Genome-Wide Analysis of Small RNAs Expressed by Yersinia pestis Identifies a Regulator of the Yop-Ysc Type III Secretion System Chelsea A. Schiano, a Jovanka T. Koo, a * Matthew J. Schipma, b Adam J. Caulfield, a Nadereh Jafari, b Wyndham W. Lathem a Northwestern University Feinberg School of Medicine, Department of Microbiology-Immunology, Chicago, Illinois, USA a ; Northwestern University Feinberg School of Medicine, Center for Genetic Medicine, Chicago, Illinois, USA b Small noncoding RNA (sRNA) molecules are integral components of the regulatory machinery for many bacterial species and are known to posttranscriptionally regulate metabolic and stress-response pathways, quorum sensing, virulence factors, and more. The Yop-Ysc type III secretion system (T3SS) is a critical virulence component for the pathogenic Yersinia species, and the regu- lation of this system is tightly controlled at each step from transcription to translocation of effectors into host cells. The contri- bution of sRNAs to the regulation of the T3SS in Yersinia has been largely unstudied, however. Previously, our lab identified a role for the sRNA chaperone protein Hfq in the regulation of components of the T3SS in the gastrointestinal pathogen Yersinia pseudotuberculosis. Here we present data demonstrating a similar requirement for Hfq in the closely related species Yersinia pestis. Through deep sequencing analysis of the Y. pestis sRNA-ome, we found 63 previously unidentified putative sRNAs in this species. We identified a Yersinia-specific sRNA, Ysr141, carried by the T3SS plasmid pCD1 that is required for the production of multiple T3SS proteins. In addition, we show that Ysr141 targets an untranslated region upstream of yopJ to posttranscription- ally activate the synthesis of the YopJ protein. Furthermore, Ysr141 may be an unstable and/or processed sRNA, which could contribute to its function in the regulation of the T3SS. The discovery of an sRNA that influences the synthesis of the T3SS adds an additional layer of regulation to this tightly controlled virulence determinant of Y. pestis. Y ersinia pestis is the causative agent of plague, a disease thought to be responsible for the mortality of over 200 million people throughout history and one that has continued to infect thou- sands of people worldwide throughout the 21st century (1–3). The plague bacillus lives a dual lifestyle, cycling between an arthropod vector and mammalian hosts, which provides a robust model for studying host-pathogen interactions and the regulation of viru- lence determinants (3–5). The coordinated production of viru- lence factors by Y. pestis as it transitions from flea to rat and back again must be stringently regulated in order for this pathogen to be successful in the host (6–9). One critical virulence determinant under tight regulation in Y. pestis is the Yop-Ysc type III secretion system (T3SS), the genes for which are carried on the plasmid pCD1 (10). This T3SS is respon- sible for delivery of effector proteins, called Yops, to the cytosol of host macrophages, neutrophils, and dendritic cells, and Yops co- ordinately work to subvert the innate immune system (11). Reg- ulation of the T3SS occurs at the transcriptional, posttranscrip- tional, and posttranslational levels (12). Activation of the T3SS is induced at mammalian host temperatures via conformational changes in the promoter regions of many yop-ysc genes (13). In addition, growth at 37°C permits the transcription and translation of LcrF, the master transcriptional activator of T3SS genes. Tran- scription of lcrF is prohibited at lower temperatures by DNA bends in its promoter and through association with the nucleoid- associated protein YmoA; upon temperature upshift, YmoA is de- graded by Clp/Lon proteases to alleviate the transcriptional re- pression of the yscW-lcrF operon (13, 14). Furthermore, synthesis of the LcrF protein is repressed by a two-stem-loop structure in the transcript that sequesters the ribosome binding site (RBS) at 26°C, which then melts at 37°C as part of a thermosensing mech- anism (15). In addition to the posttranscriptional regulation of LcrF, Yersinia modulates the synthesis of effector Yops in a post- transcriptional manner. In conjunction with the secretion chap- erone LcrH, the pore-forming protein YopD has been shown to bind the yopK mRNA in the 5= untranslated region (UTR) of the transcript (16). This binding may repress the translation of YopK, either by promoting the degradation of the yopK transcript or by competing with the ribosome for binding (17). Moreover, the half-lives of the yopH, yopE, and yscB transcripts are longer in a yopD deletion mutant of Y. pestis than the wild type, which implies that the posttranscriptional regulation of secreted effectors by YopD is not only limited to effects on yopK. Binding of the YopD- LcrH complex to target transcripts requires two specific AU-rich regions of mRNA common to many, but not all, yop transcripts, and the distance of the AU-rich regions from the RBS seems to affect the affinity of YopD for the transcript, suggesting a mecha- nism for a hierarchy of translation (17). Additionally, Y. pestis can degrade extracellular and/or mistargeted T3SS proteins via the activity of the Y. pestis-specific plasminogen activator protease Pla, thus providing a posttranslational layer of regulation to the T3SS in this species (18, 19). Previous work from our laboratory has shown that the small noncoding RNA (sRNA) chaperone Hfq also contributes to the posttranscriptional regulation of the Y. pseudotuberculosis T3SS (20). sRNAs are important regulatory elements for many bacterial Received 17 December 2013 Accepted 8 February 2014 Published ahead of print 14 February 2014 Address correspondence to Wyndham W. Lathem, [email protected]. * Present address: Jovanka T. Koo, Wheaton College, Department of Biology, Wheaton, Illinois, USA. Supplemental material for this article may be found at http://dx.doi.org/10.1128 /JB.01456-13. Copyright © 2014, American Society for Microbiology. All Rights Reserved. doi:10.1128/JB.01456-13 May 2014 Volume 196 Number 9 Journal of Bacteriology p. 1659 –1670 jb.asm.org 1659 on June 1, 2019 by guest http://jb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Genome-Wide Analysis of Small RNAs Expressed by Yersinia pestisIdentifies a Regulator of the Yop-Ysc Type III Secretion System
Chelsea A. Schiano,a Jovanka T. Koo,a* Matthew J. Schipma,b Adam J. Caulfield,a Nadereh Jafari,b Wyndham W. Lathema
Northwestern University Feinberg School of Medicine, Department of Microbiology-Immunology, Chicago, Illinois, USAa; Northwestern University Feinberg School ofMedicine, Center for Genetic Medicine, Chicago, Illinois, USAb
Small noncoding RNA (sRNA) molecules are integral components of the regulatory machinery for many bacterial species and areknown to posttranscriptionally regulate metabolic and stress-response pathways, quorum sensing, virulence factors, and more.The Yop-Ysc type III secretion system (T3SS) is a critical virulence component for the pathogenic Yersinia species, and the regu-lation of this system is tightly controlled at each step from transcription to translocation of effectors into host cells. The contri-bution of sRNAs to the regulation of the T3SS in Yersinia has been largely unstudied, however. Previously, our lab identified arole for the sRNA chaperone protein Hfq in the regulation of components of the T3SS in the gastrointestinal pathogen Yersiniapseudotuberculosis. Here we present data demonstrating a similar requirement for Hfq in the closely related species Yersiniapestis. Through deep sequencing analysis of the Y. pestis sRNA-ome, we found 63 previously unidentified putative sRNAs in thisspecies. We identified a Yersinia-specific sRNA, Ysr141, carried by the T3SS plasmid pCD1 that is required for the production ofmultiple T3SS proteins. In addition, we show that Ysr141 targets an untranslated region upstream of yopJ to posttranscription-ally activate the synthesis of the YopJ protein. Furthermore, Ysr141 may be an unstable and/or processed sRNA, which couldcontribute to its function in the regulation of the T3SS. The discovery of an sRNA that influences the synthesis of the T3SS addsan additional layer of regulation to this tightly controlled virulence determinant of Y. pestis.
Yersinia pestis is the causative agent of plague, a disease thoughtto be responsible for the mortality of over 200 million people
throughout history and one that has continued to infect thou-sands of people worldwide throughout the 21st century (1–3). Theplague bacillus lives a dual lifestyle, cycling between an arthropodvector and mammalian hosts, which provides a robust model forstudying host-pathogen interactions and the regulation of viru-lence determinants (3–5). The coordinated production of viru-lence factors by Y. pestis as it transitions from flea to rat and backagain must be stringently regulated in order for this pathogen tobe successful in the host (6–9).
One critical virulence determinant under tight regulation in Y.pestis is the Yop-Ysc type III secretion system (T3SS), the genes forwhich are carried on the plasmid pCD1 (10). This T3SS is respon-sible for delivery of effector proteins, called Yops, to the cytosol ofhost macrophages, neutrophils, and dendritic cells, and Yops co-ordinately work to subvert the innate immune system (11). Reg-ulation of the T3SS occurs at the transcriptional, posttranscrip-tional, and posttranslational levels (12). Activation of the T3SS isinduced at mammalian host temperatures via conformationalchanges in the promoter regions of many yop-ysc genes (13). Inaddition, growth at 37°C permits the transcription and translationof LcrF, the master transcriptional activator of T3SS genes. Tran-scription of lcrF is prohibited at lower temperatures by DNAbends in its promoter and through association with the nucleoid-associated protein YmoA; upon temperature upshift, YmoA is de-graded by Clp/Lon proteases to alleviate the transcriptional re-pression of the yscW-lcrF operon (13, 14). Furthermore, synthesisof the LcrF protein is repressed by a two-stem-loop structure inthe transcript that sequesters the ribosome binding site (RBS) at26°C, which then melts at 37°C as part of a thermosensing mech-anism (15). In addition to the posttranscriptional regulation ofLcrF, Yersinia modulates the synthesis of effector Yops in a post-transcriptional manner. In conjunction with the secretion chap-
erone LcrH, the pore-forming protein YopD has been shown tobind the yopK mRNA in the 5= untranslated region (UTR) of thetranscript (16). This binding may repress the translation of YopK,either by promoting the degradation of the yopK transcript or bycompeting with the ribosome for binding (17). Moreover, thehalf-lives of the yopH, yopE, and yscB transcripts are longer in ayopD deletion mutant of Y. pestis than the wild type, which impliesthat the posttranscriptional regulation of secreted effectors byYopD is not only limited to effects on yopK. Binding of the YopD-LcrH complex to target transcripts requires two specific AU-richregions of mRNA common to many, but not all, yop transcripts,and the distance of the AU-rich regions from the RBS seems toaffect the affinity of YopD for the transcript, suggesting a mecha-nism for a hierarchy of translation (17). Additionally, Y. pestis candegrade extracellular and/or mistargeted T3SS proteins via theactivity of the Y. pestis-specific plasminogen activator protease Pla,thus providing a posttranslational layer of regulation to the T3SSin this species (18, 19).
Previous work from our laboratory has shown that the smallnoncoding RNA (sRNA) chaperone Hfq also contributes to theposttranscriptional regulation of the Y. pseudotuberculosis T3SS(20). sRNAs are important regulatory elements for many bacterial
Received 17 December 2013 Accepted 8 February 2014
Published ahead of print 14 February 2014
Address correspondence to Wyndham W. Lathem, [email protected].
* Present address: Jovanka T. Koo, Wheaton College, Department of Biology,Wheaton, Illinois, USA.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.01456-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JB.01456-13
May 2014 Volume 196 Number 9 Journal of Bacteriology p. 1659 –1670 jb.asm.org 1659
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

species, are generally 50 to 500 nucleotides (nt) in length, areencoded within intergenic regions, are transcribed from theirown promoters, and contain Rho-independent terminators (21).These sRNAs frequently depend on Hfq in order to exert theirregulatory effects, which generally occur through imperfect basepairing within the 5= UTR of target mRNA sequences (22, 23).This can result in a variety of consequences, including translationinitiation or repression, altered mRNA stability, or changes inprotein activity. For example, in Escherichia coli the sRNA MicCbinds within the untranslated leader sequence of the ompC mRNAadjacent to the RBS, which represses the synthesis of the OmpCprotein (24), while the Vibrio cholerae Qrr sRNAs activate thetranslation of a diguanylate cyclase by relieving an inhibitory sec-ondary structure in the 5=UTR of this mRNA (25). Early work inthis field established the significant impact of sRNAs on the regu-lation of outer membrane protein synthesis in E. coli, and morerecently, the posttranscriptional regulation of virulence factors byHfq-dependent sRNAs in bacterial pathogens has been described.For instance, Hfq and sRNAs participate in the regulation of bio-films produced by uropathogenic E. coli, V. cholerae, and Y. pestis,as well as in the regulation of alpha-toxin synthesis in Staphylococ-cus aureus (23, 25–29). Indeed, Hfq and sRNAs have been shownto contribute to the virulence of a number of pathogens, includingboth Y. pestis and Y. pseudotuberculosis (20, 30–32).
Recently, several studies have catalogued multiple cohorts ofsRNAs expressed by Y. pestis and Y. pseudotuberculosis under var-ious conditions (30, 31, 33–35), but a specific sRNA that contrib-utes to the regulation of the T3SS in Y. pestis has not been identi-fied. Here we present evidence for the posttranscriptionalregulation of the synthesis of the T3SS effector protein YopJ by theYersinia-specific sRNA Ysr141, as well as describe the identifica-tion of 63 additional putative sRNAs expressed by Y. pestis.
MATERIALS AND METHODSReagents, bacterial strains, and growth conditions. All reagents werepurchased from Sigma-Aldrich (St. Louis, MO) unless stated otherwise.The bacterial strains used in this study are listed in Table S1 in the supple-mental material. The oligonucleotide sequences used in this study arelisted in Table S2 in the supplemental material. The plasmids used in thisstudy are listed in Table S3 in the supplemental material. Y. pestis CO92(36) and its derivatives were routinely grown at 26°C in liquid brain heartinfusion (BHI) broth (Difco) or on BHI agar. The attenuated pgm-nega-tive derivative of Y. pestis was used for this study, unless otherwise indi-cated. E. coli strains were grown at 37°C in Luria-Bertani (LB) broth or onLB agar. When necessary, these media were supplemented with kanamy-cin (50 �g/ml) or ampicillin (100 �g/ml).
Mutagenesis. To eliminate the effects of Pla on the T3SS, we generateda deletion of pla in the pgm-negative variant of Y. pestis using a modifiedform of lambda red recombination as described previously (37). A Y.pestis pgm-negative �hfq mutant strain was created in the �pla back-ground as described previously (29). A Y. pestis pgm-negative �pla�ysr141 double mutant was also generated by lambda red recombinationusing the primers ysr141 5=-509 and P1 ysr141 3=-1 to amplify a regionapproximately 500 bp upstream of ysr141 and primers ysr141 3=�535 andP4 ysr141 5=�1 to amplify a region approximately 500 bp downstream ofysr141.
Complementation of hfq and restoration of ysr141. The hfq deletionwas complemented by amplifying the coding sequence (CDS) of hfq and aregion including 492 bp upstream of the gene using primers hfq 5=-492Xho and hfq 3=-305 Bam. This construct was cloned into pUC18-mini-Tn7-R6K-Km (38) and then integrated into the chromosome at theattTn7 site. The kanamycin resistance cassette, which was flanked by FLP
recombination target sites and used for the selection of recombinants, wasexcised with the FLP recombinase as described previously (29). The ysr141deletion was restored to the wild type through homologous recombina-tion. A 1,171-bp region spanning approximately 500 bp on either side ofthe deleted portion of ysr141, including the deleted bases, was PCR am-plified and cloned into the bacteriophage lambda pir-dependent vectorpSR47S (39), the sequence of which was verified. The plasmid was subse-quently electroporated into the �ysr141 strain. Transconjugants were se-lected for with kanamycin and resolved to the wild type by passage onsucrose. The restoration of ysr141 was confirmed by PCR.
Growth curves. Bacteria were subcultured from overnight culturesinto 10 ml BHI (supplemented with 2.5 mM CaCl2, where indicated) in125-ml Erlenmeyer flasks at an optical density at 620 nm (OD620) of 0.1.Cultures were grown at 26°C or 37°C for 12 h with shaking at 250 rpm.Aliquots were removed, and the OD620 was measured every 2 h.
Generation of the YopJ antibody. A 26-mer peptide (amino acid se-quence CKNPLPHDKLDPYLPVTFYKHTQGKK) of YopJ from Y. pestisconjugated to the immunogenic carrier protein keyhole limpet hemocy-anin was synthesized by Princeton BioMolecules. The peptide was admin-istered to specific-pathogen-free rabbits, and polyclonal antibodies wereraised by Covance.
Immunoblot analysis. Y. pestis strains were cultured in BHI withmagnesium oxalate (MOX; 0.02 M Na2C2O4, 0.02 M MgCl2) to induce theT3SS in Erlenmeyer flasks at 26°C for 3 h and then transferred to 37°C foran additional 2 h (secretion-inducing conditions). Bacterial cells werecollected by centrifugation, and culture supernatants were filteredthrough 0.2-�m-pore-size syringe filters. Volumes of filtered superna-tants with equivalent ODs were precipitated by the addition of trichloro-acetic acid to 10%. Precipitated proteins were resuspended in equal vol-umes of 1 M Tris (pH 9.0) and sample buffer, separated by SDS-PAGE,and transferred to nitrocellulose. Bacterial cell pellets were washed withphosphate-buffered saline (PBS), resuspended in PBS plus lysozyme (0.5mg/ml), and incubated on ice for 30 min. Cells were lysed by sonication(three 30-s pulses), and cellular debris was removed by centrifugation.Whole-cell lysates mixed with sample buffer were separated by SDS-PAGE and transferred to nitrocellulose. Immunoblot analyses were per-formed using antibodies to YopE (J. Bliska), YopD (G. Plano), YopH (J.Bliska), YopJ, YopK (M. Marketon), YopM (S. Straley), YpkA (K.Schesser), YscF (M. Marketon), LcrF (G. Plano), and RpoA (as a loadingcontrol; M. Marketon).
RNA isolation for deep sequencing. For RNA isolation, overnightcultures of virulent Y. pestis CO92 were subcultured in BHI broth supple-mented with 2.5 mM CaCl2 to an OD620 of 0.1 and were grown to early-logphase (OD620, 0.2; 2.5 h), mid-log phase (OD620, 0.8; 5.5 h), late-log phase(OD620, 1.8; 8.5 h), and stationary phase (OD620, 4.5; 15 h) at 26°C and at37°C in a CDC-approved biosafety level 3 facility at the Ricketts RegionalBiocontainment Laboratory, Argonne National Labs. At each time point,aliquots of bacteria were removed and immediately added to 2 volumes ofRNAprotect Bacteria reagent (Qiagen). RNA was isolated and enrichedfor sRNAs using phenol-chloroform extraction, as previously described(31). Notably, the columns provided by the RiboPure Bacteria kit (Am-bion) were omitted from the isolation procedure, as they are designed toeliminate small RNA species. The quality of the RNA was verified by use ofan Experion automated electrophoresis system (Bio-Rad).
sRNA library preparation and deep sequencing. sRNA-enriched li-braries were prepared according to a previously published protocol (31).Cluster generation was performed according to the manufacturer’s in-structions (Illumina), and 36-nt single-end reads were generated on aSolexa Genome Analyzer at the Institute for Genomics and Systems Bio-logy (Argonne National Labs). The Solexa reads that passed the purityfiltering and had a unique alignment were mapped to the Y. pestis CO92reference chromosome (GenBank accession no. NC_003143) and plas-mids (pCD1, GenBank accession no. NC_003131; pPCP1, GenBank ac-cession no. NC_003132; pMT1, GenBank accession no. NC_003134).
Schiano et al.
1660 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Bioinformatics analysis of deep sequencing data. Bioinformatics wasperformed essentially as described previously (31). Illumina sequencingreads that overlap the annotated miscellaneous RNA, mRNA, rRNA,transfer-messenger RNA (tmRNA), and tRNA genes (based on theGenBank records of the Y. pestis CO92 chromosome and plasmids pre-sented above) were extracted and counted. The reads not overlappingthese annotations were considered to be intergenic and included 5= and 3=UTRs as well as potential sRNAs. Clusters of at least 50 bp in length thatformed a continuous region of coverage were extracted from the inter-genic category of reads on each strand of the chromosome or plasmids.Generated cluster sets were then analyzed using the Integrated GenomeBrowser (IGB; http://www.bioviz.org/igb/). Predicted sRNAs were exam-ined for the presence of promoters and Rho-independent terminatorsusing the BProm and TermFind/RNAFold programs (Softberry).
Northern blot analysis. sRNA-enriched total RNA isolated as de-scribed below (5 to 10 �g) was treated with DNase I (Ambion) and sepa-rated on 6% acrylamide gels (SequaGel reagents; National Diagnostics).RNA was transferred to Hybond N� membranes; the hybridization con-ditions were those described previously (31), and washes and autoradiog-raphy were performed as described previously (31). The biotinylated oli-gonucleotide probes used to detect the Ysrs are listed in Table S2 in thesupplemental material.
RNA isolation and qRT-PCR analysis of T3SS transcripts. Y. pestisstrains were grown at 37°C under secretion-inducing conditions as de-scribed above for immunoblot analysis. Aliquots of the cultures were re-moved and immediately mixed with 2 volumes of RNAprotect Bacteriareagent. RNA was isolated using a RiboPure Bacteria kit and treated withDNase, and cDNA was synthesized using SuperScript II reverse transcrip-tase (Invitrogen) and random primers (Invitrogen). Quantitative reversetranscription-PCR (qRT-PCR) of the target genes was performed in trip-licate with the SYBR green dye (Bio-Rad) using the primers listed in TableS2 in the supplemental material. The calculated threshold cycle (CT) wasnormalized to the CT of the gyrB gene (6) from the same cDNA samplebefore calculation of the fold changes using the ��CT method (40).
Plasmid copy number determination. From bacteria cultured at 37°Cunder secretion-inducing conditions, as described above, genomic DNA(gDNA) was isolated using a Wizard gDNA isolation kit (Promega) ac-cording to the manufacturer’s instructions. Quantitative PCR was per-formed on gDNA with primers specific for gyrB, pspA, and lcrV (see TableS2 in the supplemental material). Relative plasmid copy number was de-termined by the formula 2��CT, where �CT is the threshold cycle differ-ence between gyrB and the gene of interest. Data are the average andstandard error of the mean (SEM) of 3 biological replicates.
5=/3= RACE. Y. pestis was cultured under secretion-inducing condi-tions, and RNA was extracted as described above for deep sequencing. Thequality of the RNA was verified by Experion. Simultaneous determinationof the 5= and 3= ends of Ysr141 and RybB was performed using rapidamplification of cDNA ends (RACE) essentially as described previously(41). Clones were sequenced using M13 primers.
sRNA half-life determination. The decay rate and half-life of Ysr141and RybB were determined as described previously (29). Briefly, Y. pestiswas cultured in triplicate under secretion-inducing conditions as de-scribed above. After 2 h of growth at 37°C, rifampin (50 �g/ml) was addedto the cultures to stop de novo transcription. Aliquots of bacteria wereremoved at the indicated times, RNA was extracted, cDNA was synthe-sized, and the relative levels of the transcripts at each time point weremeasured by qRT-PCR. The half-life was determined by plotting the per-centage of the transcript remaining relative to the amount at time zero foreach sRNA on a linear plot. The data points were then fit with a linearcurve, and the equation t1/2 � 0.693/k was used, where t1/2 is the half-lifeand k is the slope of the line. Data are the average and SEM of 3 biologicalreplicates. Statistical analysis was performed using the Student t test.
Ysr141 target identification. Reporter strains for target identificationwere constructed as described previously (29). Briefly, the PtetO promotersequence lacking the 5= UTR (42) was PCR amplified from pWL213, to
which the 5=UTR of sycO (43) or the 34 bases upstream of the start codonof yopJ were added via the 3= primer used for amplification of PtetO. Theseproducts were subsequently joined to the CDS of the green fluorescentprotein (GFP) gene by splicing by overlap extension (SOE)-PCR andcloned into the suicide vector pWL212 (37). All sequences were con-firmed, and the reporter constructs were then introduced onto the chro-mosome of Y. pestis, Y. pestis �hfq, Y. pestis �ysr141, or Y. pestis �ysr141restored with ysr141 via Tn7-based integration, as described above. Over-night cultures of each strain were subcultured into 3 ml BHI in 15-mlculture tubes at an OD620 of 0.1. Bacteria were cultured for 2 h at 26°C;anhydrous tetracycline (ATc; 0.25 �g/ml) was then added, and the bacte-ria were subsequently cultured for an additional 4 h at 37°C. The OD620
and fluorescence of each culture were measured in a Tecan Safire2 micro-plate reader. The assay was performed in biological triplicate, and theresults are representative of those from at least 3 unique assays. Statisticalanalysis was performed with the Student t test.
RESULTSHfq participates in the regulation of the Y. pestis T3SS. Previ-ously, we demonstrated that Hfq is required for the production ofmultiple T3SS effector proteins by Y. pseudotuberculosis (20). Asthe Yop-Ysc T3SS is conserved between Y. pestis and Y. pseudotu-berculosis, we hypothesized that Hfq may also contribute to thesynthesis of Yops by Y. pestis. Multiple studies have shown, how-ever, that the Pla protease of Y. pestis degrades extracellular Yopsand, furthermore, that the levels of Pla are indirectly regulated viaHfq (1, 18, 19, 32). Therefore, to eliminate the effects of Pla on theT3SS, we deleted the CDS of pla from the pgm-negative strain of Y.pestis. In this context, we then generated an isogenic mutant lack-ing hfq; the mutation was complemented in trans by placing thenative promoter and coding sequence of hfq onto the chromo-some via the Tn7 transposase at the attTn7 site located down-stream of glmS. To test the effects of Hfq on T3SS protein produc-tion and secretion, the parental strain, the �hfq strain, and the�hfq strain complemented with hfq were cultured under type IIIsecretion-inducing conditions, cell lysates and culture superna-tants were collected, and immunoblot analyses were performedusing antibodies to YopD, YopE, YopH, YopJ, YopK, YopM,YpkA, YscF, and LcrF. The loss of Hfq resulted in reduced levels ofall T3SS proteins, with the exceptions of YopM and LcrF, exam-ined in both the cell lysate and the culture supernatant (�30%difference) (Fig. 1A). The levels of YopM appeared to be un-changed in the �hfq mutant compared to those in the wild type inthe culture supernatant, were elevated in the cell lysate, and werepartially restored by the hfq complement. We observed that theabundance of LcrF in cell lysates was also slightly increased in theabsence of Hfq.
LcrF is the master transcriptional activator of the T3SS effectorand structural genes (44). Given that LcrF levels are elevated in the�hfq mutant, despite the decreased levels of other T3SS proteins,we hypothesized that the effects of Hfq are not due to decreasedtranscript levels. To verify this, we measured the relative steady-state levels of the yopD, yopE, yopH, yopJ, yopK, yopM, ypkA, yscF,and lcrF transcripts in the �hfq strain compared to those in wild-type bacteria by qRT-PCR. In the absence of Hfq, we found thatthe transcript levels for most genes were increased (1.5- to 12-fold) compared to those in the wild type, suggesting that the de-creased protein levels are due to a posttranscriptional, Hfq-depen-dent mechanism(s) (Fig. 1B). For instance, there were 4- and6-fold increases in the levels of the yopJ and yscF transcripts, re-spectively (Fig. 1B). This highlights the posttranscriptional role of
A Yersinia-Specific sRNA Regulates the T3SS
May 2014 Volume 196 Number 9 jb.asm.org 1661
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

Hfq in the production of these proteins (Fig. 1A). In the absence ofHfq, transcript levels of yopM were increased over 6-fold com-pared to those in the wild type. Additionally, lcrF transcript levelswere elevated in the �hfq mutant, which is consistent with theincrease in protein levels observed by immunoblotting. Comple-
mentation of hfq resulted in a reduction of transcript levels com-pared to those in the �hfq mutant, and these levels were similar tothose found in wild-type bacteria (Fig. 1B). Taken together, thesedata demonstrate that Hfq contributes to the regulation of theT3SS and suggest that an Hfq-dependent sRNA (or group ofsRNAs) may be responsible for posttranscriptional regulation ofthe T3SS proteins tested in Y. pestis.
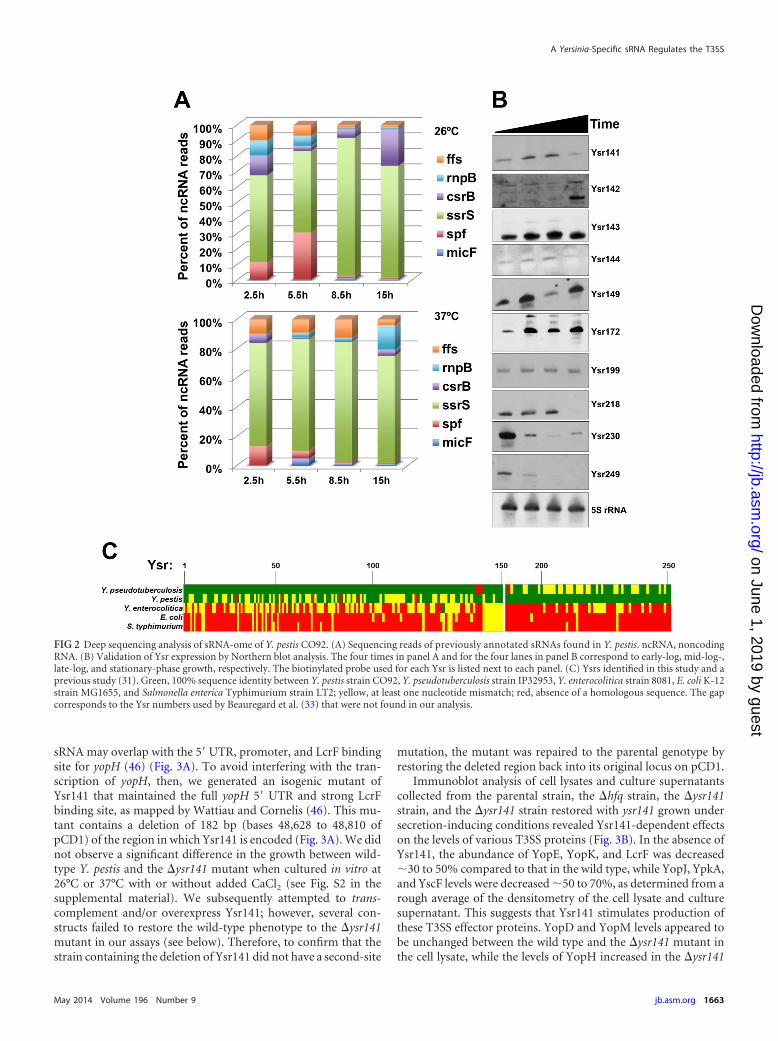
Identification of sRNAs expressed by Y. pestis. The observa-tion that Hfq is required for the regulation of components of theT3SS in Y. pestis prompted us to search for sRNAs that couldparticipate in this process. In order to identify potential candidatesRNAs (known as Ysrs, for Yersinia small RNAs), we performed adeep sequencing analysis of the sRNA-ome of Y. pestis cultured atboth 26°C and 37°C in rich broth. In addition to finding regulatorsof the T3SS, we hoped to extend our previous study of globalsRNA profiling in the related species Y. pseudotuberculosis (31).Size-selected cDNA libraries prepared from cultures grown to 4distinct growth phases at both temperatures were analyzed byIllumina-based sequencing as described previously (31). Deep se-quencing of these libraries resulted in 10 million to 16 million totalreads per sample, of which 60 to 70% corresponded to mRNAs,and 25 to 43% of the reads were aligned to the intergenic regions(see Fig. S1 in the supplemental material). The size selection andlibrary preparation were sufficient to keep the numbers of rRNAand tRNA reads low (0.05 to 0.7% and 0.5 to 1.2% of total reads,respectively). Prior to this study, 29 noncoding RNAs had beenidentified in the genome of Y. pestis. We detected 17 of these RNAsin our deep sequencing data set (see Data Set S1 in the supplemen-tal material), and of the remaining 12, 3 were antisense RNAs, 1was the tmRNA, and 1 was the 6S RNA, all of which were excludedby our filtering algorithm. Of these previously described sRNAsdetected in our study, SsrS appeared to be the most abundant at allgrowth phases and temperatures (Fig. 2A).
In order to identify additional putative sRNAs expressed by Y.pestis strain CO92, we used a modified version of the filteringalgorithm described in our previous study (31), in that the 3-folddifference in expression of RNAs between the adjacent open read-ing frames (ORFs) and the intergenic clusters was omitted. By thismethod, we uncovered 63 new potential sRNAs and generatedsequencing reads that corresponded to 144 sRNAs in Y. pestis thatwe previously identified in Y. pseudotuberculosis. We verified theexpression of 10 of these newly identified sRNAs in Y. pestis byNorthern blot analysis (Fig. 2B). Expression of most of the sRNAsvaried over the course of the growth of the bacteria, while others,such as Ysr199, remained relatively constant (Fig. 2B). In addi-tion, a significant number of the newly discovered sRNAs wereabsent from the genomes of or contained mismatches with theequivalent sequences in Y. enterocolitica, E. coli, and Salmonellaenterica serovar Typhimurium (Fig. 2C).
Ysr141 contributes to the production of T3SS proteins in Y.pestis. One sRNA gene identified by our deep sequencing analysis,Ysr141, mapped to T3SS-carrying plasmid pCD1, encoded on theopposite strand within the intergenic region between yopH andthe gene YPCD1.68c (Fig. 3A; see Data Set S1 in the supplementalmaterial). On the basis of the genomic location of Ysr141, ourability to verify its expression by Northern blotting, and theknowledge that sRNA genes sometimes occur near their targetgenes (45), we hypothesized that Ysr141may contribute to theposttranscriptional regulation of the T3SS. The transcribed regionof ysr141 predicted by our deep sequencing data suggests that this
FIG 1 Hfq is required for the synthesis of T3SS proteins. (A) Immunoblotanalysis of cell lysates and culture supernatants from the Y. pestis wild type(WT), Y. pestis �hfq, and Y. pestis �hfq complemented with hfq cultured undersecretion-inducing conditions. The antibody used to detect each protein isindicated to the left of the corresponding panel. The density of each bandrelative to that of the wild type is indicated beneath the corresponding panel.Data are representative of those from at least 3 biological replicates. (B) Rela-tive fold change of T3SS transcripts from Y. pestis �hfq and Y. pestis �hfqcomplemented with hfq, measured by qRT-PCR, compared to the level for thewild type (set equal to 1). cDNA was generated from RNA isolated from strainscultured under secretion-inducing conditions. Error bars represent the SEMsof 3 biological replicates.
Schiano et al.
1662 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
sRNA may overlap with the 5= UTR, promoter, and LcrF bindingsite for yopH (46) (Fig. 3A). To avoid interfering with the tran-scription of yopH, then, we generated an isogenic mutant ofYsr141 that maintained the full yopH 5= UTR and strong LcrFbinding site, as mapped by Wattiau and Cornelis (46). This mu-tant contains a deletion of 182 bp (bases 48,628 to 48,810 ofpCD1) of the region in which Ysr141 is encoded (Fig. 3A). We didnot observe a significant difference in the growth between wild-type Y. pestis and the �ysr141 mutant when cultured in vitro at26°C or 37°C with or without added CaCl2 (see Fig. S2 in thesupplemental material). We subsequently attempted to trans-complement and/or overexpress Ysr141; however, several con-structs failed to restore the wild-type phenotype to the �ysr141mutant in our assays (see below). Therefore, to confirm that thestrain containing the deletion of Ysr141 did not have a second-site
mutation, the mutant was repaired to the parental genotype byrestoring the deleted region back into its original locus on pCD1.
Immunoblot analysis of cell lysates and culture supernatantscollected from the parental strain, the �hfq strain, the �ysr141strain, and the �ysr141 strain restored with ysr141 grown undersecretion-inducing conditions revealed Ysr141-dependent effectson the levels of various T3SS proteins (Fig. 3B). In the absence ofYsr141, the abundance of YopE, YopK, and LcrF was decreased�30 to 50% compared to that in the wild type, while YopJ, YpkA,and YscF levels were decreased �50 to 70%, as determined from arough average of the densitometry of the cell lysate and culturesupernatant. This suggests that Ysr141 stimulates production ofthese T3SS effector proteins. YopD and YopM levels appeared tobe unchanged between the wild type and the �ysr141 mutant inthe cell lysate, while the levels of YopH increased in the �ysr141
FIG 2 Deep sequencing analysis of sRNA-ome of Y. pestis CO92. (A) Sequencing reads of previously annotated sRNAs found in Y. pestis. ncRNA, noncodingRNA. (B) Validation of Ysr expression by Northern blot analysis. The four times in panel A and for the four lanes in panel B correspond to early-log, mid-log-,late-log, and stationary-phase growth, respectively. The biotinylated probe used for each Ysr is listed next to each panel. (C) Ysrs identified in this study and aprevious study (31). Green, 100% sequence identity between Y. pestis strain CO92, Y. pseudotuberculosis strain IP32953, Y. enterocolitica strain 8081, E. coli K-12strain MG1655, and Salmonella enterica Typhimurium strain LT2; yellow, at least one nucleotide mismatch; red, absence of a homologous sequence. The gapcorresponds to the Ysr numbers used by Beauregard et al. (33) that were not found in our analysis.
A Yersinia-Specific sRNA Regulates the T3SS
May 2014 Volume 196 Number 9 jb.asm.org 1663
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

mutant in both the cell lysate and culture supernatant. Theamount of Yop proteins detected in the culture supernatant wasreduced in the absence of Ysr141 for most of the examined pro-teins, especially with YopJ, for example (Fig. 3B). Reduced pro-duction of YscF, the needle protein, in the �ysr141 mutant couldbe responsible for this phenotype. The level of production of allthe Yops examined was returned to wild-type levels with theYsr141-restored strain (Fig. 3B).
In order to determine the impact of Ysr141 on the steady-stateabundance of these T3SS transcripts, we compared the relativemRNA levels between the wild-type and �ysr141 strains by qRT-PCR. We found that 7 of 9 transcripts were not decreased morethan 2-fold (our threshold for biological significance) in the ab-sence of Ysr141 compared to the level in the wild type, which isconsistent with posttranscriptional effects (Fig. 3C). One excep-tion was found with the transcriptional activator lcrF, the level ofwhich was decreased 2.4-fold compared to the level in the wildtype. In total, these data indicate that Ysr141 contributes to theproduction of multiple T3SS proteins and suggest that, with theexception of LcrF, these effects may occur at the posttranscrip-tional level. Both the immunoblot and transcript analyses were
performed with a second independently derived mutant ofYsr141; this strain behaved identically to the original deletionstrain in these experiments (data not shown). To confirm thatthe changes that we observed are not due to an effect of Ysr141on the plasmid copy number of pCD1, we measured the abun-dance of the lcrV gene (carried by pCD1) in the �ysr141 mutantrelative to that in the wild type and observed no significantdifferences (Fig. 3D).
Ysr141-dependent posttranscriptional regulation at the yopJ5= UTR. As sRNAs often exert their effects though interactionswithin the 5= UTRs of regulated mRNA transcripts, we hypothe-sized that Ysr141 may target such sequences of the T3SS effectorproteins that are decreased in the absence of Ysr141. To examinethis, we chose to investigate the regulation of YopJ synthesis. TheyopJ gene is the third gene in an operon with the T3S chaperonegene sycO and the gene encoding the secreted effector ypkA, andthe transcriptional start site has been mapped to 28 bases up-stream of the sycO translational start site (43). There is also a396-bp transcribed, noncoding region between the stop codon ofypkA and the start codon of yopJ. Bioinformatic analysis revealedthat within this region there is a putative promoter spanning 34
FIG 3 Ysr141 is a positive regulator of the T3SS. (A) Diagram illustrating the genomic context of predicted ysr141 coordinates and the ysr141 deletion. Numbersindicate coordinates on Y. pestis CO92 pCD1, as determined by Parkhill et al. (77). (B) Immunoblot analysis of cell lysates and culture supernatants from Y. pestis,Y. pestis �ysr141, Y. pestis �ysr141 restored with ysr141, and Y. pestis �hfq cultured under secretion-inducing conditions. The antibody used to detect each proteinis indicated to the left of the corresponding panel. The density of each band relative to that in the wild type is indicated beneath the corresponding panel. Dataare representative of those from at least 3 biological replicates. (C) Relative fold change of T3SS transcripts from Y. pestis �ysr141 and Y. pestis �ysr141 restoredwith ysr141, measured by qRT-PCR, compared to that for the wild type (set equal to 1). cDNA was generated from RNA isolated from strains cultured undersecretion-inducing conditions. Error bars represent the SEMs of 2 biological replicates. (D) Relative copy number of pCD1 (represented by lcrV) in Y. pestis, Y.pestis �hfq, and Y. pestis �ysr141 compared to that in the wild type. pspA was used as a control for a chromosomal gene. Relative copy number was determinedfrom gDNA isolated from cultures used in immunoblot/qRT-PCR analyses. Error bars represent the SEMs of 3 biological replicates.
Schiano et al.
1664 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
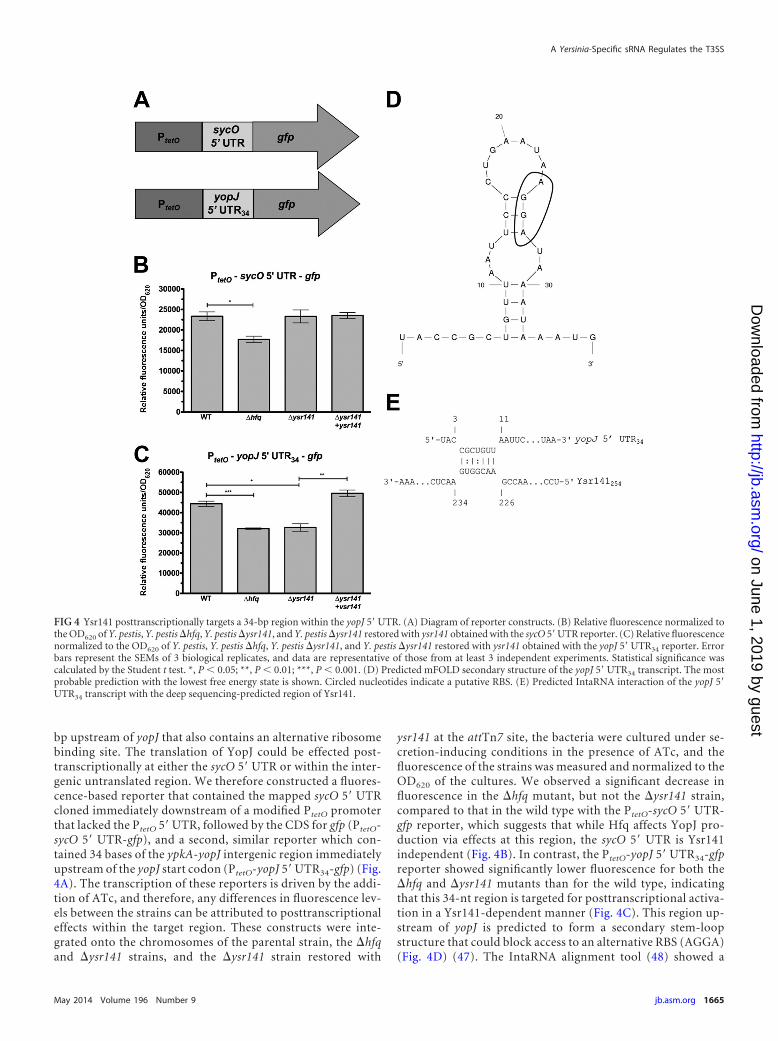
bp upstream of yopJ that also contains an alternative ribosomebinding site. The translation of YopJ could be effected post-transcriptionally at either the sycO 5= UTR or within the inter-genic untranslated region. We therefore constructed a fluores-cence-based reporter that contained the mapped sycO 5= UTRcloned immediately downstream of a modified PtetO promoterthat lacked the PtetO 5=UTR, followed by the CDS for gfp (PtetO-sycO 5= UTR-gfp), and a second, similar reporter which con-tained 34 bases of the ypkA-yopJ intergenic region immediatelyupstream of the yopJ start codon (PtetO-yopJ 5=UTR34-gfp) (Fig.4A). The transcription of these reporters is driven by the addi-tion of ATc, and therefore, any differences in fluorescence lev-els between the strains can be attributed to posttranscriptionaleffects within the target region. These constructs were inte-grated onto the chromosomes of the parental strain, the �hfqand �ysr141 strains, and the �ysr141 strain restored with
ysr141 at the attTn7 site, the bacteria were cultured under se-cretion-inducing conditions in the presence of ATc, and thefluorescence of the strains was measured and normalized to theOD620 of the cultures. We observed a significant decrease influorescence in the �hfq mutant, but not the �ysr141 strain,compared to that in the wild type with the PtetO-sycO 5= UTR-gfp reporter, which suggests that while Hfq affects YopJ pro-duction via effects at this region, the sycO 5= UTR is Ysr141independent (Fig. 4B). In contrast, the PtetO-yopJ 5= UTR34-gfpreporter showed significantly lower fluorescence for both the�hfq and �ysr141 mutants than for the wild type, indicatingthat this 34-nt region is targeted for posttranscriptional activa-tion in a Ysr141-dependent manner (Fig. 4C). This region up-stream of yopJ is predicted to form a secondary stem-loopstructure that could block access to an alternative RBS (AGGA)(Fig. 4D) (47). The IntaRNA alignment tool (48) showed a
FIG 4 Ysr141 posttranscriptionally targets a 34-bp region within the yopJ 5=UTR. (A) Diagram of reporter constructs. (B) Relative fluorescence normalized tothe OD620 of Y. pestis, Y. pestis �hfq, Y. pestis �ysr141, and Y. pestis �ysr141 restored with ysr141 obtained with the sycO 5=UTR reporter. (C) Relative fluorescencenormalized to the OD620 of Y. pestis, Y. pestis �hfq, Y. pestis �ysr141, and Y. pestis �ysr141 restored with ysr141 obtained with the yopJ 5=UTR34 reporter. Errorbars represent the SEMs of 3 biological replicates, and data are representative of those from at least 3 independent experiments. Statistical significance wascalculated by the Student t test. *, P 0.05; **, P 0.01; ***, P 0.001. (D) Predicted mFOLD secondary structure of the yopJ 5= UTR34 transcript. The mostprobable prediction with the lowest free energy state is shown. Circled nucleotides indicate a putative RBS. (E) Predicted IntaRNA interaction of the yopJ 5=UTR34 transcript with the deep sequencing-predicted region of Ysr141.
A Yersinia-Specific sRNA Regulates the T3SS
May 2014 Volume 196 Number 9 jb.asm.org 1665
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
potential 7-nt-long base-pairing interaction between Ysr141and the yopJ 5= UTR34 region immediately upstream of thestem-loop (Fig. 4E).
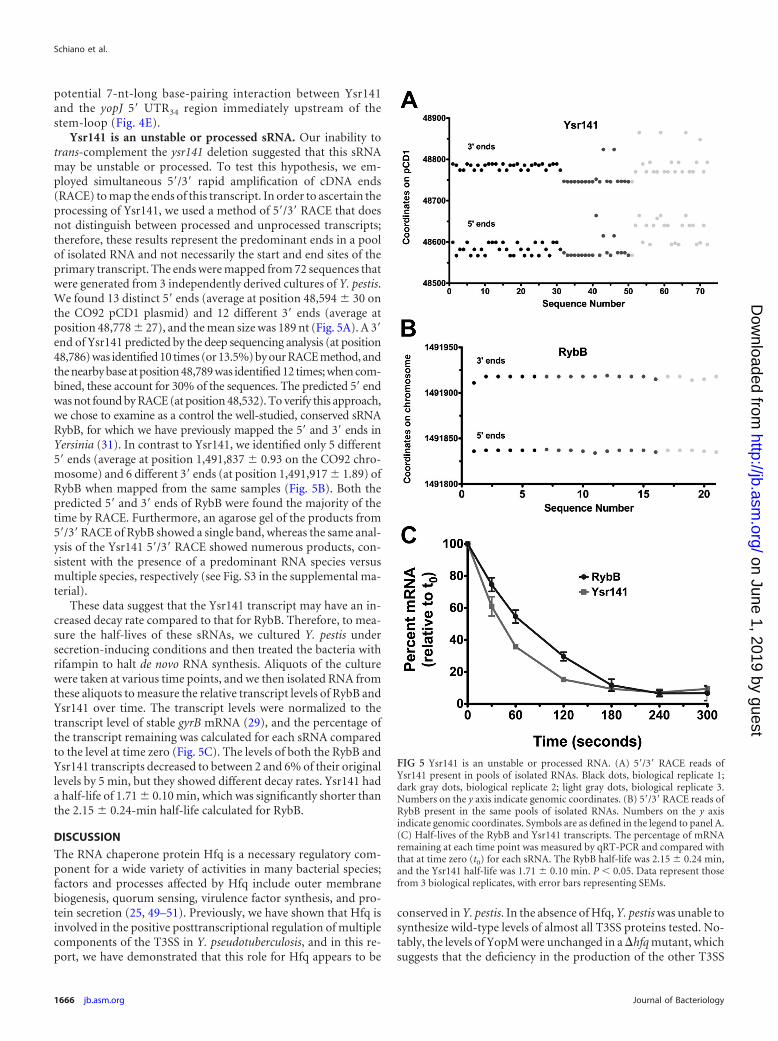
Ysr141 is an unstable or processed sRNA. Our inability totrans-complement the ysr141 deletion suggested that this sRNAmay be unstable or processed. To test this hypothesis, we em-ployed simultaneous 5=/3= rapid amplification of cDNA ends(RACE) to map the ends of this transcript. In order to ascertain theprocessing of Ysr141, we used a method of 5=/3= RACE that doesnot distinguish between processed and unprocessed transcripts;therefore, these results represent the predominant ends in a poolof isolated RNA and not necessarily the start and end sites of theprimary transcript. The ends were mapped from 72 sequences thatwere generated from 3 independently derived cultures of Y. pestis.We found 13 distinct 5= ends (average at position 48,594 30 onthe CO92 pCD1 plasmid) and 12 different 3= ends (average atposition 48,778 27), and the mean size was 189 nt (Fig. 5A). A 3=end of Ysr141 predicted by the deep sequencing analysis (at position48,786) was identified 10 times (or 13.5%) by our RACE method, andthe nearby base at position 48,789 was identified 12 times; when com-bined, these account for 30% of the sequences. The predicted 5= endwas not found by RACE (at position 48,532). To verify this approach,we chose to examine as a control the well-studied, conserved sRNARybB, for which we have previously mapped the 5= and 3= ends inYersinia (31). In contrast to Ysr141, we identified only 5 different5= ends (average at position 1,491,837 0.93 on the CO92 chro-mosome) and 6 different 3= ends (at position 1,491,917 1.89) ofRybB when mapped from the same samples (Fig. 5B). Both thepredicted 5= and 3= ends of RybB were found the majority of thetime by RACE. Furthermore, an agarose gel of the products from5=/3=RACE of RybB showed a single band, whereas the same anal-ysis of the Ysr141 5=/3= RACE showed numerous products, con-sistent with the presence of a predominant RNA species versusmultiple species, respectively (see Fig. S3 in the supplemental ma-terial).
These data suggest that the Ysr141 transcript may have an in-creased decay rate compared to that for RybB. Therefore, to mea-sure the half-lives of these sRNAs, we cultured Y. pestis undersecretion-inducing conditions and then treated the bacteria withrifampin to halt de novo RNA synthesis. Aliquots of the culturewere taken at various time points, and we then isolated RNA fromthese aliquots to measure the relative transcript levels of RybB andYsr141 over time. The transcript levels were normalized to thetranscript level of stable gyrB mRNA (29), and the percentage ofthe transcript remaining was calculated for each sRNA comparedto the level at time zero (Fig. 5C). The levels of both the RybB andYsr141 transcripts decreased to between 2 and 6% of their originallevels by 5 min, but they showed different decay rates. Ysr141 hada half-life of 1.71 0.10 min, which was significantly shorter thanthe 2.15 0.24-min half-life calculated for RybB.
DISCUSSION
The RNA chaperone protein Hfq is a necessary regulatory com-ponent for a wide variety of activities in many bacterial species;factors and processes affected by Hfq include outer membranebiogenesis, quorum sensing, virulence factor synthesis, and pro-tein secretion (25, 49–51). Previously, we have shown that Hfq isinvolved in the positive posttranscriptional regulation of multiplecomponents of the T3SS in Y. pseudotuberculosis, and in this re-port, we have demonstrated that this role for Hfq appears to be
conserved in Y. pestis. In the absence of Hfq, Y. pestis was unable tosynthesize wild-type levels of almost all T3SS proteins tested. No-tably, the levels of YopM were unchanged in a �hfq mutant, whichsuggests that the deficiency in the production of the other T3SS
FIG 5 Ysr141 is an unstable or processed RNA. (A) 5=/3= RACE reads ofYsr141 present in pools of isolated RNAs. Black dots, biological replicate 1;dark gray dots, biological replicate 2; light gray dots, biological replicate 3.Numbers on the y axis indicate genomic coordinates. (B) 5=/3= RACE reads ofRybB present in the same pools of isolated RNAs. Numbers on the y axisindicate genomic coordinates. Symbols are as defined in the legend to panel A.(C) Half-lives of the RybB and Ysr141 transcripts. The percentage of mRNAremaining at each time point was measured by qRT-PCR and compared withthat at time zero (t0) for each sRNA. The RybB half-life was 2.15 0.24 min,and the Ysr141 half-life was 1.71 0.10 min. P 0.05. Data represent thosefrom 3 biological replicates, with error bars representing SEMs.
Schiano et al.
1666 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

proteins may not be due to alterations in a pan-T3SS regulatorymechanism. In addition, the increase in transcript levels, particu-larly in the lcrF mRNA, in the absence of Hfq correlated with themicroarray data presented in an earlier report (30). Geng et al.speculated that Hfq participates in the repression of this systemunder certain conditions (30); however, our study illustrates thatthe T3SS of Y. pestis is activated in an Hfq-dependent mannerunder the conditions examined here. Interestingly, the loss of Hfqdoes not influence the synthesis of pYV-encoded T3SS proteins inY. enterocolitica, suggesting differences in the Hfq-dependentposttranscriptional control of virulence determinants among Yer-sinia species (52).
The transcription of T3SS genes is regulated through a nega-tive-feedback loop, whereby low levels of T3SS proteins in the cellresult in increased transcription, and vice versa (53, 54). We pos-tulate that in the absence of Hfq, Y. pestis is unable to synthesizeadequate levels of T3SS proteins, a condition which signals to thecell that more T3SS transcripts are needed. This leads to the ele-vated mRNA levels that both we and Geng et al. (30) observed byqRT-PCR and microarray analysis, respectively. The increasedsynthesis of the T3SS master transcriptional activator LcrF by the�hfq mutant of Y. pestis may be an effort to enhance yop-ysc tran-scription; however, the end result is still a net decrease in proteinlevels, suggesting that Hfq and one or more sRNAs are actingdownstream of LcrF to regulate the T3SS.
As Hfq typically acts in conjunction with sRNAs to exert itsregulatory effects on the cell, we employed a genome-wide deepsequencing analysis to catalog the sRNA-ome of Y. pestis strainCO92 and, consequently, identified 63 additional putative sRNAsin this species. These data build upon our previous study as well asthose from several other groups that have identified sRNAs inYersinia (31, 33–35). While there exists some overlap in the sRNAsets discovered in each of these studies, there are a large number ofpotential sRNAs that are unique to each study. Recently, Beaure-gard et al. identified 15 previously unannotated sRNAs in theKIM6� strain of Y. pestis (33). This group also identified severalpreviously annotated sRNAs and multiple Ysrs described in ourprevious study; we subsequently detected in Y. pestis CO92 theexpression of 2 Ysrs found in the KIM6� strain (Ysr164 andYsr172). A second report identified 104 sRNAs in Y. pestis strain201 from both in vitro- and in vivo-derived samples, and 56 ofthese were not identified by our previous study or by the study ofBeauregard et al. (33). In the current study, only one of the sRNAsidentified (Ysr222 or sR049) was also detected by Yan et al. (35).The predicted region of a second sRNA (Ysr196) identified in thecurrent study overlaps significantly with an sRNA identified byYan et al. (35) (sR064); however, it is encoded on the oppositestrand.
The variability in these data sets may stem from the differentstrains and methodologies used for each study. Indeed, all strainsexamined are associated with distinct biovars, each of which hasunique genetic traits (55). The RNA used in our deep sequencinganalysis was isolated from the fully virulent strain of Y. pestisCO92, which is a clinical isolate from a fatal case of pneumonicplague (56). Use of this strain allowed us to uncover the sRNAsthat are expressed under the full genetic battery of a bacteriumthat is virulent to humans and allowed the identification of plas-mid-encoded Ysrs, specifically, Ysr141, which could not have beenidentified using a strain that lacks the pCD1 plasmid (such asKIM6�).
This report brings the total number of potential sRNAs that wehave identified between Y. pestis and Y. pseudotuberculosis to 216.As our current analysis and the study in which we identifiedsRNAs expressed by Y. pseudotuberculosis (31) were performedessentially identically, our data sets permit a direct comparison ofsRNAs between these highly similar species and will be useful forunderstanding the evolution of Yersinia species through changesin sRNA content and expression. While the majority of the candi-date sRNA genes that we identified are common to both Y. pestisand Y. pseudotuberculosis, the loci encoding 5 sRNAs expressed byY. pestis CO92 are missing from the Y. pseudotuberculosis IP32953genome. When one considers that Y. pseudotuberculosis also con-tains six sRNA genes that are absent from Y. pestis, one of which isrequired for virulence (31), these differences suggest the possibil-ity that the disparate disease manifestations caused by each speciesmay be due in part to their divergent sRNA-omes (57). It is alsopossible that the targets of conserved sRNAs are differentially con-trolled between the two species, as the expression patterns of theconserved sRNAs are in some cases distinct. For example, Ysr214is encoded on the genomes of both Y. pestis and Y. pseudotubercu-losis but is detectable both by deep sequencing and by Northernblotting only in Y. pestis (see Data Set S1 in the supplementalmaterial). Additionally, thermal control of sRNA gene expressionis likely to contribute to differences, as temperature plays an im-portant role in Yersinia pathogenesis. Furthermore, many of thesRNAs that are encoded and expressed by both species containsingle nucleotide variations. Regulation of common targets bysRNAs with differences in nucleotide sequence could result indistinct interactions with the mRNA between the species.
This is the first report of a specific sRNA linked to the regula-tion of the T3SS in Yersinia. Transcript levels of the T3SS genesgenerally did not change more than 2-fold, considered the thresh-old for biological significance, between the �ysr141 and wild-typestrains. This indicates that the decreases in protein levels in theabsence of Ysr141 may not be entirely attributable to decreasedmRNA but, rather, may be attributable to a deficiency in proteinproduction through transcript degradation, translation efficiency,or posttranslational effects. Whether Ysr141 targets transcriptsdirectly or through an intermediate factor that then acts posttran-scriptionally on the targets is not yet known. Furthermore, whilewe have demonstrated a link between Ysr141 and the 34 mostproximal bases of the yopJ 5=UTR, our assays also indicate that the5= UTR of the sycO-ypkA-yopJ operon is not activated in anYsr141-dependent manner, yet YpkA levels are decreased in theabsence of Ysr141. However, the actions of sRNAs are not alwaysconfined to the 5=UTR; therefore, Ysr141 or another sRNA couldaffect ypkA translation by base pairing within the coding region;indeed, discoordinate regulation of operons by sRNAs has beendemonstrated in other bacterial species (58–60). Alternatively, thetranslation of YopJ could be the only direct target of Ysr141, andthe other observed changes could be due to the dysregulation ofYopJ production, as the Yersinia T3SS is known to be sensitive tochanges in effector protein levels (61).
In addition, the disparity between the �ysr141 and �hfq mu-tant phenotypes suggests that there may be additional sRNAs thatcontribute to the regulation of the T3SS. A second pCD1-encodedputative sRNA gene that could be an additional regulator of thesystem was identified by Yan et al. (35). Alternatively, the �hfqmutant has a general growth defect that is not seen with the�ysr141 strain, and the elaboration of the T3SS is known to be an
A Yersinia-Specific sRNA Regulates the T3SS
May 2014 Volume 196 Number 9 jb.asm.org 1667
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

energy-rich process, which could account for the significant de-crease in T3SS protein production in this mutant. Interestingly,Ysr141 does not seem to affect YopM levels, which indicates thatthis effector is not part of the Ysr141-controlled regulon.
Several examples of Hfq- and sRNA-dependent regulation oftype III secretion systems in other bacteria have recently beenreported. For example, Hfq has been shown to be involved in theregulation of locus of enterocyte effacement-encoding T3SS genesin enterohemorrhagic E. coli (EHEC) (62, 63). In addition, in theplant pathogen Xanthomonas campestris pv. vesicatoria, the sRNAxS13, which is responsive to high salt concentrations and hightemperatures, has been suggested to be an indirect positive regu-lator of secretion by affecting the levels of a key transcriptionalregulator of the T3SS (64). In Shigella dysenteriae, the iron-re-sponsive sRNA RyhB has been implicated in the repression of thetranscriptional activator VirB, which leads to reduced transcrip-tion of T3SS genes (65). Interestingly, Y. pestis encodes 2 RyhBsRNAs (RyhB1 and RyhB2), which could also influence the regu-lation of the T3SS (31, 66). It is of great interest to determine howYsr141 links the posttranscriptional activation of the T3SS to en-vironmental signals in Yersinia and whether these mechanisms aresimilar to those found in other species that regulate their T3SSs viaHfq and sRNAs. In addition, as some sRNAs are known to havemultiple targets, it is possible that Ysr141 may contribute to theregulation of factors outside the T3SS.
We have shown through RACE analysis that the processed 5=and 3= ends of Ysr141 are variable. Our data are consistent withfindings from other groups that have mapped the ends of sRNAsin Yersinia and in other bacterial species, insofar as many individ-ual sRNAs seem to have multiple start and end sites within the cellat any given time (33, 67). The prevailing theory is that, unliketheir eukaryotic counterparts, microRNAs, bacterial sRNAs arenot generally processed (21). However, the fact that sRNAs aretranscribed from their own promoters and do not depend on aDicer homolog to become functional does not preclude thesesRNAs from being processed as an integral part of their regulatorymechanism. For example, several sRNAs (SraC, SraF, GlmZ) aretranscribed as long primary transcripts that are then processedinto shorter active forms (68–70), and multiple active forms ofDsrA and GadY can be detected in a cell, despite being transcribedfrom a single promoter (71–73). We were unable to identify thedeep sequencing-predicted 5= end of Ysr141, which could be dueto misidentification of the end by deep sequencing analysis, or it ispossible that rapid processing of this transcript may prevent de-tection by our 5=/3= RACE method. Processing or degradationcould remove Ysr141 when it is no longer needed, allow it to folddifferently upon cleavage of its 5= or 3= ends, or transform it intoan active species. In addition, the 3 replicate bacterial culturesgrown for 5=/3= RACE each had distinct patterns of 5= and 3= endsfor Ysr141, even though the ends of RybB were considerably morestable, suggesting that small changes in culture conditions, me-dium composition, or other variables can have a profound effecton the stability of Ysr141. In addition, our inability to trans-com-plement the deletion of ysr141, either by use of a plasmid or on thechromosome, may be due to its transient stability or complexprocessing strategy. If Ysr141 is as unstable as our data indicate,trans-complementation may produce the sRNA at intracellularlocations too distal from the target transcripts, which would hin-der the ability of Ysr141 to interact with them before it is degraded.However, we cannot exclude the possibility that the deletion of
Ysr141 may also have polar effects, such as modification of theDNA coiling near this site.
The Yop-Ysc T3SS is a critical component of virulence forpathogenic Yersinia species (10). It is regulated in a specific, mul-tilayered manner to ensure that effectors are synthesized and se-creted only when their delivery to host cells is needed. In addition,Yersinia, through the action of YopK, ensures that only a certainamount of effectors is delivered to each cell (74). Within thisframework, then, it is not surprising that sRNAs participate in theregulation of effector production. The regulation of Yop levels byYsr141 could contribute to the virulence of Y. pestis, given that anexcess or dearth of a given effector can disrupt the balance thatYersinia achieves during the host-pathogen interaction (75). Fur-thermore, the gene encoding Ysr141 is also present in Y. pseudo-tuberculosis and Y. enterocolitica, which raises the possibility thatthe function of this sRNA may be a mechanism to regulate theT3SS conserved among the pathogenic Yersinia species. Ysr141may link environmental cues to the modulation of the T3SS inthese species, and although the exact role of host cell contact inthe induction of translocation is still not fully understood (76), theinclusion of sRNAs such as Ysr141 in the model may aid in theresolution of this question.
ACKNOWLEDGMENTS
We thank members of the W. W. Lathem laboratory for insightful discus-sions of this work and Kurt Schesser, Melanie Marketon, James Bliska,Susan Straley, and Greg Plano for the kind donation of antibodies. Wethank Trevis Alleyne, Hannah Imlay, Lauren Bellows, and Jay Schroederfor their contributions to this work. Finally, we thank Laurianne Queneefor the isolation of Y. pestis for sRNA identification and Marc Domanusfor Illumina-Solexa sequencing.
This work used resources of the Northwestern University StructuralBiology Facility, which is generously supported by NCICCSG P30CA060553 awarded to the Robert H. Lurie Comprehensive Cancer Cen-ter. This work was sponsored by the NIH/NIAID Regional Center of Ex-cellence for Bio-Defense and Emerging Infectious Diseases Research(RCE) Program. We acknowledge membership within and support fromthe Region V Great Lakes RCE (NIH award U54 AI057153). This workwas also supported by NIH/NIAID grants R21 AI103658 and R01AI093727 to W.W.L.
REFERENCES1. Perry RD, Fetherston JD. 1997. Yersinia pestis— etiologic agent of plague.
Clin. Microbiol. Rev. 10:35– 66.2. Begier EM, Asiki G, Anywaine Z, Yockey B, Schriefer ME, Aleti P,
Ogden-Odoi A, Staples JE, Sexton C, Bearden SW, Kool JL. 2006.Pneumonic plague cluster, Uganda, 2004. Emerg. Infect. Dis. 12:460 – 467.http://dx.doi.org/10.3201/eid1203.051051.
3. Butler T. 2013. Plague gives surprises in the first decade of the 21st centuryin the United States and worldwide. Am. J. Trop. Med. Hyg. 89:788 –793.http://dx.doi.org/10.4269/ajtmh.13-0191.
4. Jarrett CO, Deak E, Isherwood KE, Oyston PC, Fischer ER, WhitneyAR, Kobayashi SD, DeLeo FR, Hinnebusch BJ. 2004. Transmission ofYersinia pestis from an infectious biofilm in the flea vector. J. Infect. Dis.190:783–792. http://dx.doi.org/10.1086/422695.
5. Lorange EA, Race BL, Sebbane F, Hinnebusch BJ. 2005. Poor vectorcompetence of fleas and the evolution of hypervirulence in Yersinia pestis.J. Infect. Dis. 191:1907–1912. http://dx.doi.org/10.1086/429931.
6. Lathem WW, Crosby SD, Miller VL, Goldman WE. 2005. Progression ofprimary pneumonic plague: a mouse model of infection, pathology, andbacterial transcriptional activity. Proc. Natl. Acad. Sci. U. S. A. 102:17786 –17791. http://dx.doi.org/10.1073/pnas.0506840102.
7. Sebbane F, Jarrett CO, Gardner D, Long D, Hinnebusch BJ. 2006. Roleof the Yersinia pestis plasminogen activator in the incidence of distinctsepticemic and bubonic forms of flea-borne plague. Proc. Natl. Acad. Sci.U. S. A. 103:5526 –5530. http://dx.doi.org/10.1073/pnas.0509544103.
Schiano et al.
1668 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

8. Vadyvaloo V, Jarrett C, Sturdevant DE, Sebbane F, Hinnebusch BJ.2010. Transit through the flea vector induces a pretransmission innateimmunity resistance phenotype in Yersinia pestis. PLoS Pathog.6:e1000783. http://dx.doi.org/10.1371/journal.ppat.1000783.
9. Motin VL, Georgescu AM, Fitch JP, Gu PP, Nelson DO, Mabery SL,Garnham JB, Sokhansanj BA, Ott LL, Coleman MA, Elliott JM, Keg-elmeyer LM, Wyrobek AJ, Slezak TR, Brubaker RR, Garcia E. 2004.Temporal global changes in gene expression during temperature transi-tion in Yersinia pestis. J. Bacteriol. 186:6298 – 6305. http://dx.doi.org/10.1128/JB.186.18.6298-6305.2004.
10. Cornelis GR, Boland A, Boyd AP, Geuijen C, Iriarte M, Neyt C, SoryMP, Stainier I. 1998. The virulence plasmid of Yersinia, an antihost ge-nome. Microbiol. Mol. Biol. Rev. 62:1315–1352.
11. Marketon MM, DePaolo RW, DeBord KL, Jabri B, Schneewind O. 2005.Plague bacteria target immune cells during infection. Science 309:1739 –1741. http://dx.doi.org/10.1126/science.1114580.
12. Schiano CA, Lathem WW. 2012. Post-transcriptional regulation of geneexpression in Yersinia species. Front. Cell. Infect. Microbiol. 2:129. http://dx.doi.org/10.3389/fcimb.2012.00129.
13. Rohde JR, Luan XS, Rohde H, Fox JM, Minnich SA. 1999. The Yersiniaenterocolitica pYV virulence plasmid contains multiple intrinsic DNAbends which melt at 37°C. J. Bacteriol. 181:4198 – 4204.
14. Jackson MW, Silva-Herzog E, Plano GV. 2004. The ATP-dependentClpXP and Lon proteases regulate expression of the Yersinia pestis type IIIsecretion system via regulated proteolysis of YmoA, a small histone-likeprotein. Mol. Microbiol. 54:1364 –1378. http://dx.doi.org/10.1111/j.1365-2958.2004.04353.x.
15. Bohme K, Steinmann R, Kortmann J, Seekircher S, Heroven AK, BergerE, Pisano F, Thiermann T, Wolf-Watz H, Narberhaus F, Dersch P.2012. Concerted actions of a thermo-labile regulator and a unique inter-genic RNA thermosensor control Yersinia virulence. PLoS Pathog.8:e1002518. http://dx.doi.org/10.1371/journal.ppat.1002518.
16. Anderson DM, Ramamurthi KS, Tam C, Schneewind O. 2002. YopDand LcrH regulate expression of Yersinia enterocolitica YopQ by a post-transcriptional mechanism and bind to yopQ RNA. J. Bacteriol. 184:1287–1295. http://dx.doi.org/10.1128/JB.184.5.1287-1295.2002.
17. Chen Y, Anderson DM. 2011. Expression hierarchy in the Yersinia typeIII secretion system established through YopD recognition of RNA.Mol. Microbiol. 80:966 –980. http://dx.doi.org/10.1111/j.1365-2958.2011.07623.x.
18. Caulfield AJ, Lathem WW. 2012. Substrates of the plasminogen activatorprotease of Yersinia pestis. Adv. Exp. Med. Biol. 954:253–260. http://dx.doi.org/10.1007/978-1-4614-3561-7_32.
19. Sodeinde OA, Sample AK, Brubaker RR, Goguen JD. 1988. Plasminogenactivator/coagulase gene of Yersinia pestis is responsible for degradation ofplasmid-encoded outer membrane proteins. Infect. Immun. 56:2749 –2752.
20. Schiano CA, Bellows LE, Lathem WW. 2010. The small RNA chaperoneHfq is required for the virulence of Yersinia pseudotuberculosis. Infect.Immun. 78:2034 –2044. http://dx.doi.org/10.1128/IAI.01046-09.
21. Gottesman S. 2005. Micros for microbes: non-coding regulatory RNAs inbacteria. Trends Genet. 21:399 – 404. http://dx.doi.org/10.1016/j.tig.2005.05.008.
22. Updegrove T, Wilf N, Sun X, Wartell RM. 2008. Effect of Hfq onRprA-rpoS mRNA pairing: Hfq-RNA binding and the influence of the 5=rpoS mRNA leader region. Biochemistry 47:11184 –11195. http://dx.doi.org/10.1021/bi800479p.
23. Bardill JP, Zhao X, Hammer BK. 2011. The Vibrio cholerae quorumsensing response is mediated by Hfq-dependent sRNA/mRNA base pair-ing interactions. Mol. Microbiol. 80:1381–1394. http://dx.doi.org/10.1111/j.1365-2958.2011.07655.x.
24. Chen S, Zhang A, Blyn LB, Storz G. 2004. MicC, a second small-RNAregulator of Omp protein expression in Escherichia coli. J. Bacteriol. 186:6689 – 6697. http://dx.doi.org/10.1128/JB.186.20.6689-6697.2004.
25. Zhao X, Koestler BJ, Waters CM, Hammer BK. 2013. Post-transcriptional activation of a diguanylate cyclase by quorum sensingsmall RNAs promotes biofilm formation in Vibrio cholerae. Mol. Micro-biol. 89:989 –1002. http://dx.doi.org/10.1111/mmi.12325.
26. Kulesus RR, Diaz-Perez K, Slechta ES, Eto DS, Mulvey MA. 2008.Impact of the RNA chaperone Hfq on the fitness and virulence potential ofuropathogenic Escherichia coli. Infect. Immun. 76:3019 –3026. http://dx.doi.org/10.1128/IAI.00022-08.
27. Xue T, Zhang X, Sun H, Sun B. 2014. ArtR, a novel sRNA of Staphylo-
coccus aureus, regulates alpha-toxin expression by targeting the 5=UTR ofsarT mRNA. Med. Microbiol. Immunol. 203:1–12. http://dx.doi.org/10.1007/s00430-013-0307-0.
28. Rempe KA, Hinz AK, Vadyvaloo V. 2012. Hfq regulates biofilm gutblockage that facilitates flea-borne transmission of Yersinia pestis. J. Bac-teriol. 194:2036 –2040. http://dx.doi.org/10.1128/JB.06568-11.
29. Bellows LE, Koestler BJ, Karaba SM, Waters CM, Lathem WW. 2012.Hfq-dependent, co-ordinate control of cyclic diguanylate synthesis andcatabolism in the plague pathogen Yersinia pestis. Mol. Microbiol. 86:661–674. http://dx.doi.org/10.1111/mmi.12011.
30. Geng J, Song Y, Yang L, Feng Y, Qiu Y, Li G, Guo J, Bi Y, Qu Y, WangW, Wang X, Guo Z, Yang R, Han Y. 2009. Involvement of the post-transcriptional regulator Hfq in Yersinia pestis virulence. PLoS One4:e6213. http://dx.doi.org/10.1371/journal.pone.0006213.
31. Koo JT, Alleyne TM, Schiano CA, Jafari N, Lathem WW. 2011. Globaldiscovery of small RNAs in Yersinia pseudotuberculosis identifies Yersinia-specific small, noncoding RNAs required for virulence. Proc. Natl. Acad.Sci. U. S. A. 108:E709 –E717. http://dx.doi.org/10.1073/pnas.1101655108.
32. Lathem WW, Schroeder JA, Bellows LE, Ritzert JT, Koo JT, Price PA,Caulfield AJ, Goldman WE. 2014. Posttranscriptional regulation of theYersinia pestis cAMP receptor protein Crp and impact on virulence. mBio5(1):e01038 –13. http://dx.doi.org/10.1128/mBio.01038-13.
33. Beauregard A, Smith EA, Petrone BL, Singh N, Karch C, McDonough KA,Wade JT. 2013. Identification and characterization of small RNAs in Yersiniapestis. RNA Biol. 10:397– 405. http://dx.doi.org/10.4161/rna.23590.
34. Qu Y, Bi L, Ji X, Deng Z, Zhang H, Yan Y, Wang M, Li A, Huang X,Yang R, Han Y. 2012. Identification by cDNA cloning of abundant sRNAsin a human-avirulent Yersinia pestis strain grown under five differentgrowth conditions. Future Microbiol. 7:535–547. http://dx.doi.org/10.2217/fmb.12.13.
35. Yan Y, Su S, Meng X, Ji X, Qu Y, Liu Z, Wang X, Cui Y, Deng Z, ZhouD, Jiang W, Yang R, Han Y. 2013. Determination of sRNA expressions byRNA-seq in Yersinia pestis grown in vitro and during infection. PLoS One8:e74495. http://dx.doi.org/10.1371/journal.pone.0074495.
36. Welkos SL, Friedlander AM, Davis KJ. 1997. Studies on the role ofplasminogen activator in systemic infection by virulent Yersinia pestisstrain CO92. Microb. Pathog. 23:211–223. http://dx.doi.org/10.1006/mpat.1997.0154.
37. Lathem WW, Price PA, Miller VL, Goldman WE. 2007. A plasminogen-activating protease specifically controls the development of primary pneu-monic plague. Science 315:509 –513. http://dx.doi.org/10.1126/science.1137195.
38. Choi KH, Gaynor JB, White KG, Lopez C, Bosio CM, Karkhoff-Schweizer RR, Schweizer HP. 2005. A Tn7-based broad-range bacterialcloning and expression system. Nat. Methods 2:443– 448. http://dx.doi.org/10.1038/nmeth765.
39. Merriam JJ, Mathur R, Maxfield-Boumil R, Isberg RR. 1997. Analysis ofthe Legionella pneumophila fliI gene: intracellular growth of a defined mu-tant defective for flagellum biosynthesis. Infect. Immun. 65:2497–2501.
40. Applied Biosystems. 1997. ABI Prism 7700 sequence detection systemuser bulletin 2. Applied Biosystems, Foster City, CA.
41. Toledo-Arana A, Dussurget O, Nikitas G, Sesto N, Guet-Revillet H,Balestrino D, Loh E, Gripenland J, Tiensuu T, Vaitkevicius K, Bar-thelemy M, Vergassola M, Nahori MA, Soubigou G, Regnault B, Cop-pee JY, Lecuit M, Johansson J, Cossart P. 2009. The Listeria transcrip-tional landscape from saprophytism to virulence. Nature 459:950 –956.http://dx.doi.org/10.1038/nature08080.
42. Lutz R, Bujard H. 1997. Independent and tight regulation of transcrip-tional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2regulatory elements. Nucleic Acids Res. 25:1203–1210. http://dx.doi.org/10.1093/nar/25.6.1203.
43. Zhan L, Yang L, Zhou L, Li Y, Gao H, Guo Z, Zhang L, Qin C, ZhouD, Yang R. 2009. Direct and negative regulation of the sycO-ypkA-yopJoperon by cyclic AMP receptor protein (CRP) in Yersinia pestis. BMCMicrobiol. 9:178. http://dx.doi.org/10.1186/1471-2180-9-178.
44. Cornelis GR, Biot T, Lambert de Rouvroit C, Michiels T, Mulder B,Sluiters C, Sory MP, Van Bouchaute M, Vanooteghem JC. 1989. TheYersinia yop regulon. Mol. Microbiol. 3:1455–1459. http://dx.doi.org/10.1111/j.1365-2958.1989.tb00129.x.
45. Jager D, Pernitzsch SR, Richter AS, Backofen R, Sharma CM, SchmitzRA. 2012. An archaeal sRNA targeting cis- and trans-encoded mRNAs viatwo distinct domains. Nucleic Acids Res. 40:10964 –10979. http://dx.doi.org/10.1093/nar/gks847.
A Yersinia-Specific sRNA Regulates the T3SS
May 2014 Volume 196 Number 9 jb.asm.org 1669
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

46. Wattiau P, Cornelis GR. 1994. Identification of DNA sequences recog-nized by VirF, the transcriptional activator of the Yersinia yop regulon. J.Bacteriol. 176:3878 –3884.
47. Hoe NP, Goguen JD. 1993. Temperature sensing in Yersinia pestis: trans-lation of the LcrF activator protein is thermally regulated. J. Bacteriol.175:7901–7909.
48. Busch A, Richter AS, Backofen R. 2008. IntaRNA: efficient prediction of bacte-rial sRNA targets incorporating target site accessibility and seed regions. Bioinfor-matics 24:2849–2856. http://dx.doi.org/10.1093/bioinformatics/btn544.
49. Tsai CS, Winans SC. 2011. The quorum-hindered transcription factorYenR of Yersinia enterocolitica inhibits pheromone production and pro-motes motility via a small non-coding RNA. Mol. Microbiol. 80:556 –571.http://dx.doi.org/10.1111/j.1365-2958.2011.07595.x.
50. Pfeiffer V, Sittka A, Tomer R, Tedin K, Brinkmann V, Vogel J. 2007. A smallnon-coding RNA of the invasion gene island (SPI-1) represses outer membraneprotein synthesis from the Salmonella core genome. Mol. Microbiol. 66:1174 –1191. http://dx.doi.org/10.1111/j.1365-2958.2007.05991.x.
51. Storz G, Vogel J, Wassarman KM. 2011. Regulation by small RNAs inbacteria: expanding frontiers. Mol. Cell 43:880 – 891. http://dx.doi.org/10.1016/j.molcel.2011.08.022.
52. Kakoschke T, Kakoschke S, Magistro G, Schubert S, Borath M, Heese-mann J, Rossier O. 2014. The RNA chaperone Hfq impacts growth,metabolism and production of virulence factors in Yersinia enterocolitica.PLoS One 9:e86113. http://dx.doi.org/10.1371/journal.pone.0086113.
53. Pettersson J, Nordfelth R, Dubinina E, Bergman T, Gustafsson M,Magnusson KE, Wolf-Watz H. 1996. Modulation of virulence factorexpression by pathogen target cell contact. Science 273:1231–1233. http://dx.doi.org/10.1126/science.273.5279.1231.
54. Cornelis G, Vanootegem JC, Sluiters C. 1987. Transcription of theyop regulon from Y. enterocolitica requires trans acting pYV and chromo-somal genes. Microb. Pathog. 2:367–379. http://dx.doi.org/10.1016/0882-4010(87)90078-7.
55. Zhou D, Tong Z, Song Y, Han Y, Pei D, Pang X, Zhai J, Li M, Cui B,Qi Z, Jin L, Dai R, Du Z, Wang J, Guo Z, Wang J, Huang P, Yang R.2004. Genetics of metabolic variations between Yersinia pestis biovars andthe proposal of a new biovar, microtus. J. Bacteriol. 186:5147–5152. http://dx.doi.org/10.1128/JB.186.15.5147-5152.2004.
56. Doll JM, Zeitz PS, Ettestad P, Bucholtz AL, Davis T, Gage K. 1994.Cat-transmitted fatal pneumonic plague in a person who traveled fromColorado to Arizona. Am. J. Trop. Med. Hyg. 51:109 –114.
57. Lathem WW. 2012. Identification of small, noncoding RNAs in patho-genic Yersinia species: implications for evolution and virulence. Virulence3:154 –156. http://dx.doi.org/10.4161/viru.19155.
58. Bouvier M, Sharma CM, Mika F, Nierhaus KH, Vogel J. 2008. SmallRNA binding to 5=mRNA coding region inhibits translational initiation.Mol. Cell 32:827– 837. http://dx.doi.org/10.1016/j.molcel.2008.10.027.
59. Hayashi G, Hong C, Hagihara M, Nakatani K. 2012. Activation ofprokaryotic translation by antisense oligonucleotides binding to codingregion of mRNA. Biochem. Biophys. Res. Commun. 429:105–110. http://dx.doi.org/10.1016/j.bbrc.2012.10.072.
60. Vanderpool CK, Balasubramanian D, Lloyd CR. 2011. Dual-functionRNA regulators in bacteria. Biochimie 93:1943–1949. http://dx.doi.org/10.1016/j.biochi.2011.07.016.
61. Stainier I, Iriarte M, Cornelis GR. 1997. YscM1 and YscM2, twoYersinia enterocolitica proteins causing downregulation of yop transcrip-tion. Mol. Microbiol. 26:833– 843. http://dx.doi.org/10.1046/j.1365-2958.1997.6281995.x.
62. Kendall MM, Gruber CC, Rasko DA, Hughes DT, Sperandio V. 2011.Hfq virulence regulation in enterohemorrhagic Escherichia coli O157:H7strain 86-24. J. Bacteriol. 193:6843– 6851. http://dx.doi.org/10.1128/JB.06141-11.
63. Hansen AM, Kaper JB. 2009. Hfq affects the expression of the LEE patho-genicity island in enterohaemorrhagic Escherichia coli. Mol. Microbiol.73:446 – 465. http://dx.doi.org/10.1111/j.1365-2958.2009.06781.x.
64. Schmidtke C, Abendroth U, Brock J, Serrania J, Becker A, Bonas U.2013. Small RNA sX13: a multifaceted regulator of virulence in the plantpathogen Xanthomonas. PLoS Pathog. 9:e1003626. http://dx.doi.org/10.1371/journal.ppat.1003626.
65. Murphy ER, Payne SM. 2007. RyhB, an iron-responsive small RNAmolecule, regulates Shigella dysenteriae virulence. Infect. Immun. 75:3470 –3477. http://dx.doi.org/10.1128/IAI.00112-07.
66. Deng Z, Meng X, Su S, Liu Z, Ji X, Zhang Y, Zhao X, Wang X, Yang R,Han Y. 2012. Two sRNA RyhB homologs from Yersinia pestis biovarmicrotus expressed in vivo have differential Hfq-dependent stability. Res.Microbiol. 163:413– 418. http://dx.doi.org/10.1016/j.resmic.2012.05.006.
67. Soutourina OA, Monot M, Boudry P, Saujet L, Pichon C, Sismeiro O,Semenova E, Severinov K, Le Bouguenec C, Coppee JY, Dupuy B,Martin-Verstraete I. 2013. Genome-wide identification of regulatoryRNAs in the human pathogen Clostridium difficile. PLoS Genet.9:e1003493. http://dx.doi.org/10.1371/journal.pgen.1003493.
68. Argaman L, Hershberg R, Vogel J, Bejerano G, Wagner EG, Margalit H,Altuvia S. 2001. Novel small RNA-encoding genes in the intergenic re-gions of Escherichia coli. Curr. Biol. 11:941–950. http://dx.doi.org/10.1016/S0960-9822(01)00270-6.
69. Vogel J, Bartels V, Tang TH, Churakov G, Slagter-Jager JG, Hutten-hofer A, Wagner EG. 2003. RNomics in Escherichia coli detects new sRNAspecies and indicates parallel transcriptional output in bacteria. NucleicAcids Res. 31:6435– 6443. http://dx.doi.org/10.1093/nar/gkg867.
70. Viegas SC, Arraiano CM. 2008. Regulating the regulators: how ribonu-cleases dictate the rules in the control of small non-coding RNAs. RNABiol. 5:230 –243.
71. Majdalani N, Cunning C, Sledjeski D, Elliott T, Gottesman S. 1998.DsrA RNA regulates translation of RpoS message by an anti-antisensemechanism, independent of its action as an antisilencer of transcription.Proc. Natl. Acad. Sci. U. S. A. 95:12462–12467. http://dx.doi.org/10.1073/pnas.95.21.12462.
72. Li LC, Okino ST, Zhao H, Pookot D, Place RF, Urakami S, Enokida H,Dahiya R. 2006. Small dsRNAs induce transcriptional activation in hu-man cells. Proc. Natl. Acad. Sci. U. S. A. 103:17337–17342. http://dx.doi.org/10.1073/pnas.0607015103.
73. Repoila F, Gottesman S. 2001. Signal transduction cascade for regulationof RpoS: temperature regulation of DsrA. J. Bacteriol. 183:4012– 4023.http://dx.doi.org/10.1128/JB.183.13.4012-4023.2001.
74. Dewoody R, Merritt PM, Houppert AS, Marketon MM. 2011. YopKregulates the Yersinia pestis type III secretion system from within hostcells. Mol. Microbiol. 79:1445–1461. http://dx.doi.org/10.1111/j.1365-2958.2011.07534.x.
75. Brodsky IE, Medzhitov R. 2008. Reduced secretion of YopJ by Yersinialimits in vivo cell death but enhances bacterial virulence. PLoS Pathog.4:e1000067. http://dx.doi.org/10.1371/journal.ppat.1000067.
76. Plano GV, Schesser K. 2013. The Yersinia pestis type III secretion system:expression, assembly and role in the evasion of host defenses. Immunol.Res. 57:237–245. http://dx.doi.org/10.1007/s12026-013-8454-3.
77. Parkhill J, Wren BW, Thomson NR, Titball RW, Holden MT, Prentice MB,Sebaihia M, James KD, Churcher C, Mungall KL, Baker S, Basham D, BentleySD, Brooks K, Cerdeno-Tarraga AM, Chillingworth T, Cronin A, Davies RM,Davis P, Dougan G, Feltwell T, Hamlin N, Holroyd S, Jagels K, Karlyshev AV,Leather S, Moule S, Oyston PC, Quail M, Rutherford K, Simmonds M, SkeltonJ, Stevens K, Whitehead S, Barrell BG. 2001. Genome sequence of Yersiniapestis, the causative agent of plague. Nature 413:523–527. http://dx.doi.org/10.1038/35097083.
Schiano et al.
1670 jb.asm.org Journal of Bacteriology
on June 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
Related Documents