Molecular Microbiology (2000) 38(5), 955–970 Genome organization and characterization of mycobacteriophage Bxb1 Jose ´ Mediavilla, 1 Shruti Jain, 1 Jordon Kriakov, 2 Michael E. Ford, 1 Robert L. Duda, 1 William R. Jacobs Jr, 2 Roger W. Hendrix 1 and Graham F. Hatfull 1 * 1 Pittsburgh Bacteriophage Institute and Department of Biological Sciences, University of Pittsburgh, Pittsburgh, PA 15260, USA. 2 Howard Hughes Medical Institute, Department of Microbiology and Immunology, Albert Einstein College of Medicine, Bronx, New York, NY 10461, USA. Summary Mycobacteriophage Bxb1 is a temperate phage of Mycobacterium smegmatis. The morphology of Bxb1 particles is similar to that of mycobacteriophages L5 and D29, although Bxb1 differs from these phages in other respects. First, it is heteroimmune with L5 and efficiently forms plaques on an L5 lysogen. Secondly, it has a different host range and fails to infect slow- growing mycobacteria, using a receptor system that is apparently different from that of L5 and D29. Thirdly, it is the first mycobacteriophage to be described that forms a large prominent halo around plaques on a lawn of M. smegmatis. The sequence of the Bxb1 genome shows that it possesses a similar overall organization to the genomes of L5 and D29 and shares weak but detectable DNA sequence similarity to these phages within the structural genes. However, Bxb1 uses a different system of integration and excision, a repressor with different specificity to that of L5 and encodes a large number of novel gene products including several with enzymatic functions that could degrade or modify the mycobacterial cell wall. Introduction Mycobacteriophages have proved to be extremely useful tools for the development of mycobacterial genetics, as well as revealing novel aspects of viral evolution (Hendrix et al., 1999), site-specific recombination (Pen ˜a et al., 1999), gene regulation (Brown et al., 1997) and generat- ing tools for the clinical diagnosis of tuberculosis (Jacobs et al., 1993; Sarkis et al., 1995). Although over 250 mycobacteriophages have been described (Hatfull and Jacobs, 1994), only a small number have been studied in any detail; these include L5 (Hatfull and Sarkis, 1993), D29 (Ford et al., 1998a) and TM4 (Ford et al., 1998b). Mycobacteriophage L5 is a temperate phage that infects both fast- and slow-growing mycobacteria (Fullner and Hatfull, 1997). It has a 52 297 bp genome containing at least 86 protein-coding and three tRNA genes and can be divided into left and right arms (Hatfull and Sarkis, 1993); the left arm genes (between the leftmost cos site and the attachment site attP) encode the virion structure and assembly functions, whereas the right arm genes (between attP and the rightmost cos) code for DNA metabolism and regulatory activities. The right arm genes are expressed early in lytic growth and are transcribed leftwards, whereas the left arm genes are expressed late in lytic growth and are transcribed rightwards (see also Fig. 4). L5 forms stable lysogens in which the phage genome is integrated into a chromosomal attB site via integrase- mediated site-specific recombination between attP and attB (Snapper et al., 1988; Lee et al., 1991). The integrase protein (gp33) is a member of the tyrosine-recombinase family of site-specific recombinases and acts together with the mycobacterial integration host factor (mIHF) to catalyse integration (Lee et al., 1991; Pedulla et al., 1996). Prophage excision uses the same two proteins, but also requires a phage-encoded excisionase, the product of gene 36 (Lewis and Hatfull, 2000). Lysogeny of L5 is maintained through the action of the phage repressor, encoded by gene 71 (Donnelly Wu et al., 1993). L5 gp71 is a 183-residue protein that binds to DNA as a monomer via a helix–turn–helix motif near the N-terminus and regulates the early lytic promoter, P left , located at the right end of the genome, which promotes leftwards transcription for early lytic growth (Nesbit et al., 1995). However, L5 gp71 appears to play a complex role in regulating the phage life cycles. In addition to its binding site at the P left promoter, there are at least another 29 closely related sites, 23 of which have been shown to be substrates for gp71 binding in vitro (Brown et al., 1997). A comparison of the binding sites shows that they each conform closely to a 13 bp asymmetric consensus sequence (5 0 -GGTGGc/aTGTCAAG) and are located in only one orientation with respect to transcription. The binding of L5 gp71 appears to prevent the progress of the Q 2000 Blackwell Science Ltd Accepted 8 September, 2000. *For correspondence. E-mail gfh@pitt. edu; Tel. (11) 412 624 6975; Fax (11) 412 624 4870.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Microbiology (2000) 38(5), 955±970
Genome organization and characterization ofmycobacteriophage Bxb1
Jose Mediavilla,1 Shruti Jain,1 Jordon Kriakov,2
Michael E. Ford,1 Robert L. Duda,1
William R. Jacobs Jr,2 Roger W. Hendrix1
and Graham F. Hatfull1*1Pittsburgh Bacteriophage Institute and Department of
Biological Sciences, University of Pittsburgh, Pittsburgh,
PA 15260, USA.2Howard Hughes Medical Institute, Department of
Microbiology and Immunology, Albert Einstein College of
Medicine, Bronx, New York, NY 10461, USA.
Summary
Mycobacteriophage Bxb1 is a temperate phage of
Mycobacterium smegmatis. The morphology of Bxb1
particles is similar to that of mycobacteriophages L5
and D29, although Bxb1 differs from these phages in
other respects. First, it is heteroimmune with L5 and
efficiently forms plaques on an L5 lysogen. Secondly,
it has a different host range and fails to infect slow-
growing mycobacteria, using a receptor system that
is apparently different from that of L5 and D29.
Thirdly, it is the first mycobacteriophage to be
described that forms a large prominent halo around
plaques on a lawn of M. smegmatis. The sequence of
the Bxb1 genome shows that it possesses a similar
overall organization to the genomes of L5 and D29
and shares weak but detectable DNA sequence
similarity to these phages within the structural genes.
However, Bxb1 uses a different system of integration
and excision, a repressor with different specificity to
that of L5 and encodes a large number of novel gene
products including several with enzymatic functions
that could degrade or modify the mycobacterial cell
wall.
Introduction
Mycobacteriophages have proved to be extremely useful
tools for the development of mycobacterial genetics, as
well as revealing novel aspects of viral evolution (Hendrix
et al., 1999), site-specific recombination (PenÄa et al.,
1999), gene regulation (Brown et al., 1997) and generat-
ing tools for the clinical diagnosis of tuberculosis (Jacobs
et al., 1993; Sarkis et al., 1995). Although over 250
mycobacteriophages have been described (Hatfull and
Jacobs, 1994), only a small number have been studied in
any detail; these include L5 (Hatfull and Sarkis, 1993),
D29 (Ford et al., 1998a) and TM4 (Ford et al., 1998b).
Mycobacteriophage L5 is a temperate phage that
infects both fast- and slow-growing mycobacteria (Fullner
and Hatfull, 1997). It has a 52 297 bp genome containing
at least 86 protein-coding and three tRNA genes and can
be divided into left and right arms (Hatfull and Sarkis,
1993); the left arm genes (between the leftmost cos site
and the attachment site attP) encode the virion structure
and assembly functions, whereas the right arm genes
(between attP and the rightmost cos) code for DNA
metabolism and regulatory activities. The right arm genes
are expressed early in lytic growth and are transcribed
leftwards, whereas the left arm genes are expressed late
in lytic growth and are transcribed rightwards (see also
Fig. 4).
L5 forms stable lysogens in which the phage genome is
integrated into a chromosomal attB site via integrase-
mediated site-specific recombination between attP and
attB (Snapper et al., 1988; Lee et al., 1991). The integrase
protein (gp33) is a member of the tyrosine-recombinase
family of site-specific recombinases and acts together
with the mycobacterial integration host factor (mIHF) to
catalyse integration (Lee et al., 1991; Pedulla et al.,
1996). Prophage excision uses the same two proteins, but
also requires a phage-encoded excisionase, the product
of gene 36 (Lewis and Hatfull, 2000).
Lysogeny of L5 is maintained through the action of the
phage repressor, encoded by gene 71 (Donnelly Wu et al.,
1993). L5 gp71 is a 183-residue protein that binds to
DNA as a monomer via a helix±turn±helix motif near the
N-terminus and regulates the early lytic promoter, Pleft,
located at the right end of the genome, which promotes
leftwards transcription for early lytic growth (Nesbit et al.,
1995). However, L5 gp71 appears to play a complex role
in regulating the phage life cycles. In addition to its binding
site at the Pleft promoter, there are at least another 29
closely related sites, 23 of which have been shown to be
substrates for gp71 binding in vitro (Brown et al., 1997).
A comparison of the binding sites shows that they
each conform closely to a 13 bp asymmetric consensus
sequence (5 0-GGTGGc/aTGTCAAG) and are located in
only one orientation with respect to transcription. The
binding of L5 gp71 appears to prevent the progress of the
Q 2000 Blackwell Science Ltd
Accepted 8 September, 2000. *For correspondence. E-mail [email protected]; Tel. (11) 412 624 6975; Fax (11) 412 624 4870.

transcriptional apparatus through these sites, which are
thus referred to as `stoperator' sites (Brown et al., 1997).
This unusual action of the repressor represents a
mechanism for the global silencing of the L5 prophage
genome.
Mycobacteriophage D29 is not a temperate phage,
although its genome organization indicates that it is a
recent derivative of an L5-like temperate parent (Ford
et al., 1998a). The left arm genes are very similar to those
in L5 (<85% identity at the DNA level), whereas the right
arm is more distantly related to L5, with homologous
genes interspersed with unrelated segments. The non-
temperate nature of D29 is accounted for by a 3.6 kbp
deletion in D29 that removes most of the putative
repressor gene. Nevertheless, D29, which shares immu-
nity with L5, has essentially the same distribution of
repressor binding sites that conform to the same
consensus sequences and also has an active integration
system (Ribeiro et al., 1997; Ford et al., 1998a; PenÄa
et al., 1998).
Mycobacteriophage TM4 is a much more distant
relative of L5 and D29 (Ford et al., 1998b). The genome
organization is different, and all the genes are transcribed
in the rightwards direction. There is no significant level of
sequence similarity at the nucleic acid level, but several of
the gene products are related to those encoded by L5
and/or D29; this is particularly true of the structure and
assembly genes (1±26). However, there is no evidence of
a repressor gene, multiple binding sites or an integration
system, indicating that TM4 is not a temperate phage nor
a recent derivative of one.
The genomic analyses of these mycobacteriophages
have provided useful insights into the mechanisms of viral
evolution. In particular, it was noted that the overall
genome organization of L5 (and D29) is reminiscent of
that of phage lambda, even though there is no clear
sequence similarity at either the nucleic acid or the protein
level (Hatfull and Sarkis, 1993; Hatfull and Jacobs, 1994;
Ford et al., 1998a). However, as more phage genomes
have been sequenced, it appears as though all the tailed
bacteriophages have probably shared common ancestry
and have evolved through extensive horizontal transfer of
DNA (Hendrix et al., 1999). Phages that form closely
related groups (such as the lambda-like phages or L5 and
D29) often have a mosaic nature, with similar (and
obviously homologous) segments interspersed with unre-
lated regions, the junctions usually occurring at gene
boundaries. Comparison of the lambda-like phages also
reveals the presence of `morons', small segments of DNA
typically containing an open reading frame (ORF), a
promoter and a transcription terminator, which appear to
have been acquired in relatively recent evolutionary time
(Juhala et al., 2000). The moron genes are probably
expressed from the prophage state, but do not interfere
with lytic growth because of the presence of antiter-
minator systems that enable transcription through the
terminators. Morons have not yet been identified in the
mycobacteriophage genomes, and their acquisition may
be thwarted by the absence of such antiterminator
systems.
Mycobacteriophage Bxb1 is a recently discovered
phage isolated in the Bronx, New York, USA. It infects
Mycobacterium smegmatis but does not form plaques on
any of the slow-growing strains. As we describe here, it
has some unusual features including the ability to form
large halos around infected cells on a lawn of M.
smegmatis. We also describe the genome organization
of Bxb1 which shows it to be clearly related to L5 and D29,
with some segments having obvious DNA sequence
similarity. However, Bxb1 exhibits a number of interesting
departures from L5 and D29 at the genomic level, uses a
different integration system and enters its host cells via a
different cellular receptor.
Results
Halo formation by Bxb1
When Bxb1 was first isolated, it was noted that it forms
plaques with large halos, hence its name, `the Bronx
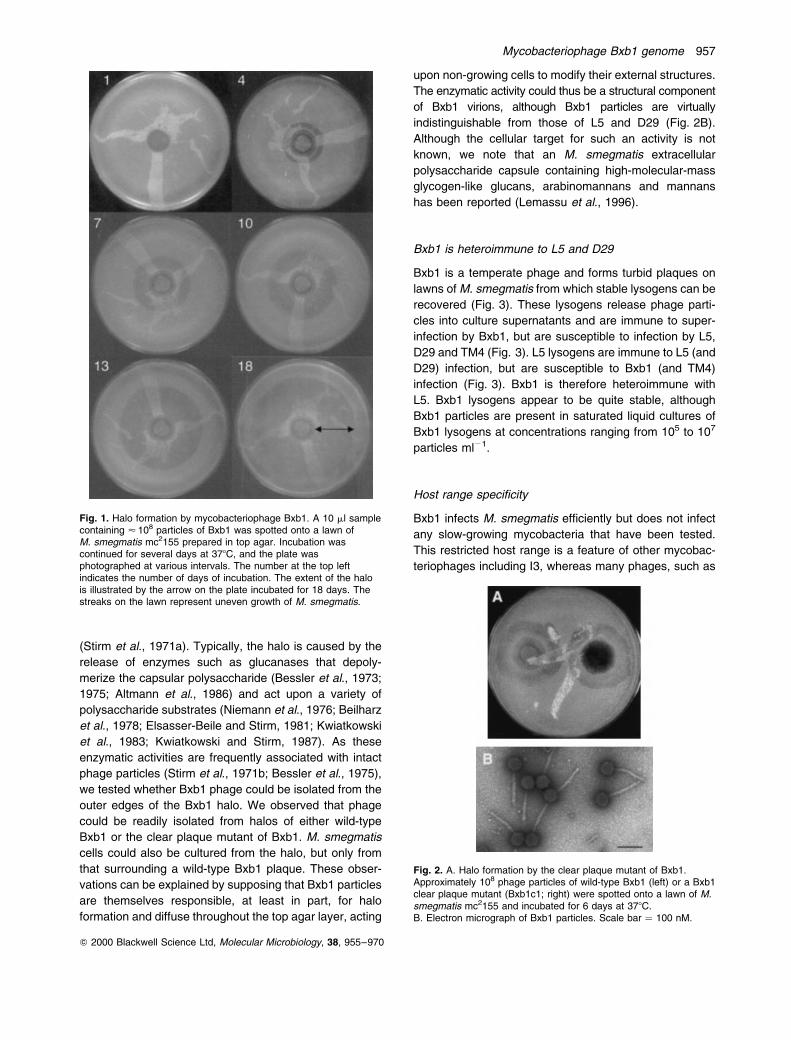
bomber' or Bxb1, as shown in Fig. 1. Formation of the
halo begins after 3 days of incubation, once the infected
area is fully formed and the M. smegmatis cells represent
a thick lawn covering the agar layer. Upon further
incubation, the halo grows in size and spreads out across
the M. smegmatis lawn until, after 18 days, it encom-
passes a large part of the entire lawn (Fig. 1). Presum-
ably, a diffusible agent is generated from the infected
area, spreads outwards through the top agar layer and
either arrests or modifies the bacterial cells. However, the
halo continues to spread after a confluent lawn of cells
has grown and, therefore, this putative agent presumably
acts upon cells that are no longer actively growing.
As Bxb1 is a temperate phage, this putative diffusible
agent could be either a product of lytic growth or secreted
by the growing Bxb1 lysogens within the plaque. To test
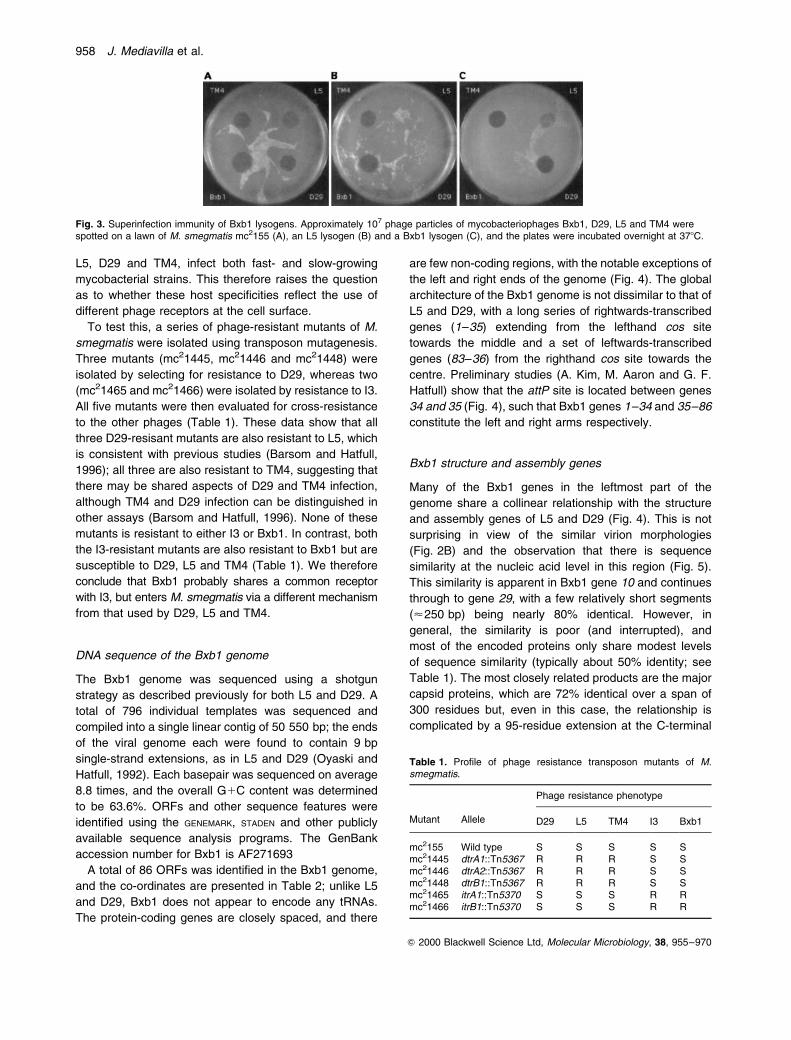
this, a clear plaque mutant of Bxb1, which is unable to
form stable lysogens, was isolated and tested for halo
formation. This clear plaque mutant forms a halo that is
indistinguishable from that of wild-type Bxb1, indicating
that the putative diffusible agent is generated by lytic
infection of Bxb1 and not by growth of Bxb1 lysogens
(Fig. 2A).
Although formation of halos around phage plaques has
not been described for other mycobacteriophages, it has
been observed for a number of other phages, particularly
those that infect encapsulated and/or mucoid Enterobac-
teriaceae such as Escherichia coli and Klebsiella spp.
956 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
(Stirm et al., 1971a). Typically, the halo is caused by the
release of enzymes such as glucanases that depoly-
merize the capsular polysaccharide (Bessler et al., 1973;
1975; Altmann et al., 1986) and act upon a variety of
polysaccharide substrates (Niemann et al., 1976; Beilharz
et al., 1978; Elsasser-Beile and Stirm, 1981; Kwiatkowski
et al., 1983; Kwiatkowski and Stirm, 1987). As these
enzymatic activities are frequently associated with intact
phage particles (Stirm et al., 1971b; Bessler et al., 1975),
we tested whether Bxb1 phage could be isolated from the
outer edges of the Bxb1 halo. We observed that phage
could be readily isolated from halos of either wild-type
Bxb1 or the clear plaque mutant of Bxb1. M. smegmatis
cells could also be cultured from the halo, but only from
that surrounding a wild-type Bxb1 plaque. These obser-
vations can be explained by supposing that Bxb1 particles
are themselves responsible, at least in part, for halo
formation and diffuse throughout the top agar layer, acting
upon non-growing cells to modify their external structures.
The enzymatic activity could thus be a structural component
of Bxb1 virions, although Bxb1 particles are virtually
indistinguishable from those of L5 and D29 (Fig. 2B).
Although the cellular target for such an activity is not
known, we note that an M. smegmatis extracellular
polysaccharide capsule containing high-molecular-mass
glycogen-like glucans, arabinomannans and mannans
has been reported (Lemassu et al., 1996).
Bxb1 is heteroimmune to L5 and D29
Bxb1 is a temperate phage and forms turbid plaques on
lawns of M. smegmatis from which stable lysogens can be
recovered (Fig. 3). These lysogens release phage parti-
cles into culture supernatants and are immune to super-
infection by Bxb1, but are susceptible to infection by L5,
D29 and TM4 (Fig. 3). L5 lysogens are immune to L5 (and
D29) infection, but are susceptible to Bxb1 (and TM4)
infection (Fig. 3). Bxb1 is therefore heteroimmune with
L5. Bxb1 lysogens appear to be quite stable, although
Bxb1 particles are present in saturated liquid cultures of
Bxb1 lysogens at concentrations ranging from 105 to 107
particles ml21.
Host range specificity
Bxb1 infects M. smegmatis efficiently but does not infect
any slow-growing mycobacteria that have been tested.
This restricted host range is a feature of other mycobac-
teriophages including I3, whereas many phages, such as
Fig. 2. A. Halo formation by the clear plaque mutant of Bxb1.Approximately 108 phage particles of wild-type Bxb1 (left) or a Bxb1clear plaque mutant (Bxb1c1; right) were spotted onto a lawn of M.smegmatis mc2155 and incubated for 6 days at 378C.B. Electron micrograph of Bxb1 particles. Scale bar � 100 nM.
Fig. 1. Halo formation by mycobacteriophage Bxb1. A 10 ml samplecontaining < 108 particles of Bxb1 was spotted onto a lawn ofM. smegmatis mc2155 prepared in top agar. Incubation wascontinued for several days at 378C, and the plate wasphotographed at various intervals. The number at the top leftindicates the number of days of incubation. The extent of the halois illustrated by the arrow on the plate incubated for 18 days. Thestreaks on the lawn represent uneven growth of M. smegmatis.
Mycobacteriophage Bxb1 genome 957
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
L5, D29 and TM4, infect both fast- and slow-growing
mycobacterial strains. This therefore raises the question
as to whether these host specificities reflect the use of
different phage receptors at the cell surface.
To test this, a series of phage-resistant mutants of M.
smegmatis were isolated using transposon mutagenesis.
Three mutants (mc21445, mc21446 and mc21448) were
isolated by selecting for resistance to D29, whereas two
(mc21465 and mc21466) were isolated by resistance to I3.
All five mutants were then evaluated for cross-resistance
to the other phages (Table 1). These data show that all
three D29-resisant mutants are also resistant to L5, which
is consistent with previous studies (Barsom and Hatfull,
1996); all three are also resistant to TM4, suggesting that
there may be shared aspects of D29 and TM4 infection,
although TM4 and D29 infection can be distinguished in
other assays (Barsom and Hatfull, 1996). None of these
mutants is resistant to either I3 or Bxb1. In contrast, both
the I3-resistant mutants are also resistant to Bxb1 but are
susceptible to D29, L5 and TM4 (Table 1). We therefore
conclude that Bxb1 probably shares a common receptor
with I3, but enters M. smegmatis via a different mechanism
from that used by D29, L5 and TM4.
DNA sequence of the Bxb1 genome
The Bxb1 genome was sequenced using a shotgun
strategy as described previously for both L5 and D29. A
total of 796 individual templates was sequenced and
compiled into a single linear contig of 50 550 bp; the ends
of the viral genome each were found to contain 9 bp
single-strand extensions, as in L5 and D29 (Oyaski and
Hatfull, 1992). Each basepair was sequenced on average
8.8 times, and the overall G1C content was determined
to be 63.6%. ORFs and other sequence features were
identified using the GENEMARK, STADEN and other publicly
available sequence analysis programs. The GenBank
accession number for Bxb1 is AF271693
A total of 86 ORFs was identified in the Bxb1 genome,
and the co-ordinates are presented in Table 2; unlike L5
and D29, Bxb1 does not appear to encode any tRNAs.
The protein-coding genes are closely spaced, and there
are few non-coding regions, with the notable exceptions of
the left and right ends of the genome (Fig. 4). The global
architecture of the Bxb1 genome is not dissimilar to that of
L5 and D29, with a long series of rightwards-transcribed
genes (1±35) extending from the lefthand cos site
towards the middle and a set of leftwards-transcribed
genes (83±36) from the righthand cos site towards the
centre. Preliminary studies (A. Kim, M. Aaron and G. F.
Hatfull) show that the attP site is located between genes
34 and 35 (Fig. 4), such that Bxb1 genes 1±34 and 35±86
constitute the left and right arms respectively.
Bxb1 structure and assembly genes
Many of the Bxb1 genes in the leftmost part of the
genome share a collinear relationship with the structure
and assembly genes of L5 and D29 (Fig. 4). This is not
surprising in view of the similar virion morphologies
(Fig. 2B) and the observation that there is sequence
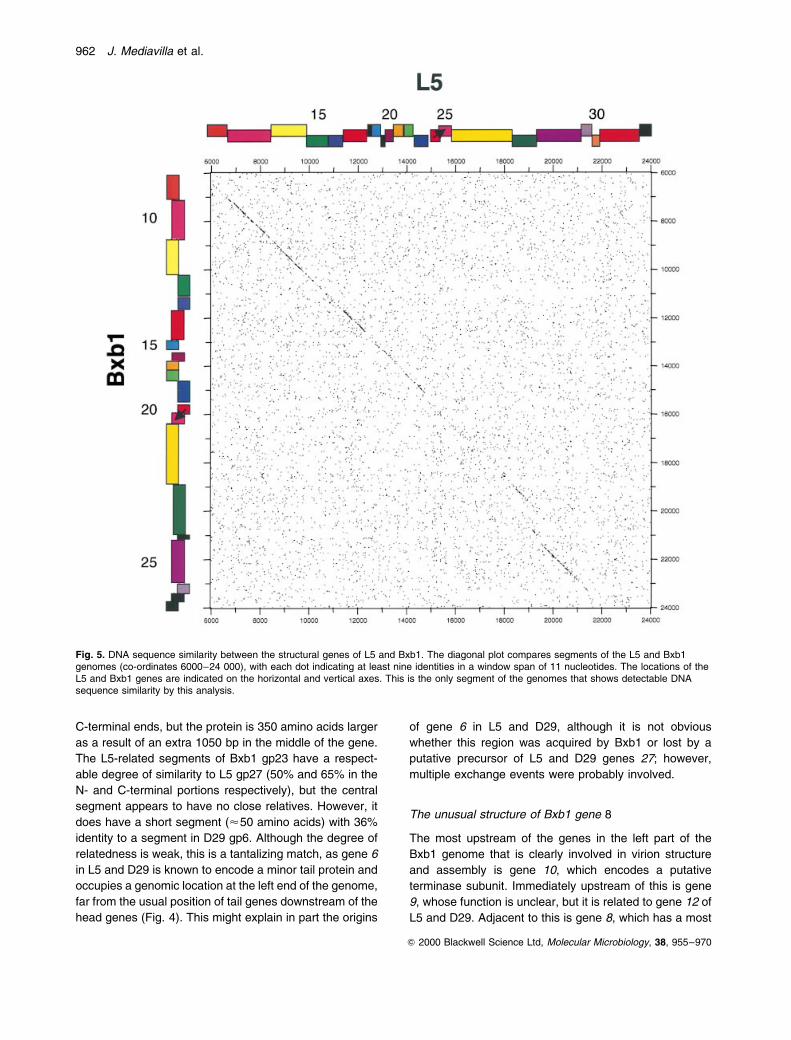
similarity at the nucleic acid level in this region (Fig. 5).
This similarity is apparent in Bxb1 gene 10 and continues
through to gene 29, with a few relatively short segments
(<250 bp) being nearly 80% identical. However, in
general, the similarity is poor (and interrupted), and
most of the encoded proteins only share modest levels
of sequence similarity (typically about 50% identity; see
Table 1). The most closely related products are the major
capsid proteins, which are 72% identical over a span of
300 residues but, even in this case, the relationship is
complicated by a 95-residue extension at the C-terminal
Table 1. Profile of phage resistance transposon mutants of M.smegmatis.
Mutant Allele
Phage resistance phenotype
D29 L5 TM4 I3 Bxb1
mc2155 Wild type S S S S Smc21445 dtrA1::Tn5367 R R R S Smc21446 dtrA2::Tn5367 R R R S Smc21448 dtrB1::Tn5367 R R R S Smc21465 itrA1::Tn5370 S S S R Rmc21466 itrB1::Tn5370 S S S R R
Fig. 3. Superinfection immunity of Bxb1 lysogens. Approximately 107 phage particles of mycobacteriophages Bxb1, D29, L5 and TM4 werespotted on a lawn of M. smegmatis mc2155 (A), an L5 lysogen (B) and a Bxb1 lysogen (C), and the plates were incubated overnight at 378C.
958 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
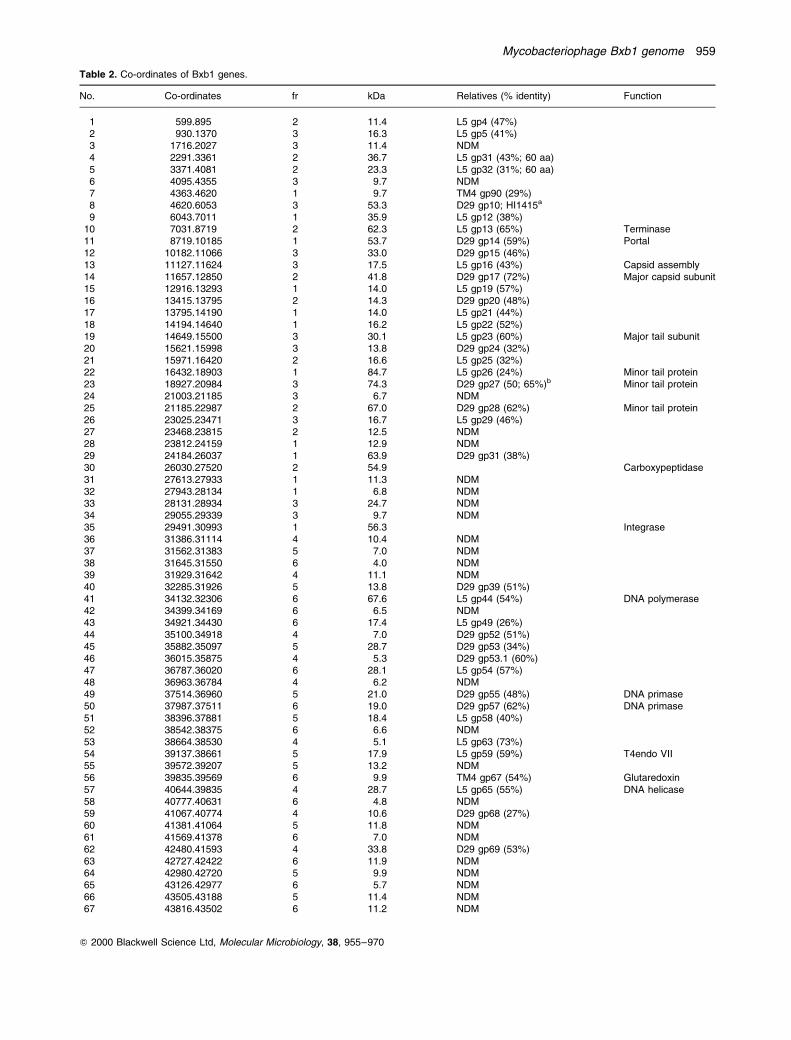
Table 2. Co-ordinates of Bxb1 genes.
No. Co-ordinates fr kDa Relatives (% identity) Function
1 599.895 2 11.4 L5 gp4 (47%)2 930.1370 3 16.3 L5 gp5 (41%)3 1716.2027 3 11.4 NDM4 2291.3361 2 36.7 L5 gp31 (43%; 60 aa)5 3371.4081 2 23.3 L5 gp32 (31%; 60 aa)6 4095.4355 3 9.7 NDM7 4363.4620 1 9.7 TM4 gp90 (29%)8 4620.6053 3 53.3 D29 gp10; HI1415a
9 6043.7011 1 35.9 L5 gp12 (38%)10 7031.8719 2 62.3 L5 gp13 (65%) Terminase11 8719.10185 1 53.7 D29 gp14 (59%) Portal12 10182.11066 3 33.0 D29 gp15 (46%)13 11127.11624 3 17.5 L5 gp16 (43%) Capsid assembly14 11657.12850 2 41.8 D29 gp17 (72%) Major capsid subunit15 12916.13293 1 14.0 L5 gp19 (57%)16 13415.13795 2 14.3 D29 gp20 (48%)17 13795.14190 1 14.0 L5 gp21 (44%)18 14194.14640 1 16.2 L5 gp22 (52%)19 14649.15500 3 30.1 L5 gp23 (60%) Major tail subunit20 15621.15998 3 13.8 D29 gp24 (32%)21 15971.16420 2 16.6 L5 gp25 (32%)22 16432.18903 1 84.7 L5 gp26 (24%) Minor tail protein23 18927.20984 3 74.3 D29 gp27 (50; 65%)b Minor tail protein24 21003.21185 3 6.7 NDM25 21185.22987 2 67.0 D29 gp28 (62%) Minor tail protein26 23025.23471 3 16.7 L5 gp29 (46%)27 23468.23815 2 12.5 NDM28 23812.24159 1 12.9 NDM29 24184.26037 1 63.9 D29 gp31 (38%)30 26030.27520 2 54.9 Carboxypeptidase31 27613.27933 1 11.3 NDM32 27943.28134 1 6.8 NDM33 28131.28934 3 24.7 NDM34 29055.29339 3 9.7 NDM35 29491.30993 1 56.3 Integrase36 31386.31114 4 10.4 NDM37 31562.31383 5 7.0 NDM38 31645.31550 6 4.0 NDM39 31929.31642 4 11.1 NDM40 32285.31926 5 13.8 D29 gp39 (51%)41 34132.32306 6 67.6 L5 gp44 (54%) DNA polymerase42 34399.34169 6 6.5 NDM43 34921.34430 6 17.4 L5 gp49 (26%)44 35100.34918 4 7.0 D29 gp52 (51%)45 35882.35097 5 28.7 D29 gp53 (34%)46 36015.35875 4 5.3 D29 gp53.1 (60%)47 36787.36020 6 28.1 L5 gp54 (57%)48 36963.36784 4 6.2 NDM49 37514.36960 5 21.0 D29 gp55 (48%) DNA primase50 37987.37511 6 19.0 D29 gp57 (62%) DNA primase51 38396.37881 5 18.4 L5 gp58 (40%)52 38542.38375 6 6.6 NDM53 38664.38530 4 5.1 L5 gp63 (73%)54 39137.38661 5 17.9 L5 gp59 (59%) T4endo VII55 39572.39207 5 13.2 NDM56 39835.39569 6 9.9 TM4 gp67 (54%) Glutaredoxin57 40644.39835 4 28.7 L5 gp65 (55%) DNA helicase58 40777.40631 6 4.8 NDM59 41067.40774 4 10.6 D29 gp68 (27%)60 41381.41064 5 11.8 NDM61 41569.41378 6 7.0 NDM62 42480.41593 4 33.8 D29 gp69 (53%)63 42727.42422 6 11.9 NDM64 42980.42720 5 9.9 NDM65 43126.42977 6 5.7 NDM66 43505.43188 5 11.4 NDM67 43816.43502 6 11.2 NDM
Mycobacteriophage Bxb1 genome 959
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

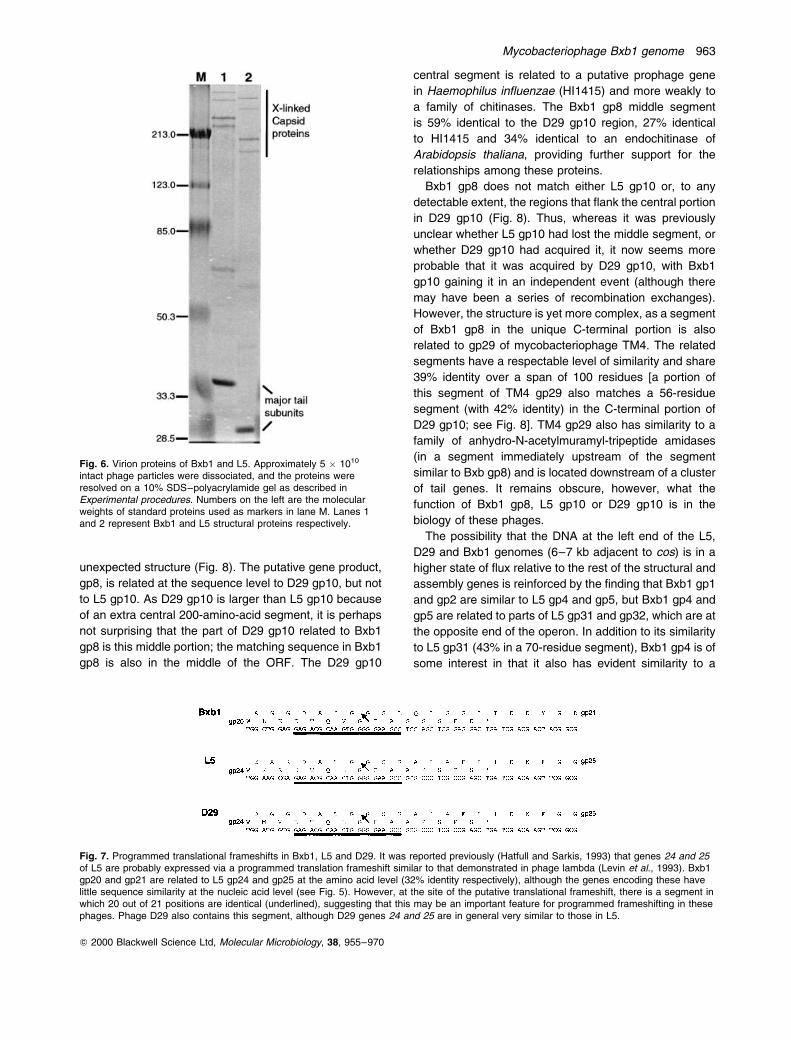
end of Bxb1 gp14. The major capsid protein does not
appear to be processed, however, as the forms of the
capsid subunits isolated from virions are clearly larger
than those from L5 (Fig. 6). Like L5 gp17, Bxb1 gp14
does appear to be extensively covalently cross-linked
in the mature phage particles, and there is little or no
free gp14 present (Fig. 6). Both L5 and Bxb1 contain a
sequence element downstream of their capsid subunit
genes that has the potential to form a stem±loop structure
at the RNA level, although their roles in the regulation of
gene expression is not known.
Bxb1 gene 19 encodes the putative major tail subunit,
which shares 60% amino acid identity with L5 gp23, the
major tail subunit (Fig. 4). However, like the major head
subunit, there is a long C-terminal extension (87 amino
acids) for which there is no counterpart in L5 or D29 gp17.
Again, the protein does not appear to be processed, as
the major tail subunit isolated from Bxb1 virions is
substantially larger than that from L5 particles, although
both migrate rather more slowly than their predicted
mobilities in SDS±PAGE (Fig. 6), a phenomenon that is
also observed with the major tail subunit of Listeria phage
A118 (Loessner et al., 2000). In contrast to these
relationships, the minor tail proteins that determine the
length of the tail (L5 gp26 and Bxb1 gp22) share only 24%
amino acid identity, although the sizes of the proteins are
very similar (823 amino acids and 837 amino acids in
Bxb1 and L5 respectively), which is reflected in the similar
lengths of the phage tails (Fig. 2B).
In phage lambda, it has been shown that the two ORFs
(G and T) located between the major tail subunit gene (V)
and the tail length determinant gene (H) are expressed via
a programmed translational frameshift to generate the
products gpG and gpG-T (Levin et al., 1993). This unusual
feature is an important component of the structure and
assembly machinery, as it is highly conserved among
phages with related organizations of structural genes, even
in the absence of any sequence relatedness (J. Xu and R.
Hendrix, unpublished observations); in L5, the ORFs 24
and 25 are analogues of lambda G and T (Hatfull and
Sarkis, 1993) and are also expressed via a translational
frameshift (J. Xu and R. Hendrix, unpublished observa-
tions). Bxb1 gp20 and gp21 are related at the sequence
level to L5 gp24 and gp25, albeit weakly (32% identity),
and are thus also likely to be translated via a programmed
frameshift. Although there is little detectable DNA sequence
similarity between these genes (Fig. 5), there is a segment
in which 20 out of 21 bases are identical (Fig. 7).
Interestingly, this region corresponds to the precise point
of the predicted translational frameshift and thus most
probably reflects the conservation of important structural
features that are required for translational frameshifting
(Fig. 7).
There are several notable differences between the
Bxb1 and L5 structural gene clusters (Fig. 4). First, there
are several small genes that are predicted to be present in
Bxb1 but absent from L5 (24, 27 and 28), and some genes
that are present in L5 and D29 but absent from Bxb1 (18,
19.1 and 30). Although the functions of these are not
clear, their general proximity to other tail genes suggests
that at least some of these may also be tail associated. A
dramatic departure is seen in Bxb1 gene 23, which is
related to L5 gene 27, a known component of the L5
tail. Bxb1 gp23 is related to L5 gp27 at both its N- and
Table 2. continued
No. Co-ordinates fr kDa Relatives (% identity) Function
68 44208.43813 4 15.3 D29 gp70 (34%)69 44762.44250 5 20.0 L5 gp71 (45%) Repressor70 45323.45024 5 11.0 L5 gp76 (47%)71 45483.45316 4 6.6 NDM72 45749.45480 5 9.9 NDM73 45949.45746 6 8.0 NDM74 46134.45946 4 6.6 NDM75 46301.46134 5 5.7 NDM76 46426.46298 6 4.5 NDM77 46596.46447 4 5.3 NDM78 47003.46641 5 13.2 NDM79 47202.47029 4 6.6 NDM80 47598.47359 4 9.1 NDM81 47928.47773 4 5.4 NDM82 48504.48205 4 10.9 NDM83 48689.48504 5 6.8 NDM84 48903.49112 3 7.2 L5 gp79 (76%)85 49109.49267 2 5.6 NDM86 49573.49761 1 6.8 NDM
aa, amino acids.a. See Fig. 5.b. Residues 9±265, 50%; 514±685, 65%.
960 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
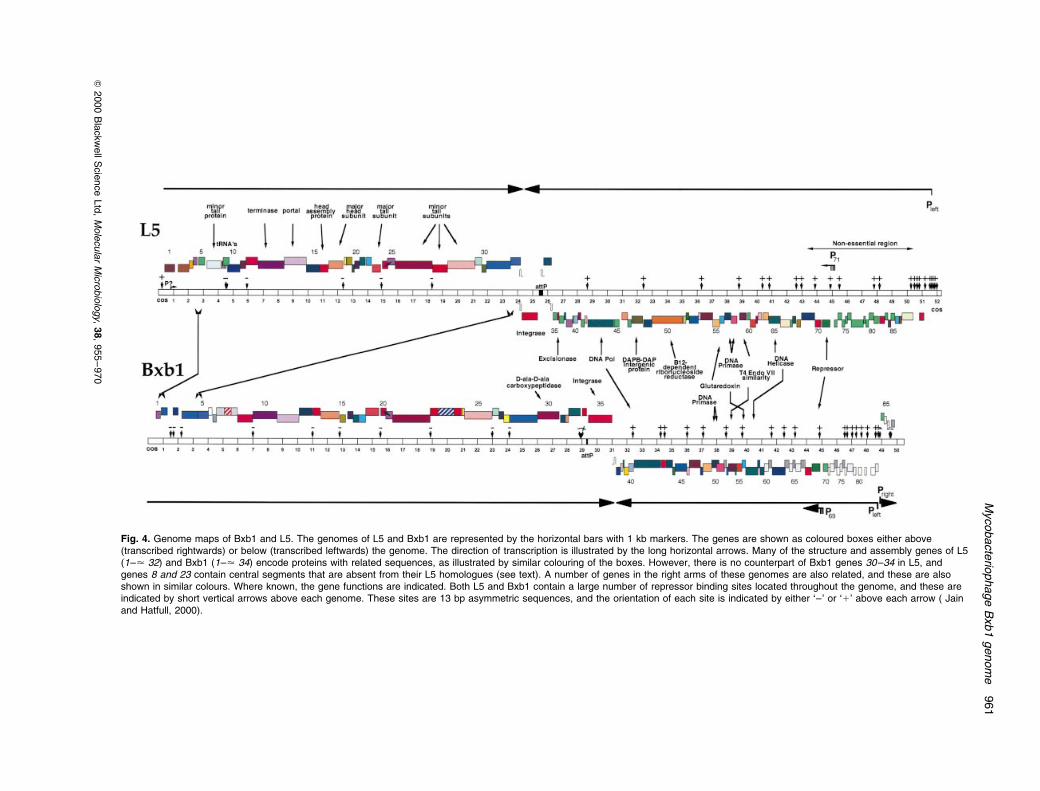
Fig. 4. Genome maps of Bxb1 and L5. The genomes of L5 and Bxb1 are represented by the horizontal bars with 1 kb markers. The genes are shown as coloured boxes either above(transcribed rightwards) or below (transcribed leftwards) the genome. The direction of transcription is illustrated by the long horizontal arrows. Many of the structure and assembly genes of L5(1±< 32) and Bxb1 (1±< 34) encode proteins with related sequences, as illustrated by similar colouring of the boxes. However, there is no counterpart of Bxb1 genes 30±34 in L5, andgenes 8 and 23 contain central segments that are absent from their L5 homologues (see text). A number of genes in the right arms of these genomes are also related, and these are alsoshown in similar colours. Where known, the gene functions are indicated. Both L5 and Bxb1 contain a large number of repressor binding sites located throughout the genome, and these areindicated by short vertical arrows above each genome. These sites are 13 bp asymmetric sequences, and the orientation of each site is indicated by either `±' or `1' above each arrow ( Jainand Hatfull, 2000).
Mycobacte
riophage
Bxb1
genom
e961
Q2000
Bla
ckw
ell
Scie
nce
Ltd
,M
ole
cu
lar
Mic
robio
logy,
38,
955
±970
C-terminal ends, but the protein is 350 amino acids larger
as a result of an extra 1050 bp in the middle of the gene.
The L5-related segments of Bxb1 gp23 have a respect-
able degree of similarity to L5 gp27 (50% and 65% in the
N- and C-terminal portions respectively), but the central
segment appears to have no close relatives. However, it
does have a short segment (<50 amino acids) with 36%
identity to a segment in D29 gp6. Although the degree of
relatedness is weak, this is a tantalizing match, as gene 6
in L5 and D29 is known to encode a minor tail protein and
occupies a genomic location at the left end of the genome,
far from the usual position of tail genes downstream of the
head genes (Fig. 4). This might explain in part the origins
of gene 6 in L5 and D29, although it is not obvious
whether this region was acquired by Bxb1 or lost by a
putative precursor of L5 and D29 genes 27; however,
multiple exchange events were probably involved.
The unusual structure of Bxb1 gene 8
The most upstream of the genes in the left part of the
Bxb1 genome that is clearly involved in virion structure
and assembly is gene 10, which encodes a putative
terminase subunit. Immediately upstream of this is gene
9, whose function is unclear, but it is related to gene 12 of
L5 and D29. Adjacent to this is gene 8, which has a most
Fig. 5. DNA sequence similarity between the structural genes of L5 and Bxb1. The diagonal plot compares segments of the L5 and Bxb1genomes (co-ordinates 6000±24 000), with each dot indicating at least nine identities in a window span of 11 nucleotides. The locations of theL5 and Bxb1 genes are indicated on the horizontal and vertical axes. This is the only segment of the genomes that shows detectable DNAsequence similarity by this analysis.
962 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
unexpected structure (Fig. 8). The putative gene product,
gp8, is related at the sequence level to D29 gp10, but not
to L5 gp10. As D29 gp10 is larger than L5 gp10 because
of an extra central 200-amino-acid segment, it is perhaps
not surprising that the part of D29 gp10 related to Bxb1
gp8 is this middle portion; the matching sequence in Bxb1
gp8 is also in the middle of the ORF. The D29 gp10
central segment is related to a putative prophage gene
in Haemophilus influenzae (HI1415) and more weakly to
a family of chitinases. The Bxb1 gp8 middle segment
is 59% identical to the D29 gp10 region, 27% identical
to HI1415 and 34% identical to an endochitinase of
Arabidopsis thaliana, providing further support for the
relationships among these proteins.
Bxb1 gp8 does not match either L5 gp10 or, to any
detectable extent, the regions that flank the central portion
in D29 gp10 (Fig. 8). Thus, whereas it was previously
unclear whether L5 gp10 had lost the middle segment, or
whether D29 gp10 had acquired it, it now seems more
probable that it was acquired by D29 gp10, with Bxb1
gp10 gaining it in an independent event (although there
may have been a series of recombination exchanges).
However, the structure is yet more complex, as a segment
of Bxb1 gp8 in the unique C-terminal portion is also
related to gp29 of mycobacteriophage TM4. The related
segments have a respectable level of similarity and share
39% identity over a span of 100 residues [a portion of
this segment of TM4 gp29 also matches a 56-residue
segment (with 42% identity) in the C-terminal portion of
D29 gp10; see Fig. 8]. TM4 gp29 also has similarity to a
family of anhydro-N-acetylmuramyl-tripeptide amidases
(in a segment immediately upstream of the segment
similar to Bxb gp8) and is located downstream of a cluster
of tail genes. It remains obscure, however, what the
function of Bxb1 gp8, L5 gp10 or D29 gp10 is in the
biology of these phages.
The possibility that the DNA at the left end of the L5,
D29 and Bxb1 genomes (6±7 kb adjacent to cos) is in a
higher state of flux relative to the rest of the structural and
assembly genes is reinforced by the finding that Bxb1 gp1
and gp2 are similar to L5 gp4 and gp5, but Bxb1 gp4 and
gp5 are related to parts of L5 gp31 and gp32, which are at
the opposite end of the operon. In addition to its similarity
to L5 gp31 (43% in a 70-residue segment), Bxb1 gp4 is of
some interest in that it also has evident similarity to a
Fig. 6. Virion proteins of Bxb1 and L5. Approximately 5 � 1010
intact phage particles were dissociated, and the proteins wereresolved on a 10% SDS±polyacrylamide gel as described inExperimental procedures. Numbers on the left are the molecularweights of standard proteins used as markers in lane M. Lanes 1and 2 represent Bxb1 and L5 structural proteins respectively.
Fig. 7. Programmed translational frameshifts in Bxb1, L5 and D29. It was reported previously (Hatfull and Sarkis, 1993) that genes 24 and 25of L5 are probably expressed via a programmed translation frameshift similar to that demonstrated in phage lambda (Levin et al., 1993). Bxb1gp20 and gp21 are related to L5 gp24 and gp25 at the amino acid level (32% identity respectively), although the genes encoding these havelittle sequence similarity at the nucleic acid level (see Fig. 5). However, at the site of the putative translational frameshift, there is a segment inwhich 20 out of 21 positions are identical (underlined), suggesting that this may be an important feature for programmed frameshifting in thesephages. Phage D29 also contains this segment, although D29 genes 24 and 25 are in general very similar to those in L5.
Mycobacteriophage Bxb1 genome 963
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

family of phage tail fibre proteins including the tail fibre
protein of phage P1 (27% identity in a 160-residue span).
As this family also includes proteins associated with
recombinase-mediated DNA inversion (e.g. the Bcv and
Bcv 0 proteins of Shigella boydii), it raises the intriguing
possibility that site-specific recombination events could
account for the non-cognate organization of genes seen
at the left ends of these mycobacterial genomes.
Integration and excision
The last of the rightwards-transcribed genes (35) appears
to encode the phage integrase (Fig. 4). However, gp35
is clearly not a member of the tyrosine site-specific
recombinases, which includes the lambda and L5 inte-
grases, but is related to the serine recombinases, a
family that includes the transposon resolvases and DNA
invertases. However, several phages (fFC1, TP901-1,
TP21, R4, f105, A118 and fC31) and genomic (pre-
sumably prophage) genes encode serine-recombinase
integrases. The closest relative to Bxb1 gp35 is a
recombinase identified in the Streptomyces genome
sequencing project that shares 29% identity over 416
residues; the closest from a mycobacterial source is
Rv1586c, encoded by the Mycobacterium tuberculosis
phage-like element fRv1; (Cole et al., 1998; Hendrix et al.,
1999), which has 27% identity over 324 residues.
Although rather little is known about this class of phage
integrases, they all appear to have an N-terminal domain
(<140 residues) that is similar to the catalytic domain of
the resolvases, coupled to a large C-terminal segment
that is presumably involved in DNA binding or protein±
protein interactions. It is also noteworthy that the Bxb1
integrase gene is positioned at the end of the structural
genes and transcribed rightwards, in contrast to the L5
integrase, which is transcribed leftwards and is at the end
of the early genes (Fig. 4). The attP site is located
immediately upstream of the integrase gene (35) in a
similar position (relative to int) to that in phage fC31
(Thorpe and Smith, 1998).
Immunity and repressor functions
As Bxb1 is a temperate phage, it presumably encodes
a repressor protein, but one that must be sufficiently
dissimilar to that of L5 (gp71) so that they are hetero-
immune (see Fig. 3). Gene 69 is an excellent candidate
for being the repressor, as it is in a collinear position to L5
gene 71 and shares 40% identity with gp71. Bxb1 gp69
has been overexpressed and characterized further and is
Fig. 8. Organization of Bxb1 gp8 and related proteins. Bxb1 gp8 has little or no obvious sequence similarity to L5 gp10. Although D29 gp10 issimilar to L5 gp10 (related segments shown in blue), it contains a central segment that is similar to a central segment in Bxb1 gp8 (59%identity; shown in green). This central portion is also related to HI1415 (27% identity) as well as to a family of chitinases including anendochitinase from Arabidopsis thaliana (34% identity). To the right of the central portion in Bxb1 gp8 is another segment (shown in purple)that is related to a 105-residue segment in mycobacteriophage TM4 gp29 (39% identity), which in turn has weak similarity to the downstreamsegment in D29 gp10 (40% identity in an < 50-residue segment).
964 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

described in detail in the accompanying paper (Jain and
Hatfull, 2000). DNA binding studies with Bxb1 gp69 show
that it recognizes a 13 bp asymmetric sequence that is
similar to, yet distinct from, that in L5. There are at least
34 occurrences of this or closely related sequences in the
Bxb1 genome, and these have similar attributes to the L5
gp71 binding sites, in that they are located throughout the
genome, are predominantly in small intergenic regions
and occupy only one orientation relative to the direction of
transcription (Fig. 4). Presumably, the Bxb1 binding sites
play a similar role in global silencing of the Bxb1 prophage
genome (Brown et al., 1997). The L5 and Bxb1 repressors
have substantially greater affinities for their cognate
binding sites, which thus provides a molecular basis for
their heteroimmunity (Jain and Hatfull, 2000). The specific
locations of the binding sites and their context also
provides insights into the evolutionary development of this
unusual system of immunity.
Other Bxb1 genes of interest
Many, but not all, of the leftwards-transcribed genes are
related to early lytic L5-encoded proteins. For example,
gene 41 encodes a DNA polymerase, gene 54 encodes a
T4 endoVII-like protein that is 59% identical to L5 gp59,
gene 56 encodes a glutaredoxin that is 54% identical to
TM4 gp67 and 36% identical to L5 gp56, gene 57 encodes
a DNA helicase, and genes 50 and 51 encode DNA
primase-related proteins. Bxb1 gp50 and gp51 are related
to gp57 and gp58, respectively, of L5 and D29, although
the relationship is rather complex. In particular, it appears
as though Bxb1 genes 50 and 51 overlap extensively, with
the start codon of gene 50 residing either 35 codons
(assuming the same start as that predicted for L5 and D29
gene 57) or 52 codons (using an alternative start codon
that is also present in L5 and D29) within the coding
region for gene 51 (Fig. 4). One simple interpretation of
the organizations of these genes is that functional DNA
primase proteins (L5 gp58-57, D29 gp58-57 and Bxb1
gp51-50) are all expressed via programmed translational
frameshifts. This possibility is further supported by the
observation that both Bxb1 gp50 and Bxb1 gp51 contain
segments that are related to the single polypeptide DNA
primase encoded by Pseudomonas putida.
Between the most downstream of the L5-related
structural genes (29) and the Bxb1 integrase (35) are
five ORFs (30±34). Four of these (31±34) are of unknown
function and are not related to known proteins. However,
gene 30 encodes a 525-residue protein that matches
several proteins with known enzymatic activities. The
most striking match is a segment of Bxb1 gp35 from
residues 100 to 430 that is similar to a family of D-ala-D-ala
carboxypeptidases (the degree of relatedness is low, with
the best match to the Streptomyces fradiae enzyme to
which it has 25% identity over a span of 315 residues, with
a BLAST E-value of e213). However, the N-terminal 100
amino acids appears, at least in part, to be a fibronectin
type III (Fn3) domain, with good similarities to related
domains found in a variety of bacterial chitinases,
cellulases, amylases and poly(3-hydroxybutyrate) depo-
lymerases (Little et al., 1994). Bxb1 gp30 is the first
enzyme to our knowledge that has an Fn3 domain fused
to a D-ala-D-ala carboxypeptidase domain. The observation
that the gene is probably expressed in lytic growth (see
below) and is unique to Bxb1 (and not present in L5, D29
or TM4) would be consistent with the product playing a
role in alteration of the cell surface of M. smegmatis and
production of the halo that surrounds Bxb1 plaques.
Moreover, it lies within a location that is not inconsistent
with it encoding a structural component of the phage tails.
Bxb1 gp30 is also a candidate for involvement in cell lysis.
Genes 84±86
In mycobacteriophages L5 and D29, all the genes in the
right arm are transcribed leftwards (see Fig. 4). In Bxb1,
although this is also true for most of the genes, there are
three small ORFs (84, 85 and 86) at the extreme right end
of the genome that are transcribed rightwards (Fig. 4).
Although all three putative genes are small (coding for
predicted proteins of 7.2, 5.6 and 6.8 kDa), at least 84 and
86 have quite respectable coding potential (as determined
by codon usage). However, the most compelling evidence
for the veracity of these assignments is that gp84 has
evident sequence similarity to L5 gp79 (76% identity),
which is encoded within the non-essential region in L5 and
is absent from D29.
The gene 84±85 segment has features that suggest
that it may be similar to the morons identified in the
lambda-related phages (Juhala et al., 2000). Morons have
been identified as small genomic segments, typically
containing one or more ORFs flanked by an upstream
promoter and a downstream terminator, which are found
in one genome but absent from others, even though the
flanking sequences are conserved (Juhala et al., 2000).
In most cases, the moron ORF is probably expressed
from the prophage, with the terminator acting to prevent
readthrough transcription of downstream genes. The
terminators are tolerated in the genome, as the presence
of antiterminator systems prevents them from interfering
with lytic growth. Morons of this type have not been
identified previously in mycobacteriophage genomes, which
could be explained, in part, by a lack of antiterminator
mechanisms in these phages. Although the gene 84±85
segment does not lie between genomic segments that are
closely related to L5, we note that there is a sequence
immediately downstream of 85 that could form an RNA
structure with a 12 bp GC-rich stem with a 4 base loop,
Mycobacteriophage Bxb1 genome 965
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

followed by the sequence 5 0-UUUCU, which could act as
a rho-independent transcription terminator in M. smegmatis,
and preliminary evidence suggests that there is a promoter
located upstream of gene 84 (Jain and Hatfull, 2000).
Gene expression
We have analysed the patterns of protein synthesis during
Bxb1 lytic growth by labelling the synthesized proteins
with 35S-methionine at various times after infection of
M. smegmatis with Bxb1 particles (Fig. 9). In general,
the pattern of gene expression observed is similar to
that seen with L5 (Hatfull and Sarkis, 1993). Soon after
infection, there is a reduction in the expression of the host
genes, as seen by a diminution in the intensity of the
smear representing the multitude of bacterial proteins.
However, by 15 min, the first of the phage proteins can
be seen, with the most prominent being < 33 kDa and
29 kDa in size (Fig. 9). The synthesis of these is maximal
at 20 min after infection and then diminishes until it is
barely detectable after another 20 min. These can there-
fore be classified as early lytic proteins. We do not know
which genes encode these proteins, but note that gene 62
codes for a putative 33.8 kDa product and genes 45, 47
and 57 all code for proteins about 28 kDa in size. By
comparison with L5, it seems likely that the genes
constituting the long leftwards-transcribed operon are
expressed at this early time (Hatfull and Sarkis, 1993).
However, in Bxb1, the great majority of these proteins are
small, with 44 of the possible 51 putative products being
less than 20 kDa. The only predicted products larger than
20 kDa are gp41 (67.6 kDa), gp45 (28.7), gp47 (28.1 kDa),
gp49 (21 kDa), gp57 (28.7 kDa), gp62 (33.8 kDa) and gp69
(22 kDa). Thus, only the absence of bands corresponding
to gp41, gp49 and gp69 (the repressor) cannot be readily
accounted for.
Beginning 20 min after infection, a different set of
proteins is expressed, and the pattern remains fairly
constant throughout the remainder of the infection
(Fig. 9). These are therefore the late proteins and
presumably include those involved in virion structure and
assembly. The slow-migrating highly cross-linked capsid
subunits are particularly prominent (the largest predicted
primary gene product is gp22, which is 84.7 kDa). The
specific identities of most of the other bands are not
clear, and we note that the prominent band migrating
slightly more slowly than the 50.3 kDa marker (see
Fig. 9) could be gp8 (53.3 kDa), gp11 (53.7 kDa) or
gp30 (54.9 kDa). In the expression of L5 late lytic
proteins, the most prominent protein seen is the major
capsid protein, which is predicted to be 41.8 kDa in Bxb1
(Table 2). It seems unlikely that this would migrate more
slowly than the 50.3 kDa marker, although this possibility
cannot be excluded.
The large number of genes in the Bxb1 genome are
probably expressed from just a few transcriptional promo-
ters. The genome organization suggests that there must be
at least three promoters: a leftwards-facing promoter for
early lytic gene expression; a second leftwards-facing
promoter for repressor synthesis; and at least one right-
wards-facing promoter. Promoter active regions have been
identified for all three of these requirements (Jain and
Hatfull, 2000). For example, the Pleft promoter, situated in
the 83±84 intergenic region, initiates transcription (at co-
ordinate 48 802) for early lytic expression and is analogous
to the Pleft promoter of L5 (Nesbit et al., 1995). Like the L5
Pleft promoter, Bxb1 Pleft has a repressor binding site
overlapping the 235 region and is directly repressed by
Bxb1 gp69 (Jain and Hatfull, 2000). The region immediately
upstream of the repressor gene (69) has also been shown
to be transcriptionally active with two transcription initiation
sites at co-ordinates 44 778 and 44 886; these correspond
to two of the three promoters upstream of L5 gene 71
(Nesbit et al., 1995). We have also identified a rightwards
promoter, which (like Bxb1 Pleft) is located within the 83±84
intergenic region and is presumably responsible for the
expression of genes 84±86 (Jain and Hatfull, 2000).
However, this promoter (Pright) is under repressor control
and is therefore activated at early stages of lytic growth.
Thus, it is not clear whether the structural genes expressed
late in lytic growth are expressed from this promoter via an
Fig. 9. Synthesis of Bxb1 proteins after infection of M. smegmatis.A culture of M. smegmatis mc2155 was infected with Bxb1 at anMOI of 100. Proteins were pulse labelled with [35S]-methionineeither 5 min before infection (25) or at the time points indicated ontop of each lane (in min) and separated by SDS±PAGE. Themolecular weights of standards are shown (in kDa) on the right.
966 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

unidentified antitermination system or if there is an additional
rightwards promoter located downstream of gene 86.
Discussion
Bxb1 is one of the more remarkable mycobacterio-
phages to be studied to date. A most unusual feature is
the formation of halos that surround the plaques and can
expand to cover a large area of uninfected cells.
Although phages of E. coli and Klebsiella have pre-
viously been shown to form halos around plaques, these
halos are typically more modest in size than the Bxb1
halos (but could possibly enlarge upon longer incubation
times). The simplest explanation for Bxb1 halo formation
is that the phage particles contain an enzyme activity
associated with a component of the phage tails and, as
phage particles diffuse through the top agar layer, this
activity modifies the external features of the non-growing
mycobacterial cells.
Although it is not obvious from examination of the Bxb1
genome which genes might be involved in halo formation,
genes 8 and 30 have some interesting properties. Bxb1
gp8 is a 53 kDa protein, of which part has sequence
similarity to chitinases, and could conceivably be involved
in degrading complex sugars on the surface of the cell.
However, D29 gp10 has a similar segment, and D29 does
not form halos. Although this does not rule out Bxb1 gp8
as a candidate for this activity, it is unclear why D29 gp10
would not confer a similar phenotype. In contrast, Bxb1
gp30 has no counterpart in any of the other well-studied
mycobacteriophages, and its sequence similarities to
fibronectin type III and D-ala-D-ala carboxypeptidase
domains would be consistent with an activity affecting
the glycopeptides in the M. smegmatis cell wall. More-
over, gene 30 is situated immediately downstream of
other tail genes, and it is therefore reasonable to suppose
that gp30 may also be a structural component of Bxb1
tails, as described for known phage-encoded capsule
depolymerases (Bessler et al., 1973; 1975).
The host ranges of the mycobacteriophages are likely
to be determined mainly by the structural features at the
tips of the tails, which are presumed to come into direct
contact with the bacterial cell surface. In L5, the minor tail
proteins, gp27 and gp28, and perhaps those encoded
immediately downstream (gp30, gp31 and gp32) are good
candidates for forming these structures. As Bxb1 appears
to use different receptors from L5 and D29, differences
in this part of the genome are good candidates for
contributing to host specificity. The most obvious depart-
ures from L5 are that Bxb1 gp23 has a large central
additional segment compared with its relative, gp27 in L5
and D29, and there are several Bxb1 genes (e.g. 24, 27
and 28) that have no counterparts in L5, D29 or TM4. It
is reasonable to suppose that these differences confer
differently structured virion tails, but which of these, if any,
confers host specificity remains to be evaluated.
Bxb1 is the first of the mycobacteriophages to be
studied that uses a serine-recombinase as the phage
integrase, although the phage-like element, fRv1, also
encodes a similar recombinase (Cole et al., 1998; Hendrix
et al., 1999). The organization of the integration apparatus
in Bxb1 is also different from that in L5, in that the Bxb1
integrase gene (35) is expressed in the same direction as
the late lytic genes. It therefore seems probable that there
is a promoter upstream of gene 35 that expresses
integrase for the establishment of lysogeny. There are
two plausible locations for such a promoter, within the
intergenic gap between genes 34 and 35 (where the attP
site is thought to be) or perhaps within the 120 bp gap
between genes 33 and 34. There is a plausible promoter
sequence within this region that overlaps the stoperator
site at co-ordinates 28 941±28 953, suggesting that it
would therefore be under gp69 regulation, which is
unexpected but not implausible. We have not been
able to identify any candidates for a gene encoding an
excisionase function, although phage excisionases are a
highly diverse group of proteins that are not easy to identify
through sequence analysis alone (Lewis and Hatfull, 2000).
Bxb1 provides several intriguing insights into bacter-
iophage evolution. First, it helps to establish that the
complex mosaic relationships seen among the lambda-
like phages of E. coli (Juhala et al.. 2000) and other phage
families (Brussow et al., 1998; Desiere et al., 1999;
Lucchini et al., 1999) are not unique to those groups,
but are also seen among the mycobacteriophages. In
particular, the Bxb1, L5 and D29 genomes share
regions of commonality (mostly seen as related protein
sequences, but also seen in segments of DNA sequence
similarity) that are interspersed with apparently unrelated
segments. The majority of these discontinuities are likely
to have occurred by illegitimate recombination events
at gene boundaries between distant relatives of these
phages. However, the insertions within the middle
portions of Bxb1 genes 8 and 23 illustrate that there are
alternative mechanisms of generating genome variation,
which may also involve illegitimate recombination events
or other processes that are not yet understood.
Antitermination systems have not been identified in
mycobacteriophages L5 (Hatfull and Sarkis, 1993), D29
(Ford et al., 1998a) or TM4 (Ford et al., 1998b). However,
there are few factor-independent terminators in these
genomes, and the few that are present are located at the
very end of the leftwards and rightwards operons (Fig. 4);
there is therefore no obvious demand for antitermination
(there could, of course, be numerous factor-dependent
terminators that have not been identified). A similar
argument can be made for Bxb1, except for the presence
of a putative terminator located between genes 85 and 86,
Mycobacteriophage Bxb1 genome 967
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

which have no counterpart in L5 or D29. If there is no
antitermination system in Bxb1, then transcription of
genes 84 and 85 presumably ends there and another
rightwards promoter must be responsible for late tran-
scription. However, we cannot rule out the intriguing
possibility that this promoter is also used for expression of
the late genes and that there is an antiterminator that
enables passage through the terminator. This scenario is
analogous to the Q-mediated regulation of transcription
from the pR 0 promoter of lambda.
Bxb1 should represent a rich resource for mycobacter-
ial genetics. Many of its novel features will enhance our
understanding of its mycobacterial hosts when investi-
gated further. For example, elucidation of the phage-
encoded enzymes that are responsible for halo formation
should provide insights into the structure of the myco-
bacterial envelope, and the molecular basis for its host
range (especially in comparison with that of other myco-
bacteriophages) will provide information on cell mem-
brane and wall structure. The serine-recombinase
integrase (gp35) presents a new model system for
understanding the mechanisms of site-specific recombi-
nation as well as enabling the construction of new
integration-proficient vectors that are compatible with
those described previously from L5 and D29 (Lee et al.,
1991; Ribeiro et al., 1997; PenÄa et al., 1998). Finally,
although the number of mycobacteriophage genomes
sequenced to date is still small, it is remarkable that three
(L5, D29 and Bxb1) share similar genomic architectures
(in spite of their diverse geographical origins), form a
small group and are more similar to each other than they
are to TM4. As additional mycobacteriophage genome
structures are determined, it will be of interest to see how
this group relates to the larger population structure of
mycobacteriophages.
Experimental procedures
Bacteria and phages
Mycobacteriophage Bxb1 was isolated by one of us (W.R.J.)from a soil sample after enrichment on a culture of M.smegmatis mc2155. Stocks prepared from a single plaquewere used for all subsequent studies. MycobacteriophagesL5, D29, TM4 and all bacterial strains used were laboratorystocks. A clear plaque mutant of Bxb1 (designated Bxb1c1)was isolated as a spontaneous mutant on a lawn of M.smegmatis mc2155.
DNA sequence determination
DNA sequencing was determined using a shotgun strategysimilar to that described previously for the sequencedetermination of the genomes of mycobacteriophages D29(Ford et al., 1998a) and TM4 (Ford et al., 1998b). A librarywas generated by cleaving Bxb1 DNA with DNase I, cloning
the fragments into the EcoRV site of pBluescript SK±(Stratagene) and thermocycling sequencing reactions withfluorescently labelled dideoxy terminators performed onrandomly chosen clones. Sequences were determined onan ABI377 sequencer (Perkin-Elmer Applied Biosystems)and compiled and edited in the program SEQUENCHER (GeneCodes). Some additional clones were made by randomcloning of restriction fragments, and the sequence of someparts of the genome was determined by direct priming witholigonucleotides on Bxb1 DNA. All parts of the genome weresequenced on both strands.
Isolation and characterization of Bxb1 lysogens
Bxb1 lysogens were isolated essentially as described bySarkis and Hatfull (1998). A 0.5 ml sample of a saturated M.smegmatis mc2155 culture was mixed with 5 ml of tryptic soytop agar and poured onto a Middlebrook 7H10 agar plate.After solidification of top agar, 10 ml of serial dilutions of aBxb1 phage stock (1012 pfu ml21) was spotted and the dropsallowed to dry. After incubation overnight at 378C, cells fromthe centre of a turbid spot were streaked for single colonieson Middlebrook 7H10 agar plates. Individual colonies werepurified and tested for immunity to Bxb1 and phage release.Bxb1 lysogens with these phenotypes were used forsubsequent studies.
SDS±PAGE analysis of phage particles
A 50 ml aliquot from a 1012 pfu ml21 stock of density gradient-purified Bxb1 phage was centrifuged at 14 000 r.p.m. for20 min at room temperature to pellet the phage particles. Thephage pellet was suspended in 75 ml of buffer containing20 mM dithiothreitol (DTT) and 12.5 mM EDTA by vigorousvortexing. The suspension was freeze±thawed in liquidnitrogen five times, followed by heating at 808C for 5 min.The suspension turned viscous as a result of the release ofchromosomal DNA from the virions. The phage DNA wassheared by sonication for 1 min. Next, 25 ml of 1 � SDSsample buffer (60 mM Tris-Cl, pH 6.8, 1% SDS, 350 mM b-mercaptoethanol, 10% glycerol) was added to the sample,and it was boiled for 2 min in a water bath. Finally, 25 ml ofthe resulting solution was electrophoresed through a 10%SDS±polyacrylamide gel (acrylamide±bisacrylamide, 100:1),and the resolved proteins were visualized by staining the gelwith Coomassie brilliant blue dye.
Isolation and characterization of phage-resistant mutants
Phage-resistant mutants were isolated by screening librariesof M. smegmatis mc2155 transposon mutants for resistanceto either D29 or I3. Transposon libraries were constructed asdescribed previously using either Tn5367 (Bardarov et al.,1997) or Tn5370 (Cox et al., 1999) delivered using aconditionally replicating TM4 phage. M. smegmatis mc2155was infected with phAE87 containing either Tn5367 (kanr) orTn5370 (hygr) and plated on media containing kanamycinor hygromycin at 378C. Approximately 2000 independentmutants for each transposon were inoculated into 96-wellMicrowell plates (Nunc) containing LB broth plus antibiotics.
968 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

Individual M. smegmatis clones from the transposon muta-genesis collection were transferred with a 96-spike replicator(Nunc-TSP) onto plates (Nunc) with Middlebrook 7H10 solidmedium, containing 1010 pfu per plate of either D29 or I3phages. Single colonies were purified from phage-resistantcandidates and retested. Chromosomal DNAs were isolatedfrom each mutant, and a short DNA sequence adjacent to thetransposon was determined, revealing that each transposoninsertion was unique. Each mutant was screened for its abilityto plaque Bxb1, I3, D29, TM4 and L5.
35S labelling of phage-encoded proteins
A 15 ml culture of M. smegmatis mc2155 was grown inMiddlebrook 7H9 medium (without ADC supplement) to anA600 of 1.5, and the cells were harvested by centrifugation atroom temperature. Cells were washed twice with 7H9medium lacking ADC and finally suspended in 7H9 mediumcontaining 2% glucose and 1 mM CaCl2 to an A600 of 0.5. A12 ml aliquot of this cell suspension was transferred to anautoclaved 100 ml flask, which was incubated at 378C withshaking. Caesium chloride-purified Bxb1 phages were addedto this cell suspension to a multiplicity of infection (MOI) of100. Aliquots (1 ml) were removed at time points 0, 5, 10, 15,20, 25, 30, 40, 50 and 60 min as well as 5 min beforeinfection. At each time point, the proteins were pulse labelledfor 3 min with 8.5 pmol of [35S]-methionine (10 mCi; NENLife Science Products), immediately frozen on dry ice andsubsequently collected by centrifugation at 14 000 r.p.m. at48C for 30 min. One millilitre of 0.2% trichloroacetic acid(TCA) was added to each pellet, followed by centrifugation.The pellets were washed twice with chilled acetone toremove TCA and air dried. Total cell proteins were solubilizedby boiling the pellets in 100 ml of 2 � SDS sample buffer for15 min and resolved by electrophoresis through a 10% SDS±polyacrylamide gel. The gel was transferred into 500 ml ofgel-destaining solution (25% methanol, 7% acetic acid) for15 min to remove SDS, followed by drying under vacuum ona sheet of Whatman paper. Labelled proteins were visualizedby autoradiography.
Acknowledgements
This work was supported by NIH grants AI28927 andGM51975. We would like to thank Aisha Mitchell for excellenttechnical assistance, and Marty Pavelka for helpful discussions.
References
Altmann, F., Kwiatkowski, B., Stirm, S., Marz, L., and Unger,F.M. (1986) A bacteriophage-associated glycanase cleavingbeta-pyranosidic linkages of 3-deoxy-D-manno-2-octulosonicacid (KDO). Biochem Biophys Res Commun 136: 329±335.
Bardarov, S., Kriakov, J., Carriere, C., Yu, S., Vaamonde, C.,McAdam, R.A., et al. (1997) Conditionally replicatingmycobacteriophages: a system for transposon delivery toMycobacterium tuberculosis. Proc Natl Acad Sci USA 94:10961±10966.
Barsom, E.K., and Hatfull, G.F. (1996) Characterization ofMycobacterium smegmatis gene that confers resistance to
phages L5 and D29 when overexpressed. Mol Microbiol 21:159±170.
Beilharz, H., Kwiatkowski, B., and Stirm, S. (1978) On thedeacetylase activity of Vi bacteriophage III particles. ActaBiochim Pol 25: 207±219.
Bessler, W., Freund-Molbert, E., Knufermann, H., Rudolph,C., Thurow, H., and Stirm, S. (1973) A bacteriophage-induced depolymerase active on Klebsiella K11 capsularpolysaccharide. Virology 56: 134±151.
Bessler, W., Fehmel, F., Freund-Molbert, E., Knufermann, H.,and Stirm, S. (1975) Escherichia coli capsule bacteriophages.IV. Free capsule depolymerase 29. J Virol 15: 976±984.
Brown, K.L., Sarkis, G.J., Wadsworth, C., and Hatfull, G.F.(1997) Transcriptional silencing by the mycobacteriophageL5 repressor. EMBO J 16: 5914±5921.
Brussow, H., Bruttin, A., Desiere, F., Lucchini, S., and Foley,S. (1998) Molecular ecology and evolution of Streptococ-cus thermophilus bacteriophages ± a review. Virus Genes16: 95±109.
Cole, S.T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C.,Harris, D., et al. (1998) Deciphering the biology ofMycobacterium tuberculosis from the complete genomesequence. Nature 393: 537±544.
Cox, J.S., Chen, B., McNeil, M., and Jacobs, W.R., Jr (1999)Complex lipid determines tissue-specific replication ofMycobacterium tuberculosis in mice. Nature 402: 79±83.
Desiere, F., Lucchini, S., and Brussow, H. (1999) Compara-tive sequence analysis of the DNA packaging, head, andtail morphogenesis modules in the temperate cos-siteStreptococcus thermophilus bacteriophage Sfi21. Virology260: 244±253.
Donnelly-Wu, M.K., Jacobs, W.R., Jr, and Hatfull, G.F.(1993) Superinfection immunity of mycobacteriophage L5:applications for genetic transformation of mycobacteria.Mol Microbiol 7: 407±417.
Elsasser-Beile, U., and Stirm, S. (1981) Substrate specificityof the glycanase activity associated with particles ofKlebsiella bacteriophage no. 6. Carbohydr Res 88: 315±322.
Ford, M.E., Sarkis, G.J., Belanger, A.E., Hendrix, R.W., andHatfull, G.F. (1998a) Genome structure of mycobacter-iophage D29: implications for phage evolution. J Mol Biol279: 143±164.
Ford, M.E., Stenstrom, C., Hendrix, R.W., and Hatfull, G.F.(1998b) Mycobacteriophage TM4: genome structure andgene expression. Tubercle Lung Dis 79: 63±73.
Fullner, K.J., and Hatfull, G.F. (1997) Mycobacteriophage L5infection of Mycobacterium bovis BCG: implications forphage genetics in the slow-growing mycobacteria. MolMicrobiol 26: 755±766.
Hatfull, G.F., and Jacobs, W.R., Jr (1994) Mycobacterio-phages: cornerstones of mycobacterial research. In Tuber-culosis: Pathogenesis, Protection and Control. Bloom, B.R.(ed. ). Washington: American Society for MicrobiologyPress, pp. 165±183.
Hatfull, G.F., and Sarkis, G.J. (1993) DNA sequence,structure and gene expression of mycobacteriophage L5:a phage system for mycobacterial genetics. Mol Microbiol7: 395±405.
Hendrix, R.W., Smith, M.C., Burns, R.N., Ford, M.E., andHatfull, G.F. (1999) Evolutionary relationships among
Mycobacteriophage Bxb1 genome 969
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970

diverse bacteriophages and prophages: all the world's aphage. Proc Natl Acad Sci USA 96: 2192±2197.
Jacobs, W.R., Jr, Barletta, R.G., Udani, R., Chan, J., Kalkut,G., Sosne, G., et al. (1993) Rapid assessment of drugsusceptibilities of Mycobacterium tuberculosis by means ofluciferase reporter phages. Science 260: 819±822.
Jain, S., and Hatfull, G.F. (2000) Transcriptional regulationand immunity in mycobacteriophage Bxb1. Mol Microbiol38: 971±985.
Juhala, R.J., Ford, M.E., Duda, R.L., Youton, A., Hatfull,G.F., and Hendrix, R.W. (2000) Genomic sequences ofbacteriophages HK97 and HK022: pervasive geneticmosaicism in the lambdoid bacteriophages. J Mol Biol299: 27±51.
Kwiatkowski, B., and Stirm, S. (1987) Polysialic aciddepolymerase. Methods Enzymol 138: 786±792.
Kwiatkowski, B., Boschek, B., Thiele, H., and Stirm, S. (1983)Substrate specificity of two bacteriophage-associatedendo-N-acetylneuraminidases. J Virol 45: 367±374.
Lee, M.H., Pascopella, L., Jacobs, W.R., Jr, and Hatfull, G.F.(1991) Site-specific integration of mycobacteriophage L5:integration-proficient vectors for Mycobacterium smeg-matis, Mycobacterium tuberculosis, and bacille Calmette-Guerin. Proc Natl Acad Sci USA 88: 3111±3115.
Lemassu, A., Ortalo-Magne, A., Bardou, F., Silve, G.,Laneelle, M.A., and Daffe, M. (1996) Extracellular andsurface-exposed polysaccharides of non-tuberculousmycobacteria. Microbiology 142: 1513±1520.
Levin, M.E., Hendrix, R.W., and Casjens, S.R. (1993) Aprogrammed translational frameshift is required for thesynthesis of a bacteriophage lambda tail assembly protein.J Mol Biol 234: 124±139.
Lewis, J.A., and Hatfull, G.F. (2000) Identification andcharacterization of mycobacteriophage L5 excisionase.Mol Microbiol 35: 350±360.
Little, E., Bork, P., and Doolittle, R.F. (1994) Tracing thespread of fibronectin type III domains in bacterial glycohy-drolases. J Mol Evol 39: 631±643.
Loessner, M.J., Inman, R.B., Lauer, P., and Calendar, R.(2000) Complete nucleotide sequence, molecular analysisand genome structure of bacteriophage A118 of Listeriamonocytogenes: implications for phage evolution. MolMicrobiol 35: 324±340.
Lucchini, S., Desiere, F., and Brussow, H. (1999) Compara-tive genomics of Streptococcus thermophilus phagespecies supports a modular evolution theory. J Virol 73:8647±8656.
Nesbit, C.E., Levin, M.E., Donnelly-Wu, M.K., and Hatfull,G.F. (1995) Transcriptional regulation of repressor synth-esis in mycobacteriophage L5. Mol Microbiol 17: 1045±1056.
Niemann, H., Birch-Andersen, A., Kjems, E., Mansa, B., andStirm, S. (1976) Streptococcal bacteriophage 12/12-bornehyaluronidase and its characterization as a lyase (EC4.2.99.1) by means of streptococcal hyaluronic acid andpurified bacteriophage suspensions. Acta Pathol MicrobiolScand B 84: 145±153.
Oyaski, M., and Hatfull, G.F. (1992) The cohesive ends ofmycobacteriophage L5 DNA. Nucleic Acids Res 20: 3251.
Pedulla, M.L., Lee, M.H., Lever, D.C., and Hatfull, G.F.(1996) A novel host factor for integration of mycobacter-iophage L5. Proc Natl Acad Sci USA 93: 15411±15416.
PenÄa, C.E., Stoner, J., and Hatfull, G.F. (1998) Mycobacter-iophage D29 integrase-mediated recombination: specificityof mycobacteriophage integration. Gene 225: 143±151.
PenÄa, C.E.A., Kahlenberg, J.M., and Hatfull, G.F. (1999)Protein±DNA complexes in mycobacteriophage L5 inte-grative recombination. J Bacteriol 181: 454±461.
Ribeiro, G., Viveiros, M., David, H.L., and Costa, J.V. (1997)Mycobacteriophage D29 contains an integration systemsimilar to that of the temperate mycobacteriophage L5.Microbiology 143: 2701±2708.
Sarkis, G.J., and Hatfull, G.F. (1998) Mycobacteriophages.Methods Mol Biol 101: 145±173.
Sarkis, G.J., Jacobs, W.R., Jr, and Hatfull, G.F. (1995) L5luciferase reporter mycobacteriophages: a sensitive tool forthe detection and assay of live mycobacteria. Mol Microbiol15: 1055±1067.
Snapper, S.B., Lugosi, L., Jekkel, A., Melton, R.E., Kieser, T.,Bloom, B.R., and Jacobs, W.R., Jr (1988) Lysogeny andtransformation in mycobacteria: stable expression offoreign genes. Proc Natl Acad Sci USA 85: 6987±6991.
Stirm, S., Bessler, W., Fehmel, F., and Freund-Molbert, E.(1971a) Bacteriophage particles with endo-glycosidaseactivity. J Virol 8: 343±346.
Stirm, S., Bessler, W., Fehmel, F., Freund-Molbert, E., andThurow, H. (1971b) Isolation of spike-formed particles frombacteriophage lysates. Virology 45: 303±308.
Thorpe, H.M., and Smith, M.C. (1998) In vitro site-specificintegration of bacteriophage DNA catalyzed by a recombi-nase of the resolvase/invertase family. Proc Natl Acad SciUSA 95: 5505±5510.
970 J. Mediavilla et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 38, 955±970
Related Documents











