Geographical variations of odour and pollinators, and test for local adaptation by reciprocal transplant of two European Arum species Marion Chartier 1,2 , Laurent P elozuelo 3 , Bruno Buatois 4 , Jean-Marie Bessi ere 4 and Marc Gibernau* ,1 1 Joint Research Unit Ecology of Guiana Forests, CNRS-UMR 8172, Campus agronomique, BP 316, 97379 Kourou cedex, France; 2 Laboratory of Evolution and Biological Diversity, Bât. 4R1, Université Paul Sabatier, 118 route de Narbonne, 31062 Toulouse cedex 9, France; 3 Laboratory of Functional Ecology and Environment, Bât. 4R1, Université Paul Sabatier, 118 route de Narbonne, 31062 Toulouse cedex 9, France; and 4 Center for Functional and Evolutive Ecology, Université Montpellier 2, 1919 route de Mende, 34293 Montpellier, France Summary 1. Interactions between entomophilous plants and their pollinators are one of the major factors shaping the evolution of floral features. As species are distributed in more or less connected populations, they have evolved in a geographical mosaic of co-evolution were the outcome of the plant–pollinator interaction is likely to vary as a result of local adaptations. 2. Arum italicum and Arum maculatum are two species of Araceae which deceive their fly poll- inators by mimicking the odour of their oviposition sites. Whereas A. italicum is known to be pollinated by flies belonging to different families (i.e. opportunist), A. maculatum relies on only two pollinating species of the family Psychodidae throughout its European repartition area (i.e. specialist). 3. The interannual and geographical variations of pollinators and pollinator-attractive odours were described in several populations of the two species over two consecutive years. Further- more, local adaptation to pollinators was tested by transplanting inflorescence-bearing plants between two different sites and by recording the number and composition of the insect fauna trapped inside the inflorescences during anthesis as a measure of a fitness component. 4. Pollinators and pollinator-attractive odours of the two Arum species varied in time and space, but there was no clear odour structure between populations. When transplanted, inflo- rescences of both species trapped the same composition and number of insects as native inflo- rescences at a given site; this indicates that pollinator composition is highly dependent on the local availability of insects. 5. No pattern of local adaptation was found for these two species, but local pollination condi- tions were shown to strongly affect the degree of geographical variations of these interactions. The lack of a clear odour geographical structure might be due to high gene flow or to similar selective pressures exerted by pollinators, and the high interindividual odour variation may be linked to the deceptive strategy adopted by the two plant species. Key-words: deception, diptera, floral scent, geographical mosaic, Psychodidae, sapromyophily, transplant experiment Introduction It is believed that the interactions that evolved between many plants species and their insect pollinators have shaped angiosperm diversification (reviewed by Johnson 2006; Herrera, Castellanos & Medrano 2006). For instance, the observation that plants pollinated by the same group of pollinators often converge in their traits, such as flower colour, odour, size and/or shape, leads to the concept of pollination syndromes (Stebbins 1970; Faegri & Van Der Pijl 1971; Proctor, Leo & Lack 1996; Fenster et al. 2004). Pollinators of a given species can vary among different populations, leading to plant and *Correspondence author. E-mail: [email protected] © 2013 The Authors. Functional Ecology © 2013 British Ecological Society Functional Ecology 2013 doi: 10.1111/1365-2435.12122

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Geographical variations of odour and pollinators, andtest for local adaptation by reciprocal transplant of twoEuropean Arum speciesMarion Chartier1,2, Laurent P�elozuelo3, Bruno Buatois4, Jean-Marie Bessi�ere4
and Marc Gibernau*,1
1Joint Research Unit Ecology of Guiana Forests, CNRS-UMR 8172, Campus agronomique, BP 316, 97379 Kouroucedex, France; 2Laboratory of Evolution and Biological Diversity, Bât. 4R1, Université Paul Sabatier, 118 route deNarbonne, 31062 Toulouse cedex 9, France; 3Laboratory of Functional Ecology and Environment, Bât. 4R1, UniversitéPaul Sabatier, 118 route de Narbonne, 31062 Toulouse cedex 9, France; and 4Center for Functional and EvolutiveEcology, Université Montpellier 2, 1919 route de Mende, 34293 Montpellier, France
Summary
1. Interactions between entomophilous plants and their pollinators are one of the major
factors shaping the evolution of floral features. As species are distributed in more or less
connected populations, they have evolved in a geographical mosaic of co-evolution were the
outcome of the plant–pollinator interaction is likely to vary as a result of local adaptations.
2. Arum italicum and Arum maculatum are two species of Araceae which deceive their fly poll-
inators by mimicking the odour of their oviposition sites. Whereas A. italicum is known to be
pollinated by flies belonging to different families (i.e. opportunist), A. maculatum relies on only
two pollinating species of the family Psychodidae throughout its European repartition area
(i.e. specialist).
3. The interannual and geographical variations of pollinators and pollinator-attractive odours
were described in several populations of the two species over two consecutive years. Further-
more, local adaptation to pollinators was tested by transplanting inflorescence-bearing plants
between two different sites and by recording the number and composition of the insect fauna
trapped inside the inflorescences during anthesis as a measure of a fitness component.
4. Pollinators and pollinator-attractive odours of the two Arum species varied in time and
space, but there was no clear odour structure between populations. When transplanted, inflo-
rescences of both species trapped the same composition and number of insects as native inflo-
rescences at a given site; this indicates that pollinator composition is highly dependent on the
local availability of insects.
5. No pattern of local adaptation was found for these two species, but local pollination condi-
tions were shown to strongly affect the degree of geographical variations of these interactions.
The lack of a clear odour geographical structure might be due to high gene flow or to similar
selective pressures exerted by pollinators, and the high interindividual odour variation may be
linked to the deceptive strategy adopted by the two plant species.
Key-words: deception, diptera, floral scent, geographical mosaic, Psychodidae, sapromyophily,
transplant experiment
Introduction
It is believed that the interactions that evolved between
many plants species and their insect pollinators have
shaped angiosperm diversification (reviewed by Johnson
2006; Herrera, Castellanos & Medrano 2006). For
instance, the observation that plants pollinated by the
same group of pollinators often converge in their traits,
such as flower colour, odour, size and/or shape, leads to
the concept of pollination syndromes (Stebbins 1970;
Faegri & Van Der Pijl 1971; Proctor, Leo & Lack 1996;
Fenster et al. 2004). Pollinators of a given species can
vary among different populations, leading to plant and*Correspondence author. E-mail: [email protected]
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society
Functional Ecology 2013 doi: 10.1111/1365-2435.12122

pollinator species interactions among populations under
different selective pressures, resulting in a geographical
mosaic of co-evolution (Thompson 2005; Gomez et al.
2009). These populations are more or less interconnected
by gene flows that control the degree of differentiation
among populations. In the case of sufficient divergence of
one of these populations, speciation is likely to occur
(Levin 2000; Thompson 2005). This process may for
instance occur through pollinator shifts: pollinators,
through visitation preference, can isolate a plant genotype
from another (Gould & Johnston 1972; Kiester, Lande &
Schemske 1984; Bradshaw & Schemske 2003). As pollina-
tors are likely to vary from a site to another, they may be
an important factor shaping geographical variations of flo-
ral traits due to local adaptation. Thus, studies on those
variations among populations might help understand the
mechanisms leading to speciation and evolution (Herrera,
Castellanos & Medrano 2006). Recently, some of these
studies revealed striking correlations between pollinator
shifts or changes in pollinator traits and variations of
floral features (e.g. Kato et al. 2000; Johnson & Steiner
1997; Elle & Carney 2003; Valiente-Banuet et al. 2004;
Anderson & Johnson 2007; Gomez et al. 2008; Cosacov,
Nattero & Cocucci 2008; Schlumpberger et al. 2009;
Brown, Downs & Johnson 2011; but see Ellis & Johnson
2009; Nattero, Cocussi & Medel 2010). It was also shown
that floral traits or pollinator composition can vary accord-
ing to abiotic conditions (Blionis & Vokou 2002; Hodgins
& Barrett 2008; Esp�ındola, Pellissier & Alvarez 2011).
Here, we investigated the geographical variations of a
plant–pollinator interaction, focusing on the main attrac-
tive signal: floral odour. The emission of floral odours is a
widespread trait in flowers, and its major function is plant
recognition by pollinators (Pichersky & Gershenzon 2002;
Knudsen et al. 2006; Schaeffer & Ruxton 2011). In some
plant species, it has been highly suspected or shown that
floral odours are the main attractive cues. Such is the case,
for example, in the very specialized interactions between
fig and fig wasps (Gibernau 1997), orchids and male bees
(Schiestl & Schl€uter 2009), yucca and yucca moth (Svens-
son, Pellmyr & Raguso 2011) or Araceae and euglossine
bees (Hentrich, Kaiser & Gottsberger 2010). This is also
the case for flowers attracting pollinators at dusk or during
the night, when visual cues are poorly informative (Maia
et al. 2012, 2013). Floral odours are labile and can vary in
compound composition, in the relative amount of com-
pounds, or their overall quantities (D€otterl, Wolfe &
J€urgens 2005; Raguso 2008). In the context of the
geographical mosaic of co-evolution (as described by
Thompson 2005), local adaptation to pollinators may thus
cause floral scent variations. Floral scent variations were
tested in a few studies, among which gene flow and
geographical distance have been more often proposed to
explain the observed odour variations or stability than
adaptation to local pollinator preferences (Ackerman,
Mel�endez-Ackerman & Salguero-Faria 1997; Knudsen
2002; Solers et al. 2011). So far, a limited number of stud-
ies have investigated geographical variations of both poll-
inators and floral scents (Svensson et al. 2005; Svensson
et al. 2006; Pettersson & Knudsen 2001; Schlumpberger &
Raguso 2008; Ibanez et al. 2010).
Arum italicum Mill. and Arum maculatum L. (Fig. 1) are
two species from the Araceae family, found in temperate
woodlands on the forest floor (Boyce 2006). They are polli-
nated by insects (mainly Diptera) which they lure with
floral odours mimicking ovipositing sites (sapromyophilous)
and entrap within isolated floral chambers for about 24 h
over 2 days (Lack & Diaz 1991; Albre, Quilichini & Giber-
nau 2003; Gibernau, Macquart & Przetak 2004). Up to
now, it has been shown that A. maculatum is mainly polli-
nated by one of only two species from the Psychodidae
family, Psychoda phalaenoides and Psycha grisescens,
according to the population (Prime 1960; Rohacek, Beck-
Haug & Dobat 1990; Lack & Diaz 1991; Diaz & Kite
2002; Chartier, Pelozuelo & Gibernau 2011; Esp�ındola,
Pellissier & Alvarez 2011). Contrastingly, the insect com-
position found in association with inflorescences of A. ital-
icum fluctuates greatly between sites (Gibernau, Macquart
& Przetak 2004; Chartier, Pelozuelo & Gibernau 2011):
different Psychodidae species were found in Spain and in
the South of France, as well as diverse Diptera species
from the families Ceratopogonidae, Sciaridae and Chiro-
nomidae (M�endez & Obeso 1992; Diaz & Kite 2002; Albre,
Quilichini & Gibernau 2003; Albre & Gibernau 2008;
Chartier, Pelozuelo & Gibernau 2011).
The pollinator-attractive odours of A. italicum and
A. maculatum have been studied in England (Kite 1995; Kite
et al. 1998; Diaz & Kite 2002). Recently, it has been pro-
posed that geographical variations of the odour profiles of
these two species were linked to their different degree of spec-
ificity in several populations in France (Chartier, Pelozuelo
& Gibernau 2011): floral odour profiles of A. italicum were
not geographically structured among populations, suggesting
(a) (b)
Fig. 1. Inflorescence of (a) Arum maculatum L., (b) A. italicum
Mill. in Bagn�eres-de-Bigorre, France. (Pictures: M. Chartier).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
2 M. Chartier et al.

a high level of gene flow or adaptation to a fluctuant guild of
pollinators or to different pollinators having the same olfac-
tory preferences. On the other hand, odour profiles ofA. mac-
ulatum varied between studied populations, suggesting a
lower level of gene flow or adaptation to different local polli-
nator preferences. These hypotheses, nonetheless, remained
to be tested, and the chemical composition of the floral blends
was yet to be elucidated in these populations (Chartier,
Pelozuelo&Gibernau 2011).
In this paper, floral odours and pollinators of A. itali-
cum and A. maculatum were studied in native populations
within a wide geographical range. Furthermore, local
adaptation to pollinators was tested by (i) comparing
odours and pollinator geographical structures for both
species, and (ii) transplanting inflorescences between two
different sites and recording the number and the composi-
tion of insects trapped by the transplanted and native
inflorescences, as a measure of a fitness component. Hence,
if local adaptation to pollinators does occur (through vari-
ations in the attractive odour between populations as an
adaptation to local pollinator preferences), we would
expect that the fitness is lower for the transplanted plants
than for the native plants in each site.
The purpose of this long-term work was to answer three
main questions: (i) How do pollinator and odour composi-
tions vary in time and space in Arum italicum and A. mac-
ulatum? (ii) Is there a link between odour and pollinator
variations for both species? (iii) Are pollinator variations
among sites due to differences in plant attractiveness or in
pollinator availability?
Materials and methods
INFLORESCENCE V IS ITORS
Insects trapped inside the floral chambers of A. italicum and
A. maculatum were collected at seven locations: inflorescences of
A. italicum were sampled in 2009 and 2010 in Chantonnay
(Vend�ee, France, 46°40′N 1°06′O), Smarves (Vienne, France,
46°30′N 0°22′E), Toulouse (Haute-Garonne, France, 43°33′N1°28′E), and Bagn�eres-de-Bigorre (Midi-Pyr�en�ees, France, 43°04′N0°09′E), and in 2009 additionally in Pierrelatte (Drome, France,
44°22′N 4°14′E) and Igeldo (Gipuzkoa, Spain, 43°18′N 2°04′O). A
total of three populations of A. maculatum were sampled in La
Loubati�ere (2009, Aude, France, 43°24′N 2°15′E), Bagn�eres-
de-Bigorre and Smarves (2009 and 2010). Note that Bagn�eres-de-Bigorre and Smarves are sites where A. maculatum and
A. italicum grow intermingled.
Inflorescence visitors were collected in each population in the
morning of the second day of flowering. At this phenological
stage, insects are captive inside the floral chamber. Trapped insects
were collected by pouring 70% ethanol into the floral chamber
and then opening the spathes with a scalpel. The insects were con-
served in 70% ethanol until determination at the family level
under a stereomicroscope, with precious help from a Diptera
taxonomist (Prof. Alain Thomas, University of Toulouse).
As Psychodidae were the most numerous insects trapped, and
the main pollinators of A. maculatum, we estimated their diversity
in each population. When available, four Psychodidae females per
inflorescence from five inflorescences per population were identi-
fied at the species level under a microscope based on their genitalia
and antenna shapes (Vaillant 1988; Withers 1989; Je�zek 1990). In
three populations of A. italicum, <20 Psychodidae were trapped in
all the collected inflorescences. In these cases, all the Psychodidae
were identified.
FLORAL SCENT COLLECT ION
Odours of four to 10 inflorescences of A. maculatum and A. itali-
cum from each of the studied sites were collected in 2009 and 2010.
Inflorescence odours were collected in the evening, between 8 p.m.
and 11 p.m. At this stage, the spathe is widely open, the appendix
is warm and the floral odour is strong. For odour collecting, each
sampled inflorescence (spathe and spadix) was wrapped in a plastic
inert bag (Nalophan NA colourless, calibre 90, available from ETS
Charles-Fr�eres, Saint-Etienne, France) to create an ‘open static
headspace’: the bottom of the bag was kept close under the floral
chamber with a bond, isolating the inflorescence from the leaves
and soil. The top of the bag was kept open 10 cm above the spathe,
to avoid any condensation due to the heat of the appendix. Volatile
organic compounds (VOCs) were collected by solid phase microex-
traction (SPME): VOCs are absorbed and desorbed from a fibre
stored within the needle of a modified syringe. We used Stable-
FlexTM SPME Fibre, 65 lm Polydimethylsiloxane/Divinylben-
zene, handled with a manual holder (available from Supelco,
Bellefonte, PA, USA). The fibre was introduced in the nalophan
bag through a little slit and maintained 0�5–1�0 cm distant from
the appendix for 20 min. Closed empty bags containing ambient
air from 3–4 m away the inflorescences were used as controls
to discard putative VOCs not originating from inflorescences.
Fibres were stored in a freezer (�20 °C) until analyses by gas
chromatography–mass spectometry.
FLORAL SCENT ANALYSES
Gas chromatography–mass spectometry analyses were performed
at the Platform for Chemical Analyses in Ecology of the ‘SFR 119
Montpellier Environnement Biodiversit�e’, at the ‘Centre d’�Ecolo-
gie Fonctionnelle et �Evolutive (Montpellier, France)’. SPME
Fibres were desorbed for 5 min at 250 °C into a 1177 Split/Split-
less injector of a CP-3800 gas chromatograph (Varian Inc., Palo
Alto, CA, USA) coupled to a Saturn 2000 ion trap mass spec-
trometer (Varian Inc.). The carrier gas was helium with a constant
flow rate of 1�0 mL min�1. A split ratio of 1:4 was used. The
temperature of the column (fused silica capillary column, 30 m
9 0�25 mm 9 0�25 lm, CP-Sil 8 CB lowbleed MS, Varian Inc. in
2009; Optima 5 Accent, Macherey-Nagel, D€uren, Germany in
2010) was maintained at 50 °C for 2 min after injection, increased
to 200 °C at a rate of 5 °C min�1, and then increased to 250 °Cat a rate of 10 °C min�1 and maintained at 250 °C for 1 min.
Mass spectra were recorded in scan mode from 38 to 300 m/z with
an electronic impact (EI) at 70 eV. The chemical compounds were
identified by comparison with the NIST98 MS and Adams 2007
mass spectral libraries, and retention indices were found in
libraries and published data (Adams 2007). All chromatogram
peaks were manually integrated and the relative percentage area
of each peak was calculated for each chromatogram (i.e. each
sampled inflorescence). Only peaks representing more than 1% of
the total peaks area per chromatogram were kept for the analyses.
RECIPROCAL TRANSPLANT EXPER IMENT
Plants were transplanted between the sites of Bagn�eres-de-Bigorre
and Toulouse in 2008 and 2009. Entire plants bearing an inflores-
cence were put in pots in the morning, before the opening of the
spathes and transported between sites in the early afternoon. Pots
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 3

were randomly disposed among the native plants to avoid any
environmental bias. Inflorescences opened in the evening, and the
trapped insects were collected in the following morning. Insects
were preserved in 70% ethanol until taxonomic determination at
the family level. In 2008, plants of A. italicum were transplanted
from Toulouse to Bagn�eres-de-Bigorre and plants of A. macula-
tum were moved from Bagn�eres-de-Bigorre to Toulouse. In 2009,
plants of A. italicum were transplanted from Toulouse to
Bagn�eres-de-Bigorre and plants of the two species from Bagn�eres-
de-Bigorre to Toulouse. The composition and number of the
insects trapped inside inflorescences of these transplanted plants
were compared to those recorded in inflorescences of native plants
at the same sites and during the same days (data from native inflo-
rescences in 2008 are from Chartier, Pelozuelo & Gibernau 2011;
data from 2009 are from the present study).
The composition of Psychodidae trapped inside the trans-
planted inflorescences was estimated in 2009 by identifying female
specimens at the species level under a microscope based on their
genitalia and antenna shapes (Vaillant 1988; Withers 1989; Je�zek
1990). Four female specimens were selected per inflorescence, from
five inflorescences per treatment.
Note that all trapped insects were harvested from inside the
transplanted inflorescences; thus, no pollen from these inflores-
cences could be transported to native inflorescences.
STAT IST ICAL ANALYSES
The quantity of insects trapped inside inflorescences was com-
pared between species or populations of Arum with Wilcoxon
Mann–Whitney (WMW) and Kruskal–Wallis (KW) tests using the
functions wilcox.test() and kruskal.test() from stats package in R
version 2.15.1 (R Core Team, 2012). Insect proportions were com-
pared between groups by nonparametric multivariate analyses of
variance (npMANOVA) with the function adonis() from vegan pack-
age in R (Anderson 2001) using the factors ‘population’, ‘year’
and ‘species’ and their interactions.
For transplantation tests, the insect composition was compared
between groups (native and transplanted inflorescences) with the
same test for each transplantation event. Post hoc tests consisted
of npMANOVAs with a Bonferroni correction. In the results section,
only the statistic values for global tests are given.
To visualize individual variations of odours, VOCs were
grouped by metabolic pathways into four classes: benzenoids,
monoterpenoids, sesquiterpenoids and aliphatics. As indole, a
nitrogen-containing compound, was one of the main compounds
emitted by A. maculatum, it was added as a fifth compound class.
For each species, the relative amount of each compound class for
all individuals was compared between each group (plants grouped
per species, population and year) with a npMANOVA using the func-
tion adonis() in R.
Floral scents were compared between species and between
populations for each species and year according to the relative
percentage of each compound. The significance of the differences
between the different groups was assessed with a npMANOVA using
the function adonis() in R and the factors ‘population’, ‘year’ and
‘species’ and their interactions. NpMANOVAs were used as post hoc,
with a Bonferroni correction.
In addition, the odour variability was compared between the
two species by comparing the mean Jaccard distances among
individuals per species (Ackerman, Cuevas & Hof 2011).
To assess whether there was a correlation between scent compo-
sition and pollinator assemblage, a Mantel test was performed on
the insect and the odour matrices from inflorescences for which
both the odour and pollinators had been identified (across all
populations). As A. maculatum inflorescences trapped primarily
P. phalaenoides and P. grisescens, an initial analysis was per-
formed taking into account the non-Psychodidae families plus the
total number of Psychodidae trapped per inflorescence; in a
second analysis, the total number of Psychodidae was replaced by
an estimation of the number of P. phalaenoides and of P. grises-
cens trapped per inflorescence (extrapolated for each inflorescence
from the percentage of insects from both Psychodidae species
identified in five inflorescences per populations, see section ‘Inflo-
rescence Visitors’). The tests were performed with 10 000 repeti-
tions with the function mantel.randtest() from ade4 package in
R on three data sets: A. italicum and A. maculatum, A. italicum
alone and A. maculatum alone.
Results
OVERALL POLL INATOR VAR IAT IONS
In all studied populations in 2009 and 2010, inflorescences
of Arum italicum and A. maculatum trapped mainly
Diptera from nematoceran families: Psychodidae (10 964
insects), Chironomidae (2007), Ceratopogonidae (107),
Sciaridae (34), plus some Brachycera (209, Diptera) and
Staphylinidae (62, Coleoptera). A total of 51 other insects
were not identified. Inflorescences of A. italicum trapped in
average 18�1 � 2�87 insects (n = 240), including 8�70 � 1�27Psychodidae and 8�01 � 2�09 Chironomidae. Inflorescences
of A. maculatum trapped significantly more insects (WMW:
W = 15�866, P < 1�10�3), with a mean of 84�93 � 17�07insects per inflorescence (n = 107), including 82�94 � 17�02Psychodidae.
The number of insects trapped by inflorescences of both
A. italicum and A. maculatum varied geographically
between populations, but also interannually for a given
population (Table 1). In 2009, inflorescences of A. italicum
trapped two to 49 times more insects in Smarves,
Bagn�eres-de-Bigorre and Igeldo than in Chantonnay, Pier-
relatte and Toulouse (KW: v25, 153 = 65�86, P < 1�10�12).
In 2010, the inflorescences of Bagn�eres-de-Bigorre trapped
51 to 67 times more insects than in the other studied popula-
tions of Toulouse, Chantonnay and Smarves (KW: v23, 85 =50�85, P < 1�10�10). Moreover, interannual variations were
documented in A. italicum for the three studied popula-
tions. Hence, the number of insects trapped per inflores-
cence in 2010 compared to 2009 was three times higher in
Bagn�eres-de-Bigorre (WMW: W = 209, P = 0�003), 33
times lower in Smarves (WMW: W = 343, P < 1�10�6) and
four times lower in Chantonnay (WMW: W = 252,
P < 1�10�3). Geographical variations also occurred for
A. maculatum: inflorescences of this species trapped 11 to
99 times more insects in Bagn�eres-de-Bigorre in 2009 (KW:
v22, 49 = 13�24, P < 1�10�3) and 2010 (MW: W = 788,
P < 1�10�8) than in Smarves and la Loubati�ere. Interannu-
al variations also occurred: the number of insects trapped
per inflorescence in 2010 compared to 2009 was 1�5 times
higher in Bagn�eres-de-Bigorre (WMW: W = 183,
P = 0�031) and 13 times lower in Smarves (WMW:
W = 341, P < 1�10�4).
Pollinator composition exhibited significant differences
between species, populations and years (Figs 2 and 3).
Geographical and interannual variations of pollinator
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
4 M. Chartier et al.
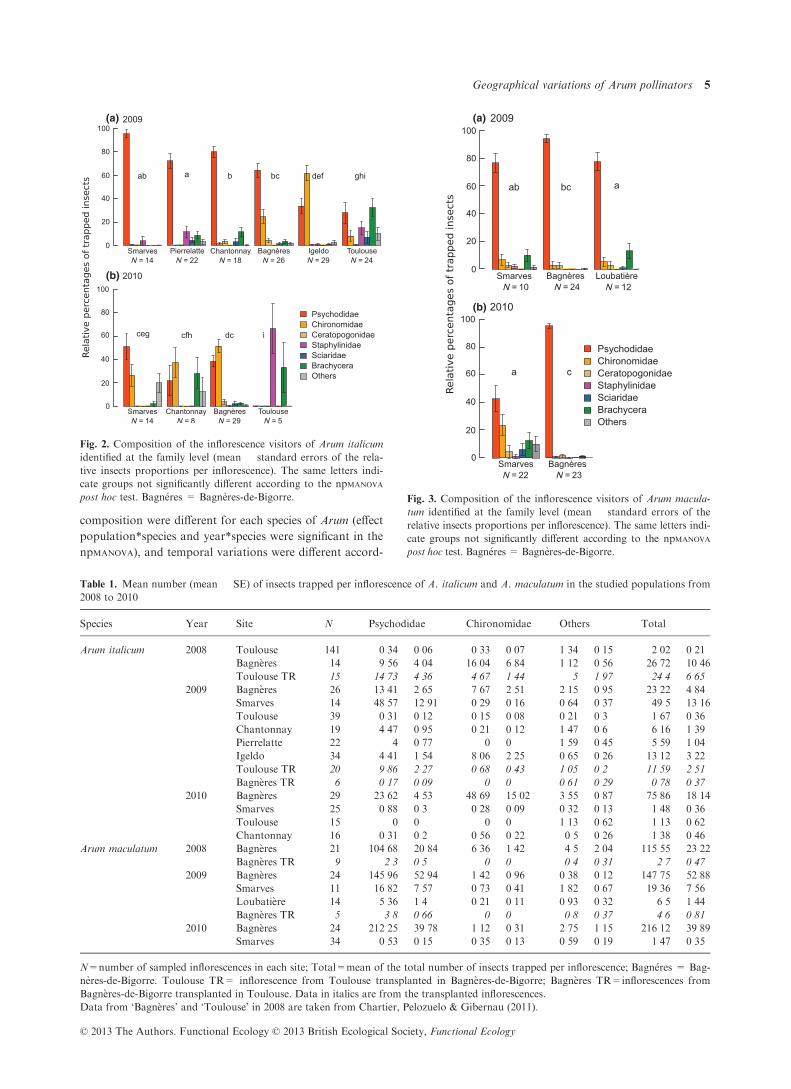
composition were different for each species of Arum (effect
population*species and year*species were significant in the
npMANOVA), and temporal variations were different accord-
Table 1. Mean number (mean � SE) of insects trapped per inflorescence of A. italicum and A. maculatum in the studied populations from
2008 to 2010
Species Year Site N Psychodidae Chironomidae Others Total
Arum italicum 2008 Toulouse 141 0�34 � 0�06 0�33 � 0�07 1�34 � 0�15 2�02 � 0�21Bagn�eres 14 9�56 � 4�04 16�04 � 6�84 1�12 � 0�56 26�72 � 10�46Toulouse TR 15 14�73 � 4�36 4�67 � 1�44 5 � 1�97 24�4 � 6�65
2009 Bagn�eres 26 13�41 � 2�65 7�67 � 2�51 2�15 � 0�95 23�22 � 4�84Smarves 14 48�57 � 12�91 0�29 � 0�16 0�64 � 0�37 49�5 � 13�16Toulouse 39 0�31 � 0�12 0�15 � 0�08 0�21 � 0�3 1�67 � 0�36Chantonnay 19 4�47 � 0�95 0�21 � 0�12 1�47 � 0�6 6�16 � 1�39Pierrelatte 22 4 � 0�77 0 � 0 1�59 � 0�45 5�59 � 1�04Igeldo 34 4�41 � 1�54 8�06 � 2�25 0�65 � 0�26 13�12 � 3�22Toulouse TR 20 9�86 � 2�27 0�68 � 0�43 1�05 � 0�2 11�59 � 2�51Bagn�eres TR 6 0�17 � 0�09 0 � 0 0�61 � 0�29 0�78 � 0�37
2010 Bagn�eres 29 23�62 � 4�53 48�69 � 15�02 3�55 � 0�87 75�86 � 18�14Smarves 25 0�88 � 0�3 0�28 � 0�09 0�32 � 0�13 1�48 � 0�36Toulouse 15 0 � 0 0 � 0 1�13 � 0�62 1�13 � 0�62Chantonnay 16 0�31 � 0�2 0�56 � 0�22 0�5 � 0�26 1�38 � 0�46
Arum maculatum 2008 Bagn�eres 21 104�68 � 20�84 6�36 � 1�42 4�5 � 2�04 115�55 � 23�22Bagn�eres TR 9 2�3 � 0�5 0 � 0 0�4 � 0�31 2�7 � 0�47
2009 Bagn�eres 24 145�96 � 52�94 1�42 � 0�96 0�38 � 0�12 147�75 � 52�88Smarves 11 16�82 � 7�57 0�73 � 0�41 1�82 � 0�67 19�36 � 7�56Loubati�ere 14 5�36 � 1�4 0�21 � 0�11 0�93 � 0�32 6�5 � 1�44Bagn�eres TR 5 3�8 � 0�66 0 � 0 0�8 � 0�37 4�6 � 0�81
2010 Bagn�eres 24 212�25 � 39�78 1�12 � 0�31 2�75 � 1�15 216�12 � 39�89Smarves 34 0�53 � 0�15 0�35 � 0�13 0�59 � 0�19 1�47 � 0�35
N=number of sampled inflorescences in each site; Total=mean of the total number of insects trapped per inflorescence; Bagn�eres = Bag-
neres-de-Bigorre. Toulouse TR= inflorescence from Toulouse transplanted in Bagn�eres-de-Bigorre; Bagn�eres TR=inflorescences from
Bagn�eres-de-Bigorre transplanted in Toulouse. Data in italics are from the transplanted inflorescences.
Data from ‘Bagn�eres’ and ‘Toulouse’ in 2008 are taken from Chartier, Pelozuelo & Gibernau (2011).
(a)
(b)
Fig. 2. Composition of the inflorescence visitors of Arum italicum
identified at the family level (mean � standard errors of the rela-
tive insects proportions per inflorescence). The same letters indi-
cate groups not significantly different according to the npMANOVA
post hoc test. Bagn�eres = Bagneres-de-Bigorre.
(a)
(b)
Fig. 3. Composition of the inflorescence visitors of Arum macula-
tum identified at the family level (mean � standard errors of the
relative insects proportions per inflorescence). The same letters indi-
cate groups not significantly different according to the npMANOVA
post hoc test. Bagn�eres = Bagneres-de-Bigorre.
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 5
ing to populations (effect population*year was significant,
Table 2). There was no significant combined effect of year,
population and species (npMANOVA: df = 1,280, F = 0�88,r2 = 0�002, P = 0�44) and this factor was thus removed
from the model.
In 2009, different populations of A. italicum showed dif-
ferent associated pollinator compositions. Inflorescences
trapped 64–95% of Psychodidae in Smarves, Chantonnay,
Pierrelatte and Bagn�eres-de-Bigorre (Fig. 2a). Insect com-
position found in association with A. italicum inflorescences
in Igeldo and Toulouse was significantly different than that
of all other populations: inflorescences trapped mainly
Chironomidae in Igeldo (61�75%), whereas in Toulouse,
they trapped few Psychodidae (27�22%) and Chironomidae
(7�98%), but a high diversity of other insects (64�8% of all
other categories). In 2010, populations of A. italicum
attracted a lower proportion of Psychodidae than in 2009
and showed two different tendencies (Fig. 2b). In Smarves,
Chantonnay and Bagn�eres-de-Bigorre, inflorescences of
A. italicum attracted a majority of Psychodidae and Chiro-
nomidae, with a high proportion of Brachycera in Chanton-
nay (28�1%) and of unidentified insects in Smarves (20�0%).
The insect composition found in inflorescences from Tou-
louse was significantly different than that of all other popu-
lations, with 66�7% of Sciaridae and 33�3% of Brachycera
(Fig. 2b). Insect compositions varied significantly between
2009 and 2010 in Smarves and Chantonnay, and marginally
significantly in Bagn�eres-de-Bigorre and Toulouse (Fig. 2).
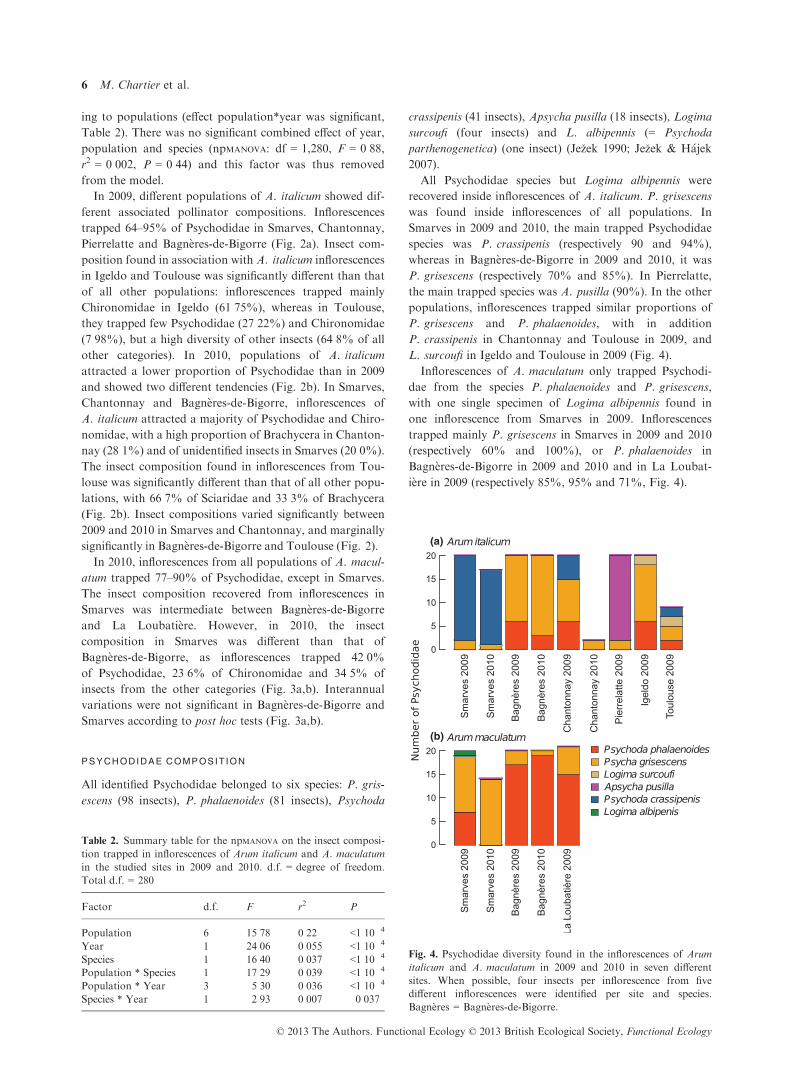
In 2010, inflorescences from all populations of A. macul-
atum trapped 77–90% of Psychodidae, except in Smarves.
The insect composition recovered from inflorescences in
Smarves was intermediate between Bagn�eres-de-Bigorre
and La Loubati�ere. However, in 2010, the insect
composition in Smarves was different than that of
Bagn�eres-de-Bigorre, as inflorescences trapped 42�0%of Psychodidae, 23�6% of Chironomidae and 34�5% of
insects from the other categories (Fig. 3a,b). Interannual
variations were not significant in Bagn�eres-de-Bigorre and
Smarves according to post hoc tests (Fig. 3a,b).
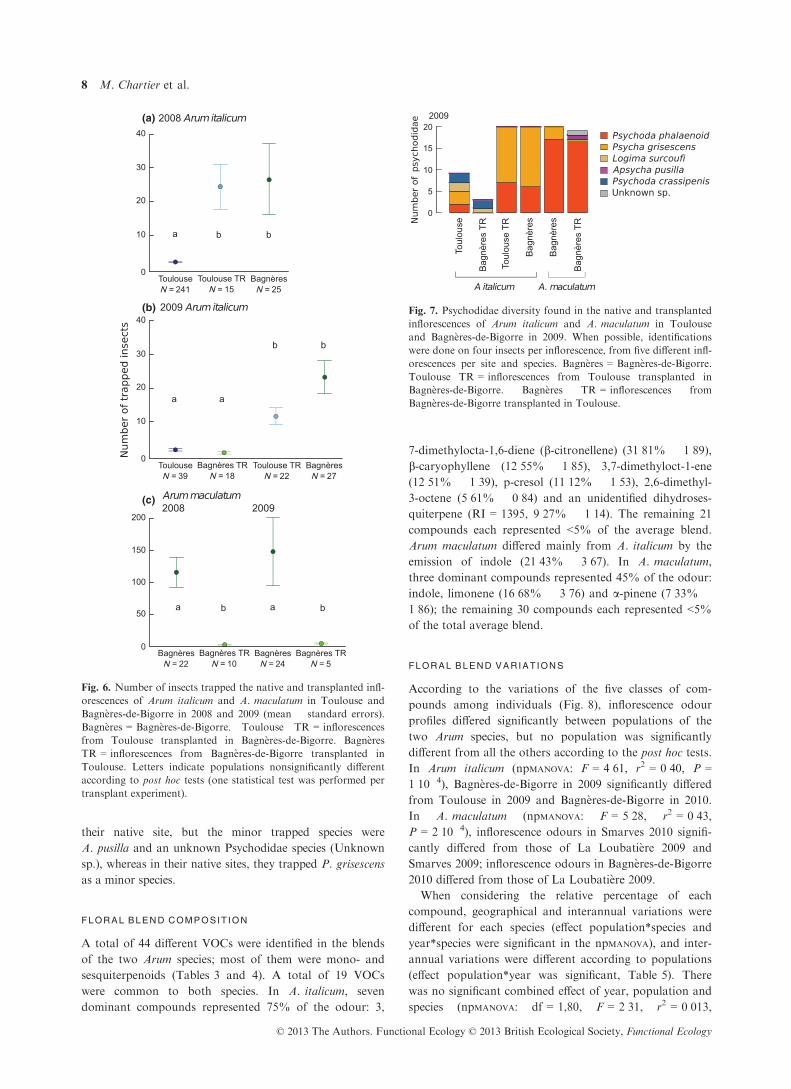
PSYCHODIDAE COMPOSIT ION
All identified Psychodidae belonged to six species: P. gris-
escens (98 insects), P. phalaenoides (81 insects), Psychoda
crassipenis (41 insects), Apsycha pusilla (18 insects), Logima
surcoufi (four insects) and L. albipennis (= Psychoda
parthenogenetica) (one insect) (Je�zek 1990; Je�zek & H�ajek
2007).
All Psychodidae species but Logima albipennis were
recovered inside inflorescences of A. italicum. P. grisescens
was found inside inflorescences of all populations. In
Smarves in 2009 and 2010, the main trapped Psychodidae
species was P. crassipenis (respectively 90 and 94%),
whereas in Bagn�eres-de-Bigorre in 2009 and 2010, it was
P. grisescens (respectively 70% and 85%). In Pierrelatte,
the main trapped species was A. pusilla (90%). In the other
populations, inflorescences trapped similar proportions of
P. grisescens and P. phalaenoides, with in addition
P. crassipenis in Chantonnay and Toulouse in 2009, and
L. surcoufi in Igeldo and Toulouse in 2009 (Fig. 4).
Inflorescences of A. maculatum only trapped Psychodi-
dae from the species P. phalaenoides and P. grisescens,
with one single specimen of Logima albipennis found in
one inflorescence from Smarves in 2009. Inflorescences
trapped mainly P. grisescens in Smarves in 2009 and 2010
(respectively 60% and 100%), or P. phalaenoides in
Bagn�eres-de-Bigorre in 2009 and 2010 and in La Loubat-
i�ere in 2009 (respectively 85%, 95% and 71%, Fig. 4).
(a)
(b)
Fig. 4. Psychodidae diversity found in the inflorescences of Arum
italicum and A. maculatum in 2009 and 2010 in seven different
sites. When possible, four insects per inflorescence from five
different inflorescences were identified per site and species.
Bagn�eres = Bagn�eres-de-Bigorre.
Table 2. Summary table for the npMANOVA on the insect composi-
tion trapped in inflorescences of Arum italicum and A. maculatum
in the studied sites in 2009 and 2010. d.f. = degree of freedom.
Total d.f. = 280
Factor d.f. F r2 P
Population 6 15�78 0�22 <1�10�4
Year 1 24�06 0�055 <1�10�4
Species 1 16�40 0�037 <1�10�4
Population * Species 1 17�29 0�039 <1�10�4
Population * Year 3 5�30 0�036 <1�10�4
Species * Year 1 2�93 0�007 0�037
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
6 M. Chartier et al.

RECIPROCAL TRANSPLANT EXPER IMENT : OVERALL
INSECT COMPOSIT ION
In 2008 and 2009, Arum italicum inflorescences trans-
planted from Toulouse to Bagn�eres-de-Bigorre trapped the
same composition of insects as the native A. italicum inflo-
rescences in Bagn�eres-de-Bigorre, and a significantly differ-
ent composition than in their native site (overall test
npMANOVA: F = 10�64, r2 = 0�113, P < 1�10�4 in 2008,
F = 9�67, r2 = 0�29, P < 1�10�4 in 2009; Fig. 5). In
Bagn�eres-de-Bigorre, transplanted and native inflorescenc-
es trapped a high percentage of Psychodidae (62�6% and
48�71%), contrary to native inflorescences in Toulouse
(19�7%). In both years, the proportion of Chironomidae
trapped in the transplanted inflorescences in Bagn�eres-
de-Bigorre (23�5% in 2008, 6�4% in 2009) was slightly lower
than in the native inflorescences (41�7% in 2008, 24�5% in
2009), as they attracted a significantly lower number of
Chironomidae (KW: v22 = 30�8, P < 1�10�6 in 2008,
v22 = 58�0, P < 1�10�11 in 2009). In both years, the total
number of insects trapped was six to 16 times higher in
Bagn�eres-de-Bigorre in the transplanted and native inflo-
rescences than in the native inflorescences in Toulouse
(KW: v22 = 30�8, P < 1�10�6 in 2008, v22 = 58�0,P < 1�10�11 in 2009, Fig. 6).
In the same way, Arum italicum inflorescences trans-
planted from Bagn�eres-de-Bigorre to Toulouse in 2009
trapped the same insect composition as the native A. itali-
cum inflorescences in Toulouse, and a significantly different
composition of insects than in their native site (overall test
npMANOVA: F = 9�67, r2 = 0�29, P < 1�10�4, Fig. 5). They
trapped a low proportion of Psychodidae (16�7%) and 30
times less insects than in their native site (MW: v22 = 58�0,P < 1�10�11, Fig. 6).
The insect composition recovered from inflorescences of
A. maculatum transplanted from Bagn�eres-de-Bigorre to
Toulouse was not significantly different than that of native
A. maculatum inflorescences in Bagn�eres-de-Bigorre (Fig. 5c;
npMANOVA: F = 2�61, r2 = 0�09, P = 0�063 in 2008; F = 3�35,r2 = 0�11, P = 0�051 in 2009). In all cases, inflorescences
trapped more than 85% of Psychodidae. In both years, the
number of insects trapped by the transplanted inflorescenc-
es in Toulouse was 25–55 times lower than that recovered
inside native inflorescences in Bagn�eres-de-Bigorre
(Table 1, Fig. 6). Note that A. maculatum does not
naturally occur in Toulouse, but the transplanted inflores-
cences of this species trapped more insects than those of
A. italicum in this locality.
RECIPROCAL TRANSPLANT EXPER IMENT :
PSYCHODIDAE COMPOSIT ION
Inflorescences of A. italicum transplanted from Toulouse
to Bagn�eres-de-Bigorre trapped the same proportions of
Psychodidae species as the native A. italicum inflorescences
in Bagn�eres-de-Bigorre (65% of P. grisescens and 35% of
P. phalaenoides, Fig. 7). As inflorescences of A. italicum
trapped few Psychodidae in Toulouse, we were only able to
determine three specimens from inflorescences of Bagn�eres-
de-Bigorre transplanted in Toulouse: one L. surcoufi and
two P. crassipenis, which was consistent with the composi-
tion of Psychodidae found in the native inflorescences of
A. italicum in Toulouse. The inflorescences of A. maculatum
transplanted from Bagn�eres-de-Bigorre to Toulouse trapped
89% of P. phalaenoides, similarly to what was observed in
(a)
(b)
(c)
Fig. 5. Composition of the visitors of the native and transplanted
inflorescences of Arum italicum and A. maculatum in Toulouse
and Bagn�eres-de-Bigorre in 2008 and 2009 (mean � standard
errors of the relative insects proportions per inflorescence).
Bagn�eres = Bagn�eres-de-Bigorre. Toulouse TR = inflorescences
from Toulouse transplanted in Bagn�eres-de-Bigorre. Bagn�eres
TR = inflorescences from Bagn�eres-de-Bigorre transplanted in
Toulouse. The same letters indicate groups not significantly differ-
ent (one statistical npMANOVA test was performed per transplant
experiment).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 7
their native site, but the minor trapped species were
A. pusilla and an unknown Psychodidae species (Unknown
sp.), whereas in their native sites, they trapped P. grisescens
as a minor species.
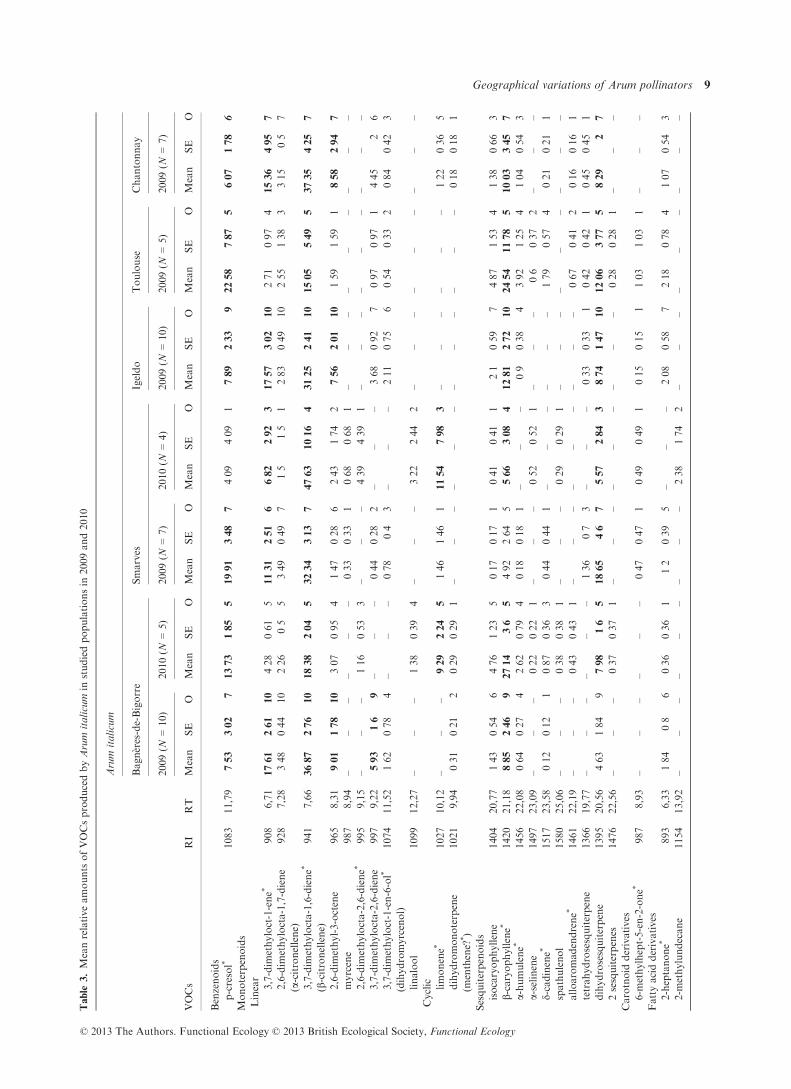
FLORAL BLEND COMPOSIT ION
A total of 44 different VOCs were identified in the blends
of the two Arum species; most of them were mono- and
sesquiterpenoids (Tables 3 and 4). A total of 19 VOCs
were common to both species. In A. italicum, seven
dominant compounds represented 75% of the odour: 3,
7-dimethylocta-1,6-diene (b-citronellene) (31�81% � 1�89),b-caryophyllene (12�55% � 1�85), 3,7-dimethyloct-1-ene
(12�51% � 1�39), p-cresol (11�12% � 1�53), 2,6-dimethyl-
3-octene (5�61% � 0�84) and an unidentified dihydroses-
quiterpene (RI = 1395, 9�27% � 1�14). The remaining 21
compounds each represented <5% of the average blend.
Arum maculatum differed mainly from A. italicum by the
emission of indole (21�43% � 3�67). In A. maculatum,
three dominant compounds represented 45% of the odour:
indole, limonene (16�68% � 3�76) and a-pinene (7�33% �1�86); the remaining 30 compounds each represented <5%of the total average blend.
FLORAL BLEND VAR IAT IONS
According to the variations of the five classes of com-
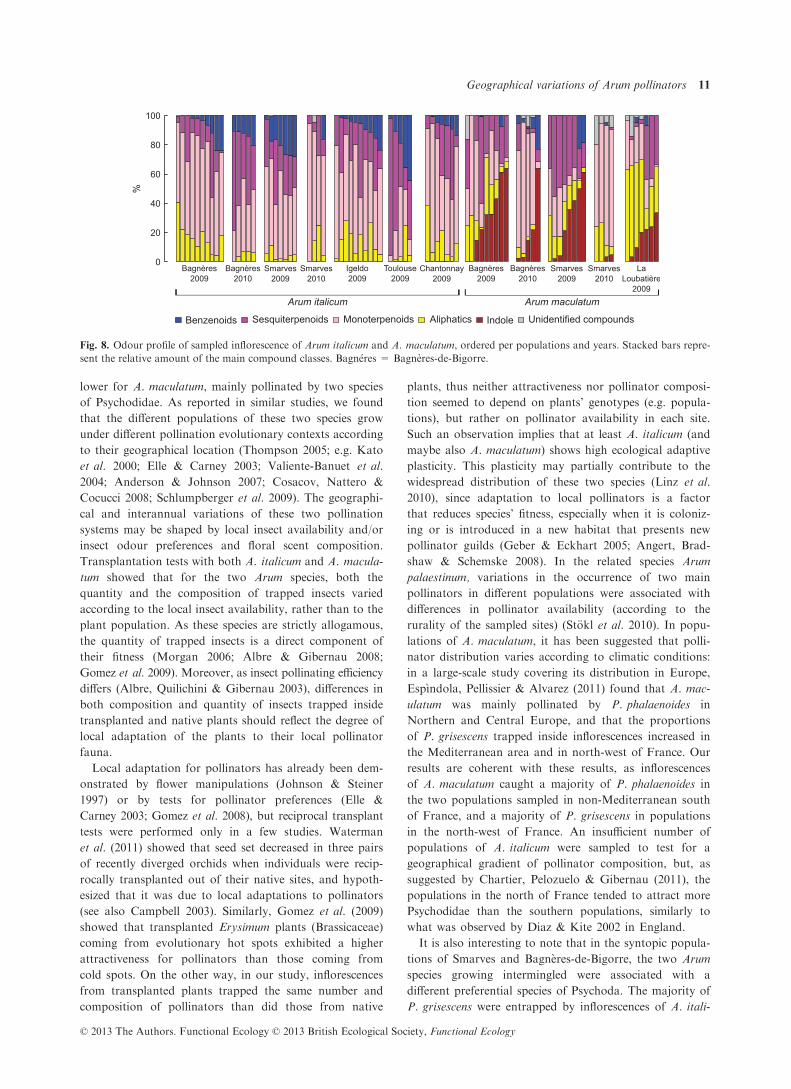
pounds among individuals (Fig. 8), inflorescence odour
profiles differed significantly between populations of the
two Arum species, but no population was significantly
different from all the others according to the post hoc tests.
In Arum italicum (npMANOVA: F = 4�61, r2 = 0�40, P =1�10�4), Bagn�eres-de-Bigorre in 2009 significantly differed
from Toulouse in 2009 and Bagn�eres-de-Bigorre in 2010.
In A. maculatum (npMANOVA: F = 5�28, r2 = 0�43,P = 2�10�4), inflorescence odours in Smarves 2010 signifi-
cantly differed from those of La Loubati�ere 2009 and
Smarves 2009; inflorescence odours in Bagn�eres-de-Bigorre
2010 differed from those of La Loubati�ere 2009.
When considering the relative percentage of each
compound, geographical and interannual variations were
different for each species (effect population*species and
year*species were significant in the npMANOVA), and inter-
annual variations were different according to populations
(effect population*year was significant, Table 5). There
was no significant combined effect of year, population and
species (npMANOVA: df = 1,80, F = 2�31, r2 = 0�013,
Fig. 7. Psychodidae diversity found in the native and transplanted
inflorescences of Arum italicum and A. maculatum in Toulouse
and Bagn�eres-de-Bigorre in 2009. When possible, identifications
were done on four insects per inflorescence, from five different infl-
orescences per site and species. Bagn�eres = Bagn�eres-de-Bigorre.
Toulouse TR = inflorescences from Toulouse transplanted in
Bagn�eres-de-Bigorre. Bagn�eres TR = inflorescences from
Bagn�eres-de-Bigorre transplanted in Toulouse.
(a)
(b)
(c)
Fig. 6. Number of insects trapped the native and transplanted infl-
orescences of Arum italicum and A. maculatum in Toulouse and
Bagn�eres-de-Bigorre in 2008 and 2009 (mean � standard errors).
Bagn�eres = Bagn�eres-de-Bigorre. Toulouse TR = inflorescences
from Toulouse transplanted in Bagn�eres-de-Bigorre. Bagn�eres
TR = inflorescences from Bagn�eres-de-Bigorre transplanted in
Toulouse. Letters indicate populations nonsignificantly different
according to post hoc tests (one statistical test was performed per
transplant experiment).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
8 M. Chartier et al.
Table
3.Meanrelativeamounts
ofVOCsproducedbyArum
italicum
instudiedpopulationsin
2009and2010
VOCs
RI
RT
Arum
italicum
Bagn� eres-de-Bigorre
Smarves
Igeldo
Toulouse
Chantonnay
2009(N
=10)
2010(N
=5)
2009(N
=7)
2010(N
=4)
2009(N
=10)
2009(N
=5)
2009(N
=7)
Mean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
O
Benzenoids
p-cresol*
1083
11,79
7�53
3�02
713�73
1�85
519�91
3�48
74�09
4�09
17�89
2�33
922�58
7�87
56�07
1�78
6
Monoterpenoids
Linear
3,7-dim
ethyloct-1-ene*
908
6,71
17�61
2�61
10
4�28
0�61
511�31
2�51
66�82
2�92
317�57
3�02
10
2�71
0�97
415�36
4�95
7
2,6-dim
ethylocta-1,7-diene
(a-citronellene)
928
7,28
3�48
0�44
10
2�26
0�5
53�49
0�49
71�5
1�5
12�83
0�49
10
2�55
1�38
33�15
0�5
7
3,7-dim
ethylocta-1,6-diene*
(b-citronellene)
941
7,66
36�87
2�76
10
18�38
2�04
532�34
3�13
747�63
10�16
431�25
2�41
10
15�05
5�49
537�35
4�25
7
2,6-dim
ethyl-3-octene
965
8,31
9�01
1�78
10
3�07
0�95
41�47
0�28
62�43
1�74
27�56
2�01
10
1�59
1�59
18�58
2�94
7
myrcene
987
8,94
––
––
––
0�33
0�33
10�68
0�68
1–
––
––
––
––
2,6-dim
ethylocta-2,6-diene*
995
9,15
––
–1�16
0�53
3–
––
4�39
4�39
1–
––
––
––
––
3,7-dim
ethylocta-2,6-diene
997
9,22
5�93
1�6
9–
––
0�44
0�28
2–
––
3�68
0�92
70�97
0�97
14�45
26
3,7-dim
ethyloct-1-en-6-ol*
(dihydromyrcenol)
1074
11,52
1�62
0�78
4–
––
0�78
0�4
3–
––
2�11
0�75
60�54
0�33
20�84
0�42
3
linalool
1099
12,27
––
–1�38
0�39
4–
––
3�22
2�44
2–
––
––
––
––
Cyclic limonene*
1027
10,12
––
–9�29
2�24
51�46
1�46
111�54
7�98
3–
––
––
–1�22
0�36
5
dihydromonoterpene
(menthene?
*)
1021
9,94
0�31
0�21
20�29
0�29
1–
––
––
––
––
––
–0�18
0�18
1
Sesquiterpenoids
isocaryophyllene
1404
20,77
1�43
0�54
64�76
1�23
50�17
0�17
10�41
0�41
12�1
0�59
74�87
1�53
41�38
0�66
3
b-caryophyllene*
1420
21,18
8�85
2�46
927�14
3�6
54�92
2�64
55�66
3�08
412�81
2�72
10
24�54
11�78
510�03
3�45
7
a-humulene*
1456
22,08
0�64
0�27
42�62
0�79
40�18
0�18
1–
––
0�9
0�38
43�92
1�25
41�04
0�54
3
a-selinene
1497
23,09
––
–0�22
0�22
1–
––
0�52
0�52
1–
––
0�6
0�37
2–
––
d-cadinene*
1517
23,58
0�12
0�12
10�87
0�36
30�44
0�44
1–
––
––
–1�79
0�57
40�21
0�21
1
spathulenol
1580
25,06
––
–0�38
0�38
1–
––
0�29
0�29
1–
––
––
––
––
alloaromadendrene*
1461
22,19
––
–0�43
0�43
1–
––
––
––
––
0�67
0�41
20�16
0�16
1
tetrahydrosesquiterpene
1366
19,77
––
––
––
1�36
0�7
3–
––
0�33
0�33
10�42
0�42
10�45
0�45
1
dihydrosesquiterpene
1395
20,56
4�63
1�84
97�98
1�6
518�65
4�6
75�57
2�84
38�74
1�47
10
12�06
3�77
58�29
27
2sesquiterpenes
1476
22,56
––
–0�37
0�37
1–
––
––
––
––
0�28
0�28
1–
––
Carotnoid
derivatives
6-m
ethylhept-5-en-2-one*
987
8,93
––
––
––
0�47
0�47
10�49
0�49
10�15
0�15
11�03
1�03
1–
––
Fattyacidderivatives
2-heptanone*
893
6,33
1�84
0�8
60�36
0�36
11�2
0�39
5–
––
2�08
0�58
72�18
0�78
41�07
0�54
3
2-m
ethylundecane
1154
13,92
––
––
––
––
–2�38
1�74
2–
––
––
––
––
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 9

P = 0�05) and this factor was thus removed from the
model. No population was different from all the others
when considering post hoc tests for both species.
In 2009, the inflorescence odours of A. italicum were dif-
ferent in Bagn�eres-de-Bigorre, Smarves and Chantonnay
when compared to Igeldo; and in Toulouse, they were
intermediate between the latter and Chantonnay. In 2010,
there was no significant difference between inflorescence
odours in Smarves and Bagn�eres-de-Bigorre. Floral odour
did not vary significantly between 2009 and 2010 in Smar-
ves, but did so in Bagn�eres-de-Bigorre, a result which may
be due to an increase of the emission of limonene and b-caryophyllene (Table 3).
In 2009, there was no significant difference in the inflo-
rescence odours of A. maculatum between the populations
of Bagn�eres-de-Bigorre, Smarves and La Loubati�ere, nor
in 2010 between Smarves and Bagn�eres-de-Bigorre. Odours
varied significantly between 2009 and 2010 in Smarves, but
not in Bagn�eres-de-Bigorre (Table 4).
The mean Jaccard distances among individuals were 0�44� 0�01 for A. italicum and 0�68 � 0�01 for A. maculatum.
CORRELAT ION BETWEEN SCENT AND POLL INATOR
COMPOSIT IONS
When integrating the total number of Psychodidae in the
analysis, we found a significant correlation between scent
dissimilarities and pollinator dissimilarities for A. italicum
alone (Mantel test: n = 38, obs = 0�217, P = 0�003), but
not for A. maculatum alone (Mantel test: n = 28,
obs = 0�156, P = 0�200). There was also no significant
correlation when integrating data from both Arum species
(Mantel test: n = 66, obs = 0�035, P = 0�207). When inte-
grating the number of P. phalaenoides and P. grisescens in
the analysis, we found a significant correlation between
scent dissimilarities and pollinator dissimilarities for
A. maculatum alone (Mantel test: n = 14, obs = 0�229,P = 0�045) and when integrating data from both Arum
species (Mantel test: n = 31, obs = 0�515, P < 10�3); there
was no significant correlation for A. italicum alone
(Mantel test: n = 17, obs = 0�380, P = 0�073).
Discussion
In this study, it was demonstrated that differences in floral
scent composition of two species of Arum influenced the
composition of the pollinator fauna associated with them.
However, there was no clear correlation between pollinator
variations among populations and odour composition. In
fact, pollinator attraction in each site was highly depen-
dent on local insect availability and diversity during the
transplant experiments, indicating no clear pattern of local
adaptation of the two Arum species to pollinators. The
two studied pollination systems varied between years and
sites. These variations were high for A. italicum, pollinated
by insects belonging to six different families (and by five
different Psychodidae species according to the site), andTable
3(C
ontinued)
VOCs
RI
RT
Arum
italicum
Bagn� eres-de-Bigorre
Smarves
Igeldo
Toulouse
Chantonnay
2009(N
=10)
2010(N
=5)
2009(N
=7)
2010(N
=4)
2009(N
=10)
2009(N
=5)
2009(N
=7)
Mean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
OMean
SE
O
decanal
1203
15,36
––
––
––
––
–1�12
1�12
1–
––
0�56
0�56
1–
––
Unknowncompounds
unknown1
1645
26,52
0�12
0�12
1–
––
0�18
0�18
1–
––
––
––
––
––
–Groups
ab
acd
bde
efbcf
ab
RI=
retentionindex;RT=
retentiontime;
N=
number
ofsampledinflorescences;
SE=
standard
error;
O=
number
ofchromatogrammswheretheVOC
wasrecorded;VOC
=volatile
organic
compounds.
Compoundsrepresentingmore
than5%
oftheaveragetotalblendare
highlightedin
bold.In
thebottom
line,
thesamelettersindicate
groupsnotsignificantlydifferent.
*Compoundsalsodescribed
byKiteet
al.(1998)orDiaz&
Kite(2002).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
10 M. Chartier et al.
lower for A. maculatum, mainly pollinated by two species
of Psychodidae. As reported in similar studies, we found
that the different populations of these two species grow
under different pollination evolutionary contexts according
to their geographical location (Thompson 2005; e.g. Kato
et al. 2000; Elle & Carney 2003; Valiente-Banuet et al.
2004; Anderson & Johnson 2007; Cosacov, Nattero &
Cocucci 2008; Schlumpberger et al. 2009). The geographi-
cal and interannual variations of these two pollination
systems may be shaped by local insect availability and/or
insect odour preferences and floral scent composition.
Transplantation tests with both A. italicum and A. macula-
tum showed that for the two Arum species, both the
quantity and the composition of trapped insects varied
according to the local insect availability, rather than to the
plant population. As these species are strictly allogamous,
the quantity of trapped insects is a direct component of
their fitness (Morgan 2006; Albre & Gibernau 2008;
Gomez et al. 2009). Moreover, as insect pollinating efficiency
differs (Albre, Quilichini & Gibernau 2003), differences in
both composition and quantity of insects trapped inside
transplanted and native plants should reflect the degree of
local adaptation of the plants to their local pollinator
fauna.
Local adaptation for pollinators has already been dem-
onstrated by flower manipulations (Johnson & Steiner
1997) or by tests for pollinator preferences (Elle &
Carney 2003; Gomez et al. 2008), but reciprocal transplant
tests were performed only in a few studies. Waterman
et al. (2011) showed that seed set decreased in three pairs
of recently diverged orchids when individuals were recip-
rocally transplanted out of their native sites, and hypoth-
esized that it was due to local adaptations to pollinators
(see also Campbell 2003). Similarly, Gomez et al. (2009)
showed that transplanted Erysimum plants (Brassicaceae)
coming from evolutionary hot spots exhibited a higher
attractiveness for pollinators than those coming from
cold spots. On the other way, in our study, inflorescences
from transplanted plants trapped the same number and
composition of pollinators than did those from native
plants, thus neither attractiveness nor pollinator composi-
tion seemed to depend on plants’ genotypes (e.g. popula-
tions), but rather on pollinator availability in each site.
Such an observation implies that at least A. italicum (and
maybe also A. maculatum) shows high ecological adaptive
plasticity. This plasticity may partially contribute to the
widespread distribution of these two species (Linz et al.
2010), since adaptation to local pollinators is a factor
that reduces species’ fitness, especially when it is coloniz-
ing or is introduced in a new habitat that presents new
pollinator guilds (Geber & Eckhart 2005; Angert, Brad-
shaw & Schemske 2008). In the related species Arum
palaestinum, variations in the occurrence of two main
pollinators in different populations were associated with
differences in pollinator availability (according to the
rurality of the sampled sites) (St€okl et al. 2010). In popu-
lations of A. maculatum, it has been suggested that polli-
nator distribution varies according to climatic conditions:
in a large-scale study covering its distribution in Europe,
Esp�ındola, Pellissier & Alvarez (2011) found that A. mac-
ulatum was mainly pollinated by P. phalaenoides in
Northern and Central Europe, and that the proportions
of P. grisescens trapped inside inflorescences increased in
the Mediterranean area and in north-west of France. Our
results are coherent with these results, as inflorescences
of A. maculatum caught a majority of P. phalaenoides in
the two populations sampled in non-Mediterranean south
of France, and a majority of P. grisescens in populations
in the north-west of France. An insufficient number of
populations of A. italicum were sampled to test for a
geographical gradient of pollinator composition, but, as
suggested by Chartier, Pelozuelo & Gibernau (2011), the
populations in the north of France tended to attract more
Psychodidae than the southern populations, similarly to
what was observed by Diaz & Kite 2002 in England.
It is also interesting to note that in the syntopic popula-
tions of Smarves and Bagn�eres-de-Bigorre, the two Arum
species growing intermingled were associated with a
different preferential species of Psychoda. The majority of
P. grisescens were entrapped by inflorescences of A. itali-
Fig. 8. Odour profile of sampled inflorescence of Arum italicum and A. maculatum, ordered per populations and years. Stacked bars repre-
sent the relative amount of the main compound classes. Bagn�eres = Bagneres-de-Bigorre.
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 11
cum in Bagn�eres-de-Bigorre and by inflorescences of
A. maculatum in Smarves. This suggests that there might
be a mechanism of avoidance of hybridization or/and
competition, which could reinforce interspecific odour
divergence, favouring pollinator shift when the two species
grow in syntopy (Levin & Anderson 1970; Waser 1978;
Mitchell et al. 2009). Odour variations between sympatric
related species and their hybrids can be involved in specia-
tion and competition avoidance and are thus particularly
interesting in the context of the geographical mosaic of
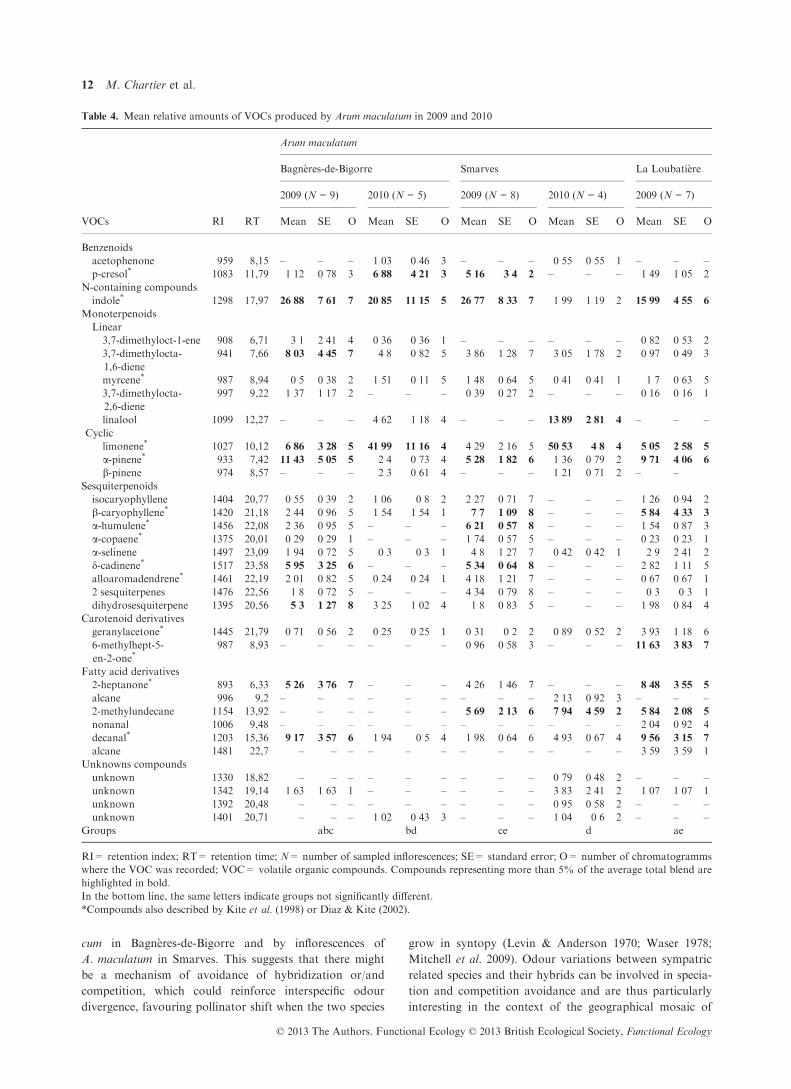
Table 4. Mean relative amounts of VOCs produced by Arum maculatum in 2009 and 2010
VOCs RI RT
Arum maculatum
Bagn�eres-de-Bigorre Smarves La Loubati�ere
2009 (N = 9) 2010 (N = 5) 2009 (N = 8) 2010 (N = 4) 2009 (N = 7)
Mean SE O Mean SE O Mean SE O Mean SE O Mean SE O
Benzenoids
acetophenone 959 8,15 – – – 1�03 0�46 3 – – – 0�55 0�55 1 – – –p-cresol* 1083 11,79 1�12 0�78 3 6�88 4�21 3 5�16 3�4 2 – – – 1�49 1�05 2
N-containing compounds
indole* 1298 17,97 26�88 7�61 7 20�85 11�15 5 26�77 8�33 7 1�99 1�19 2 15�99 4�55 6
Monoterpenoids
Linear
3,7-dimethyloct-1-ene 908 6,71 3�1 2�41 4 0�36 0�36 1 – – – – – – 0�82 0�53 2
3,7-dimethylocta-
1,6-diene
941 7,66 8�03 4�45 7 4�8 0�82 5 3�86 1�28 7 3�05 1�78 2 0�97 0�49 3
myrcene* 987 8,94 0�5 0�38 2 1�51 0�11 5 1�48 0�64 5 0�41 0�41 1 1�7 0�63 5
3,7-dimethylocta-
2,6-diene
997 9,22 1�37 1�17 2 – – – 0�39 0�27 2 – – – 0�16 0�16 1
linalool 1099 12,27 – – – 4�62 1�18 4 – – – 13�89 2�81 4 – – –Cyclic
limonene* 1027 10,12 6�86 3�28 5 41�99 11�16 4 4�29 2�16 5 50�53 4�8 4 5�05 2�58 5
a-pinene* 933 7,42 11�43 5�05 5 2�4 0�73 4 5�28 1�82 6 1�36 0�79 2 9�71 4�06 6
b-pinene 974 8,57 – – – 2�3 0�61 4 – – – 1�21 0�71 2 – –Sesquiterpenoids
isocaryophyllene 1404 20,77 0�55 0�39 2 1�06 0�8 2 2�27 0�71 7 – – – 1�26 0�94 2
b-caryophyllene* 1420 21,18 2�44 0�96 5 1�54 1�54 1 7�7 1�09 8 – – – 5�84 4�33 3
a-humulene* 1456 22,08 2�36 0�95 5 – – – 6�21 0�57 8 – – – 1�54 0�87 3
a-copaene* 1375 20,01 0�29 0�29 1 – – – 1�74 0�57 5 – – – 0�23 0�23 1
a-selinene 1497 23,09 1�94 0�72 5 0�3 0�3 1 4�8 1�27 7 0�42 0�42 1 2�9 2�41 2
d-cadinene* 1517 23,58 5�95 3�25 6 – – – 5�34 0�64 8 – – – 2�82 1�11 5
alloaromadendrene* 1461 22,19 2�01 0�82 5 0�24 0�24 1 4�18 1�21 7 – – – 0�67 0�67 1
2 sesquiterpenes 1476 22,56 1�8 0�72 5 – – – 4�34 0�79 8 – – – 0�3 0�3 1
dihydrosesquiterpene 1395 20,56 5�3 1�27 8 3�25 1�02 4 1�8 0�83 5 – – – 1�98 0�84 4
Carotenoid derivatives
geranylacetone* 1445 21,79 0�71 0�56 2 0�25 0�25 1 0�31 0�2 2 0�89 0�52 2 3�93 1�18 6
6-methylhept-5-
en-2-one*987 8,93 – – – – – – 0�96 0�58 3 – – – 11�63 3�83 7
Fatty acid derivatives
2-heptanone* 893 6,33 5�26 3�76 7 – – – 4�26 1�46 7 – – – 8�48 3�55 5
alcane 996 9,2 – – – – – – – – – 2�13 0�92 3 – – –2-methylundecane 1154 13,92 – – – – – – 5�69 2�13 6 7�94 4�59 2 5�84 2�08 5
nonanal 1006 9,48 – – – – – – – – – – – – 2�04 0�92 4
decanal* 1203 15,36 9�17 3�57 6 1�94 0�5 4 1�98 0�64 6 4�93 0�67 4 9�56 3�15 7
alcane 1481 22,7 – – – – – – – – – – – – 3�59 3�59 1
Unknowns compounds
unknown 1330 18,82 – – – – – – – – – 0�79 0�48 2 – – –unknown 1342 19,14 1�63 1�63 1 – – – – – – 3�83 2�41 2 1�07 1�07 1
unknown 1392 20,48 – – – – – – – – – 0�95 0�58 2 – – –unknown 1401 20,71 – – – 1�02 0�43 3 – – – 1�04 0�6 2 – – –
Groups abc bd ce d ae
RI= retention index; RT= retention time; N= number of sampled inflorescences; SE= standard error; O= number of chromatogramms
where the VOC was recorded; VOC= volatile organic compounds. Compounds representing more than 5% of the average total blend are
highlighted in bold.
In the bottom line, the same letters indicate groups not significantly different.
*Compounds also described by Kite et al. (1998) or Diaz & Kite (2002).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
12 M. Chartier et al.

co-evolution. Such is the case, for example, of some decep-
tive orchids (e.g. Schiestl & Ayasse 2002; St€okl et al. 2008)
and Silene species (Waelti et al. 2008).
Whereas the lack of clear geographical structure of
odour is consistent with the results from Chartier, Pelozu-
elo & Gibernau (2011) for A. italicum, we found no
geographical structure in the odour of A. maculatum either
in 2009 and 2010, surprisingly in contradiction with our
results from 2008 (Chartier, Pelozuelo & Gibernau 2011).
The geographical structure found in 2008 would have been
consistent with the hypothesis of Esp�ındola, Pellissier &
Alvarez (2011), suggesting that the floral odour of A. mac-
ulatum could be adapted to attract P. grisescens in some
populations and P. phalaenoides in others. This geographi-
cal structure was anyway poorly supported as only two
populations were sampled for odours in 2008, one of them
represented by only three individuals (Chartier, Pelozuelo
& Gibernau 2011). Instead, we found no clear odour varia-
tion/structure between populations of the two Arum
species, but their interindividual variations were high. The
lack of interpopulation odour variations can be either
explained as (i) a consequence of high gene flow among
populations of the two species, preventing a genetic differ-
entiation and thus the divergence of odour in some popu-
lations (Knudsen 2002; e.g. Svensson et al. 2005); or (ii) a
consequence of similar selective pressures for pollinator
attraction in distant populations (Svensson et al. 2006).
This could be the case for the attraction of P. phalaenoides
by inflorescences of A. italicum: the five major compounds
dominating the floral odour of A. italicum in our study
only slightly differed from the ones identified in British
populations primarily pollinated by Psychodidae (mainly
P. phalaenoides) and Chironomidae (Smittia pratorum)
(Diaz & Kite 2002).
The mean Jaccard distance indices between individual
floral odours of A. italicum (0�44) and A. maculatum (0�68)were both closer to the mean interindividual Jaccard index
of deceptive flowers (0�55) than to rewarding flowers
(0�28). These results were issued in a comparative study
over 12 species of rewarding flowers vs. deceptive flowers
(Ackerman, Cuevas & Hof 2011), indicating that deception
might be a factor selecting for high interindividual varia-
tions in the attractive odour. The odours of both Arum
species were mainly composed of sesquiterpenoids and
monoterpenoids, with some aliphatics (in higher propor-
tions in the floral odour of A. maculatum), and an aromatic
heterocyclic organic compound containing nitrogen, indole,
exclusively found in the floral odour of A. maculatum.
Among these compounds, some are very common
components of flower scents, such as a-pinene, limonene
and b-caryophyllene (Knudsen et al. 2006), and 2-hepta-
none, p-cresol, indole, a-pinene, limonene and b-caryo-phyllene are typical of the odours of sapromyophilous
plants (J€urgens, D€otterl & Meve 2006; Urru, Stensmyr &
Hansson 2011), and were also found in the odour of dung
from sheep, cow, horse or boar (Kite 1995; Dormont et al.
2010; Johnson & J€urgens 2010). In addition, 2-heptanone,
p-cresol and indole were shown to be attractive to psycho-
did flies, even more when they were mixed together (Kite
et al. 1998). St€okl et al. (2010), in their detailed study of
the pollination mechanism of the deceiving A. palaestinum,
proposed that the deceptive fermentation odour of the
plant was constituted of various attractive compounds
forming a ‘super-attractive mixture’ acting on Drosophila
innate preferences for fermentation-associated volatiles.
The mechanism is likely to be similar in inflorescences of
A. italicum and A. maculatum. The sapromyophilous flies
they deceive are known to look for a resource to oviposit,
but apparently inflorescences of neither studied species are
breeding sites for attracted flies, as no eggs, larvae or
pupae were ever observed in them (M. Chartier, pers.
obs.). This might suggest that a strong selective pressure is
likely to influence the behaviour of insects associated with
these aroids, favouring the ones able to ‘recognize’ and
avoid the inflorescences (Renner 2006). Arum italicum and
A. maculatum might thus be subjected to a balancing selec-
tion in favour of interindividual variations in the composi-
tion of the ‘super-attractive mixture’, to compensate any
evolutionary response from pollinators (Thompson 2005;
Renner 2006). In other plant species, it has been suggested
that high odour variations between individuals resulted
from few selective pressures from pollinators, for instance
if other cues play an important role in pollinator attraction
(Ibanez et al. 2010). This is not likely the case of Arum
italicum and A. maculatum were the major pollinator
attractive feature is known to be the floral odour (Lack &
Diaz 1991; reviewed by Gibernau, Macquart & Przetak
2004 and Urru, Stensmyr & Hansson 2011). High individ-
ual odour variations can also conceal the stability of physi-
ologically active compounds, which may represent a more
or less important proportion of the total variable odour
(Ayasse et al. 2000; Mant, Peakall & Schiestl 2005; Ibanez
et al. 2010), or be an adaptive response to high variations
of pollinators from year to year (Geber & Moeller 2006).
These two hypotheses could also explain the pattern found
in A. italicum and A. maculatum.
Future research should focus on coupling biotests on
pollinators and accumulating genetic information on plants
across a broad geographical population range. This data
will be decisive in assessing which VOCs are actually
attractive for pollinators, and what are the relative effects
of gene flow/genetic drift and pollinator selective pressures
Table 5. Summary table for the npMANOVA on the odour variabil-
ity for A. italicum and A. maculatum inflorescences in the studied
sites in 2009 and 2010. d.f. = degree of freedom, Total d.f. = 80
Factor d.f. F r2 P-value
Species 1 37�374 0�374 <1�10�4
Population 5 2�726 0�076 <1�10�4
Year 1 12�7 0�07 <1�10�4
Species * Population 1 3�398 0�019 0�009Population * Year 1 5�178 0�029 0�001Species * Year 1 8�085 0�045 <1�10�4
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 13

on odour variability among populations and individuals
(Volis 2011).
Acknowledgements
The authors thank Gael Grenouillet for his help doing the statistical analy-
ses, Professor Alain Thomas for Diptera identification, Josselin Cornuault
and Suzanne Liagre for their help in the field. This work received funding
from the CNRS-GDREC (French National Research Group in Chemical
Ecology). We also thank Stefan D€otterl, Artur Maia and an anonymous
reviewer for their constructive comments on the manuscript.
References
Ackerman, J.D., Cuevas, A.A. & Hof, D. (2011) Are deception-pollinated
species more variable than those offering a reward? Plant Systematics
and Evolution, 293, 91–99.Ackerman, J.D., Mel�endez-Ackerman, E.J. & Salguero-Faria, J. (1997)
Variation in pollinator abundance and selection on fragrance phenotypes
in an epiphytic orchid. American Journal of Botany, 84, 1383–1390.Adams, R.P. (2007) Identification of Essential Oil Components by Gas Chro-
matography/Mass Spectroscopy, 4th edn. Allured Publishing, Carol
Stream.
Albre, J. & Gibernau, M. (2008) Reproductive biology of Arum italicum
(Araceae) in the South of France. Botanical Journal of the Linnean
Society, 156, 43–49.Albre, J., Quilichini, A. & Gibernau, M. (2003) Pollination ecology of
Arum italicum (Araceae). Botanical Journal of the Linnean Society, 141,
205–214.Anderson, M.J. (2001) A new method for non-parametric multivariate
analysis of variance. Austral Ecology, 26, 32–46.Anderson, B. & Johnson, S.F. (2007) The geographic mosaic of coevolution
in a plant-pollinator mutualism. Evolution, 62, 220–225.Angert, A.L., Bradshaw, H.D. & Schemske, D.W. (2008) Using experimen-
tal evolution to investigate geographic range limits in monkey flowers.
Evolution, 62, 2660–2675.Ayasse, M., Schiestl, F.P., Paulus, H.F., L€ofstedt, C., Hannson, B., Ibarra, F.
& Francke, W. (2000) Evolution of reproductive strategies in the sexually
deceptive orchid Ophrys sphegodes: how does flower-specific variation of
odor signals influence reproductive success? Evolution, 54, 1995–2006.Blionis, G.J. & Vokou, D. (2002) Structural and functional divergence of
Campanula spatulata subspecies on Mt Olympos (Greece). Plant System-
atics and Evolution, 232, 89–105.Boyce, P.C. (2006) Arum – a decade of change. Aroideana, 29, 132–137.Bradshaw, H.D. & Schemske, D.W. (2003) Allele substitution at a flower
colour locus produces a pollinator shift in monkeyflowers. Nature, 426,
176–178.Brown, M., Downs, C.T. & Johnson, S.D. (2011) Covariation of flower traits
and bird pollinator assemblages among pollinations of Kniphofia linearifo-
lia (Asphodelaceae). Plant Systematics and Evolution, 294, 199–206.Campbell, D.R. (2003) Natural selection in Ipomopsis hybrid zones: impli-
cations for ecological speciation. New Phytologist, 161, 83–90.Chartier, M., Pelozuelo, L. & Gibernau, M. (2011) Do floral odor profiles
geographically vary with the degree of specificity for pollinators? Investi-
gation in two sapromyophilous Arum species (Araceae). Annales de la
Soci�et�e Entomologique de France, 47, 71–77.Cosacov, A., Nattero, J. & Cocucci, A.A. (2008) Variation of pollinator
assemblages and pollen limitation in a locally specialized system: the oil-
producing Nierembergia liariifolia (Solanaceae). Annals of Botany, 102,
723–734.Diaz, A. & Kite, G.C. (2002) A comparison of the pollination ecology of
Arum maculatum and A. italicum in England. Watsonia, 24, 171–181.Dormont, L., Jay-Robert, P., Bessi�ere, J.-M., Rapior, S. & Lumaret, J.-P.
(2010) Innate olfactory preferences in dung beetles. The Journal of
Experimental Biology, 213, 3177–3185.D€otterl, S., Wolfe, L.M. & J€urgens, A. (2005) Qualitative and quantitative
analyses of flower scent in Silene latifolia. Phytochemistry, 66, 203–213.Elle, E. & Carney, R. (2003) Reproductive assurance varies with flower size
in Collinsia parviflora (Scrophulariaceae). American Journal of Botany,
90, 888–896.Ellis, A.G. & Johnson, S.D. (2009) The evolution of floral variation with-
out pollinator shifts in Gorteria diffusa (Asteraceae). American Journal of
Botany, 96, 793–801.
Esp�ındola, A., Pellissier, L. & Alvarez, N. (2011) Variation in the propor-
tion of flore visitors of Arum maculatum along its distributional range in
relation with community-based climatic niche analyses. Oikos, 120,
728–734.Faegri, K. & Van Der Pijl, L. (1971) The Principles of Pollination Ecology.
Pergamon Press, Oxford.
Fenster, C.B., Armbruster, W.S., Wilson, P., Dudash, M.R. & Thomson,
J.D. (2004) Pollination syndromes and floral specialization. Annual
Reviews of Ecology, Evolution and Systematics, 35, 375–403.Geber, M.A. & Eckhart, V.M. (2005) Experimental studies of adaptation
in Clarkia xantiana. II. Fitness variation across a subspecies border.
Evolution, 59, 521–531.Geber, M.A. & Moeller, D.A. (2006) Pollinator responses to plant commu-
nities and implications for reproductive character evolution. Ecology and
Evolution of Flowers (eds L.D. Harder & S.C.H. Barrett), pp. 102–119.Oxford University Press, Oxford.
Gibernau, M. (1997) Odeurs et sp�ecificit�e dans les mutualismes figuier polli-
nisateur: Le cas de Ficus carica L. et de Blastophaga psenes L. PhD
thesis, University of Montpellier, Montpellier.
Gibernau, M., Macquart, D. & Przetak, G. (2004) Pollination in the genus
Arum – a review. Aroideana, 27, 148–166.Gomez, J.M., Bosh, J., Perfectti, F., Fern�andez, J.D., Abdelaziz, M. &
Camacho, J.P.M. (2008) Spatial variation in selection on corolla shape
in a generalist plant is promoted by the preference patterns of its local
pollinators. Proceedings of the Royal Society B, 275, 2241–2249.Gomez, J.M., Abdelaziz, M., Camacho, J.P.M., Munoz-Pajares, A.J. &
Perfectti, F. (2009) Local adaptation and maladaptation to pollinators in
a generalist geographic mosaic. Ecology Letters, 12, 672–682.Gould, S.J. & Johnston, R.F. (1972) Geographic variation. Annual Review
of Ecology, Evolution, and Systematics, 3, 457–498.Hentrich, H., Kaiser, R. & Gottsberger, G. (2010) Floral biology and
reproductive isolation by floral scent in three sympatric Aroid species in
French Guyana. Plant Biology, 12, 587–596.Herrera, C.M., Castellanos, M.C. & Medrano, M. (2006) Geographical
context of floral evolution: towards an improved research programme in
floral diversification. Ecology and Evolution of Flowers (eds L.D. Harder
& S.C.H. Barrett), pp. 278–294. Oxford University Press, Oxford.
Hodgins, K.A. & Barrett, C.H. (2008) Geographic variation in floral mor-
phology and style-morph ratios in a sexually polymorphic daffodil.
American Journal of Botany, 95, 185–195.Ibanez, S., D€otterl, S., Anstett, M.-C., Baudino, S., Caissard, J.C., Gallet,
C. & Despr�es, L. (2010) The role of volatile organic compounds, mor-
phology and pigments of globeflowers in the attraction of their specific
pollinating flies. New Phytologist, 188, 451–463.Je�zek, J. (1990) Redescriptions of nine common palaeartic and holartic
species of Psychodini End. (Diptera: Psychodidae). Acta Entomologica
Musei Nationalis Pragae, 43, 33–83.Je�zek, J. & H�ajek, J. (2007) Psychodidae (Diptera) of the Orlick�e hory
protected landscape area and neighbouring areas with descriptions of
two new species from the Czech Republic. Acta Entomologica Musei
Nationalis Pragae, 47, 237–285.Johnson, S.D. (2006) Pollinator-driven speciation in plants. Ecology and
Evolution of Flowers (eds L.D. Harder & S.C.H. Barrett), pp. 295–310.Oxford University Press, Oxford.
Johnson, S.D. & J€urgens, A. (2010) Convergent evolution of carrion and
faecal scent mimicry in fly-pollinated angiosperm flowers and a stink-
horn fungus. South African Journal of Botany, 76, 796–807.Johnson, S.D. & Steiner, K.E. (1997) Long-tongued fly pollination and evo-
lution of floral spur length in the Disa draconis complex (Orchidaceae).
Evolution, 51, 45–53.J€urgens, A., D€otterl, S. & Meve, U. (2006) The chemical nature of fetid flo-
ral odours in stapeliads (Apocynaceae–Asclepiadoideae–Ceropegieae).New Phytologist, 172, 452–468.
Kato, M., Itioka, T., Sakai, S., Momose, K., Yamane, S., Hamid, A.A. &
Inoue, T. (2000) Various population fluctuation patterns of light-
attracted beetles in a tropical lowland dipterocarp forest in Sarawak.
Population Ecology, 42, 97–104.Kiester, A.R., Lande, R. & Schemske, D.W. (1984) Models of coevolution
and speciation in plants and their pollinators. The American Naturalist,
1242, 220–243.Kite, G.C. (1995) The floral odour of Arum maculatum. Biochemical Sys-
tematics and Ecology, 23, 343–354.Kite, G.C., Hetterscheid, W.L.A., Lewis, M.J., Boyce, P.C., Ollerton, J.,
Cocklin, E., Diaz, A. & Simmonds, M.S.J. (1998) Inflorescence odours
and pollinators of Arum and Amorphophallus (Araceae). Reproductive
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
14 M. Chartier et al.

Biology (eds S.J. Owens & P.J. Rudall), pp. 295–315. Royal Botanical
Gardens, Kew.
Knudsen, J.T. (2002) Variation in floral scent composition within and
between populations of Geonoma macrostachys (Arecaceae) in the wes-
tern Amazon. American Journal of Botany, 89, 1772–1778.Knudsen, J.T., Eriksson, R., Gershenzon, J. & St�ahl, B. (2006) Diversity
and distribution of floral scent. The Botanical Review, 72, 1–120.Lack, A.J. & Diaz, A. (1991) The pollination of Arum maculatum L. – a
historical review and new observations. Watsonia, 18, 333–342.Levin, D.A. (2000) The Origin, Expansion and Demise of Plant Species.
Oxford University Press, Oxford.
Levin, D.A. & Anderson, W.W. (1970) Competition for pollinators between
simultaneously flowering species. The American Naturalist, 104, 455–467.Linz, J., St€okl, J., Urru, I., Kr€ugel, T., Stensmyr, M.C. & Hansson, B.S.
(2010) Molecular phylogeny of the genus Arum (Araceae) inferred from
multi-locus sequence data and AFLPs. Taxon, 59, 405–415.Maia, A.C.D., D€otterl, S., Kaiser, R., Silberbauer-Gottsberger, I., Teichert,
H., Gibernau, M., Navarro, D.M.D.F., Schlindwein, C. & Gottsberger,
G. (2012) The key role of 4-methyl-5-vinylthiazole in the attraction of
scarab beetle pollinators: a unique olfactory floral signal shared by An-
nonaceae and Araceae. Journal of Chemical Ecology, 38, 1072–1080.Maia, A.C.D., Gibernau, M., D€otterl, S., Navarro, D.M.do.A.F., Seifert,
K., M€uller, T. & Schlindwein, C. (2013) The floral scent of Taccarum
ulei (araceae): attraction of scarab beetle pollinators to an unusual ali-
phatic acyloin. Phytochemistry, http://dx.doi.org/10.1016/j.phytochem.
2013.03.005. (in press).
Mant, J., Peakall, R. & Schiestl, F.P. (2005) Does selection on floral odor
promote differentiation among populations and species of the sexually
deceptive orchid genus Ophrys? Evolution, 59, 1449–1463.M�endez, M. & Obeso, J.R. (1992) Influencia del osmoforo en la produccion
de infrutescencias en Arum italicum Miller (Araceae). Anales del Jard�ın
Bot�anico de Madrid, 50, 229–237.Mitchell, R.J., Flanagan, R.J., Brown, B.J., Waser, N.M. & Karron, J.D.
(2009) New frontiers in competition for pollination. Annals of Botany,
103, 1403–1413.Morgan, M.T. (2006) Selection on reproductive characters: conceptual
foundations and their extension to pollinator interactions. Ecology and
Evolution of Flowers (eds L.D. Harder & S.C.H. Barrett), pp. 25–40.Oxford University Press, Oxford.
Nattero, J., Cocussi, A.A. & Medel, R. (2010) Pollinator-mediated selection
in a specialized pollination system: matches and mismatches across
populations. Journal of Evolutionary Biology, 23, 1957–1968.Pettersson, S. & Knudsen, J.T. (2001) Floral scent and nectar production in
Parkia biglobosa Jacq. (Leguminosae: Mimosoideae). Botanical Journal
of the Linnean Society, 135, 97–106.Pichersky, E. & Gershenzon, J. (2002) The formation and function of plant
volatiles: perfumes for pollinator attraction and defense. Plant Biology,
5, 237–243.Prime, C.T. (1960) Lords and Ladies. Harper Collins Publishers, London.
Proctor, M., Leo, P. & Lack, A. (1996) Natural History of Pollination.
Harper Collins Publishers, London.
Raguso, R.A. (2008) Wake up and smell the roses: the ecology and evolu-
tion of floral scent. Annual Review of Ecology, Evolution, and Systemat-
ics, 39, 549–569.R Core Team (2012) R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria
ISBN 3-900051-07-0, URL http://www.R-project.org/University of
Chicago Press, Chicago, IL.
Renner, S.S. (2006) Rewardless flowers in the Angiosperms and the role of
insect cognition in their evolution. Plant–Pollinator Interactions: From
Specialization to Generalization (eds N.M. Waser & J. Ollerton),
pp. 123–144. University of Chicago Press, Chicago, IL.
Rohacek, J., Beck-Haug, I. & Dobat, K. (1990) Sphaeroceridae associated
with flowering Arum maculatum (Araceae) in the vicinity of T€ubingen,
SW-Germany. Senckenbergiana biologie, 71, 259–268.Schaeffer, H.M. & Ruxton, G.D. (2011) Plant-Animal Communication.
Oxford University Press, Oxford.
Schiestl, F.P. & Ayasse, M. (2002) Do changes in floral odor cause speciation in
sexually deceptive orchids? Plant Systematics and Evolution, 234, 111–119.Schiestl, F.P. & Schl€uter, P.M. (2009) Floral isolation, specialized pollina-
tion, and pollinator behavior in Orchids. Annual Review of Entomology,
54, 425–446.
Schlumpberger, B.O. & Raguso, R.A. (2008) Geographic variation in floral
scent of Echinopsis ancistrophora (Cactaceae); evidence for constraints
on hawkmoth attraction. Oikos, 117, 801–814.Schlumpberger, B.O., Cocussi, A.A., Mor�e, M., S�ersic, A.N. & Raguso,
R.A. (2009) Extreme variation in floral characters and its consequences
for pollinator attraction among populations of an Andean cactus. Annals
of Botany, 103, 1489–1500.Solers, C., Hossaert-McKey, M., Buatois, B., Bessi�ere, J.-M., Schatz, B. &
Proffit, M. (2011) Geographic variation of floral scent in a highly special-
ized pollination mutualism. Phytochemistry, 72, 74–81.Stebbins, G.L. (1970) Adaptive radiation of reproductive characteristics in
Angiosperms, I: pollination mechanisms. Annual Review of Ecology and
Systematics, 1, 307–326.St€okl, J., Strutz, A., Dafni, A., Svatos, A., Doubsky, J., Knaden, M.,
Sachse, S., Hansson, B.S. & Stensmyr, M.C. (2010) A deceptive pollina-
tion system targeting drosophilids through olfactory mimicry of yeast.
Current Biology, 20, 1846–1852.St€okl, J., Schl€uter, P.M., Stuessy, T.F., Paulus, H.F., Assum, G. & Ayasse,
M. (2008) Scent variation and hybridization cause the displacement of a
sexually deceptive orchid species. American Journal of Botany, 95,
472–481.Svensson, G.P., Pellmyr, O. & Raguso, R.A. (2006) Strong conservation of
floral scent composition in two allopatric yuccas. Journal of Chemical
Ecology, 32, 2657–2665.Svensson, G.P., Pellmyr, O. & Raguso, R.A. (2011) Pollinator attraction to
volatiles from virgin and pollinated host flowers in a yucca/moth obli-
gate mutualism. Oikos, 120, 1577–1583.Svensson, G.P., Hickman, M.O., Bartram, S., Boland, W., Pellmyr, O. &
Raguso, R.A. (2005) Chemistry and geographic variation of floral scent
in Yucca filamentosa (Agavaceae). American Journal of Botany, 92,
1624–1631.Thompson, J.N. (2005) The Geographic Mosaic of Coevolution. University
Chicago Press, Chicago, IL.
Urru, I., Stensmyr, M.C. & Hansson, B.S. (2011) Pollination by brood-site
deception. Phytochemistry, 72, 1655–1666.Vaillant, F. (1988) Dipt�eres psychodidae coprophiles et coprobiontes d’Eur-
ope. Bulletin Romand d’Entomologie, 6, 1–43.Valiente-Banuet, A., Molina-Freaner, F., Torres, A., del Coro Arizmendi,
M. & Casas, A. (2004) Geographic differentiation in the pollination sys-
tem of the columnar cactus Pachycereus pecten-aboriginum. American
Journal of Botany, 91, 850–855.Volis, S. (2011) Adaptive genetic differentiation in a predominantly self-pol-
linating species analyzed by transplanting into natural environment,
crossbreeding and Qst-Fst test. New Phytologist, 192, 237–248.Waelti, M.O., Muhlemann, J.K., Widmer, A. & Schiestl, F.P. (2008) Floral
odour and reproductive isolation in two species of Silene. Journal of
Evolutionary Biology, 21, 111–121.Waser, N.M. (1978) Interspecific pollen transfer and competition between
co-occurring plant species. Oecologia, 36, 223–236.Waterman, R.J., Bidartondo, M.I., Stofberg, J., Combs, J.K., Gebauer, G.,
Savolainen, V., Barraclough, T.G. & Pauw, A. (2011) The effects of
above- and belowground mutualisms on orchid speciation and coexis-
tence. The American Naturalist, 177, E54–E68.Withers, P. (1989) Roth flies. Diptera: Psychodidae.Dipterist Digest, 4, 1–83.
Received 21 June 2012; accepted 22 April 2013
Handling Editor: Manfred Ayasse
Supporting Information
Additional Supporting information may be found in the online
version of this article:
Table S1. Percentages of insects (mean � SE) caught in the
A. italicum and A. maculatum inflorescences from all studied sites
in 2009 and 2010, and from 2008 (data from Chartier, Pelozuelo
& Gibernau 2011).
© 2013 The Authors. Functional Ecology © 2013 British Ecological Society, Functional Ecology
Geographical variations of Arum pollinators 15
Related Documents











