Genomics and Gene Recognition CIS 667 April 27, 2004

Genomics and Gene Recognition CIS 667 April 27, 2004.
Dec 14, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Genomics and Gene Recognition
CIS 667 April 27, 2004

Genomics and Gene Recognition
• How do we recognize the genes given the raw sequence data?
• Two different cases: Prokaryotes: relatively easy Eukaryotes: relatively difficult
Much “junk DNA” to search through
• Signals determine the beginnings and ends of genes Need to find the signals

Prokaryotic Genomes
• Genomic information of prokaryotes dedicated mainly to basic tasks Make and replicate DNA Make new proteins Obtain and store energy
• Over 60 prokaryotic genomes have been completely sequenced since mid-1990s
Prokaryotic Genomes
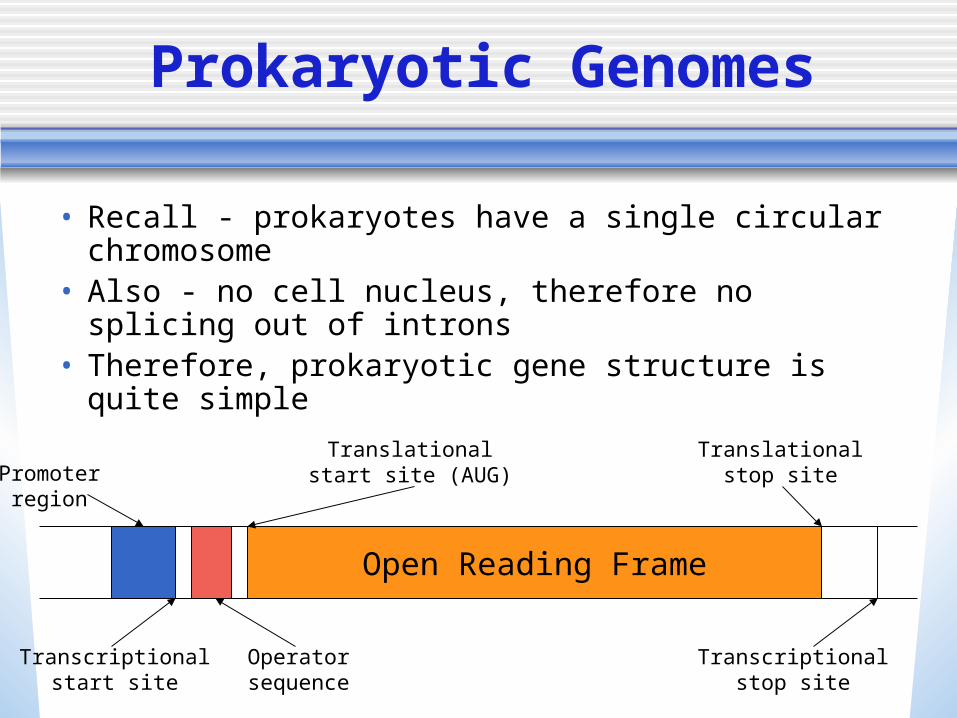
• Recall - prokaryotes have a single circular chromosome
• Also - no cell nucleus, therefore no splicing out of introns
• Therefore, prokaryotic gene structure is quite simple
Transcriptionalstart site
Promoterregion
Operatorsequence
Open Reading Frame
Transcriptionalstop site
Translationalstart site (AUG)
Translationalstop site

Promoter Elements
• Gene expression begins with transcription RNA copy of a gene made by an RNA
polymerase Prokaryotic RNA polymerases are
assemblies of several different proteins ’ protein binds to DNA template protein links nucleotides protein holds subunits together protein recognizes specific nucleotide
sequences of promoters

Promoter Elements
• ’, and often very similar from one bacterial species to another
• can vary (less well conserved) Several variants often found in a cell The ability to use several different
factors allows a cell to turn on or off expression of whole sets of genes For example, 32 turns on gene expressions
for genes associated with heat shock while does the same for nitrogen stress and genes that always need to be expressed are transcribed by polymerases with

Promoter Elements
• Each factor recognizes a particular sequence of nucleotides upstream from the gene looks for -35 sequence TTGACA and -
10 sequence TATAAT Other factors look for other -35 and -
10 sequences The match need not always be exact The better the match, the more likely
transcription will be initiated

Promoter Elements
• Protein products from some genes are always used in tandem with those from some other genes These related genes may share a single
promoter in prokaryotic genomes and be arranged in an operon
When one gene is transcribed, so are all of the others - one polycistronic RNA molecule is produced
The lactose operon contains three genes involved in metabolism of the sugar lactose in bacterial cells

Operon
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

Operon
• The protein encoded by the regulatory gene (pLacI) can bind to lactose or to the operator sequence of the operon So when lactose is abundant, less likely
to bind to operator sequence When it does, it blocks transcription, thus
acting as a negative regulator Even without negative regulation, we have
low levels of operon expression due to poor match of consensus sequence for the factor• A positive regulator (CRP) promotes expression

Operon
QuickTime™ and aAnimation decompressor
are needed to see this picture.

Lac Operon
QuickTime™ and a decompressor
are needed to see this picture.

Open Reading Frames
• Recall - 3 of the 64 codons are stop codons (UAA, UAG, UGA) - they cause translation to stop
• Most prokaryotic proteins are longer than 60 amino acids Since on average we expect to find a stop
codon once in every 21 (3/64) codons, the presence of a run of 30 or more codons with no stop codons (an Open Reading Frame - ORF) is good evidence that we are looking at the coding sequence of a prokaryotic gene

Open Reading Frames
• AUG is a start codon Defines where translation begins If no likely promoter sequences are
found upstream of a start codon at the start of an ORF before the end of the preceding ORF, assume the two genes are part of an operon whose promoter sequence is further upstream

Termination Sequence
• Most prokaryotic operons contain specific signals for the termination of transcription called intrinsic terminators Must have a sequence of nucleotides that
includes an inverted repeat followed by A run of roughly six uracils The inverted repeat allows the RNA to form a
loop structure that greatly slows down RNA synthesis Together with the chemical properties of uracil, this is
enough to end transcription

Termination Sequence
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

GC Content in Prokaryotic Genomes
• For every G within a double-stranded DNA genome there must be a C - likewise an A for every T Only constraint on fraction of nucleotides that
are G/C as opposed to A/T is that the two must add to 100%
Can use genomic GC content to identify bacterial species (ranges from 25% to 75%)
Can also use GC content to identify genes that have been obtained from other bacteria by horizontal gene transfer

Prokaryotic Gene Densities
• Gene density within prokaryotic genomes is very high Between 85% and 88% of the
nucleotides are typically associated with coding regions of genes
Just as large portions of chromosomes can be acquired, they can also be deleted Portions left are those which code for
essential genes

Gene Recognition in Prokaryotes
• Long ORFs (60 or more codons)• Matches to simple promoter
sequences• Recognizable transcriptional
termination signal (inverted repeats followed by run or uracils)
• Comparison with nucleotide (or amino acid) sequences of known protein coding regions from other organisms

Eukaryotic Genomes
• Much more complex Internal membrane-bound
compartments allows wide variety of chemical environments in each cell
Multicellular organisms Each cell type has distinct gene expression
Size of genome may be larger Allows for “junk DNA”
• Gene expression more complex and flexible than in prokaryotes

Eukaryotic Gene Structure

Promoter Elements
• Each different cell type requires different gene expression Therefore eukaryotes have elaborate
mechanisms for starting transcription Prokaryotes have a single RNA
polymerase - eukaryotes have three RNA polymerase I - Ribosomal RNAs RNA polymerase II - Protein-coding genes RNA polymerase III - tRNAs, other small RNAs

Promoter Elements
• Most RNA polymerase II promoters contain a set of sequences known as a basal promoter where an initiation complex is assembled and transcription begins
• Also have several upstream promoter elements (typically at least 5) to which other proteins bind Without the proteins binding
upstream, initiation complex assembly is difficult
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

Promoter Elements
• RNA polymerase II does not directly recognize the basal sequences of promoters Basal transcription factors
including a TATA-binding protein (TBP) and at least 12 TBP-associated factors bind to the promoter in a specific order, facilitating binding of RNA polymerase TATA-box 5’-TATAWAW-3’ (W
is A or T) at -25 relative to transcriptional start site
Initiator sequence 5’-YYCARR-3’ (Y is C or T and R is G or A) at transcriptional start site
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

TranscriptionQuickTime™ and a decompressor
are needed to see this picture.

Regulatory Protein Binding Sites
• Transcription initiation in eukaryotes relies heavily on positive regulation Constitutive factors work on many genes
and don’t respond to external signals Regulatory factors have limited number
of genes and respond to external signals Response factors (e.g. heat shock) Cell-specific factors (e.g. pituitary cells only) Developmental factors (e.g. early embryo
organization)

Open Reading Frames
• Before translation, a heterogeneous RNA (hnRNA) is transformed into mRNA by being Capped
5’ end chemically altered
Spliced Various splicings can occur
Polyadenylated Long stretch of A’s added at 3’ end

Introns and Exons
• The introns are spliced out of the hnRNA Protein-coding genes conform to the GU-
AG rule These are the nucleotides at the 5’ and 3’
end of the intron Other nucleotides are examined as well
• Most of these are inside the intron• These signals constrain introns to be at least 60 bp
long - but there is no upper limit

Alternative Splicing
• About 20% of human genes give rise to more than one type of mRNA sequence due to alternative splicing
• Splice junctions can be masked, causing an exon to be spliced out
• The following slide shows how alternative splicing based on different splicing factors (proteins) can stop a useful protein from being produced

Alternative Splicing

GC Content
• Overall GC content between different genomes does not vary as much in eukaryotes as in prokaryotes However variations in GC content within
a genome can help us to recognize genes Of all of the pairs of nucleotides,
statistically, CG is found only at 20% of its expected value No other pair is under or over represented

GC Content
• The expected levels of are found, however, in stretches of 1 -2 kbp at the end of the 5’ ends of many human genes These are called CpG islands and are
associated with methylation Can cause make it easy for CG to mutate to
TG or CA High levels of methylation imply low levels of
acetylation of histones (a protein which, when acetylated makes transcription of DNA possible)

Isochores
• Vertebrates and plants display a level of organization called isochores that is intermediate between that of genes and chromosomes The GC content of an isochore is relatively
uniform throughout There are five classes of isochores depending
on the level of GC content Those with high GC content also have high gene
density The types of genes found in different classes differs as
well

Codon Usage Bias
• Another hint for gene hunting can be derived from the fact that every organism prefers some equivalent triplet codon to code for proteins
• Real exons generally reflect the bias while randomly chosen strings of triplets do not

Gene Recognition
• In summary, useful DNA sequence features for gene hunting include Known promoter elements (I.e. TATA
boxes) CpG islands Splicing signals associated with introns ORFs with characteristic codon utilization Similarity to the sequences of ESTs or
genes from other organisms.

Gene Expression
• Expression varies greatly however• Tools for determining gene
expression levels include cDNAs and ESTs Complementary DNAs are synthesized
from mRNAs and can be used to provide expressed sequence tags useful for contig assembly or gene recognition

cDNA

Microarrays
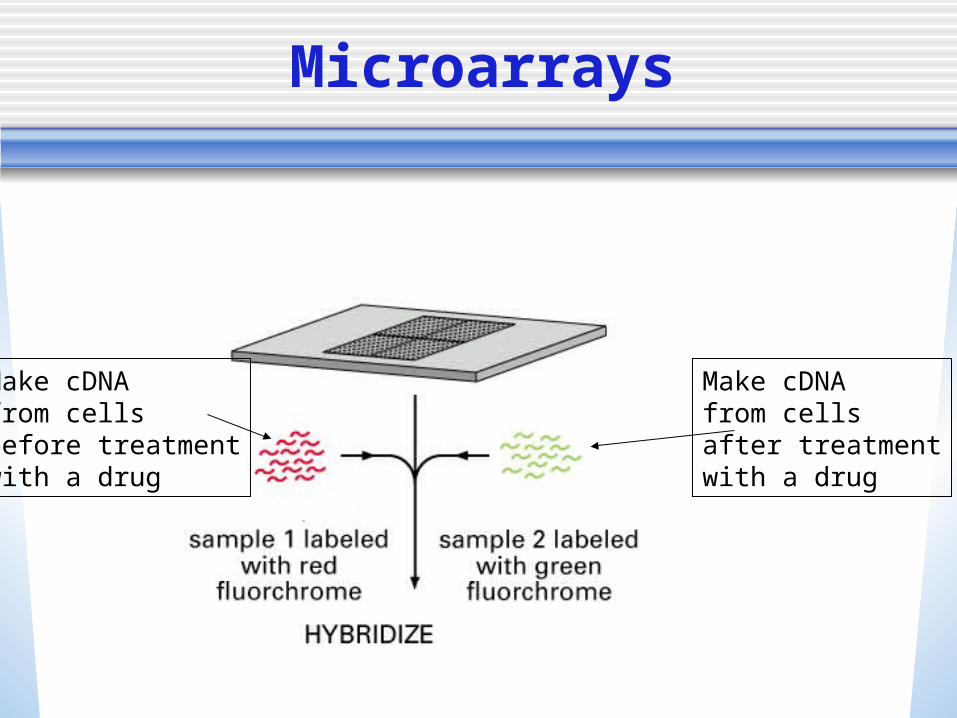
• Gene expression patterns can be studied using microarrays Small silica (glass) chips covered with
thousands of short sequences of nucleotides of known sequence
The microarray can then be used to compare the expression of all of the genes in the genome simultaneously
A gene is represented by a set of 16 probes

Microarrays
• The probes representing genes are arranged in a grid on the chip
• Fluorescently labeled cDNA from the tissue/organism we want to test is washed over the chip from the tissue/organism we want to test
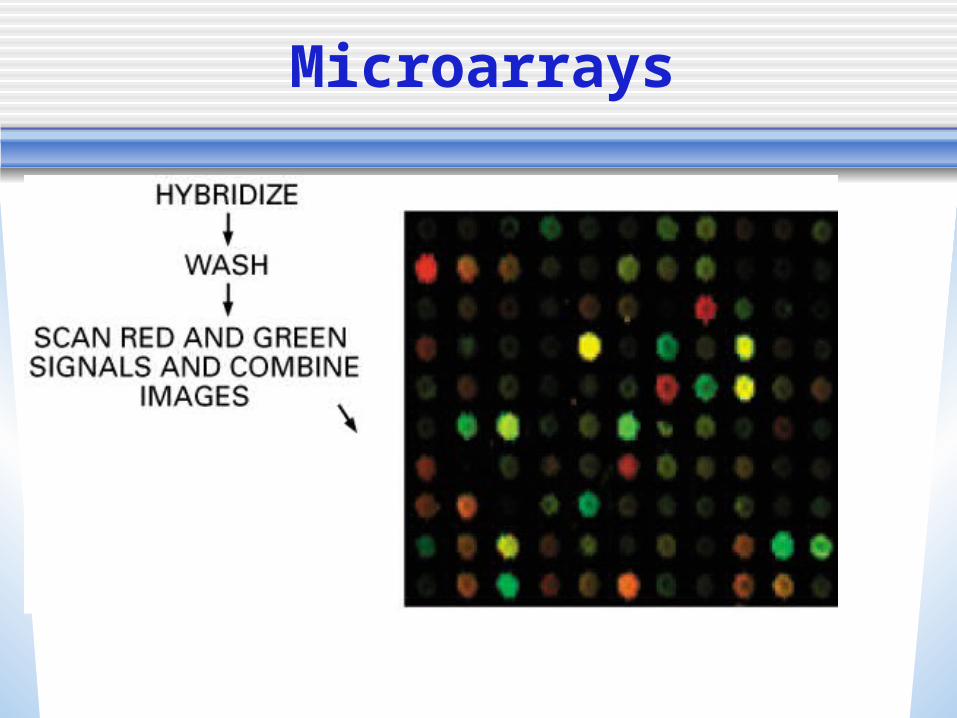
• If a gene is expressed, it will bind to the genes tags
• We can detect this through pattern recognition
Microarrays
Make cDNAfrom cells after treatmentwith a drug
Make cDNAfrom cells before treatmentwith a drug
Microarrays

Transposition
• Transposons result from insertion of duplicate sequence from another part of the genome aided by a transposase enzyme If inserted in “junk DNA”, not harmful More common are retrotransposons
which are by retroviruses (encapsulated RNA and reverse transcriptase which use a host to duplicate) like HIV

Retrovirus Replication
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

Virus Replication
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
Related Documents











