Palaeogeography, Palaeoclimatology, Palaeoecology, 69 (1989): 245-266 245 Elsevier Science Publishers B.V., Amsterdam -- Printed in The Netherlands GEOCHEMICAL AND PALEOENVIRONMENTAL VARIATIONS ACROSS THE CRETACEOUS/TERTIARY BOUNDARY AT BRAGGS, ALABAMA JAMES C. ZACHOS 1, MICHAEL A. ARTHUR 1 and WALTER E. DEAN 2 ~ Graduate School of Oceanography, University of Rhode Island, Narragansett, RI 02882 (U.S.A.) 2U.S. Geologic Survey, P.O. Box 25046, MS 939, Federal Center, Denver, CO 80225 (U.S.A.) (Received March 9, 1988; revised and accepted August 8, 1988) Abstract Zachos, J. C., Arthur, M.A. and Dean, W.E., 1989. Geochemical and paleoenvironmental variations across the Cretaceous/Tertiary boundary at Braggs, Alabama. Palaeogeogr., Palaeoclimatol., Palaeoecol., 69:245 266. The Cretaceous/Tertiary (K/T) boundary in southern Alabama occurs in a sequence of interbedded shallow-marine limestones and marls deposited during a Late Maastrichtian regression and subsequent Danian transgression. The presence of a diverse assemblage of Cretaceous and Paleocene benthic micro- and macrofossils has allowed detailed examination of paleoenvironmental changes in this shallow-marine setting at the time of the K/T boundary extinctions. Although extensive diagenesis has resulted in the recrystallization and cementation of whole rock carbonate, a few molluscan macrofossils have retained their original calcitic structure. The oxygen isotopic record of these macrofossils shows a gradual cooling of 2 3°C over a 3 m.y. period which began in the late Maastrichtian and continued into the Danian. The characteristic depletion in 513C across the K/T boundary displayed by planktonic microfossils from pelagic sequences is not recorded at Braggs. Major changes in whole rock trace and minor element chemistry reflect the decrease in terrigenous mineral fluxes to this location brought about by the rapid Early Paleocene transgression. Introduction During the last decade the majority of geochemical evidence for marine environmen- tal changes across the Cretaceous/Tertiary boundary has come primarily from DSDP (Deep Sea Drilling Project) and land-based pelagic sequences. With this evidence a num- ber of investigators have shown that dramatic changes in oceanic primary productivity oc- curred coeval with the major marine plankton extinctions as well as with the deposition of chemical anomalies suggestive of a bolide impact event (e.g. Hsfi et al., 1982; Perch- Nielsen et al., 1982; Shackleton and Hall, 1984; Zachos and Arthur, 1986). The carbon isotope evidence upon which the productivity records were based suggest that a sharp reduction in oceanic productivity occurred in a relatively short period (< 10,000 yr). An increasing body of paleontological evidence from shallower marine sequences (Kauffman, 1984; Mount et al., 1986), however, suggests that while the extinctions among the planktonic organisms may have been rapid, the extinction of the marine benthic invertebrates may have occurred in a more gradual, step-like manner, raising serious doubts about the timing of and very nature of the extinctions (Officer and Drake, 1983; Hal- lam, 1987; Hut et al., 1987; Officer et al., 1987). Another unresolved aspect of the K/T transi- tion is the nature of possible climate changes. Paleotemperature reconstructions on the basis of oxygen isotope data from DSDP sequences 0031-0182/89/$03.50 ~(:) 1989 Elsevier Science Publishers B.V.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology, 69 (1989): 245-266 245 Elsevier Science Publishers B.V., Amsterdam - - Printed in The Netherlands
GEOCHEMICAL AND PALEOENVIRONMENTAL VARIATIONS ACROSS THE CRETACEOUS/TERTIARY BOUNDARY AT
BRAGGS, ALABAMA
J A M E S C. ZACHOS 1, M I C H A E L A. A R T H U R 1 and W A L T E R E. D E A N 2
~ Graduate School of Oceanography, University of Rhode Island, Narragansett, RI 02882 (U.S.A.) 2U.S. Geologic Survey, P.O. Box 25046, MS 939, Federal Center, Denver, CO 80225 (U.S.A.)
(Received March 9, 1988; revised and accepted August 8, 1988)
Abstract
Zachos, J. C., Arthur, M.A. and Dean, W.E., 1989. Geochemical and paleoenvironmental variations across the Cretaceous/Tertiary boundary at Braggs, Alabama. Palaeogeogr., Palaeoclimatol., Palaeoecol., 69:245 266.
The Cretaceous/Tertiary (K/T) boundary in southern Alabama occurs in a sequence of interbedded shallow-marine limestones and marls deposited during a Late Maastrichtian regression and subsequent Danian transgression. The presence of a diverse assemblage of Cretaceous and Paleocene benthic micro- and macrofossils has allowed detailed examination of paleoenvironmental changes in this shallow-marine setting at the time of the K/T boundary extinctions.
Although extensive diagenesis has resulted in the recrystallization and cementation of whole rock carbonate, a few molluscan macrofossils have retained their original calcitic structure. The oxygen isotopic record of these macrofossils shows a gradual cooling of 2 3°C over a 3 m.y. period which began in the late Maastrichtian and continued into the Danian. The characteristic depletion in 513C across the K/T boundary displayed by planktonic microfossils from pelagic sequences is not recorded at Braggs. Major changes in whole rock trace and minor element chemistry reflect the decrease in terrigenous mineral fluxes to this location brought about by the rapid Early Paleocene transgression.
Introduct ion
D u r i n g the l as t decade the m a j o r i t y of
g e o c h e m i c a l ev idence for m a r i n e env i ronmen-
ta l changes ac ross the C r e t a c e o u s / T e r t i a r y
b o u n d a r y has come p r i m a r i l y f rom DSDP
(Deep Sea D r i l l i n g P ro jec t ) and l and-based
pe lag ic sequences . W i t h th i s ev idence a num-
ber of i n v e s t i g a t o r s have shown t h a t d r a m a t i c
changes in ocean i c p r i m a r y p r o d u c t i v i t y oc-
cu r r ed coeva l w i th the m a j o r m a r i n e p l a n k t o n
e x t i n c t i o n s as wel l as wi th the depos i t i on of chemica l a n o m a l i e s sugges t i ve of a bo l ide
impac t even t (e.g. Hsfi et al., 1982; Perch- Nie l sen et al., 1982; S h a c k l e t o n and Hal l , 1984;
Zachos and A r t h u r , 1986). The c a r b o n i so tope ev idence upon wh ich the p r o d u c t i v i t y r eco rds
were based sugges t t h a t a sha rp r educ t ion in
ocean ic p r oduc t i v i t y occu r red in a r e l a t ive ly
shor t per iod ( < 10,000 yr). An inc reas ing body of
pa l e on t o l og i c a l ev idence from sha l lower mar ine
sequences (Kauffman, 1984; M o u n t et al., 1986),
however , sugges ts t ha t whi le the ex t inc t ions
among the p l a n k t o n i c o rgan i sms may have been rapid , the ex t inc t ion of the mar ine ben th ic
i n v e r t e b r a t e s may have occur red in a more g radua l , s tep- l ike manner , r a i s ing ser ious
doubts abou t the t iming of and very n a t u r e of
the ex t inc t ions (Officer and Drake , 1983; Hal- lam, 1987; Hu t et al., 1987; Officer et al., 1987).
A n o t h e r u n r e s o l v e d a spec t of the K/T t rans i - t ion is the n a t u r e of poss ib le c l i m a t e changes .
P a l e o t e m p e r a t u r e r e c o n s t r u c t i o n s on the bas is of oxygen i so tope d a t a from DSDP sequences
0031-0182/89/$03.50 ~(:) 1989 Elsevier Science Publishers B.V.

246
have proved contradictory, providing neither a consistent picture of nor any evidence of major global temperature trends or events at or near the K/T boundary. The few available isotopic paleotemperature records from land-based hemipelagic and shallow-marine sequences sug- gest large excursions across the K/T boundary (Perch-Nielsen et al., 1982; Margolis et al., 1987). The oxygen isotope data upon which these temperature estimates are based, how- ever, may be compromised by diagenetic re- crystallization.
The available evidence suggests that the rate and magnitude of environmental change may have differed between pelagic and shal- low-marine environments. However, because very little adequate stable isotope and other geochemical data exists for shallow shelf sequences spanning the K/T boundary we have been unable to make direct comparisons of the environmental and paleontological changes that have been recorded for the open ocean with those that may have occurred on the shelf.
The main purpose of this investigation was to determine if the changes in productivity and temperature recorded in pelagic sequences could be recognized in shallow-marine se- quences, and if so, what relationship these changes bear in terms of timing to the main marine invertebrate extinctions. Employing the "holistic" approach utilized during in- vestigations of DSDP boundary sequences, high-resolution geochemical (this paper), paleontologic (Bryan and Jones, this issue), and paleomagnetic records (Channell and Dob- son, this issue) were assembled for a shallow marine K/T boundary sequence located in Braggs, southern Alabama (Fig.l), an initial summary of which was provided by Jones et al. (1987). We chose to focus our study on this particular sequence for several reasons. First, a well developed biostratigraphy, including both macro- and microfauna and -flora, was already established by previous investigators (Cepek et al., 1968; Worsley, 1974; Jarzen, 1978, Smith, 1978; Thierstein, 1981; Donovan et al., in press). Secondly, a sea level history has been developed for the region (Donovan et al., in
Lowndes County, Alabama
Mootevi l lc Chalk
Dcmopol is Chalk
Riplcy Format ion 5 km
Prairie Bluff Chalk
Providence Sand
Clayton Format ion
Alabama
! . t / ' 'x , ~
.r. ] ~ Ala.2l
i . , . . . . . ' " " " ' 1
i ' . B R A G G S
i.":. .... t : ~ ' . . . . . . . . . . . . i .... , . . . . . . . . . . . : /
%
Ala, 2 6 3
Fig.1. The K/T boundary exposure located along Alabama State highway 263 in Lowndes County, on the south side of the roadway about 7.4 km southeast of Braggs, Alabama. The location of Lowndes County is shown in the inset map. The outcrop occurs along the contact between the Clayton Formation (Early Paleocene) to the southwest and the Prairie Bluff Formation (Late Cretaceous) to the northeast (after Scott, 1968; Copeland and Mancini, 1986; Donovan et al., in press).
press) perhaps allowing us to separate events related to the environmental effects of sea level change from other processes. And finally, Braggs is one of the few nearly complete K/T sections representative of a shallow shelf environment available.
This contribution presents a detailed de- scription and summary of the geochemical results of that investigation. The dataset includes stable carbon and oxygen isotope, total carbonate and inorganic geochemical data from whole rock samples as well as isotope data from individual species of calcare- ous macro- and microfossils. In addition, a petrographic investigation was conducted to elucidate sediment composition and diagenetic history. The carbon isotope and faunal/floral data are used to explore possible changes in regional fertility and their association with the K/T boundary biotic extinctions. A paleotem- perature curve is developed from the benthic foraminifera and macrofossil oxygen isotope profiles. In addition, major and minor element

chemistry are utilized to characterize changes in sediment mineral assemblages across the boundary. The changes in composition and mass flux of detri tal minerals to the region reflect local or regional variat ions in r iverine input, sea level and/or climate.
M e t h o d s
Carbon and oxygen stable isotope analyses were conducted on bulk sediment, <53 and < 31 ~tm carbonate fractions, benthic foramini- fera and macrofossils (predominantly oysters). Samples were disaggregated in warm calgon solution, wet sieved through 53 ~m sieves, and then washed. The <31 pm fraction was separated by settling techniques (Paull and Thierstein, 1987). Benthic foraminifera were sonicated in methyl alcohol to remove adher- ing particles.
Calcareous samples, which had been roasted in vacuo at 390°C for 1 hr, were reacted in 100% phosphoric acid at 50°C. The resulting CO~ was analyzed in a VG micromass 602D mass spectrometer. All data are corrected for machine effects (Craig, 1957) and reported in per mil (%o) notation relative to the PDB stand- ard. Precision of duplicate analyses was bet ter than + 0.05%0 oxygen and _0.03%o for carbon.
Carbonate contents were determined by the carbonate bomb method as outlined by Mfiller and Gastner (1971). Whole rock samples were analyzed for 9 major and minor elements by X-ray fluorescence (XRF) (Taggert et al., 1980) and 19 trace elements by induction coupled argon plasma spectrometry (ICP) (Floyd et al., 1980). Individual samples were ground to less than 63 pm. Data are reported as % oxides for major and minor elements and ppm for t race elements.
D e p o s i t i o n a l s e t t i n g a n d l i t h o s t r a t i g r a p h y
During the Late Cretaceous/Early Tert iary, shallow seas covered most of what is now the Gulf Coastal Plain and deposited relat ively thick sequences of marine shelf chalks and sands from Texas to Florida. In southern
247
Alabama these sequences strike in a general east-west direction and dip seaward with the oldest deposits exposed toward the north (Fig.l). From west to east there is a gradation from chalk sequences into sands. The majority of terr igenous sand and clay sediments were supplied from the north (for a more detailed description see Scott, 1968; Copeland and Mancini, 1986).
The sedimentary sequence exposed at Braggs, Alabama consists of two principal units, the upper portion of the Prairie Bluff Chalk (Late Cretaceous) and the Pine Barren Member of the Clayton Formation (Paleocene). The Prair ie Bluff Chalk extends up to the middle of bed 3 in our measured section (Fig.2; See Copeland and Maneini, 1986 for original strat igraphy) and generally consists of dark- gray to black silty to sandy, micaceous, cal-
Braggs, Alabama
(m) ~ ~ Zones ~ 5 1 2 3 4 5 1 6 1 7
-_-~_-
3 - " ~ " "~ " _ _ ~. . - . :-:';':
• - . ., =. ,4 ~
1 ~ ~ c . . . - . . _
• ~ ' v
"~ ~ ~ " = ~ = "r. ~ ;~ '~ :~-.--0-.2 .~-. ~"" ~ ~,~ ~ ~ = % -
-2 ~ ~ .d ~ ---2"~ - 1 . & 2. C o p e l a n d & M a n c i n i (1986) 3. C o p e l a n d & M a n c i n i (1986) 4. 5. Donovan et al. (in press) 6. Channell & Dobson (1988) 7.
L,,
(m) 5
--4
:q':-~:_ [ ] C~ay ~'Z6-2~:5 - - 3 ~ Calcareous
• ~ Clay
[ ] Sa~ 2 [ ] Limestone
f"~'] Macrofossils
- - I [ ] Glauconitc
' ~ Phosphale Nodules
- 0
--I
-2
Worsley (1974) according to Donovan et al. (in press)
Modified from Copeland & Mancini (1986); Jones et al., (1986)
Fig.2. Lithobiomagnetostratigraphy of the Braggs se- quence, Alabama (Worsley, 1974; Copeland and Mancini, 1986; Jones et al., 1986; Channel and Dobson, 1988; Donovan et al., in press).
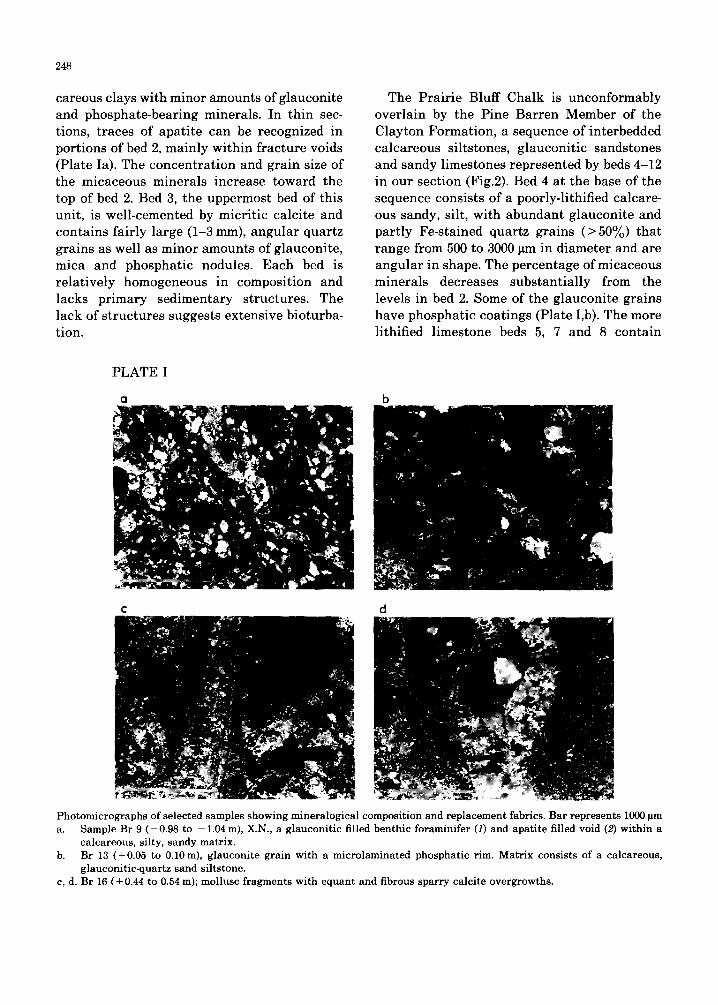
248
careous clays with minor amounts of g lauconi te and phospha te -bear ing minerals . In th in sec- tions, t races of apa t i t e can be recognized in por t ions of bed 2, main ly wi th in f r ac tu re voids (Plate Ia). The c oncen t r a t i on and gra in size of the micaceous minera l s increase toward the top of bed 2. Bed 3, the uppermos t bed of this unit , is wel l -cemented by micr i t ic calc i te and conta ins fa i r ly large (1-3 mm), angu l a r quar tz grains as well as minor amoun t s of g lauconi te , mica and phospha t ic nodules. Each bed is re la t ive ly homogeneous in composi t ion and lacks p r imary sed imen ta ry s t ruc tures . The lack of s t ruc tu re s suggests ex tens ive bioturba- t ion.
The Pra i r i e Bluff Chalk is unconfo rmab ly over la in by the P ine Ba r r en Member of the Clay ton Format ion , a sequence of in te rbedded ca lcareous si l tstones, g lauconi t i c sands tones and sandy l imestones represen ted by beds 4-12 in our sect ion (Fig.2). Bed 4 at the base of the sequence consists of a poorly-l i thif ied calcare- ous sandy, silt, wi th ab u n d an t g lauconi te and par t ly Fe-s ta ined quar tz grains ( > 5 0 % ) tha t r ange from 500 to 3000 pm in d iamete r and are angu la r in shape. The pe rcen tage of micaceous minera l s decreases subs tan t ia l ly from the levels in bed 2. Some of the g lauconi te grains have phospha t ic coat ings (Plate I,b). The more li thified l imestone beds 5, 7 and 8 conta in
P L A T E I
o b
¢ d
Photomicrographs of selected samples showing mineralogical composition and replacement fabrics. Bar represents 1000 pm a. Sample Br 9 (-0.98 to -1.04 m), X.N., a glauconitic filled benthic foraminifer (1) and apatite filled void (2) within a
calcareous, silty, sandy matrix. b. Br 13 (+ 0.05 to 0.10 m), glauconite grain with a microlaminated phosphatic rim. Matrix consists of a calcareous,
glauconitic-quartz sand siltstone. c, d. Br 16 (+0.44 to 0.54 m); mollusc fragments with equant and fibrous sparry calcite overgrowths.

a b u n d a n t macro- and microfossi ls . The shell m a t e r i a l of the macrofoss i l s , p r i m a r i l y mol- luscs, is mos t ly recrys ta l l ized . The r e p l a c e m e n t fabr ics cons is t of o v e r g r o w t h s and f ibrous and e q u a n t spa r ry ca lc i te infi l l ings of d i s so lu t ion voids (P la tes I,c,d and II,a). As in lower beds, the m a t r i x m a t e r i a l is a c a l ca r eous muds tone . Loba t e and v e r m i c u l a r g l aucon i t e grains , which occur as pel le ts and infi l l ings of ben th ic fo ramin i fe ra , a re fa i r ly a b u n d a n t in these beds, cons t i t u t i ng up to 10% of the to ta l ma t e r i a l (see P la te s Ia, and II,b,c). Micas a re v i r tua l ly absen t f rom these more l i thified units . In beds 7 and 8, the g l aucon i t e g ra ins exhibi t i nc reas ing ly more Fe-oxide s ta in ing . In ~dditlnn, the a m o u n t and a v e r a g e gra in size of
P L A T E II
a b
249
quar tz g ra ins in these beds d imin ishes con- s iderably . Ev idence of b u r r o w i n g and the r e m a i n s of se rpul id worm tubes are found n e a r the tops of beds 5, 7 and 8 (Pla te II,d). Our samples of bed 9 a re h igh ly wea the red . Fe-oxide s t a in ing is common on mos t grains , espec ia l ly g l aucon i t e (Pla te II,c). Bed 10 con- sists of unconso l ida t ed g laucon i t i c sands t h a t make -up more t h a n 75% of the to ta l sed iment t o w a r d the top of the bed. Bed 11, a s l ight ly more l i thified and ca l ca r eous unit , also con- t a ins la rge a m o u n t s of g lauconi te . The major- i ty of the g l aucon i t e a t Braggs appea r s to h a v e been reworked . Almos t all the g ra ins a re oval in shape and well rounded, poss ibly ind ica t ing smoo th ing f rom ag i t a t i on and /or
d
Photomicrographs of selected samples showing mineralogical composition and replacement fabrics. Bar represents 1000 ~m. a. Br 16 (+ 0.44 to 0.54 m), X.N., mollusc shell fragments with sparry and microsparry calcite replacement texture. b. Br 23 (+ 1.22 to 1.30 m), X.N., vermicular and lobate shaped glauconite grains. c. Br 26 (+ 1.82 to 2.01 m), X.N, Fe-oxide coated glauconite grains. d. Br 18 (÷ 0.52 to 0.60 m), X.N., serpulid worm tube filled with assorted debris.

250
transport. In addition, the size range of glau- conite grains, especially in the sandy layers, is small implying sorting by wave or tidal action. Bed 12 is composed of a mineral assemblage similar to that found near the top of bed 2 described earlier•
Jones et al. (1987) and Donovan et al (in press) have interpreted the succession of de- posits at Braggs in terms of sea level change suggesting that the contact between the Prairie Bluff and Clayton Formation, which locally shows signs of erosional t runcat ion and sub-aerial exposure, was produced during a eustatic fall in the Late Maastrichtian. The beds above, predominantly muddy sandstones and somewhat more pelagic fossiliferous lime- stones, were probably deposited during a subsequent step-like transgression which re- duced the amount of terrigenous sediment reaching the locality. The maximum point of the high stand is suggested to be marked by the
deposition of glauconite rich bed 10 (Donovan et al., in press)•
Carbonate-stratigraphy
For the 6 m interval analyzed here in some detail, carbonate contents of bulk samples vary from 15 to 75% (Fig.3). The more lithified beds 3, 7, 8, 9 and 11 have average carbonate contents of greater than 50% whereas the carbonate contents of the unlithified sandy beds 2, 4, 6, 10 and 12 average less than 30%•
Chronostratigraphy
An adequate chronostrat igraphy is neces- sary in order to compare the timing of environ- mental changes that occur at Braggs with those that occurred in the open ocean. In pelagic sequences the position of the boundary has been constrained by planktonic foramini-
~. ~ 4
7 . . . . . . . . . 2
8 ",:,i,:,:,:I
o
m
-2 0 20 40 60 80 ~Clay [ ] Calcareo~ [ ] Macrofossils
Clay [ ] Cl~.~o.i~ % CaCO3 [ ] sa~
Limestone [ ] NodulesPh°sphate
Braggs, Alabama
-5
Bulk Robulus Gavelinella
i
• -~, -3 " -2 ' "I " I () I
618 O (PDB) 613 C (PDB)
• -e-. Molluscs [] <53 um Frac X <31urn Frac
Fig.3. Age, and lithology, percent CaCO 3 and oxygen and carbon stable isotope geochemistry plotted versus depth intervals (m) relative to the contact between the Prairie Bluff Formation and the Pine Barren member of the Clayton Formation. Stable isotope data are of whole rock, macrofossils, benthic foraminifera (Robulus and Gavelinella), and < 53 and < 31 I~m calcareous size fractions.

fera and nannofossil zonations (e.g. Thierstein, 1981; Smit, 1982). Cepek and others (1968) indicated that the uppermost Cretaceous planktonic foraminifer Abathomphalus maya- roensis is not present at Braggs and thus they ultimately assigned the Prairie Bluff Forma- tion to the Middle Maastr icht ian Globotrun- cana gansseri zone (Fig.2). However, based on studies of the abundant nannofossil assem- blages, Worsley (1974) and Thierstein (1981) estimated a Late Maastr icht ian age for the upper portion of the Prairie Bluff unit. Species representative of the upper Maastrichtian, Nephrolithus frequens and Micula murus as well as Lithraphidites quadratus, are all pres- ent at the top of the Prairie Bluff Chalk and indicate that the hiatus in the Late Maas- tr ichtian at the top of bed 2 represents less than 0.5 x l06 yr.
Cepek and others (1968) also noted that fossils representative of the earliest Paleocene planktonic foraminifer zone, Globigerina eugu- bina, are not present in the lowermost Pine Barren Member, whereas species which have been assigned to the Subbotina pseudobuUo- ides zone, such as Globoconusa daubjergensis and Subbotina triloculinoides are common. In contrast, nannofossils representative of the earliest Paleocene nannoplankton zones (NP-1 and NP-2 zones of Berggren et al., 1985) such as Zygodiscus sigmoides, Markalius inversus, Thoracosphaera and Cruciplacolithus tenuis are present in the Pine Barren Member of the Clayton Formation (Fig.2) (Worsley, 1974; Thierstein, 1981; Donovan et al., in press) indicating that the K/T transit ion is nearly continuous.
The absence of various index planktonic foraminifera across the K/T boundary at Braggs is not unique, having been recognized at a second Gulf coast shallow marine K/T boundary sequence located along the Brazos River, Texas (Hansen et al., 1987). The similar- ity in the depositional environments of these two locations indicates that the absence of certain species may have resulted from en- vironmental conditions which prevented cer- tain planktonic foraminifera from inhabiting
251
local surface waters. For example, A. mayaro- ensis is likely a deeper dwelling species (e.g. Boersma, 1984) which may have been excluded from the shallower shelf waters that character- ized the Late Cretaceous depositional environ- ments of Braggs and Brazos River (Hansen et al., 1987). In addition, preservational changes may be responsible for the absence of certain species of foraminifera such as G. eugubina. This is suggested by the gradual upward increase in the abundance of solution-resistant calcareous nannoplankton which Thierstein (1981) asserts is an artifact of dissolution.
Although it is generally accepted that nan- nofossils representative of the NP-1 zone are present in the Pine Barren Member, there appears to have been some confusion as to the exact placement of the first coccoliths of Paleocene age (Zone NP-1). We have used the age assignments of Donovan et al. (in press) because they have carefully tied their sampling to the l i thostratigraphy of Copeland and Man- cini (1986) to which we have referred our own observations. In the scheme of Donovan et al. (in press), the nannofossil K/T boundary is placed at the top of Bed 5 (Fig.2), approxi- mately 100 cm above the lithologic break at the top of bed 2. Although Worsley (1974) and Thierstein (1981) have studied the calcareous nannofossils in some detail, it is difficult from the information available to relate their micro- paleontologic observations directly to the li thostratigraphy. This is problematic because it would appear from the Worsley (1974) study that he picked the K/T boundary at the top of bed 2 (Fig.2). However, Donovan et al. (in press), after having closely reviewed the data of Worsley (1974), suggested that Worsley's original placement of the boundary is near that of theirs at the top of bed 5 (Fig.2). This selection places the nannofossil K/T boundary just above the boundary of Copeland and Mancini 's (1986) which was placed within bed 4 on the basis of planktonic foraminifera (Fig.2). However, as mentioned above the discrepancy between the nannofossil and foraminifera stra- tigraphies is likely an artifact of local environ- mental effects which excluded certain index

252
foraminifera from inhabiting the region as well as selective dissolution which may have re- moved solution susceptible fossils. Thus, it appears that the Prairie Bluff Formation and lowermost 60 cm of the Pine Barren Member as defined by Copeland and Mancini (1986) are late Maastr icht ian in age.
Jones et al. (1986) and Bryan and Jones (this issue) demonstrated that the majority of Creta- ceous ammonites, as well as other molluscan genera, disappear abruptly in the transit ion zone between beds 2 and 3. Although greater than half these species fail to reappear any- where in the Paleocene, the discrepancy in timing between the molluscan and nanno- plankton disappearance in the Braggs se- quence apparently results not from a discor- dant extinction event, but primarily from a local facies change related to a rapid drop in sea level (Bryan and Jones, this issue; Donovan et al., in press).
Paleomagnetic data of Channell and Dobson (this issue) confirm the relative position of the boundary as determined by the Donovan et al. (in press) nannofossil data. A reversed polarity zone extends from the upper part of bed 2 to the bottom of bed 9 and is confirmed to represent reversed magnetic Chron 29 (Fig.2) by the presence of M. murus and N: frequens. In comparison with paleomagnetic strat igraphy from other K/T sequences, a disproportionate part of the reversed zone occurs above the nannofossil boundary lending further support for erosional t runcat ion of part of the Late Maastrichtian sequence, possibly during the brief late Maastrichtian regression or the sub- sequent rapid transgression (Jones et al., 1987).
S e d i m e n t a t i o n rates
Utilizing the above biomagnetostratigraphy and the time scales of Berggren et al. (1985) and Kent and Gradstein (1985), Maastr icht ian and Danian sedimentation rates were estab- lished for the Braggs locality. Because of the absence of index planktonic foraminifera and the normal correlation problems associated with unconformities using benthic fossils, we
have based all age estimates on the nannofossil stratigraphy. No corrections are made for physical and chemical compaction. Using the FAD of L. quadratus ( -34.0m; Thierstein, 1981) and assuming a maximum hiatus in deposition of 0.5 x 106 yr, a rate of 8.1 m/m.y. was derived for the Late Maastrichtian. Alter- natively, if we ignore the hiatus the rate would be slightly less at 7.2 m/m.y. For the interval from the base of NP-2 (÷ 3.10 m; Worsley, 1974; Donovan et al., in press) to the K/T boundary (+ 0.60 m; Donovan et al., in press) a sedimen- tation rate of 5.0 m/m.y, was estimated, in good agreement with that estimated by Channell and Dobson (this issue). Utilizing the magnetic stratigraphy they determined a sedimentation rate of 4.6 (_1.75) m/m.y, over the interval represented by Chron 29R.
The relatively low sedimentation rates re- corded at Braggs clearly reflect reduced rate of clastic input during the Early Paleocene transgression and, would help to explain the extensive bioturbation and mineralization (e.g. glauconite and phosphorite) that is observed there. Unfortunately, a result of these low sedi- mentation rates is the lack of a high resolution record of environmental change. However, be- cause of the relative completeness of this sequence, it does enable us to examine longer term changes in shallow-marine paleotempera- tures, paleoecology and fossil evolution that occur in response to the K/T boundary event.
Stable i sotope results
The oxygen isotopic ratios of the bulk samples display large amplitude variations ranging from - 4.5 to - 1.0%oo up section (Fig.3). Samples from the calcareous, sandy bed 2 of the Prairie Bluff Chalk yielded values in the range of -2 .8 to -1.7%o, whereas highly depleted values of -4 .5 to -3.0%0 were re- corded for samples from overlying lithified, more carbonate-rich bed 3. This pattern of 180 enriched values from the unconsolidated sandy beds and depleted values from the more indu- rated, carbonate-rich units is repeated in overlying beds. Although not as obvious, a
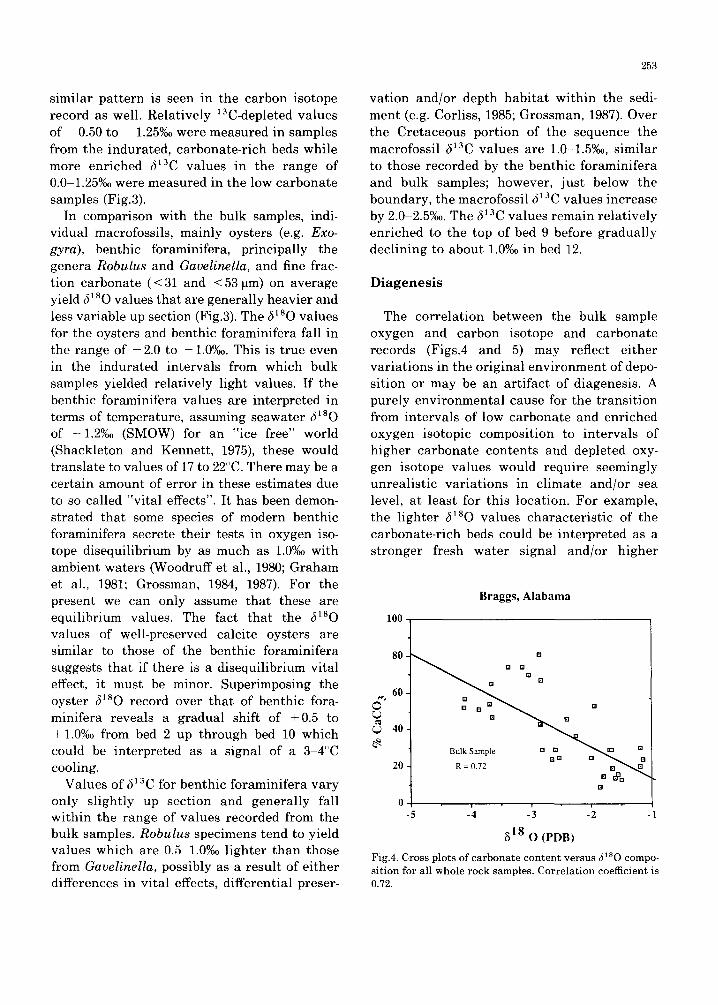
similar pat tern is seen in the carbon isotope record as well. Relatively 13C-depleted values of - 0.50 to - 1.25%o were measured in samples from the indurated, carbonate-rich beds while more enriched 513C values in the range of 0.0 1.25%o were measured in the low carbonate samples (Fig.3).
In comparison with the bulk samples, indi- vidual macrofossils, mainly oysters (e.g. Exo- gyra), benthic foraminifera, principally the genera Robulus and Gavelinella, and fine frac- tion carbonate (< 31 and < 53 pm) on average yield 51 sO values that are generally heavier and less variable up section (Fig.3). The 51sO values for the oysters and benthic foraminifera fall in the range of -2 .0 to -1.0%0. This is true even in the indurated intervals from which bulk samples yielded relatively light values. If the benthic foraminifera values are interpreted in terms of temperature, assuming seawater (~lsO of -1.2%o (SMOW) for an "ice free" world (Shackleton and Kennett , 1975), these would translate to values of 17 to 22°C. There may be a certain amount of error in these estimates due to so called "vital effects". It has been demon- strated that some species of modern benthic foraminifera secrete their tests in oxygen iso- tope disequilibrium by as much as 1.0%o with ambient waters (Woodruff et al., 1980; Graham et al., 1981; Grossman, 1984, 1987). For the present we can only assume that these are equilibrium values. The fact that the ~ sO values of well-preserved calcite oysters are similar to those of the benthic foraminifera suggests that if there is a disequilibrium vital effect, it must be minor. Superimposing the oyster ~lsO record over that of benthic fora- minifera reveals a gradual shift of +0.5 to + 1.0%o from bed 2 up through bed 10 which could be interpreted as a signal of a 3 4°C cooling.
Values of 513C for benthic foraminifera vary only slightly up section and generally fall within the range of values recorded from the bulk samples. Robulus specimens tend to yield values which are 0.5 1.0%0 lighter than those from Gavelinella, possibly as a result of ei ther differences in vital effects, differential preser-
253
vation and/or depth habitat within the sedi- ment (e.g. Corliss, 1985; Grossman, 1987). Over the Cretaceous portion of the sequence the macrofossil 51sC values are 1.0-1.5%0o, similar to those recorded by the benthic foraminifera and bulk samples; however, just below the boundary, the macrofossil ~13C values increase by 2.0 2.5%0. The 513C values remain relatively enriched to the top of bed 9 before gradually declining to about 1.0%o in bed 12.
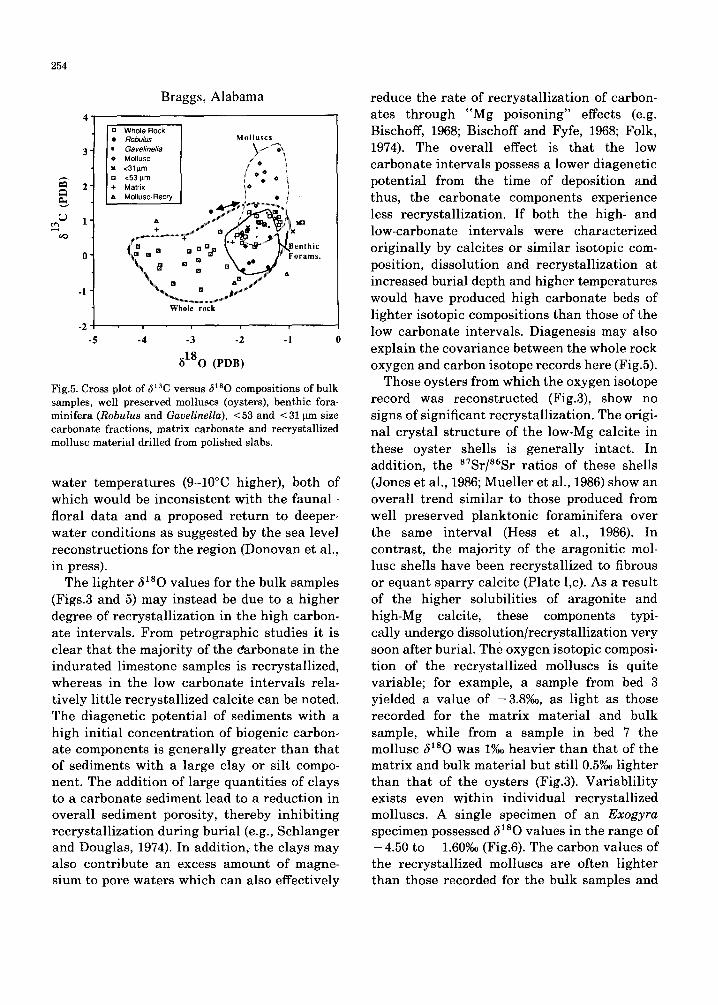
Diagenesis
The correlat ion between the bulk sample oxygen and carbon isotope and carbonate records (Figs.4 and 5) may reflect either variat ions in the original environment of depo- sition or may be an art ifact of diagenesis. A purely environmental cause for the transi t ion from intervals of low carbonate and enriched oxygen isotopic composition to intervals of higher carbonate contents and depleted oxy- gen isotope values would require seemingly unrealist ic variat ions in climate and/or sea level, at least for this location. For example, the lighter ~lsO values character is t ic of the carbonate-rich beds could be interpreted as a stronger fresh water signal and/or higher
100
Braggs, Alabama
80
60
~.~ 40
20
[] [][]
[] [] []
[] ca : [] ca
Bulk Sarnple c a : [ ] • ml
-5 -4 -3 -2 -1
518 O (PDB)
Fig.4. Cross p lots of c a r b o n a t e c o n t e n t v e r s u s ~lsO compo- s i t ion for all whole rock samples . Cor re l a t i on coefficient is 0.72.
254
Braggs, Alabama
4 "
3 " : Flobulus M o l l u s c s Gavelinolla ~ r ~ " - ' - Mollusc f e <31~m / <53 ~m o : •
2 Matrix ie Mollusc-Recry f e
°
1
II1%; ° A MM B ~l~"' -
W h o l e r o c k
- 2 i i i r
-5 -4 -3 -2 -1 0
5 1 8 0 (PDB)
Fig.5. Cross plot of &13C versus ~180 compositions of bulk samples, well preserved molluscs (oysters), benthic fora- minifera (Robulus and Gavelinella), < 53 and < 31 ttm size carbonate fractions, matr ix carbonate and recrystallized mollusc mater ial drilled from polished slabs.
water temperatures (9-10°C higher), both of which would be inconsistent with the faunal - floral data and a proposed return to deeper- water conditions as suggested by the sea level reconstructions for the region (Donovan et al., in press).
The lighter ~lSO values for the bulk samples (Figs.3 and 5) may instead be due to a higher degree of recrystallization in the high carbon- ate intervals. From petrographic studies it is clear that the majority of the darbonate in the indurated limestone samples is recrystallized, whereas in the low carbonate intervals rela- tively little recrystallized calcite can be noted. The diagenetic potential of sediments with a high initial concentrat ion of biogenic carbon- ate components is generally greater than that of sediments with a large clay or silt compo- nent. The addition of large quantities of clays to a carbonate sediment lead to a reduction in overall sediment porosity, thereby inhibiting recrystallization during burial (e.g., Schlanger and Douglas, 1974). In addition, the clays may also contribute an excess amount of magne- sium to pore waters which can also effectively
reduce the rate of recrystallization of carbon- ates through "Mg poisoning" effects (e.g. Bischoff, 1968; Bischoff and Fyfe, 1968; Folk, 1974). The overall effect is that the low carbonate intervals possess a lower diagenetic potential from the time of deposition and thus, the carbonate components experience less recrystallization. If both the high- and low-carbonate intervals were characterized originally by calcites or similar isotopic com- position, dissolution and recrystallization at increased burial depth and higher temperatures would have produced high carbonate beds of lighter isotopic compositions than those of the low carbonate intervals. Diagenesis may also explain the covariance between the whole rock oxygen and carbon isotope records here (Fig.5).
Those oysters from which the oxygen isotope record was reconstructed (Fig.3), show no signs of significant recrystallization. The origi- nal crystal s tructure of the low-Mg calcite in these oyster shells is generally intact. In addition, the STSr/S6Sr ratios of these shells (Jones et al., 1986; Mueller et al., 1986) show an overall trend similar to those produced from well preserved planktonic foraminifera over the same interval (Hess et al., 1986). In contrast, the majority of the aragonitic mol- lusc shells have been recrystallized to fibrous or equant sparry calcite (Plate I,c). As a result of the higher solubilities of aragonite and high-Mg calcite, these components typi- cally undergo dissolution/recrystallization very soon after burial. The oxygen isotopic composi- tion of the recrystallized molluscs is quite variable; for example, a sample from bed 3 yielded a value of -3.8%0, as light as those recorded for the matrix material and bulk sample, while from a sample in bed 7 the mollusc &lSO was 1%o heavier than that of the matrix and bulk material but still 0.5000 lighter than that of the oysters (Fig.3). Variablility exists even within individual recrystallized molluscs. A single specimen of an Exogyra specimen possessed &1so values in the range of
- 4.50 to - 1.60%o (Fig.6). The carbon values of the recrystallized molluscs are often lighter than those recorded for the bulk samples and

255
Pelecypod: Exogyra Upper Maastrichtian
N;q [] Voids filled with "fringe cements"
Matrix, Cemented Siltstone
[] Dark calcite bands [ ] Light calcite bands
/
Fig.6. Cross section of an Exogyra shell showing the locations of samples drilled for stable isotope analyses. The upper and lower values in each box represent 51sO and ~13C, respectively, in %o relative to PDB standard. The shell was recovered from an Upper Maastrichtian bed and consists of alternating light and dark bands with fringe cement lined voids. The surrounding matrix material from which the shell was recovered, a calcareous siltstone, was also analyzed.
for well p rese rved mol luscs ind ica t ing t h a t the d i s so lu t i on / r ep l acem en t of the a r a g o n i t i c species m a y h a v e occu r r ed du r ing ea r ly bur ia l , n e a r the s e d i m e n t / w a t e r in t e r face and in the p resence of decay ing o rgan ic ma t t e r .
A l though the d iagene t i c h i s to ry is compli- ca ted and not comple te ly resolved, i t appea r s t h a t the bu lk sample i so topic v a r i a t i o n s m a y be more an a r t i f ac t of d i f fe ren t ia l d iagenes i s r a t h e r t h a n p r i m a r y e n v i r o n m e n t a l s ignals . Some molluscs , p r e d o m i n a n t l y the pe lecypod oysters , h a v e r e t a ined the i r o r ig ina l i so topic compos i t ion whi le those mol luscs which h a v e u n d e r g o n e r ep lacemen t , m a y have done so ve ry soon a f t e r bur ia l .
Major and m i n o r e l e m e n t g e o c h e m i s t r y
The re l a t ive c o n c e n t r a t i o n s of m a j o r and mino r e l ements m a y be used as an ind ica to r of m ine r a log i ca l va r i a t ion . V a r i a t i o n s in m i ne ra l a s semblages of s e d i m e n t a r y rocks in t u rn prov ide i n fo rma t ion on the m e c h a n i s m s which r egu la t e the supply and de l ive ry of sed iments to a si te and thus m a y be a useful p a r a m e t e r for
e v a l u a t i n g r eg iona l changes in c l imate and /o r sea level and the subs t r a t e redox condi t ion a t the t ime of deposi t ion.
The whole rock ino rgan ic geochemica l da t a f rom Braggs are p resen ted in Tab le II. Concen- t r a t i ons of se lec ted e lements were ca lcu la t ed on a c a r b o n a t e free basis for the purpose of e l imina t ing d i lu t ion effects b r o u g h t abou t by the va r i ab l e flux of b iogenic ca rbona te s . Car- b o n a t e free c o n c e n t r a t i o n s of se lected ele- men t s and ra t ios of e l ements a re p lo t ted versus dep th re la t ive to the l i thologic K/T b o u n d a r y in Fig.7.
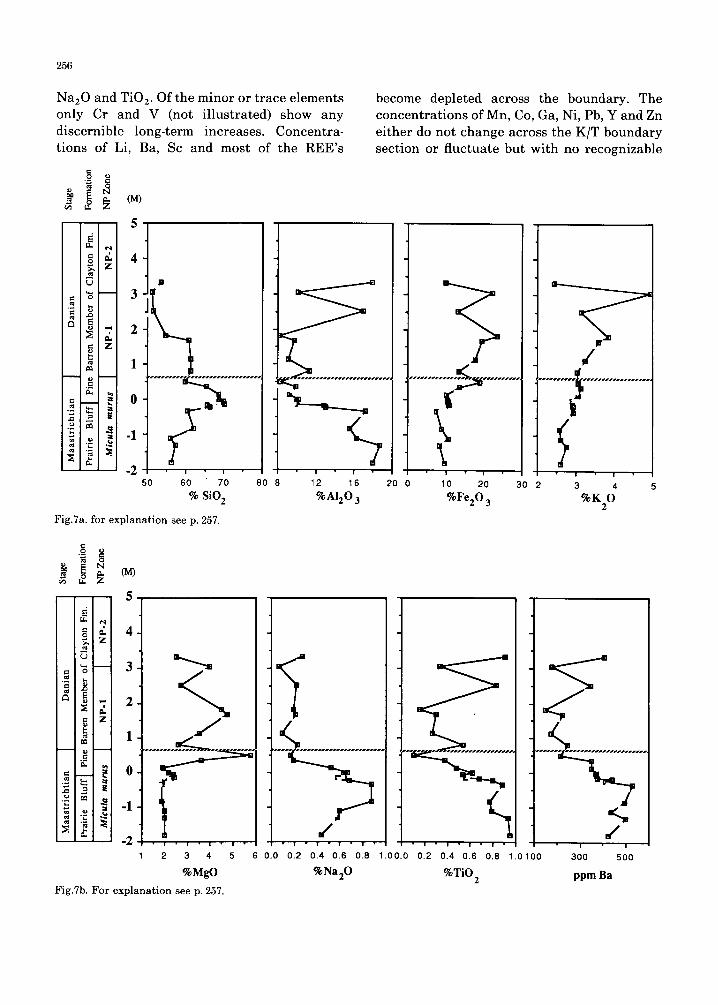
At Braggs , the c o n c e n t r a t i o n s of mos t m a j o r and m i n o r e l ements show cons ide rab le change across the K/T bounda ry . Ma jo r e l ements which inc rease by a f ac to r of a b o u t 2 f rom La te C r e t a c e o u s to Ea r ly P a l e o c e n e samples in- c lude FeEO 3, MgO and K 2 0 (Fig.7). The mos t a b u n d a n t c o m p o n e n t of the n o n - c a r b o n a t e f r ac t ion is SiO2 which inc reases f rom 55 to 70% f rom the base of bed 2 to the top of bed 3 but then g radua l ly decreases in c o n c e n t r a t i o n up sect ion. M a j o r e l emen t c o n c e n t r a t i o n s t h a t decrease above the b o u n d a r y inc lude AlEO 3,
256
Na:O and TiO:. Of the minor or trace elements only Cr and V (not illustrated) show any discernible long-term increases. Concentra- tions of Li, Ba, Sc and most of the REE's
n
~ 9
~ 3
~ ~ ~ 2
1
~ 0
"'~ ~ -1
(M)
5
5 0 6O 7 0
% SiO 2
Fig.7a. for explanation see p. 257.
8 0 8 12 16
%AI20 3
become depleted across the boundary. The concentrat ions of Mn, Co, Ga, Ni, Pb, Y and Zn either do not change across the K/T boundary section or fluctuate but with no recognizable
,~l~o~ur~rauu~o!r~__~ I~ao~or~o~
2O 0 10 2 0
%Fe20 3
w w
! i
3 0 2 3 4
%K O 2
"=
5
I
~ 4
~ 2
! ' Z ~ ~ 0
:~ =~ -2 . , . , . , . , .
2 3 4 5
%MgO
Fig.7b. For explanation see p. 257.
• I • I " 1 " ! • 0 .0 0 .2 0 .4 0 . 6 0 .8 1 . 0 0 . 0 0 .2 0 .4 0 . 6 0 .8 1 . 0 1 0 0
%Na20 %TiO 2
i • i • i • i • 3O0
i !
5OO
ppm Ba
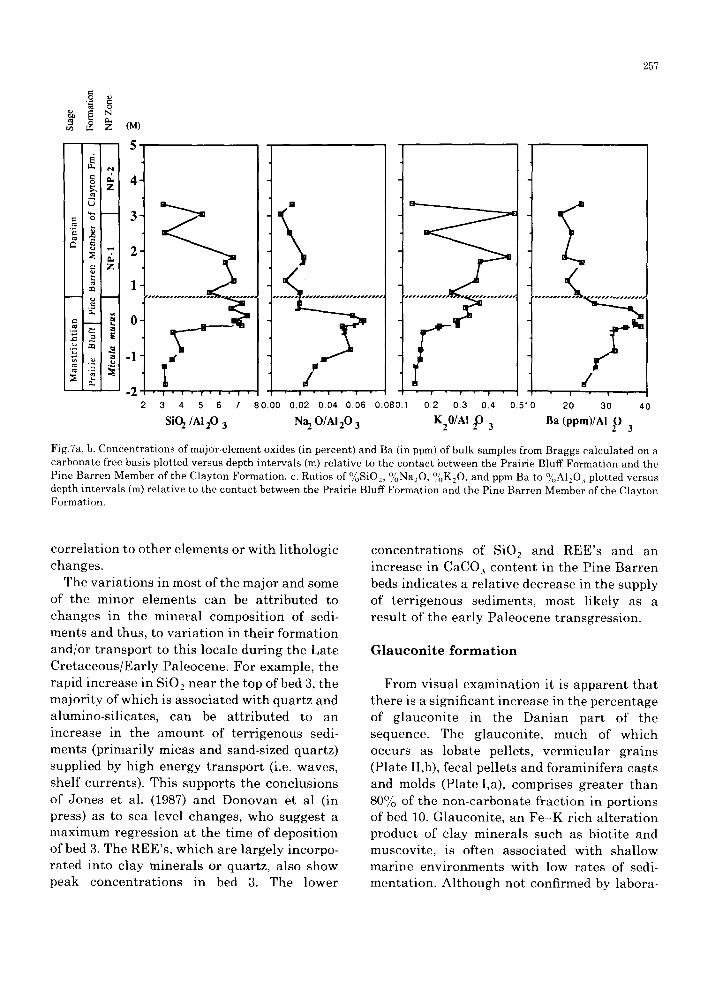
257
~ N
.~ z
o
g
.! " N E
N
(g)
1
,+ ...... 7+- 1 I
• I ' I • I ' I • I '
3 4 5 6 7 80.00 0.02 0.04 0.06
/ I I I
SiO 2/AI 2 ° 3 Na 2 O/A! 2 ° 3
0.080.1 I I I
0.2 0.3 0.4
K20/AI 2 ° 3
J ! I
0.510 20 30 40
Ba (ppm)/AI 20 3
Fig.7a, b. Concentrations of major-element oxides (in percent) and Ba (in ppm) of bulk samples from Braggs calculated on a carbonate free basis plotted versus depth intervals (m) relative to the contact between the Prairie Bluff Formation and the Pine Barren Member of the Clayton Formation. c. Ratios of °/oSiO2, °/oNa20, °/oK20 , and ppm Ba to °/oA1203 plotted versus depth intervals (m) relative to the contact between the Prairie Bluff Formation and the Pine Barren Member of the Clayton Formation.
correlat ion to other elements or with lithologic changes.
The variat ions in most of the major and some of the minor elements can be at tr ibuted to changes in the mineral composition of sedi- ments and thus, to variat ion in their formation and/or t ransport to this locale during the Late Cretaceous/Early Paleocene. For example, the rapid increase in SiO2 near the top of bed 3, the majority of which is associated with quartz and alumino-silicates, can be at t r ibuted to an increase in the amount of terr igenous sedi- ments (primarily micas and sand-sized quartz) supplied by high energy t ransport (i.e. waves, shelf currents). This supports the conclusions of Jones et al. (1987) and Donovan et al (in press) as to sea level changes, who suggest a maximum regression at the time of deposition of bed 3. The REE's, which are largely incorpo- rated into clay minerals or quartz, also show peak concentrat ions in bed 3. The lower
concentrat ions of SiO 2 and REE's and an increase in CaCO 3 content in the Pine Barren beds indicates a relative decrease in the supply of terr igenous sediments, most likely as a result of the early Paleocene transgression.
G l a u c o n i t e f o r m a t i o n
From visual examination it is apparent that there is a significant increase in the percentage of glauconite in the Danian part of the sequence. The glauconite, much of which occurs as lobate pellets, vermicular grains (Plate II,b), fecal pellets and foraminifera casts and molds (Plate I,a), comprises greater than 80% of the non-carbonate fraction in portions of bed 10. Glauconite, an Fe -K rich al terat ion product of clay minerals such as biotite and muscovite, is often associated with shallow marine environments with low rates of sedi- mentation. Although not confirmed by labora-
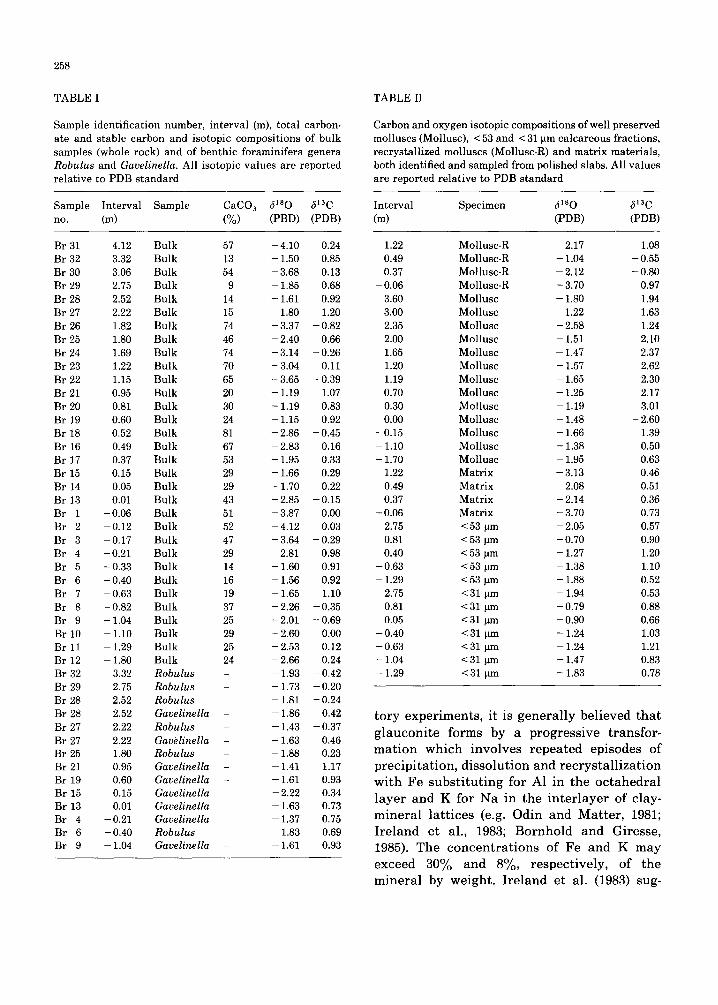
258
TABLE I
Sample i den t i f i ca t i on number , i n t e r v a l (m), t o t a l carbon-
a te and s t ab le ca rbon and i so top ic compos i t i ons of b u l k
samples (whole rock) and of be n th i c fo ramin i fe ra gene ra
Robulus and Gavelinella. All i so top ic v a l u e s a re r epor t ed
r e l a t i v e to PDB s t a n d a r d
Sample I n t e r v a l Sample CaCO 3 5 ' s O 51aC
no. (m) (%) (PBD) (PDB)
Br 31 4.12 B u l k 57 - 4 . 1 0 0.24
Br 32 3.32 Bu lk 13 - 1.50 0.85
Br 30 3.06 Bu lk 54 - 3.68 0.13
Br 29 2.75 B u l k 9 - 1.85 0.68
Br 28 2.52 B u l k 14 - 1.61 0.92
Br 27 2.22 B u l k 15 - 1.80 1.20
Br 26 1.82 B u l k 74 - 3 . 3 7 - 0 . 8 2
Br 25 1.80 Bu lk 46 - 2.40 0.66
Br 24 1.69 Bu lk 74 - 3 . 1 4 - 0 . 2 6
Br 23 1.22 B u l k 70 - 3.04 0.11
Br 22 1.15 Bu lk 65 - 3 . 6 5 - 0 . 3 9
Br 21 0.95 Bu lk 20 - 1.19 1.07
Br 20 0.81 B u l k 30 - 1.19 0.83
Br 19 0.60 B u l k 24 - 1.15 0.92
Br 18 0.52 B u l k 81 - 2 . 8 6 - 0 . 4 5
Br 16 0.49 B u l k 67 - 2.83 0.16
Br 17 0.37 B u l k 53 - 1.95 0.33 Br 15 0.15 Bu lk 29 - 1.66 0.29
Br 14 0.05 B u l k 29 - 1.70 0.22
Br 13 0.01 B u l k 43 - 2 . 8 5 - 0 . 1 5
Br 1 - 0 . 0 6 Bu lk 51 - 3 . 8 7 0.00
Br 2 - 0 . 1 2 Bu lk 52 - 4 . 1 2 0.03
Br 3 - 0 . 1 7 B u l k 47 - 3 . 6 4 - 0 . 2 9
Br 4 - 0 . 2 1 B u l k 29 - 2 . 8 1 - 0 . 9 8
Br 5 - 0 . 3 3 Bu lk 14 - 1 . 6 0 0.91
Br 6 - 0 . 4 0 B u l k 16 - 1 . 5 6 0.92
Br 7 - 0 . 6 3 B u l k 19 - 1 . 6 5 1.10
Br 8 - 0 . 8 2 Bu lk 37 - 2 . 2 6 - 0 . 3 5 Br 9 - 1.04 B u l k 25 - 2 . 0 1 - 0 . 6 9
Br 10 - 1.10 B u l k 29 - 2.60 0.00
Br 11 - 1.29 B u l k 25 - 2.53 0.12 Br 12 - 1.80 Bu lk 24 - 2.66 0.24
Br 32 3.32 Robulus - - 1 . 9 3 - 0 . 4 2
Br 29 2.75 Robulus - - 1.73 - 0 . 2 0 Br 28 2.52 Robulus - 1.81 - 0 . 2 4
Br 28 2.52 Gavelinella - - 1.86 0.42
Br 27 2.22 Robulus - 1 . 4 3 - 0 . 3 7
Br 27 2.22 Gavelinella - 1.63 0.46 Br 25 1.80 Robulus - 1.88 0.23 Br 21 0.95 Gavelinella - - 1.41 1.17 Br 19 0.60 Gavelinella - - 1.61 0.93
Br 15 0.15 GavelineUa - 2.22 0.34 Br 13 0.01 Gavelinella - 1.63 0.73 Br 4 -0 .21 Gavelinella - 1 . 3 7 0.75
Br 6 - 0 . 4 0 Robulus - 1.83 0.69 Br 9 - 1.04 Gavelinella - 1.61 0.93
TABLE II
Carbon and oxygen i so topic compos i t ions of wel l p rese rved
mol luscs (Mollusc) , < 53 and < 31 pm ca l ca r eous f rac t ions ,
r e c ry s t a l l i z ed mo l lu sc s (Mollusc-R) and m a t r i x ma te r i a l s ,
bo th ident i f ied and sampled from po l i shed slabs. All va lue s
a re r epo r t ed r e l a t i v e to PDB s t a n d a r d
I n t e r v a l Spec imen (f i s O 513 C
(m) (PDB) (PDB)
1.22 Mol lusc-R - 2.17 - 1.08
0.49 Mol lusc-R - 1.04 - 0.55
0.37 Mol lusc-R - 2.12 - 0.80
- 0.06 Mol lusc-R - 3.70 0.97
3.60 Mol lusc - 1.80 1.94
3.00 Mol lusc - 1.22 1.63
2.35 Mol lusc - 2.58 1.24
2.00 Mol lusc - 1.51 2.10
1.65 Mol lusc - 1.47 2.37
1.20 Mol lusc - 1.57 2.62
1.19 Mol lusc - 1.65 2.30
0.70 Mol lusc - 1.25 2.17
0.30 Mol lusc - 1.19 3.01
0.00 Mol lusc - 1.48 - 2.60
- 0.15 Mol lusc - 1.66 1.39 - 1.10 Mol lusc - 1.38 0.50
- 1.70 Mol lusc - 1.95 0.63
1.22 M a t r i x - 3.13 0.46
0.49 M a t r i x - 2.08 0.51
0.37 M a t r i x - 2.14 0.36
- 0.06 M a t r i x - 3.70 0.73
2.75 < 53 pm - 2.05 0.57
0.81 < 53 pm - 0.70 0.90
- 0.40 < 53 pm - 1.27 1.20 - 0 . 6 3 <53 ~m - 1.38 1.10
- 1.29 < 53 pm - 1.88 0.52 2.75 < 31 ~m - 1.94 0.53
0.81 < 31 pm - 0.79 0.88
0.05 < 31 pm - 0.90 0.66
- 0.40 < 31 9m - 1.24 1.03 - 0.63 < 31 pm - 1.24 1.21
- 1.04 < 31 pm - 1.47 0.83
- 1.29 < 31 pm - 1.83 0.78
t o r y e x p e r i m e n t s , i t i s g e n e r a l l y b e l i e v e d t h a t
g l a u c o n i t e f o r m s b y a p r o g r e s s i v e t r a n s f o r -
m a t i o n w h i c h i n v o l v e s r e p e a t e d e p i s o d e s o f
p r e c i p i t a t i o n , d i s s o l u t i o n a n d r e c r y s t a l l i z a t i o n
w i t h F e s u b s t i t u t i n g f o r A1 i n t h e o c t a h e d r a l
l a y e r a n d K f o r N a i n t h e i n t e r l a y e r o f c l a y -
m i n e r a l l a t t i c e s ( e . g . O d i n a n d M a t t e r , 1 9 8 1 ;
I r e l a n d e t a l . , 1983 ; B o r n h o l d a n d G i r e s s e ,
1985 ) . T h e c o n c e n t r a t i o n s o f F e a n d K m a y
e x c e e d 3 0 % a n d 8 % , r e s p e c t i v e l y , o f t h e
m i n e r a l b y w e i g h t . I r e l a n d e t a l . ( 1 9 8 3 ) s u g -
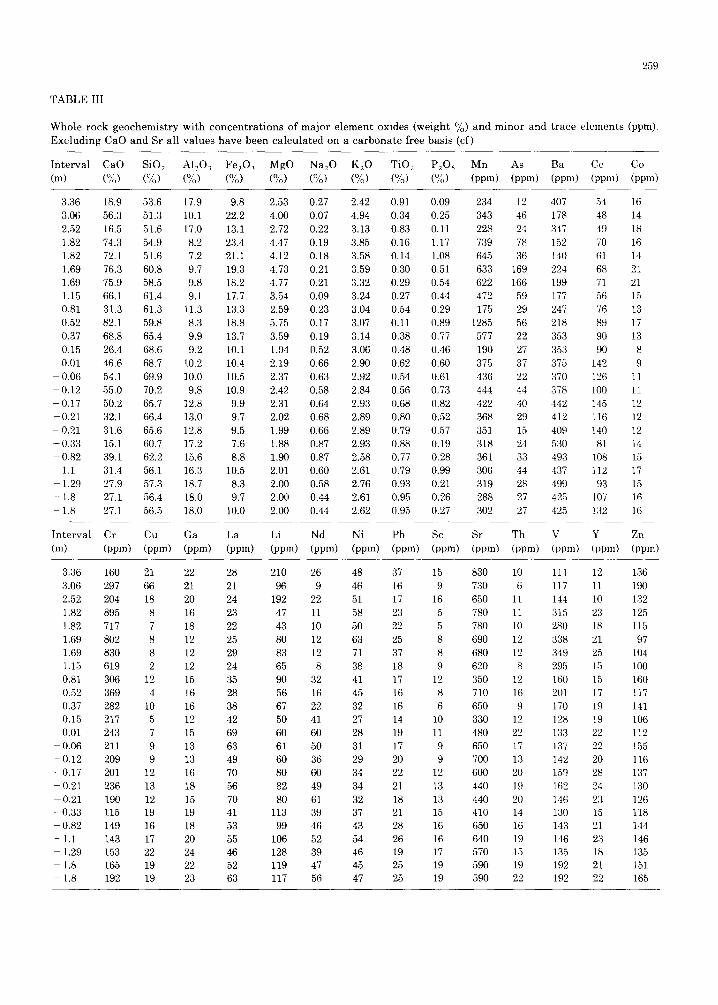
TABLE III
Whole rock geochemistry wi th concen t ra t ions of major element oxides (weight %) and minor and Excluding CaO and Sr all values have been calculated on a carbonate free basis (cf)
259
trace elements (ppm).
In terval CaO SiO 2 A1203 Fe203 MgO Na20 K20 TiO 2 P205 Mn As Ba Ce Co (m) (o/~,) (%,) (0/o) (%) (%) (%) (%) (%) (%) (ppm) (ppm) (ppm) (ppm) (ppm)
3.36 18.9 53.6 17.9 9.8 2.53 0.27 2.42 0.91 0.09 234 12 407 54 16 3.06 56.3 51.3 10.1 22.2 4.00 0.07 4.94 0.34 0.25 343 46 178 48 14 2.52 16.5 51.6 17.0 13.1 2.72 0.22 3.13 0.83 0.11 228 24 347 49 18 1.82 74.3 54.9 8.2 23.4 4.47 0.19 3.85 0.16 1.17 739 78 152 70 16 1.82 72.1 51.6 7.2 21.1 4.12 0.18 3.58 0.14 1.08 645 36 140 61 14 1.69 76.3 60.8 9.7 19.3 4.73 0.21 3.59 0.30 0.51 633 169 224 68 21 1.69 75.9 58.5 9.8 18.2 4.77 0.21 3.32 0.29 0.54 622 166 199 71 21 1.15 66.1 61.4 9.1 17.7 3.54 0.09 3.24 0.27 0.44 472 59 177 56 15 0.81 31.3 61.3 11.3 13.3 2.59 0.23 3.04 0.54 0.29 175 29 247 76 13 0.52 82.1 59.8 8.3 18.8 5.75 0.17 3.07 0.11 0.89 1285 56 218 89 17 0.37 68.8 65.4 9.9 13.7 3.59 0.19 3.14 0.38 0.77 577 22 353 90 13 0.15 26.4 68.6 9.2 10.1 1.94 0.52 3.06 0.48 0.46 190 27 353 90 8 0.01 46.6 68.7 10.2 10.4 2.19 0.66 2.90 0.62 0.60 375 37 375 142 9 0.06 54.1 69.9 10.0 10.5 2.37 0.63 2.92 0.54 0.61 436 22 370 126 11
- 0.12 55.0 70.2 9.8 10.9 2.42 0.58 2.84 0.56 0.73 444 44 378 100 11 - 0.17 50.2 65.7 12.8 9.9 2.31 0.64 2.93 0.68 0.82 422 40 442 145 12 - 0.21 32.1 66.4 13.0 9.7 2.02 0.68 2.89 0.80 0.52 368 29 412 116 12
- 0.21 31.6 65.6 12.8 9.5 1.99 0.66 2.89 0.79 0.57 351 15 409 140 12 - 0.33 15.1 60.7 17.2 7.6 1.88 0.87 2.93 0.88 0.19 318 24 530 81 14
0.82 39.1 62.2 15.6 8.8 1.90 0.87 2.58 0.77 0.28 361 33 493 108 15 - 1.1 31.4 56.1 16.3 10.5 2.01 0.60 2.61 0.79 0.99 306 44 437 112 17
1.29 27.9 57.3 18.7 8.3 2.00 0.58 2.76 0.93 0.21 319 28 499 93 15 - 1.8 27.1 56.4 18.0 9.7 2.00 0.44 2.61 0.95 0.26 288 27 425 107 16
1.8 27.1 56.5 18.0 10.0 2.00 0.44 2.62 0.95 0.27 302 27 425 132 16
Interval Cr Cu Ga La Li Nd Ni Pb Sc Sr Th V Y Zn (m) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm) (ppm)
3.36 160 21 22 28 210 26 48 37 15 830 10 i l l 12 136 3.06 297 66 21 21 96 9 46 16 9 730 6 117 11 190 2.52 204 18 20 24 192 22 51 17 16 650 11 144 10 132 1.82 895 8 16 23 47 11 58 23 5 780 11 315 23 125 1.82 717 7 18 22 43 10 50 22 5 780 10 280 18 115 1.69 802 8 12 25 80 12 63 25 8 690 12 338 21 97 1.69 830 8 12 29 83 12 71 37 8 680 12 349 25 104 1.15 619 2 12 24 65 8 38 18 9 620 8 295 15 100 0.81 306 12 15 35 90 32 41 17 12 350 12 160 15 160 0.52 369 4 16 28 56 16 45 16 8 710 16 201 17 117 0.37 282 10 16 38 67 22 32 16 6 650 9 170 19 141 0.15 217 5 12 42 50 41 27 14 10 330 12 128 19 106 0.01 243 7 15 69 60 60 28 19 11 480 22 133 22 112
- 0.06 211 9 13 63 61 50 31 17 9 650 17 137 22 155 - 0.12 209 9 13 49 60 36 29 20 9 700 13 142 20 116 - 0.17 201 12 16 70 80 60 34 22 12 600 20 159 28 137 - 0.21 236 13 18 56 82 49 34 21 13 440 19 162 24 130
0,21 190 12 15 70 80 61 32 18 13 440 20 146 23 126 -0 .33 115 19 19 41 113 39 37 21 15 410 14 130 15 118 - 0,82 149 16 18 53 99 46 43 28 16 650 16 143 21 144 - 1,1 143 17 20 55 106 52 54 26 16 640 19 146 23 146 - 1,29 153 22 24 46 128 39 46 19 17 570 15 135 18 135 - 1,8 165 19 22 52 119 47 45 25 19 590 19 192 21 151 - 1.8 192 19 23 63 117 56 47 25 19 590 22 192 22 165
260
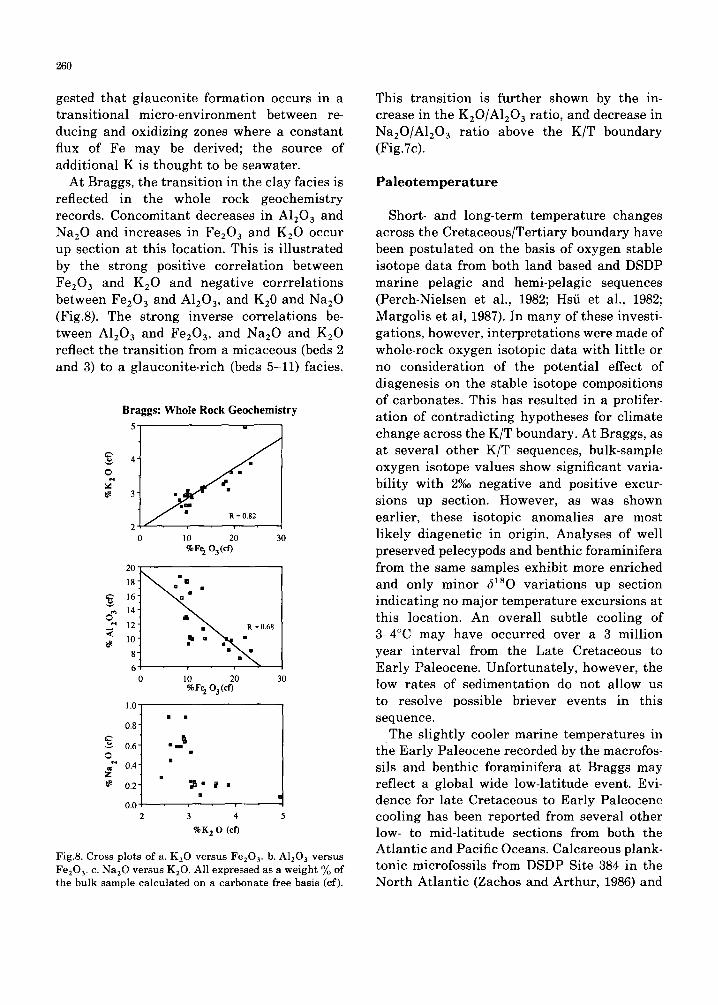
gested that glauconite formation occurs in a transitional micro-environment between re- ducing and oxidizing zones where a constant flux of Fe may be derived; the source of additional K is thought to be seawater.
At Braggs, the transition in the clay facies is reflected in the whole rock geochemistry records. Concomitant decreases in A1203 and Na20 and increases in FezO 3 and KEO occur up section at this location. This is i l lustrated by the strong positive correlation between Fe203 and K20 and negative corrrelations between Fe203 and A1203, and K20 and Na20 (Fig.8). The strong inverse correlations be- tween A1203 and Fe203, and Na20 and K20 reflect the transition from a micaceous (beds 2 and 3) to a glauconite-rich (beds 5-11) facies.
0
3"
2
Braggs: Whole Rock Geochemistry 5
ii Ii
•Il l
R = 0.82
10 20 30 %Fe~ 03 (cf)
2 0
18'
~ 1 6
"~ 12
O~ 1 0
8'
] .0-
0.8"
0.6-
c~ 0.4" Z
~ 0.2"
0.C 2
•i•R = 0.68
10 20 30 Fe 2 0 3(c0
. . t i
m
la
[]
3 4
% K 2 0 (cf)
Fig.8. Cross plots of a. K20 versus Fe203. b. A1203 versus Fe203. c. Na20 versus K20. All expressed as a weight % of the bulk sample calculated on a carbonate free basis (cf).
This transition is further shown by the in- crease in the K20/A120 a ratio, and decrease in Na20/A1203 ratio above the K/T boundary (Fig.7c).
P a l e o t e m p e r a t u r e
Short- and long-term temperature changes across the Cretaceous/Tertiary boundary have been postulated on the basis of oxygen stable isotope data from both land based and DSDP marine pelagic and hemi-pelagic sequences (Perch-Nielsen et al., 1982; Hsii et al., 1982; Margolis et al, 1987). In many of these investi- gations, however, interpretations were made of whole-rock oxygen isotopic data with little or no consideration of the potential effect of diagenesis on the stable isotope compositions of carbonates. This has resulted in a prolifer- ation of contradicting hypotheses for climate change across the K/T boundary. At Braggs, as at several other K/T sequences, bulk-sample oxygen isotope values show significant varia- bility with 2%0 negative and positive excur- sions up section. However, as was shown earlier, these isotopic anomalies are most likely diagenetic in origin. Analyses of well preserved pelecypods and benthic foraminifera from the same samples exhibit more enriched and only minor 6 '80 variations up section indicating no major temperature excursions at this location. An overall subtle cooling of 3-4°C may have occurred over a 3 million year interval from the Late Cretaceous to Early Paleocene. Unfortunately, however, the low rates of sedimentation do not allow us to resolve possible briever events in this sequence.
The slightly cooler marine temperatures in the Early Paleocene recorded by the macrofos- sils and benthic foraminifera at Braggs may reflect a global wide low-latitude event. Evi- dence for late Cretaceous to Early Paleocene cooling has been reported from several other low- to mid-latitude sections from both the Atlantic and Pacific Oceans. Calcareous plank- tonic microfossils from DSDP Site 384 in the North Atlantic (Zachos and Arthur, 1986) and

Site 527 in the South Atlantic (Shackleton et al., 1984) and several of the European sequences (e.g. Perch-Nielsen et al., 1982) also display isotopic enrichments during the Late Cretaceous to Early Paleocene transition. The duration and magnitude of the shifts vary considerably at each locality. For this reason and the fact that a number of other sequences, [DSDP Site 465 in the Pacific (Boersma and Shackleton, 1981) Sites 524 (Hsfi et al., 1982) and 356 in the South Atlantic, and Site 577 in the Pacific (Zachos and Arthur, 1986)], show climatic stasis, warming or general instability, it is difficult to support any one scenario of climate change for the K/T transition. This by no means precludes the possibility of a major climate event. The lack of reasonable correla- tion of sedimentary oxygen isotope records may result from a number of factors:
(1) As mentioned above, diagenesis may have removed or enhanced blso anomalies at some sections, especially in hemipelagic sequences that have been buried to depths in excess of 300 m. Unfortunately, the evidence for diagen- esis is not always obvious and anomalous isotopic signals resulting from diagenesis are often interpreted in terms of primary temper- ature variation.
(2) The types of calcareous components ana- lyzed for ~urface water temperature estimates, including ibulk samples, fine fraction carbon- ate, both of which are assumed to consist mainly of coccoliths, and various species of planktonic and shallow shelf benthic foramini- fera, are quite variable from sequence to sequence. As a result, differences in isotopic compositions may be related to vital effects and/or depth habitat variability (Goodney et al., 1980; Duplessy et al., 1981; Fairbanks et al., 1982; Dudley et al., 1986; Paull and Thierstein, 1987).
(3) The stratigraphic time resolution repre- sented by each isotope record is variable due to differences in the sedimentation rate, bioturba- tion, and/or the sampling density.
A further possibility is that the data are correct and that the temperature response was variable from region to region. From recon-
261
structions we know that Late Cretaceous continental configurations, circulation pat- terns, sea level, lati tudinal temperature gradi- ents and albedo were much different from today (e.g. Brass et al., 1977; Matsumoto, 1980; Barron, 1983, 1987; Barron et al., 1984). Re- sponse to climatic forcing on the modern earth is quite variable with separate regions experi- encing different effects (e.g. E1 Nifio events). Such may have been the case during the K/T boundary event.
Therefore, on the basis of the available oxygen isotope evidence [here], we might at best conclude that a long term gradual cooling of 3-4cC took place at Braggs and that major and more rapid shifts in temperature did not occur during the K/T transition. The low rates of sedimentation and the presence of a hiatus below the boundary prevent the determination of short term (< 100 kyr) climatic events.
Paleoecology
Following the K/T boundary event, it is clear from the fossil record that major changes occurred in the trophic structure of the benthic ecosystem at the Braggs locality. The abun- dance, diversity, and average size of macrofos- sils all decreased during this period (Bryan and Jones, this issue). The Cretaceous molluscan assemblages were dominated by suspension- filter feeders whereas the Early Paleocene assemblages consisted mainly of deposit feed- ers (Arthur et al., 1987). Similar evolutionary patterns have been observed at other Gulf Coast K/T boundary sequences (Hansen et al., 1987). This shift in the character of benthic macrofossil assemblages suggests that the food supply for suspension feeders may have been reduced allowing only those species which normally feed on the sediment reservoirs of Corg, the deposit feeders, to survive (Sheehan and Hansen, 1986; Arthur et al., 1987). If there was substantial reduction in the flux of Cot ~ to the substrate one may expect to see evidence for this in the geochemical record. Previous geochemical investigations of pelagic se- quences have revealed a negative shift of about

262
1.5%o in the ~13C compositions of calcareous plankton and slight enrichment in the 5~3C of benthic foraminifera at the time of the major K/T boundary extinctions (e.g. Zachos and Arthur, 1986). This has been interpreted to represent a complete breakdown of the surface- to deep-water organic carbon transfer charac- teristic of normally productive oceans. At Braggs, however, no such negative shift in carbon isotopes is recorded by any of the components analyzed. In fact, the only pertur- bation within the carbon isotope record other than the shifts related to diagenesis in the bulk samples is the positive enrichment of the molluscs.
On the basis of investigations of modern molluscs it has been demonstrated that the carbon isotopic composition of the shells may be controlled by a number of parameters, including ambient water TDC ~13C, food 513C, sexual maturity, and to a lesser extent water temperature (Killingley and Berger, 1979; Jones et al., 1983; Tanaka et al., 1986; Krantz et al., 1987; Romanek et al., 1987). The relative importance of each parameter may vary considerably from species to species.
Epifaunal molluscs may precipitate their shells in equilibrium with bottom water TDC, whereas infaunal species precipitate carbonate in equilibrium with pore water TDC (e.g. Stevens and Vella, 1981; Krantz et al., 1987), the composition of which is relatively depleted with respect to ~3C due to the in-situ decay of isotopically light organic carbon (McCorkle et al., 1985). As a result, one may expect slightly lighter isotopic compositions for the infaunal species. If a decrease in the flux of organic matter to the sediment were to occur, the difference in carbon composition between the epi- and infaunal species should diminish.
Indirect evidence for a decrease in the flux of organic matter across the K/T boundary at Braggs comes from both barium and organic carbon data. In the modern marine environ- ment the distribution of Ba resembles that of the major nutrients, especially silica (Turek- ian, 1968; Chan et al., 1977). Ba has been linked with suspended organic matter distribution
and flux to the sediments (Dehairs et al., 1980, 1987; Dymond, 1986). Sedimentary organic matter is readily oxidized and only a small fraction (< 1%) of the organic matter produced is buried in sediments. Because sedimentary pore waters are near saturat ion with respect to barite, it is more readily preserved than organic matter (Church, 1979). As a result, the Ba/A1 ratio has been used as a proxy indicator of organic matter flux and paleoproductivity (Schmitz, 1987). At Braggs, the Ba:A1 and Ba:Ca ratios show a substantial decrease just above the boundary. The decrease in accumu- lation of Ba in the lower Paleocene probably reflects a reduction in the accumulation rate of organic matter. The reduction in the preserva- tion of organic matter can be brought about by many factors that affect both production and destruction of that organic matter. The most obvious means of reduction would be by a decrease in marine primary productivity in this region related to the decrease in open ocean productivity at this time (Zachos and Arthur, 1986). The Ba data support this inter- pretation.
Alternatively, a decrease in the accumu- lation of organic matter could be produced by simply changing sedimentation rates. At lower sedimentation rates organic matter is buried less quickly and thus is more likely to undergo oxidation at or near the seafloor. A decrease in terrigenous sediment flux occurs at this time as a result of the Early Paleocene transgression along the Gulf Coast (Donovan et al., in press) and thus may have led to a reduction in the rate of Corg burial at this location. The more enriched molluscan ~13C values correlate well with the interval of deep shelf carbonate-rich beds (5-9) deposited during the transgression and, therefore, may reflect the relative reduc- tion in sedimentation rates and, hence, organic carbon burial. The result would be lower pore water 513C gradients. Also, during transgres- sion sources of terrestrial organic matter are more distal further reducing the input of Corg to a given site.
Other factors which may be considered in explaining the carbon shift of the macrofossils,

is that of physiological growth changes related to sexual maturity. Other calcareous secreting organisms which have been studied in detail, such as planktonic foraminifera Neogloboqua- drina dutertrei and Globorotalia menardii, dis- play an intraspecific, size- dependent fractiona- tion effect, possibly reflecting a change in the rate of metabolism through the lifetime of an organism (Bouvier-Soumagnac and Duplessy, 1985). Similarly, the ratio of metabolic carbon to sea water TDC incorporated into a mollusc shell may change over the lifetime of an organism (e.g. Jones et al., 1983; Krantz et al., 1987; Romanek et al., 1987). Analyses of shells of modern molluscs have demonstrated that there is a gradual depletion in 513C with age of up to 2.0%0 along the axis of growth. During early rapid growth the composition of the shell may be near equilibrium with the TDC. In later stages of growth, the organism begins to secrete calcite that contains a higher percentage of metabolic carbon. As a result, the ~13C of each subsequent growth ring becomes progressively lighter. As the molluscan species assemblages change, the average specimen size decreases across the K/T boundary at Braggs (Bryan and Jones, 1988) thus leaving size-related disequilibrium effects as a possible cause of the 51~C enrichment measured in the molluscs.
Regardless of the exact origin of the 13C anomaly measured in the molluscs, it does coincide with the near complete taxonomic turnover in the benthic and planktonic fauna and flora in this sequence, suggesting a signifi- cant change in the local ecology across the K/T boundary interval. Furthermore, over the interval examined the stable isotopic and paleontological records (Jones et al., 1987; Bryan and Jones, this issue) exhibit little change prior to the main disappearance of taxa at the depositional hiatus near the base of bed 4. This indicates that environmental stress and step wise extinctions did not occur at Braggs during the Late Cretaceous prior to 66.9 Ma [0.5x106 years is the maximum possible amount of time thought to be represented by the erosional unconformity].
263
Summary and conclusions
From the geochemical and petrographic evidence presented here we are able to demon- strate that a substantial amount of sediment diagenesis has occurred at the K/T boundary sequence exposed at Braggs, Alabama. Despite the alteration of bulk material, however, enough well preserved molluscs and benthic foraminifera were recovered to demonstrate that:
(1) Shallow shelf marine temperatures along the Gulf coast averaged around 19 ° during the late Cretaceous/early Paleocene transition.
(2) At most there was a gradual 2-3°C cooling in marine temperature across the K/T boundary at Braggs. No major temperature change is associated with the marine plankton extinctions, at least on time scales of greater than 100,000 years.
(3) Significant changes in the local ecology did occur as reflected in the shift in the carbon isotopic compositions of molluscs across the K/T boundary. However, the exact nature of the change is uncertain and may have been brought about by one or a combination of several factors, including the early Danian sea level rise, decreased sedimentation rates, or a decline in primary productivity.
From the whole rock geochemical and petro- graphic data we are able to show that;
(4) The influx of somewhat coarser-grained sediments, primarily sand sized quartz indica- tive of higher energy, decreased during the early Paleocene at Braggs. As a result, more locally derived biogenic and authigenic min- erals, such as calcite and glauconite, became the major sedimentary components. The tran- sition in mineral assemblages is the result of a sealevel rise during this period.
Acknowledgements
The authors would like to thank P. Abell, D. Fastovsky and D. Jones for providing helpful comments on this manuscript. Financial sup- port for this investigation came from NSF grant EAR 83-06561.

264
References
Arthur, M. A., Zachos, J. C. and Jones, D. S., 1987. Primary productivity and the Cretaceous/Tertiary boundary event in the oceans. Cretaceous Res., 8: 43-54.
Barron, E.J . , 1983. A warm equable Cretaceous: The nature of the problem. Earth Sci. Rev., 19: 305-338.
Barron, E. J., 1987. Cretaceous plate tectonic reconstruc- tions. Palaeogeogr., Palaeoclimatol., Palaeoecol., 59: 3-29.
Barron, E.J. , Thompson, S.L. and Hay, W.W., 1984. Continental distribution as a forcing factor for global- scale temperature. Nature, 310: 574-575.
Baum, G. R., Bleschschmidt, G. L., Hardenbol, J., Loutit, T.S., Vail, P.R. and Wright, R.C., 1984. The Maas- trichtian/Danian boundary in Alabama: A strati- graphically condensed section. Geol. Soc. Am. Abstr. with Programs, 16: 440.
Berggren, W. A., Kent, D. V., Flynn, J. J. and Van Couver- ing, J. A., 1985. Cenozoic geochronology. Geol. Soc. Am. Bull., 96:1407 1418.
Bischoff, J. L., 1968. Kinetics of calcite nucleation: Mag- nesium ion inhibition and ionic strength catalysts. J. Geophys. Res., 73: 3315-3322.
Bischoff, J. L. and Fyfe, W. S., 1968. Catalysis, inhibition, and the calci te-aragonite problem. I. The aragonite- calcite transformation. Am. J. Sci., 266:65 79.
Boersma, A., 1984. Campanian through Paleocene paleo- temperature and carbon isotope sequence and the Cretaceous/Tertiary boundary in the Atlantic ocean. In: W.A. Berggren and J .A. Van Couvering (Editors), Catastrophism and Earth History, The New Uniformi- tarianism. Princeton Univ. Press, Princeton, N.J., pp. 247 278.
Boersma, A. and Shackleton, N.J. , 1981. Oxygen and carbon isotope variations and planktonic-foraminifer depth habitats, Late Cretaceous to Paleocene, Central Pacific. In: J. Thiede, T. L. Vallier, et al., Initial Reports of the Deep Sea Drilling Project, 43. U.S. Government Printing Office, Washington, D.C., pp. 513 526.
Bornhold, B.D. and Giresse, P., 1985. Glauconitic sedi- ments on the continental shelf off Vancouver Island, British Columbia, Canada. J. Sediment. Petrol., 55: 653-664.
Bouvier-Soumagnac, Y. and Duplessy, J.-C., 1985. Carbon and oxygen isotopic composition of planktonic foramini- fera from laboratory culture, plankton tows and recent sediment: Implications for the reconstruction of paleo- climatic conditions and of the global carbon cycle. J. Foraminiferal Res., 15: 302-320.
Brass, G.W., Southam, J .R. and Peterson, W.H., 1977. Warm saline bottom water in the ancient ocean. Nature, 296:620 623.
Cepek, P., Hay, W.W., Masters, B. A. and Worsley, T. R., 1968. Calcareous plankton in samples from field trip stops. In: J. C. Scott (Chairman), Facies Changes in the Selma Group in Central and Eastern Alabama, Alabama Geol. Soc. Guidebook, 6th fieldtrip, Ala. Geol. Surv., Tuscaloosa, pp. 33-40.
Chan, L. H., Drummond, D., Edmond, J. M. and Grant, B.,
1977. On the barium data for the Atlantic GEOSECS expedition. Deep-Sea Res., 24: 613-649.
Church, T.M., 1979. Marine barite. In: R.G. Burns (Editor), Marine Minerals. Mineral. Soc. Am., Short Course Notes, 6: 175-207.
Copeland, C.W. and Mancini, E.A., 1986. Cretaceous Tertiary boundary southeast of Braggs, Lowndes County, Alabama. In: T.L. Neathery (Editor), South- eastern Section of the Geological Society of America, Centennial Field Guide, 6, pp. 369-372.
Corliss, B. H., 1985. Microhabitats of benthic foraminifera within deep-sea sediments. Nature, 314:435 438.
Craig, H., 1957. Isotopic standards for carbon and oxygen and correction factors for mass-spectro-metric analysis of carbon dioxide. Geochim. Cosmochim. Acta, 12: 133-149.
Dehairs, F., Chesselet, R. and Jedwab, J., 1980. Discrete suspended particles of barite and the barium cycle in the open ocean. Earth Planet. Sci. Lett., 49: 528-550.
Dehairs, F., Lambert, C. E., Chesselet, R. and Risler, N., 1987. The biological cycle in the Western Mediterranean Sea. Biogeochemistry, 4: 119-139.
Donovan, A. D. and Vail, P. R., 1986. Sequence stratigraphy of the K-T boundary in Alabama: A noncatastrophic alternative. Geol. Soc. Am. Abstr. with Programs, 18: 587.
Donovan, A. D., Baum, G. R., Bleschschmidt, G. L., Loutit, T.S., Pflum, C.E. and Vail, P.R., in press. Sequence stratigraphic setting of the Cretaceous-Tertiary bound- ary in Central Alabama. Soc. Econ. Paleontol. Mineral. Spec. Publ.
Dudley, W. C., Blackwelder, P., Brand, L. and Duplessy, J.-C., 1986. Stable isotopic composition of coccoliths. Mar. Micropaleontol., 10:1 8.
Duplessy, J.-C., Blanc, P.-L. and B~, A.W.H. , 1981. Oxygen-18 enrichment of planktonic foraminifera due to gammetogenic calcification below the euphotic zone. Science, 213:1247 1250.
Dymond, J., 1986. Particulate barium fluxes in the oceans: An indicator of new productivity. In: Abstr. Ocean Sciences Meet. Am. Geophys. Union, New Orleans, La., 1986, p. 972.
Fairbanks, R. G., Sverdlove, M., Free, R., Wiebe, R. H. and B~, A. W. H., 1982. Vertical distribution and isotopic fractionation of living planktonic foraminifera from the Panama Basin. Nature, 298: 841-844.
Floyd, M.A., Fassel, V.A. and D'Silva, A.P., 1980. Computer-controlled scanning monochrometer for the determination of 50 elements in geochemical and en- vironmental samples by inductively coupled plasma- atomic emission spectrometry. Anal. Chem., 52: 2168 2173.
Folk, R. L., 1974. The natural history of crystalline calcium carbonate: effect of magnesium content and salinity. J. Sediment. Petrol., 44:40 53.
Goodney, D. E., Margolis, S. V., Dudley, W. C., Kroopnick, P. and Williams, D. F., 1980. Oxygen and carbon isotopes of recent calcareous nannofossils as paleoceanographic indicators. Mar. Micropaleontol., 5: 31-42.
Graham, D. W., Corliss, B. H., Bender, M. L. and Keigwin, L. D., 1981. Carbon and oxygen isotope disequilibrium of

recent deep sea benthic foraminifera. Mar. Micropaleon- tol., 6:438 497.
Grossman, E. L., 1984. Stable isotope fract ionat ion in live benthic foraminifera from the Southern California Borderland. Palaeogeogr., Palaeoclimatol. , Palaeoecol., 47:301 327.
Grossman, E. L., 1987. Stable isotopes in modern benthic foraminifera: A study of vital effect. J. Foraminiferal Res., 17:48 61.
Hallam, A., 1987. End-Cretaceous mass ext inct ion event: Argument for terrestr ial causation. Science, 238: 1237 1242.
Hansen, T.A., Farrand, R.B., Montgomery, H.A., Bill- man, H. G. and Bleschschmidt, G., 1987. Sedimentology and ext inct ion pat terns across the Cretaceous Tert iary boundary interval in East Texas. Cretaceous Res., 8: 229 252.
Hess, J., Bender, M. L. and Schilling, J.-G., 1986. Evolut ion of the ratio of strontium-87 to strontium-86 in seawater from Cretaceous to present. Science, 231:979 984.
Hs/l, K.J. , He, Q., McKenzie, J .A. , Weissert, C., Perch- Nielsen, K., Oberhansli , H., Kelts, K., LaBr6cque, J., Tauxe, L., Krahenb6hl , U., Percival, S. F., Wright, R., Darposoff, A. M., Peterson, N., Tucker, P., Poore, R. Z., Gombos, A. M., Posciotto, K., Carman, M. F. and Schrei- ber, E., 1982. Mass mortal i ty and its environmental and evolut ionary consequences. Science, 216:249 256.
Hut, P., Alvarez, W., Elder, W. P., Hansen, T., Kauffman, E. G., Keller, G., Shoemaker, E. M. and Weissman, P. R., 1987. Comet showers as a cause of mass extinctions. Nature, 329:118 126.
Ireland, B.J., Curtis, C.C. and Whiteman, J .A. , 1983. Compositional var ia t ion within some glauconites and illites and implication for thei r stabil i ty and origin. Sedimentology, 30:769 786.
Jarzen, D.M., 1978. The terrestr ial palynoflora from the Cretaceous Tert iary transi t ion, Alabama, U.S.A. Pollen Spores, 20:535 553.
Jones, D.S., Williams, D.F. and Arthur, M.A., 1983. Growth history and ecology of the Atlant ic surf claim, Spisula solidissima (Dillwin), as revealed by stable isotopes and annual shell increments. J. Exp. Mar. Biol. Ecol., 73:225 242.
Jones, D.S., Mueller, P.A., Bryan, J .R. , Dobson, J .P . , Channel, J. E. T., Zachos, J. C. and Arthur, M. A., 1987. Biotic, geochemical and paleomagnetic changes across the Cretaceous/Tert iary boundary at Braggs, Alabama. Geology, 15:311 315.
Kauffman, E.G., 1984. The fabric of Cretaceous marine extinctions. In: W. A. Berggren and J. A. Van Couvering (Editors), Catastrophism and Ear th History, The New Uniformitarianism. Pr inceton Univ. Press, Princeton, N.J., pp. 151 246.
Kent, D. V. and Gradstein, F. M., 1985. A Cretaceous and ,Jurassic geochronology. Geol. Soc. Am. Bull., 96: 1419 1427.
Killingley, J. S. and Berger, W. H., 1979. Stable isotopes in mollusk shell: Detection of upwelling events. Science, 205:186 188.
Krantz, D.E., Williams, D.F. and Jones, D.F., 1987.
265
Ecological and paleoenvironmental information using stable isotope profiles from living and fossil molluscs. Palaeogeogr., Palaeoclimatol. , Palaeoecol., 58:249 266.
Margolis, S. V., Mount, J. F., Doehne, E., Showers, W. and Ward, P., 1987. The Cretaceous/Tert iary boundary carbon and oxygen isotope strat igraphy, diagenesis, and paleoceanography at Zumaya, Spain. Paleoceanography, 2:361 378.
Matsumoto, T., 1980. Inter-regional correlat ion of trans- gressions and regressions in the Cretaceous Period. Cretaceous Res., 1:359 373.
McCorkle, D.C., Emerson, S.R. and Quay, P.D., 1985. Stable carbon isotopes in marine porewaters. Ear th Planet. Sci. Lett., 74:13 26.
Mount, J. F., Margolis, S. V., Showers, S. V., Ward, P. and Doehne, E., 1986. Carbon and oxygen isotope stratigra- phy of the Upper Maastr icht ian, Zumaya Spain: A record of oceanographic and biological changes at the end of the Cretaceous Period. Palaios, 1:87 92.
Mueller, P., Jones, D, S., Arthur, M.A. and Zachos, J., 1986. Sr, C, and O isotopic var ia t ions across the K/T boundary at Braggs, Alabama. In: Geol. Soc. Am. Meet., Abstr. with Programs, 18: 700.
M/iller, G. and Gastner, M., 1971. The "Karbonate Bombe": A simple device for the determinat ion of the carbonate content in sediments, soils and other materials. Neues Jahrb. Miner. Monatsh., 10:466 469.
Odin, G. S. and Matter, A., 1981. De glauconaiarum origine. (The origin of glauconite). Sedimentology, 28:611 641.
Officer, C. B. and Drake, C.L., 1983. The Cretaceous/Ter- t iary transit ion. Science, 219:1383 1390.
Officer, C. B., Hallam, A., Drake, C. L. and Devine, J. B., 1985. Late Cretaceous and paroxysmal Cretaceous/Ter- t iary extinctions. Nature, 326:143 149.
Paull, C.K. and Thierstein, H.R., 1987. Stable isotopic fract ionat ion among particles in Quar ternary coccolith- sized deep-sea sediments. Paleoceanography, 2:423 430.
Perch-Nielsen, K., McKenzie, J. and He, Q., 1982. Biostrati- graphic and isotope s t ra t igraphy and the "catas t rophic" extinction of calcareous nannoplank ton at the Creta- ceous/Tertiary boundary. In: L.T. Silver and P.H. Schultz (Editors), Geological Implications of Large Asteroids and Comets on the Earth. Geol. Soc. Am. Spec. Publ., 190:353 371.
Romanek, C. S., Jones, D. S., William, D. F., Krantz, D. E. and Radtke, R., 1987. Stable isotopic invest igation of physiological and environmental changes recorded in shell carbonate from the giant clam Tridacna maxima. Mar. Biol., 91:385 393.
Schlanger, S.O. and Douglas, R.G., 1974. The pelagic ooze chalk limestone t ransi t ion and its implication for marine strat igraphy. Spec. Publ. Int. Assoc. Sedimentol., 1:177 210.
Schmitz, B., 1987. Barium, equatorial high productivity, and the nor thward wandering of the Indian Continent. Paleoceanography, 2: 63-- 78.
Scott, J .C. (Editor), 1968. Facies changes in the Selma Group in central and eastern Alabama. In: Alabama Geol. Soc. Guidebook, 6th fieldtrip. Ala. Geol. Surv., Tuscaloosa, pp. 33 40.

266
Shackleton, N. J. and Kennett, J. P., 1975. Paleotempera- ture history of the Cenozoic and the initiation of Antarctic glaciation: Oxygen and carbon isotope analy- sis of DSDP Sites 277, 279 and 281. In: Initial Reports of the Deep Sea Drilling Project, 29. U.S. Government Printing Office, Washington, D.C., pp. 743 755.
Shackleton, N.J., Hall, M.A. and Boersma, A., 1984. Oxygen and carbon isotope data from Leg 74 Foramini- fers. In: Initial Reports of the Deep Sea Drilling Project, 74. U.S. Government Printing Office, Washington, D.C., pp. 599 612.
Sheehan, P. M. and Hansen, T. A., 1986. Detrital feeding as a buffer to exinction at the end of the Cretaceous. Geology, 14:868 870.
Smit, J., 1982. Extinction and evolution of planktonic foraminifera at the Cretaceous/Tertiary boundary after a major impact. In: L.T. Silver and P.H. Schultz (Editors), Geological Implications of Large Asteroids and Comets on the Earth. Geol. Soc. Am. Spec. Publ., 190: 329-352.
Smith, G.K., 1978. Ostracoda of the Prairie Bluff Chalk, Upper Cretaceous (Maestrichtian) and Pine Barren member of the Clayton formation, Lower Paleocene (Danian) from exposures along Alabama State Highway 263 in Lowndes County, Alabama. Gulf Coast Assoc. Geol. Soc. Trans., 28:539 580.
Stevens, K. F. and Vella, P., 1981. Palaeoclimatic interpre- tation of stable isotope ratios in molluscan fossils from Middle Pleistocene strata, Eanganui, New Zealand. Palaeogeogr., Palaeoclimatol., Palaeoecol., 34: 257-266.
Taggert, J.E., Wahlberg, J.S. and Taylor, H.E., 1980. X-ray spectrometric major-element analyses of tephra
samples from the May 18, 1980 eruption of Mount St. Helens samples collected from Washington, Idaho, and Montana. Geol. Surv. Open-file Rep., pp. 80 1130.
Tanaka, N., Monaghan, M.C. and Rye, D.M., 1986. Contribution of metabolic carbon to mollusc and bar- nacle shell carbonate. Nature, 320: 520-523.
Thierstein, H.R., 1981. Late Cretaceous nannoplankton and the change at the Cretaceous-Tertiary boundary. Soc. Econ. Paleontol. Mineral. Spec. Publ., 32: 355-394.
Turekian, K., 1968. Deep-sea deposition of barium, cobalt and silver. Geochim. Cosmochim. Acta, 32: 603-612.
Vail, P.R., Baum, G.R., Loutit, T.S., Donovan, A.D., Mancini, E.A. and Tew, B.H., Jr., 1987. Sequence Stratigraphy of the Upper-most Cretaceous and Paleo- gene of the Alabama/Western Georgia Coastal Plain. In: Fieldtrip Guidebook for the Lafayette Geological Society. Lafayette Geol. Soc., Lafayette, La., 214 pp.
Ward, P., Wiedman, J. and Mount, J.F., 1986. Maas- trichtian molluscan biostratigraphy and extinction pat- tern in a Cretaceous/Tertiary boundary section exposed at Zumaya, Spain. Geology, 14:899 903.
Woodruff, F., Savin, S. and Douglas, R., 1980. Bio- logical fractionation of oxygen and carbon isotopes by recent benthic foraminifera. Mar. Micropaleontol., 5: 3-11.
Worsley, T.R., 1974. The Cretaceous-Tertiary boundary event in the ocean. Soc. Econ. Paleontol. and Mineral. Spec. Publ., 20: 94-125.
Zachos, J. C. and Arthur, M. A., 1986. Paleoceanography of the Cretaceous/Tertiary boundary event: Inferences from stable isotopic and other data. Paleoceanography, 1: 5-26.
Related Documents











