesface-to-volume relations if the release or influx is constant have different properties; they have a delay at cell bordersfor a given membrane area. However, subsequent analysis and persist in Ca2/-free solutions (61). They are, however,suggests the situation is more complex. The localization of dependent on Ca2/ release from the internal stores; theCa2/ release sites may reflect a higher concentration of Ca2/ depletion of stores with thapsigargin blocks wave propa-signaling molecules in distal parts of glial cells in regions gation (60). The mechanism of intercellular propagationwhere they are in more intimate contact with neurons. Cul- also involves gap junctions; the waves are inhibited bytured astrocytes display more complex spatial [Ca2/]i sig- octanol, halothane (115, 126), or by an endogenous deriva-nals, developing long-lasting [Ca2/]i waves (450, 451). These tive of arachidonic acid, anandamide (427). In the C6 gli-waves are associated with several intracellular loci, each oma cell line, intercellular Ca2/ wave propagation wascharacterized by its own oscillatory pattern with [Ca2/]i observed only in cells that were transfected with the genewaves propagating between them (450, 451). Several hypoth- for the gap junction protein connexin-43 (62). Thus aeses have been proposed to explain [Ca2/]i wave initiation likely mechanism for Ca2/ wave propagation involves in-and propagation. These include periodic fluctuations of cyto- tercellular (via gap junctions) diffusion of InsP3, whichplasmic InsP3 level, [Ca2/]i-dependent regulation of InsP3- generates Ca2/ release in one cell after another (126, 390);gated Ca2/ release channel open probability, and an inter- the degree of intercellular coupling would, presumably,play between InsP3-induced and Ca2/-induced Ca2/ release determine the wave path. The important role of intracellu-(see Refs. 6, 36, 259 for review). lar Ca2/ stores and InsP3 production is substantiated by
In cultured astrocytes obtained from the visual cor- the finding that treatment of astrocytic cultures with thap-tex, the glutamate-evoked [Ca2/]i oscillations demon- sigargin or with PLC blocker U-73122 completely pre-strated a long-lasting modulation in frequency. The second vented the spread of intercellular Ca2/ waves (428). Inter-application of glutamate arriving 1–60 min after the first estingly, interglial communications via gap junctions maychallenge always triggered [Ca2/]i oscillation with higher be regulated by various physiological stimuli, e.g., it in-frequency (332). These changes of frequency were exclu- creases upon glutamate and high K/ treatment (113, 150)sively confined to glutamate-induced Ca2/ signals; [Ca2/]i or even after action potentials in adjacent axons (277).oscillations triggered by other agonists did not show such This susceptibility of interglial gap junctions to externallong-term plasticity. regulation may influence interglial Ca2/ signaling.
Alternative mechanisms for Ca2/ wave propagationB. Intercellular Ca2/ Waves
may involve an extracellular messenger released by stimu-lated cells. An extracellular pathway for Ca2/ wave propa-In cultured astrocytes, a stimulus-evoked intracellu-
lar Ca2/ wave can cross cell boundaries and travel within gation was observed by Hassinger et al. (167). To test the
FIG. 8. Ca2/ wave propagation in glial networks. A: extracellular component in astrocytic Ca2/ wave propagation.Images show an electrically evoked astrocytic Ca2/ wave traversing a cell-free lane. a: Phase-contrast image of a bedof astrocytes in which a cell-free lane had been created by scratching with a micropipette 6 h earlier. Also in view isextracellular stimulating electrode 10 mm above astrocyte upper surface. Red square delineates field of fluorescentimaging of fluo 3 signals. b: Cellular outlines showing astrocytic bed, cell-free lane, and all of astrocytes participatingin Ca2/ wave during observation time depicted on left. c: Pseudocolor images of change in fluorescent signal (F/Fo) inastrocytes at selected time points. Images were collected at 3, 16, 22, and 36 s after stimulation. Note that in thirdframe from left, Ca2/ wave jumps across cell-free lane and subsequently spreads to neighboring astrocytes (4th frame).Although magnitude of spread, amplitude of response, and number of participating cells can vary in each experiment,most preparations with lanes of õ100 mm show responses similar to that shown. Bar Å 50 mm. [From Hassinger et al.(167). Copyright 1997 National Academy of Sciences, USA.] B: propagation of Ca2/ waves in glial cells in acutely isolatedrat retina. Spread of a Ca2/ wave initiated by a mechanical stimulus. Fluorescence image is shown in black and white.Superimposed yellow rings mark leading edge of Ca2/ wave (where change in fluorescence between successive panelsexceeded a threshold value). Interval between panels Å 0.93 s; scale bars in first frame Å 50 mm. C: Ca2/ waves inretinal glia originate from intracellular stores. a: Ca2/ wave evoked by an electrical stimulus. b: Different region ofretina shown in a, stimulated after 31 min in 0 mM Ca2/ and 0.5 mM EGTA. c: Ca2/ wave evoked by an electricalstimulus under control conditions. d: Different region of retina shown in c, stimulated 16 min after addition of 1.5 mMthapsigargin. Scale bar Å 50 mm. [From Newman and Zahs (312). Copyright 1997 American Association for the Advance-ment of Science.]
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78128
importance of the extracellular pathway, they created a [Ca2/]i in glial cells have been carried out with astrocytescell-free lane in confluent astrocytic cultures by mechani- in tissue culture. These cells have membrane receptorscally eliminating astrocytes with a glass pipette. As shown for many neurotransmitters, and their response often in-in Fig. 8A, electrically induced Ca2/ waves appeared to cludes changes in [Ca2/]i . It is tempting to assume thatcross cell free lanes narrower than 120 mm. Moreover, the when the transmitters are released from the neurons thatvelocity of wave propagation through the cell-free lane they provoke responses not only in a postsynaptic cell butdid not differ significantly from the velocity of the wave also in the closely apposed glial cells. Moreover, neuronsspreading via the astrocytic network. The nature of the release a variety of substances, e.g., amines, peptides, andextracellular messenger was not clarified, although gluta- hormones, which diffuse through the extracellular spacemate can be excluded; the wave spreading over cell-free bathing other neurons and glial cells. Among the questionsregions was not modified by glutamate receptor antago- that need to be addressed are the following: 1) Whichnists. Whether the extracellular mechanism may work in receptors are expressed in glial cells in the intact nervousconcert with gap junction propagation or it may take the system? 2) Can a glial cell distinguish between stimulantsleading role in certain brain regions remains totally un- that all increase [Ca2/]i? 3) What is the functional re-clear. sponse of the glial cell after it has undergone an increase
The existence of interastrocytic [Ca2/]i waves in in [Ca2/]i? 4) What other changes within the glial cellbrain tissue has only recently been demonstrated. A prop- result from neuronal activity in addition to an increase inagating wave was observed by Dani et al. (90) in organo- [Ca2/]i?typic hippocampal slice cultures. These waves were initi- Because glial Ca2/ signaling appears to be a ubiqui-ated by stimulation of mossy fibers, which are believed tous consequence of the activation of glial neurotransmit-to utilize glutamate as a neurotransmitter. However, the ter receptor systems, it is of interest to explore whetherintercellular Ca2/ waves observed in these experiments neuronal activity leads to [Ca2/]i fluctuations in glial cells.usually propagated for much shorter distances (2–3 Indeed, experiments performed recently in both periph-astrocyte diameters) than observed in cultures. This might eral and central macroglial cells demonstrate that gliareflect a higher degree of coupling of cultured astrocytes sense nervous activity, and the latter initiates Ca2/ signalsthan astrocytes in situ. More recently, propagating Ca2/
in glial cells. The peripheral glia, the Schwann cells, gener-waves were recorded in glial cells from acutely isolated ate [Ca2/]i transients in response to electrical stimulationrat retina (312). These Ca2/ waves were initiated by either of peripheral nerves (200, 258, 363). In periaxonallocal mechanical or electrical stimulation or by focal ap- Schwann cells, the Ca2/ signal probably results from de-plication of ATP, carbachol, or phenylephrine (Fig. 8, B
polarization occurring in response to increases in [K/]oand C). Interestingly, focal applications of glutamate didresulting from nerve activity; this depolarization, in turn,
not trigger Ca2/ waves in retinal glial networks, althoughtriggers Ca2/ entry via voltage-gated channels and subse-
incubation of the whole preparation in glutamate potenti-quent activation of Ca2/-induced Ca2/ release from inter-ated Ca2/ waves induced by other stimuli. The propaga-nal stores (258). In contrast, in perisynaptic Schwanntion velocity of Ca2/ wave in retinal preparation was Ç25cells, Ca2/ signals were mediated by either muscarinic or
mm/s for all types of stimulation. The retinal Ca2/ wavesP2 purinergic receptors (200, 363) activated by neurotrans-obviously originated from intracellular Ca2/ release, beingmitters released from the nerve ending.preserved in Ca2/-free solution and blocked by thapsigar-
Neuronal activity has been found to induce [Ca2/]igin as shown in Figure 8C.elevation in glial cells from the leech nervous system(371). At least in part, these glial Ca2/ signals were medi-ated through activation of GluR. Similarly, in brain whiteVII. GLIAL CALCIUM SIGNALING AND
matter, electrical stimulation of the optic nerve evokedNEURON-GLIAL INTERACTIONS
oscillatory Ca2/ responses in periaxonal glial cells (65,247); the frequency of [Ca2/]i oscillations correlated withThe coordination of neuronal and glial activity re-stimulation frequency. The possible mechanism underly-quires that appropriate signals pass from one cell type toing axon-glial communication in the optic nerve may in-the other. The study of these signals is a central theme involve glutamate release from the stimulated nerve due toneuroglial research (82, 220, 389). These interactionsreversal of Na//glutamate transporter (65).might involve short-term functions such as ion regulation,
In astrocytes of the gray matter, neuronal-inducedrelease of substrates, or transmitter clearance or muchCa2/ signaling has also been found in both neuronal-gliallonger term processes as might be involved in neuronalcocultures and in hippocampal slice preparations. In neu-guidance during development, myelination, mitosis, andronal-glial cocultures, selective stimulation of neuronsregeneration. The question of how changes in glial [Ca2/]i
with NMDA triggered [Ca2/]i spikes and [Ca2/]i oscilla-serve to couple neuronal and glial functions is a majortions in the neighboring astrocytes (91). Using in situ prep-question raised by the work that has been reviewed and,
unfortunately, largely unanswered. Most of the studies of aration, Dani et al. (90) observed [Ca2/]i transients evoked
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 129
by mossy fiber stimulation in astrocytes in hippocampal concentrations, whereas oligodendroglia die when chal-lenged with 200 mM glutamate (326). This result is proba-cultured slices. Similarly, stimulation of Schaffer collater-
als triggered [Ca2/]i elevation in astrocytes located in CA1 bly because the most abundant astrocytic ionotropicGluRs are of AMPA type, characterized by rapid desensiti-stratum radiatum region (351). In hippocampal and visual
cortex slices, electrical stimulation of presynaptic affer- zation. Removal of AMPA receptor desensitization bycyclothiazide dramatically increased astrocytic vulnera-ents triggered [Ca2/]i oscillations in astrocytes (331);
moreover, the frequency of astrocytic [Ca2/]i oscillations bility to glutamate (93). The high resistivity of astrocytesto glutamate coincides with their role as a major gluta-increased while increasing the intensity or frequency of
afferent stimulation. In all cases, [Ca2/]i transients in as- mate sink in the brain; it is well established that astrocytesare mainly responsible for glutamate removal from thetrocytes were mediated by glutamate presumably released
from neuronal terminals, since astrocytic Ca2/ responses extracellular space (212), being thus one of the importantcomponents of nerve tissue defense against glutamate ex-were mimicked by (1S,3R)-ACPD (331) and were inhibited
by either nonselective ionotropic GluR blocker kinurenic citotoxicity.Injury or brain pathology leads to a complex reactionacid (90, 351) or by mGluR blocker a-methyl-4-carboxy-
phenylglycine (351). from the astrocytes and microglial cells that are an im-portant part of the brain’s defenses (145, 252). AstrocytesIn cocultures, it has been found that not only may
neurons initiate Ca2/ signals in glial cells, but also glial respond with a variety of biochemical, structural, and pro-liferative changes that transform them into so-called ‘‘ac-[Ca2/]i waves may trigger Ca2/ signals in neurons. First,
Nedergaard (310) found that [Ca2/]i waves in astrocytes tive astrocytes.’’ Microglia also undergo dramatic morpho-logical and biochemical changes transforming them intoin rat forebrain cortical cultures triggered [Ca2/]i spikes
in neurons. This astrocyte-to-neuron Ca2/ signaling was phagocytotic macrophages. The initial signals producingthese long-term changes are not well understood; there issensitive to gap junction inhibitors, suggesting the direct
spread of signal from astroglia to neuronal cells. As such, some evidence that Ca2/ signaling is an important (ormaybe even triggering) element in the glial response togap junctions have not been demonstrated in vivo; this
result can be considered an artifact of the culture system. these brain insults. Studies in both in vitro and in vivoexperimental models used to study ischemic damage re-However, in neuronal-glial cocultures from visual cortex
(329), hippocampus (166), and forebrain (59), glial [Ca2/]i vealed that brief exposures of astrocytes to hypoxic/hypo-glycemic conditions trigger a [Ca2/]i increase as a resultwaves induced by mechanical (59, 166), electrical (166),
or agonist (BK; Ref. 329) stimulation resulted in Ca2/ sig- of both activation of voltage-gated Ca2/ channels and Ca2/
release from internal pools (105, 169). The ischemia-in-nals in neurons. In these experiments, glial-to-neuronalCa2/ signaling was sensitive to ionotropic GluRs antago- duced [Ca2/]i increase was activated either by a propagat-
ing wave of raised [K/]o , accompanying injury-inducednists, suggesting a primary role of glutamate released fromglial cells in this form of signaling. spreading depression, or by a massive release of neuro-
transmitters. Interestingly, [Ca2/]i was much more sensi-Therefore, glial Ca2/ signaling is involved, at least inculture systems, in bidirectional neuronal-glial signaling. tive to ischemia in astrocytes in situ, in hippocampal
slices, than in the same cells acutely isolated (105). ThisThe questions of the physiological role of these signals,their precise cellular mechanism, and their occurrence in suggests that the ischemia-induced increase in [Ca2/]i in
astroglia results from factors released by the damagedthe intact nervous system remain unanswered and de-serve further study. tissue (e.g., excess of neurotransmitters or elevated K/
concentration). Intracellular Ca2/ increases in astrocytesinduced by brain damage might change the functional
VIII. GLIAL CALCIUM AND BRAIN PATHOLOGY state of the cell and lead to the release of growth factorsor result in astrocytic volume changes.
It is a well-established paradigm that cellular Ca2/ Another potential pathological Ca2/ signal in glia isoverload is a key trigger in causing cell death (66). Much triggered by the human immunodeficiency virus-1 enve-is known about Ca2/ toxicity in neurons; however, the lope protein gp120. It induces [Ca2/]i increases in bothproblem of [Ca2/]i damage to glial cells is much less ex- cultured astrocytes and oligodendrocytes from cerebel-plored. Experiments on cultured cells demonstrated that lum (76) and cortex (77), but not in neurons (72). TheseCa2/ overload of Schwann cells (291) and oligodendro- [Ca2/]i responses varied between single spikes and [Ca2/]i
cytes (29, 377) might severely damage or kill them. In oscillations. An [Ca2/]i elevation was also observed in cul-neurons exposed to hypoxia or ischemia, the Ca2/ over- tured astrocytes attacked by a fragment (so-called PrPload results mainly from overactivation of GluRs pro- 106–126) of another pathogenic protein, infectious prionduced by excessive glutamate release (glutamate excito- protein. This [Ca2/]i increase was prevented by the dihy-toxicity). Astrocytes but not oligodendrocytes are gener- dropyridine Ca2/ antagonist nicardipine or by Ca2/ re-ally believed to be more resistant to excitotoxic insults; moval from the incubation media, suggesting the leading
role of Ca2/ influx through voltage-gated channels (130).astrocytes can tolerate a very high (up to 5 mM) glutamate
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78130
There have been only a few studies of changes in changes in [Ca2/]i , can it respond appropriately to eachsignal? There are experimental results to indicate that aoligodendrocyte [Ca2/]i after brain damage. A possible
link between [Ca2/]i in oligodendroglia and brain pathol- single cell either in tissue culture (128) or in situ (235,239, 419) responds to individual ligands with a cell-specificogy was suggested by experiments demonstrating that the
addition of complement triggered [Ca2/]i oscillations in characteristic increase in [Ca2/]i that has a different spa-tiotemporal pattern. Thus an individual cell may respondprimary cultured oligodendrocytes (446). These oscilla-
tions were blocked by thapsigargin, indicating an im- to ATP with a brief spike and to norepinephrine or endo-thelin with an oscillatory or sustained increase in [Ca2/]i .portant role of intracellular Ca2/ stores in this phenome-
non. The relative increase may vary in different cellular re-gions, e.g., processes vs. soma, depending on the stimulusMicroglia detect and respond to brain pathology
(246). Damage to the CNS triggers a complex cascade and lead to quite different functional responses. More-over, the exact source of the Ca2/, voltage-dependentleading to a transformation of microglial cells from a rest-
ing form to an active form, termed a macrophage (145). channels versus release from the ER, may create largespatial Ca2/ gradients within the glial cell. Therefore, al-The mechanisms of [Ca2/]i homeostasis and the role of
Ca2/ signaling in microglial function are not known in though the results require confirmation, it appears thatCa2/ signaling exhibits the flexibility to enable the cellsdetail. Microglial cells do not respond to the ‘‘classical’’
neurotransmitters such as glutamate or GABA. They, how- to respond in an appropriate stimulus-specific manner.These considerations do not rule out the possibility thatever, are responsive to signaling molecules from the im-
mune system such as cytokines or chemokines. For exam- there are other important intracellular signaling sub-stances in glial cells that have not been well characterizedple, activation of complement receptors C5a or C3a trig-
gers a Ca2/ increase in cultured microglial cells (292, 318). for lack of appropriate detectors. There can be no ques-tion but that the ready measurement of [Ca2/]i with fluo-The microglial [Ca2/]i transients were also observed in
response to extracellular application of lipopolysaccaride, rescent probes has greatly accelerated the appreciationof the role of this ion in the signaling chain.a potent activator of immune cells (16). The study of the
Ca2/ signaling in microglial cells is at an early stage: therole of [Ca2/]i in microglia activation remains obscure.
X. CONCLUDING REMARKS: CALCIUM
SIGNALS ARE A CONSEQUENCE OF
GLIAL EXCITABILITYIX. CALCIUM SIGNALS AND GLIAL FUNCTION
Glial cells exhibit various mechanisms for controlling The classical view that glia are inexcitable refers totheir inability to respond to electrical stimulation with aand varying [Ca2/]i . As described above, modulation of a
number of intracellular cascades and membrane pathways propagated electrical response, i.e., an action potential.However, as documented throughout this review, glialenables a variety of external stimuli to induce changes in
[Ca2/]i , the Ca2/ signals. Glial cells are able to sense and cells are not passive; they display diverse temporal andspatial increases in [Ca2/]i in response to a variety ofreact to various neuroactive substances; their reactions
almost always involve changes in [Ca2/]i . What glial func- stimuli, e.g., chemical, electrical, or mechanical. These[Ca2/]i increases may outlast the stimulus and exhibit ago-tions are modulated or initiated by such signals? A number
of studies indicate that activation of glial Ca2/ cascades nist-specific spatiotemporal patterns, thus providing apossible means for information coding. The glial [Ca2/]iaffects glial K/ channels (82, 359), thereby modulating
ionic buffering properties of the glial syncytium. Further- signals are apparently also capable of propagating withoutdecrement via gap junctions into neighboring cells andmore, increases in [Ca2/]i promote glycogen breakdown
(343) and gene expression (264, 342) and trigger release even to neurons.The ability of glial cells to actively respond to exter-of neuroactive substances (322, 330). Calcium influx into
glial cell precursors triggers a phosphorylation of cAMP nal stimuli makes them excitable. To avoid confusing thisexcitability with electrical excitability, it is best to referresponse element binding protein (341) that is known to
be a key factor in Ca2/-dependent gene expression. Nor- to glial cells as ‘‘internally calcium excitable.’’ It may evenindeed be appropriate to describe all electrically inexcit-epinephrine, which increases [Ca2/]i , also regulates the
uptake of glutamate and GABA into astrocytes (164). In able cells that do respond to stimuli via second messen-gers as ‘‘internally second messenger excitable.’’ As de-addition, the propagation of Ca2/ waves through syncytial
networks of neuroglia via gap junctions can serve to coor- tailed above, there is quite a bit known about the physiol-ogy of glial Ca2/ signaling. A beginning has been made indinate glial activity and to transfer information between
neurons apposed to different parts of the network. This the attempt to characterize in detail the molecular cas-cades involved in [Ca2/]i homeostasis and shaping ofsignal transfer can participate in the integrative functions
of both glial and neuronal networks. [Ca2/]i signals in glia. The questions that remain to beanswered are as follows: How is Ca2/ signaling involvedWhen a glial cell has multiple receptors that lead to
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 131
17. BAIMBRIDGE, K. G., M. R. CELIO, AND J. H. ROGERS. Calcium-in the integrative function of glial cells? Is glial Ca2/ signal-binding proteins in the nervous system. Trends Neurosci. 15: 303–
ing involved in information processing in glial networks? 308, 1992.Finally, how is glial Ca2/ signaling involved in brain func- 18. BALLANYI, K., AND W.-R. SCHULE. Direct effects of carbachol on
membrane potential and ion activities in leech glial cells. Glia 1:tion? These intriguing questions will be the subject of165–167, 1988.
continued study in this area. 19. BALLERINI, P., M. P. RATHBONE, P. DI IORIO, A. RENZETTI, P.GIULIANI, I. D’ALIMONTE, O. TRUBIANI, F. CACIAGLI, AND R.
We are grateful to Professor R. C. Thomas and Dr. Frank CICCARELLI. Rat astroglial P2Z (P2X7) receptors regulate intracellu-Kirchhoff for helpful comments. lar calcium and purine release. Neuroreport 7: 2533–2537, 1996.
20. BARG, J., M. M. BELCHEVA, R. ZIMLICHMAN, R. LEVY, D. SAYA,A. Verkhratsky and H. Kettenmann’s research is supportedR. J. MCHALE, F. E. JOHNSON, C. J. COSCIA, AND Z. VOGEL. Opi-by grants from SonderForschungsBereich 1534. R. K. Orkand isoids inhibit endothelin-mediated DNA synthesis, phosphoinositidesupported by National Science Foundation (EPSCoR) and the turnover, and Ca2/ mobilization in rat C6 glioma cells. J. Neurosci.
National Institutes of Health (National Institute of Neurological 14: 5858–5864, 1994.Disorders and Stroke, Minority Institutional Research Develop- 21. BARNUM, S. R. Complement biosynthesis in the central nervous
system. Crit. Rev. Oral Biol. Med. 6: 132–146, 1995.ment Program, and Fogarty Foundation).22. BARONE, F. C., M.-Y.-T. GLOBUS, W. J. PRICE, R. F. WHITE, B. L.
STORER, G. Z. FEUERSTEIN, R. BUSTO, AND E. H. OHLSTEIN.Endothelin levels increase in rat focal and global ischaemia. J.
REFERENCES Cereb. Blood Flow Metab. 14: 337–342, 1993.23. BARRES, B. A., L. L. CHUN, AND D. P. COREY. Ion channel expres-
sion by white matter glia. I. Type 2 astrocytes and oligodendrocytes.1. AHMED, Z., C. A. LEWIS, AND D. S. FABER. Glutamate stimulatesGlia 1: 10–30, 1988.release of Ca2/ from internal stores in astroglia. Brain Res. 516:
24. BARRES, B. A., L. L. CHUN, AND D. P. COREY. Calcium current in165–169, 1990.cortical astrocytes: induction by cAMP and neurotransmitters and2. AKOPIAN, G., K. KRESSIN, A. DEROUICHE, AND C.permissive effect of serum factors. J. Neurosci. 9: 3169–3175, 1989.STEINHA
¨USER. Identified glial cells in the early postnatal mouse
25. BARRES, B. A., W. J. KOROSHETZ, L. L. CHUN, AND D. P. COREY.hippocampus display different types of Ca2/ currents. Glia 17: 181–Ion channel expression by white matter glia: the type-1 astrocyte.194, 1996.Neuron 5: 527–544, 1990.3. ALBRECHT, J., B. PEARCE, AND S. MURPHY. Evidence for an
26. BAX, W. A., AND P. R. SAXENA. The current endothelin receptorinteraction between GABAB and glutamate receptors in astrocytesclassification: time for reconsideration? Trends Pharmacol. Sci. 15:as revealed by changes in Ca2/ flux. Eur. J. Pharmacol. 125: 463–379–386, 1994.464, 1986.
27. BAZAN, N. G. Platelet-activating factor is a synapse messenger and4. AMEDEE, T., AND S. DESPEYROUX. ATP activates cationic andan intracellular modulator of gene expression. J. Lipid. Mediat.anionic conductances in Schwann cells cultured from dorsal rootCell Signal. 10: 83–86, 1994.ganglia of the mouse. Proc. R. Soc. Lond. B Biol. Sci. 259: 277–
28. BEAUDU, C., S. DESPEYROUX, AND T. AMEDEE. 8-(4-Chlorophe-284, 1995.nylthio)-cAMP has opposite long-term effects on potassium and5. AMEDEE, T., E. ELLIE, B. DUPOUY, AND J. D. VINCENT. Voltage-calcium channel currents in cultured mouse Schwann cells (Ab-dependent calcium and potassium channels in Schwann cells cul-stract). J. Physiol. (Lond.) 494: 53P, 1996.tured from dorsal root ganglia of the mouse. J. Physiol. (Lond.)
29. BENJAMINS, J. A., AND L. NEDELKOSKA. Release of intracellular441: 35–56, 1991.calcium stores leads to retraction of membrane sheets and cell6. AMUNDSON, J., AND D. CLAPHAM. Calcium waves. Curr. Opin.death in mature mouse oligodendrocytes. Neurochem. Res. 21: 471–
Neurobiol. 3: 375–382, 1993.479, 1996.7. ANANTH U. S., U. LELI, AND G. HAUSER. Stimulation of phospho-
30. BENNINGER, C., J. KADIS, AND D. A. PRINCE. Extracellular cal-inositides hydrolysis by serotonin in C6 glioma cells. J. Neurochem.cium and potassium changes in hippocampal slices. Brain Res.48: 253–261, 1987.187: 165–182, 1980.8. ANDRE, C., G. DOS SANTOS, AND A. KOULAKOFF. Muscarinic
31. BERGER, T., J. SCHNITZER, P. M. ORKAND, AND H. KETTEN-receptor profiles of mouse brain astrocytes in culture vary withMANN. Sodium and calcium currents in glial cells of the mousetheir tissue of origin but differ from those of neurons. Eur. J.corpus callosum slice. Eur. J. Neurosci. 4: 1271–1284, 1992.Neurosci. 6: 1702–1709, 1994.
32. BERGER, T., W. WALZ, J. SCHNITZER, AND H. KETTENMANN.9. ANSSELIN, A. D., D. F. DAVEY, AND D. G. ALLEN. ExtracellularGABA- and glutamate-activated currents in glial cells of the mouseATP increases intracellular calcium in cultured adult Schwanncorpus callosum slice. J. Neurosci. Res. 31: 21–27, 1992.cells. Neuroscience 76: 947–955, 1997.
33. BERNSTEIN, M., S. A. LYONS, T. MOLLER, AND H. KETTENMANN.10. ARBONES, L., F. PICATOSTE, AND A. GARCIA. Histamine H1-re-Receptor-mediated calcium signalling in glial cells from mouse cor-ceptors mediate phosphoinositide hydrolysis in astrocyte-enrichedpus callosum slices. J. Neurosci. Res. 46: 152–163, 1996.primary cultures. Brain Res. 450: 144–152, 1988.
34. BERRIDGE, M. J. Inositol trisphosphate and calcium signalling. Na-11. ASANO, S., T. MATSUDA, K. TAKUMA, H. S. KIM, T. SATO, T.ture 361: 315–325, 1993.NISHIKAWA, AND A. BABA. Nitroprusside and cyclic GMP stimu-
35. BERRIDGE, M. J. Capacitative calcium entry. Biochem. J. 312: 1–late Na/-Ca2/ exchange activity in neuronal preparations and cul-11, 1995.tured rat astrocytes. J. Neurochem. 64: 2437–2441, 1995.
36. BERRIDGE, M. J., AND A. GALIONE. Cytosolic calcium oscillators.12. ATTWELL, D., AND P. MOBBS. Neurotransmitter transporters.FASEB J. 2: 3074–3082, 1988.Curr. Opin. Neurobiol. 4: 353–359, 1994.
37. BEVAN, S., S. Y. CHIU, P. T. GRAY, AND J. M. RITCHIE. The pres-13. AUGUSTINE, G. J. Combining patch-clamp and optical methods inence of voltage-gated sodium, potassium and chloride channels inbrain slices. J. Neurosci. Methods 54: 163–169, 1994.rat cultured astrocytes. Proc. R. Soc. Lond. B Biol. Sci. 225: 299–14. BACKUS, K. H., AND T. BERGER. Developmental variation of the313, 1985.permeability to Ca2/ of AMPA receptors in presumed hilar glial
38. BITO, H., Z. HONDA, M. NAKAMURA, AND T. SHIMIZU. Cloning,recursor cells. Pflu¨gers Arch. 431: 244–252, 1995.
expression and tissue distribution of rat platelet-activating factor15. BACKUS, K. H., T. BERGER, AND H. KETTENMANN. Activation ofreceptor cDNA. Eur. J. Biochem. 221: 211–218, 1994.neurokinin receptors modulates K/ and Cl0 channel activity in
39. BITO, H., M. NAKAMURA, Z. HONDA, T. IZUMI, T. IWATSUBO, Y.cultured astrocytes from rat cortex. Brain Res. 541: 103–109, 1991.SEYAMA, A. OGURA, Y. KUDO, AND T. SHIMIZU. Platelet-activating16. BADER, M. F., L. TAUPENOT, G. ULRICH, D. AUNIS, AND J. CIE-factor (PAF) receptor in rat brain: PAF mobilizes intracellular Ca2/SIELSKI TRESKA. Bacterial endotoxin induces [Ca2/]i transientsin hippocampal neurons. Neuron 9: 285–294, 1992.and changes the organization of actin in microglia. Glia 11: 336–
344, 1994. 40. BLAUSTEIN, M. P., W. F. GOLDMAN, G. FONTANA, B. K.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78132
KRUEGER, E. SANTIAGO, T. D. STEEL, D. N. WEISS, AND P. J. a therapeutically relevant lithium concentration. Brain Res. 711:245–248, 1996.YAROWSKY. Physiological role of the sodium-calcium exchanger
in nerve and muscle. Ann. NY Acad. Sci. 639: 254–274, 1991. 64. CHEN, Y., L. PENG, X. ZHANG, J. U. STOLZENBURG, AND L.HERTZ. Further evidence that fluoxetine interacts with a 5-HT2C41. BONNER, T. I., N. J. BUCKLEY, A. C. YOUNG, AND M. R. BRANN.
Identification of a family of muscarinic acetylcholine receptor receptor in glial cells. Brain Res. Bull. 38: 153–159, 1995.genes. Science 237: 527–532, 1987. 65. CHIU, S. Y., AND S. KRIEGLER. Neurotransmitter-mediated signal-
ing between axons and glial cells. Glia 11: 191–200, 1994.42. BOOTMAN, M. D., AND M. J. BERRIDGE. The elemental principlesof calcium signalling. Cell 83: 675–678, 1995. 66. CHOI, D. W. Calcium: still center-stage in hypoxim-ischaemic neu-
ronal death. Trends Neurosci. 18: 58–60, 1995.43. BORDEY, A., P. FELTZ, AND J. TROUSLARD. Mobilization of intra-cellular calcium by substance P in a human astrocytoma cell line 67. CHOLEWINSKI, A. J., M. R. HANLEY, AND G. P. WILKIN. A phos-(U-373 MG). Glia 11: 277–283, 1994. phoinositide-linked peptide response in astrocytes: evidence for
regional heterogeneity. Neurochem. Res. 13: 389–394, 1988.44. BORGES, K., C. OHLEMEYER, J. TROTTER, AND H. KETTEN-MANN. AMPA/kainate receptor activation in murine oligodendro- 68. CHOLEWINSKI, A. J., G. STEVENS, A. M. MCDERMOTT, AND G. P.cyte precursor cells leads to activation of a cation conductance, WILKIN. Identification of B2 bradykinin binding sites on culturedcalcium influx and blockade of delayed rectifying K/ channels. cortical astrocytes. J. Neurochem. 57: 1456–1458, 1991.Neuroscience 63: 135–149, 1994. 69. CHOLEWINSKI, A. J., AND G. P. WILKIN. Bradykinin and vasopres-
45. BORGES, K., G. WOLSWIJK, C. OHLEMEYER, AND H. KETTEN- sin both stimulate phosphoinositide turnover in astrocytes in highMANN. Adult rat optic nerve oligodendrocyte progenitor cells ex- serum- and low serum-containing media but have no effect onpress a distinct repertoire of voltage- and ligand-gated ion channels. [3H]thymidine incorporation. Neurochem. Int. 13: 393–396, 1988.J. Neurosci. Res. 40: 591–605, 1995. 70. CHUANG, D. M., W. W. LIN, AND C. Y. LEE. Endothelin-induced
46. BOWMAN, C. L., AND H. K. KIMELBERG. Excitatory amino acids activation of phosphoinositide turnover, calcium mobilization, anddirectly depolarize rat brain astrocytes in primary culture. Nature transmitter release in cultured neurons and neurally related cell311: 656–659, 1984. types. J. Cardiovasc. Pharmacol. 17, Suppl. 7: S85–S88, 1991.
47. BRISMAR, T. Physiology of transformed glial cells. Glia 15: 231– 71. CHUN, L. L. Y., B. A. BARRES, AND D. P. COREY. Induction of a243, 1995. calcium channel in astrocytes by cAMP. Soc. Neurosci. Abstr. 12:
1346, 1986.48. BRODIE, C. Functional PAF receptors in glia cells: binding parame-ters and regulation of expression. Int. J. Dev. Neurosci. 12: 631– 72. CIARDO, A., AND J. MELDOLESI. Effects of the HIV-1 envelope640, 1994. glycoprotein gp120 in cerebellar cultures. [Ca2/]i increases in a glial
cell subpopulation. Eur. J. Neurosci. 5: 1711–1718, 1993.49. BRUNE, T., AND J. W. DEITMER. Intracellular acidification andCa2/ transients in cultured rat cerebellar astrocytes evoked by glu- 73. CLAPHAM, D. E. Calcium signaling. Cell 80: 259–268, 1995.tamate agonists and noradrenaline. Glia 14: 153–161, 1995. 74. CLARK, B., AND P. MOBBS. Transmitter-operated channels in rabbit
50. BRUNER, G., AND S. MURPHY. UTP activates multiple second mes- retinal astrocytes studied in situ by whole-cell patch clamping. J.
senger systems in cultured rat astrocytes. Neurosci. Lett. 162: 105– Neurosci. 12: 664–673, 1992.108, 1993. 75. CLEMENTI, E., AND J. MELDOLESI. Pharmacological and func-
51. BURGESS, S. K., AND K. D. MCCARTHY. Autoradiographic quantita- tional properties of voltage-independent Ca2/ channels. Cell Cal-
tion of beta-adrenergic receptors on neural cells in primary cul- cium 19: 269–279, 1996.tures. I. Pharmacological studies of [125I]pindolol binding of individ- 76. CODAZZI, F., A. MENEGON, D. ZACCHETTI, A. CIARDO, F. GRO-ual astroglial cells. Brain Res. 335: 1–9, 1985. HOVAZ, AND J. MELDOLESI. HIV-1 gp120 glycoprotein induces
52. BURNASHEV, N. Calcium permeability of glutamate gated channels [Ca2/]i responses not only in type-2 but also type-1 astrocytes andin the central nervous system. Curr. Opin. Neurobiol. 6: 311–317, oligodendrocytes of the rat cerebellum. Eur. J. Neurosci. 7: 1333–1996. 1341, 1995.
53. BURNASHEV, N., A. KHODOROVA, P. JONAS, P. J. HELM, W. WIS- 77. CODAZZI, F., G. RACCHETTI, F. GROHOVAZ, AND J. MELDOLESI.DEN, H. MONYER, P. H. SEEBURG, AND B. SAKMANN. Calcium- Transduction signals induced in rat brain cortex astrocytes by thepermeable AMPA-kainate receptors in fusiform cerebellar glial HIV-1 gp120 glycoprotein. FEBS Lett. 384: 135–137, 1996.cells. Science 256: 1566–1570, 1992. 78. COHEN, R. I., AND G. ALMAZAN. Rat oligodendrocytes express
54. BURNSTOCK, G., AND C. M. BROWN. An introduction to purinergic muscarinic receptors coupled to phosphoinositide hydrolysis andreceptors. In: Purinergic Receptors, edited by G. Burnstock. Lon- adenylyl cyclase. Eur. J. Neurosci. 6: 1213–1224, 1994.don: Chapman & Hall, 1981, p. 1–46. 79. COHEN, R. I., E. MOLINA-HOLGADO, AND G. ALMAZAN. Carba-
chol stimulates c-fos expression and proliferation in oligodendro-55. BURNSTOCK, G., AND B. F. KING. Numbering of cloned P2 recep-tors. Drug Dev. Res. 38: 67–71, 1996. cyte progenitors. Mol. Brain Res. 43: 193–201, 1996.
80. COLMERS, W. F., AND D. BLEAKMAN. Effects of neuropeptide Y56. BYLUND, D. B., D. C. EIKENBERG, J. P. HIEBLE, S. Z. LANGER,R. J. LEFKOWITZ, K. P. MINNEMAN, P. B. MOLINOFF, R. R. RUF- on the electrical properties of neurons. Trends Neurosci. 17: 373–
379, 1994.FOLO, JR., AND U. TRENDELENBURG. International Union of Phar-macology nomenclature of adrenoceptors. Pharmacol. Rev. 46: 81. CONDORELLI, D. F., P. DELL’ALBANI, M. CORSARO, V. BARRESI,121–136, 1994. AND A. M. GIUFFRIDA STELLA. AMPA-selective glutamate recep-
tor subunits in astroglial cultures. J. Neurosci. Res. 36: 344–356,57. CARAFOLI, E. Calcium pump of the plasma membrane. Physiol.
Rev. 71: 129–153, 1992. 1993.82. COOPER, M. S. Intercellular signaling in neuronal-glial networks.58. CHANGEUX, J. P. Thudichum Medal Lecture. The acetylcholine
receptor: a model for allosteric membrane proteins. Biochem. Soc. Biosystems 34: 65–85, 1995.Trans. 23: 195–205, 1995. 83. CORNELL BELL, A. H., S. M. FINKBEINER, M. S. COOPER, AND
S. J. SMITH. Glutamate induces calcium waves in cultured astro-59. CHARLES, A. C. Glia-neuron intercellular calcium signaling. Dev.
Neurosci. 16: 196–206, 1994. cytes: long-range glial signaling. Science 247: 470–473, 1990.84. CORNELL BELL, A. H., AND S. M. FINKBEINER. Ca2/ waves in60. CHARLES, A. C., E. R. DIRKSEN, J. E. MERRILL, AND M. J. SAND-
ERSON. Mechanisms of intercellular calcium signaling in glial cells astrocytes. Cell Calcium 12: 185–204, 1991.studied with dantrolene and thapsigargin. Glia 7: 134–145, 1993. 85. CORVALAN, V., R. COLE, J. DE VELLIS, AND S. HAGIWARA. Neu-
ronal modulation of calcium channel activity in cultured rat astro-61. CHARLES, A. C., J. E. MERRILL, E. R. DIRKSEN, AND M. J. SAND-ERSON. Intercellular signaling in glial cells: calcium waves and cytes. Proc. Natl. Acad. Sci. USA 87: 4345–4348, 1990.oscillations in response to mechanical stimulation and glutamate. 86. COURAUD, P. O., O. DURIEU TRAUTMANN, D. L. NGUYEN, P.Neuron 6: 983–992, 1991. MARIN, F. GLIBERT, AND A. D. STROSBERG. Functional endo-
thelin-1 receptors in rat astrocytoma C6. Eur. J. Pharmacol. 206:62. CHARLES, A. C., C. C. G. NAUS, D. ZHU, G. M. KIDDER, E. R.DIRKSEN, AND M. J. SANDERSON. Intercellular calcium signalling 191–198, 1991.via gap junctions in glioma cells. J. Cell Biol. 118: 195–201, 1992. 87. CZUBAYKO, U., AND G. REISER. [Ca2/]i oscillations in single rat
glioma cells induced by thrombin through activation of cell surface63. CHEN, Y., AND L. HERTZ. Inhibition of noradrenaline stimulatedincrease in [Ca2/]i in cultured astrocytes by chronic treatment with receptors. Neuroreport 6: 1249–1252, 1995.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 133
88. DALZIEL, H. H., AND D. P. WESTFALL. Receptors for adenine nu- of neurokinin-1 receptors on human U373MG astrocytoma cells.Glia 6: 89–95, 1992.cleotides and nucleosides: subclassification, distribution, and mo-
lecular characterization. Pharmacol. Rev. 46: 449–466, 1994. 111. ELSHOURBAGY, N. A., J. A. LEE, D. R. KORMAN, P. NUTHALA-GANTI, D. R. SYLVESTER, A. G. DILELLA, J. A. SUTIPHONG, AND89. DAM, T. V., E. ESCHER, AND R. QUIRION. Evidence for the exis-
tence of three classes of neurokinin receptors in brain. Differential C. S. KUMAR. Molecular cloning and characterization of the majorendothelin receptor subtype in porcine cerebellum. Mol. Pharma-ontogeny of neurokinin-1, neurokinin-2 and neurokinin-3 binding
sites in rat cerebral cortex. Brain Res. 453: 372–376, 1988. col. 41: 465–473, 1992.112. ENKVIST, M. O., I. HOLOPAINEN, AND K. E. AKERMAN. Glutamate90. DANI, J. W., A. CHERNJAVSKY, AND S. J. SMITH. Neuronal activity
triggers calcium waves in hippocampal astrocyte networks. Neuron receptor-linked changes in membrane potential and intracellularCa2/ in primary rat astrocytes. Glia 2: 397–402, 1989.8: 429–440, 1992.
91. DANI, J. W., AND S. J. SMITH. The triggering of astrocytic calcium 113. ENKVIST, M. O., AND K. D. MCCARTHY. Astroglial gap junctioncommunication is increased by treatment with either glutamate orwaves by NMDA-induced neuronal activation. Ciba Found. Symp.
188: 195–205, 1995. high K/ concentration. J. Neurochem. 62: 489–495, 1994.114. ENKVIST, M. O. K., H. HAMALAINEN, C. C. JANSSON, J. P. KUK-92. DAVE, V., G. W. GORDON, AND K. D. MCCARTHY. Cerebral type 2
astroglia are heterogeneous with respect to their ability to respond KONEN, R. HAUTALA, M. J. COURTNEY, AND K. E. O. AKERMAN.Coupling of astroglial a2-adrenoreceptors to second messengerto neuroligands linked to calcium mobilization. Glia 4: 440–447,
1991. pathways. J. Neurochem. 66: 2394–2401, 1996.115. ENKVIST, M. O. K., AND K. D. MCCARTHY. Activation of protein93. DAVID, J. C., K. A. YAMADA, M. R. BAGWE, AND M. P. GOLDBERG.
AMPA receptor activation is rapidly toxic to cortical astrocytes kinase C blocks astroglial gap junction communication and inhibitsthe spread of calcium waves. J. Neurochem. 59: 519–526, 1992.when desensitisation is blocked. J. Neurosci. 16: 200–209, 1996.
94. DEECHER, D. C., B. D. WILCOX, V. DAVE, P. A. ROSSMAN, AND 116. ERIKSSON, P. S., M. NILSSON, M. WAGBERG, E. HANSSON, AND
L. RONNBACK. Kappa-opioid receptors on astrocytes stimulate L-H. K. KIMELBERG. Detection of 5-hydroxytryptamine2 receptorsby radioligand binding, Northern blot analysis, and Ca2/ responses type Ca2/ channels. Neuroscience 54: 401–407, 1993.in rat primary astrocyte cultures. J. Neurosci. Res. 35: 246–256, 117. EVANS, P. D., V. REALE, R. M. MERZON, AND J. VILLEGAS. N-1993. methyl-D-aspartate (NMDA) and non-NMDA type glutamate recep-
tors are present on squid giant axon Schwann cells. J. Exp. Biol.95. DEL RIO-HORTEGA, P. La microglia y su transformation en celulasan basoncito y cuerpos granulo-adiposos. Trab. Lab. Invest. Biol. 157: 593–600, 1991.Madrid 18: 37–82, 1920. 118. EVANS, P. D., V. REALE, R. M. MERZON, AND J. VILLEGAS. N-
methyl-D-aspartate (NMDA) and non-NMDA (metabotropic) type96. DELUMEAU, J. C., F. PETITET, J. CORDIER, J. GLOWINSKI, AND
J. PREMONT. Synergistic regulation of cytosolic Ca2/ concentra- glutamate receptors modulate the membrane potential of theSchwann cell of the squid giant nerve fibre. J. Exp. Biol. 173: 229–tion in mouse astrocytes by NK1 tachykinin and adenosine agonists.
J. Neurochem. 57: 2026–2035, 1991. 249, 1992.119. FATATIS, A., L. A. HOLTZCLAW, R. AVIDOR, D. E. BRENNEMAN,97. DEMEMES, D., A. LLEIXA, AND C. J. DECHESNE. Cellular and
subcellular localization of AMPA-selective glutamate receptors in AND J. T. RUSSELL. Vasoactive intestinal peptide increases intracel-lular calcium in astroglia: synergism with alpha-adrenergic recep-the mammalian peripheral vestibular system. Brain Res. 671: 83–
94, 1995. tors. Proc. Natl. Acad. Sci. USA 91: 2036–2040, 1994.120. FATATIS, A., AND R. J. MILLER. Sphingosine and sphingosine 1-98. DENT, M. A. R., G. RAISMAN, AND F. A. LAI. Expression of type 1
inositol 1,4,5 trisphosphate receptor during axogenesis and synap- phosphate differentially modulate platelet-derived growth factor-BB-induced Ca2/ signaling in transformed oligodendrocytes. J.tic contact in the central and peripheral nervous system of devel-
oping rat. Development 122: 1029–1039, 1996. Biol. Chem. 271: 295–301, 1996.121. FATATIS, A., AND J. T. RUSSELL. Spontaneous changes in intracel-99. DI SCALA GUENOT, D., D. MOUGINOT, AND M. T. STROSSER.
Increase of intracellular calcium induced by oxytocin in hypotha- lular calcium concentration in type I astrocytes from rat cerebralcortex in primary culture. Glia 5: 95–104, 1992.lamic cultured astrocytes. Glia 11: 269–276, 1994.
100. DI SCALA GUENOT, D., AND M. T. STROSSER. Oxytocin receptors 122. FELDER, C. C. Muscarinic acetylcholine receptors: signal transduc-tion through multiple effectors. FASEB J. 9: 619–625, 1995.on cultured astroglial cells. Kinetic and pharmacological character-
ization of oxytocin-binding sites on intact hypothalamic and hippo- 123. FERNANDEZ-DURANGO, R., J. A. DE JUAN, H. ZIMMAN, F. J.campic cells from foetal rat brain. Biochem. J. 284: 491–497, 1992. MOYA, M. GARCIA DE LA COBA, AND A. FERNANDEZ-CRUZ. Iden-
101. DI SCALA GUENOT, D., AND M. T. STROSSER. Oxytocin receptors tification of endothelin receptor subtype (ETB) in human cerebralon cultured astroglial cells. Regulation by a guanine-nucleotide- cortex using subtype-selective ligands. J. Neurochem. 62: 1482–binding protein and effect of Mg2/. Biochem. J. 284: 499–505, 1992. 1488, 1994.
102. DRURY, A. N., AND A. SZENT-GYORGYI. The physiological activity 124. FERRARI, D., M. VILLALBA, P. CHIOZZI, S. FALZONI, P. RIC-of adenine compounds with special reference to their action upon CIARDI CASTAGNOLI, AND F. DI VIRGILIO. Mouse microglial cellsthe mammalian heart. J. Physiol. (Lond.) 68: 213–237, 1929. express a plasma membrane pore gated by extracellular ATP. J.
Immunol. 156: 1531–1539, 1996.103. DUFFY, S., AND B. A. MACVICAR. Potassium-dependent calciuminflux in acutely isolated hippocampal astrocytes. Neuroscience 61: 125. FEUERSTEIN, G., J.-L. GU, E. H. OHLSTEIN, F. C. BARONE, AND
51–61, 1994. T.-L. YUE. Peptidic endothelin-1 receptor antagonist, BQ-123, andneuroprotection. Peptides 15: 467–469, 1994.104. DUFFY, S., AND B. A. MACVICAR. Adrenergic calcium signaling in
astrocyte networks within the hippocampal slice. J. Neurosci. 15: 126. FINKBEINER, S. Calcium waves in astrocytes—filling in the gaps.5535–5550, 1995. Neuron 8: 1101–1108, 1992.
105. DUFFY, S., AND B. A. MACVICAR. In vitro ischemia promotes cal- 127. FINKBEINER, S., AND M. E. GREENBERG. Ca2/-dependent routescium influx and intracellular calcium release in hippocampal astro- to ras: mechanisms for neuronal survival, differentiation, and plas-cytes. J. Neurosci. 16: 71–81, 1996. ticity? Neuron 16: 233–236, 1996.
106. DUMONT, Y., J. C. MARTEL, A. FOURNIER, S. ST. PIERRE, AND R. 128. FINKBEINER, S. M. Glial calcium. Glia 9: 83–104, 1993.QUIRION. Neuropeptide Y and neuropeptide Y receptor subtypes in 129. FINKBEINER, S. M. Modulation and control of intracellular cal-brain and peripheral tissues. Prog. Neurobiol. 38: 125–167, 1992. cium. In: Neuroglia, edited by H. Kettenmann and B. R. Ransom.
107. DYER, C. A., AND J. A. BENJAMINS. Glycolipids and transmem- New York: Oxford Univ. Press, 1995, p. 273–288.brane signaling: antibodies to galactocerebroside cause an influx 130. FLORIO, T., M. GRIMALDI, A. SCORZIELLO, M. SALMONA, O.of calcium in oligodendrocytes. J. Cell Biol. 111: 625–633, 1990. BUGIANI, F. TAGLIAVINI, G. FORLONI, AND G. SCHETTINI. Intra-
108. EDWARDS F. A. ATP receptors. Curr. Opin. Neurobiol. 4: 347– cellular calcium rise through L-type calcium channels, as molecular352, 1994. mechanism for prion protein fragment 106–126-induced astroglial
proliferation. Biochem. Biophys. Res. Commun. 228: 397–405,109. EDWARDS, F. A., AND A. J. GIBB. ATP—a fast neurotransmitter.FEBS Lett. 325: 86–89, 1993. 1996.
131. FRASER, D. D., S. DUFFY, K. J. ANGELIDES, J. L. PEREZ-VELAZ-110. EISTETTER, H. R., A. MILLS, R. BREWSTER, S. ALOUANI, C.RAMBOSSON, AND E. KAWASHIMA. Functional characterization QUEZ, H. KETTENMANN, AND B. A. MACVICAR. GABAA/benzodi-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78134
azepine receptors in acutely isolated hippocampal astrocytes. J. and other vasoactive peptides in the brain of Mongolian gerbil. J.
Cardiovasc. Pharmacol. 20, Suppl.: S41–S44, 1992.Neurosci. 15: 2720–2732, 1995.154. GLAUM, S. R., J. A. HOLZWARTH, AND R. J. MILLER. Glutamate132. FRASER, D. D., L. A. MUDRICK DONNON, AND B. A. MACVICAR.
receptors activate Ca2/ mobilization and Ca2/ influx into astro-Astrocytic GABA receptors. Glia 11: 83–93, 1994.cytes. Proc. Natl. Acad. Sci. USA 87: 3454–3458, 1990.133. FREDHOLM, B. B., M. P. ABBRACCHIO, G. BURNSTOCK, J. W.
155. GOLDMAN, R. S., S. M. FINKBEINER, AND S. J. SMITH. EndothelinDALY, T. K. HARDEN, K. A. JACOBSON, P. LEFF, AND M. WIL-induces a sustained rise in intracellular calcium in hippocampalLIAMS. Nomenclature and classification of purinoceptors. Pharma-astrocytes. Neurosci. Lett. 123: 4–8, 1991.col. Rev. 46: 143–156, 1994.
156. GOLDMAN, W. F., P. J. YAROWSKY, M. JUHASZOVA, B. K.134. FRIEL, D. D. Calcium oscillations in neurons. Ciba Found. Symp.KRUEGER, AND M. P. BLAUSTEIN. Sodium/calcium exchange in188: 210–223, 1995.rat cortical astrocytes. J. Neurosci. 14: 5834–5843, 1994.135. FUJIWARA, Y., C. R. MANTIONE, AND H. L. YAMAMURA. Identifi-
157. GOLDSTEIN, I. M. Complement: biologically active products. In:cation of B2 bradykinin binding sites in guinea-pig brain. Eur. J.Inflammation, edited by J. I. Gallin, I. M. Goldstein, and R. Sny-Pharmacol. 147: 487–488, 1988.derman. New York: Raven, 1988, p. 55–74.136. FUKUI, H., N. INAGAKI, S. ITO, A. KUBO, H. KONDOH, A. YAMA-
158. GOLOVINA, V. A., L. L. BAMBRICK, P. J. YAROWSKY, B. K.TODANI, AND H. WADA. Histamine H1-receptors on astrocytes inKRUEGER, AND M. P. BLAUSTEIN. Modulation of two functionallyprimary cultures: a possible target for histaminergic neurones.distinct Ca2/ stores in astrocytes: role of the plasmalemmal Na/CaAgents Actions Suppl. 33: 161–180, 1991.exchanger. Glia 16: 296–305, 1996.137. FULTON, B. P., J. F. BURNE, AND M. C. RAFF. Visualization of O-
159. GRYNKIEWICZ, G., M. POENIE, AND R. Y. TSIEN. A new generation2A progenitor cells in developing and adult rat optic nerve by quis-of Ca2/ indicators with greatly improved fluorescent properties. J.qualate-stimulated cobalt uptake. J. Neurosci. 12: 4816–4833, 1992.Biol. Chem. 260: 3440–3450, 1985.138. FURUICHI, T., D. FURUTAMA, Y. HAKAMATA, J. NAKAI, H.
160. HAAS, S., J. A. BROCKHAUS, A. VERKHRATSKY, AND H. KETTEN-TAKESHIMA, AND K. MIKOSHIBA. Multiple types of ryanodine re-MANN. ATP-induced membrane currents in ameboid microgliaceptor/Ca2/ release channels are differentially expressed in rabbitacutely isolated from mouse brain slices. Neuroscience 75: 257–brain. J. Neurosci. 14: 4794–4805, 1994.261, 1996.139. FURUICHI, T., K. KOHDA, A. MIYAWAKI, AND K. MIKOSHIBA.
161. HALL, J. Bradykinin receptors: pharmacological and biologicalIntracellular channels. Curr. Opin. Neurobiol. 4: 294–303, 1994.roles. Pharmacol. Ther. 56: 131–190, 1992.140. GABELLINI, N., L. FACCI, D. MILANI, A. NEGRO, L. CALLEGARO,
162. HAMA, H., T. SAKURAI, Y. KASUYA, M. FUJIKI, T. MASAKI, ANDS. D. SKAPER, AND A. LEON. Differences in induction of c-fos tran-K. GOTO. Action of endothelin-1 on rat astrocytes through the ETBscription by cholera toxin-derived cyclic AMP and Ca2/ signals inreceptor. Biochem. Biophys. Res. Commun. 186: 355–362, 1992.astrocytes and 3T3 fibroblasts. Exp. Cell Res. 194: 210–217, 1991.
163. HAMLYN, J. M., M. P. BLAISTEIN, S. BOVA, D. W. DUCHARME,141. GALIONE, A. Cyclic ADP-ribose: a new way to control calcium.D. W. HARRIS, F. MANDEL, W. R. MATHEWS, AND J. H. LUDENS.
Science 259: 325–326, 1993.Identification and characterization of a ouabain-like compaund142. GALLO, V., AND J. T. RUSSELL. Excitatory amino acid receptors infrom human plasma. Proc. Natl. Acad. Sci. USA 81: 6259–6263,glia: different subtypes for distinct functions? J. Neurosci. Res. 42:1991.1–8, 1995.
164. HANSSON, E., AND L. RONNBACK. Receptor regulation of the glu-143. GAMBETTI, P., S. E. ERULKAR, A. P. SOMLYO, AND N. K. GON-tamate, GABA and taurine high-affinity uptake into astrocytes in
TAS. Calcium-containing structures in vertebrate glial cells. Ultra- primary culture. Brain Res. 548: 215–221, 1991.structural and microprobe analysis. J. Cell Biol. 64: 322–330, 1975. 165. HART, I. K., W. D. RICHARDSON, S. R. BOLSOVER, AND M. C.
144. GASQUE, P., P. CHAN, M. FONTAINE, A. ISHENKO, M. LAMACZ, RAFF. PDGF and intracellular signaling in the timing of oligoden-O. GOTZE, AND B. P. MORGAN. Identification and characterization drocyte differentiation. J. Cell Biol. 109: 3411–3417, 1989.of the complement C5a anaphylatoxin receptor on human astro- 166. HASSINGER, T. D., P. B. ATKINSON, G. J. STRECKER, L. R.cytes. J. Immunol. 155: 4882–4889, 1995. WHALEN, F. E. DUDEK, A. H. KOSSEL, AND S. B. KATER. Evidence
145. GEHRMANN, J., Y. MATSUMOTO, AND G. W. KREUTZBERG. Mi- for glutamate-mediated activation of hippocampal neurons by glialcroglia: intrinsic immuneffector cell of the brain. Brain Res. Rev. calcium waves. J. Neurobiol. 28: 159–170, 1995.20: 269–287, 1995. 167. HASSINGER, T. D., P. B. GUTHRIE, P. B. ATKINSON, M. V. L. BEN-
146. GEIGER, J. R., T. MELCHER, D. S. KOH, B. SAKMANN, P. H. SEE- NETT, AND S. B. KATER. An extracellular signaling component inBURG, P. JONAS, AND H. MONYER. Relative abundance of subunit propagation of astrocytic calcium waves. Proc. Natl. Acad. Sci.mRNAs determines gating and Ca2/ permeability of AMPA recep- USA 93: 13268–13273, 1997.tors in principal neurons and interneurons in rat CNS. Neuron 15: 168. HATTON, G. I., R. J. BICKNELL, J. HOYLAND, R. BUNTING, AND193–204, 1995. W. T. MASON. Arginine vasopressin mobilises intracellular calcium
147. GHOSH, A., AND M. E. GREENBERG. Calcium signaling in neurons: via V1-receptor activation in astrocytes (pituicytes) cultured frommolecular mechanisms and cellular consequences. Science 268: adult rat neural lobes. Brain Res. 588: 75–83, 1992.239–247, 1995. 169. HAUN, S. E., E. J. MURPHY, C. M. BATES, AND L. A. HARROCKS.
148. GIAID, A., S. J. GIBSON, M. T. HERRERO, S. GENTLEMAN, S. Extracellular calcium is a mediator of astroglial injury during com-LEGON, M. YANAGISAWA, T. MASAKI, N. B. N. IBRAHIM, G. W. bined glucose-oxygen deprivation. Brain Res. 593: 45–50, 1992.ROBERTS, M. L. ROSSI, AND J. M. POLAK. Topographical localiza- 170. HAUSER, K. F., A. STEINEMARTIN, M. P. MATTSON, R. P. ELDE,tion of endothelin mRNA and peptide immunoreactivity in neu- S. E. RYAN, AND C. C. GODLESKE. m-Opioid receptor induced Ca2/
rones of the human brain. Histochemistry 95: 303–314, 1991. mobilization and astroglial development: morphine inhibits DNA149. GIANNINI, G., E. CLEMENTI, R. CECI, G. MARZIALI, AND V. SOR- synthesis and stimulates cellular hypertrophy through a Ca2/ de-
RENTINO. Expression of ryanodine receptor-Ca2/ channel that is pendent mechanism. Brain Res. 720: 191–203, 1996.regulated by TGF-b. Science 257: 91–94, 1992. 171. HE, M., D. G. HOWE, AND K. D. MCCARTHY. Oligodendroglial signal
150. GIAUME, C., AND K. D. MCCARTHY. Control of gap-junctional com- transduction systems are regulated by neuronal contact. J. Neuro-munication in astrocytic networks. Trends Neurosci. 19: 319–325, chem. 67: 1491–1499, 1996.1996. 172. HEATH, M. J., M. D. WOMACK, AND A. B. MACDERMOTT. Sub-
151. GIMPL, G., F. KIRCHHOFF, R. E. LANG, AND H. KETTENMANN. stance P elevates intracellular calcium in both neurons and glialIdentification of neuropeptide Y receptors in cultured astrocytes cells from the dorsal horn of the spinal cord. J. Neurophysiol. 72:from neonatal rat brain. J. Neurosci. Res. 34: 198–205, 1993. 1192–1198, 1994.
152. GIMPL, G., W. WALZ, C. OHLEMEYER, AND H. KETTENMANN. 173. HEDLEY, L., S. B. PHAGOO, AND I. F. JAMES. Measurement ofBradykinin receptors in cultured astrocytes from neonatal rat brain intracellular calcium in cell populations loaded with aequorin: neu-are linked to physiological responses. Neurosci. Lett. 144: 139–142, rokinin 1 responses in U373MG cells. Anal. Biochem. 236: 270–1992. 274, 1996.
153. GIUFFRIDA, R., M. BELLOMO, G. POLIZZI, AND L. S. MALATINO. 174. HELDIN, C. H. Structural and functional studies on platelet-derivedgrowth factor. EMBO J. 11: 4251–4260, 1992.Ischemia-induced chnages in the immunoreactivity for endothelin
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 135
175. HILDEBRANDT, J. P., AND P. HILDEBRANDT. Lysophosphatidic 198. ITO, S., K. SUGAMA, N. INAGAKI, H. FUKUI, H. GILES, H. WADA,AND O. HAYAISHI. Type-1 and type-2 astrocytes are distinct targetsacid depletes intracellular calcium stores different from those me-
diating capacitative calcium entry in C6 rat glioma cells. Glia 19: for prostaglandins D2, E2, and F2alpha. Glia 6: 67–74, 1992.67–73, 1997. 199. JABS, R., F. KIRCHHOFF, H. KETTENMANN, AND C.
STEINHAUSER. Kainate activates Ca2/-permeable glutamate re-176. HO, C., J. HICKS, AND M. W. SALTER. A novel P2-purinoceptorexpressed by a subpopulation of astrocytes from the dorsal spinal ceptors and blocks voltage-gated K/ currents in glial cells of mouse
hippocampal slices. Pflu¨gers Arch. 426: 310–319, 1994.cord of the rat. Br. J. Pharmacol. 116: 2909–2918, 1995.
177. HOFMANN, F., M. BIEL, AND V. FLOCKERZI. Molecular basis for 200. JAHROMI, B. S., R. ROBITAILLE, AND M. P. CHARLTON. Transmit-ter release increases intracellular calcium in perisynaptic SchwannCa2/ channel diversity. Annu. Rev. Neurosci. 17: 399–418, 1994.cells in situ. Neuron 8: 1069–1077, 1992.178. HOLGADO, A., AND L. BEAUGE. The Na/-Ca2/ exchange system
in rat glial cells in culture: activation by external monovalent cat- 201. JAISWAL, N., E. A. TALLANT, D. I. DIZ, M. C. KHOSLA, AND C. M.FERRARIO. Subtype 2 angiotensin receptors mediate prostaglandinions. Glia 14: 77–86, 1995.synthesis in human astrocytes. Hypertension 17: 1115–1120, 1991.179. HOLLMANN, M., M. HARTLEY, AND S. HEINEMANN. Ca2/ perme-
ability of KA-AMPA-gated glutamate receptor channels depends on 202. JANIGRO, D., R. WENDER, G. RANSOM, D. L. TINKLEPAUGH,AND H. R. WINN. Adenosine induced release of nitric oxide fromsubunit composition. Science 252: 851–853, 1991.cortical astrocytes. Neuroreport 7: 1640–1644, 1996.180. HOLLMANN, M., AND S. HEINEMANN. Cloned glutamate receptors.
Annu. Rev. Neurosci. 17: 31–108, 1994. 203. JENSEN, A. M., AND S. Y. CHIU. Fluorescence measurement ofchanges in intracellular calcium induced by excitatory amino acids181. HOLTZCLAW, L. A., V. GALLO, AND J. T. RUSSELL. AMPA recep-in cultured cortical astrocytes. J. Neurosci. 10: 1165–1175, 1990.tors shape Ca2/ responses in cortical oligodendrocyte progenitors
and CG-4 cells. J. Neurosci. Res. 42: 124–130, 1995. 204. JENSEN, A. M., AND S. Y. CHIU. Differential intracellular calciumresponses to glutamate in type 1 and type 2 cultured brain astro-182. HOLZWARTH, J. A., S. J. GIBBONS, J. R. BRORSON, L. H. PHIL-cytes. J. Neurosci. 11: 1674–1684, 1991.IPSON, AND R. J. MILLER. Glutamate receptor agonists stimulate
diverse calcium responses in different types of cultured rat cortical 205. JENSEN, A. M., AND S. Y. CHIU. Expression of glutamate receptorgenes in white matter: developing and adult rat optic nerve. J.glial cells. J. Neurosci. 14: 1879–1891, 1994.Neurosci. 13: 1664–1675, 1993.183. HOLZWARTH, J. A., S. R. GLAUM, AND R. J. MILLER. Activation of
endothelin receptors by sarafotoxin regulates Ca2/ homeostasis in 206. JOHNSTON, G. A. GABAC receptors. Prog. Brain Res. 100: 61–65,1994.cerebellar astrocytes. Glia 5: 239–250, 1992.
184. HOSLI, E., AND L. HOSLI. Evidence for the existence of alpha- 207. JONAS, P., AND N. BURNASHEV. Molecular mechanisms control-ling calcium entry through AMPA-type glutamate receptor chan-and beta-adrenoceptors on neurones and glial cells of cultured rat
central nervous system—an autoradiographic study. Neuroscience nels. Neuron 15: 987–990, 1995.7: 2873–2881, 1982. 208. JONES, C. R., C. R. HILEY, J. T. PELTON, AND M. MOHR. Autora-
diographic visualization of the binding sites for [125I]endothelin in185. HOSLI, E., AND L. HOSLI. Autoradiographic localization of bindingsites for muscarinic and nicotinic agonists and antagonists on cul- rat and human brain. Neurosci. Lett. 97: 276–279, 1989.tured astrocytes. Exp. Brain Res. 71: 450–454, 1988. 209. JOU, M. J., T. I. PENG, AND S. S. SHEU. Histamine induced oscilla-
tions of mitochondrial free Ca2/ concentration in single cultured186. HOSLI, E., AND L. HOSLI. Autoradiographic evidence for endothelinreceptors on astrocytes of rat cerebellum, brainstem and spinal rat brain astrocytes. J. Physiol. (Lond.) 497: 299–308, 1996.cord. Neurosci. Lett. 129: 55–58, 1991. 210. JURZAK, M., A. R. MULLER, AND R. GERSTBERGER. Characteriza-
tion of vasopressin receptors in cultured cells derived from the187. HOSLI, L., E. HOSLI, U. SCHNEIDER, AND W. WIGET. Evidencefor the existence of histamine H1- and H2-receptors on astrocytes region of rat brain circumventricular organs. Neuroscience 65:
1145–1159, 1995.of cultured rat central nervous system. Neurosci. Lett. 48: 287–291, 1984. 211. JURZAK, M., A. R. MULLER, AND R. GERSTBERGER. AVP-frag-
ment peptides induce Ca2/ transients in cells cultured from rat188. HOTH, M., AND R. PENNER. Calcium-release-activated calcium cur-rent in rat mast cells. J. Physiol. (Lond.) 465: 359–386, 1993. circumventricular organs. Brain Res. 673: 349–355, 1995.
212. KANAI, Y., C. P. SMITH, AND M. A. HEDIGER. The elusive trans-189. HOYER, D., D. E. CLARKE, J. R. FOZARD, P. R. HARTIG, G. R.MARTIN, E. J. MYLECHARANE, P. R. SAXENA, AND P. P. HUM- porters with a high affinity for glutamate. Trends Neurosci. 16:
365–370, 1993.PHREY. International Union of Pharmacology classification of re-ceptors for 5-hydroxytryptamine (serotonin). Pharmacol. Rev. 46: 213. KASTRITSIS, C. H., AND K. D. MCCARTHY. Oligodendroglial lineage
cells express neuroligand receptors. Glia 8: 106–113, 1993.157–203, 1994.190. HUGUENARD, J. R. Low threshold calcium currents in central ner- 214. KASTRITSIS, C. H., A. K. SALM, AND K. MCCARTHY. Stimulation
of the P2Y purinergic receptor on type 1 astroglia results in inositolvous system. Annu. Rev. Physiol. 58: 329–348, 1996.phosphate formation and calcium mobilization. J. Neurochem. 58:191. IINO, M., S. OZAWA, AND K. TSUZUKI. Permeation of calcium1277–1284, 1992.through excitatory amino acid receptor channels in cultured rat
hippocampal neurones. J. Physiol. (Lond.) 424: 151–165, 1990. 215. KATO, K., G. D. CLARK, N. G. BAZAN, AND C. F. ZORUMSKI. Plate-let-activating factor as a potential retrograde messenger in CA1192. IMOTO, K., H. TAKEMURA, M. KANEKO, AND H. OSHIKA. Lackhippocampal long-term potentiation. Nature 367: 175–179, 1994.of voltage operated calcium channel in rat glioma C6 cells. Res.
Commun. Mol. Pathol. Pharmacol. 93: 249–256, 1996. 216. KAWABATA, S., R. TSUTSUMI, A. KOHARA, T. YAMAGUCHI, S.NAKANISHI, AND M. OKADA. Control of calcium oscillations by193. INAGAKI, N., H. FUKUI, S. ITO, AND H. WADA. Type-2 astrocytesphosphorylation of metabotropic glutamate receptors. Nature 383:show intracellular Ca2/ elevation in response to various neuroac-89–92, 1996.tive substances. Neurosci. Lett. 128: 257–260, 1991.
217. KEICHER, E., K. MAGGIO, M.-L. HERNANDEZ-NICAISE, AND G.194. INAGAKI, N., H. FUKUI, S. ITO, A. YAMATODANI, AND H. WADA.NICAISE. The lacunar glial zone at the periphery of Aplysia giantSingle type-2 astrocytes show multiple independent sites of Ca2/
neuron: volume of extracellular space and total calcium content ofsignaling in response to histamine. Proc. Natl. Acad. Sci. USA 88:gliagrana. Neuroscience 42: 593–601, 1991.4215–4219, 1991.
218. KEICHER, E., K. MAGGIO, M.-L. HERNAHNEZ-NICAISE, AND G.195. INAGAKI, N., H. FUKUI, Y. TAGUCHI, N. P. WANG, A. YAMATO-NICAISE. The abundance of Aplysia gliagrana depends on Ca2/DANI, AND H. WADA. Characterization of histamine H1-receptorsand/or Na/ concentrations in seawater. Glia 5: 131–138, 1992.on astrocytes in primary culture: [3H]mepyramine binding studies.
Eur. J. Pharmacol. 173: 43–51, 1989. 219. KEIRSTEAD, S. A., AND R. F. MILLER. Calcium waves in dissoci-ated retinal glial (Muller) cells are evoked by release of calcium196. INAGAKI, N., AND H. WADA. Histamine and prostanoid receptorsfrom intracellular stores. Glia 14: 14–22, 1995.on glial cells. Glia 11: 102–109, 1994.
220. KETTENMANN, H. Beyond the neuronal circuitry. Trends Neu-197. INOUE, A., M. YANAGISAWA, S. KIMURA, Y. KASUYA, T. MIYAU-rosci. 19: 305–306, 1996.CHI, K. GOTO, AND T. MASAKI. The human endothelin family: three
structurally and pharmacologically distinct isopeptides predicted 221. KETTENMANN, H., K. H. BACKUS, AND M. SCHACHNER. Aspar-tate, glutamate and g-aminobutyric acid depolarize cultured astro-by three separate genes. Proc. Natl. Acad. Sci. USA 86: 2863–2867,
1989. cytes. Neurosci. Lett. 52: 25–29, 1984.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78136
222. KETTENMANN, H., K. H. BACKUS, AND M. SCHACHNER. g-Amino- type-2 astrocytes from neonatal rat cerebral cortex. Neurosci. Lett.
180: 131–134, 1994.butyric acid opens Cl0 channels in cultured astrocytes. Brain Res.
404: 1–9, 1987. 244. KONDOU, H., N. INAGAKI, H. FUKUI, Y. KOYAMA, A. KANAMURA,AND H. WADA. Histamine-induced inositol phosphate accumulation223. KETTENMANN, H., AND B. R. RANSOM. Neuroglia. New York: Ox-
ford Univ. Press, 1995. in type-2 astrocytes. Biochem. Biophys. Res. Commun. 177: 734–738, 1991.224. KHODAKHAH, K., AND D. OGDEN. Functional heterogeneity of
calcium release by inositol trisphosphate in single Purkinje neu- 245. KOSTYUK, P. G., AND A. N. VERKHRATSKY. Calcium Signalling
in the Nervous System. Chichester, UK: Wiley, 1995.rones, cultured cerebellar astrocytes, and peripheral tissues. Proc.
Natl. Acad. Sci. USA 90: 4976–4980, 1993. 246. KREUTZBERG, G. W. Microglia: a sensor for pathological eventsin the CNS. Trends Neurosci. 19: 312–318, 1996.225. KIM, W. T., M. G. RIOULT, AND A. H. CORNELL BELL. Glutamate-
induced calcium signaling in astrocytes. Glia 11: 173–184, 1994. 247. KRIEGLER, S., AND S. Y. CHIU. Calcium signaling of glial cells alongmammalian axons. J. Neurosci. 13: 4229–4245, 1993.226. KIMBALL, B. C., AND M. W. MULHOLLAND. Enteric glia exhibit P2U
receptors that increase cytosolic calcium by a phospholipase C 248. KUFFLER, S. W., J. G. NICHOLLS, AND R. K. ORKAND. Physiologi-dependent mechanism. J. Neurochem. 66: 604–612, 1996. cal properties of glial cells in the central nervous system of am-
phibia. J. Neurophysiol. 29: 768–787, 1966.227. KIMELBERG, H. K., E. ANDERSON, AND H. KETTENMANN. Swell-ing-induced changes in electrophysiological properties of cultured 249. KUFFLER, S. W., AND D. D. POTTER. Glia in the leech central ner-astrocytes and oligodendrocytes. II. Whole-cell currents. Brain vous system: physiological properties and neuron-glia relationship.Res. 529: 262–268, 1990. J. Neurophysiol. 27: 290–320, 1964.
228. KING, B. F., J. T. NEARY, Q. ZHU, S. WANG, M. D. NORENBERG, 250. KUNYSZ, E. A., A. D. MICHEL, R. L. WHITING, AND K. WOODS.AND G. BURNSTOCK. P2 purinoreceptors in rat cortical astrocytes: The human astrocytoma cell line 1321 N1 contains M2-glandularexpression, calcium-imaging and signalling studies. Neuroscience type muscarinic receptors linked to phosphoinositide turnover. Br.
74: 1187–1196, 1996. J. Pharmacol. 96: 271–278, 1989.229. KIRCHHOFF, F., I. BLUMCKE, M. R. CELIO, AND H. KETTEN- 251. LAMACZ, M., M. TONON, F. SMIT-ROUET, C. PATTE, P. GASQUE,
MANN. Patch-clamp analysis of glial cells in brain slices of murine M. FONTAINE, AND H. VAUDRY. The endogeneous bezodiazepineneocortex. Soc. Neurosci. Abstr. 19: 686, 1993. receptor ligand ODN increases cytosolic calcium in cultured rat
astrocytes. Mol. Brain Res. 37: 290–296, 1996.230. KIRCHHOFF, F., AND H. KETTENMANN. GABA triggers a [Ca2/]i
increase in murine precursor cells of oligodendrocyte lineage. Eur. 252. LANDIS, D. M. D. The early reactions of non-neuronal cells to brainJ. Neurosci. 4: 1049–1058, 1992. injury. Annu. Rev. Neurosci. 17: 133–151, 1994.
231. KIRCHHOFF, F., C. MULHARDT, A. PASTOR, C.-M. BECKER, AND 253. LANGLEY, D., AND B. PEARCE. Ryanodine-induced intracellularH. KETTENMANN. Expression of glycine receptor subunits in glial calcium mobilisation in cultured astrocytes. Glia 12: 128–134, 1994.cells of the rat spinal cord. J. Neurochem. 66: 1383–1390, 1996. 254. LAZAREWITCH, J. W., M. KANJE, A. SELLSTROM, AND A. HAM-
232. KIRISCHUK, S., H. KETTENMANN, AND A. VERKHRATSKY. Metab- BERGER. Calcium fluxes in cultured and bulk isolated neuronalotropic receptors involved in calcium signalling in mouse Berg- and glial cells. J. Neurochem. 29: 495–502, 1977.mann glial cell (Abstract). J. Physiol. (Lond.) 493: 46P, 1996. 255. LEITCH, B., B. L. WATKINS, AND M. BURROWS. Distribution of
233. KIRISCHUK, S., H. KETTENMANN, AND A. VERKHRATSKY. Na// acetylcholine receptors in the central nervous system of adult lo-Ca2/ exchanger modulates kainate-triggered Ca2/ signalling in custs. J. Comp. Neurol. 334: 47–58, 1993.Bergmann glial cells in situ. FASEB J. 11: 566–572, 1997. 256. LEREA, L. S., AND K. D. MCCARTHY. Astroglial cells in vitro are
234. KIRISCHUK, S., V. MATIASH, A. KULIK, N. VOITENKO, P. KOS- heterogeneous with respect to expression of the alpha1-adrenergicTYUK, AND A. VERKHRATSKY. Activation of P2-purino-, a1-adreno receptor. Glia 2: 135–147, 1989.and H1-histamine receptors triggers cytoplasmic calcium signalling 257. LEURS, R., M. J. SMIT, AND H. TIMMERMAN. Molecular pharmaco-in cerebellar Purkinje neurones. Neuroscience 73: 643–647, 1996. logical aspects of histamine receptors. Pharmacol. Ther. 66: 413–
235. KIRISCHUK, S., T. MO¨
LLER, N. VOITENKO, H. KETTENMANN, 463, 1995.AND A. VERKHRATSKY. ATP-triggered calcium mobilization in cer- 258. LEV RAM, V., AND M. H. ELLISMAN. Axonal activation-induced cal-ebellar Bergmann glial cells. J. Neurosci. 15: 7861–7871, 1995. cium transients in myelinating Schwann cells, sources, and mecha-
236. KIRISCHUK, S., J. NEUHAUS, A. VERKHRATSKY, AND H. KETTEN- nisms. J. Neurosci. 15: 2628–2637, 1995.MANN. Preferential localization of active mitochondria in process 259. LI, Y. X., J. KEIZER, S. S. STOJILKOVIC, AND J. RINZEL. Ca2/ excit-
ability of the ER membrane: an explanation for IP3-induced Ca2/tips of immature retinal oligodendrocytes. Neuroreport 6: 737–741,oscillations. Am. J. Physiol. 269 (Cell Physiol. 38): C1079–C1092,1995.1995.237. KIRISCHUK, S., J. SCHERER, H. KETTENMANN, AND A. VERK-
260. LIEBERMAN, E. M. Role of glutamate in axon-Schwann cell signal-HRATSKY. Activation of P2-purinoreceptors triggered Ca2/ releaseing in the squid. Ann. NY Acad. Sci. 633: 448–457, 1991.from InsP3-sensitive internal stores in mammalian oligodendro-
cytes. J. Physiol. (Lond.) 483: 41–57, 1995. 261. LIEBERMAN, E. M., N. J. ABBOTT, AND S. HASSAN. Evidence thatglutamate mediates axon-to-Schwann cell signaling in the squid.238. KIRISCHUK, S., J. SCHERER, T. MOLLER, A. VERKHRATSKY, AND
Glia 2: 94–102, 1989.H. KETTENMANN. Subcellular heterogeneity of voltage-gated Ca2/
channels in cells of the oligodendrocyte lineage. Glia 13: 1–12, 262. LIEBERMAN, E. M., AND E. SANZENBACHER. Mechanisms of glu-tamate activation of axon-to-Schwann cell signaling in the squid.1995.Neuroscience 47: 931–939, 1992.239. KIRISCHUK, S., S. TUSCHICK, A. VERKHRATSKY, AND H. KET-
TENMANN. Calcium signalling in mouse Bergmann glial cells medi- 263. LIN, W.-W., J. G. KIANG, AND D.-M. CHUANG. Pharmacologicalcharacterization of endothelin-stimulated phosphoinositide break-ated by a1-adrenoreceptors and H1 histamine receptors. Eur. J.
Neurosci. 8: 1198–1208, 1996. down and cytosolic free Ca2/ rise in rat C6 glioma cells. J. Neurosci.
12: 1077–1085, 1992.240. KIRISCHUK, S., AND A. VERKHRATSKY. Ca2/ recordings from neu-ral cells in acutely isolated cerebellar slices employing differential 264. LIU, H. N., AND G. ALMAZAN. Glutamate induces c-fos protoonco-
gene expression and inhibits proliferation in oligodendrocyte pro-loading of membrane permeant form of calcium indicator fura-2.Pflu
¨gers Arch. 431: 977–983, 1996. genitors: receptor characterization. Eur. J. Neurosci. 7: 2355–2363,
1995.241. KITANAKA, J., AND A. BABA. Effect of dibutyryl cyclic AMP-treat-ment on prostaglandin F2a-stimulated phosphoinositide hydrolysis 265. LLANO, I., J. DREESSEN, M. KANO, AND A. KONNERTH. Intraden-
dritic release of calcium induced by glutamate in cerebellar Pur-in cultured rat astrocytes. Jpn. J. Pharmacol. 58: 189–192, 1992.kinje cells. Neuron 7: 577–583, 1991.242. KNAUS, H. G., T. MOSHAMMER, H. C. KANG, R. P. HAUGLAND,
AND H. GLOSSMANN. A unique fluorescent phenylalkylamine probe 266. LUQUE, J. M., AND J. G. RICHARDS. Expression of NMDA 2B recep-tor subunit mRNA in Bergmann cells. Glia 13: 228–232, 1995.for L-type Ca2/ channels. Coupling of phenylalkylamine receptors
to Ca2/ and dihydropyridine binding sites. J. Biol. Chem. 267: 2179– 267. LYONS, S. A., P. MORELL, AND K. D. MCCARTHY. Schwann cells2189, 1992. exhibit P2Y purinergic receptors that regulate intracellular calcium
and are up-regulated by cyclic AMP analogues. J. Neurochem. 63:243. KONDOU, H., N. INAGAKI, AND H. FUKUI. Formation of inositolphosphates mediated by M3 muscarinic receptors in type-1 and 552–560, 1994.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 137
268. LYONS, S. A., P. MORELL, AND K. D. MCCARTHY. Schwann cell containing only rat Schwann cells and sensory neurones. Brain
Res. 525: 267–274, 1990.ATP-mediated calcium increases in vitro and in situ are dependent292. MO
¨LLER, T., C. NOLTE, R. BURGER, A. VERKHRATSKY, AND H.on contact with neurons. Glia 13: 27–38, 1995.
KETTENMANN. Mechanisms of C5a and C3a complement frag-269. MA, K. C., X. J. NIE, A. HOOG, Y. OLSSEN, AND W. W. ZANG. Reac-ments induced [Ca2/]i signalling in mouse microglia. J. Neurosci.tive astrocytes in viral infections of the human brain express endo-17: 615–624, 1997.thelin-like immunoreactivity. J. Neurol. Sci. 126: 184–192, 1994.
293. MO¨
LLER, T., C. NOLTE, A. VERKHRATSKY, AND H. KETTEN-270. MACCUMBER, M. W., C. A. ROSS, AND S. H. SNYDER. EndothelinMANN. Agonist-induced calcium signalling in cultured mouse mi-in the brain: receptors, mitogenesis, and biosynthesis in glial cells.croglia: intracellular Ca2/ release and capacitative Ca2/ entry (Ab-Proc. Natl. Acad. Sci. USA 87: 2359–2363, 1990.stract). J. Physiol. (Lond.) 493: 81P, 1996.271. MACVICAR, B. A. Voltage-dependent calcium channels in glial cells.
294. MONK, P. N., AND L. J. PARTRIDGE. Characterization of a comple-Science 226: 1345–1347, 1984.ment-fragment-C5a-stimulated calcium-influx mechanism in U937272. MACVICAR, B. A., D. HOCHMAN, M. J. DELAY, AND S. WEISS. Mod-monocytic cells. Biochem. J. 295: 679–684, 1993.ulation of intracellular Ca2/ in cultured astrocytes by influx through
295. MONYER, H., N. BURNASHEV, D. J. LAURIE, B. SAKMANN, ANDvoltage-activated Ca2/ channels. Glia 4: 448–455, 1991.P. H. SEEBURG. Developmental and regional expression in the rat273. MACVICAR, B. A., AND F. W. TSE. Norepinephrine and cyclic adeno-brain and functional properties of four NMDA receptors. Neuronsine 3*:5*-cyclic monophosphate enhance a nifedipine-sensitive cal-12: 529–540, 1994.cium current in cultured rat astrocytes. Glia 1: 359–365, 1988.
296. MOORMAN, S. J., AND R. I. HUME. Omega-conotoxin prevents my-274. MAGOSKI, N. S., AND W. WALZ. Ionic dependence of a P2-purino-elin evoked growth cone collapse in neonatal-rat brainstem neu-ceptor mediated depolarization of cultured astrocytes. J. Neurosci.rons in vitro. J. Neurosci. 13: 4727–4736, 1993.Res. 32: 530–538, 1992.
297. MOORMAN, S. J., AND R. I. HUME. Contact with myelin evokes a275. MANEO, H., H. KIYAMA, AND M. TOHYAMA. Distribution of therelease of calcium from internal stores in neonatal rat oligodendro-substance P receptor (NK-1) receptor in the central nervous sys-cytes in vitro. Glia 10: 202–210, 1994.tem. Mol. Brain Res. 18: 43–58, 1993.
298. MORI, M., M. AIHARA, K. KUME, M. HAMANOUE, S. KOSHAKA,276. MANTYH, P. W., S. D. ROGERS, C. J. ALLEN, M. D. CATTON, M. D.AND T. SHIMIZU. Predominant expression of platelet-activating fac-GHILARDI, L. A. LEVIN, J. E. MAGGIO, AND S. R. VIGNA. b2-Adren-tor receptor in the rat brain microglia. J. Neurosci. 16: 3590–3600,ergic receptors are expressed by glia in vivo in the normal and1996.injured central nervous system in the rat, rabbit, and human. J.
299. MORTON, A. J., AND A. P. DAVENPORT. Cerebellar neurons andNeurosci. 15: 152–164, 1995.glia respond differentially to endothelins and sarafotoxin S6b.277. MARRERO, H., AND R. K. ORKAND. Nerve impulses increase glialBrain Res. 581: 299–306, 1992.intercellular permeability. Glia 16: 285–289, 1996.
300. MU¨
LLER, T., J. GROSCHE, C. OHLEMEYER, AND H. KETTEN-278. MARRIOTT, D. R., AND G. P. WILKIN. Substance P receptors on O-MANN. NMDA-activated currents in Bergmann glial cells. Neurore-2A progenitor cells and type-2 astrocytes in vitro. J. Neurochem.port 4: 671–674, 1993.61: 826–834, 1993.
301. MU¨
LLER, T., T. MO¨
LLER, T. BERGER, J. SCHNITZER, AND H.279. MARRIOTT, D. R., G. P. WILKIN, AND J. N. WOOD. Substance P-KETTENMANN. Calcium entry through kainate receptors and re-induced release of prostaglandins from astrocytes: regional spe-sulting potassium-channel blockade in Bergmann glial cells. Sci-cialisation and correlation with phosphoinositol metabolism. J.ence 256: 1563–1566, 1992.
Neurochem. 56: 259–265, 1991.302. MURPHY, S., B. PEARCE, AND C. MORROW. Astrocytes have both
280. MARSAULT, R., P. VIGNE, J. P. BREITTMAYER, AND C. FRELIN. M1 and M2 muscarinic receptor subtypes. Brain Res. 364: 177–180,Astrocytes are target cells for endothelins and sarafotoxin. J. Neu- 1986.rochem. 54: 2142–2144, 1990. 303. NADAL, A., E. FUENTES, AND P. A. MCNAUGHTON. Albumin stim-
281. MARTIN, F. C., A. C. CHARLES, M. J. SANDERSON, AND J. E. MER- ulates uptake of calcium into subcellular stores in rat cortical astro-RILL. Substance P stimulates IL-1 production by astrocytes via cytes. J. Physiol. (Lond.) 492: 737–750, 1996.intracellular calcium. Brain Res. 599: 13–18, 1992. 304. NADAL, A., E. FUENTES, J. PASTOR, AND P. A. MCNAUGHTON.
282. MATSUDA, T., K. TAKUMA, E. NISHIGUCHI, S. ASANO, H. HASHI- Plasma albumin is a potent trigger of calcium signals and DNAMOTO, J. AZUMA, AND A. BABA. Involvement of Na/-Ca2/ ex- synthesis in cortical astrocytes. Proc. Natl. Acad. Sci. USA 92:changer in reperfusion-induced delayed cell death of cultured rat 1426–1430, 1995.astrocytes. Eur. J. Neurosci. 8: 951–958, 1996. 305. NAKAHATA, N., M. T. ABE, I. MATSUOKA, T. ONO, AND H. NAKAN-
283. MCCARTHY, K. D., AND J. DEVELLIS. Preparation of separate astro- ISHI. Adenosine inhibits histamine-induced phosphoinositide hy-glial and oligodendroglial cell cultures from rat cerebral tissue. J. drolysis mediated via pertussis toxin-sensitive G protein in humanCell Biol. 85: 890–902, 1980. astrocytoma cells. J. Neurochem. 57: 963–969, 1991.
284. MCCARTHY, K. D., K. ENKVIST, AND Y. SHAO. Astroglial adrenergic 306. NAKAHATA, N., AND T. K. HARDEN. Regulation of inositol tris-receptors: expression and function. In: Neuroglia, edited by H. phosphate accumulation by muscarinic cholinergic and H1-hista-Kettenmann and B. R. Ransom. Oxford, UK: Oxford Univ. Press, mine receptors on human astrocytoma cells. Differential induction1995, p. 354–366. of desensitization by agonists. Biochem. J. 241: 337–344, 1987.
285. MCCARTHY, K. D., AND A. K. SALM. Pharmacologically-distinct 307. NAKANISHI, S. Metabotropic glutamate receptors: synaptic trans-subsets of astroglia can be identified by their calcium response to mission, modulation, and plasticity. Neuron 13: 1031–1037, 1994.neuroligands. Neuroscience 41: 325–333, 1991. 308. NAKAYASU, H., H. KIMURA, AND K. KURIYAMA. Cerebral GABAA
286. MCDONOUGH, P. M., J. H. EUBANKS, AND J. H. BROWN. Desensiti- and GABAB receptors. Structure and function. Ann. NY Acad. Sci.zation and recovery of muscarinic and histaminergic Ca2/ mobiliza- 757: 516–527, 1995.tion in 1321N1 astrocytoma cells. Biochem. J. 249: 135–141, 1988. 309. NEARY, J. T., C. VAN BREEMEN, E. FORSTER, L. O. NOREN-
287. MEISSNER, G. Ryanodine receptor/Ca2/ release channels and their BERG, AND M. D. NORENBERG. ATP stimulates calcium influx inregulation by endogeneous effectors. Annu. Rev. Physiol. 56: 485– primary astrocyte cultures. Biochem. Biophys. Res. Commun. 157:508, 1994. 1410–1416, 1988.
288. MEUCCI, O., A. FATATIS, J. A. HOLZWARTH, AND R. J. MILLER. 310. NEDERGAARD, M. Direct signaling from astrocytes to neurons inDevelopmental regulation of the toxin sensitivity of Ca2/-permeable cultures of mammalian brain cells. Science 263: 1768–1771, 1994.AMPA receptors in cortical glia. J. Neurosci. 16: 519–530, 1996. 311. NEWMAN, E. A. Voltage-dependent calcium and potassium chan-
289. MILANI, D., L. FACCI, D. GUIDOLIN, A. LEON, AND S. D. SKAPER. nels in retinal glial cells. Nature 317: 809–811, 1985.Activation of polyphosphoinositide metabolism as a signal-trans- 312. NEWMAN, E. A., AND K. R. ZAHS. Calcium waves in retinal glialducing system coupled to excitatory amino acid receptors in astro- cells. Science 275: 844–847, 1997.glial cells. Glia 2: 161–169, 1989. 313. NICHOLSON, C., G. T. BRUGGENCATE, H. STOCKLE, AND R.
290. MILLER, R. J. The control of neuronal Ca2/ homeostasis. Prog. STEINBERG. Calcium and potassium changes in extracellular mi-Neurobiol. 37: 255–285, 1991. croenvironment of cat cerebellar cortex. J. Neurophysiol. 41: 1026–
1039, 1978.291. MITHEN, F. A., AND R. BIRCHEM. Effects of A23187 in cultures
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78138
314. NILSSON, M., P. S. ERIKSSON, L. RONNBACK, AND E. HANSSON. 335. PEAKMAN, M. C., AND S. J. HILL. Adenosine A1 receptor-mediatedchanges in basal and histamine-stimulated levels of intracellularGABA induces Ca2/ transients in astrocytes. Neuroscience 54: 605–
614, 1993. calcium in primary rat astrocytes. Br. J. Pharmacol. 115: 801–810,1995.315. NILSSON, M., E. HANSSON, AND L. RONNBACK. Adrenergic and
5-HT2 receptors on the same astroglial cell. A microspectrofluori- 336. PEARCE, B., J. ALBRECHT, C. MORROW, AND S. MURPHY.Astrocyte glutamate receptor activation promotes inositol phos-metric study on cytosolic Ca2/ responses in single cells in primary
culture. Brain Res. 63: 33–41, 1991. pholipid turnover and calcium flux. Neurosci. Lett. 72: 335–340,1986.316. NILSSON, M., E. HANSSON, AND L. RONNBACK. Heterogeneity
among astroglial cells with respect to 5-HT-evoked cytosolic Ca2/ 337. PEARCE, B., M. CAMBRAY DEAKIN, C. MORROW, J. GRIMBLE,AND S. MURPHY. Activation of muscarinic and of a1-adrenergicresponses. A microspectrofluorimetric study on single cells in pri-
mary culture. Life Sci. 49: 1339–1350, 1991. receptors on astrocytes results in the accumulation of inositol phos-phates. J. Neurochem. 45: 1534–1540, 1985.317. NILSSON, M., E. HANSSON, AND L. RONNBACK. Agonist-evoked
Ca2/ transients in primary astroglial cultures—modulatory effects 338. PEARCE, B., AND D. LANGLEY. Purine- and pyrimidine-stimulatedphosphoinositide breakdown and intracellular calcium mobilisa-of valproic acid. Glia 5: 201–209, 1992.tion in astrocytes. Brain Res. 660: 329–332, 1994.318. NOLTE, C., T. MO
¨LLER, T. WALTER, AND H. KETTENMANN. Com-
plement C5a controls motility of murine microglial cells in vitro 339. PEARCE, B., C. MORROW, AND S. MURPHY. Receptor-mediatedinositol phospholipid hydrolysis in astrocytes. Eur. J. Pharmacol.via activation of an inhibitory G-protein and the rearrangement of
the actin cytoskeleton. Neuroscience 73: 1091–1107, 1996. 121: 231–243, 1986.340. PEARCE, B., S. MURPHY, J. JEREMY, C. MORROW, AND P. DAN-319. NORGAUER, J., G. DOBOS, E. KOWNATZKI, C. DAHINDEN, R.
BURGER, R. KUPPER, AND P. GIERSCHIK. Complement fragment DONA. ATP-evoked Ca2/ mobilisation and prostanoid release fromastrocytes: P2-purinergic receptors linked to phosphoinositide hy-C3a stimulates Ca2/ influx in neutrophils via a pertussis-toxin-sensi-
tive G protein. Eur. J. Biochem. 217: 289–294, 1993. drolysis. J. Neurochem. 52: 971–977, 1989.341. PENDE, M., T. L. FISHER, P. B. SIMPSON, J. T. RUSSELL, J.320. NORONHA-BLOB, L., C. RICHARD, AND D. C. U’PRICHARD. Cal-
cium mobilization by muscarinic receptors in human astrocytoma BLENIS, AND V. GALLO. Neurotransmitter- and growth factor-in-duced cAMP response element binding protein phosphorylation incells: measurements with quin 2. Biochem. Biophys. Res. Commun.
147: 182–188, 1987. glial cell progenitors: role of calcium ions, protein kinase C, andmitogen-activated protein kinase/ribosomal S6 kinase pathway. J.321. NUNEZ, L., AND J. GARCIA-SANCHO. Two different constituentsNeurosci. 17: 1291–1301, 1997.of plasma increase cytosolic calcium selectively in neurones or glia
of primary rat cerebellar cultures. J. Physiol. (Lond.) 490: 577– 342. PENDE, M., L. A. HOLTZCLAW, J. L. CURTIS, J. T. RUSSELL, AND
V. GALLO. Glutamate regulates intracellular calcium and gene ex-583, 1996.pression in oligodendrocyte progenitors through the activation of322. O’CONNOR, E. R., AND H. K. KIMELBERG. Role of calcium inDL-alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid re-astrocyte volume regulation and in release of ions and amino acids.ceptors. Proc. Natl. Acad. Sci. USA 91: 3215–3219, 1994.J. Neurosci. 13: 2638–2650, 1993.
343. PENTREATH, V. W., L. H. SEAL, J. H. MORRISON, AND P. J. MA-323. OERTEL, W. H. Neurotransmitters in the cerebellum. Scientific as-GISTRETTI. Transmitter mediated regulation of energy metabolismpects and clinical relevance. Adv. Neurol. 61: 33–75, 1993.in nervous tissue at the cellular level. Neurochem. Int. 9: 1–10,324. OGATA, T., Y. NAKAMURA, K. TSUJI, T. SHIBATA, K. KATAOKA,1986.AND P. SCHUBERT. Adenosine enhances intracellular Ca2/ mobili-
344. PERRY, D. C., AND S. H. SNYDER. Identification of bradykinin inzation in conjunction with metabotropic glutamate receptor activa-mammalian brain. J. Neurochem. 43: 1072–1080, 1984.tion by t-ACPD in cultured hippocampal astrocytes. Neurosci. Lett.
170: 5–8, 1994. 345. PETERSEN, O. H., C. C. H. PETERSEN, AND H. KASAI. Calciumand hormone action. Annu. Rev. Physiol. 56: 297–319, 1994.325. OGURA, A., K. OZAKI, Y. KUDO, AND T. AMANO. Cytosolic calcium
elevation and cGMP production induced by serotonin in a clonal 346. PETRONI, A., M. SALAMI, M. BLASEVICH, N. PAPINI, G. GA-LELLA, C. COLOMBO, AND C. GALLI. Eicosanoid and inositol phos-cell of glial origin. J. Neurosci. 6: 2489–2494, 1986.phate response to platelet-activating factor (PAF) and to PAF antag-326. OKA, A., M. J. BELLIVEAU, P. A. ROSENBERG, AND J. J. VOLPE.onist in rat astroglial cells. Brain Res. 78: 169–174, 1994.Vulnerability of oligodendroglia to glutamate: pharmacology, mech-
anisms and prevention. J. Neurosci. 13: 1441–1453, 1993. 347. PEUCHEN, S., J. B. CLARK, AND M. R. DUCHEN. Mechanisms ofintracellular calcium regulation in adult astrocytes. Neuroscience327. ORELLANA, S. A., P. A. SOLSKI, AND J. H. BROWN. Phorbol ester71: 871–883, 1996.inhibits phosphoinositide hydrolysis and calcium mobilization in
cultured astrocytoma cells. J. Biol. Chem. 260: 5236–5239, 1985. 348. PIN, J. P., AND R. DUVOISIN. The metabotropic glutamate recep-tors: structure and functions. Neuropharmacology 34: 1–26, 1995.328. ORKAND, R. K., P. M. ORKAND, AND C. M. TANG. Membrane prop-
erties of neuroglia in the optic nerve of Necturus. J. Exp. Biol. 95: 349. PORTER, J. T., AND K. D. MCCARTHY. Adenosine receptors modu-late [Ca2/]i in hippocampal astrocytes in situ. J. Neurochem. 65:49–59, 1981.1515–1523, 1995.329. PARPURA, V., T. A. BASARSKY, F. LIU, K. JEFTINIJA, S. JEFTIN-
IJA, AND P. G. HAYDON. Glutamate-mediated astrocyte-neuron sig- 350. PORTER, J. T., AND K. D. MCCARTHY. GFAP-positive hippocampalastrocytes in situ respond to glutamatergic neuroligands with in-nalling. Nature 369: 744–747, 1994.creases in [Ca2/]i . Glia 13: 101–112, 1995.330. PARPURA, V., F. LIU, K. V. JEFTINIJA, P. G. HAYDON, AND S. D.
JEFTINIJA. Neuroligand-evoked calcium-dependent release of ex- 351. PORTER, J. T., AND K. D. MCCARTHY. Hippocampal astrocytes insitu respond to glutamate released from synaptic terminals. J. Neu-citatory amino acids from Schwann cells. J. Neurosci. 15: 5831–
5839, 1995. rosci. 16: 5073–5081, 1996.352. POZZAN, T., R. RIZZUTO, P. VOLPE, AND J. MELDOLESI. Molecular331. PASTI, L., G. CARMIGNOTO, P. GIUSTI, AND T. POZZAN. The fre-
quency of [Ca2/]i oscillations in cortical and hippocampal astro- and cellular physiology of intracellular calcium stores. Physiol.
Rev. 74: 595–636, 1994.cytes is modulated by neuronal activity. Soc. Neurosci. Abstr. 22:311, 1996. 353. PRADIER, L., E. HEUILLET, J. P. HUBERT, M. LAVILLE, S. LE
GUERN, AND A. DOBLE. Substance P-evoked calcium mobilization332. PASTI, L., T. POZZAN, AND G. CARMIGNOTO. Long-lasting changesof calcium oscillations in astrocytes. A new form of glutamate- and ionic current activation in the human astrocytoma cell line U
373 MG: pharmacological characterization. J. Neurochem. 61:mediated plasticity. J. Biol. Chem. 270: 15203–15210, 1995.1850–1858, 1993.333. PASTOR, A., A. CHVATAL, E. SYKOVA, AND H. KETTENMANN.
Glycine- and GABA-activated currents in identified glial cells of the 354. PRIVAT, A., M. GIMENEZ-RIBOTTA, AND J. L. RIDET. Morphologyof astrocytes. In: Neuroglia, edited by H. Kettenmann and B. R.developing rat spinal cord slice. Eur. J. Neurosci. 7: 1188–1198,
1995. Ransom. New York: Oxford Univ. Press, 1995, p. 3–22.355. PRUSS, R. M., R. L. AKESON, M. M. RACKE, AND J. L. WILBURN.334. PATNEAU, D. K., P. W. WRIGHT, C. WINTERS, M. L. MAYER, AND
V. GALLO. Glial cells of the oligodendrocyte lineage express both Agonist-activated cobalt uptake identifies divalent cation-perme-able kainate receptors on neurons and glial cells. Neuron 7: 509–kainate- and AMPA-preferring subtypes of glutamate receptor. Neu-
ron 12: 357–371, 1994. 518, 1991.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 139
356. PURO, D. G. A calcium-activated, calcium-permeable ion channel response in individual astroglial cells. Mol. Pharmacol. 44: 247–254, 1993.in human retinal glial cells: modulation by basic fibroblast growth
380. SHAO, Y., AND K. D. MCCARTHY. Regulation of astroglial respon-factor. Brain Res. 548: 329–333, 1991.siveness to neuroligands in primary culture. Neuroscience 55: 991–357. PURO, D. G., J. J. HWANG, O. J. KWON, AND H. M. CHIN. Charac-1001, 1993.terization of an L type calcium channel expressed by human retinal
381. SHAO, Y., AND K. D. MCCARTHY. Plasticity of astrocytes. Glia 11:Muller (glial) cells. Mol. Brain Res. 37: 41–48, 1996.147–155, 1994.358. PURO, D. G., J. P. YUAN, AND N. J. SUCHER. Activation of NMDA
382. SHAO, Y., AND K. D. MCCARTHY. Receptor-mediated calcium sig-receptor channels in human retinal Muller cells inhibits inwardnals in astroglia: multiple receptors, common stores and all-or-rectifying potassium currents. Vis. Neurosci. 13: 319–326, 1996.nothing responses. Cell Calcium 17: 187–196, 1995.359. QUANDT, F. N., AND B. A. MACVICAR. Calcium activated potas-
383. SHAO, Y., AND J. SUTIN. Expression of adrenergic receptors insium channels in cultured astrocytes. Neuroscience 19: 29–41, 1986.individual astrocytes and motor neurones isolated from the adult360. RAIVICH, G., AND W. KREUTZBERG. Pathophysiology of glialrat brain. Glia 6: 108–117, 1992.growth factor receptors. Glia 11: 129–146, 1994.
384. SHIGEMOTO, R., Y. YOKOTA, K. TSUCHIDA, AND S. NAKANISHI.361. RANSOM, B. R., AND S. GOLDRING. Ionic determinants of mem-Cloning and expression of a rat neuromedin K receptor cDNA. J.brane potential of cells presumed to be glia in cerebral cortex ofBiol. Chem. 265: 623–628, 1990.cat. J. Neurophysiol. 36: 855–868, 1973.
385. SHIMOMURA, O., F. N. JOHNSON, AND Y. SAIGA. Extraction, puri-362. REETZ, G., AND G. REISER. [Ca2/]i oscillations induced by bradyki-fication and properties of aequorin, a bioluminiscent protein fromnin in rat glioma cells associated with Ca2/ store-dependent Ca2/
the luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol. 59:influx are controlled by cell volume and by membrane potential.233–239, 1962.Cell Calcium 19: 143–156, 1996.
386. SIEGHART, W. Structure and pharmacology of g-aminobutyric acid363. REIST, N. E., AND S. J. SMITH. Neurally evoked calcium transientsA receptor subtypes. Pharmacol. Rev. 47: 181–234, 1995.in terminal Schwann cells at the neuromuscular junction. Proc.
387. SIMPSON, P. B., R. A. CHALLISS, AND S. R. NAHORSKI. NeuronalNatl. Acad. Sci. USA 89: 7625–7629, 1992.Ca2/ stores: activation and function. Trends Neurosci. 18: 299–306,364. RESINK, T., J. SCOTT-BURDEN, C. BOULANGER, E. WEBER, AND1995.F. R. BUHLER. Internalization of endothelin by cultured human
388. SIMPSON, P. B., AND J. T. RUSSELL. Mitochondria support inositolvascular smooth muscle cells: characterization and physiological1,4,5 trisphosphate medited Ca2/ waves in cultured oligodendro-significance. Mol. Pharmacol. 38: 244–252, 1990.cytes. J. Biol. Chem. 271: 33493–33501, 1996.365. RIGHI, M., O. LETARI, P. SACERDOTE, F. MARANGONI, A. MI-
389. SMITH, S. J. Neural signalling. Neuromodulatory astrocytes. Curr.OZZO, AND S. NICOSIA. Myc-immortalized microglial cells expressBiol. 4: 807–810, 1994.a functional platelet-activating factor receptor. J. Neurochem. 64:
390. SNEYD, J., A. C. CHARLES, AND M. J. SANDERSON. A model for121–129, 1995.the propagation of intercellular calcium waves. Am. J. Physiol. 266366. RITCHIE, T., R. COLE, H. S. KIM, J. DE VELLIS, AND E. P. NOBLE.(Cell Physiol. 35): C293–C302, 1994.Inositol phospholipid hydrolysis in cultured astrocytes and oligo-
391. SOLIVEN, B., M. TAKEDA, T. SHANDY, AND D. J. NELSON. Arachi-dendrocytes. Life Sci. 41: 31–39, 1987.donic acid and its metabolites increase Cai in cultured rat oligoden-367. ROBITAILLE, R. Purinergic receptors and their activation by endog-drocytes. Am. J. Physiol. 264 (Cell Physiol. 33): C632–C640, 1993.enous purines at perisynaptic glial cells of the frog neuromuscular
392. SONTHEIMER, H. Voltage-dependent ion channels in glial cells.junction. J. Neurosci. 15: 7121–7131, 1995.Glia 11: 156–172, 1994.
368. ROBITAILLE, R., M.-J. BORQUE, AND S. VANDAELE. Localization 393. SONTHEIMER, H., J. A. BLACK, AND S. G. WAXMAN. Voltage-gatedof L-type Ca2/ channels at perisynaptic glial cells of the frog neuro- Na/ channels in glia: properties and possible functions. Trendsmuscular junction. J. Neurosci. 16: 148–158, 1996.
Neurosci. 19: 325–331, 1996.369. ROGERS, M., AND J. A. DANI. Comparison of quantitative calcium 394. SONTHEIMER, H., H. KETTENMANN, K. H. BACKUS, AND M.
flux through NMDA, ATP, and ACh receptor channels. Biophys. J. SCHACHNER. Glutamate opens Na//K/ channels in cultured astro-68: 501–506, 1995. cytes. Glia 1: 328–336, 1988.
370. ROMANO, C., M. A. SESMA, C. T. MCDONALD, K. O’MALLEY, A. N. 395. SORRENTINO, V., AND P. VOLPE. Ryanodine receptors: how many,VAN DEN POL, AND J. W. OLNEY. Distribution of metabotropic why and where? Trends Pharmacol. Sci. 14: 98–103, 1993.glutamate receptor mGluR5 immunoreactivity in rat brain. J. Comp. 396. STANIMIROVIC, D. B., R. BALL, G. MEALING, P. MORLEY, ANDNeurol. 355: 455–469, 1995. J. P. DURKIN. The role of intracellular calcium and protein kinase
371. ROSE, C. R., C. LOHR, AND J. W. DEITMER. Activity induced Ca2/C in endothelin-stimulated proliferation of rat type I astrocytes.
transients in nerve and glial cells in the leech CNS. Neuroreport 6: Glia 15: 119–130, 1995.642–644, 1995. 397. STEINHA
¨USER, C., AND V. GALLO. News on glutamate receptors
372. RUBANYI, G. M., AND M. A. POLOKOFF. Endothelins: molecular in glial cells. Trends Neurosci. 19: 339–345, 1996.biology, biochemistry, pharmacology, physiology, and pathophysi- 398. STEINHA
¨USER, C., R. JABS, AND H. KETTENMANN. Properties of
ology. Pharmacol. Rev. 46: 325–415, 1994. GABA and glutamate responses in identified glial cells of the mouse373. SALM, A. K., AND K. D. MCCARTHY. Norepinephrine-evoked cal- hippocampal slice. Hippocampus 4: 19–35, 1994.
cium transients in cultured cerebral type 1 astroglia. Glia 3: 529– 399. STEPHENS, G. J., A. J. CHOLEWINSKI, G. P. WILKIN, AND M. B.538, 1990. DJAMGOZ. Calcium-mobilizing and electrophysiological effects of
374. SALTER, M. W., AND J. L. HICKS. ATP-evoked increases in intracel- bradykinin on cortical astrocyte subtypes in culture. Glia 9: 269–lular calcium in neurons and glia from the dorsal spinal cord. J. 279, 1993.Neurosci. 14: 1563–1575, 1994. 400. STEPHENS, G. J., D. R. MARRIOTT, M. B. A. DJAMGOZ, AND G. P.
375. SALTER, M. W., AND J. L. HICKS. ATP causes release of intracellu- WILKIN. Electrophysiological and biochemical evidence for brady-lar Ca2/ via the phospholipase Cb/IP3 pathway in astrocytes from kinin receptors on cultured cortical oligodendrocytes. Neurosci.the dorsal spinal cord. J. Neurosci. 15: 2961–2971, 1995. Lett. 153: 223–226, 1993.
376. SCHLUE, W. R., AND P. HOCHSTRATE. Ca2/ influx into leech neu- 401. STIENE MARTIN, A., M. P. MATTSON, AND K. F. HAUSER. Opiatesropile glial cells mediated by nicotinic acetylcholine receptors. Glia selectively increase intracellular calcium in developing type-1 astro-15: 43–53, 1995. cytes: role of calcium in morphine-induced morphologic differentia-
377. SCOLDING, N. J., B. P. MORGAN, A. K. CAMBELL, AND D. A. S. tion. Brain Res. 76: 189–196, 1993.COMPSTON. The role of calcium in rat oligodendrocyte injury and 402. SUGINO, H., A. OGURA, Y. KUDO, AND T. AMANO. Intracellularrepair. Neurosci. Lett. 135: 199–211, 1992. Ca2/ elevation induced by a neurotransmitter in a glial cell clone.
378. SEIFERT, G., AND C. STEINHA¨USER. Glial cells in the mouse hippo- Brain Res. 322: 127–130, 1984.
campus express AMPA receptors with an intermediate Ca2/ perme- 403. SULLIVAN, A. M., AND A. J. MORTON. Endothelins induce FOS ex-ability. Eur. J. Neurosci. 7: 1872–1881, 1995. pression in neurones and glia in organotypic cultures of rat cerebel-
lum. J. Neurochem. 67: 1409–1418, 1996.379. SHAO, Y., AND K. D. MCCARTHY. Quantitative relationship betweena1-adrenergic receptor density and the receptor-mediated calcium 404. SUPATTAPONE, S., A. W. SIMPSON, AND C. C. ASHLEY. Free cal-
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

VERKHRATSKY, ORKAND, AND KETTENMANN Volume 78140
cium rise and mitogenesis in glial cells caused by endothelin. Bio- glutamate receptor mGluR5 subcellular distribution and develop-mental expression in hypothalamus. J. Comp. Neurol. 362: 134–chem. Biophys. Res. Commun. 165: 1115–1122, 1989.150, 1995.405. SYKOVA, E. Ionic and Volume Changes in the Microenvironment
of Nerve and Receptor Cells. Heidelberg, Germany: Springer-Verlag, 427. VENANCE, L., D. PIOMELLI, J. GLOWINSKI, AND C. GIAUME. Inhi-bition by anandamide of gap junctions and intercellular calcium1992.signalling in striatal astrocytes. Nature 376: 590–594, 1995.406. SZUCHET, S. The morphology and ultrastructure of oligodendro-
cytes and their functional implications. In: Neuroglia, edited by H. 428. VENANCE, L., N. STELLA, J. GLOWINSKI, AND C. GIAUME. Mecha-nism involved in initiation and propagation of receptor-inducedKettenmann and B. R. Ransom. New York: Oxford Univ. Press,
1995, p. 23–43. intercellular calcium signalling in cultured rat astrocytes. J. Neu-
rosci. 17: 1981–1992, 1997.407. TAKEDA, M., D. J. NELSON, AND B. SOLIVEN. Calcium signalingin cultured rat oligodendrocytes. Glia 14: 225–236, 1995. 429. VERKHRATSKY, A., AND H. KETTENMANN. Calcium signalling in
glial cells. Trends Neurosci. 19: 346–352, 1996.408. TAKEUCHI, N. Some properties of the conductance changes at theend-plate membrane during the action of acetylcholine. J. Physiol. 430. VERKHRATSKY, A., AND A. SHMIGOL. Calcium-induced calcium
release in neurones. Cell Calcium 19: 1–14, 1996.(Lond.) 167: 128–140, 1963.409. TAKUMA, K., T. MATSUDA, S. ASANO, AND A. BABA. Intracellular 431. VERKHRATSKY, A. N., J. TROTTER, AND H. KETTENMANN. Cul-
tured glial precursor cells from mouse cortex express two typesascorbic acid inhibits the Na/-Ca2/ exchanger in cultured rat astro-cytes. J. Neurochem. 64: 1536–1540, 1995. of calcium currents. Neurosci. Lett. 112: 194–198, 1990.
432. VERNADAKIS, A. Glia-neuron intercommunications and synaptic410. TAKUMA, K., T. MATSUDA, H. HASHIMOTO, S. ASANO, AND A.BABA. Cultured rat astrocytes possess Na/-Ca2/ exchanger. Glia plasticity. Prog. Neurobiol. 49: 185–214, 1996.12: 336–342, 1994. 433. VERNINO, S., M. ROGERS, K. A. RADCLIFFE, AND J. A. DANI.
Quantitative measurement of calcium flux through muscle and neu-411. TAKUMA, K., T. MATSUDA, H. HASHIMOTO, J. KITANAKA, S.ASANO, Y. KISHIDA, AND A. BABA. Role of Na/-Ca2/ exchanger ronal nicotinic acetylcholine receptors. J. Neurosci. 14: 5514–5524,
1994.in agonist-induced Ca2/ signaling in cultured rat astrocytes. J. Neu-
rochem. 67: 1840–1845, 1996. 434. VILLEGAS, J. Cholinergic systems in axon-Schwann cell interac-tions. Trends Neurosci. 1: 66–68, 1978.412. TALLANT, E. A., N. JAISWAL, D. I. DIZ, AND C. M. FERRARIO. Hu-
man astrocytes contain two distinct angiotensin receptor subtypes. 435. VON BLANKENFELD, G., AND H. KETTENMANN. Glutamate andHypertension 18: 32–39, 1991. GABA receptors in vertebrate glial cells. Mol. Neurobiol. 5: 31–43,
1991.413. TANABE, Y., A. NOMURA, M. MASU, R. SHIGEMOTO, N. MIZUNO,AND S. NAKANISHI. Signal transduction, pharmacological proper- 436. VON BLANKENFELD, G., A. VERKHRATSKY, AND H. KETTEN-ties, and expression patterns of two rat metabotropic glutamate MANN. Ca2/ channel expression in the oligodendrocyte lineage.receptors, mGluR3 and mGluR4. J. Neurosci. 13: 1372–1378, 1993. Eur. J. Neurosci. 4: 1035–1048, 1992.
414. TANG, C. M., AND R. K. ORKAND. Glutamate depolarization of glial 437. WAKAKURA, M., AND N. YAMAMOTO. Cytosolic calcium transientcells in Necturus optic nerve. Neurosci. Lett. 63: 300–304, 1986. increase through the AMPA/kainate receptor in cultured Muller
cells. Vision Res. 34: 1105–1109, 1994.415. TATSUNO, I., P. E. GOTTSCHALL, AND A. ARIMURA. Specific bind-ing sites for pituitary adenylate cyclase activating polypeptide (PA- 438. WALZ, W., G. GIMPL, C. OHLEMEYER, AND H. KETTENMANN.CAP) in rat cultured astrocytes: molecular identification and inter- Extracellular ATP-induced currents in astrocytes: involvement ofaction with vasoactive intestinal peptide (VIP). Peptides 12: 617– a cation channel. J. Neurosci. Res. 38: 12–18, 1994.621, 1991. 439. WALZ, W., S. ILSCHNER, C. OHLEMEYER, R. BANATI, AND H.
416. TOESCU, E. C., AND A. VERKHRATSKY. Dynamical relationship KETTENMANN. Extracellular ATP activates a cation conductancebetween intracellular Ca2/ stores and stores-operated Ca2/ entry and a K/ conductance in cultured microglial cells from mousepathway in mouse brain microglia: long-term activation of capacita- brain. J. Neurosci. 13: 4403–4411, 1993.tive Ca2/ entry (Abstract). J. Physiol. (Lond.) 501P: 21P, 1997. 440. WALZ, W., AND B. MACVICAR. Electrophysiological properties of
417. TSIEN, R. Y. Fluorescent indicators of ion concentrations. Methods glial cells: comparison of brain slices with primary cultures. Brain
Cell Biol. 30: 127–156, 1989. Res. 443: 321–324, 1988.418. TUSCHICK, S., S. KIRISCHUK, F. KIRCHHOFF, L. LIEFELD, M. 441. WANG, D., J. R. MARTENS, P. POSNER, C. SUMNERS, AND C. H.
PAUL, A. VERKHRATSKY, AND H. KETTENMANN. Bergmann glial GELBAND. Angiotensin II regulation of intracellular calcium inastroglia cultured from rat hypothalamus and brain stem. J. Neuro-cells in situ express endothelinB receptors linked to cytoplasmicchem. 67: 996–1004, 1996.calcium signals. Cell Calcium 21: 408–419, 1997.
442. WATSON, S., AND D. GIRDLESTONE. Receptor nomenclature sup-419. TUSCHICK, S., S. KIRISCHUK, A. VERKHRATSKY, AND H. KET-plement. Trends Pharmacol. Sci. 17, Suppl.: 54–56, 1996.TENMANN. Mouse cerebellar Bergmann glial cells express ETB
endothelin receptors linked to the intracellular Ca2/ release from 443. WETTSTEIN, J. G., B. EARLEY, AND J. L. JUNIEN. Central nervoussystem pharmacology of neuropeptide Y. Pharmacol. Ther. 65: 397–InsP3-sensitive intracellular stores (Abstract). J. Physiol. (Lond.)
493: 72P–73P, 1996. 414, 1995.444. WIENRICH, M., AND H. KETTENMANN. Activation of substance420. TZENG, S.-F., G. E. DEIBLER, AND G. H. DEVRIES. Exogenous my-
elin basic protein promotes oligodendrocyte death via increased P receptors leads to membrane potential responses in culturedastrocytes. Glia 2: 155–160, 1989.calcium influx. J. Neurosci. Res. 42: 768–774, 1995.
421. UCHIHORI, Y., AND D. G. PURO. Glutamate as a neuron-to-glial 445. WILLARD, A. L. Excitatory and neurotoxic actions of platelet-acti-vating factor on rat myenteric neurones in cell culture. Ann. NYsignal for mitogenesis: role of glial N-methyl-D-aspartate receptors.
Brain Res. 613: 212–220, 1993. Acad. Sci. 664: 284–292, 1992.446. WOOD, A., M. G. WING, C. D. BENHAM, AND D. A. COMPSTON.422. USOWICZ, M. M., V. GALLO, AND S. G. CULL CANDY. Multiple con-
ductance channels in type-2 cerebellar astrocytes activated by ex- Specific induction of intracellular calcium oscillations by comple-ment membrane attack on oligodendroglia. J. Neurosci. 13: 3319–citatory amino acids. Nature 339: 380–383, 1989.3332, 1993.423. VALERA, S., N. HUSSY, R. J. EVANS, N. ADAMI, R. A. NORTH, A.
SURPRENANT, AND G. BUELL. A new class of ligand-gated ion 447. WOODS, M. J., AND D. C. WILLIAMS. Multiple forms and locationsfor the peripheral-type benzodiazepine receptor. Biochem. Phar-channel defined by P2X receptor for extracellular ATP. Nature 371:
516–519, 1994. macol. 52: 1805–1814, 1996.448. WYLLIE, D. J., AND S. G. CULL CANDY. A comparison of non-424. VANDAELE, S., M. FOSSET, J. P. GALIZZI, AND M. LAZDUNSKI.
Monoclonal antibodies that coimmunoprecipitate the 1,4-dihydro- NMDA receptor channels in type-2 astrocytes and granule cellsfrom rat cerebellum. J. Physiol. (Lond.) 475: 95–114, 1994.pyridine and phenylalkylamine receptors and reveal the Ca2/ chan-
nel structure. Biochemistry 26: 5–9, 1987. 449. WYLLIE, D. J., A. MATHIE, C. J. SYMONDS, AND S. G. CULLCANDY. Activation of glutamate receptors and glutamate uptake425. VAN DEN POL, A. N., S. M. FINKBEINER, AND A. H. CORNELL
BELL. Calcium excitability and oscillations in suprachiasmatic nu- in identified macroglial cells in rat cerebellar cultures. J. Physiol.
(Lond.) 432: 235–258, 1991.cleus neurons and glia in vitro. J. Neurosci. 12: 2648–2664, 1992.426. VAN DEN POL, A. N., C. ROMANO, AND P. GHOSH. Metabotropic 450. YAGODIN, S., L. A. HOLTZCLAW, AND J. T. RUSSELL. Subcellular
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev

January 1998 CALCIUM SIGNALING IN GLIA 141
calcium oscillators and calcium influx support agonist-induced cal- induced by endothelins and sarafotoxin in human astrocytomacells (1321N1): comparison of histamine-induced calcium mobi-cium waves in cultured astrocytes. Mol. Cell. Biochem. 149–150:
137–144, 1995. lization. Naunun-Schmiedebergs Arch. Pharmacol. 346: 51–56,1992.451. YAGODIN, S. V., L. HOLTZCLAW, C. A. SHEPPARD, AND J. T. RUS-
SELL. Nonlinear propagation of agonist-induced cytoplasmic cal- 457. YODER, E. J., H. TAMIR, AND M. H. ELLISMAN. 5-Hydroxytrypta-mine2A receptors on cultured rat Schwann cells. Glia 17: 15–27,cium waves in single astrocytes. J. Neurobiol. 25: 265–280, 1994.
452. YAKEL, J. L., R. A. WARREN, S. M. REPPERT, AND R. A. NORTH. 1996.458. YOKOTA, Y., Y. SASAI, K. TANAKA, T. FUJIWARA, K. TSUCHIDA,Functional expression of adenosine A2b receptor in Xenopus oo-
cytes. Mol. Pharmacol. 43: 277–280, 1993. R. SHIGEMOTO, A. KAKIZUKA, H. OHKUBO, AND S. NAKANISHI.Molecular characterization of a functional cDNA for rat substance453. YAMAMOTO HINO, M., A. MIYAWAKI, H. KAWANO, T. SUGI-
YAMA, T. FURUICHI, M. HASEGAWA, AND K. MIKOSHIBA. Immu- P receptor. J. Biol. Chem. 264: 17649–17652, 1989.459. ZHANG, W., N. SAKAI, H. YAMADA, T. FU, AND Y. NOZAWA. Endo-nohistochemical study of inositol 1,4,5-trisphosphate receptor type
3 in rat central nervous system. Neuroreport 6: 273–276, 1995. thelin 1 induces intracellular calcium rise and inositol-1,4,5-trispho-sphate formation in cultured rat and human glioma cells. Neurosci.454. YAMASHITA, K., M. NIWA, Y. KATAOKA, K. SHIGEMATSU, A.
HIMENO, K. TSUTSUMI, M. NAKANO NAKASHIMA, Y. SAKURAI, Lett. 112: 199–204, 1990.460. ZHANG, W. W., T. BADONIC, A. HOOG, M. H. JIANG, K. C. MA,Y. YAMASHITA, S. SHIBATA, AND K. J. TANIYAMA. Microglia with
an endothelin ETB receptor aggregate in rat hippocampus CA1 sub- X. J. NIE, AND Y. OLSSON. Astrocytes in Alzheimer disease expressimmunoreactivity to the vasoconstrictor endothelin-1. J. Neurol.fields following transient forebrain ischemia. J. Neurochem. 63:
1042–1051, 1994. Sci. 122: 90–96, 1994.461. ZHAO, Z., W. E. CODE, AND L. HERTZ. Dexmedetomidine, a potent455. YANAGISAWA, M., H. KURIHARA, S. KIMURA, Y. TOMOBE, M.
KOBAYASHI, Y. MITSUI, Y. YAZAKI, K. GOTO, AND T. MASAKI. and highly specific alpha2 agonist, evokes cytosolic calcium surgein astrocytes but not in neurons. Neuropharmacology 31: 1077–Endothelin: a novel potent vasoconstrictor peptide produced by
vascular endothelial cells. Nature 332: 411–415, 1988. 1079, 1992.462. ZIFA, E., AND G. FILLION. 5-Hydroxytryptamine receptors. Phar-456. YANAI, K., K. MAEYAMA, N. NAKAHATA, H. NAKANISHI, AND
T. WATANABE. Calcium mobilization and its desensitization macol. Rev. 44: 401–458, 1992.
P21-7/ 9j07$$ja04 01-13-98 12:09:44 pral APS-Phys Rev
Related Documents











