Review The genetics of sex differences in brain and behavior Tuck C. Ngun 1 , Negar Ghahramani 1 , Francisco J. Sánchez, Sven Bocklandt, Eric Vilain ⇑ David Geffen School of Medicine at UCLA, Gonda Center, Room 5506, 695 Charles Young Drive South, Los Angeles, CA 90095-7088, United States article info Article history: Available online 15 October 2010 Keywords: Sexual differentiation Brain anatomy Sex differences Sexual orientation Gender identity Sex chromosomes SRY Dopamine Behavior abstract Biological differences between men and women contribute to many sex-specific illnesses and disorders. Historically, it was argued that such differences were largely, if not exclusively, due to gonadal hormone secretions. However, emerging research has shown that some differences are mediated by mechanisms other than the action of these hormone secretions and in particular by products of genes located on the X and Y chromosomes, which we refer to as direct genetic effects. This paper reviews the evidence for direct genetic effects in behavioral and brain sex differences. We highlight the ‘four core genotypes’ model and sex differences in the midbrain dopaminergic system, specifically focusing on the role of Sry. We also discuss novel research being done on unique populations including people attracted to the same sex and people with a cross-gender identity. As science continues to advance our understanding of biological sex differences, a new field is emerging that is aimed at better addressing the needs of both sexes: gender-based biology and medicine. Ultimately, the study of the biological basis for sex differences will improve healthcare for both men and women. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction Men and women are different in many ways. These differences include both biological phenotypes [e.g. 191] and psychological traits [e.g. 200]. Some of these differences are influenced by envi- ronmental factors [1,340]. Yet, there are fundamental differences between the sexes that are rooted in biology. Of particular interest are sex differences that have been identi- fied in the brain. Although the brains of men and women are highly similar, they show consistent differences that have important implications for each sex. That is, brain sex differences uniquely af- fect biochemical processes, may contribute to the susceptibility to specific diseases, and may influence specific behaviors. Such bio- logical differences should never be used to justify discrimination or sexism. However, we believe that a thorough understanding of these differences can inform researchers and clinicians so that they can better address important issues. Two examples include how genetic sex can lead to differences between the sexes in the etiol- ogy and the progression of disease and how differences in neural development may result in differences in cognition and behavior. In this paper, we will review sex differences in brain and behav- ior that are not due to the action of hormones secreted by the gonads—which has been the dominant mechanism associated with such differences—but to what we term ‘direct genetic effects.’ These are effects that arise from the expression of X and Y genes within non-gonadal cells and result in sex differences in the func- tions of those cells. First, we will highlight some sex differences at the biological level and at the psychological level. Then, we will review the ‘classic’ view that dominated the field of sex differ- ences—that most sex differences, especially those concerned with reproductive physiology and behavior, were due to the action of hormones produced by the gonads. Next, we will present the emerging view that ‘direct genetic effects’ play an important role as well. Finally, we will discuss novel approaches to studying sex differences by focusing on unique groups of individuals: people with sex-chromosome variations (e.g., Klinefelter Syndrome and Turner Syndrome), people with genetic mutations in the sexual development pathway, people with an atypical sexual-orientation, and people who experience a cross-gender identity. 2. Biological sex differences There are many biological differences between males and females that are beyond the obvious differences at a gross, macro level (e.g., height, weight, and external genitalia). Specifically, there are several important physiological differences that have critical implications including the susceptibility to different diseases and the ability to metabolize different medications. In this section we will highlight some sex differences in neuroanatomy and neurochemistry. 2.1. Neuroanatomy The two sexes have similar but not identical brains. Most brain studies have focused on gross manifestations of these 0091-3022/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.yfrne.2010.10.001 ⇑ Corresponding author. Fax: +1 (310) 794 5446. E-mail address: [email protected] (E. Vilain). 1 These authors contributed equally to this work. Frontiers in Neuroendocrinology 32 (2011) 227–246 Contents lists available at ScienceDirect Frontiers in Neuroendocrinology journal homepage: www.elsevier.com/locate/yfrne

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Frontiers in Neuroendocrinology 32 (2011) 227–246
Contents lists available at ScienceDirect
Frontiers in Neuroendocrinology
journal homepage: www.elsevier .com/locate /yfrne
Review
The genetics of sex differences in brain and behavior
Tuck C. Ngun 1, Negar Ghahramani 1, Francisco J. Sánchez, Sven Bocklandt, Eric Vilain ⇑David Geffen School of Medicine at UCLA, Gonda Center, Room 5506, 695 Charles Young Drive South, Los Angeles, CA 90095-7088, United States
a r t i c l e i n f o
Article history:Available online 15 October 2010
Keywords:Sexual differentiationBrain anatomySex differencesSexual orientationGender identitySex chromosomesSRYDopamineBehavior
0091-3022/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.yfrne.2010.10.001
⇑ Corresponding author. Fax: +1 (310) 794 5446.E-mail address: [email protected] (E. Vilain).
1 These authors contributed equally to this work.
a b s t r a c t
Biological differences between men and women contribute to many sex-specific illnesses and disorders.Historically, it was argued that such differences were largely, if not exclusively, due to gonadal hormonesecretions. However, emerging research has shown that some differences are mediated by mechanismsother than the action of these hormone secretions and in particular by products of genes located onthe X and Y chromosomes, which we refer to as direct genetic effects. This paper reviews the evidencefor direct genetic effects in behavioral and brain sex differences. We highlight the ‘four core genotypes’model and sex differences in the midbrain dopaminergic system, specifically focusing on the role ofSry. We also discuss novel research being done on unique populations including people attracted tothe same sex and people with a cross-gender identity. As science continues to advance our understandingof biological sex differences, a new field is emerging that is aimed at better addressing the needs of bothsexes: gender-based biology and medicine. Ultimately, the study of the biological basis for sex differenceswill improve healthcare for both men and women.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Men and women are different in many ways. These differencesinclude both biological phenotypes [e.g. 191] and psychologicaltraits [e.g. 200]. Some of these differences are influenced by envi-ronmental factors [1,340]. Yet, there are fundamental differencesbetween the sexes that are rooted in biology.
Of particular interest are sex differences that have been identi-fied in the brain. Although the brains of men and women are highlysimilar, they show consistent differences that have importantimplications for each sex. That is, brain sex differences uniquely af-fect biochemical processes, may contribute to the susceptibility tospecific diseases, and may influence specific behaviors. Such bio-logical differences should never be used to justify discriminationor sexism. However, we believe that a thorough understanding ofthese differences can inform researchers and clinicians so that theycan better address important issues. Two examples include howgenetic sex can lead to differences between the sexes in the etiol-ogy and the progression of disease and how differences in neuraldevelopment may result in differences in cognition and behavior.
In this paper, we will review sex differences in brain and behav-ior that are not due to the action of hormones secreted by thegonads—which has been the dominant mechanism associated withsuch differences—but to what we term ‘direct genetic effects.’These are effects that arise from the expression of X and Y genes
ll rights reserved.
within non-gonadal cells and result in sex differences in the func-tions of those cells. First, we will highlight some sex differences atthe biological level and at the psychological level. Then, we willreview the ‘classic’ view that dominated the field of sex differ-ences—that most sex differences, especially those concerned withreproductive physiology and behavior, were due to the action ofhormones produced by the gonads. Next, we will present theemerging view that ‘direct genetic effects’ play an important roleas well. Finally, we will discuss novel approaches to studying sexdifferences by focusing on unique groups of individuals: peoplewith sex-chromosome variations (e.g., Klinefelter Syndrome andTurner Syndrome), people with genetic mutations in the sexualdevelopment pathway, people with an atypical sexual-orientation,and people who experience a cross-gender identity.
2. Biological sex differences
There are many biological differences between males and femalesthat are beyond the obvious differences at a gross, macro level (e.g.,height, weight, and external genitalia). Specifically, there are severalimportant physiological differences that have critical implicationsincluding the susceptibility to different diseases and the ability tometabolize different medications. In this section we will highlightsome sex differences in neuroanatomy and neurochemistry.
2.1. Neuroanatomy
The two sexes have similar but not identical brains. Mostbrain studies have focused on gross manifestations of these

Table 1Selected neuroanatomical sex differences in the rat.
Structure/region Known roles Sex difference Basis of difference
Sexually dimorphicnucleus of thepreoptic area(SDN-POA)
The POA is implicated in the regulation of male copulatorybehavior [225]. Lesions of the SDN alone slow acquisitionof this behavior. Potential human equivalent is INAH-3 [4]
2.6 times larger in males [118] Perinatal aromatized androgen decreasesneuronal apoptotic rates in males [317]
Anteroventralperiventricularnucleus (AVPV)
Involved in regulating the luteinizing hormone surge infemales [317] and male copulatory behavior [262]
2.2 times larger in females with ahigher cell density [45]
Degeneration of cells in this region is greaterin males [308] due to prenatal action ofandrogen
Bed nucleus of striaterminalis(BNST)
Plays a role in the control of male sexual behavior [100],release of gonadotropin [32], and modulation of stress[329,134]
The principal nucleus (BNSTp) islarger in volume in males [85]
The larger volume in males is due tosexually different apoptotic rates caused bytestosterone [109]
Corpus callosum Conducts information between the two halves of thecortex [304]
Larger in neonatal males [351] Organizational effects of testosterone leadto masculinization while feminizationappears to be dependent on estrogens[106,105]
Arcuate nucleus(ARC)
Helps regulate the estrus cycle [203], appetite and bodyweight [217]
Neurokin-B neurons innervatecapillary vessels in theventromedial ARC in post-pubertalmales only [66]
Dihydrotestosterone is responsible for themasculine projection pattern [67]
Amygdala Strongly associated with emotion, decision-making andPavlovian conditioning [288]
Adult males have a larger medialnucleus than adult females [221]
Treatment of females with estradiolmasculinizes this nucleus [221]
The posterodorsal aspect of themedial amygdala is 65% larger inmales [148]
Activational effects of circulating androgensaccount for the larger region in males [73]
Cerebral cortex Connected to a wide range of processes from memory [20]to language [33] to emotional processing [237]
Right posterior cortex is thickerthan left but only in males [90]
Gonadal hormones play a role inmaintaining the sex difference (ovariectomymasculinizes the cortex of females) [90]
Ventromedialhypothalamicnucleus (VMN)
Involved in the control of lordosis, mounting, andnorepinephrine release [102]. High concentrations ofsteroid receptor mRNA have been observed in theventrolateral VMN [297]
Females have less synapses in theventrolateral VMN compared tomales [211]
Organizational effects of aromatizedtestosterone appear to be crucial inestablishing the masculine trait [253]
Substantia nigrapars compacta
Made up almost entirely of dopaminergic neurons.Dopamine is involved in control of motor activity [123]
Females have 20% fewerdopaminergic neurons [86]
A genetic component has beendemonstrated in mice [60]
*Note: This table highlights some prominent sex differences in the rat brain but it is by no means exhaustive. Conflicting evidence concerning the examples reported here(particularly in the SDN-POA) exist, and the interpretation of the data is often more complicated than this summary implies.
228 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
differences—namely the size of specific regions or nuclei. Yet, thereis mounting evidence of sex differences at a finer level includingdifferences in synaptic patterns [120,66] and neuronal density[117,211,338]. It is beyond the scope of this article to provide acomprehensive review of all known neuoranatomical differences.We have provided notable sex differences in the rat brain inTable 1. There are also excellent resources for those who are inter-ested in delving deeper into this topic [146,98,28].
We have chosen to focus on neuroanatomical differences in therat because the biological significance and origins of these differ-ences are much clearer than in humans. Neuroanatomical differ-ences in humans are also well-studied although ethical reasonspreclude the experimental manipulations that have led to the find-ings detailed in Table 1. This significantly limits the conclusionsthat can be drawn from any observations made in humans.
Although these neuroanatomical differences are intriguing,most are limited because the practical or functional significanceof these findings are unknown. Discovering the significance ofthese differences is often difficult, even in rodents. de Vries andSodersten have eloquently outlined the challenges facing research-ers who want to understand the link between sex differences instructure and behavior [82]. A highly relevant case study high-lighted in their review concerns the sexually dimorphic nucleusof the preoptic area (SDN-POA). The preoptic area (POA) has beenimplicated in the regulation of male copulatory behavior [225],but the link (if any) between the sex difference in SDN-POA sizeand behavior remains elusive. Masculinizing the size of theSDN-POA in female rats does not result in a correspondingmasculinization and defeminization of behavior [159]. Instead,the SDN-POA may be related to inhibition of female sexual behav-iors [252,141], which might not have been an obvious hypothesisgiven what was known about the POA previously. As science and
technology continue to advance, we will eventually know how tomake sense of the mounting evidence of sex differences in thebrain. For now, it is reasonable to suspect that such differencesmay help account for observed sex differences in behavior, neuro-logical diseases, and cognitive abilities.
2.2. Neurochemistry
Males and females exhibit different patterns of transmitting,regulating, and processing biomolecules. Table 2 presents someof the neurochemical sex differences that have been identified.As a specific example, we focus below on the monoaminergic sys-tem, which has been implicated in several neurological diseasesand mental disorders that differentially affect men and women.
Monoamines are a class of small-molecule neurotransmittersthat are involved in the control of a variety of processes includingreproduction and sexual behavior [183,170], respiration [112], andstress responses [163]. Monoamines have also been implicated innumerous mental disorders, including ones that differentially af-fect men and women [283,303]. Likewise, sex differences in themonoaminergic systems in the rat are well-documented. Reisertand Pilgrim provided a comprehensive review of arguments forthe genetic bases of these differences [259].
Monoamines are subdivided into two groups—catecholaminesand indolamines—based on their molecular structure. The maincatecholamines are dopamine (DA), norepinephrine (NE) and epi-nephrine, which are synthesized from the amino acid tyrosine.Fig. 1 highlights some of the known sex differences of the dopami-nergic system. Regulation of dopamine can potentially control thelevels of the other two catecholamines as they are derived fromdopamine.

Table 2Selected neurochemical sex differences in the brain.
Neurochemicalsystem/pathway
Known roles Species Selected sex differences
Catecholamines(also seeFig. 1)
Involved in the control of a variety of processes includingreproduction and sexual behavior [183,170], respiration [112], andstress responses [163]
Rat Male have higher norepinephrine (NE) levels in the amygdala andhypothalamus at day 25. Direction of this sex difference is reversedat day 300 [296]In response to chronic physical stress, dopamine (DA) activity isupregulated only in males whereas NE activity is increased only infemales [201]
Human Women appear to be more dependent than men on NE for long-term emotional memory formation [323]
Serotonin Modulates a wide variety of processes including mood, aggression,perception, reward, and attention [39]
Ratandhuman
Sex differences in the serotonergic system are found at multiplelevels [234,333,348,305,220]. See Fig. 2 for an illustration of some ofthese differences
Aromatase Plays a key role in sexual differentiation of the brain by convertingtestosterone to 17b-estradiol [231]
Rat Aromatase activity is higher in males than females in many regionsincluding the anterior hypothalamus, BNST and POA [269]Only males experience spikes in the expression of brain-specific andtotal aromatase during embryonic development and shortly after[71]
Vasopressin(VP)
VP in the central nervous system (CNS) has been linked to learning,memory and motor behavior [263]. It has also been connected tothe control of social behaviors such as pair-bonding, parenting andaggression [151]
Rat The number of vasopressin-positive cells is two to three timeshigher in males than in females [81]Vasopressin-positive projections are also two to three times denserin males [81]Intrahypothalamic release of VP due to an increase of plasmaosmolality is higher in females [239]
Human Some studies have found that plasma VP concentrations are higherin men than in women [263]
Cholinergicsystem
The cholinergic system helps regulate the sleep-wake cycle andmodulates synaptic plasticity implicated in memory, learning, anddevelopment [77,165]. Sex differences are found at many points inthe cholinergic system (reviewed in Rhodes and Rubin [263])
Rat Levels of acetylcholine (ACh) are higher in females, regardless ofestrous cycle, than in males [153]. The maximal level of ACh infemales was found at proestrusThe binding affinity of muscarinic ACh receptors is lower in femalesthan in males [18]. Estrogens appear to modulate the bindingactivity of these receptors [96]
Human Men are more sensitive to cholinergic stimulation than women[273]
Opioid system Opioids are a class of chemical for which receptors are foundthroughout the CNS [346,206]. Opioids exert an analgesic effect andalso play a role in stress response and reproduction [315]
Ratandmouse
Generally, l and j class opioids seem more effective in males thanfemales although in some cases the effectiveness is equal [76]. In aminority of cases, they are more effective in females
Human l-opioids appear more effective in women than in men [76]l-opioids show significantly higher binding potential in women inthe amygdala, thalamus and the cerebellum [352]. The sexdifference in the first two regions is reversed after menopause
T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 229
Catecholamines are released by the adrenal glands usually in re-sponse to stress, which affects males and females differently. Forinstance, chronic physical stress impairs memory in male rats only[201]. The sexes also show differing neurochemical responses:Dopamine activity is upregulated in males only whereas norepi-nephrine is upregulated in females only (Fig. 1A). Sex differenceshave also been found in the regulation and modification of dopa-mine (see Fig. 1B and C). Specifically, the enzyme tyrosine hydrox-ylase (TH), which is involved in dopamine synthesis [193], isregulated by Sry—the male sex determination gene—which is notpresent in females. Additionally, levels of norepinehrine in theamygdala differ between the sexes as a result of age. Thus, it islikely that brain catecholaminergic responses to stress might alsodiffer between the sexes.
Another monoamine is serotonin, which is an indolamine. Un-like catecholamines, serotonin is derived from the amino acid tryp-tophan. The serotonergic system shows sex differences (Fig. 2),though many of these differences remain unlinked to behavioraldifferences between men and women. Nevertheless, differencesin this system likely have consequences given the link betweenserotonin and numerous mental disorders [48,275].
3. Psychological and behavioral sex differences
In addition to biological differences, men and women differ inmany psychological and behavioral aspects. For instance, men per-
form better on specific visuospatial aspects (e.g., mental rotation)compared to women; and women perform better on specific verbaltasks (e.g., verbal fluency) compared to men [155]. Furthermore,there is a large sex difference in sexual interests and behaviors, suchas interest in casual sex, interest in multiple sex partners, and inter-est in visual-sexual stimuli (e.g., pornography) [198,281]. Otherexamples are summarized in Table 3.
Some contend that these differences are due to social systemsand gender socialization [cf. 310,53,238]. Nevertheless, biologicaltraits likely contribute to many sex differences. Thus, a thoroughunderstanding of the main determinants involved in expressionof such sex differences can help us better explain the relationshipbetween brain, behavior, and environment. In addition, it allows usto determine how one’s sex potentially influences the risk of devel-oping disorders that manifest and progress differently in men andwomen. Such knowledge can better inform the treatment of thesediseases. Tables 3 and 4 illustrate several factors (e.g. hormonesand genes) that may be causally linked to expression of sex differ-ences in behavior and disease, respectively.
4. The classical view on sex differences
Researchers have examined what contributes to the differenceswe see between males and females. Certainly for humans, socialenvironments influence some of these differences. For instance,social stratifications (e.g., social class and the distribution of social
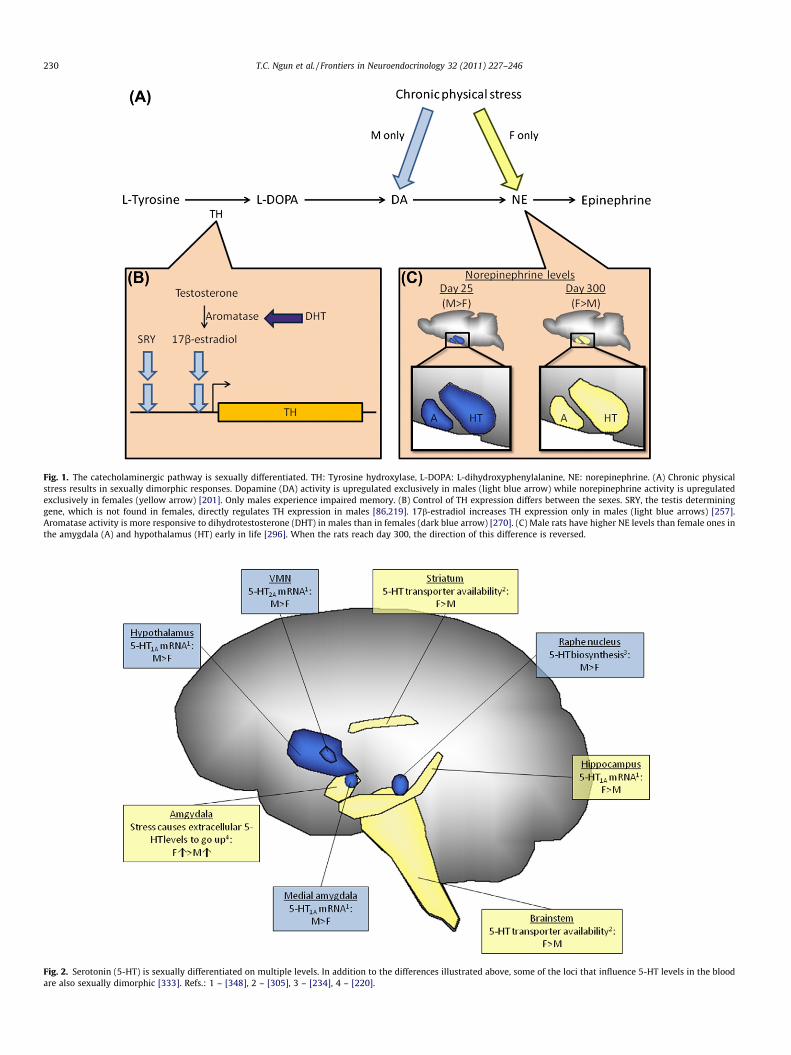
Fig. 1. The catecholaminergic pathway is sexually differentiated. TH: Tyrosine hydroxylase, L-DOPA: L-dihydroxyphenylalanine, NE: norepinephrine. (A) Chronic physicalstress results in sexually dimorphic responses. Dopamine (DA) activity is upregulated exclusively in males (light blue arrow) while norepinephrine activity is upregulatedexclusively in females (yellow arrow) [201]. Only males experience impaired memory. (B) Control of TH expression differs between the sexes. SRY, the testis determininggene, which is not found in females, directly regulates TH expression in males [86,219]. 17b-estradiol increases TH expression only in males (light blue arrows) [257].Aromatase activity is more responsive to dihydrotestosterone (DHT) in males than in females (dark blue arrow) [270]. (C) Male rats have higher NE levels than female ones inthe amygdala (A) and hypothalamus (HT) early in life [296]. When the rats reach day 300, the direction of this difference is reversed.
Fig. 2. Serotonin (5-HT) is sexually differentiated on multiple levels. In addition to the differences illustrated above, some of the loci that influence 5-HT levels in the bloodare also sexually dimorphic [333]. Refs.: 1 – [348], 2 – [305], 3 – [234], 4 – [220].
230 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246

Table 3Sex differences in behavioral traits in humans.
Trait Sex bias Evidence for the role of hormones Evidence for the role of genetic factors Other factors affecting sex differences in behavior
Cognition Men do better at spatial tasks [326] andmathematical problem solving [34]. Womendo better on verbal fluency, articulation, andverbal memory tests [28]
Prenatal hormone effects shown fromstudies of CAH, Turner’s and AndrogenInsensitivity Syndromes [130]
No reliable evidence for the effect of sex-chromosomegenes proven from studies of Turner and XX males[131]
Greater brain asymmetry in men for bothverbal and non-verbal tasks [137,179]
Playbehavior–movement
There are sex differences in choice oftoys, gender of the play partner, socialplay [147] and movement [5,78,248]
Testosterone influences juvenile play[17]
Genetics sex seems to affect play behavior morethan prenatal hormone exposure [17]
Parents and other socializing agents (i.e. peers,community, and child’s own cognitive processes) [28]
Prenatal androgen levels affect playbehavior and movement [235,38]
Developmental experience [129], visual information[316] affect movement organization
Language Women perform better on episodic memory[144] and verbal fluency tasks, men are betterat visuospatial processing [178,195,292]
Estrogen influences word anddeclarative memory abilities in women[291,246,205,298,240,294,228,341,280]
Single nucleotide polymorphisms in the gene,brain derived neurotrophic factor (BDNF)affecting BDNF secretion rates, partly accountingfor greater dependence of females on declarativememory and the sex differences observed inlanguage-related tasks [95]
Greater degrees of left hemispheric lateralization ofbrain for language in males and the bilateral languageprocessing in females [171]
Greater dependence of females on declarativememory and males on procedural memory[320,139]
Testosterone influences word memoryin men [101]
Faster development of hippocampal brainregions in girls, activation of certain brainregions such as hippocampus andparahippocampal gyrus [245,29]
Prenatal testosterone levels relate tolanguage processing in girls [122]
Aggression Foul language, imitation of aggressivemodels, violence and physical aggressionmore common in males [40]
Estradiol and progesterone influencingthe serotonergic system [336,274]
Association between serotonin transporter genepolymorphisms and greater impulsivity in malesbut not females [207]
Low self-control, high impulsivity and negativeemotionality [187]
Weak association between testosteroneand aggression in both sexes [8,49]
Polymorphisms in monoamine oxidase-A (MAOA)gene associated with antisocial personality disorderand aggression in males [62]
Sex-specific disparities in the neural circuitry ofimpulse control and emotion regulation,as well as serotonergic systems [233]
High testosterone levels leading toincreased verbal aggression andimpulsivity in women [61,325]
Larger orbitofrontal cortexes in women [125]
T.C.Ngun
etal./Frontiers
inN
euroendocrinology32
(2011)227–
246231
232 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
power) and social rules (e.g., customs and traditions) may affectthe ability of people to access educational resources or to engagein certain behaviors [156,339]. However, social factors alone donot explain all differences seen between males and females—especially regarding biological differences [72].
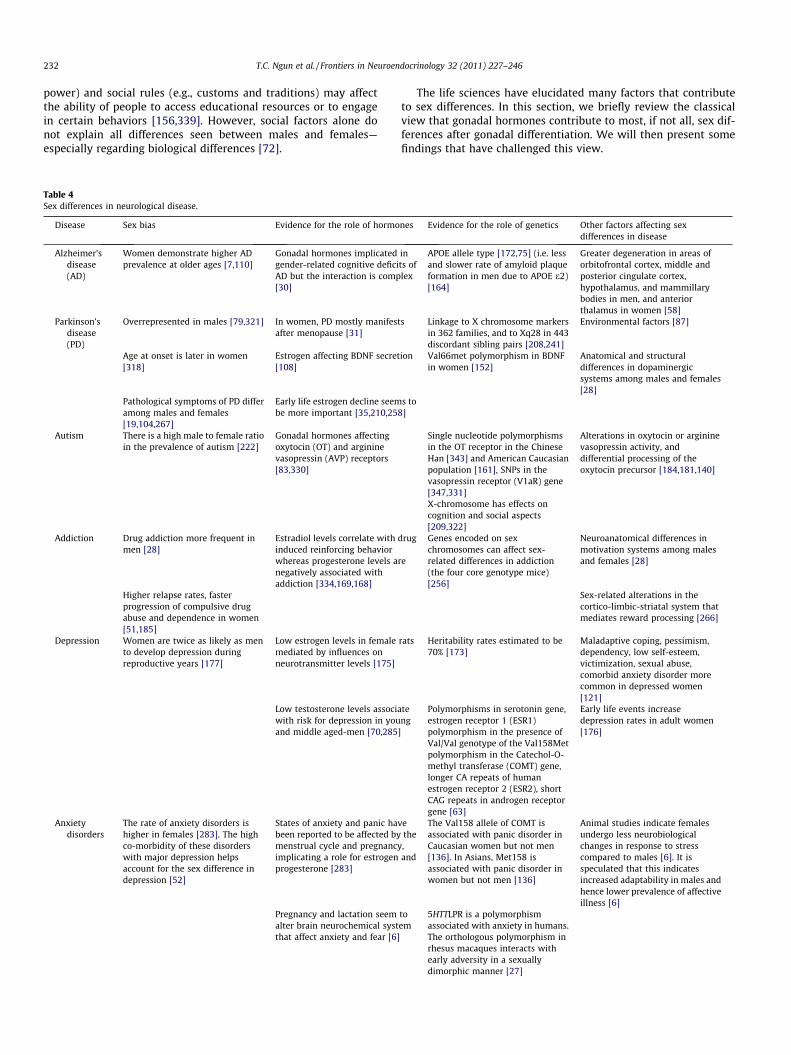
Table 4Sex differences in neurological disease.
Disease Sex bias Evidence for the role of hormo
Alzheimer’sdisease(AD)
Women demonstrate higher ADprevalence at older ages [7,110]
Gonadal hormones implicatedgender-related cognitive deficiAD but the interaction is comp[30]
Parkinson’sdisease(PD)
Overrepresented in males [79,321] In women, PD mostly manifestafter menopause [31]
Age at onset is later in women[318]
Estrogen affecting BDNF secret[108]
Pathological symptoms of PD differamong males and females[19,104,267]
Early life estrogen decline seembe more important [35,210,258
Autism There is a high male to female ratioin the prevalence of autism [222]
Gonadal hormones affectingoxytocin (OT) and argininevasopressin (AVP) receptors[83,330]
Addiction Drug addiction more frequent inmen [28]
Estradiol levels correlate withinduced reinforcing behaviorwhereas progesterone levels anegatively associated withaddiction [334,169,168]
Higher relapse rates, fasterprogression of compulsive drugabuse and dependence in women[51,185]
Depression Women are twice as likely as mento develop depression duringreproductive years [177]
Low estrogen levels in femalemediated by influences onneurotransmitter levels [175]
Low testosterone levels associawith risk for depression in youand middle aged-men [70,285
Anxietydisorders
The rate of anxiety disorders ishigher in females [283]. The highco-morbidity of these disorderswith major depression helpsaccount for the sex difference indepression [52]
States of anxiety and panic havbeen reported to be affected bymenstrual cycle and pregnancyimplicating a role for estrogenprogesterone [283]
Pregnancy and lactation seemalter brain neurochemical systthat affect anxiety and fear [6]
The life sciences have elucidated many factors that contributeto sex differences. In this section, we briefly review the classicalview that gonadal hormones contribute to most, if not all, sex dif-ferences after gonadal differentiation. We will then present somefindings that have challenged this view.
nes Evidence for the role of genetics Other factors affecting sexdifferences in disease
ints oflex
APOE allele type [172,75] (i.e. lessand slower rate of amyloid plaqueformation in men due to APOE e2)[164]
Greater degeneration in areas oforbitofrontal cortex, middle andposterior cingulate cortex,hypothalamus, and mammillarybodies in men, and anteriorthalamus in women [58]
s Linkage to X chromosome markersin 362 families, and to Xq28 in 443discordant sibling pairs [208,241]
Environmental factors [87]
ion Val66met polymorphism in BDNFin women [152]
Anatomical and structuraldifferences in dopaminergicsystems among males and females[28]
s to]
Single nucleotide polymorphismsin the OT receptor in the ChineseHan [343] and American Caucasianpopulation [161], SNPs in thevasopressin receptor (V1aR) gene[347,331]
Alterations in oxytocin or argininevasopressin activity, anddifferential processing of theoxytocin precursor [184,181,140]
X-chromosome has effects oncognition and social aspects[209,322]
drug
re
Genes encoded on sexchromosomes can affect sex-related differences in addiction(the four core genotype mice)[256]
Neuroanatomical differences inmotivation systems among malesand females [28]
Sex-related alterations in thecortico-limbic-striatal system thatmediates reward processing [266]
rats Heritability rates estimated to be70% [173]
Maladaptive coping, pessimism,dependency, low self-esteem,victimization, sexual abuse,comorbid anxiety disorder morecommon in depressed women[121]
teng
]
Polymorphisms in serotonin gene,estrogen receptor 1 (ESR1)polymorphism in the presence ofVal/Val genotype of the Val158Metpolymorphism in the Catechol-O-methyl transferase (COMT) gene,longer CA repeats of humanestrogen receptor 2 (ESR2), shortCAG repeats in androgen receptorgene [63]
Early life events increasedepression rates in adult women[176]
ethe,and
The Val158 allele of COMT isassociated with panic disorder inCaucasian women but not men[136]. In Asians, Met158 isassociated with panic disorder inwomen but not men [136]
Animal studies indicate femalesundergo less neurobiologicalchanges in response to stresscompared to males [6]. It isspeculated that this indicatesincreased adaptability in males andhence lower prevalence of affectiveillness [6]
toem
5HTTLPR is a polymorphismassociated with anxiety in humans.The orthologous polymorphism inrhesus macaques interacts withearly adversity in a sexuallydimorphic manner [27]

Table 4 (continued)
Disease Sex bias Evidence for the role of hormones Evidence for the role of genetics Other factors affecting sexdifferences in disease
Schizophrenia More common in men than inwomen [160]
This disease is not common beforeadolescence and puberty [88]
Eight ultra-rare variants in eightdistinct miRNA genes in 4% ofanalyzed males with schizophrenia[103]
Anatomical and structural braindifferences among males andfemales [115]
Age at onset is later in women,another smaller peak of onsetduring peri- and post-menopause[160,254]
Male schizophrenics have higherlevels of luteinizing hormone (LH)and testosterone than healthysubjects, and femaleschizophrenics higher levels of LHand lower levels of estrogen [186]
Relatives of females withschizophrenia demonstrate higherlevels of the psychotic formswhereas relatives of schizophrenicmen express lower rates ofpsychosis suggesting the presenceof genetic heterogeneity [116]
Higher cortisol levels in males ascompared to females according tosome studies [115]
Pathological symptoms ofschizophrenia differ among malesand females (males experiencemore negative symptoms, greaterdecrease in emotion expressionand recognition; greater paranoiddelusions in women) [115]
Higher rate of CAG repeatexpansions among families offemale patients and not malepatients [224]
Higher sensitivity of the dopaminesystem in men as compared towomen (normal males producemore striatal dopamine inresponse to an amphetaminechallenge as compared to females)[227]
Lower chances of full recovery, anda poorer prognosis in men[160,254]Anatomical brain differencesbetween male and female patients
T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 233
4.1. The role of gonadal hormones
Sexual development in mammals can be divided into two maincomponents: sex determination and sex differentiation [324]. ‘Sexdetermination’ is the process by which the bipotential gonaddevelops into either a testis or an ovary, which depends exclusivelyon genetics. ‘Sex differentiation’ is the development of other inter-nal reproductive structures, the external genitalia, and non-gona-dal sex differences. Unlike sex determination, sex differentiationis driven by gonadal hormones. It was widely believed that sex dif-ferences that emerged after sex determination were largely due tothe actions of gonadal hormones. Examples of this pervasive viewinclude writings from Lillie in 1939 (‘‘[T]he mechanism of sex dif-ferentiation is taken over by extracellular agents, the male and fe-male hormones.” [197]), Jost in 1970 (‘‘The developmental analysisof the body sex characteristics reveals a hormonal control.” [167]),Morris et al. in 2004 (‘‘[A] single factor—the steroid hormone tes-tosterone—accounts for most, and perhaps all, of the known sexdifferences. . .” [225]) and Zhao et al. in 2010 (‘‘[T]he sexual pheno-type of individuals is dependent on the gonad. . .” [349]). We willuse the term ‘classical view’ to refer to this hypothesis.
The classical view was based on decades of compelling researchdemonstrating the organizational and activational effects of gona-dal hormones in vertebrates [196,12]. ‘Organizational effects’ referto the permanent, irreversible changes during development thatorganize the body in either a male- or female-typical pattern. Forinstance, the neonatal surge of testosterone in male rodents leadsto life-long changes in the synaptic pattern of the ventrolateralVMN [253]. ‘Activational effects’ refer to the short-term changesthat occur in the body depending on the presence or absence ofspecific hormones. An example of this is the requirement for thepresence of both estrogen and progesterone to induce or ‘‘activate”lordosis in female rats [251].
Recently, it was found that gonadal hormones might not be thesole contributor to male- and female-typical development. Genesencoded on the sex chromosomes that directly act on the brainto influence neural developmental and sex-specific behaviors havebeen identified—an example of what we describe as direct geneticeffects [113,84]. When we use this term, we refer to effects arisingfrom the expression of X and Y genes within non-gonadal cells that
result in sex differences in the functions of those cells or targetcells. Such direct genetic actions are wide-ranging and can includeeffects of locally produced hormones or other non-hormonal mes-senger molecules. For example, sex differences arising in the brainfrom differential paracrine secretion of neurosteroids would beconsidered a direct genetic effect. The commonality among theseactions is that they are not dependent on mediation by hormonessecreted by the gonads. In many cases, the identity of the messen-ger molecules have yet to be determined. This review will now fo-cus on examples in which sex differences in behaviors are unlikelyto be influenced by only the action of gonadal hormonal secretionsand may in fact be due to direct genetic effects.
4.2. Exceptions to the classical view
The idea that factors other than the gonadal hormone milieucould account for sex differences first gained credence from re-search performed on the zebra finch. In zebra finches, males exhi-bit courtship behaviors that are unique to their sex. Specifically,they possess the ability to sing a distinct courtship song. Thismale-specific ability has been attributed to several brain regionsthat are larger in males compared to females [10,236]. Given thehypothesis that such differences must have been the result ofsex-specific hormones, several researchers unsuccessfully at-tempted to alter the courtship behavior of finches by manipulatinghormone levels [244]. For example, it was shown that castratedmale zebra finches were not significantly different from intactmale zebra finches in terms of song development [9]. Furthermore,female zebra finches that developed testes continued to developfeminine song circuitry and did not exhibit masculine song behav-ior [328,327].
Several other experimental manipulations led researchers toquestion the role of hormones. For instance, Jacobs et al. treatedfemale zebra finches with estrogen at the beginning of hatchinggiven that estrogen induces male sexual differentiation in thezebra finch neural song system [126]. Interestingly, estrogen treat-ment was not able to cause full masculinization of the neuralcircuitry of the zebra finch song system (the song circuitry was stillsmaller compared to control males) [162,299] and supraphysiolog-ical doses of estrogen were required for full masculinization [13].

Fig. 3. 2 � 2 comparison in the four core genotypes model. In this comparison, thefactors are gonadal sex and sex chromosome complement.
234 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
Similarly, it was shown that inhibiting the action of estrogen byusing aromatase blockers in males did not completely preventthe male differentiation pathway [10,91,150,11,16].
The discovery of a rare type of zebra finch provided further sup-port for a new hypothesis regarding sexual differentiation: Thebilateral gynandromorphic finch has male-typical phenotypes onone half of the body (e.g., plumage, testis, and song circuitry) andfemale-typical phenotypes on the other half of the body. Each halfof such finches is either entirely genetically male or genetically fe-male. Thus, each side contains the sex-specific genes necessary forthe development of the corresponding sex-specific traits. In thismodel, while the gonadal hormonal actions in producing sex differ-ences in the brain cannot be completely ruled out (both sides of theneural song system were larger than that of normal females), theirinfluences cannot fully explain the differences observed betweenthe left and right sides of the brain. Given this explanation, themost reasonable theory is that endogenous genetic differences inthe brain cells themselves can also contribute to the unequal dif-ferentiation of the two sides producing sex differences throughtheir local action within the brain [2].
Recent work on gynandromorphic chickens strengthens thecase that the classical view largely does not apply to sexual differ-entiation in birds. Zhao et al. showed that the ‘sex identity’ (or theexpression of sex-specific phenotypes) of somatic cells in birds isdetermined by the sex chromosome complement of those cellsand not the gonadal hormonal environment [349,2]. In mammals,transplantation of somatic cells from one sex into the gonad ofthe other sex reverses the sex identity of the donor somatic cells.For example, XX cells can develop into functioning Sertoli cellswhile XY cells can become functioning granulosa cells [242,56].However, this is not the case in the chicken as male donor cellsintroduced into the developing ovary continued to express amale-specific marker and were excluded from ‘functional’ struc-tures of the host gonad. The host and donor somatic cells were ex-posed to the same hormones, but they responded differently basedon their respective sex chromosome complement.
A second exception to the classical view that we highlight be-low concerns the development of the tammar wallaby. As withbrain development, gonadal hormones drive the sex-specific devel-opment of the external genitalia in most mammals. Specifically,androgens promote the development of male genitalia. However,the formation of reproductive structures in the tammar wallabyappears to be independent of gonadal hormone control and is so-lely due to the effect of sex chromosome complement.
The tammar wallaby is a marsupial that is much smaller thanthe kangaroo. During fetal development, the production of testos-terone, which would typically masculinize mammalian fetuses,does not occur in these marsupials until about the fourth or fifthday after birth [293,260,261,337]. Yet, signs of sex-specific repro-ductive structures (e.g., scrotum, mammary gland, and pouchformation) can be observed as early as several days before birth.In mammals, the development of male-specific structures, isthought to be completely dependent on the action of androgens[324]. Experiments that increased or decreased the action of tes-tosterone or estrogen in the tammar wallaby had no significanteffect on the development of the external genitalia [199,290]. Thissuggested that such differences were not under gonadal hormonecontrol.
A case similar to the gynandromorphic zebra finch has also beenreported in tammar wallabies: This consists of wallabies that areXX on one side of the body and XY on the other side of the body.Such wallabies develop a hemipouch on the XX side and a hemi-scrotum on the XY side even after exposure to circulating gonadalhormones [13,289,74]. As with the zebra finch, such cases chal-lenged the view that all sex differences were due to hormones pro-duced by the gonads.
5. An expanded view on sex differences
In light of scientific findings such as the ones presented above(the zebra finch and the tammar wallaby), the field of sex differ-ences has now come to encompass studies that examine gonadalhormone as well as genetic origins of these differences. One ofthe most significant challenges in studying the establishment ofsex differences in animal models has been the difficulty in separat-ing gonadal sex from chromosomal sex. These two parameters al-most always correlate in an animal.
In the following section, we highlight the ‘four core genotypes’model, which has proven to be a powerful tool in teasing out theeffects of gonadal versus chromosomal sex and enabling research-ers to overcome this confound. We then discuss in-depth sexualdifferentiation and sex differences in the midbrain dopaminergicsystem, focusing specifically on the role of Sry. We discuss theimplications that these differences may have on the developmentof this system as well as neurological health implications.
5.1. The ‘four core genotypes’ model
A 2 � 2 mouse-model was developed to separate the effects ofgonadal sex from chromosomal sex. This model, known as the ‘fourcore genotypes’ (FCG), allows researchers to establish the relativecontribution of sex chromosomes and hormones in sexual differen-tiation as well as the interaction between the two. Arnold and Chenrecently reviewed this model [14]. Here we highlight some of themodel’s basic concepts.
Fig. 3 depicts the effect of the presence or absence of Sry—a12 kb region on the Y chromosome that is responsible for testisdetermination—using the FCG model. An XY mouse should developtestes; however, if Sry is deleted from the Y chromosome (symbol-ized by Y–) then the mouse will develop ovaries [124]. If the Srygene is inserted into any chromosome of an XX mouse (symbolizedby XXSry), then the mouse will develop testes. Finally, if Sry is de-leted from the Y chromosome of an XY mouse and then insertedinto one of its autosomes (symbolized by XY–Sry), then it will devel-op testes.
XY–Sry mice are fully fertile because the presence of Sry pro-motes testes development. XXSry mice lack some of the genes re-quired for sperm production, which are found on the Ychromosome [214], and therefore do not appear to be fertile.

T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 235
However, they have small testes and are fully masculinized interms of measures of male copulatory behavior, social explorationbehavior, and sexually dimorphic neuroanatomical structures inthe septum, and lumbar spinal cord.
XY–Sry mice can be mated with XX females to produce the fourtypes of offspring (XX, XY–, XXSry, and XY–Sry) that can then be usedto assess the impact of a mouse’s chromosomal and gonadal sex ondifferent phenotypes. That is, if there is a difference between micethat carry the Sry gene (i.e., XXSry and XY–Sry) versus those that donot (i.e., XX and XY–), then the observed difference can be attrib-uted to the gonadal type and/or presence of Sry. On the other hand,if there is a difference between mice that have the Y chromosome(i.e., XY– and XY–Sry) versus those that do not (i.e., XX and XXSry),then the observed difference can be attributed to complement ofsex chromosomes (XX versus XY).
5.1.1. Limitations of the FCG modelSome possible limits to the FCG model have been suggested. One
possible limit is that the size, morphology, and function of thegonads are not exactly the same in XX and XY mice of the samegonadal type (e.g. XXSry versus XY-Sry). Consequently, the level ofgonadal hormone secretions in FCG mice may differ during criticalperiods of development—a confound that has yet to be investigated.Yet, numerous phenotypes that are responsive to the organizationaleffects of gonadal hormones (including sexually dimorphic brainstructures) do not differ in XX and XY mice of same gonadal type[288,259,18,33], indicating that XX and XY mice of the same sexare likely experiencing similar levels of gonadal secretions. Forexample, measurements of circulating testosterone in XX and XYmales found no difference in testosterone levels between the groups[15].
A second limit relates to the biochemical and molecular envi-ronment. That is, one cannot rule out the effect of prenatal hor-monal secretions, the influence of adult circulating hormonesproduced by the gonads or other tissues, acute fluctuations in hor-monal levels, and the influence of the Sry transgene (i.e., the poten-tially higher expression-level of the Sry transgene in XY–Sry animalsversus XY mice). For example, the Sry transgene could hasten theearly stages of testis organogenesis in XY–Sry males. Furthermore,several phenotypes have been found to differ between XY andXY–Sry males. However, it is not known whether these differencesare caused by the effect of Sry on androgen production or by someother mechanisms that are not mediated through the action of go-nadal hormones.
To address these limits, it is best to rule out the effect of circu-lating gonadal hormones. An effective approach would be to firstgonadectomize the mice followed by an administration of equiva-lent doses of gonadal steroid hormones. This is particularly impor-tant in the case of XY– females since their level of ovarian steroidhormones differ from that in the XX wild type females [15]. Never-theless, a major limitation still remains: It will not be obviouswhether the sex difference attributed to the complement of sexchromosomes within cells is caused by (a) gene or genes encodedon the Y chromosome; (b) higher dosage of X genes particularly theones that escape X inactivation in XX animals [59]; or (c) the pater-nal imprint of the genes encoded on the X chromosome in XX ani-mals, which changes the expression of these genes to exhibit afemale-specific pattern [244,345]. If one determines that the sexdifference in phenotype is due to the sex chromosome comple-ment, then the next step would be to discover the nature of thegene or genes involved and identify whether those genes are en-coded on the X or Y chromosome and how and where they mediatetheir role [84].
Notwithstanding these potential limitations, a variety of sex dif-ferences have been examined using the FCG model. We reviewthree of these.
5.1.2. Lateral septumOne clear example of the role of sex-chromosome genes in brain
phenotypes can be found in the lateral septum. The lateral septumis part of the limbic system and is involved in stress-related behav-iors. This nucleus is denser in male brains compared to femalebrains. However, it was found that the vasopressin fiber densitywas greater in the lateral septum of XY–Sry and XY– mice comparedto XX and XXSry mice [113]. In addition, an examination of vaso-pressin fiber densities in animals with the same sex chromosomecomplement indicated a role for the action of gonadal steroid hor-mones. No interaction was observed between gonadal sex and sexchromosomes [84].
5.1.3. AddictionOn average, women use addictive drugs at lower levels than
men, but women become addicted to drugs more rapidly thanmen [138]. Based on the FCG model, Quinn et al. showed that thisdifference could be attributed to the differences in the complementof the sex chromosomes and not to the gonadal secretions and/orthe expression of the Sry gene. XX mice developed habitual behav-ior more rapidly than the XY animals independently of their gona-dal phenotype and even after gonadectomy. This implies thatneither gonadal sex nor circulating steroid hormones exert majoreffects on the development of habit-driven behavior in mice [256].
5.1.4. AggressionMales typically exhibit more aggressive behaviors compared to
females [229,69,311]. Recent reports have shown that aggressionlatencies are strongly influenced by the simultaneous action of go-nadal hormones and sex chromosomes. Using the four core geno-types model, it was found that a significant interaction existsbetween the two variables. In this model, the XX females appearedto be slower at displaying aggressive behavior on their firstencounter with an intruder compared to animals in all othergroups [113].
5.2. Direct role of Sry in brain sex differences
Sex differences in the brain may contribute to some of the psy-chological and behavioral differences we observe between thesexes. Furthermore, they may influence the susceptibility to differ-ent diseases. For instance, Parkinson’s disease—a neurodegenera-tive disease that impairs motor function and speech—affectsmore men than women. Research has established a link betweenParkinson’s disease and a loss of dopaminergic neurons in the sub-stantia nigra [114]. Such losses disrupt dopamine pathways, whichleads to many of the symptoms associated with Parkinson’sdisease.
Robust sex differences have been observed in the development,activity, and number of dopaminergic neurons. The data describedbelow represents a clear example of a sex difference in the brainthat has a strong genetic component.
5.2.1. Dopaminergic neurons in rodentsSex differences in dopaminergic neurons have been found prior
to exposure to gonadal steroid hormones. During in utero develop-ment, rat embryos are exposed to a plasma surge of hormonesaround embryonic day 17 or 18 (E17 or E18). Yet, as early asE14, dissociated cell cultures of dopaminergic neurons obtainedfrom male and female rat brainstems were found to be fundamen-tally different in their morphology and function prior to exposureto gonadal steroid hormones [259]. Furthermore, females hadhigher numbers of dopaminergic, tyrosine hydroxylase-immuno-reactive (TH-ir) cells in the midbrain; and their mesencepahlicand diencepahlic neurons produced more dopamine whencompared to males. On the other hand, soma measurements of

236 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
diencephalic neurons showed that male cultures contained largerdopaminergic neurons. Although it is difficult to make accuratemeasurements of hormonal levels in the embryonic brain, it isunlikely that there is a huge sex difference due to gonadal hormoneexposure at this stage as the rat gonad only begins to differentiateat this point. Therefore, this suggests a contribution of sex chromo-some complement and/or sex-specific gene expression.
These differences are not altered even when gonadal hormonelevels are manipulated. Specifically, treatment with estradiol andtestosterone does not eliminate the observed sex differences innumber, size, or function of the dopaminergic cells. These findingswere later replicated in a study using mesencephalic cultures fromthe NMRI strain of mice [295]. Collectively, these observationsstrongly support the idea that some of the sex-specific propertiesof the dopaminergic neurons are under the control of non-hor-monal mechanisms.
5.2.2. The Y chromosome’s role in dopaminergic neuron developmentA study utilizing the four core genotypes model further
strengthened the case that a genetic component largely accountsfor these sex differences. Carruth et al. cultured mesencephalicneurons from E14 animals representing each of the groups fromthe four core genotypes [60]. Cultures from XY– and XY–Sry animalsdeveloped significantly more TH-ir neurons compared to the XXand XXSry animals. However, gonadal sex did have a small effect:Animals that had Sry (and hence testes) were associated with ahigher number TH-ir cells compared to those without Sry. Due tothe design of the four core genotypes model, it is difficult to sepa-rate the direct effects of Sry from its indirect effects on dopaminer-gic neurons or their precursors (e.g., through testis determinationand the subsequent hormonal secretions).
The data pertaining to sex differences in dopaminergic neurondevelopment show sex differences in distinct directions and soare difficult to interpret. In cultures from E14 rats and NMRI mice,the sex difference is the reverse of what was seen with the fourcore genotypes. However, rather than invalidating the findings,the conflicting information highlights the complex interactions be-tween genetics and gonadal hormones in leading to the sex differ-ences that are observed. First, the differences between data fromNMRI mice and the four core genotypes may be attributable tostrain differences. Data from the former study indicate that geneticbackground can significantly affect whether a sex difference is ob-served [295]. Carruth et al. had outbred their mice onto the MF1background [60]. In regards to the differences seen between cul-tures from rats and the four core genotypes, one possible explana-tion is that both androgens and the Y chromosome are needed tolead to the number of dopaminergic neurons being higher in males.Support for this hypothesis comes from our own studies where wesee that the number of dopaminergic neurons in the rat substantianigra is higher in adult males [86]. Additionally, the finding that thepresence of testis was associated with a higher number of theseneurons fits with our hypothesis. There are also important distinc-tions in the timing of the cultures in relation to gonadal develop-ment: while both studies cultured E14 neurons, the bipotentialgonad has already differentiated into a testis to a larger extent inthe mice [97,182] than in the rat at this gestational stage [204].As such, the hormonal environment from which these cultureswere derived may not be the same, which could account for someof the disparity in the direction of the sex difference. An elabora-tion of the hypothesis presented above is that it is not just the tes-tes and androgens that are essential but also Sry, the gene thatinitiates testicular development. Using a rat model, our laboratoryhas found evidence that this may be the case and showed that Sryhas a direct effect on the expression of TH in the substantia nigra[86].
5.2.3. Sry is a direct effector of TH expressionSry is the gene on the Y chromosome that directs the bipotential
mammalian gonad to develop as testes—hence, its name: Sex-determining region on Y. Sry is the founding member of the Soxfamily of proteins, which play a major role in a wide range of bio-logical processes such as neurogenesis, hematopoiesis, and neuralcrest development [189]. Sry contains a high mobility group(HMG) box domain and shows little conservation from mouse tohuman outside this stretch of about 80 amino acids [335].
The HMG box forms a domain that induces a sharp bend in theDNA [332]. It is proposed that this bending of the DNA enhancesrecruitment of specific transcriptional factors. In line with thishypothesis, Sry has two nuclear localization signals within theHMG domain [307] and its ability to activate transcriptionin vitro has been demonstrated [93].
Recently, researchers have used genome-wide surveys to iden-tify targets of Sry in the gonad. The most widely known target ofSry is Sox9 [286]. Two other notable targets of Sry are Cbln4 [50],which codes for the cerebellin precursor; and MAO A, which codesfor monoamine oxidase A [344]. Wu and colleagues also found thatMAO A was upregulated by Sry in the BE(2)C neuroblastoma cellline suggesting that MAO A may be a neural Sry target [344].
Most studies on Sry expression have focused on the gonad andSry’s subsequent effects on sex determination and differentiation[312]. In the developing mouse embryo, Sry is expressed betweenE 10.5 and E 12.5 in the developing genital ridge, prior to overt tes-tis differentiation [128]. Until recently, it was thought that Sry hadno role other than sex determination. However, Sry expression hasbeen found in numerous tissues outside of the testis (see below)and this expression in the adult male rat is now known to have bio-logically significant effects. Sry’s crucial role in the regulation of thecatecholaminergic system is one of the best examples of a directgenetic regulator of a trait that differs between the sexes.
5.2.4. SRY in the brainClépet et al. were the first to perform a survey of SRY expression
in human tissue outside of the gonads [68]. In fetal tissue, SRY wasseen in the brain, adrenal, heart, and pancreas. In adults, transcrip-tion was detected in the kidney, heart, and liver. This study alsoshowed that SRY was expressed in the teratocarcinoma cell lineNT2/D1, which was derived from adult male tissue and whichcan be used as a model for dopaminergic neurons. When NT2/D1was induced to differentiate into neurons by retinoic acid, SRYexpression remained.
SRY expression in the human adult brain was not surveyed until1998. Mayer et al. showed that SRY mRNA was present in the hypo-thalamus, frontal, and temporal cortex of only the adult male[212]. Sry mRNA is also found in the adult male mouse brain whereit can be detected in the midbrain (including the substantia nigra)and hypothalamus in all developmental stages [213].
5.2.5. SRY and the regulation of TH expressionSry has a biologically significant role in the brain in at least one
instance—the regulation of tyrosine hydroxylase (TH) [86,219]. In a2004 study, Milsted et al. found that Sry is a regulator of TH genetranscription [219]. The study looked at Sry’s role in relation toTH in both the brain and the adrenal medulla. They demonstratedthat Sry and TH mRNA were co-localized in the locus coeruleus,substantia nigra, and ventral tegmental area of the male rat(Fig. 4A). They then used a luciferase reporter assay to show thatSry’s ability to upregulate TH expression is dependent on the AP1binding sites in the promoter of TH.
The in vivo significance of those findings was shown and ex-panded upon by a study from our laboratory. By in situ hybridiza-tion, we were able to determine the spatial distribution of SrymRNA within the rodent brain [86]. Specific labeling of Sry was ob-
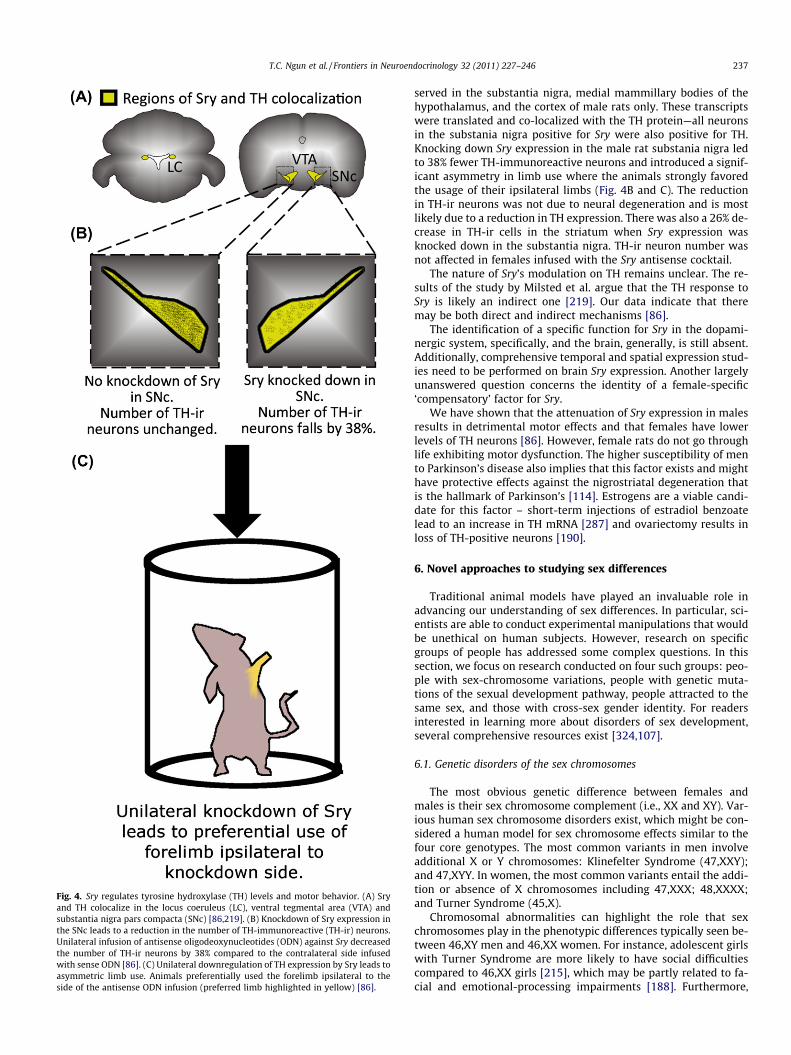
Fig. 4. Sry regulates tyrosine hydroxylase (TH) levels and motor behavior. (A) Sryand TH colocalize in the locus coeruleus (LC), ventral tegmental area (VTA) andsubstantia nigra pars compacta (SNc) [86,219]. (B) Knockdown of Sry expression inthe SNc leads to a reduction in the number of TH-immunoreactive (TH-ir) neurons.Unilateral infusion of antisense oligodeoxynucleotides (ODN) against Sry decreasedthe number of TH-ir neurons by 38% compared to the contralateral side infusedwith sense ODN [86]. (C) Unilateral downregulation of TH expression by Sry leads toasymmetric limb use. Animals preferentially used the forelimb ipsilateral to theside of the antisense ODN infusion (preferred limb highlighted in yellow) [86].
T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 237
served in the substantia nigra, medial mammillary bodies of thehypothalamus, and the cortex of male rats only. These transcriptswere translated and co-localized with the TH protein—all neuronsin the substania nigra positive for Sry were also positive for TH.Knocking down Sry expression in the male rat substania nigra ledto 38% fewer TH-immunoreactive neurons and introduced a signif-icant asymmetry in limb use where the animals strongly favoredthe usage of their ipsilateral limbs (Fig. 4B and C). The reductionin TH-ir neurons was not due to neural degeneration and is mostlikely due to a reduction in TH expression. There was also a 26% de-crease in TH-ir cells in the striatum when Sry expression wasknocked down in the substantia nigra. TH-ir neuron number wasnot affected in females infused with the Sry antisense cocktail.
The nature of Sry’s modulation on TH remains unclear. The re-sults of the study by Milsted et al. argue that the TH response toSry is likely an indirect one [219]. Our data indicate that theremay be both direct and indirect mechanisms [86].
The identification of a specific function for Sry in the dopami-nergic system, specifically, and the brain, generally, is still absent.Additionally, comprehensive temporal and spatial expression stud-ies need to be performed on brain Sry expression. Another largelyunanswered question concerns the identity of a female-specific‘compensatory’ factor for Sry.
We have shown that the attenuation of Sry expression in malesresults in detrimental motor effects and that females have lowerlevels of TH neurons [86]. However, female rats do not go throughlife exhibiting motor dysfunction. The higher susceptibility of mento Parkinson’s disease also implies that this factor exists and mighthave protective effects against the nigrostriatal degeneration thatis the hallmark of Parkinson’s [114]. Estrogens are a viable candi-date for this factor – short-term injections of estradiol benzoatelead to an increase in TH mRNA [287] and ovariectomy results inloss of TH-positive neurons [190].
6. Novel approaches to studying sex differences
Traditional animal models have played an invaluable role inadvancing our understanding of sex differences. In particular, sci-entists are able to conduct experimental manipulations that wouldbe unethical on human subjects. However, research on specificgroups of people has addressed some complex questions. In thissection, we focus on research conducted on four such groups: peo-ple with sex-chromosome variations, people with genetic muta-tions of the sexual development pathway, people attracted to thesame sex, and those with cross-sex gender identity. For readersinterested in learning more about disorders of sex development,several comprehensive resources exist [324,107].
6.1. Genetic disorders of the sex chromosomes
The most obvious genetic difference between females andmales is their sex chromosome complement (i.e., XX and XY). Var-ious human sex chromosome disorders exist, which might be con-sidered a human model for sex chromosome effects similar to thefour core genotypes. The most common variants in men involveadditional X or Y chromosomes: Klinefelter Syndrome (47,XXY);and 47,XYY. In women, the most common variants entail the addi-tion or absence of X chromosomes including 47,XXX; 48,XXXX;and Turner Syndrome (45,X).
Chromosomal abnormalities can highlight the role that sexchromosomes play in the phenotypic differences typically seen be-tween 46,XY men and 46,XX women. For instance, adolescent girlswith Turner Syndrome are more likely to have social difficultiescompared to 46,XX girls [215], which may be partly related to fa-cial and emotional-processing impairments [188]. Furthermore,

238 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
46,XX girls score better than boys on tests of social cognitive skills[300]. The fact that both 46,XY boys and 45,X girls experience moresocial adjustment problems compared to 46,XX girls suggests thepresence of a genetic locus involved in social cognitive skills onthe X chromosome. Data from Skuse et al. [301] suggest that thislocus may be subject to imprinting. Significant differences between45,Xp Turner-syndrome girls (in which the X was of paternal ori-gin) and 45,Xm girls (in which the X was maternally derived) interms of social skills have been reported. 45,Xp had superior socialcompetence and better social skills than 45,Xm girls suggestingthat the genes in this locus are expressed only from the paternalX. This could potentially be one of the reasons why boys are moresusceptible to disorders such as autism that affect social adjust-ment and social skills such as language. In boys the X is only ofmaternal origin and therefore this locus would be silenced.
In other cases, however, the characteristics of individuals withsex chromosome abnormalities may augment the expected sex dif-ference. On average, men in the general population have better vis-uospatial skills than women, and women have better verbal skillsthan men—which suggests that increased dosage of X chromosomegenes may contribute to these skills. However, women with TurnerSyndrome have impaired visuospatial abilities yet greater languageskills compared to control women [272,314].
The role of the Y chromosome in psychosexual differentiation isstill unclear. Work by McCarty et al. indicates that genes on the Ychromosome outside of SRY and the pseudoautosomal region have‘‘no obvious role. . .on psychosexual differentiation in geneticmales [214].” This is difficult to ascertain because (a) the incidenceof XY gonadal dysgenesis is extremely rare, estimated to be 1 in20,000 [64]; and (b) these individuals have been poorly studiedin regards to sexual differentiation of the brain.
6.2. Androgen Insensitivity Syndrome
The role of the androgen receptor (AR) in brain sexual differen-tiation has been discussed in patients with Androgen InsensitivitySyndrome (AIS). AIS is an X-linked recessive disorder that is seen in1 out of 20,400 live male births [250]. There are two forms of AIS:Complete (cAIS) and Partial (pAIS). People with Complete AIS aregenetically male (46,XY with undescended testes) but phenotypi-cally female. However, individuals with Partial AIS typically haveambiguous genitalia.
AIS is caused by mutations in the androgen receptor (AR) gene[255]. There is an important difference in the behavioral phenotypebetween humans and rats with Complete AIS. Humans with Com-plete AIS are female-typical in their play behavior and sexual ori-entation [353]. In contrast, XY rats with AR mutations behavesexually like wild-type males and have a male-typical partner pref-erence [353]. The reasons behind this difference remain unclearalthough the implication is that androgens play an important rolein masculinizing the human brain. However, we cannot completelydiscount the role of estradiol as the expression of aromatase(which converts testosterone to estradiol) is dependent on andro-gen signaling via AR [26].
6.3. Sexual orientation
Of all behavioral differences between males and females, part-ner choice is one of the most pronounced. With very few excep-tions in the Animal Kingdom, males typically choose females tomate with, and females typically choose males to mate with.Although sexual selection is a driving force of evolution, little isknown about the molecular basis of partner preference.
Human sexual orientation is a complex phenotype to study. Partof this difficulty comes from the accurate assessment of sexual ori-entation [218,223], especially when researchers depend on self-
identification, which may be mediated by numerous social andpsychological factors [65,143,194]. Nevertheless, most peoplereport primarily opposite-sex or heterosexual attractions. Yet, asignificant number of people (approximately 2–6%) report pre-dominantly homosexual attractions [89].
The distribution of sexual behavior differs between men andwomen. In men, the distribution is largely bimodal [133]. That is,men are either attracted to one sex or the other. Although thereis disagreement regarding bisexuality among men [342], physio-logical research has found that very few men (even those whoopenly identify as bisexual) show comparable physical attractionto both men and women [265]. The distribution is more complexin women, in which the fraction of women that show exclusivesame-sex attraction is lower than men (1–3%), but many more wo-men than men report erotic fantasies towards both sexes [154].
In this section we will highlight some of the biological researchthat has focused on same-sex attraction. A more thorough reviewis available for interested readers [278].
6.3.1. Neuroanatomy differences in sexual orientationNeuroanatomical differences have been reported for three brain
regions based on sexual orientation in human males: the argininevasopressin neuronal population of the suprachiasmatic nucleus,which is larger in gay men than in male and female controls[309]; the third interstitial nucleus of the anterior hypothalamus(INAH-3), which is smaller in gay men and more similar in sizeto female controls [192]; and the anterior commissure, which islarger in gay men than in control males and females [3]. The mostdiscussed anatomical finding was in INAH-3 [192]. Although sub-sequent researchers reported inconsistent findings [57], a compa-rable difference was found in sheep [271].
Approximately 8–10% of the domestic ram population has beenfound to sexually prefer other males. Unlike other animal modelsshowing atypical sexual behavior, these male-oriented ramsmount and ejaculate on other males versus simply exhibiting apassive stance (i.e., lordosis). Consequently, they are an ideal ani-mal model of male homosexuality because their coital behavioris masculine but their sexual partner preference is feminine.
An analog of the sexually dimorphic nucleus (ovine SDN oroSDN)—a hypothalamic nucleus thought to be involved in mateselection—was identified in the sheep brain [271]. The oSDN wasfound to be larger in female-oriented rams compared to male-oriented rams, and ewes; the latter two groups had oSDN’s compa-rable in size. It was hypothesized that the oSDN corresponds withhuman INAH-3, which suggests that the relevant neuroanatomicalpathways are conserved between mammalian species.
6.3.2. The role of prenatal androgensOne of the main hypotheses on the determinants of sexual ori-
entation was that same-sex attraction was the result of atypicalsex-hormone levels during gestation. Studies in rodents and ferretsshowed that pre- or perinatal hormonal manipulation could lead tochanges in partner preference, sexual behavior, and coital perfor-mance largely controlled by the hypothalamus [92,306]. Yet,extending this hypothesis from animal research to humans is diffi-cult in our opinion. Atypical sexual behavior in rodents is hard toequate to human sexuality. For example, the induction of lordosisin male rats does not change their partner preference. Instead, whatchanges is the rat’s entire sexual behavior, which is different fromsexual orientation. Rather, an animal that consistently choosessame-sex partners—such as the above-mentioned ram whose adulthormone levels are within the male-typical range [268]—would bea better model. Furthermore, the treatment necessary to changethe sexual behavior of rodents goes far beyond any naturally occur-ring variation in androgen levels [247], and as such is unlikely toreflect natural causes of human variation in sexual orientation.

T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 239
Additionally, hormonal manipulations have failed to make maleanimals mount other males.
Case studies on humans with various genetic defects in theandrogen pathway show only limited support for the hypothesis.There are no reports showing an increase in attraction to men inhypovirilized XY individuals relative to the general population.This implies that disruption of the androgen pathway does nothave a strong effect on male sexual orientation. The role of andro-gens in female sexual orientation appears more complex. Womenwith congenital adrenal hyperplasia (CAH) experience abnormalactivity of the embryonic adrenal glands. This leads to a muchhigher exposure of female fetuses to androgens, greatly exceedingfemale-typical levels. The exposure is often high enough to causesome degree of genital masculinization. Several studies have foundthat CAH women reported more same-sex sexual activity and thatmore self-identified as homosexual compared to the general popu-lation, which suggests that typical female sexual development isdisrupted by extreme prenatal androgen exposure [149]. It isimportant to note that while women with CAH reported more gen-der atypical attitudes, interests, and behavior, the majority stillidentified as heterosexual. The role of androgens in the sexual ori-entation of lesbian women who have no genital masculinization isstill unclear.
Two studies looked at genetic variation in genes related to thesteroid pathway. A candidate gene study on the human androgenreceptor gene [202] and one on the aromatase gene (CYP19) [94]found no evidence that variations in these genes play a role invariations in human sexual orientation. A variety of anthropomor-phic measures have been used as indirect measures of prenatalandrogen exposure, but results have been inconsistent. A recentprospective study showed no correlation between maternal circu-lating androgen concentration at 18 and 34 weeks of gestationand digit ratio in girls [145]. An in-depth discussion of these stud-ies and a speculation on their widely varying results falls outsideof the scope of this manuscript. For a review on the often cited2D:4D finger-length ratio in sexual orientation, see McFaddenet al. [216]. Finally, studies retrospectively examining the influ-ence of stressful events during pregnancy have been inconclusive[23,99]. Therefore we believe that there is little evidence that nat-urally occurring variations of prenatal circulating gonadal hor-mones within one sex play a role in determining variants ofsexual orientation although diverging views have been expressedon this topic.
6.3.3. The genetics of sexual orientationEvidence is mounting that there is a strong genetic component
influencing sexual orientation. Family studies [249,21,243,24] havefound an increased rate of homosexuality among siblings and inthe maternal uncles of gay men (a median rate of 9% for brothersof gay men) [22]. Although the concordance rates of homosexualityin monozygotic twins vary depending on ascertainment methods[21,25,174,180], twin studies have found that there is a substantialgenetic component in the development of sexual orientation.
There has been limited molecular genetics research. In 1993,Hamer et al. reported that male homosexuality was more oftenon the mother’s side of the family versus the father’s side [133].A linkage scan showed significant linkage of male homosexualityto the X-chromosome region Xq28 [133]. This finding was subse-quently replicated by two studies [154,279] but not by an indepen-dent group [264]. However, a meta-analysis of the results across allfour studies yielded an estimated level of Xq28 allele sharing be-tween gay brothers of 64% instead of the expected 50% [132]. Nev-ertheless, the exact gene(s) involved has (have) yet to be identified.
A different method also implicated the role of the X-chromo-some. Unlike male cells, female cells contain two X-chromosomes.Consequently, each female cell randomly inactivates one X-chro-
mosome during embryogenesis to create dosage compensation:The inactive chromosome remains inactive in all resulting daugh-ter cells [54]. If inactivation is completely random, this means thatin a population of female cells the maternal X will be inactivated in50% of the cells whereas the paternal X is inactive in the remaininghalf. If a particular X chromosome, whether maternal or paternal, isinactivated in more than 90% of cells, that individual is consideredto be extremely skewed in regards to X-inactivation. Mothers withgay sons were found to have extreme skewing of X-inactivationwhen compared to mothers with no gay sons: Skewing in motherswith one gay son = 13/97 or 13%; skewing in mothers with two ormore gay sons = 10/44 or 23% [47]. This suggested an involvementof the X chromosome in the molecular mechanisms of sexual ori-entation. Arguably, the effect of the X-chromosome gene(s) ormechanisms that influence sexual orientation in the sons is visiblein the blood of their mothers.
A genome-wide linkage scan on gay-brother pairs showed sug-gestive linkage to loci on chromosome 7 and 8 [230]. A maternal ori-gin effect was found near marker D10S217, located at 10q26, withsignificant linkage for maternal meioses but no paternal contribu-tion. This result suggested the presence of a maternally expressed,paternally-silenced imprinted gene for sexual orientation in10q26. The relatively small sample size (N = 456) likely underpow-ered this study. However, a larger linkage scan on 1000 homosexualmale sibling pairs is currently underway (A.R. Sanders, personalcommunication).
The presence of a possible imprinted gene on chromosome 10 isparticularly interesting. Previously reported evidence of maternalloading of sexual orientation transmission was initially used toimplicate the X-chromosome in human sexual orientation, but itcould just as well indicate epigenetic factors acting on autosomalgenes. A role for imprinted genes in human sexual orientationwas hypothesized earlier [46].
One of the most replicated findings in sexual orientation re-search is known as the ‘fraternal birth order effect’: Each olderbrother increases the odds of male homosexuality by approxi-mately 33% [41,166]. This is relative to the baseline frequency ofhomosexuality, and the odds of being homosexual are about twiceas high for the fourth-born son relative to the first-born son. Yet,this finding is not so simple: The effect is only influenced by olderbrothers born via the same mother, it is not influenced by the num-ber of older sisters, and it only seems to be true for right-handedhomosexual men [44]. The dominant hypothesis for this effect,which lacks empirical support, is that each successive male preg-nancies increases the mother’s immunity against male-specificantigens expressed by the fetus [42,43], and this immune responseaffects any subsequent male fetuses. Although this phenomenondoes not directly implicate genetics, at the very least it demon-strates a biological basis for human male sexual orientation andsuggests the immune system as an alternative, gonadal hormone-independent mechanism through which sex differences can bemediated.
Altogether, there is mounting evidence for a genetic role of hu-man sexual orientation. The overwhelming dominance of hetero-sexual behavior in the animal kingdom points at a tightmolecular regulation of this trait.
6.4. Gender identity
Gender identity, or our sense of maleness or femaleness, playsan important role throughout our development affecting bothour sense of self and our relationships [278,277,282]. Our genderidentity and the roles ascribed to that gender are heavily influ-enced by social factors [e.g. 226]. Most people adopt a gender iden-tity congruent with the sex assigned at birth, which remainsconstant throughout life [339]. The best approach to study the

240 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
biological basis of gender identity is to study individuals who de-velop a cross-gender identity—in particular transsexuals.
Zhou et al. were the first to describe a sex difference in the cen-tral subdivision bed nucleus of the stria terminalis (BSTc) in hu-mans and a potential biological marker for gender identity [350].The type and direction of the sex difference mirrored that of therat: the volume of the BSTc is larger in men than in women. Thestudy also found that the BSTc of male-to-female (MtF) transsexu-als is female-sized but the interpretation of this finding is compli-cated. The MtF subjects used in the study had all received estrogentherapy so it remains unclear if the sex difference is related to gen-der identity or hormonal exposure since estrogens can modify thestructure of the brain. A second confound is the relatively smallsize of the sample pool as the authors were only able to gain accessto tissue from six MtF transsexuals.
The literature on the genetic basis of transsexualism is extre-mely limited. Although there are reports of families where severalmembers identify as transsexuals [119], such reports are rare.There are few twin case studies, and they have reported differingconcordance rates for transsexualism [284,111,157,276]. Since nosystematic twin study has been reported, it is impossible to sepa-rate genetic from environmental influences. Consequently, there isno clear support for a genetic basis of transsexualism at this point.
A number of chromosomal abnormalities have been reported intranssexuals [127,55,313,302]. In all cases, sex chromosomes wereinvolved. The most common association was with disomy-Y(47,XYY). However, because of the relatively high frequency ofsex chromosome aneuploidy (1 in 900 males for XYY; [232] a sta-tistically significant association with transsexualism has not beenshown.
A small number of candidate genes have been studied for trans-sexualism. A recent study looked at a polymorphism in the genecoding for 5-alpha reductase and found no association in a sampleof MtF and female-to-male (FtM) transsexuals [36]. The samegroup found a significant association between a single nucleotidepolymorphism in the CYP17 gene (which encodes the 17a-hydrox-ylase enzyme) in FtM transsexuals but not MtF transsexuals [37].However, their sample size was small and they reported a signifi-cant difference in allele distribution between male and female con-trols as well, shedding doubts on these results.
A small study of MtF transsexuals in a Swedish population stud-ied repeat sequences in or near the androgen receptor gene, theestrogen receptor beta gene, and the aromatase gene. They foundan association between MtF transsexualism and a dinucleotideCA polymorphism in the estrogen receptor beta (ERb) gene [142].However, a larger study of MtF transsexuals failed to replicatethe ERb results and instead found a significant association withthe androgen receptor repeat [135]. This, too, was not replicatedby a subsequent research team [319]. Overall, the significance ofthese genetics studies remains unclear.
7. Conclusion
There are many differences between men and women. In thisreview, we have focused on brain sex differences because of therole that they play in people’s health and behavior. Historically,it was believed that such differences were solely due to gonadalhormone secretions. Yet, emerging research is also implicating di-rect genetic effects.
The next challenge will be to first elucidate the molecular mech-anisms by which these direct genetic effects on sex differences arise.One way to address this question will be to manipulate gene dosagein specific tissues and at specific times of development by using tar-geted approaches in genetically modified animals. Another criticalissue will be to understand how compensation mechanisms be-tween the sexes operate. de Vries proposed that observed molecular
sex differences may compensate for other sex differences ratherthan generating differences between males and females themselves[80]. It will be crucial to tease out whether small imbalances influ-ence specific differences in neuropsychological function. Finally, itwill be essential to translate our knowledge of sex differences toimprove the quality of medical and psychological care.
As science continues to advance our understanding of sex dif-ferences, a new field is emerging focused on better addressingthe needs of men and women: gender-based biology and medicine.The ultimate aim of this field is to translate scientific data intopractical applications that are effective for each sex [158]. Fromtailoring preventive screenings to treating sex-specific illnesses,this field recognizes that ‘‘one-size-fits-all” healthcare has its lim-its. Rather, our biological sex is an important variable that must beconsidered in our mental and physical health.
Acknowledgment
This work was funded in part by the National Institutes ofHealth (1R01MH075046).
References
[1] M.E. Addis, A.K. Mansfield, M.R. Syzdek, Is ‘‘masculinity” a problem?: framingthe effects of gendered social learning in men, Psychol. Men Masc. 11 (2010)77–90.
[2] R.J. Agate, W. Grisham, J. Wade, S. Mann, J. Wingfield, C. Schanen, A. Palotie,A.P. Arnold, Neural, not gonadal, origin of brain sex differences in agynandromorphic finch, Proc. Natl. Acad. Sci. USA 100 (2003) 4873–4878.
[3] L.S. Allen, R.A. Gorski, Sexual orientation and the size of the anteriorcommissure in the human brain, Proc. Natl. Acad. Sci. USA 89 (1992) 7199–7202.
[4] L.S. Allen, M. Hines, J.E. Shryne, R.A. Gorski, Two sexually dimorphic cellgroups in the human brain, J. Neurosci. 9 (1989) 497–506.
[5] C.R. Almli, R.H. Ball, M.E. Wheeler, Human fetal and neonatal movementpatterns: gender differences and fetal-to-neonatal continuity, Dev.Psychobiol. 38 (2001) 252–273.
[6] M. Altemus, Sex differences in depression and anxiety disorders: potentialbiological determinants, Horm. Behav. 50 (2006) 534–538.
[7] K. Andersen, L.J. Launer, M.E. Dewey, L. Letenneur, A. Ott, J.R. Copeland, J.F.Dartigues, P. Kragh-Sorensen, M. Baldereschi, C. Brayne, A. Lobo, J.M.Martinez-Lage, T. Stijnen, A. Hofman, Gender differences in the incidence ofAD and vascular dementia: the EURODEM studies. EURODEM incidenceresearch group, Neurology 53 (1999) 1992–1997.
[8] J. Archer, N. Graham-Kevan, M. Davies, Testosterone and aggression: areanalysis of book, Starzyk and Quinsey’s (2001) study, Aggress. ViolentBehav. 10 (2005) 241–261.
[9] A.P. Arnold, The effects of castration on song development in zebra finches(Poephila guttata), J. Exp. Zool. 191 (1975) 261–278.
[10] A.P. Arnold, Genetically triggered sexual differentiation of brain and behavior,Horm. Behav. 30 (1996) 495–505.
[11] A.P. Arnold, Sexual differentiation of the zebra finch song system: positiveevidence, negative evidence, null hypotheses, and a paradigm shift, J.Neurobiol. 33 (1997) 572–584.
[12] A.P. Arnold, S.M. Breedlove, Organizational and activational effects of sexsteroids on brain and behavior: a reanalysis, Horm. Behav. 19 (1985) 469–498.
[13] A.P. Arnold, P.S. Burgoyne, Are XX and XY brain cells intrinsically different?,Trends Endocrinol Metab. 15 (2004) 6–11.
[14] A.P. Arnold, X. Chen, What does the ‘‘four core genotypes” mouse model tellus about sex differences in the brain and other tissues?, FrontNeuroendocrinol. 30 (2009) 1–9.
[15] A.P. Arnold, X. Chen, What does the ‘‘four core genotypes” mouse model tellus about sex differences in the brain and other tissues?, FrontNeuroendocrinol. 30 (2009) 1–9.
[16] A.P. Arnold, J. Wade, W. Grisham, E.C. Jacobs, A.T. Campagnoni, Sexualdifferentiation of the brain in songbirds, Dev. Neurosci. 18 (1996) 124–136.
[17] B. Auyeung, S. Baron-Cohen, E. Ashwin, R. Knickmeyer, K. Taylor, G. Hackett,M. Hines, Fetal testosterone predicts sexually differentiated childhoodbehavior in girls and in boys, Psychol. Sci. 20 (2009) 144–148.
[18] S. Avissar, Y. Egozi, M. Sokolovsky, Studies on muscarinic receptors in mouseand rat hypothalamus: a comparison of sex and cyclical differences,Neuroendocrinology 32 (1981) 295–302.
[19] Y. Baba, J.D. Putzke, N.R. Whaley, Z.K. Wszolek, R.J. Uitti, Gender and theParkinson’s disease phenotype, J. Neurol. 252 (2005) 1201–1205.
[20] D. Badre, A.D. Wagner, Semantic retrieval, mnemonic control, and prefrontalcortex, Behav. Cogn. Neurosci. Rev. 1 (2002) 206–218.
[21] J.M. Bailey, R.C. Pillard, A genetic study of male sexual orientation, Arch. Gen.Psychiatry 48 (1991) 1089–1096.

T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 241
[22] J.M. Bailey, R.C. Pillard, Genetics of human sexual orientation, Annu. Rev. SexRes. (1995) 126–150.
[23] J.M. Bailey, L. Willerman, C. Parks, A test of the maternal stress theory ofhuman male homosexuality, Arch. Sex. Behav. 20 (1991) 277–293.
[24] J.M. Bailey, R.C. Pillard, K. Dawood, M.B. Miller, L.A. Farrer, S. Trivedi, R.L.Murphy, A family history study of male sexual orientation using threeindependent samples, Behav. Genet. 29 (1999) 79–86.
[25] J.M. Bailey, M.P. Dunne, N.G. Martin, Genetic and environmental influences onsexual orientation and its correlates in an Australian twin sample, J. Pers. Soc.Psychol. 78 (2000) 524–536.
[26] J. Balthazart, G.F. Ball, New insights into the regulation and function of brainestrogen synthase (aromatase), Trends Neurosci. 21 (1998) 243–249.
[27] C.S. Barr, T.K. Newman, M. Schwandt, C. Shannon, R.L. Dvoskin, S.G. Lindell, J.Taubman, B. Thompson, M. Champoux, K.P. Lesch, D. Goldman, S.J. Suomi, J.D.Higley, Sexual dichotomy of an interaction between early adversity and theserotonin transporter gene promoter variant in rhesus macaques, Proc. Natl.Acad. Sci. USA 101 (2004) 12358–12363.
[28] J.B. Becker, Sex Differences in the Brain: From Genes to Behavior, OxfordUniversity Press, Oxford; New York, 2008.
[29] J.B. Becker, Sex Differences in the Brain: From Genes to Behavior, OxfordUniversity Press, Oxford; New York, 2008. p. 427.
[30] J.B. Becker, Sex Differences in the Brain: From Genes to Behavior, OxfordUniversity Press, Oxford; New York, 2008. pp. 442–445.
[31] J.B. Becker, Sex Differences in the Brain: From Genes to Behavior, OxfordUniversity Press, Oxford; New York, 2008. p. 458.
[32] C. Beltramino, S. Taleisnik, Dual action of electrochemical stimulation of thebed nucleus of the stria terminalis on the release of LH, Neuroendocrinology30 (1980) 238–242.
[33] D. Ben Shalom, D. Poeppel, Functional anatomic models of language:assembling the pieces, Neuroscientist 14 (2008) 119–127.
[34] C.P. Benbow, D. Lubinski, D.L. Shea, H. Eftekhari-Sanjani, Sex differences inmathematical reasoning ability at age 13: their status 20 years later, Psychol.Sci. 11 (2000) 474–480.
[35] M.D. Benedetti, D.M. Maraganore, J.H. Bower, S.K. McDonnell, B.J. Peterson,J.E. Ahlskog, D.J. Schaid, W.A. Rocca, Hysterectomy, menopause, and estrogenuse preceding Parkinson’s disease: an exploratory case-control study, Mov.Disord. 16 (2001) 830–837.
[36] E.K. Bentz, C. Schneeberger, L.A. Hefler, M. van Trotsenburg, U. Kaufmann, J.C.Huber, C.B. Tempfer, A common polymorphism of the SRD5A2 gene andtranssexualism, Reprod. Sci. 14 (2007) 705–709.
[37] E.K. Bentz, L.A. Hefler, U. Kaufmann, J.C. Huber, A. Kolbus, C.B. Tempfer, Apolymorphism of the CYP17 gene related to sex steroid metabolism isassociated with female-to-male but not male-to-female transsexualism,Fertil. Steril. 90 (2008) 56–59.
[38] S.A. Berenbaum, S.C. Duck, K. Bryk, Behavioral effects of prenatal versuspostnatal androgen excess in children with 21-hydroxylase-deficientcongenital adrenal hyperplasia, J. Clin. Endocrinol. Metab. 85 (2000) 727–733.
[39] M. Berger, J.A. Gray, B.L. Roth, The expanded biology of serotonin, Annu. Rev.Med. 60 (2009) 355–366.
[40] B.A. Bettencourt, N. Miller, Gender differences in aggression as a function ofprovocation: a meta-analysis, Psychol. Bull. 119 (1996) 422–447.
[41] R. Blanchard, Birth order and sibling sex ration in homosexual versusheterosexual males and females, Annu. Rev. Sex Res. 8 (1997) 27–67.
[42] R. Blanchard, A.F. Bogaert, Homosexuality in men and number of olderbrothers, Am. J. Psychiatry 153 (1996) 27–31.
[43] R. Blanchard, P. Klassen, HY antigen and homosexuality in men, J. Theor. Biol.185 (1997) 373–378.
[44] R. Blanchard, J.M. Cantor, A.F. Bogaert, S.M. Breedlove, L. Ellis, Interaction offraternal birth order and handedness in the development of malehomosexuality, Horm. Behav. 49 (2006) 405–414.
[45] R. Bleier, W. Byne, I. Siggelkow, Cytoarchitectonic sexual dimorphisms of themedial preoptic and anterior hypothalamic areas in guinea pig, rat, hamster,and mouse, J. Comp. Neurol. 212 (1982) 118–130.
[46] S. Bocklandt, D.H. Hamer, Beyond hormones: a novel hypothesis for thebiological basis of male sexual orientation, J. Endocrinol. Invest. 26 (2003) 8–12.
[47] S. Bocklandt, S. Horvath, E. Vilain, D.H. Hamer, Extreme skewing of Xchromosome inactivation in mothers of homosexual men, Hum. Genet. 118(2006) 691–694.
[48] L. Booij, A.J. Van der Does, W.J. Riedel, Monoamine depletion in psychiatricand healthy populations: review, Mol. Psychiatry 8 (2003) 951–973.
[49] A.S. Book, K.B. Starzyk, V.L. Quinsey, The relationship between testosteroneand aggression: a meta-analysis, Aggress. Violent Behav. 6 (2001) 579–599.
[50] S.T. Bradford, R. Hiramatsu, M.P. Maddugoda, P. Bernard, M.C. Chaboissier, A.Sinclair, A. Schedl, V. Harley, Y. Kanai, P. Koopman, D. Wilhelm, The cerebellin4 precursor gene is a direct target of SRY and SOX9 in mice, Biol. Reprod. 80(2009) 1178–1188.
[51] K.T. Brady, C.L. Randall, Gender differences in substance use disorders,Psychiatr. Clin. North Am. 22 (1999) 241–252.
[52] N. Breslau, L. Schultz, E. Peterson, Sex differences in depression: a role forpreexisting anxiety, Psychiatry Res. 58 (1995) 1–12.
[53] G.R. Brooks, Despite problems, ‘‘masculinity” is a vital construct, Psychol.Men Masc. 11 (2010) 107–108.
[54] C.J. Brown, W.P. Robinson, The causes and consequences of random and non-random X chromosome inactivation in humans, Clin. Genet. 58 (2000)353–363.
[55] N. Buhrich, R. Barr, P.R. Lam-Po-Tang, Two transsexuals with 47-XYYkaryotype, Br. J. Psychiatry 133 (1978) 77–81.
[56] P.S. Burgoyne, M. Buehr, A. McLaren, XY follicle cells in ovaries of XX–XYfemale mouse chimaeras, Development 104 (1988) 683–688.
[57] W. Byne, S. Tobet, L. Mattiace, M.S. Lasco, E. Kemether, M.A. Edgar, S.Morgello, M.S. Buchsbaum, L.B. Jones, The interstitial nuclei of the humananterior hypothalamus: an investigation of variation within sex, sexualorientation and HIV status, Horm. Behav. 40 (2001) 86–92.
[58] D.J. Callen, S.E. Black, C.B. Caldwell, C.L. Grady, The influence of sex on limbicvolume and perfusion in AD, Neurobiol. Aging 25 (2004) 761–770.
[59] L. Carrel, A.A. Cottle, K.C. Goglin, H.F. Willard, A first-generation X-inactivation profile of the human X chromosome, Proc. Natl. Acad. Sci. USA96 (1999) 14440–14444.
[60] L.L. Carruth, I. Reisert, A.P. Arnold, Sex chromosome genes directly affect brainsexual differentiation, Nat. Neurosci. 5 (2002) 933–934.
[61] E. Cashdan, Hormones and competitive aggression in women, Aggress. Behav.29 (2003) 107–115.
[62] A. Caspi, J. McClay, T.E. Moffitt, J. Mill, J. Martin, I.W. Craig, A. Taylor, R.Poulton, Role of genotype in the cycle of violence in maltreated children,Science 297 (2002) 851–854.
[63] A. Caspi, K. Sugden, T.E. Moffitt, A. Taylor, I.W. Craig, H. Harrington, J. McClay,J. Mill, J. Martin, A. Braithwaite, R. Poulton, Influence of life stress ondepression: moderation by a polymorphism in the 5-HTT gene, Science 301(2003) 386–389.
[64] D. Chadwick, J. Goode, The Genetics and Biology of Sex Determination, J.Wiley, Chichester, West Sussex, England, New York, NY, 2002.
[65] P.K.Y. Chow, S.T. Cheng, Shame, internalized heterosexism, lesbian identity,and coming out to others: a comparative study of lesbians in mainland Chinaand Hong Kong, J. Couns. Psychol. 57 (2010) 92–104.
[66] P. Ciofi, D. Leroy, G. Tramu, Sexual dimorphism in the organization of the rathypothalamic infundibular area, Neuroscience 141 (2006) 1731–1745.
[67] P. Ciofi, O.C. Lapirot, G. Tramu, An androgen-dependent sexual dimorphismvisible at puberty in the rat hypothalamus, Neuroscience 146 (2007) 630–642.
[68] C. Clépet, A.J. Schafer, A.H. Sinclair, M.S. Palmer, R. Lovell-Badge, P.N.Goodfellow, The human SRY transcript, Hum. Mol. Genet. 2 (1993) 2007–2012.
[69] A.M. Cohn, A. Zeicher, A.L. Seibert, Labile affect as a risk factor for aggressivebehavior in men, Psychol. Men Masc. 9 (2008) 29–39.
[70] L.A. Colangelo, L. Sharp, P. Kopp, D. Scholtens, B.C. Chiu, K. Liu, S.M. Gapstur,Total testosterone, androgen receptor polymorphism, and depressivesymptoms in young black and white men: the CARDIA male hormonestudy, Psychoneuroendocrinology 32 (2007) 951–958.
[71] A. Colciago, F. Celotti, A. Pravettoni, O. Mornati, L. Martini, P. Negri-Cesi,Dimorphic expression of testosterone metabolizing enzymes in thehypothalamic area of developing rats, Dev. Brain Res. 155 (2005) 107–116.
[72] J.C. Confers, J.A. Easton, D.S. Fleischman, C.D. Goetz, D.M. Lewis, C. Perilloux,D.M. Buss, Evolutionary psychology: controversies, questions, prospects, andlimitations, Am. Psychol. 65 (2010) 110–126.
[73] B.M. Cooke, G. Tabibnia, S.M. Breedlove, A brain sexual dimorphismcontrolled by adult circulating androgens, Proc. Natl. Acad. Sci. USA 96(1999) 7538–7540.
[74] D.W. Cooper, The evolution of sex determination, sex chromosomedimorphism, and X-inactivation in Therian mammals: a comparison ofmetatherians (marsupials) and eutherians (‘‘placentals”), in: K. Reed, J.A.M.Graves (Eds.), Sex Chromosomes and Sex-Determining Genes, HarwoodAcademic Publishers, Chur, Switzerland, Langhorne, PA, 1993, pp. 183–200.
[75] E.H. Corder, A.M. Saunders, W.J. Strittmatter, D.E. Schmechel, P.C. Gaskell,G.W. Small, A.D. Roses, J.L. Haines, M.A. Pericak-Vance, Gene dose ofapolipoprotein E type 4 allele and the risk of Alzheimer’s disease in lateonset families, Science 261 (1993) 921–923.
[76] R.M. Craft, Sex differences in opioid analgesia: ‘‘from mouse to man”, Clin. J.Pain 19 (2003) 175–186.
[77] J.A. Dani, D. Bertrand, Nicotinic acetylcholine receptors and nicotiniccholinergic mechanisms of the central nervous system, Annu. Rev.Pharmacol. Toxicol. 47 (2007) 699–729.
[78] P.L. Davies, J.D. Rose, Motor skills of typically developing adolescents:awkwardness or improvement?, Phys Occup. Ther. Pediatr. 20 (2000) 19–42.
[79] L.M. de Lau, P.C. Giesbergen, M.C. de Rijk, A. Hofman, P.J. Koudstaal, M.M.Breteler, Incidence of Parkinsonism and Parkinson disease in a generalpopulation: the Rotterdam study, Neurology 63 (2004) 1240–1244.
[80] G.J. De Vries, Minireview: sex differences in adult and developing brains:compensation, compensation, compensation, Endocrinology 145 (2004)1063–1068.
[81] G.J. De Vries, P.A. Boyle, Double duty for sex differences in the brain, Behav.Brain Res. 92 (1998) 205–213.
[82] G.J. de Vries, P. Sodersten, Sex differences in the brain: the relation betweenstructure and function, Horm. Behav. 55 (2009) 589–596.
[83] G.J. De Vries, Z. Wang, N.A. Bullock, S. Numan, Sex differences in theeffects of testosterone and its metabolites on vasopressin messenger RNAlevels in the bed nucleus of the stria terminalis of rats, J. Neurosci. 14(1994) 1789–1794.
[84] G.J. De Vries, E.F. Rissman, R.B. Simerly, L.Y. Yang, E.M. Scordalakes, C.J. Auger,A. Swain, R. Lovell-Badge, P.S. Burgoyne, A.P. Arnold, A model system forstudy of sex chromosome effects on sexually dimorphic neural andbehavioral traits, J. Neurosci. 22 (2002) 9005–9014.

242 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
[85] A. del Abril, S. Segovia, A. Guillamon, The bed nucleus of the stria terminalis inthe rat: regional sex differences controlled by gonadal steroids early afterbirth, Brain Res. 429 (1987) 295–300.
[86] P. Dewing, C.W. Chiang, K. Sinchak, H. Sim, P.O. Fernagut, S. Kelly, M.F.Chesselet, P.E. Micevych, K.H. Albrecht, V.R. Harley, E. Vilain, Direct regulationof adult brain function by the male-specific factor SRY, Curr. Biol. 16 (2006)415–420.
[87] D.A. Di Monte, M. Lavasani, A.B. Manning-Bog, Environmental factors inParkinson’s disease, Neurotoxicology 23 (2002) 487–502.
[88] T. Di Paolo, Modulation of brain dopamine transmission by sex steroids, Rev.Neurosci. 5 (1994) 27–41.
[89] M. Diamond, Homosexuality and bisexuality in different populations, Arch.Sex. Behav. 22 (1993) 291–310.
[90] M.C. Diamond, G.A. Dowling, R.E. Johnson, Morphologic cerebral corticalasymmetry in male and female rats, Exp. Neurol. 71 (1981) 261–268.
[91] F. Dittrich, Y. Feng, R. Metzdorf, M. Gahr, Estrogen-inducible, sex-specificexpression of brain-derived neurotrophic factor mRNA in a forebrain songcontrol nucleus of the juvenile zebra finch, Proc. Natl. Acad. Sci. USA 96(1999) 8241–8246.
[92] E. Dominguez-Salazar, W. Portillo, M.J. Baum, J. Bakker, R.G. Paredes, Effect ofprenatal androgen receptor antagonist or aromatase inhibitor on sexualbehavior, partner preference and neuronal Fos responses to estrous femaleodors in the rat accessory olfactory system, Physiol. Behav. 75 (2002) 337–346.
[93] R.A. Dubin, H. Ostrer, Sry is a transcriptional activator, Mol. Endocrinol. 8(1994) 1182–1192.
[94] M.G. DuPree, B.S. Mustanski, S. Bocklandt, C. Nievergelt, D.H. Hamer, Acandidate gene study of CYP19 (aromatase) and male sexual orientation,Behav. Genet. 34 (2004) 243–250.
[95] M.F. Egan, M. Kojima, J.H. Callicott, T.E. Goldberg, B.S. Kolachana, A. Bertolino,E. Zaitsev, B. Gold, D. Goldman, M. Dean, B. Lu, D.R. Weinberger, The BDNFval66met polymorphism affects activity-dependent secretion of BDNF andhuman memory and hippocampal function, Cell 112 (2003) 257–269.
[96] Y. Egozi, S. Avissar, M. Sokolovsky, Muscarinic mechanisms and sex hormonesecretion in rat adenohypophysis and preoptic area, Neuroendocrinology 35(1982) 93–97.
[97] E.M. Eicher, L.L. Washburn, J.B. Whitney 3rd, K.E. Morrow, Mus poschiavinusY chromosome in the C57BL/6J murine genome causes sex reversal, Science217 (1982) 535–537.
[98] G. Einstein, Sex and the Brain, MIT Press, Cambridge, Mass., 2007.[99] L. Ellis, S. Cole-Harding, The effects of prenatal stress, and of prenatal alcohol
and nicotine exposure, on human sexual orientation, Physiol. Behav. 74(2001) 213–226.
[100] D.E. Emery, B.D. Sachs, Copulatory behavior in male rats with lesions in thebed nucleus of the stria terminalis, Physiol. Behav. 17 (1976) 803–806.
[101] I.V. Estabrooke, K. Mordecai, P. Maki, M.T. Ullman, The effect of sex hormoneson language processing, Brain Lang. 83 (2002) 143–146.
[102] A.M. Etgen, J.C. Morales, Somatosensory stimuli evoke norepinephrine releasein the anterior ventromedial hypothalamus of sexually receptive female rats,J. Neuroendocrinol. 14 (2002) 213–218.
[103] J. Feng, G. Sun, J. Yan, K. Noltner, W. Li, C.H. Buzin, J. Longmate, L.L. Heston, J.Rossi, S.S. Sommer, Evidence for X-chromosomal schizophrenia associatedwith microRNA alterations, PLoS ONE 4 (2009) e6121.
[104] H.H. Fernandez, K.L. Lapane, B.R. Ott, J.H. Friedman, Gender differences in thefrequency and treatment of behavior problems in Parkinson’s disease. SAGEstudy group. Systematic assessment and geriatric drug use via epidemiology,Mov. Disord. 15 (2000) 490–496.
[105] R.H. Fitch, A.S. Berrebi, P.E. Cowell, L.M. Schrott, V.H. Denenberg, Corpuscallosum: effects of neonatal hormones on sexual dimorphism in the rat,Brain Res. 515 (1990) 111–116.
[106] R.H. Fitch, P.E. Cowell, L.M. Schrott, V.H. Denenberg, Corpus callosum: ovarianhormones and feminization, Brain Res. 542 (1991) 313–317.
[107] A. Fleming, E. Vilain, The endless quest for sex determination genes, Clin.Genet. 67 (2005) 15–25.
[108] T. Foltynie, S.G. Lewis, T.E. Goldberg, A.D. Blackwell, B.S. Kolachana, D.R.Weinberger, T.W. Robbins, R.A. Barker, The BDNF Val66Met polymorphismhas a gender specific influence on planning ability in Parkinson’s disease, J.Neurol. 252 (2005) 833–838.
[109] N.G. Forger, G.J. Rosen, E.M. Waters, D. Jacob, R.B. Simerly, G.J. de Vries,Deletion of Bax eliminates sex differences in the mouse forebrain, Proc. Natl.Acad. Sci. USA 101 (2004) 13666–13671.
[110] S. Gao, H.C. Hendrie, K.S. Hall, S. Hui, The relationships between age, sex, andthe incidence of dementia and Alzheimer disease: a meta-analysis, Arch. Gen.Psychiatry 55 (1998) 809–815.
[111] G.M. Garden, D.J. Rothery, A female monozygotic twin pair discordant fortranssexualism. Some theoretical implications, Br. J. Psychiatry 161 (1992)852–854.
[112] L.H. Gargaglioni, K.C. Bícego, L.G.S. Branco, Brain monoaminergic neurons andventilatory control in vertebrates, Resp. Physiol. Neurobiol. 164 (2008) 112–122.
[113] J.D. Gatewood, A. Wills, S. Shetty, J. Xu, A.P. Arnold, P.S. Burgoyne, E.F.Rissman, Sex chromosome complement and gonadal sex influence aggressiveand parental behaviors in mice, J. Neurosci. 26 (2006) 2335–2342.
[114] W.R. Gibb, Neuropathology of Parkinson’s disease and related syndromes,Neurol. Clin. 10 (1992) 361–376.
[115] J.M. Goldstein, Sex, hormones and affective arousal circuitry dysfunction inschizophrenia, Horm. Behav. 50 (2006) 612–622.
[116] J.M. Goldstein, S.V. Faraone, W.J. Chen, M.T. Tsuang, Genetic heterogeneitymay in part explain sex differences in the familial risk for schizophrenia, Biol.Psychiatry 38 (1995) 808–813.
[117] C.D. Good, I. Johnsrude, J. Ashburner, R.N.A. Henson, K.J. Friston, R.S.J.Frackowiak, Cerebral asymmetry and the effects of sex and handedness onbrain structure: a voxel-based morphometric analysis of 465 normal adulthuman brains, NeuroImage 14 (2001) 685–700.
[118] R.A. Gorski, R.E. Harlan, C.D. Jacobson, J.E. Shryne, A.M. Southam, Evidence forthe existence of a sexually dimorphic nucleus in the preoptic area of the rat, J.Comp. Neurol. 193 (1980) 529–539.
[119] R. Green, Family coocurrence of ‘‘gender dysphoria”: ten siblings or parent–child pairs, Arch. Sex. Behav. 29 (2000) 499–507.
[120] W.T. Greenough, C.S. Carter, C. Steerman, T.J. DeVoogd, Sex differences indendritic patterns in hamster preoptic area, Brain Res. 126 (1977) 63–72.
[121] S. Grigoriadis, G.E. Robinson, Gender issues in depression, Annu. Clin.Psychiatry 19 (2007) 247–255.
[122] G.M. Grimshaw, M.P. Bryden, J.A.K. Finegan, Relations between prenataltestosterone and cerebral lateralization in children, Neuropsychology 9(1995) 68–79.
[123] H.J. Groenewegen, The basal ganglia and motor control, Neural Plast. 10(2003) 107–120.
[124] J. Gubbay, P. Koopman, J. Collignon, P. Burgoyne, R. Lovell-Badge, Normalstructure and expression of Zfy genes in XY female mice mutant in Tdy,Development 109 (1990) 647–653.
[125] R.C. Gur, F. Gunning-Dixon, W.B. Bilker, R.E. Gur, Sex differences in temporo-limbic and frontal brain volumes of healthy adults, Cereb. Cortex 12 (2002)998–1003.
[126] M.E. Gurney, M. Konishi, Hormone-induced sexual differentiation of brainand behavior in zebra finches, Science 208 (1980) 1380–1383.
[127] M. Haberman, F. Hollingsworth, A. Falek, R.P. Michael, Gender identityconfusion, schizophrenia and a 47 XYY karyotype: a case report,Psychoneuroendocrinology 1 (1975) 207–209.
[128] A. Hacker, B. Capel, P. Goodfellow, R. Lovell-Badge, Expression of Sry, themouse sex determining gene, Development 121 (1995) 1603–1614.
[129] J.A. Hall, D. Kimura, Sexual orientation and performance on sexuallydimorphic motor tasks, Arch. Sex. Behav. 24 (1995) 395–407.
[130] D.F. Halpern, Sex Differences in Cognitive Abilities, Lawrence ErlbaumAssociates, 2000. pp. 159–163.
[131] D.F. Halpern, Sex Differences in Cognitive Abilities, Lawrence ErlbaumAssociates, 2000. pp. 147–148.
[132] D. Hamer, Genetics and male sexual orientation, Science 285 (1999) 803.[133] D. Hamer, S. Hu, V. Magnuson, N. Hu, A.M. Pattatucci, A linkage between DNA
markers on the X chromosome and male sexual orientation, Science 261(1993) 321–327.
[134] S.E. Hammack, K.J. Richey, L.R. Watkins, S.F. Maier, Chemical lesion of the bednucleus of the stria terminalis blocks the behavioral consequences ofuncontrollable stress, Behav. Neurosci. 118 (2004) 443–448.
[135] L. Hare, P. Bernard, F.J. Sanchez, P.N. Baird, E. Vilain, T. Kennedy, V.R. Harley,Androgen receptor repeat length polymorphism associated with male-to-female transsexualism, Biol. Psychiatry 65 (2009) 93–96.
[136] P.J. Harrison, E.M. Tunbridge, Catechol-O-methyltransferase (COMT): a genecontributing to sex differences in brain function, and to sexual dimorphism inthe predisposition to psychiatric disorders, Neuropsychopharmacology 33(2008) 3037–3045.
[137] R.A. Harshman, E. Hampson, S.A. Berenbaum, Individual differences incognitive abilities and brain organization. Part I: sex and handednessdifferences in ability, Can. J. Psychol. 37 (1983) 144–192.
[138] C.L. Hart, G.D. Smith, Alcohol consumption and mortality and hospitaladmissions in men from the Midspan collaborative cohort study, Addiction103 (2008) 1979–1986.
[139] J.K. Hartshorne, M.T. Ullman, Why girls say ‘holded’ more than boys, Dev. Sci.9 (2006) 21–32.
[140] M. Heinrichs, T. Baumgartner, C. Kirschbaum, U. Ehlert, Social support andoxytocin interact to suppress cortisol and subjective responses topsychosocial stress, Biol. Psychiatry 54 (2003) 1389–1398.
[141] A.C. Hennessey, K. Wallen, D.A. Edwards, Preoptic lesions increase the displayof lordosis by male rats, Brain Res. 370 (1986) 21–28.
[142] S. Henningsson, L. Westberg, S. Nilsson, B. Lundstrom, L. Ekselius, O. Bodlund,E. Lindstrom, M. Hellstrand, R. Rosmond, E. Eriksson, M. Landen, Sex steroid-related genes and male-to-female transsexualism, Psychoneuro-endocrinology 30 (2005) 657–664.
[143] G.M. Herek, J.R. Gillis, J.C. Cogan, Internalized stigma among sexual minorityadults: insights from a social psychological perspective, J. Couns. Psychol. 56(2009) 32–43.
[144] A. Herlitz, E. Airaksinen, E. Nordstrom, Sex differences in episodic memory: theimpact of verbal and visuospatial ability, Neuropsychology 13 (1999) 590–597.
[145] M. Hickey, D.A. Doherty, R. Hart, R.J. Norman, E. Mattes, H.C. Atkinson, D.M.Sloboda, Maternal and umbilical cord androgen concentrations do not predictdigit ratio (2D:4D) in girls: a prospective cohort study, Psychoneuro-endocrinology 35 (2010) 1235–1244.
[146] M. Hines, Brain Gender, Oxford University Press, Oxford; New York, 2004. pp.191–197.
[147] M. Hines, Brain Gender, Oxford University Press, Oxford; New York, 2004.[148] M. Hines, L.S. Allen, R.A. Gorski, Sex differences in subregions of the medial
nucleus of the amygdala and the bed nucleus of the stria terminalis of the rat,Brain Res. 579 (1992) 321–326.

T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 243
[149] M. Hines, C. Brook, G.S. Conway, Androgen and psychosexual development:core gender identity, sexual orientation and recalled childhood gender rolebehavior in women and men with congenital adrenal hyperplasia (CAH), J.Sex Res. 41 (2004) 75–81.
[150] C.C. Holloway, D.F. Clayton, Estrogen synthesis in the male brain triggersdevelopment of the avian song control pathway in vitro, Nat. Neurosci. 4(2001) 170–175.
[151] M.M. Holmes, B.D. Goldman, S.L. Goldman, M.L. Seney, N.G. Forger,Neuroendocrinology and sexual differentiation in eusocial mammals, Front.Neuroendocrinol. 30 (2009) 519–533.
[152] C.J. Hong, H.C. Liu, T.Y. Liu, C.H. Lin, C.Y. Cheng, S.J. Tsai, Brain-derivedneurotrophic factor (BDNF) Val66Met polymorphisms in Parkinson’s diseaseand age of onset, Neurosci. Lett. 353 (2003) 75–77.
[153] H. Hörtnagl, L. Hansen, G. Kindel, B. Schneider, A.E. Tamert, I. Hanin, Sexdifferences and estrous cycle-variations in the AF64A-induced cholinergicdeficit in the rat hippocampus, Brain Res. Bull. 31 (1993) 129–134.
[154] S. Hu, A. Pattatucci, C. Patterson, L. Li, D. Fulker, S. Cherny, L. Kruglyak, D.Hamer, Linkage between sexual orientation and chromosome Xq28 in malesbut not females, Nat. Genet. 11 (1995) 248–256.
[155] J.S. Hyde, The gender similarities hypothesis, Am. Psychol. 60 (2005) 581–592.
[156] J.S. Hyde, New directions in the study of gender similarities and differences,Curr. Dir. Psychol. Sci. 16 (2007) 259–263.
[157] C. Hyde, J.C. Kenna, A male MZ twin pair, concordant fortranssexualism, discordant for schizophrenia, Acta Psychiatr. Scand. 56(1977) 265–275.
[158] Institute of Medicine (US), Committee on understanding the biology of sexand gender differences, in: T.M. Wizemann, M.L. Pardue (Eds.), Exploring theBiological Contributions to Human Health: Does Sex Matter?, NationalAcademy Press, Washington, DC, 2001
[159] S. Ito, S. Murakami, K. Yamanouchi, Y. Arai, Prenatal androgen exposure,preoptic area and reproductive functions in the female rat, Brain Dev. 8(1986) 463–468.
[160] A. Jablensky, Epidemiology of schizophrenia: the global burden of disease anddisability, Eur. Arch. Psychiatry Clin. Neurosci. 250 (2000) 274–285.
[161] S. Jacob, C.W. Brune, C.S. Carter, B.L. Leventhal, C. Lord, E.H. Cook Jr.,Association of the oxytocin receptor gene (OXTR) in Caucasian children andadolescents with autism, Neurosci. Lett. 417 (2007) 6–9.
[162] E.C. Jacobs, W. Grisham, A.P. Arnold, Lack of a synergistic effect betweenestradiol and dihydrotestosterone in the masculinization of the zebra finchsong system, J. Neurobiol. 27 (1995) 513–519.
[163] S.m.R.L.o. Joca, F.R.r. Ferreira, F.S. Guimarães, Modulation of stressconsequences by hippocampal monoaminergic, glutamatergic and nitrergicneurotransmitter systems, Stress 10 (2007) 227–249.
[164] J.K. Johnson, R. McCleary, M.H. Oshita, C.W. Cotman, Initiation andpropagation stages of beta-amyloid are associated with distinctiveapolipoprotein E, age, and gender profiles, Brain Res. 798 (1998) 18–24.
[165] B.E. Jones, From waking to sleeping: neuronal and chemical substrates,Trends Pharmacol. Sci. 26 (2005) 578–586.
[166] M.B. Jones, R. Blanchard, Birth order and male homosexuality: extension ofSlater’s index, Hum. Biol. 70 (1998) 775–787.
[167] A. Jost, Hormonal factors in the sex differentiation of the mammalian foetus,Philos. Trans. R. Soc. B – Biol. Sci. 259 (1970) 119–130.
[168] A.J. Justice, H. de Wit, Acute effects of d-amphetamine during the follicularand luteal phases of the menstrual cycle in women, Psychopharmacology(Berlin) 145 (1999) 67–75.
[169] A.J. Justice, H. De Wit, Acute effects of d-amphetamine during the early andlate follicular phases of the menstrual cycle in women, Pharmacol. Biochem.Behav. 66 (2000) 509–515.
[170] S.P. Kalra, P.S. Kalra, Neural regulation of luteinizing hormone secretion in therat, Endocr. Rev. 4 (1983) 311–351.
[171] K. Kansaku, A. Yamaura, S. Kitazawa, Sex differences in lateralization revealedin the posterior language areas, Cereb. Cortex 10 (2000) 866–872.
[172] P. Kehoe, F. Wavrant-De Vrieze, R. Crook, W.S. Wu, P. Holmans, I. Fenton, G.Spurlock, N. Norton, H. Williams, N. Williams, S. Lovestone, J. Perez-Tur, M.Hutton, M.C. Chartier-Harlin, S. Shears, K. Roehl, J. Booth, W. Van Voorst, D.Ramic, J. Williams, A. Goate, J. Hardy, M.J. Owen, A full genome scan for lateonset Alzheimer’s disease, Hum. Mol. Genet. 8 (1999) 237–245.
[173] K.S. Kendler, M.C. Neale, R.C. Kessler, A.C. Heath, L.J. Eaves, A longitudinaltwin study of 1-year prevalence of major depression in women, Arch. Gen.Psychiatry 50 (1993) 843–852.
[174] K.S. Kendler, L.M. Thornton, S.E. Gilman, R.C. Kessler, Sexual orientation in aUS national sample of twin and nontwin sibling Pairs, Am. J. Psychiatry 157(2000) 1843–1846.
[175] G.A. Kennett, F. Chaouloff, M. Marcou, G. Curzon, Female rats are morevulnerable than males in an animal model of depression: the possible role ofserotonin, Brain Res. 382 (1986) 416–421.
[176] R. Kessler, Gender differences in major depression: epidemiological findings,in: E. Frank (Ed.), Gender and its Effects on Psychopathology, AmericanPsychiatric Press, Washington, 2000, pp. 61–84.
[177] R.C. Kessler, K.A. McGonagle, S. Zhao, C.B. Nelson, M. Hughes, S. Eshleman,H.U. Wittchen, K.S. Kendler, Lifetime and 12-month prevalence of DSM-III-Rpsychiatric disorders in the United States. Results from the nationalcomorbidity survey, Arch. Gen. Psychiatry 51 (1994) 8–19.
[178] D. Kimura, Sex, sexual orientation and sex hormones influence humancognitive function, Curr. Opin. Neurobiol. 6 (1996) 259–263.
[179] D. Kimura, R.A. Harshman, Sex differences in brain organization for verbaland non-verbal functions, Prog. Brain Res. 61 (1984) 423–441.
[180] K.M. Kirk, J.M. Bailey, M.P. Dunne, N.G. Martin, Measurement models forsexual orientation in a community twin sample, Behav. Genet. 30 (2000)345–356.
[181] P. Kirsch, C. Esslinger, Q. Chen, D. Mier, S. Lis, S. Siddhanti, H. Gruppe, V.S.Mattay, B. Gallhofer, A. Meyer-Lindenberg, Oxytocin modulates neuralcircuitry for social cognition and fear in humans, J. Neurosci. 25 (2005)11489–11493.
[182] P. Koopman, Gonad development: signals for sex, Curr. Biol. 11 (2001) R481–R483.
[183] C. Kordon, J. Glowinski, Role of hypothalamic monoaminergic neurones in thegonadotrophin release-regulating mechanisms, Neuropharmacology 11(1972) 153–162.
[184] M. Kosfeld, M. Heinrichs, P.J. Zak, U. Fischbacher, E. Fehr, Oxytocin increasestrust in humans, Nature 435 (2005) 673–676.
[185] T.R. Kosten, B.J. Rounsaville, H.D. Kleber, Ethnic and gender differencesamong opiate addicts, Int. J. Addict. 20 (1985) 1143–1162.
[186] J. Kulkarni, A. de Castella, D. Smith, J. Taffe, N. Keks, D. Copolov, A clinical trialof the effects of estrogen in acutely psychotic women, Schizophr. Res. 20(1996) 247–252.
[187] T.C. LaGrange, R.A. Silverman, Low self-control and opportunity: testing thegeneral theory of crime as an explanation for gender differences indelinquency, Criminology 37 (1999) 41–72.
[188] K. Lawrence, J. Kuntsi, M. Coleman, R. Campbell, D. Skuse, Face and emotionrecognition deficits in Turner syndrome: a possible role for X-linked genes inamygdala development, Neuropsychology 17 (2003) 39–49.
[189] V. Lefebvre, B. Dumitriu, A. Penzo-Mendez, Y. Han, B. Pallavi, Control of cellfate and differentiation by Sry-related high-mobility-group box (Sox)transcription factors, Int. J. Biochem. Cell Biol. 39 (2007) 2195–2214.
[190] C. Leranth, R.H. Roth, J.D. Elsworth, F. Naftolin, T.L. Horvath, D.E. Redmond Jr.,Estrogen is essential for maintaining nigrostriatal dopamine neurons inprimates: implications for Parkinson’s disease and memory, J. Neurosci. 20(2000) 8604–8609.
[191] R.F. Levant, R.J. Hall, C.M. Williams, N.T. Hasan, Gender differences inalexithymia, Psychol. Men Masc. 10 (2009) 190–203.
[192] S. LeVay, A difference in hypothalamic structure between heterosexual andhomosexual men, Science 253 (1991) 1034–1037.
[193] M. Levitt, S. Spector, A. Sjoerdsma, S. Udenfriend, Elucidation of the rate-limiting step in norepinephrine biosynthesis in the perfused guinea-pigheart, J. Pharmacol. Exp. Ther. 148 (1965) 1–8.
[194] H.M. Levitt, E. Ovrebo, M.B. Anderson-Cleveland, C. Leone, J.Y. Jeong, J.R. Arm,B.P. Bonin, J. Cicala, R. Coleman, A. Laurie, J.M. Vardaman, S.G. Horne,Balancing dangers: GLBT experience in a time of anti-GLBT legislation, J.Couns. Psychol. 56 (2009) 67–81.
[195] C. Lewin, G. Wolgers, A. Herlitz, Sex differences favoring women in verbal butnot in visuospatial episodic memory, Neuropsychology 15 (2001) 165–173.
[196] F.R. Lillie, The theory of the free-martin, Science 43 (1916) 611–613.[197] F.R. Lillie, General Biological Introduction, Williams and Wilkins Co.,
Baltimore, 1939.[198] R.A. Lippa, Sex differences in sex drive, sociosexuality, and height across 53
nations: testing evolutionary and social structural theories, Arch. Sex. Behav.38 (2009) 631–651.
[199] J.C. Lucas, M.B. Renfree, G. Shaw, C.M. Butler, The influence of the anti-androgen flutamide on early sexual differentiation of the marsupial male, J.Reprod. Fertil. 109 (1997) 205–212.
[200] E. Luders, C. Gaser, K.L. Narr, A.W. Toga, Why sex matters: brain sizeindependent differences in gray matter distributions between men andwomen, J. Neurosci. 29 (2009) 14265–14270.
[201] V. Luine, Sex differences in chronic stress effects on memory in rats, Stress 5(2002) 205–216.
[202] J.P. Macke, N. Hu, S. Hu, M. Bailey, V.L. King, T. Brown, D. Hamer, J. Nathans,Sequence variation in the androgen receptor gene is not a commondeterminant of male sexual orientation, Am. J. Hum. Genet. 53 (1993) 844–852.
[203] K. Maeda, S. Adachi, K. Inoue, S. Ohkura, H. Tsukamura, Metastin/kisspeptinand control of estrous cycle in rats, Rev. Endocr. Metab. Disord. 8 (2007) 21–29.
[204] S. Magre, A. Jost, Sertoli cells and testicular differentiation in the rat fetus, J.Electron. Microsc. Tech. 19 (1991) 172–188.
[205] P.M. Maki, S.M. Resnick, Longitudinal effects of estrogen replacement therapyon PET cerebral blood flow and cognition, Neurobiol. Aging 21 (2000) 373–383.
[206] A. Mansour, C.A. Fox, H. Akil, S.J. Watson, Opioid-receptor mRNA expressionin the rat CNS: anatomical and functional implications, Trends Neurosci. 18(1995) 22–29.
[207] S.B. Manuck, J.D. Flory, R.E. Ferrell, J.J. Mann, M.F. Muldoon, A regulatorypolymorphism of the monoamine oxidase-A gene may be associated withvariability in aggression, impulsivity, and central nervous systemserotonergic responsivity, Psychiatry Res. 95 (2000) 9–23.
[208] D.M. Maraganore, M. de Andrade, T.G. Lesnick, K.J. Strain, M.J. Farrer, W.A.Rocca, P.V. Pant, K.A. Frazer, D.R. Cox, D.G. Ballinger, High-resolution whole-genome association study of Parkinson disease, Am. J. Hum. Genet. 77 (2005)685–693.
[209] E.J. Marco, D.H. Skuse, Autism-lessons from the X chromosome, Soc. Cogn.Affect. Neurosci. 1 (2006) 183–193.

244 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
[210] E. Martignoni, R.E. Nappi, A. Citterio, D. Calandrella, E. Corengia, A. Fignon, R.Zangaglia, G. Riboldazzi, C. Pacchetti, G. Nappi, Parkinson’s disease andreproductive life events, Neurol. Sci. 23 (Suppl. 2) (2002) S85–S86.
[211] A. Matsumoto, Y. Arai, Male-female difference in synaptic organization of theventromedial nucleus of the hypothalamus in the rat, Neuroendocrinology 42(1986) 232–236.
[212] A. Mayer, G. Lahr, D.F. Swaab, C. Pilgrim, I. Reisert, The Y-chromosomal genesSRY and ZFY are transcribed in adult human brain, Neurogenetics 1 (1998)281–288.
[213] A. Mayer, G. Mosler, W. Just, C. Pilgrim, I. Reisert, Developmental profile of Srytranscripts in mouse brain, Neurogenetics 3 (2000) 25–30.
[214] B.M. McCarty, C.J. Migeon, H.F. Meyer-Bahlburg, H. Zacur, A.B. Wisniewski,Medical and psychosexual outcome in women affected by complete gonadaldysgenesis, J. Pediatr. Endocrinol. Metab. 19 (2006) 873–877.
[215] E. McCauley, P. Feuillan, H. Kushner, J.L. Ross, Psychosocial developmentin adolescents with Turner syndrome, J. Dev. Behav. Pediatr. 22 (2001)360–365.
[216] D. McFadden, J.C. Loehlin, S.M. Breedlove, R.A. Lippa, J.T. Manning, Q. Rahman,A reanalysis of five studies on sexual orientation and the relative length ofthe 2nd and 4th fingers (the 2D:4D ratio), Arch. Sex. Behav. 34 (2005) 341–356.
[217] B. Meister, Neurotransmitters in key neurons of the hypothalamus thatregulate feeding behavior and body weight, Physiol. Behav. 92 (2007) 263–271.
[218] I.H. Meyer, P.A. Wilson, Sampling lesbian, gay, and bisexual populations, J.Couns. Psychol. 56 (2009) 23–31.
[219] A. Milsted, L. Serova, E.L. Sabban, G. Dunphy, M.E. Turner, D.L. Ely, Regulationof tyrosine hydroxylase gene transcription by Sry, Neurosci. Lett. 369 (2004)203–207.
[220] D. Mitsushima, K. Yamada, K. Takase, T. Funabashi, F. Kimura, Sex differencesin the basolateral amygdala: the extracellular levels of serotonin anddopamine, and their responses to restraint stress in rats, Eur. J. NeuroSci.24 (2006) 3245–3254.
[221] S. Mizukami, M. Nishizuka, Y. Arai, Sexual difference in nuclear volume andits ontogeny in the rat amygdala, Exp. Neurol. 79 (1983) 569–575.
[222] C. Modahl, L. Green, D. Fein, M. Morris, L. Waterhouse, C. Feinstein, H. Levin,Plasma oxytocin levels in autistic children, Biol. Psychiatry 43 (1998) 270–277.
[223] B. Moradi, J.J. Mohr, R.L. Worthington, R.E. Fassinger, Counseling psychologyresearch on sexual (orientation) minority issues: conceptual andmethodological challenges and opportunities, J. Couns. Psychol. 56 (2009)5–22.
[224] A.G. Morris, E. Gaitonde, P.J. McKenna, J.D. Mollon, D.M. Hunt, CAG repeatexpansions and schizophrenia: association with disease in females and withearly age-at-onset, Hum. Mol. Genet. 4 (1995) 1957–1961.
[225] J.A. Morris, C.L. Jordan, S.M. Breedlove, Sexual differentiation of thevertebrate nervous system, Nat. Neurosci. 7 (2004) 1034–1039.
[226] C.A. Moss-Racusin, J.E. Phelan, L.A. Rudman, When men break the genderrules: status incongruity and backlash against modest men, Psychol. MenMasc. 11 (2010) 140–151.
[227] C.A. Munro, M.E. McCaul, D.F. Wong, L.M. Oswald, Y. Zhou, J. Brasic, H.Kuwabara, A. Kumar, M. Alexander, W. Ye, G.S. Wand, Sex differences instriatal dopamine release in healthy adults, Biol. Psychiatry 59 (2006) 966–974.
[228] D.D. Murphy, N.B. Cole, M. Segal, Brain-derived neurotrophic factor mediatesestradiol-induced dendritic spine formation in hippocampal neurons, Proc.Natl. Acad. Sci. USA 95 (1998) 11412–11417.
[229] C.M. Murphy, C.T. Taft, C.I. Eckhardt, Anger problem profiles among partnerviolent men: differences in clinical presentation and treatment outcome, J.Couns. Psychol. 54 (2007) 189–200.
[230] B.S. Mustanski, M.G. Dupree, C.M. Nievergelt, S. Bocklandt, N.J. Schork, D.H.Hamer, A genomewide scan of male sexual orientation, Hum. Genet. 116(2005) 272–278.
[231] F. Naftolin, Brain aromatization of androgens, J. Reprod. Med. 39 (1994) 257–261.
[232] J. Nielsen, M. Wohlert, Sex chromosome abnormalities found among 34,910newborn children: results from a 13-year incidence study in Arhus, Denmark,Birth Defects – Orig. Artic. Ser. 26 (1990) 209–223.
[233] S. Nishizawa, C. Benkelfat, S.N. Young, M. Leyton, S. Mzengeza, C.DeMontigny, P. Blier, M. Diksic, Differences between males and females inrates of serotonin synthesis in human brain, Proc. Natl. Acad. Sci. USA 94(1997) 5308–5313.
[234] S. Nishizawa, C. Benkelfat, S.N. Young, M. Leyton, S. Mzengeza, C. DeMontigny, P. Blier, M. Diksic, Differences between males and females in ratesof serotonin synthesis human brain, Proc. Natl. Acad. Sci. USA 94 (1997)5308–5313.
[235] A. Nordenstrom, A. Servin, G. Bohlin, A. Larsson, A. Wedell, Sex-typed toy playbehavior correlates with the degree of prenatal androgen exposure assessedby CYP21 genotype in girls with congenital adrenal hyperplasia, J. Clin.Endocrinol. Metab. 87 (2002) 5119–5124.
[236] F. Nottebohm, A.P. Arnold, Sexual dimorphism in vocal control areas of thesongbird brain, Science 194 (1976) 211–213.
[237] K.N. Ochsner, The social-emotional processing stream: five core constructsand their translational potential for schizophrenia and beyond, Biol.Psychiatry 64 (2008) 48–61.
[238] J.M. O’neil, Is criticism of generic masculinity, essentialism, and positive-healthy-masculinity a problem for the psychology of men?, Psychol MenMasc. 11 (2010) 98–106.
[239] M. Ota, J.T. Crofton, L. Share, Hemorrhage-induced vasopressin release in theparaventricular nucleus measured by in vivo microdialysis, Brain Res. 658(1994) 49–54.
[240] M.G. Packard, Posttraining estrogen and memory modulation, Horm. Behav.34 (1998) 126–139.
[241] N. Pankratz, W.C. Nichols, S.K. Uniacke, C. Halter, J. Murrell, A. Rudolph, C.W.Shults, P.M. Conneally, T. Foroud, Genome-wide linkage analysis andevidence of gene-by-gene interactions in a sample of 362 multiplexParkinson disease families, Hum. Mol. Genet. 12 (2003) 2599–2608.
[242] C.E. Patek, J.B. Kerr, R.G. Gosden, K.W. Jones, K. Hardy, A.L. Muggleton-Harris,A.H. Handyside, D.G. Whittingham, M.L. Hooper, Sex chimaerism, fertility andsex determination in the mouse, Development 113 (1991) 311–325.
[243] A.M.L. Pattatucci, D.H. Hamer, Development and familiarity of sexualorientation in females, Behav. Genet. 25 (1995) 407–420.
[244] D.W. Pfaff, Hormones, Brain, and Behavior, Academic Press, Amsterdam,Boston, 2002.
[245] T. Pfluger, S. Weil, S. Weis, C. Vollmar, D. Heiss, J. Egger, R. Scheck, K. Hahn,Normative volumetric data of the developing hippocampus in children basedon magnetic resonance imaging, Epilepsia 40 (1999) 414–423.
[246] S.M. Phillips, B.B. Sherwin, Effects of estrogen on memory function insurgically menopausal women, Psychoneuroendocrinology 17 (1992) 485–495.
[247] C.H. Phoenix, R.W. Goy, A.A. Gerall, W.C. Young, Organizing action ofprenatally administered testosterone propionate on the tissuesmediating mating behavior in the female guinea pig, Endocrinology65 (1959) 369–382.
[248] J.P. Piek, N. Gasson, N. Barrett, I. Case, Limb and gender differences in thedevelopment of coordination in early infancy, Hum. Mov. Sci. 21 (2002) 621–639.
[249] R.C. Pillard, J.D. Weinrich, Evidence of familial nature of male homosexuality,Arch. Gen. Psychiatry 43 (1986) 808–812.
[250] A. Poletti, P. Negri-Cesi, L. Martini, Reflections on the diseases linked tomutations of the androgen receptor, Endocrine 28 (2005) 243–262.
[251] J.B. Powers, Hormonal control of sexual receptivity during the estrous cycle ofthe rat, Physiol. Behav. 5 (1970) 831–835.
[252] B. Powers, E.S. Valenstein, Sexual receptivity: facilitation by medial preopticlesions in female rats, Science 175 (1972) 1003–1005.
[253] M.L.D. Pozzo, A. Aoki, Stereological analysis of the hypothalamicventromedial nucleus: II. Hormone-induced changes in the synaptogenicpattern, Brain Res. Dev. Brain Res. 61 (1991) 189–196.
[254] N.J. Preston, K.G. Orr, R. Date, L. Nolan, D.J. Castle, Gender differences inpremorbid adjustment of patients with first episode psychosis, Schizophr.Res. 55 (2002) 285–290.
[255] C.A. Quigley, A. De Bellis, K.B. Marschke, M.K. el-Awady, E.M. Wilson, F.S.French, Androgen receptor defects: historical, clinical, and molecularperspectives, Endocr. Rev. 16 (1995) 271–321.
[256] J.J. Quinn, P.K. Hitchcott, E.A. Umeda, A.P. Arnold, J.R. Taylor, Sex chromosomecomplement regulates habit formation, Nat. Neurosci. 10 (2007) 1398–1400.
[257] H. Raab, C. Pilgrim, I. Reisert, Effects of sex and estrogen on tyrosinehydroxylase mRNA in cultured embryonic rat mesencephalon, Brain Res. Mol.Brain Res. 33 (1995) 157–164.
[258] P. Ragonese, M. D’Amelio, G. Salemi, P. Aridon, M. Gammino, A. Epifanio, L.Morgante, G. Savettieri, Risk of Parkinson disease in women: effect ofreproductive characteristics, Neurology 62 (2004) 2010–2014.
[259] I. Reisert, C. Pilgrim, Sexual differentiation of monoaminergic neurons –genetic or epigenetic?, Trends Neurosci 14 (1991) 468–473.
[260] M.B. Renfree, R.V. Short, Sex determination in marsupials: evidence for amarsupial-eutherian dichotomy, Philos. Trans. R. Soc. B – Biol. Sci. 322 (1988)41–53.
[261] M.B. Renfree, W.S.O.R.V. Short, G. Shaw, Sexual differentiation of theurogenital system of the fetal and neonatal tammar wallaby, Macropuseugenii, Anat. Embryol. (Berl.) 194 (1996) 111–134.
[262] R.W. Rhees, H.N. Al-Saleh, E.W. Kinghorn, D.E. Fleming, E.D. Lephart,Relationship between sexual behavior and sexually dimorphic structures inthe anterior hypothalamus in control and prenatally stressed male rats, BrainRes. Bull. 50 (1999) 193–199.
[263] M.E. Rhodes, R.T. Rubin, Functional sex differences (‘sexual diergism’) ofcentral nervous system cholinergic systems, vasopressin, and hypothalamic–pituitary–adrenal axis activity in mammals: a selective review, Brain Res.Rev. 30 (1999) 135–152.
[264] G. Rice, C. Anderson, N. Risch, G. Ebers, Male homosexuality: absence oflinkage to microsatellite markers at Xq28, Science 284 (1999) 665–667.
[265] G. Rieger, M.L. Chivers, J.M. Bailey, Sexual arousal patterns of bisexual men,Psychol. Sci. 16 (2005) 579–584.
[266] S.J. Robbins, R.N. Ehrman, A.R. Childress, C.P. O’Brien, Comparing levels ofcocaine cue reactivity in male and female outpatients, Drug Alcohol Depend.53 (1999) 223–230.
[267] A. Rojo, M. Aguilar, M.T. Garolera, E. Cubo, I. Navas, S. Quintana, Depression inParkinson’s disease: clinical correlates and outcome, Parkinsonism Relat.Disord. 10 (2003) 23–28.
[268] C.E. Roselli, F. Stormshak, The neurobiology of sexual partner preferences inrams, Horm. Behav. 55 (2009) 611–620.

T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246 245
[269] C.E. Roselli, L.E. Horton, J.A. Resko, Distribution and regulation of aromataseactivity in the rat hypothalamus and limbic system, Endocrinology 117(1985) 2471–2477.
[270] C.E. Roselli, S.E. Abdelgadir, J.A. Resko, Regulation of aromatase geneexpression in the adult rat brain, Brain Res. Bull. 44 (1997) 351–357.
[271] C.E. Roselli, K. Larkin, J.A. Resko, J.N. Stellflug, F. Stormshak, The volume of asexually dimorphic nucleus in the ovine medial preoptic area/anteriorhypothalamus varies with sexual partner preference, Endocrinology 145(2004) 478–483.
[272] J.L. Ross, G.A. Stefanatos, H. Kushner, C. Bondy, L. Nelson, A. Zinn, D. Roeltgen,The effect of genetic differences and ovarian failure: intact cognitive functionin adult women with premature ovarian failure versus Turner syndrome, J.Clin. Endocrinol. Metab. 89 (2004) 1817–1822.
[273] R.T. Rubin, L.K. Sekula, S. O’Toole, M.E. Rhodes, R.K. Czambel, Pituitary–adrenal cortical responses to low-dose physostigmine and argininevasopressin administration in normal women and men,Neuropsychopharmacology 20 (1999) 434–446.
[274] D.R. Rubinow, P.J. Schmidt, C.A. Roca, Estrogen–serotonin interactions:implications for affective regulation, Biol. Psychiatry 44 (1998) 839–850.
[275] H.G. Ruhe, N.S. Mason, A.H. Schene, Mood is indirectly related to serotonin,norepinephrine and dopamine levels in humans: a meta-analysis ofmonoamine depletion studies, Mol. Psychiatry 12 (2007) 331–359.
[276] M. Sadeghi, A. Fakhrai, Transsexualism in female monozygotic twins: a casereport, Aust. NZ J. Psychiatry 34 (2000) 862–864.
[277] F.J. Sanchez, E. Vilain, Collective self-esteem as a coping resource for male-to-female transsexuals, J. Couns. Psychol. 56 (2009) 202–209.
[278] F.J. Sánchez, S. Bocklandt, E. Vilain, The biology of sexual orientation andgender identity, in: D.W. Pfaff, A.P. Arnold, A.M. Etgen, S.E. Fahrbach, R.T.Rubin (Eds.), Hormones, Brain and Behavior, Academic Press, San Diego, CA,2009, pp. 1911–1929.
[279] A.R. Sanders, K. Dawood, Nature Encyclopedia of Life Sciences, NaturePublishing Group, London, 2003.
[280] H.E. Scharfman, N.J. Maclusky, Similarities between actions of estrogen andBDNF in the hippocampus: coincidence or clue?, Trends Neurosci 28 (2005)79–85.
[281] D.P. Schmitt, Sociosexuality from Argentina to Zimbabwe: a 48-nation studyof sex, culture, and strategies of human mating, Behav. Brain. Sci. 28 (2005)247–275 (Discussion 275–311).
[282] K. Schweizer, F. Brunner, K. Schutzmann, V. Schonbucher, H. Richter-Appelt,Gender identity and coping in female 46, XY adults with androgenbiosynthesis deficiency (intersexuality/DSD), J. Couns. Psychol. 56 (2009)189–201.
[283] M.V. Seeman, Psychopathology in women and men: focus on femalehormones, Am. J. Psychiatry 154 (1997) 1641–1647.
[284] N.L. Segal, Two monozygotic twin pairs discordant for female-to-maletranssexualism, Arch. Sex. Behav. 35 (2006) 347–358.
[285] S.N. Seidman, A.B. Araujo, S.P. Roose, J.B. McKinlay, Testosterone level,androgen receptor polymorphism, and depressive symptoms in middle-agedmen, Biol. Psychiatry 50 (2001) 371–376.
[286] R. Sekido, R. Lovell-Badge, Sex determination involves synergistic actionof SRY and SF1 on a specific Sox9 enhancer, Nature 453 (2008) 930–934.
[287] L.I. Serova, S. Maharjan, A. Huang, D. Sun, G. Kaley, E.L. Sabban, Response oftyrosine hydroxylase and GTP cyclohydrolase I gene expression to estrogen inbrain catecholaminergic regions varies with mode of administration, BrainRes. 1015 (2004) 1–8.
[288] B. Seymour, R. Dolan, Emotion, decision making, and the amygdala, Neuron58 (2008) 662–671.
[289] G.B. Sharman, The chromosomal basis of sex differentiation in marsupials,Aust. J. Zool. 37 (1990) 451–466.
[290] G. Shaw, M.B. Renfree, R.V. Short, W.S. O, Experimental manipulation ofsexual differentiation in wallaby pouch young treated with exogenoussteroids, Development 104 (1988) 689–701.
[291] B.B. Sherwin, Estrogen and cognitive functioning in women, Proc. Soc. Exp.Biol. Med. 217 (1998) 17–22.
[292] B.B. Sherwin, Estrogen and cognitive functioning in women, Endocr. Rev. 24(2003) 133–151.
[293] W.S.O.R.V. Short, M.B. Renfree, G. Shaw, Primary genetic control of somaticsexual differentiation in a mammal, Nature 331 (1988) 716–717.
[294] P.J. Shughrue, P.J. Scrimo, I. Merchenthaler, Estrogen binding and estrogenreceptor characterization (ERalpha and ERbeta) in the cholinergic neurons ofthe rat basal forebrain, Neuroscience 96 (2000) 41–49.
[295] R. Sibug, E. Kuppers, C. Beyer, S.C. Maxson, C. Pilgrim, I. Reisert,Genotype-dependent sex differentiation of dopaminergic neurons inprimary cultures of embryonic mouse brain, Brain Res. Dev. Brain Res.93 (1996) 136–142.
[296] A. Siddiqui, B.H. Shah, Neonatal androgen manipulation differentially affectsthe development of monoamine systems in rat cerebral cortex, amygdala andhypothalamus, Brain Res. Dev. Brain Res. 98 (1997) 247–252.
[297] L.W.S.R.B. Simerly, C. Chang, M. Muramatsu, Distribution of androgen andestrogen receptor mRNA-containing cells in the rat brain: an in situhybridization study, J. Comp. Neurol. 294 (1990) 76–95.
[298] J.W. Simpkins, P.S. Green, K.E. Gridley, M. Singh, N.C. de Fiebre, G. Rajakumar,Role of estrogen replacement therapy in memory enhancement and theprevention of neuronal loss associated with Alzheimer’s disease, Am. J. Med.103 (1997) 19S–25S.
[299] H.B. Simpson, D.S. Vicario, Early estrogen treatment of female zebra finchesmasculinizes the brain pathway for learned vocalizations, J. Neurobiol. 22(1991) 777–793.
[300] D.H. Skuse, R.S. James, D.V. Bishop, B. Coppin, P. Dalton, r.G. Aamodt-Leepe,M. Bacarese-Hamilton, C. Creswell, R. McGurk, P.A. Jacobs, Evidence fromTurner’s syndrome of an imprinted X-linked locus affecting cognitivefunction, Nature 387 (1997).
[301] D.H. Skuse, R.S. James, D.V. Bishop, B. Coppin, P. Dalton, G. Aamodt-Leeper, M.Bacarese-Hamilton, C. Creswell, R. McGurk, P.A. Jacobs, Evidence fromTurner’s syndrome of an imprinted X-linked locus affecting cognitivefunction, Nature 387 (1997) 705–708.
[302] R.P. Snaith, S. Penhale, P. Horsfield, Male-to-female transsexual with XYYkaryotype, Lancet 337 (1991) 557–558.
[303] M.B. Solomon, J.P. Herman, Sex differences in psychopathology: of gonads,adrenals and mental illness, Physiol. Behav. 97 (2009) 250–258.
[304] R. Sperry, Some effects of disconnecting the cerebral hemispheres, Science217 (1982) 1223–1226.
[305] J.K. Staley, S. Krishnan-Sarin, S. Zoghbi, G. Tamagnan, M. Fujita, J.P. Seibyl, P.K.Maciejewski, S. O’Malley, R.B. Innis, Sex differences in [123I]beta-CIT SPECTmeasures of dopamine and serotonin transporter availability in healthysmokers and nonsmokers, Synapse 41 (2001) 275–284.
[306] E.R. Stockman, R.S. Callaghan, M.J. Baum, Effects of neonatal castration andtestosterone treatment on sexual partner preference in the ferret, Physiol.Behav. 34 (1985) 409–414.
[307] P. Sudbeck, G. Scherer, Two independent nuclear localization signals arepresent in the DNA-binding high-mobility group domains of SRY and SOX9, J.Biol. Chem. 272 (1997) 27848–27852.
[308] H. Sumida, M. Nishizuka, Y. Kano, Y. Arai, Sex differences in the anteroventralperiventricular nucleus of the preoptic area and in the related effects ofandrogen in prenatal rats, Neurosci. Lett. 151 (1993) 41–44.
[309] D.F. Swaab, J.N. Zhou, M. Fodor, M.A. Hofman, Sexual differentiation of thehuman hypothalamus: differences according to sex, sexual orientation, andtranssexuality, in: Ellis Lee, Ebertz Linda (Eds.), Sexual Orientation: TowardBiological Understanding, Praeger Publishers/Greenwood Publishing Group,Inc., Westport, CT, US, 1997, pp. 129–150 (xxii, 276 pp. see book).
[310] M. Sylvester, S.C. Hayes, Unpacking masculinity as a construct: ontology,pragmatism, and an analysis of language, Psychol. Men Masc. 11 (2010) 91–97.
[311] D. Tager, G.E. Good, S. Brammer, ‘‘Walking over ‘Em”: an exploration of relationsbetween emotion dysregulation, masculine norms, and intimate partner abusein a clinical sample of men, Psychol. Men Masc. 11 (2010) 233–239.
[312] T. Taketo, C.H. Lee, J. Zhang, Y. Li, C.Y. Lee, Y.F. Lau, Expression of SRY proteinsin both normal and sex-reversed XY fetal mouse gonads, Dev. Dyn. 233(2005) 612–622.
[313] N. Taneja, A.C. Ammini, I. Mohapatra, S. Saxena, K. Kucheria, A transsexualmale with 47,XYY karyotype, Br. J. Psychiatry 161 (1992) 698–699.
[314] C.M. Temple, Oral fluency and narrative production in children with Turner’ssyndrome, Neuropsychologia 40 (2002) 1419–1427.
[315] A.J. Tilbrook, A.I. Turner, I.J. Clarke, Stress and reproduction: centralmechanisms and sex differences in non-rodent species, Stress 5 (2002) 83–100.
[316] L.S. Tottenham, D.M. Saucier, Throwing accuracy during prism adaptation:male advantage for throwing accuracy is independent of prism adaptationrate, Percept. Mot. Skills 98 (2004) 1449–1455.
[317] Shinji Tsukahara, Sex differences and roles of sex steroids in apoptosis ofsexually dimorphic nuclei of preoptic area in postnatal rats, J.Neuroendocrinol. 9999 (2009).
[318] D. Twelves, K.S. Perkins, C. Counsell, Systematic review of incidence studies ofParkinson’s disease, Mov. Disord. 18 (2003) 19–31.
[319] H. Ujike, K. Otani, M. Nakatsuka, K. Ishii, A. Sasaki, T. Oishi, T. Sato, Y. Okahisa,Y. Matsumoto, Y. Namba, Y. Kimata, S. Kuroda, Association study of genderidentity disorder and sex hormone-related genes, Prog. Neuropsycho-pharmacol. Biol. Psychiatry 33 (2009) 1241–1244.
[320] M.T. Ullman, Contributions of memory circuits to language: the declarative/procedural model, Cognition 92 (2004) 231–270.
[321] S.K. Van Den Eeden, C.M. Tanner, A.L. Bernstein, R.D. Fross, A. Leimpeter, D.A.Bloch, L.M. Nelson, Incidence of Parkinson’s disease: variation by age, gender,and race/ethnicity, Am. J. Epidemiol. 157 (2003) 1015–1022.
[322] S. van Rijn, H. Swaab, A. Aleman, R.S. Kahn, X Chromosomal effects on socialcognitive processing and emotion regulation: a study with Klinefelter men(47,XXY), Schizophr. Res. 84 (2006) 194–203.
[323] A.H. van Stegeren, R. Goekoop, W. Everaerd, P. Scheltens, F. Barkhof, J.P.A.Kuijer, S.A.R.B. Rombouts, Noradrenaline mediates amygdala activation inmen and women during encoding of emotional material, NeuroImage 24(2005) 898–909.
[324] E. Vilain, Genetics of intersexuality, J. Gay Lesbian Psychother. 10 (2006) 9–26.
[325] B. von der Pahlen, R. Lindman, T. Sarkola, H. Makisalo, C.J.P. Eriksson, Anexploratory study on self-evaluated aggression and androgens in women,Aggress. Behav. 28 (2002) 273–280.
[326] D. Voyer, S. Voyer, M.P. Bryden, Magnitude of sex differences in spatialabilities: a meta-analysis and consideration of critical variables, Psychol. Bull.117 (1995) 250–270.
[327] J. Wade, Zebra finch sexual differentiation: the aromatization hypothesisrevisited, Microsc. Res. Tech. 54 (2001) 354–363.
[328] J. Wade, A.P. Arnold, Functional testicular tissue does not masculinizedevelopment of the zebra finch song system, Proc. Natl. Acad. Sci. USA 93(1996) 5264–5268.

246 T.C. Ngun et al. / Frontiers in Neuroendocrinology 32 (2011) 227–246
[329] D.L. Walker, D.J. Toufexis, M. Davis, Role of the bed nucleus of the striaterminalis versus the amygdala in fear, stress, and anxiety, Eur. J. Pharmacol.463 (2003) 199–216.
[330] Z. Wang, N.A. Bullock, G.J. De Vries, Sexual differentiation of vasopressinprojections of the bed nucleus of the stria terminals and medial amygdaloidnucleus in rats, Endocrinology 132 (1993) 2299–2306.
[331] T.H. Wassink, J. Piven, V.J. Vieland, J. Pietila, R.J. Goedken, S.E. Folstein, V.C.Sheffield, Examination of AVPR1a as an autism susceptibility gene, Mol.Psychiatry 9 (2004) 968–972.
[332] M.A. Weiss, Floppy SOX: mutual induced fit in HMG (high-mobility group)box-DNA recognition, Mol. Endocrinol. 15 (2001) 353–362.
[333] L.A. Weiss, M. Abney, E.H. Cook Jr., C. Ober, Sex-specific genetic architectureof whole blood serotonin levels, Am. J. Hum. Genet. 76 (2005) 33–41.
[334] F.J. White, A behavioral/systems approach to the neuroscience of drugaddiction, J. Neurosci. 22 (2002) 3303–3305.
[335] L.S. Whitfield, R. Lovell-Badge, P.N. Goodfellow, Rapid sequence evolution ofthe mammalian sex-determining gene SRY, Nature 364 (1993) 713–715.
[336] A.C. Wihlback, I.S. Poromaa, M. Bixo, P. Allard, T. Mjorndal, O. Spigset, Influenceof menstrual cycle on platelet serotonin uptake site and serotonin(2A) receptorbinding, Psychoneuroendocrinology 29 (2004) 757–766.
[337] J.D. Wilson, F.W. George, M.B. Renfree, The endocrine role in mammaliansexual differentiation, Recent Prog. Horm. Res. 50 (1995) 349–364.
[338] S.F. Witelson, Glezer II, D.L. Kigar, Women have greater density of neurons inposterior temporal cortex, J. Neurosci. 15 (1995) 3418–3428.
[339] W. Wood, A.H. Eagly, A cross-cultural analysis of the behavior of women andmen: implications for the origins of sex differences, Psychol. Bull. 128 (2002)699–727.
[340] S. Woody, P.D. McLean, S. Taylor, W.J. Koch, Treatment of major depression inthe context of panic disorder, J. Affect. Disord. 53 (1999) 163–174.
[341] C.S. Woolley, Effects of estrogen in the CNS, Curr. Opin. Neurobiol. 9 (1999)349–354.
[342] R.L. Worthington, A.L. Reynolds, Within-group differences in sexualorientation and identity, J. Couns. Psychol. 56 (2009) 44–55.
[343] S. Wu, M. Jia, Y. Ruan, J. Liu, Y. Guo, M. Shuang, X. Gong, Y. Zhang, X. Yang, D.Zhang, Positive association of the oxytocin receptor gene (OXTR) with autismin the Chinese Han population, Biol. Psychiatry 58 (2005) 74–77.
[344] J.B. Wu, K. Chen, Y. Li, Y.-F.C. Lau, J.C. Shih, Regulation of monoamine oxidaseA by the SRY gene on the Y chromosome, FASEB J. (2009) (fj.09-139097).
[345] J. Xu, P.S. Burgoyne, A.P. Arnold, Sex differences in sex chromosome geneexpression in mouse brain, Hum. Mol. Genet. 11 (2002) 1409–1419.
[346] T.L. Yaksh, Opioid receptor systems and the endorphins: a review of theirspinal organization, J. Neurosurg. 67 (1987) 157–176.
[347] N. Yirmiya, C. Rosenberg, S. Levi, S. Salomon, C. Shulman, L. Nemanov, C. Dina,R.P. Ebstein, Association between the arginine vasopressin 1a receptor(AVPR1a) gene and autism in a family-based study: mediation bysocialization skills, Mol. Psychiatry 11 (2006) 488–494.
[348] L. Zhang, W. Ma, J.L. Barker, D.R. Rubinow, Sex differences in expression ofserotonin receptors (subtypes 1A and 2A) in rat brain: a possible role oftestosterone, Neuroscience 94 (1999) 251–259.
[349] D. Zhao, D. McBride, S. Nandi, H.A. McQueen, M.J. McGrew, P.M. Hocking, P.D.Lewis, H.M. Sang, M. Clinton, Somatic sex identity is cell autonomous in thechicken, Nature 464 (2010) 237–242.
[350] J.N. Zhou, M.A. Hofman, L.J. Gooren, D.F. Swaab, A sex difference in the humanbrain and its relation to transsexuality, Nature 378 (1995) 68–70.
[351] B. Zimmerberg, L.V. Scalzi, Commissural size in neonatal rats: effects of sexand prenatal alcohol exposure, Int. J. Dev. Neurosci. 7 (1989) 81–86.
[352] J.-K. Zubieta, R.F. Dannals, J.J. Frost, Gender and age influences on humanbrain mu-opioid receptor binding measured by PET, Am. J. Psychiatry 156(1999) 842–848.
[353] D.G. Zuloaga, D.A. Puts, C.L. Jordan, S.M. Breedlove, The role of androgenreceptors in the masculinization of brain and behavior: what we’ve learnedfrom the testicular feminization mutation, Horm. Behav. 53 (2008) 613–626.
Related Documents











