International Journal of Antimicrobial Agents 39 (2012) 16–21 Contents lists available at SciVerse ScienceDirect International Journal of Antimicrobial Agents j our na l ho me p age: http://www.elsevier.com/locate/ijantimicag Genetic response of Salmonella enterica serotype Enteritidis to thioridazine rendering the organism resistant to the agent Gabriella Spengler a,b,c , Liliana Rodrigues a,b , Ana Martins a,b,1 , Marta Martins d , Matthew McCusker d , Pedro Cerca a,b , Lisa Machado a,b , Sofia Santos Costa a,b , Eleni Ntokou e , Isabel Couto a,f , Miguel Viveiros a,g , Séamus Fanning d,g , Joseph Molnar c,g , Leonard Amaral a,b,g,∗ a Grupo de Micobactérias, Unidade de Microbiologia Médica, Instituto de Higiene e Medicina Tropical (IHMT), Universidade Nova de Lisboa, Rua da Junqueira 100, 1349-008 Lisbon, Portugal b UPMM (Unidade de Parasitologia e Microbiologia Médicas), Instituto de Higiene e Medicina Tropical (IHMT), Universidade Nova de Lisboa, Rua da Junqueira 100, 1349-008 Lisbon, Portugal c Institute of Medical Microbiology and Immunobiology, Faculty of Medicine, University of Szeged, Szeged, Hungary d UCD Centre for Food Safety, School of Agriculture, Food Science and Veterinary Medicine, University College Dublin, Ireland e Department of Microbiology, Medical School, University of Thessaly, Larissa, Greece f Centro de Recursos Microbiológicos (CREM), Faculdade de Ciências e Tecnologia (FCT), Universidade Nova de Lisboa, Portugal g Cost Action BM0701 (ATENS), of the European Commission/European Science Foundation, Brussels, Belgium a r t i c l e i n f o Article history: Received 8 July 2011 Accepted 24 August 2011 Keywords: Salmonella enterica serotype Enteritidis Thioridazine AcrAB efflux pump Real-time RT-PCR Activation of genes by thioridazine ramA marA soxS rob acrB pmrA pmrB Developed resistance to thioridazine during culture a b s t r a c t Thioridazine (TZ)-induced accumulation of the universal efflux pump substrate ethidium bromide and its subsequent efflux by Salmonella strains with various degrees of overexpressed efflux pumps takes place automatically at pH 7.4, is independent of a metabolic source, is not affected by a proton ionophore and is precluded by palmitic acid. Salmonella enterica serotype Enteritidis cultured in medium containing increasing concentrations of TZ does not grow during the first 6–8 h, after which time its growth is similar to unexposed controls. At the end of a 16-h exposure period, the organism is resistant to >250 mg/L TZ. Parallel assessment by real-time reverse transcription quantitative polymerase chain reaction (RT-qPCR) of the activity of genes that regulate and code for the AcrB transporter of the main efflux pump (AcrAB) of the organism at periodic intervals suggests a sequence of activation beginning with the stress gene soxS, followed by the global regulator ramA, then by the local regulator marA and then by the transporter acrB. These activations take place during the period of no growth. By the end of a 16-h culture period, only the acrB transporter gene is still highly overexpressed. Assessment of the activity of genes of the two- component regulon PmrA/B indicates that TZ also activates this regulon. Because activation of pmrA/B also activates acrB, development of high resistance to TZ during a 16-h culture period is in part due to activation of the two-component regulon. © 2011 Elsevier B.V. and the International Society of Chemotherapy. All rights reserved. 1. Introduction According to the US Centers for Disease Control and Prevention (CDC), ‘During 1993–1997, a total of 2,751 outbreaks of food- borne disease were reported (489 in 1993, 653 in 1994, 628 in 1995, 477 in 1996, and 504 in 1997). Among outbreaks for which the etiology was determined, bacterial pathogens caused the largest percentage of outbreaks (75%) and the largest per- centage of cases (86%). Salmonella enterica serotype Enteritidis ∗ Corresponding author. Tel.: +351 21 365 2600; fax: +351 21 363 2105. E-mail address: [email protected] (L. Amaral). 1 Permanent address: Institute of Pharmacognosy, Faculty of Pharmacy, Univer- sity of Szeged, Eötvös u. 6, 6720 Szeged, Hungary. accounted for the largest number of outbreaks, cases, and deaths.’ [1]; 95% of Salmonella outbreaks were from food-borne sources [2]. Nosocomial Salmonella infections and outbreaks in hospitals result from food-borne sources, most of which can be traced to food workers (faecal-to-hand contamination) with the remainder due to transfer from patient sources (faecal soiling of bed sheets, etc.) to hospital personnel who transfer the organism to other patients, etc. [3]. Although hospital-acquired Salmonella infections are readily managed, when they take place in patients who have had a resection of the colon, the organism may penetrate into the mucosa of the colon and produce high mortality rates regardless of therapy [4]. The main reason for problematic therapy lies in the variety of responses that Salmonella activates when in a nox- ious environment, rendering the organism quite resistant to most antibiotics. 0924-8579/$ – see front matter © 2011 Elsevier B.V. and the International Society of Chemotherapy. All rights reserved. doi:10.1016/j.ijantimicag.2011.08.013

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Gr
GPMa
Pb
Pc
d
e
f
g
a
ARA
KSTARArmsrappDc
1
(biwtc
s
0d
International Journal of Antimicrobial Agents 39 (2012) 16– 21
Contents lists available at SciVerse ScienceDirect
International Journal of Antimicrobial Agents
j our na l ho me p age: ht tp : / /www.e lsev ier .com/ locate / i jant imicag
enetic response of Salmonella enterica serotype Enteritidis to thioridazineendering the organism resistant to the agent
abriella Spenglera,b,c , Liliana Rodriguesa,b , Ana Martinsa,b,1 , Marta Martinsd , Matthew McCuskerd ,edro Cercaa,b, Lisa Machadoa,b, Sofia Santos Costaa,b, Eleni Ntokoue, Isabel Coutoa,f,iguel Viveirosa,g, Séamus Fanningd,g, Joseph Molnarc,g, Leonard Amarala,b,g,∗
Grupo de Micobactérias, Unidade de Microbiologia Médica, Instituto de Higiene e Medicina Tropical (IHMT), Universidade Nova de Lisboa, Rua da Junqueira 100, 1349-008 Lisbon,ortugalUPMM (Unidade de Parasitologia e Microbiologia Médicas), Instituto de Higiene e Medicina Tropical (IHMT), Universidade Nova de Lisboa, Rua da Junqueira 100, 1349-008 Lisbon,ortugalInstitute of Medical Microbiology and Immunobiology, Faculty of Medicine, University of Szeged, Szeged, HungaryUCD Centre for Food Safety, School of Agriculture, Food Science and Veterinary Medicine, University College Dublin, IrelandDepartment of Microbiology, Medical School, University of Thessaly, Larissa, GreeceCentro de Recursos Microbiológicos (CREM), Faculdade de Ciências e Tecnologia (FCT), Universidade Nova de Lisboa, PortugalCost Action BM0701 (ATENS), of the European Commission/European Science Foundation, Brussels, Belgium
r t i c l e i n f o
rticle history:eceived 8 July 2011ccepted 24 August 2011
eywords:almonella enterica serotype EnteritidishioridazinecrAB efflux pumpeal-time RT-PCRctivation of genes by thioridazineamAarA
a b s t r a c t
Thioridazine (TZ)-induced accumulation of the universal efflux pump substrate ethidium bromide andits subsequent efflux by Salmonella strains with various degrees of overexpressed efflux pumps takesplace automatically at pH 7.4, is independent of a metabolic source, is not affected by a proton ionophoreand is precluded by palmitic acid. Salmonella enterica serotype Enteritidis cultured in medium containingincreasing concentrations of TZ does not grow during the first 6–8 h, after which time its growth is similarto unexposed controls. At the end of a 16-h exposure period, the organism is resistant to >250 mg/L TZ.Parallel assessment by real-time reverse transcription quantitative polymerase chain reaction (RT-qPCR)of the activity of genes that regulate and code for the AcrB transporter of the main efflux pump (AcrAB) ofthe organism at periodic intervals suggests a sequence of activation beginning with the stress gene soxS,followed by the global regulator ramA, then by the local regulator marA and then by the transporter acrB.These activations take place during the period of no growth. By the end of a 16-h culture period, only
oxSobcrBmrAmrB
the acrB transporter gene is still highly overexpressed. Assessment of the activity of genes of the two-component regulon PmrA/B indicates that TZ also activates this regulon. Because activation of pmrA/Balso activates acrB, development of high resistance to TZ during a 16-h culture period is in part due toactivation of the two-component regulon.
© 2011 Elsevier B.V. and the International Society of Chemotherapy. All rights reserved.
eveloped resistance to thioridazine duringulture. Introduction
According to the US Centers for Disease Control and PreventionCDC), ‘During 1993–1997, a total of 2,751 outbreaks of food-orne disease were reported (489 in 1993, 653 in 1994, 628
n 1995, 477 in 1996, and 504 in 1997). Among outbreaks for
hich the etiology was determined, bacterial pathogens causedhe largest percentage of outbreaks (75%) and the largest per-entage of cases (86%). Salmonella enterica serotype Enteritidis
∗ Corresponding author. Tel.: +351 21 365 2600; fax: +351 21 363 2105.E-mail address: [email protected] (L. Amaral).
1 Permanent address: Institute of Pharmacognosy, Faculty of Pharmacy, Univer-ity of Szeged, Eötvös u. 6, 6720 Szeged, Hungary.
924-8579/$ – see front matter © 2011 Elsevier B.V. and the International Society of Chemoi:10.1016/j.ijantimicag.2011.08.013
accounted for the largest number of outbreaks, cases, and deaths.’[1]; 95% of Salmonella outbreaks were from food-borne sources[2]. Nosocomial Salmonella infections and outbreaks in hospitalsresult from food-borne sources, most of which can be traced tofood workers (faecal-to-hand contamination) with the remainderdue to transfer from patient sources (faecal soiling of bed sheets,etc.) to hospital personnel who transfer the organism to otherpatients, etc. [3]. Although hospital-acquired Salmonella infectionsare readily managed, when they take place in patients who havehad a resection of the colon, the organism may penetrate into themucosa of the colon and produce high mortality rates regardless
of therapy [4]. The main reason for problematic therapy lies inthe variety of responses that Salmonella activates when in a nox-ious environment, rendering the organism quite resistant to mostantibiotics.otherapy. All rights reserved.

al of A
cpoPA(mmFtetmlosihpt
cmoah
Sifeipwh[minisSwEe[puAq
2
2
fcf
2
a
G. Spengler et al. / International Journ
Amongst these responses is activation of the PmrA/B two-omponent regulon, which is activated by the low pH of thehagolysosome of the neutrophil that temporarily imprisons therganism subsequent to its phagocytosis [5]. Activation of themrA/B two-step regulon eventually leads to the synthesis of lipid, which is rapidly introduced into the nascent lipopolysaccharide
LPS) layer of the outer cell membrane [5]. The increase in LPSakes the organism resistant to practically everything, includingost antibiotics and antimicrobial agents such as bactericidins [5].
ailure of antibiotic therapy is also assured by the two-step regulonhat activates PmrD, which in turn activates the global regulator offflux pumps, the MarA operon, that eventually leads to the activa-ion of acrB, the gene coding for the transporter component of the
ain efflux pump (AcrAB) of the organism [5]. Activation of acrBeads to overexpression of the AcrAB efflux pump, bestowing therganism with a multidrug-resistant (MDR) phenotype [6]. Intrin-ic resistance of Salmonella during colonisation of the colon lies ints downregulation of porins, thereby limiting the penetration ofydrophobic antibiotics, or upregulation of its main AcrAB effluxump that extrudes the antibiotic before it reaches its intendedarget [7,8].
The intrinsic resistance of Salmonella during colonisation of theolon as well as its induced resistance to degradation by the lysoso-al machinery of the neutrophil thereby limiting the effectiveness
f current antibiotics, coupled with its high frequency in food-bornend nosocomial infections, poses a public health hazard that extolsigh economic costs and human suffering.
Although the identification of overexpressed efflux pumps inalmonella clinical isolates has been scantly reported, it is antic-pated that, as is the case for other Gram-negative bacteria, therequency of multidrug resistance attributed to overexpressedfflux pumps will increase as attention to this cause of resistancencreases. Because of the role that overexpressed efflux pumpslay in problematic therapy, we have studied the mechanisms byhich efflux pumps of Gram-negative bacteria arise [9,10] andave paid particular attention to agents that inhibit their activity7,9–11]. In addition, we have developed methods for the assess-
ent of efflux pumps of bacteria and evaluation of agents thatnhibit them [12,13]. Amongst the most potent inhibitors of Gram-egative efflux pumps are phenothiazines [14,15], amongst which
s the neuroleptic phenothiazine thioridazine (TZ). In the currenttudy, the effect of TZ on the activity of the efflux pump system of. Enteritidis strains, each one differing from their parental strainith respect to the level of efflux pumps expressed, was studied.
valuation of real-time activity was conducted by the automatedthidium bromide (EtBr) method as previously described in detail12,13]. Moreover, as a consequence of assessment of the effluxump system, the effects of TZ on the activity of genes that reg-late and code for the AcrB transporter of the main efflux pumpcrAB–TolC was also studied by real-time reverse transcriptionuantitative polymerase chain reaction (RT-qPCR).
. Materials and methods
.1. Materials
Mueller–Hinton (MH) powder (Sigma, Madrid, Spain) was usedor preparation of MH agar and MH broth. EtBr, TZ and carbonylyanide m-chlorophenylhydrazone (CCCP) were also purchasedrom Sigma.
.2. Bacteria
Salmonella Enteritidis NCTC 13349, S. Enteritidis 104 and 104CIPnd S. Enteritidis 5408 and 5408CIP were studied. Salmonella
ntimicrobial Agents 39 (2012) 16– 21 17
Enteritidis 104CIP and 5408CIP strains were derived from theirrespective parental strains by gradual exposure to ciprofloxacin,achieving high-level resistance to this antibiotic. Regarding5408CIP, resistance was due to six-fold overexpression of theacrB gene that codes for the transporter of the AcrAB–TolC effluxpump; and regarding the 104CIP, resistance was due to similaroverexpression of the AcrAB–TolC efflux pump, two mutations inthe soxS gene and a mutation in gyrase 1A [16]. Bacterial strainswere provided by S. Fanning.
2.3. Methods
Bacteria were cultured on MH agar and were incubatedovernight. Thereafter, a single colony was transferred to MH broth,incubated at 37 ◦C for 16 h and the minimum inhibitory concen-tration (MIC) of TZ was determined by microbroth dilution as perClinical and Laboratory Standards Institute (CLSI) guidelines [17].It is necessary to know the MIC of compounds to be evaluatedfor effects on the efflux pump system as it enables the amount ofcompound to yield 0.25–0.5× MIC to be determined, relative con-centrations that are known to have little or no effect on the viabilityand replication of the bacterium [11].
2.3.1. Detection of efflux pump activityDetection of efflux pump activity by the Salmonella strains was
conducted by a semi-automated fluorometric method as describedpreviously [7,12,13]. Briefly, the method follows the real-time accu-mulation of EtBr by a bacterial population using a Rotor-Gene3000TM thermocycler (Corbett Research, Sydney, Australia) pro-grammed for 30–40 cycles of 1 min each for a duration of 25 min ata constant temperature of 37 ◦C. Bacteria were first grown in MHbroth to reach an optical density at 600 nm (OD600) of 0.6. Cells werethen centrifuged, washed twice with phosphate-buffered saline(PBS) (pH 8) and the OD600 adjusted to 0.6 with PBS (pH 7.4)containing 0.4% glucose. Aliquots of 0.045 mL were transferred tomicrotubes of 0.2 mL volume and immediately aliquots of 0.045 mLof saline (pH 7.4) containing EtBr and glucose to yield final concen-trations of 1 mg/L and 0.4%, respectively, with and without varyingconcentrations of TZ at concentrations well below the MIC, wereadded. The instrument was started and real-time accumulation ofEtBr (amount of relative fluorescence emitted) was followed upto 60 min. Excitation and emission wavelengths were 530 nm and585 nm, respectively.
2.3.2. Determination of growth of Salmonella strains in theabsence and presence of thioridazine
Isolated colonies of the Salmonella strains were transferred to10 mL tubes containing increasing concentrations of TZ in MHbroth. Growth was followed spectrophotometrically at 600 nm forup to 24 h.
2.3.3. Assessment of the activity of genes that regulate and codefor the AcrB transporter by real-time reverse transcriptasepolymerase chain reaction
Isolated colonies of S. Enteritidis 104 were transferred to 10 mLtubes containing a subinhibitory concentration of TZ (100 mg/L)and at intervals of 0, 1, 4, 8, 16 and 24 h of culture at 37 ◦C thetubes were centrifuged at 1200 × g for 15 min. Pellets were sus-pended in PBS, centrifuged and re-suspended in PBS and the OD600was adjusted to 0.6. From aliquots of 1.5 mL of each tube, totalRNA was isolated in an RNase-free environment using an RNeasy
Protect Mini Kit (QIAGEN, Hilden, Germany) according to the man-ufacturer’s instructions. The integrity, purity and concentration ofthe extracted RNA templates were assessed by spectrophotome-try at 260 nm and agarose gel (1.5%). Purified RNA was stored in
1 al of Antimicrobial Agents 39 (2012) 16– 21
Ru
s(mBtQR
arc
3
2css
aaeotiioeoddfboati(alcortcTpiasgtptfpd(eE
Fig. 1. (A and B) Effect of increasing concentrations of thioridazine (TZ) on accumu-lation of 1 mg/L ethidium bromide (EtBr) by Salmonella enterica serovar EnteritidisNCTC 13349 reference strain in an assay containing 0.4% glucose (A) and in theabsence of glucose (B). (C) Effect of carbonyl cyanide m-chlorophenylhydrazone(CCCP) on TZ-promoted accumulation and efflux of EtBr in medium lacking or con-taining glucose in S. Enteritidis 104. Note: the figure presents the effect of addition ofCCCP at the peak of accumulation of EtBr in the assay. Regardless of the presence ofglucose, addition of CCCP did not prevent the efflux that followed the TZ-promotedpeak accumulation of EtBr. In contrast, addition of CCCP in the assays that did not
8 G. Spengler et al. / International Journ
Nase-free water in siliconised tubes and was maintained at −20 ◦Cntil quantification was performed.
Real-time quantification of the RNA templates by real-time one-tep RT-qPCR was performed in a Rotor-Gene 3000TM thermocyclerCorbett Research) strictly adhering to the manufacturer’s recom-
endations of the QuantiTect® SYBR® Green RT-PCR Kit (QIAGEN).riefly, each 0.2 mL standard microfuge tube contained 12.5 �L ofhe 2× QuantiTect SYBR Green RT-PCR Master Mix, 0.25 �L of 10×uantiTect RT mix, 900 nM of each primer and ca. 20 ng of totalNA in RNase-free water in a final volume of 25 �L.
The forward and reverse primers used for assessment of thectivity of the global regulator ramA, the stress genes soxS andob, the local regulator marA, the transporter acrB and the two-omponent regulon pmrA and pmrB are shown in Table 1.
. Results
The MIC of TZ against the five strains of Salmonella exceeded50 mg/L and did not vary significantly for each strain. Therefore,oncentrations of TZ that approached the MIC were selected for thetudy evaluating their effects on the efflux pump system of all fivetrains.
Because efflux at pH 7 requires a source of metabolic energy [19]nd as phenothiazines inhibit the activity of many enzymes thatre involved in the generation of metabolic energy, the study of theffects of TZ on efflux was conducted in the presence and absencef glucose in order that any effects produced by the phenothiazinehat are dependent or independent of metabolic energy could bedentified. As shown in Fig. 1A, the effect of TZ concentrationsn the assay containing glucose produces increasing accumulationf EtBr in a concentration-dependent manner by the NCTC refer-nce strain. However, at a concentration of 50 mg/L the amountf accumulation peaks after 10 min, after which time it begins toecrease (efflux) to a level that is similar to the accumulation pro-uced by TZ concentrations >20 mg/L. When glucose is omittedrom the assay medium (Fig. 1B), accumulation of EtBr promotedy 50 mg/L TZ is twice as high as that produced in the presencef glucose (relative fluorescence of 80 vs. 40 in the presence andbsence of glucose, respectively) and the peak of accumulationhat takes place at 10 min is followed by efflux, ultimately reach-ng a plateau that is equal to that noted in the presence of glucoseca. relative fluorescence of 20; Fig. 1A). Concentrations of TZ thatpproach the MIC produce increasingly greater accumulation ear-ier in time, which is followed by efflux. At a concentration that islose to the MIC, efflux ends when a level of relative fluorescencef 60 is reached, after which time the level of accumulated EtBremains unchanged for the duration of the assay. The reason forhis plateau is due to the intercalation of EtBr that takes place at orlose to the MIC (extracted DNA from bacteria exposed to 200 mg/LZ in this assay showed the presence of fluorescence that disap-ears when DNase is added; data not shown). Intercalated DNA
s not subject to efflux due to its high binding constant [13]. Theutomatic efflux promoted by TZ in the absence of glucose sub-equent to the peak of EtBr accumulation was indeed surprisingiven that efflux by Salmonella [20] and other Gram-negative bac-eria is dependent upon metabolic energy in an assay where theH is ≥7 [7,12,13]. Another surprise was that CCCP, an ionophorehat by binding protons promotes the collapse of the proton-motiveorce (PMF) thereby inhibiting the activity of PMF-dependent effluxumps, when added at the peak of accumulation promoted by TZ
oes not affect the efflux that takes place in glucose-free salineFig. 1C). However, as is the case with Escherichia coli [21], the pres-nce of calcium in the assay system prevents the accumulation oftBr produced by TZ.contain TZ inhibited efflux regardless of the presence of glucose. The data have beennormalised relative to the curves of each set (with and without TZ) in order that thecomparison is more readily made.

G. Spengler et al. / International Journal of Antimicrobial Agents 39 (2012) 16– 21 19
Table 1Forward and reverse primers used for assessment of the activity of the global regulator ramA, the stress genes soxS and rob, the local regulator marA, the transporter acrB andthe two-component regulon pmrA and pmrB of Salmonella enterica serovar Enteritidis 104.
Gene Primer sequence(5′ → 3′)
Amplicon size (bp) Reference
16S rRNA Fw GCGGCAGGCCTAACACATRv GCAAGAGGCCCGAACGTC
182 [16]
ramA Fw CGTCATGCGGGGTATTCCAAGTGRv CGCGCCGCCAGTTTTAGC
107 [16]
marA Fw ATCCGCAGCCGTAAAATGACRv TGGTTCAGCGGCAGCATATA
180 [16]
soxS Fw AAATCGGGCTACTCCAAGTGRv CTACAGGCGGTGACGGTAAT
217 [16]
rob Fw CATTACGGCTGGGCGAGTTTACCRv CTGGCGGAATAGTTGGCGAATGAC
180 [16]
acrB Fw TTTTGCAGGGCGCGGTCAGAATACRv TGCGGTGCCCAGCTCAACGAT
184 [16]
pmrA Fw CGCGCAAACCGAAGGCTATG 192 This study
29
edtAnwo1entrorttp
fhEicadalastoaa
lttst[i[dCC
component of the regulon, pmrA. Activation of the PmrA/B regulonis known to activate acrB by activating pmrD [5]. Because activationof the PmrA/B regulon takes place under a very acid pH such as that
Fig. 2. Effect of palmitic acid on thioridazine (TZ)-promoted accumulation and efflux
Rv GTATCGCGGGCGGTCAGAATGpmrB Fw GCGTGGGCCAGTCATTCTCTTCC
Rv ATCAATACCCGGCCCCTCGTCTT
Previous results demonstrated that at pH 8 the activity of thefflux pump system of the Salmonella NCTC strain was totallyependent upon metabolic energy [20]. Therefore, we examinedhe effect of TZ on another MDR strain S. Enteritidis 5408 whosecrAB–TolC efflux pump is of an intrinsic type, i.e. it is present at aormal basal level of activity and may be subject to overexpressionhen the organism is exposed to a noxious agent [6]. At the high pH
f 8, TZ promotes the exact same response from the S. Enteritidis04 and 5408 strains as that evident by the Salmonella NCTC refer-nce strain when glucose is present and absent in the medium (dataot shown). In contrast, the effects of TZ on accumulation of EtBr byhe progeny of 104 and 5408 that have been induced to high-levelesistance to ciprofloxacin (104CIP and 5408CIP), do not yield a peakf accumulation that is followed by efflux (data not shown). Theseesults suggest that the overexpressed AcrAB–TolC efflux pumps ofhese strains readily extrude the phenothiazine before it reacheshe targets that promote accumulation of EtBr as is the case for thearents and for the NCTC wild-type strains.
Since metabolic energy provides the needed protons for theunction of an efflux pump at the high of pH 8 [20] and as Salmonellaas the ability to utilise fatty acids in place of glucose [22,23], thetBr assay was repeated with a concentration of 50 mg/L TZ andncreasing concentrations of palmitic acid added in place of glu-ose. As noted in Fig. 2, with increasing concentrations of palmiticcid the TZ-promoted accumulation of EtBr is proportionatelyecreased. These results are interpreted to signify that palmiticcid provides the needed energy for efflux, hence little accumu-ation of EtBr results. Moreover, because TZ inhibits enzymes thatre involved in the generation of energy from metabolic sourcesuch as glucose and their dependence on calcium [21], and becausehe effects of TZ on the accumulation of EtBr are reduced or evenbviated with palmitic acid, it may well be that the enzymes thatct on the fatty acid for the generation of energy are either notffected by the phenothiazine or are beyond its reach.
The TZ-promoted increased accumulation of EtBr that is fol-owed by efflux may be the reason for the resistance of Salmonella tohe phenothiazine. Exposure to chlorpromazine (CPZ), a neurolep-ic from which TZ is derived, inhibits the replication of S. entericaerotype Typhimurium during the first 6–8 h of culture, after whichhe organism becomes increasingly resistant to this phenothiazine24]. Others have shown that multiples of the MIC of this phenoth-azine affect genes that regulate and code for the transporter AcrB
25]. Because the latter study did not evaluate the activity of genesuring the entire culture period of exposure to the phenothiazinePZ, and because the activity of TZ on bacteria is similar to that ofPZ, we have examined the effects of increasing concentrations of9 This study
TZ on the growth of S. Enteritidis 104 and, at intervals during theculture, assessed the expression of genes that regulate and code forthe AcrB transporter.
As seen in Fig. 3, during the first 6–8 h the organism is not grow-ing. However, during this period of no growth, a sequence of genesare overexpressed compared with their control counterparts (noTZ). Briefly, by the end of 1 h of culture, the stress gene soxS is over-expressed more than five-fold compared with that of the control(no TZ) and returns to normal levels of expression by the end of4 h. The stress gene rob is not activated during the entire culture.Activation of soxS is followed by activation of the global regulatorramA, and this is followed later in the culture by activation of marA.By the end of 8 h of culture, the acrB gene that codes for the trans-porter is now overexpressed by more than 40-fold. By the end of 8 hof exposure to TZ, the organism is now able to extrude the noxiousagent and achieve growth at its normal rate. Interestingly, pmrB,the sensor gene of the two-component regulon that bestows resis-tance in Salmonella [5], is significantly overexpressed by the endof 4 h and this is followed hours later by activation of the second
of ethidium bromide (EtBr) by Salmonella enterica serovar Enteritidis NCTC 13349in medium lacking or containing glucose or palmitic acid. Note: in this assay, theconcentration of glucose was 0.6% and the effects of TZ on accumulation of EtBr werenot obviated. However, in glucose-free medium, addition of palmitic acid inhibitedthe effect of TZ on accumulation in a concentration-dependent manner.
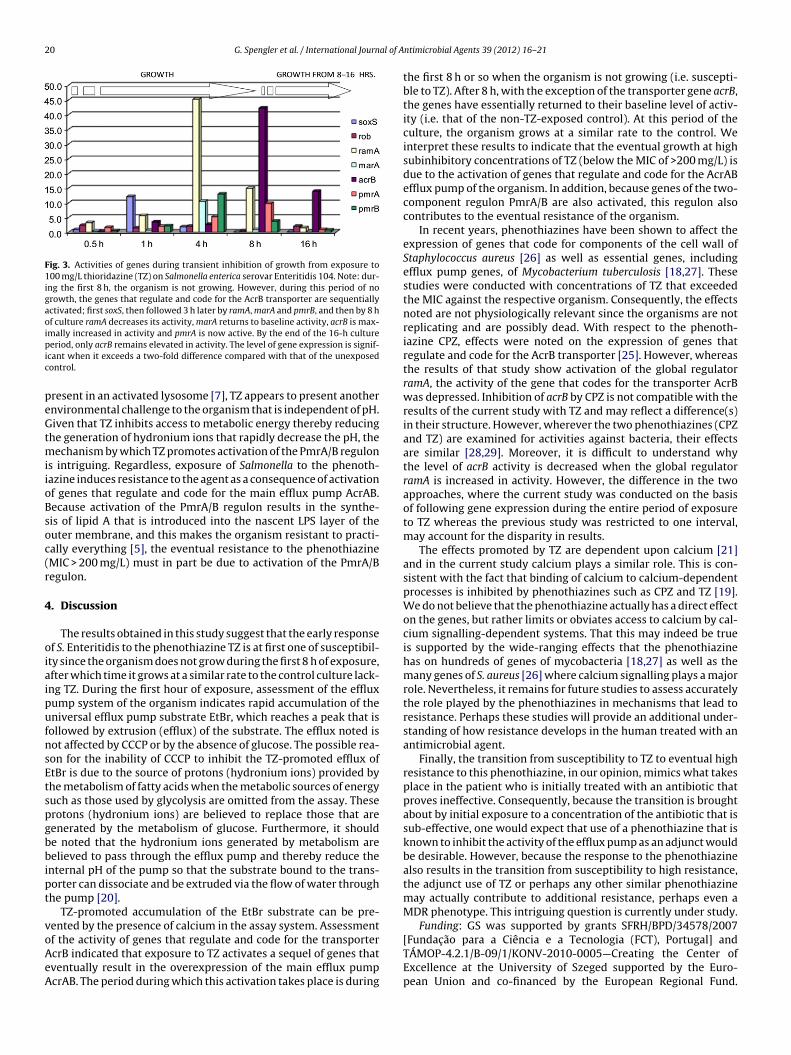
20 G. Spengler et al. / International Journal of A
Fig. 3. Activities of genes during transient inhibition of growth from exposure to100 mg/L thioridazine (TZ) on Salmonella enterica serovar Enteritidis 104. Note: dur-ing the first 8 h, the organism is not growing. However, during this period of nogrowth, the genes that regulate and code for the AcrB transporter are sequentiallyactivated; first soxS, then followed 3 h later by ramA, marA and pmrB, and then by 8 hof culture ramA decreases its activity, marA returns to baseline activity, acrB is max-imally increased in activity and pmrA is now active. By the end of the 16-h culturepic
peGtmiioBsoc(r
4
oiaipufnsEtspgbbipt
voAeA
eriod, only acrB remains elevated in activity. The level of gene expression is signif-cant when it exceeds a two-fold difference compared with that of the unexposedontrol.
resent in an activated lysosome [7], TZ appears to present anothernvironmental challenge to the organism that is independent of pH.iven that TZ inhibits access to metabolic energy thereby reducing
he generation of hydronium ions that rapidly decrease the pH, theechanism by which TZ promotes activation of the PmrA/B regulon
s intriguing. Regardless, exposure of Salmonella to the phenoth-azine induces resistance to the agent as a consequence of activationf genes that regulate and code for the main efflux pump AcrAB.ecause activation of the PmrA/B regulon results in the synthe-is of lipid A that is introduced into the nascent LPS layer of theuter membrane, and this makes the organism resistant to practi-ally everything [5], the eventual resistance to the phenothiazineMIC > 200 mg/L) must in part be due to activation of the PmrA/Begulon.
. Discussion
The results obtained in this study suggest that the early responsef S. Enteritidis to the phenothiazine TZ is at first one of susceptibil-ty since the organism does not grow during the first 8 h of exposure,fter which time it grows at a similar rate to the control culture lack-ng TZ. During the first hour of exposure, assessment of the effluxump system of the organism indicates rapid accumulation of theniversal efflux pump substrate EtBr, which reaches a peak that isollowed by extrusion (efflux) of the substrate. The efflux noted isot affected by CCCP or by the absence of glucose. The possible rea-on for the inability of CCCP to inhibit the TZ-promoted efflux oftBr is due to the source of protons (hydronium ions) provided byhe metabolism of fatty acids when the metabolic sources of energyuch as those used by glycolysis are omitted from the assay. Theserotons (hydronium ions) are believed to replace those that areenerated by the metabolism of glucose. Furthermore, it shoulde noted that the hydronium ions generated by metabolism areelieved to pass through the efflux pump and thereby reduce the
nternal pH of the pump so that the substrate bound to the trans-orter can dissociate and be extruded via the flow of water throughhe pump [20].
TZ-promoted accumulation of the EtBr substrate can be pre-ented by the presence of calcium in the assay system. Assessment
f the activity of genes that regulate and code for the transportercrB indicated that exposure to TZ activates a sequel of genes thatventually result in the overexpression of the main efflux pumpcrAB. The period during which this activation takes place is duringntimicrobial Agents 39 (2012) 16– 21
the first 8 h or so when the organism is not growing (i.e. suscepti-ble to TZ). After 8 h, with the exception of the transporter gene acrB,the genes have essentially returned to their baseline level of activ-ity (i.e. that of the non-TZ-exposed control). At this period of theculture, the organism grows at a similar rate to the control. Weinterpret these results to indicate that the eventual growth at highsubinhibitory concentrations of TZ (below the MIC of >200 mg/L) isdue to the activation of genes that regulate and code for the AcrABefflux pump of the organism. In addition, because genes of the two-component regulon PmrA/B are also activated, this regulon alsocontributes to the eventual resistance of the organism.
In recent years, phenothiazines have been shown to affect theexpression of genes that code for components of the cell wall ofStaphylococcus aureus [26] as well as essential genes, includingefflux pump genes, of Mycobacterium tuberculosis [18,27]. Thesestudies were conducted with concentrations of TZ that exceededthe MIC against the respective organism. Consequently, the effectsnoted are not physiologically relevant since the organisms are notreplicating and are possibly dead. With respect to the phenoth-iazine CPZ, effects were noted on the expression of genes thatregulate and code for the AcrB transporter [25]. However, whereasthe results of that study show activation of the global regulatorramA, the activity of the gene that codes for the transporter AcrBwas depressed. Inhibition of acrB by CPZ is not compatible with theresults of the current study with TZ and may reflect a difference(s)in their structure. However, wherever the two phenothiazines (CPZand TZ) are examined for activities against bacteria, their effectsare similar [28,29]. Moreover, it is difficult to understand whythe level of acrB activity is decreased when the global regulatorramA is increased in activity. However, the difference in the twoapproaches, where the current study was conducted on the basisof following gene expression during the entire period of exposureto TZ whereas the previous study was restricted to one interval,may account for the disparity in results.
The effects promoted by TZ are dependent upon calcium [21]and in the current study calcium plays a similar role. This is con-sistent with the fact that binding of calcium to calcium-dependentprocesses is inhibited by phenothiazines such as CPZ and TZ [19].We do not believe that the phenothiazine actually has a direct effecton the genes, but rather limits or obviates access to calcium by cal-cium signalling-dependent systems. That this may indeed be trueis supported by the wide-ranging effects that the phenothiazinehas on hundreds of genes of mycobacteria [18,27] as well as themany genes of S. aureus [26] where calcium signalling plays a majorrole. Nevertheless, it remains for future studies to assess accuratelythe role played by the phenothiazines in mechanisms that lead toresistance. Perhaps these studies will provide an additional under-standing of how resistance develops in the human treated with anantimicrobial agent.
Finally, the transition from susceptibility to TZ to eventual highresistance to this phenothiazine, in our opinion, mimics what takesplace in the patient who is initially treated with an antibiotic thatproves ineffective. Consequently, because the transition is broughtabout by initial exposure to a concentration of the antibiotic that issub-effective, one would expect that use of a phenothiazine that isknown to inhibit the activity of the efflux pump as an adjunct wouldbe desirable. However, because the response to the phenothiazinealso results in the transition from susceptibility to high resistance,the adjunct use of TZ or perhaps any other similar phenothiazinemay actually contribute to additional resistance, perhaps even aMDR phenotype. This intriguing question is currently under study.
Funding: GS was supported by grants SFRH/BPD/34578/2007
[Fundac ão para a Ciência e a Tecnologia (FCT), Portugal] andTÁMOP-4.2.1/B-09/1/KONV-2010-0005—Creating the Center ofExcellence at the University of Szeged supported by the Euro-pean Union and co-financed by the European Regional Fund.
al of A
LSspLbUMf
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
G. Spengler et al. / International Journ
R and SSC were supported by grants SFRH/BD/24931/2005 andFRH/BD/44214/2008, respectively, provided by FCT. MMa wasupported by grant SFRH/BPD/63871/2009 (FCT). EN was sup-orted by an STSM grant from COST ACTION BM0701 (ATENS).A was supported by BCC grant SFRH/BCC/51099/2010 providedy the FCT, and PTDC/SAU-FCF/102807/2008 provided by thePMM. This work was supported by EU-FSE/FEDER-PTDC/BIA-IC/105509/2008 and EU-FSE/FEDERPTDC/SAU-FCF/102807/2008
rom the FCT.Competing interests: None declared.Ethical approval: Not required.
eferences
[1] Olsen SJ, MacKinnon LC, Goulding JS, Bean NH, Slutsker L. Surveillance forfoodborne-disease outbreaks—United States, 1993–1997. MMWR CDC SurveillSumm 2000;49:1–62.
[2] Mead PS, Slutsker L, Dietz V, McCaig LF, Bresee JS, Shapiro C, et al. Food-relatedillness and death in the United States. Emerg Infect Dis 1999;5:607–25.
[3] Todd EC, Greig JD, Bartleson CA, Michaels BS. Outbreaks where food work-ers have been implicated in the spread of foodborne disease. Part 5. Sourcesof contamination and pathogen excretion from infected persons. J Food Prot2008;71:2582–95.
[4] Buzio M, Shoshtari Mahmoud K, Memore L, Cotogni P. Perforation of the smallintestine. Minerva Chir 1999;54:851–4 [in Italian].
[5] Gunn JS. The Salmonella PmrAB regulon: lipopolysaccharide modifications,antimicrobial peptide resistance and more. Trends Microbiol 2008;16:284–90.
[6] Amaral L, Viveiros M, Fanning S, Pages JM, Couto I, Spengler G, et al. Geneticregulation, physiology, assessment and inhibition of efflux pumps responsi-ble for multi-drug resistant phenotypes of bacterial pathogens. In: Bonilla AR,Muniz KP, editors. Antibiotic resistance: causes and risk factors, mechanismsand alternatives. New York, NY: Nova Science Publishers, Inc.; 2009. p. 313–32.
[7] Martins A, Spengler G, Rodrigues L, Viveiros M, Ramos J, Martins M, et al. pHmodulation of efflux pump activity of multi-drug resistant Escherichia coli: pro-tection during its passage and eventual colonization of the colon. PLoS One2009;4:e6656.
[8] Randall LP, Woodward MJ. Multiple antibiotic resistance (mar) locus inSalmonella enterica serovar Typhimurium DT104. Appl Environ Microbiol2001;67:1190–7.
[9] Viveiros M, Rodrigues L, Dupont M, Martins M, Couto I, Pagès JM, et al. Antibi-otic stress, genetic response and altered permeability of E. coli. PLoS One2007;2:e365.
10] Martins A, Iversen C, Rodrigues L, Spengler G, Ramos J, Kern WV, et al. An AcrAB-mediated multidrug-resistant phenotype is maintained following restorationof wild-type activities by efflux pump genes and their regulators. Int J Antimi-crob Agents 2009;34:602–4.
11] Pagès JM, Amaral L. Mechanisms of drug efflux and strategies to combat them:
challenging the efflux pump of Gram-negative bacteria. Biochim Biophys Acta2009;1794:826–33.12] Viveiros M, Rodrigues L, Martins M, Couto I, Spengler G, Martins A, et al. Eval-uation of efflux activity of bacteria by a semi-automated fluorometric system.Methods Mol Biol 2010;642:159–72.
[
ntimicrobial Agents 39 (2012) 16– 21 21
13] Viveiros M, Martins A, Paixão L, Rodrigues L, Martins M, Couto I, et al.Demonstration of intrinsic efflux activity of Escherichia coli K-12 AG100by an automated ethidium bromide method. Int J Antimicrob Agents2008;31:458–62.
14] Kristiansen JE, Thomsen VF, Martins A, Viveiros M, Amaral L. Non-antibiotics reverse resistance of bacteria to antibiotics. In Vivo 2010;24:751–4.
15] Takács D, Cerca P, Martins A, Riedl Z, Hajós G, Molnár J, et al. Evaluation of fortynew phenothiazine derivatives for activity against intrinsic efflux pump sys-tems of reference Escherichia coli, Salmonella Enteritidis, Enterococcus faecalisand Staphylococcus aureus strains. In Vivo 2011;25:719–24.
16] O’Regan E, Quinn T, Pagès JM, McCusker M, Piddock L, Fanning S. Multipleregulatory pathways associated with high-level ciprofloxacin and mul-tidrug resistance in Salmonella enterica serovar Enteritidis: involvement ofRamA and other global regulators. Antimicrob Agents Chemother 2009;53:1080–7.
17] Clinical and Laboratory Standards Institute. Performance standards forantimicrobial susceptibility testing; seventeenth informational supplement.Document M100-S17. Wayne, PA: CLSI; 2007.
18] Dutta NK, Mazumdar K, Dastidar SG, Karakousis PC, Amaral L. New patentableuse of an old neuroleptic compound thioridazine to combat tuberculosis:a gene regulation perspective. Recent Pat Antiinfect Drug Discov 2011;6:128–38.
19] Amaral L, Viveiros M, Molnar J. Antimicrobial activity of phenothiazines. In Vivo2004;18:725–31.
20] Amaral L, Cerca P, Spengler G, Machado L, Couto I, Viveiros M, et al. Ethidiumbromide efflux by Salmonella: modulation by metabolic energy, pH, ions andphenothiazines. Int J Antimicrob Agents 2011;38:140–5.
21] Martins A, Machado L, Costa S, Cerca P, Spengler G, Viveiros M, et al. Role ofcalcium in efflux system of Escherichia coli. Int J Antimicrob Agents 2011;37:410–4.
22] Evans MR, Fink RC, Vazquez-Torres A, Porwollik S, Jones-Carson J, McClellandM, et al. Analysis of the ArcA regulon in anaerobically grown Salmonella entericasv. Typhimurium. BMC Microbiol 2011;11:58.
23] Calhoun LN, Liyanage R, Lay Jr JO, Kwon YM. Proteomic analysis of Salmonellaenterica serovar Enteritidis following propionate adaptation. BMC Microbiol2010;10:249.
24] Amaral L, Kristiansen JE, Frolund Thomsen V, Markovich B. The effects ofchlorpromazine on the outer cell wall of Salmonella typhimurium in ensuringresistance to the drug. Int J Antimicrob Agents 2000;14:225–9.
25] Bailey AM, Paulsen IT, Piddock LJ. RamA confers multidrug resistance inSalmonella enterica via increased expression of acrB, which is inhibited by chlor-promazine. Antimicrob Agents Chemother 2008;52:3604–11.
26] Bonde M, Højland DH, Kolmos HJ, Kallipolitis BH, Klitgaard JK. Thioridazineaffects transcription of genes involved in cell wall biosynthesis in methicillin-resistant Staphylococcus aureus. FEMS Microbiol Lett 2011;318:168–76.
27] Dutta NK, Mehra S, Kaushal D. A Mycobacterium tuberculosis sigma factor net-work responds to cell-envelope damage by the promising anti-mycobacterialthioridazine. PLoS One 2010;5:e10069.
28] Amaral L, Kristiansen JE, Abebe LS, Millett W. Inhibition of the respiration ofmulti-drug resistant clinical isolates of Mycobacterium tuberculosis by thior-
idazine: potential use for initial therapy of freshly diagnosed tuberculosis. JAntimicrob Chemother 1996;38:1049–53.29] Bettencourt MV, Bosne-David S, Amaral L. Comparative in vitro activity ofphenothiazines against multidrug-resistant Mycobacterium tuberculosis. Int JAntimicrob Agents 2000;16:69–71.
Related Documents











