Genomic Characterization and Embryonic Expression of the Mouse Bigh3 (Tgfbi) Gene Daniel F. Schorderet,* , ² ,1 Maurice Menasche,‡ Sabine Morand,* Se ´bastien Bonnel,‡ Vale ´rie Bu ¨ chillier,* Dominique Marchant,‡ Katya Auderset,* Christophe Bonny,* Marc Abitbol,‡ and Francis L. Munier* , ² , § *Division autonome de ge ´ne ´tique Me ´dicale, CHUV, and ²Unite ´ d’Oculoge ´ne ´tique, Universite ´ de Lausanne, Lausanne, Switzerland; ‡CERTO, Faculte ´ Necker, Paris, France; and §Ho ˆpital Jules Gonin, Avenue de France 15, Lausanne, Switzerland Received June 19, 2000 Mutations in human BIGH3 (TGFB1), a gene identi- fied after treatment of an adenocarcinoma cell line with TGF-b, have been observed in patients with gran- ular Groenouw type I, Reis–Bu ¨ cklers, Thiel–Behnke, Avellino, and Lattice type I and IIIa, six autosomal dominant corneal dystrophies linked to chromosome 5q. In order to gain insight into the physiological role of this gene, we characterized the genomic structure of the mouse Bigh3 and its expression in murine em- bryos. The gene spans 30 kb on mouse chromosome 13 and has 17 exons. Embryonic expression of Bigh3 is observed in the mesenchyme of the first and second branchial archs as early as dpc 11.5 and is particularly strong in the mesenchyme of numerous tissues throughout all the development stages. In fetal eye, the expression is first seen at 11.5 dpc in the mesen- chyme surrounding the optic stalk, extends toward the sclera and choroid by 14.3 dpc and reaches the cornea by 17.5 dpc. Because the physiological role of BIGH3/Bigh3 is still largely unknown, embryonic ex- pression in organs like heart, vessels, and intestine may help to identify new functions which could be searched for in patients and in knock-out animal mod- els. The characterization of the murine structure is a prerequisite for the making of such models. © 2000 Academic Press Key Words: TGFBI; Bigh3; corneal dystrophy; gene expression; in situ hybridization. Although mutations in the BIGH3 gene, also known as TGFBI, have been reported in autosomal dominant corneal dystrophies linked to chromosome 5q (1, 2), the exact role of keratoepithelin (KE), its encoded protein, is largely unknown. BIGH3 has been isolated as a TGF-b induced gene and thus has been linked to pro- cesses where TGF-b is implicated (3): wound healing, adhesion and spreading of fibroblasts (4), and athero- sclerotic and restenotic vascular lesions (5). Recently, BIGH3 has been shown to be down-regulated in the embryonal-rhabdomyosarcoma cell line RD (6). As a secreted protein, KE is located to the periphery of the surface membrane and could act as a membrane- associated growth factor (4, 7). Immunohistology stud- ies have shown that KE is present in the corneal de- posits of patients with 5q31-linked corneal dystrophies (8 –10). In order to better understand the physiological role of BIGH3 and, as a first step in engineering a mouse knock-out model, we established the genomic structure of the mouse Bigh3 gene and analyzed its expression in developing mouse embryos. MATERIALS AND METHODS Human and murine intron– exon characterization. We have pre- viously characterized the genomic structure of the human BIGH3 and were able to align both sequences in order to determine each putative mouse exon. Murine PCR primers were synthesized accord- ing to the published cDNA sequence in regions close to the ends of each putative exons. Exon amplification was performed using these primers and conventional PCR. The PCR product of each exon was sequenced using the dye terminator system from Perkin–Elmer and electrophoresed on an ABI310 sequencer or the dideoxy technique and electrophoresed on an ALF automated sequencer from Pharma- cia. New PCR primers aiming toward the introns were then designed from the obtained sequences and each intron was amplified either by conventional PCR or long range PCR (Promega) and totally or par- tially sequenced depending on the size of the introns. The size of introns that were completely sequenced are given as precise values while the size of the remaining introns was estimated on gels and is given approximately. GenBank Accession No. bankit321614 (provisional). 1 To whom correspondence should be addressed at Division of Medical Genetics, CHUV, 1011 Lausanne, Switzerland. Fax: 14121 314 33 72. E-mail: [email protected]. Biochemical and Biophysical Research Communications 274, 267–274 (2000) doi:10.1006/bbrc.2000.3116, available online at http://www.idealibrary.com on 267 0006-291X/00 $35.00 Copyright © 2000 by Academic Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Go
DVM*LA
R
fiwuAd5oobaobsttctcBpmsepA
e
a
M3
Biochemical and Biophysical Research Communications 274, 267–274 (2000)
doi:10.1006/bbrc.2000.3116, available online at http://www.idealibrary.com on
enomic Characterization and Embryonic Expressionf the Mouse Bigh3 (Tgfbi) Gene
aniel F. Schorderet,*,†,1 Maurice Menasche,‡ Sabine Morand,* Sebastien Bonnel,‡alerie Buchillier,* Dominique Marchant,‡ Katya Auderset,* Christophe Bonny,*arc Abitbol,‡ and Francis L. Munier*,†,§
Division autonome de genetique Medicale, CHUV, and †Unite d’Oculogenetique, Universite de Lausanne,ausanne, Switzerland; ‡CERTO, Faculte Necker, Paris, France; and §Hopital Jules Gonin,venue de France 15, Lausanne, Switzerland
eceived June 19, 2000
ceiTcasBessaip(
okod
M
vapiepseacfctiwg
Mutations in human BIGH3 (TGFB1), a gene identi-ed after treatment of an adenocarcinoma cell lineith TGF-b, have been observed in patients with gran-lar Groenouw type I, Reis–Bucklers, Thiel–Behnke,vellino, and Lattice type I and IIIa, six autosomalominant corneal dystrophies linked to chromosomeq. In order to gain insight into the physiological rolef this gene, we characterized the genomic structuref the mouse Bigh3 and its expression in murine em-ryos. The gene spans 30 kb on mouse chromosome 13nd has 17 exons. Embryonic expression of Bigh3 isbserved in the mesenchyme of the first and secondranchial archs as early as dpc 11.5 and is particularlytrong in the mesenchyme of numerous tissueshroughout all the development stages. In fetal eye,he expression is first seen at 11.5 dpc in the mesen-hyme surrounding the optic stalk, extends towardhe sclera and choroid by 14.3 dpc and reaches theornea by 17.5 dpc. Because the physiological role ofIGH3/Bigh3 is still largely unknown, embryonic ex-ression in organs like heart, vessels, and intestineay help to identify new functions which could be
earched for in patients and in knock-out animal mod-ls. The characterization of the murine structure is arerequisite for the making of such models. © 2000
cademic Press
Key Words: TGFBI; Bigh3; corneal dystrophy; genexpression; in situ hybridization.
Although mutations in the BIGH3 gene, also knowns TGFBI, have been reported in autosomal dominant
GenBank Accession No. bankit321614 (provisional).1 To whom correspondence should be addressed at Division ofedical Genetics, CHUV, 1011 Lausanne, Switzerland. Fax: 14121
14 33 72. E-mail: [email protected].
267
orneal dystrophies linked to chromosome 5q (1, 2), thexact role of keratoepithelin (KE), its encoded protein,s largely unknown. BIGH3 has been isolated as aGF-b induced gene and thus has been linked to pro-esses where TGF-b is implicated (3): wound healing,dhesion and spreading of fibroblasts (4), and athero-clerotic and restenotic vascular lesions (5). Recently,IGH3 has been shown to be down-regulated in thembryonal-rhabdomyosarcoma cell line RD (6). As aecreted protein, KE is located to the periphery of theurface membrane and could act as a membrane-ssociated growth factor (4, 7). Immunohistology stud-es have shown that KE is present in the corneal de-osits of patients with 5q31-linked corneal dystrophies8–10).
In order to better understand the physiological rolef BIGH3 and, as a first step in engineering a mousenock-out model, we established the genomic structuref the mouse Bigh3 gene and analyzed its expression ineveloping mouse embryos.
ATERIALS AND METHODS
Human and murine intron–exon characterization. We have pre-iously characterized the genomic structure of the human BIGH3nd were able to align both sequences in order to determine eachutative mouse exon. Murine PCR primers were synthesized accord-ng to the published cDNA sequence in regions close to the ends ofach putative exons. Exon amplification was performed using theserimers and conventional PCR. The PCR product of each exon wasequenced using the dye terminator system from Perkin–Elmer andlectrophoresed on an ABI310 sequencer or the dideoxy techniquend electrophoresed on an ALF automated sequencer from Pharma-ia. New PCR primers aiming toward the introns were then designedrom the obtained sequences and each intron was amplified either byonventional PCR or long range PCR (Promega) and totally or par-ially sequenced depending on the size of the introns. The size ofntrons that were completely sequenced are given as precise valueshile the size of the remaining introns was estimated on gels and isiven approximately.
0006-291X/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.

Tissue preparation. Morphologically normal mice embryos andfpeaosfg2
mTdobc(thBat5GaG2AaA
thsampcAtcSsgte
R
G
bktitstcts
in both species. The exon–intron boundaries follow theG
I
c(lBsestcaottsva
Btadw1petlpppdtwTgDc(ii
(3l2taiwt
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
etuses ranging from 11.5 to 18.5 dpc were collected from NMRIregnant mice. Two embryos were processed for each stage. Thembryos were microdissected under the microscope, frozen in dry ice,nd stored at 280°C. Cryostat serial sections (15 mm) were mountedn slides previously coated with 2% 3-aminopropyltriethoxy-silaneolution in acetone. Sections were fixed during 30 min in 2% para-ormaldehyde in 0.1 M phosphate buffer (pH 7.4), dehydrated with araded ethanol series (50%, 75%, 95%), air-dried, and stored at80°C.
DNA probes for in situ hybridization. Oligonucleotide probes (60ers) were synthesized and purified by Genset (Les Ullis, France).hey were 39-end-labeled with [a-35S]dATP (NEN) using terminaleoxyribonucleotidyl transferase (GIBCO-BRL) at a specific activityf approximately 7 3 108 cpm/mg. The probes were purified oniospin P 30 columns (Bio-Rad). Two sense and 2 antisense oligonu-leotides were chosen according to the mouse Bigh3 cDNA sequenceGenBank Accession No. L19932), using the OLIGO software withhe following parameters: 55% GC content and DG . 24 kcal/mol forairpins and self-pairing. All sequences were compared to the Gen-ank and EMBL nucleotidic sequence databases using the FASTAnd the BLAST procedures, to make sure that they were specific tohe Bigh3 gene. The sequence of the oligomers were: Bigh3 sense 1,9-GCG CAC CAT ATT GAG ATC TGA GCC TTG GAC GGG TAGGA AGG GGT TAA GGG GAG AAA GGT-39 (position 2210–2269);ntisense 1, 59-ACC TTT CTC CCC TTA ACC CCT TCC CTA CCCTC CAA GGC TCA GAT CTC AAT ATG GTG CGC-39; Bigh3 sense, 59-CCA GCA GCT CTT CCG CTC TCA AAT CTG TAT GAG ACCTG GGA GTT GTG GGA TCG ACC ACC-39 (position 337–396);ntisense 2, 59-GGT GGT CGA TCC CAC AAC TCC CAT GGT CTCTA CAG ATT TGA GAG CGG AAG AGC TGC TGG-39.
In situ hybridization procedure. The hybridization cocktail con-ained 50% formamide, 43 SSC (standard saline citrate), 13 Den-ardt’s solution, 0.25 mg/ml yeast tRNA, 0.25 mg/ml sheared herringperm DNA, 0.25 mg/ml poly(A), 10% dextran sulfate, 100 mM DTTnd [a-35S] dATP-labeled probe at a concentration of 6 3 105 cpm/100l of final hybridization solution. 100 ml of hybridization solution wasut on each section. The sections were then covered with a parafilmoverslip and incubated in a humidified chamber at 43°C for 20 h.fter hybridization, the slides were washed twice in 13 SSC con-
aining 10 mM DTT for 15 min each at 55°C, and twice in 0.53 SSContaining 10 mM DTT for 15 min each at 55°C, and once in 0.53SC containing 10 mM DTT for 15 min at room temperature. Theections were then dipped in water, dehydrated with a series ofraded concentrations of ethanol and exposed to Amersham Be-amax X-ray films for 4 days and then to Kodak NTB2 photographicmulsion for 2 months at 14°C.
ESULTS
enomic Structure
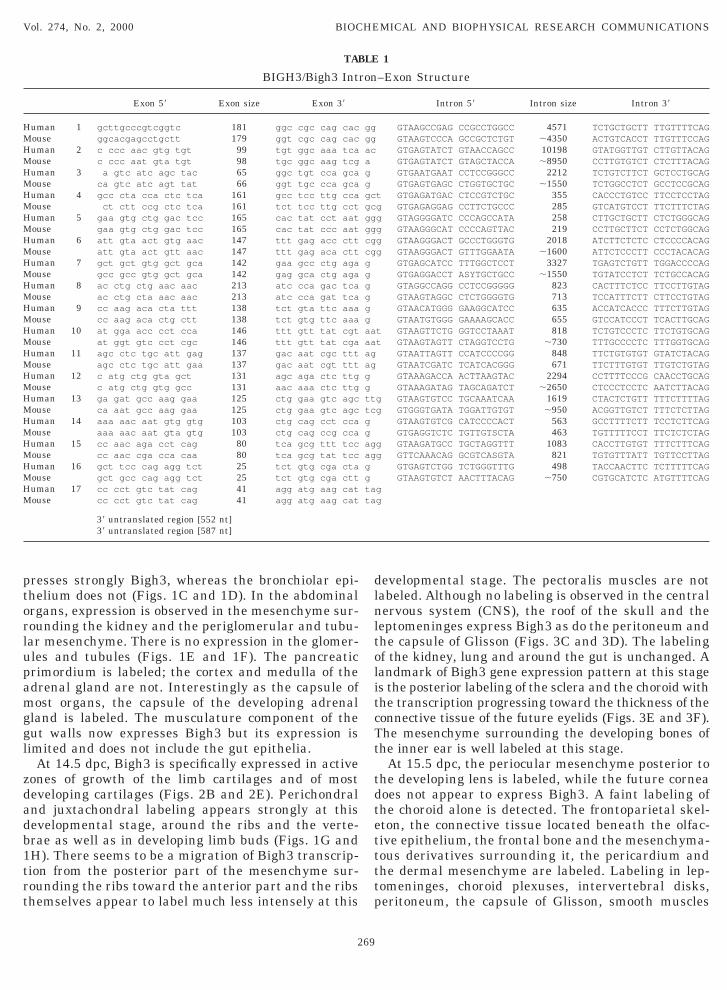
The size and structure of Bigh3 is well conservedetween human and mouse. It spans approximately 30b of genomic sequence on mouse chromosome 13 whilehe human gene is approximately 35 kb. The sequenc-ng of all the exons from genomic mouse DNA indicateshat the structure and number of the murine exons areimilar to that of the human BIGH3 and that no addi-ional exon is present. Bigh3 contains 17 exons and hasorresponding introns of approximately the same sizeo the human homolog. Like in human, intron 5 is themallest, intron 2, the largest; exon 16 is the smallest
268
T/AG rule in both genes (Table 1).
n Situ Hybridization
No labeling could be detected at embryonic day postonception 11.5 (dpc) in the central nervous systemCNS), in the skin and dermal mesenchyme, in theiver nor in the heart by in situ hybridization analysis.igh3 is expressed in the mesenchyme of the first andecond branchial arches, but not of the third. It isxpressed in developing bones like the frontal, parietal,phenoid and temporal bones. It is also expressed inhe neural crest derived mesenchyme of the VIII, IX, Xranial nerve nuclei presumably both in neural cellsnd in Schwann cells. The olfactive placode and theptic stalk are not labeled whereas the costal and ver-ebral mesenchyme and the mesenchyme surroundinghe optic stalk are. Already at this developmentaltage, the mesenchyme surrounding the developingessels is expressing Bigh3 while the endothelial cellsre not.At 12.5 dpc, dorsal root ganglia are transcribingigh3. The nonsegmented conotroncus which will
ransform into the pulmonary artery and into the aortare significantly labeled. The inferior and superior en-ocardial cushions are labeled. The heart is speciallyell labeled at this developmental stage (Figs. 1A andB), the lung starts to express Bigh3 with an exclusiveeribronchiolar labeling. The bronchiolar and the lungpithelia are not labeled at the cellular level. The myo-om derived premuscular mass (lateral mesoderm) isabeled. The mesenchyme surrounding the Ratckeouch is labeled, whereas the pouch itself is not. Ex-ression of Bigh3 is also present in the nasofrontalrimordium, the ethmoidis and the intervertebralisks. Vascular structures localized under the futureongue are labeled in the subendothelial mesenchymehile the musculature mass of the tongue itself is not.he Wolff ducts, the pharyngeal gut and all the upperut as well as the mid- and hindgut are not labeled.eveloping bones are highly expressing Bigh3: basiooc-
ipital, chondroocciput, anterior thoracic bone wallsternum and ribs primordia). The thymic primordiums well labeled. The mesenchyme of the ombilical cordiss significantly labeled, while the caudal appendix is not.
At 13.5 dpc, the CNS and the skin are not labeledFigs. 2A and 2D). The choroid plexus are labeled (Figs.A and 3B). The vertebrae, ribs as well as juxtacarti-aginous mesenchyme are still labeled (Figs. 2A andB). Bigh3 expression is visible in the primordium ofhe nasal capsule and ethmoidis, in Meckel’s cartilagend in the basis of the skull. Dorsal root ganglia label-ng persists. The vertebrae and ribs as well as thehole anterior thoracic wall are still labeled. In lungs,
he mesenchyme surrounding the bronchioles ex-
ptorlupamggl
zdadb1trt
dlnltolitcTt
tdtettttp
TABLE 1
H GM AGH AGM GH AGM AGH AGM GH AGM AGH AGM AGH AGM AGH GM GH AGM AGH AGM AGH AGM GH AGM AGH GM GH GM GH GM GH GM AGHM
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
resses strongly Bigh3, whereas the bronchiolar epi-helium does not (Figs. 1C and 1D). In the abdominalrgans, expression is observed in the mesenchyme sur-ounding the kidney and the periglomerular and tubu-ar mesenchyme. There is no expression in the glomer-les and tubules (Figs. 1E and 1F). The pancreaticrimordium is labeled; the cortex and medulla of thedrenal gland are not. Interestingly as the capsule ofost organs, the capsule of the developing adrenal
land is labeled. The musculature component of theut walls now expresses Bigh3 but its expression isimited and does not include the gut epithelia.
At 14.5 dpc, Bigh3 is specifically expressed in activeones of growth of the limb cartilages and of mosteveloping cartilages (Figs. 2B and 2E). Perichondralnd juxtachondral labeling appears strongly at thisevelopmental stage, around the ribs and the verte-rae as well as in developing limb buds (Figs. 1G andH). There seems to be a migration of Bigh3 transcrip-ion from the posterior part of the mesenchyme sur-ounding the ribs toward the anterior part and the ribshemselves appear to label much less intensely at this
BIGH3/Bigh3 Int
Exon 59 Exon size Exon 39
uman 1 gcttgcccgtcggtc 181 ggc cgc cag cac gouse ggcacgagcctgctt 179 ggt cgc cag cac guman 2 c ccc aac gtg tgt 99 tgt ggc aaa tca acouse c ccc aat gta tgt 98 tgc ggc aag tcg auman 3 a gtc atc agc tac 65 ggc tgt cca gca gouse ca gtc atc agt tat 66 ggt tgc cca gca guman 4 gcc cta cca ctc tca 161 gcc tcc ttg cca gctouse ct ctt ccg ctc tca 161 tct tcc ttg cct gcguman 5 gaa gtg ctg gac tcc 165 cac tat cct aat gggouse gaa gtg ctg gac tcc 165 cac tat ccc aat gguman 6 att gta act gtg aac 147 ttt gag acc ctt cggouse att gta act gtt aac 147 ttt gag aca ctt cgguman 7 gct gct gtg gct gca 142 gaa gcc ctg aga gouse gcc gcc gtg gct gca 142 gag gca ctg aga guman 8 ac ctg ctg aac aac 213 atc cca gac tca gouse ac ctg cta aac aac 213 atc cca gat tca guman 9 cc aag aca cta ttt 138 tct gta ttc aaa gouse cc aag aca ctg ctt 138 tct gtg ttc aaa guman 10 at gga acc cct cca 146 ttt gtt tat cgt aatouse at ggt gtc cct cgc 146 ttt gtt tat cga aatuman 11 agc ctc tgc att gag 137 gac aat cgc ttt agouse agc ctc tgc att gaa 137 gac aat cgt ttt aguman 12 c atg ctg gta gct 131 agc aga ctc ttg gouse c atg ctg gtg gcc 131 aac aaa ctc ttg guman 13 ga gat gcc aag gaa 125 ctg gaa gtc agc ttgouse ca aat gcc aag gaa 125 ctg gaa gtc agc tcuman 14 aaa aac aat gtg gtg 103 ctg cag cct cca gouse aaa aac aat gta gtg 103 ctg cag ccg cca guman 15 cc aac aga cct cag 80 tca gcg ttt tcc aggouse cc aac cga cca caa 80 tca gcg tat tcc agguman 16 gct tcc cag agg tct 25 tct gtg cga cta gouse gct gcc cag agg tct 25 tct gtg cga ctt guman 17 cc cct gtc tat cag 41 agg atg aag cat taouse cc cct gtc tat cag 41 agg atg aag cat ta
39 untranslated region [552 nt]39 untranslated region [587 nt]
269
evelopmental stage. The pectoralis muscles are notabeled. Although no labeling is observed in the centralervous system (CNS), the roof of the skull and the
eptomeninges express Bigh3 as do the peritoneum andhe capsule of Glisson (Figs. 3C and 3D). The labelingf the kidney, lung and around the gut is unchanged. Aandmark of Bigh3 gene expression pattern at this stages the posterior labeling of the sclera and the choroid withhe transcription progressing toward the thickness of theonnective tissue of the future eyelids (Figs. 3E and 3F).he mesenchyme surrounding the developing bones ofhe inner ear is well labeled at this stage.
At 15.5 dpc, the periocular mesenchyme posterior tohe developing lens is labeled, while the future corneaoes not appear to express Bigh3. A faint labeling ofhe choroid alone is detected. The frontoparietal skel-ton, the connective tissue located beneath the olfac-ive epithelium, the frontal bone and the mesenchyma-ous derivatives surrounding it, the pericardium andhe dermal mesenchyme are labeled. Labeling in lep-omeninges, choroid plexuses, intervertebral disks,eritoneum, the capsule of Glisson, smooth muscles
–Exon Structure
Intron 59 Intron size Intron 39
GTAAGCCGAG CCGCCTGGCC 4571 TCTGCTGCTT TTGTTTTCAGTAAGTCCCA GCCGCTCTGT ;4350 ACTGTCACCT TTGTTTCC
GTGAGTATCT GTAACCAGCC 10198 GTATGGTTGT CTTGTTACGTGAGTATCT GTAGCTACCA ;8950 CCTTGTGTCT CTCTTTACAGTGAATGAAT CCTCCGGGCC 2212 TCTGTCTTCT GCTCCTGCGTGAGTGAGC CTGGTGCTGC ;1550 TCTGGCCTCT GCCTCCGC
GTGAGATGAC CTCCGTCTGC 355 CACCCTGTCC TTCCTCCTTGAGAGGAG CCTTCTGCCC 285 GTCATGTCCT TTCTTTCTAGTAGGGGATC CCCAGCCATA 258 CTTGCTGCTT CTCTGGGCGTAAGGGCAT CCCCAGTTAC 219 CCTTGCTTCT CCTCTGGC
GTAAGGGACT GCCCTGGGTG 2018 ATCTTCTCTC CTCCCCACGTAAGGGACT GTTTGGAATA ;1600 ATTCTCCCTT CCCTACAC
GTGAGCATCC TTTGGCTCCT 3327 TGAGTCTGTT TGGACCCCGTGAGGACCT ASYTGCTGCC ;1550 TGTATCCTCT TCTGCCAC
GTAGGCCAGG CCTCCGGGGG 823 CACTTTCTCC TTCCTTGTAGTAAGTAGGC CTCTGGGGTG 713 TCCATTTCTT CTTCCTGTA
GTAACATGGG GAAGGCATCC 635 ACCATCACCC TTTCTTGTGTAATGTGGG GAAAAGCACC 655 GTCCATCCCT TCACTTGCAAGTTCTG GGTCCTAAAT 818 TCTGTCCCTC TTCTGTGC
TAAGTAGTT CTAGGTCCTG ;730 TTTGCCCCTC TTTGGTGCGTAATTAGTT CCATCCCCGG 848 TTCTGTGTGT GTATCTACGTAATCGATC TCATCACGGG 671 TTCTTTGTGT TTGTCTGTAGTAAAGACCA ACTTAAGTAC 2294 CCTTTTCCCG CAACCTGCGTAAAGATAG TAGCAGATCT ;2650 CTCCCTCCTC AATCTTAC
GTAAGTGTCC TGCAAATCAA 1619 CTACTCTGTT TTTCTTTTAGTGGGTGATA TGGATTGTGT ;950 ACGGTTGTCT TTTCTCTTAGTAAGTGTCG CATCCCCACT 563 GCCTTTTCTT TCCTCTTCAGTGAGGTCTC TGTTGTSCTA 463 TGTTTTTCCT TTCTCTCTA
GTAAGATGCC TGCTAGGTTT 1083 CACCTTGTGT TTTCTTTCAGTTCAAACAG GCGTCASGTA 821 TGTGTTTATT TGTTCCTTAGTGAGTCTGG TCTGGGTTTG 498 TACCAACTTC TCTTTTTCA
GTAAGTGTCT AACTTTACAG ;750 CGTGCATCTC ATGTTTTC
ron
gg
G
g
GTG
g
gg

LdD
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. (A and B) Labeling of the heart (stage 12.5 dpc). (C and D) peribronch lab, peribronchiolar labeling (stage 13.5 dpc). (E and F)abeling of the mesenchyme surrounding the kidney and the glomeruli (stage 13.5 dpc). (G) pericondral lab, perichondral labeling (stage 14.5pc). (H) cart, cartilage (stage 14.5 dpc). A, C, E, and G correspond to the dark-field aspects of the structures observed in bright field in B,, F, and H, respectively. Bar (1 cm), 150 mm for A–F; bar (1 cm), 75 mm for G and H.
270

sgttml
cttp
ctmnrcpitmt
opwmsBa
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
urrounding the oesophagus and the other parts of theut (Figs. 3G and 3H), the mesenchyme surroundinghe ribs, the vertebrae, the bronchioles and kidneyubules is still observed. Skeletal bones are in generalore diffusely labeled at this stage. The CNS is not
abeled in any of its parts.At 16.5 dpc, labeling is concentrated in the mesen-
hyme surrounding the vibrissae, the primary andhe secondary palate, the developing membranous den-al bone and the primordium of the dental bud, theerinevrum of the dental nerve, the mesenchymatous
FIG. 2. (A) Stage 13.5 dpc: coch, cochlea; rnc, roof of nopallialssification within cartilage ossification of rib; inc to, incisor tooth; spart of a rib. Bar (1 cm), 1.45 mm. (B) Stage 14.5 dpc: cns, central nerall of the embryo with transversal sections of the ribs; culp, cartilagm. (C) Stage 17.5 dpc: bone primordium. Bar (1 cm), 210 mm. (D) ch
urrounding metanephros. Bar (1 cm), 1.45 mm. (E) perioc mes, perioar (1 cm), 1.62 mm. (F) Bar (1 cm), 2.7 mm. A–C are magnified picre magnified pictures of autoradiograms.
271
omponents of the submaxillary gland and the connec-ive tissue surrounding the submaxillary acini. Ada-antine epithelium and the foramen of Bochdalek are
ot labeled. The thymus, the connective tissue sur-ounding the acini and Langerhans islands of the pan-reas are strongly labeled. Expression of Bigh3 is stillresent in several actively growing perichondral zones,n kidney and lung, in intervertebral disks as well as inhe nucleus pulposus, in connective tissues and in theusculature of the stomachal wall. The epithelium of
he stomach, the CNS and liver are not labeled.
tex; tb, cartilage primordium of turbinate bone; cart oss, cartilagepinal cord; met, medullary region of left metanephros; ppr, posteriors system; cllp, cartilage of the lower limb primordium; vwe, ventral
f the upper limb primordium; st, lumen of stomach. Bar (1 cm), 1.62dr lab, chondral labeling; vert, vertebral labeling; mst, mesenchymear mesenchyme; carp pr, cartilage primordium; ws, wall of stomach.es of embryonic sections observed at the binocular microscope; D–F
corc, svoue ooncultur

adbm
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 3. (A and B) chpl, plexus choroid (stage 13.5 dpc). (C and D) liv, liver; pcl, pericostal labeling; diaph, diaphragm; mu ap, musculatureponeurosis; pect mu, pectoralis muscle (stage 14.5 dpc). (E and F) ch lab, choroidal labeling: perioc mes, periocular mesenchyme (stage 14.5pc). (G and H) lab in and out wmdg, labeling of the inner and outer part of the wall midgut (stage 15.5 dpc). A, C, and G correspond to theright-field aspects of the structures observed in dark field in B, D, and H, respectively. E and F are dark-field aspects of the periocularesenchyme and of the retina at stage 14.5 dpc. Bar (1 cm), 150 mm.
272

iswcimTbcgsnmttppBtb
talmbkpl
D
aeimTotmhthtp
aHaas3s
Bigh3 expression during embryogenesis is mainlyclcmeIasechocBeccBopatBlecmpba
stakttieiitrsintaredapRs
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
At 17.5 dpc, the choroid, the sclera and the cornea,ncluding the future corneal epithelium and thetroma, are clearly expressing Bigh3, delineating thehole circumference of the eye (Figs. 2C and 2F) which
ontrasts with the preceding developmental stages. Asn the previous developmental stages, the retinal pig-
ent epithelium and the neural retina are not labeled.he ocular labeling prolongs itself towards the palpe-ral groove but does not extent to the lens. The mus-ular component of trachea, oesophagus and the entireut wall are labeled. Perichondral labeling is stilltrong in active zones of bone growth but dermal con-ective tissue is not labeled anymore at this develop-ental stage. The muscular component of the tongue,
he myocardial walls, the mesenchymal component ofhe developing alveolocapillar membrane, the pleura,ericardium, peritoneum and capsule of Glisson ex-ress Bigh3. Detailed analysis of the heart shows thatigh3 expression is high in the myocardium itself, in
he coronary arteries and in the valvular structures. Asefore, CNS, liver and epidermis are not labeled.At 18.5 dpc, the ocular labeling is mostly confined to
he anterior part of the cornea where the epitheliumppears more labeled than the developing stroma. Noabeling is observed in the retina. Intercostal liga-
ents, perivertebral and pericostal regions are la-eled. No labeling is detected in the lung, the liver andidney while labeling is still detectable in the choroidlexuses and the wall of the gut, mainly at the colicevel.
ISCUSSION
BIGH3 was identified as a TGF-b inducible gene inn adenocarcinoma cell like (3). The cDNA sequencencodes a 76-kDa protein that contains 4 domains ofnternal homology. KE is secreted in the extracellular
atrix where it can be seen as a 68-kDa protein (4). AsGF-b induces the synthesis of BIGH3, understandingf the function of KE has been centered around situa-ions where TGF-b is upregulated. The report thatutations in BIGH3 were associated with several in-erited corneal dystrophies came as a surprise andriggered a switch in the tissues studied. In order toave a broader approach, we decided to characterizehe genomic murine Bigh3 gene and to study its ex-ression in developing murine embryos.Recently, most of the human BIGH3 gene was char-
cterized by Munier et al. (1) and efforts from theuman Genome Organization (HUGO) have madevailable the rest of the genomic sequence. The humannd murine BIGH3 genes have very similar genomictructures. They both have a size of approximately0–35 kb and have 17 exons of comparable length. Theize of the introns is also remarkably similar.
273
oncentrated in tissues derived from the mesoderm,ike the mesenchyme of most organs: developing bones,artilage, blood vessels, peribronchiolar structure, thy-us, kidney. Besides the mesenchyme of these organs,
xpression of Bigh3 in three structures is interesting.n the heart, Bigh3 is highly expressed both in thedult heart (data not shown) and during embryogene-is. Expression is first seen in the superior and inferiorndocardial cushions, then in the other structures, in-luding the valvular system. Our in situ investigationas shown that Bigh3 is highly expressed in the devel-ping pancreas and Western blot analysis of bTC2ells, an insulin secreting mouse cell line, showed thatigh3 was expressed in b cells (data not shown). Noxpression was seen in the CNS. However, the mesen-hyme surrounding the vascular structures and thehoroid plexus do express Bigh3. In developing eye,igh3 is expressed in the mesenchyme surrounding theptic stalk as early as 11.5 dpc. Three days later, ex-ression is present in the posterior part of the sclerand the choroid and expression migrates toward thehickness of the connective tissue of the future eyelids.etween 16.5 and 17.5 dpc, the future corneal epithe-
ium, the stroma and the complete choroid and scleraxpress Bigh3. The embryonic analysis supports andomplements the analysis of MP78/70, the bovine ho-olog, in a 210-day-fetal calf. In the calf, the staining
attern of MP78/80 colocalizes to type VI collagen fi-ers in tissues such as developing nuchal ligament,orta, cornea and other tissues (11).Despite all the recent work on BIGH3, its function is
till mostly unknown. KE is expected to play a role inumorigenesis, but when mutated it aggregates to formmyloid deposits in the cornea and, to the best of ournowledge, in no other part of the body. Our report ofhe genomic structure of the mouse Bigh3 will facili-ate the making of knock-out mice and the understand-ng of the physiological role of Bigh3. The embryonicxpression study described here shows that Bigh3 is anmportant component of the mesenchyme and, as such,s expected to play a role in the development and main-enance of most organs. Mutations in Bigh3 or in itsegulatory sequences could lead to three speculativeituations: pathologies with increased expression lead-ng to exaggerated physiological processes as in reste-osis (5); pathologies with downregulation followed byhe loss of cellular shape and/or cell–cell interactionnd formation of tumors (12). The third group is rep-esented by mutations in the coding region which gen-rate abnormal KE isoforms. In the case of the cornealystrophies linked to 5q, abnormal isoforms aggregatend form amyloid or other non-fibrillar deposits de-ending on the molecular location of the mutation.ecently, we showed that abnormal KE isoforms arepecifically produced in diseased corneas as cultured

fiampfm(ptfwbeeosao
A
MS
R
2. Okada, M., Yamamoto, S., Inoue, Y., Watanabe, H., Maeda, N.,
1
1
1
1
Vol. 274, No. 2, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
broblasts from an identical cornea do no form suchberrant isoforms (Korvatska et al., 2000). This abnor-al KE turnover could be explained by corneal specific
rotein–protein interactions as it has been describedor choroideremia, another disease of the eye where theutated ubiquitously expressed Rab escort protein-1
REP-1) gene interacts with a retinal specific Rab27rotein in order to produce the disease (13). Alterna-ively, corneal specific proteases that either originaterom cellular types not present in our culture system orere lost during the many passages could be responsi-le for the specificity of the deposits. It is also to bexpected that mutations elsewhere in the gene mightxpress different phenotypes, not necessarily related tophthalmic disorders. From our embryonic expressiontudies, one can speculate that loss of function couldffect the development of cartilages and bones andther mesoderm-derived structures.
CKNOWLEDGMENTS
We thank Dr. Gerard Couly, CNRS, Hopital Necker Enfants-alades, Paris, for his help. This investigation was supported by the
wiss National Science Foundation Research Grant 31-52940.
EFERENCES
1. Munier, F. L., Korvatska, E., Djemai, A., Le Paslier, D., Zografos,L., Pescia, G., and Schorderet, D. F. (1997) Nature Genet. 15,247–251.
274
Shimomura, Y., Ishii, and Tano, Y. (1998) Invest. Ophthalmol.Visual Sci. 39, 1947–1953.
3. Skonier, J., Neubauer, M., Madisen, L., Bennett, K., Plow-man, G. D., and Purchio, A. F. (1992) DNA Cell Biol. 11,511–522.
4. LeBaron, R. G., Bezverkov, K. I., Zimber, M. P., Pavelec, R.,Skonier, J., and Purchio, A. F. (1995) J. Invest. Dermatol. 104,844–849.
5. O’Brien, E. R., Bennett, K. L., Garvin, M. R., Zderic, T. W.,Hinohara, T., Simpson, J. B., Kimura, T., Nobuyoshi, M., Miz-gala, H., Purchio, A., and Schwartz, S. M. (1996) Arterioscler.Thromb. Vasc. Biol. 16, 576–584.
6. Genini, M., Schwalbe, P., Scholl, F. A., and Schafer, B. W. (1996)Int. J. Cancer 66, 571–577.
7. Escribano, J., Hernando, N., Ghosh, S., Crabb, J., and Coca-Prados, M. (1994) J. Cell. Physiol. 160, 511–521.
8. Klintworth, G. K., Valnickova, Z., and Enghild, J. J. (1998)Am. J. Pathol. 152, 743–748.
9. Korvatska, E., Munier, F. L., Chaubert, P., Wang, M. X.,Mashima, Y., Yamada, M., Uffer, Zografos, L., and Schorde-ret, D. F. (1999) Invest. Ophthalmol. Visual Sci. 40, 2213–2219.
0. Takacs, L., Boross, P., Tozser, J., Modis, L. J., Toth, G., andBerta, A. (1998) Exp. Eye Res. 66, 739–745.
1. Gibson, M. A., Kumaratilake, J. S., and Cleary, E. G. (1997)J. Histochem. Cytochem. 45, 1683–1696.
2. Schenker, T., and Trueb, B. (1998) Exp. Cell Res. 239, 161–168.
3. Seabra, M. C., Ho, Y. K., and Anant, J. S. (1995) J. Biol. Chem.270, 24420–24427.
Related Documents











