Identification of genes conferring stress tolerance Page numbers not for citation purposes. 1 Bioresource Technology Genetic resources of extremotolerant fungi: a method for identification of genes conferring stress tolerance Cene Gostinčar a,b , Nina Gunde-Cimerman a,b , Martina Turk a * a University of Ljubljana, Biotechnical Faculty, Department of Biology, Večna pot 111, SI- 1000 Ljubljana, Slovenia b Centre of Excellence for Integrated Approaches in Chemistry and Biology of Proteins (CIPKeBiP), Jamova 39, SI-1000 Ljubljana, Slovenia *Corresponding author: Martina Turk Department of Biology Biotechnical Faculty University of Ljubljana Večna pot 111 SI-1000 Ljubljana, Slovenia Tel: +386-1-3203392; Fax: +386-1-2573390 Email: [email protected] Cene Gostinčar email: [email protected]-lj.si, [email protected] Nina Gunde-Cimerman email: [email protected] Please note that this is the “revised personal version of the text of the final journal article”, published according to the copyright policy of the publisher. If you would like to receive a reprint of the final article (the content of which is identical to this file, with the exception of the formatting), please contact the corresponding author or try to access the final article on the official website of the publisher. Cite this article as: Gostinčar, C., Gunde-Cimerman, N., Turk, M., 2012. Genetic resources of extremotolerant fungi: A method for identification of genes conferring stress tolerance. Bioresource Technology 111, 360–367.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 1
Bioresource Technology
Genetic resources of extremotolerant fungi: a method for
identification of genes conferring stress tolerance
Cene Gostinčara,b, Nina Gunde-Cimermana,b, Martina Turka*
a University of Ljubljana, Biotechnical Faculty, Department of Biology, Večna pot 111, SI-
1000 Ljubljana, Slovenia b Centre of Excellence for Integrated Approaches in Chemistry and Biology of Proteins
(CIPKeBiP), Jamova 39, SI-1000 Ljubljana, Slovenia
*Corresponding author: Martina Turk Department of Biology Biotechnical Faculty University of Ljubljana Večna pot 111 SI-1000 Ljubljana, Slovenia Tel: +386-1-3203392; Fax: +386-1-2573390 Email: [email protected]
Cene Gostinčar email: [email protected], [email protected] Nina Gunde-Cimerman email: [email protected]
Please note that this is the “revised personal version of the text of the final journal article”,
published according to the copyright policy of the publisher. If you would like to receive a
reprint of the final article (the content of which is identical to this file, with the exception of
the formatting), please contact the corresponding author or try to access the final article on
the official website of the publisher.
Cite this article as:
Gostinčar, C., Gunde-Cimerman, N., Turk, M., 2012. Genetic resources of extremotolerant fungi: A method for identification of genes conferring stress tolerance. Bioresource Technology 111, 360–367.

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 2
Abstract
Fungal species from extreme environments represent an underexploited source of stress-
resistance genes. These genes have the potential to improve stress tolerance of
economically important microorganisms and crops. An efficient high-throughput method for
the identification of biotechnologically interesting genes of extremotolerant fungi was
developed by constructing a cDNA expression library in Saccharomyces cerevisiae and
screening for gain-of-function transformants under stress conditions. The advantages and
possible modifications of this method are discussed, and its efficiency is demonstrated using
the stress-tolerant basidiomycetous yeast Rhodotorula mucilaginosa. Twelve R.
mucilaginosa genes are described that increase halotolerance in S. cerevisiae. These
include genes encoding a phosphoglucomutase and a phosphomannomutase. All twelve
investigated genes might be useful for the improvement of halotolerance in genetically
modified crops or industrial microorganisms.
Keywords: functional screening; gain-of-function method; stress-tolerance genes; stress-tolerant
fungi; halotolerance
Abbreviations: YNB – Yeast Nitrogen Base medium, CSM – Complete amino acid Supplement
Mixture, CSM-Ura – Complete amino acid Supplement Mixture without uracil, YPD – Yeast Peptone
Dextrose medium, LB – Luria - Bertani medium, SOC - Super Optimal broth with Catabolite repression
medium, MIC – Minimum Inhibitory Concentration, CDS – Coding Sequence
1. Introduction
Dehydration is a major stress factor for organisms living at subzero temperatures as
extracellular freezing leads to cell dehydration and reduced water absorption. When the ice
melts, the organisms are suddenly exposed to an abundance of water, a change that
provides an advantage to species that can adapt to a range of water activities. Similar stress
is encountered in hypersaline environments, where water activity is low, but can increase
quickly during rainfall. It is therefore not surprising that cellular adaptations to low
temperature and high salinity are often alike. Similar selection pressures in these
environments result in a significant overlap of fungal diversity (Gunde-Cimerman et al.,
2003), characterised by well-adapted species with specialised stress-tolerance mechanisms
(Gostinčar et al., 2011). One of the prominent representatives of these stress-tolerant
communities is the basidiomycetous yeast, Rhodotorula mucilaginosa. This species is
commonly found in cold and hypersaline environments, as well as in the deep sea and in
other unusual habitats (reviewed in Gostinčar et al., 2011). Due to its remarkable ability to
withstand a wide variety of stress conditions, R. mucilaginosa is a promising source of genes
that can confer stress tolerance.
Stress tolerance is a highly desirable trait in economically important organisms. Abiotic stress
can cause changes in morphology, physiology and biochemistry, affects growth and
productivity, and decreases yields. Even traditional processes, such as dough fermentation
and the production of Saccharomyes cerevisiae biomass, can expose yeast cells to freeze-
thaw, high sugar concentrations, air-drying, and oxidative stress (Shima and Takagi, 2009).

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 3
Similarly, crops are often exposed to drought, temperature extremes, and saline soils
(Bhatnagar-Mathur et al., 2008).
The production of ethanol as a biofuel and other industrial processes have increased the
need for improvements to multi-stress tolerance of S. cerevisiae and other microorganisms
(Zheng et al., 2010). Similarly, the need for increased food production calls for further
optimisation of crop yields. Salinisation of soil and lack of freshwater due to climate change
can increase drought frequency and the need for irrigation (Rozema and Flowers, 2008). The
connection between several abiotic stress factors and water deficit or osmotic stress in cells
(Holmberg and Bulow, 1998) further underlines the importance of osmotolerance.
Conventional breeding methods have so far failed to provide high levels of salt and drought
tolerance in crops (Rai et al., 2010). Therefore, genetic modifications remain the most
promising option (Bhatnagar-Mathur et al., 2008). While some moderate achievements have
already been made towards improvements in stress tolerance of some organisms, further
efforts are needed (Munns, 2002). These will have to involve elucidation of stress-tolerance
mechanisms and identification of genes that confer stress tolerance.
Several high-throughput approaches have been used to identify genes that are involved in
stress resistance, from microarrays, expressed sequence tags and transcriptome
sequencing, to two-dimensional protein electrophoresis, and others. These methods are
extremely important in basic research of stress responses. In biotechnology, however,
functional gene screening (which is also referred to as expression cloning) offers an
important advantage since it is targeted directly towards useful genes that can increase
stress tolerance of acceptor organisms. Functional gene screening does not require any prior
knowledge about the genome of the donor, and it is useful in cases of unsequenced
genomes, non-axenic cultures, and even whole communities.
There have been several studies that have used a variety of donor organisms and screening
systems. These have included: a plant cDNA library (Pisum sativum) in a bacterial screening
system (Escherichia coli; Joshi et al., 2009); a plant cDNA library (Thellungiella halophila) in
a fungal screening system (Schizosaccharomyces pombe; Chen et al., 2007); a plant cDNA
library (T. halophila) in a plant screening system (Arabidopsis thaliana; Du et al., 2008); a
fungal genomic library (Debaryomyces hansenii) in a yeast screening system (S. cerevisiae;
Prista et al., 2002); and even a metagenomic DNA library in a bacterial screening system (E.
coli; Kapardar et al., 2010). Neither these nor others have – to our knowledge – screened a
cDNA library of a halotolerant fungus in a fungal screening system.
Novel sources of stress-tolerance genes need to be considered to facilitate the engineering
of enhanced stress tolerance in various organisms (Somvanshi, 2009). Fungi have long been
accepted as a good source of extracellular enzyme encoding genes (Dalboge, 1997),
although in other fields their resources have been less well exploited. Patents concerning
fungal drought tolerance are few, and focused on S. cerevisiae (Somvanshi, 2009). However,
some other fungal species are much more tolerant to a variety of extreme stress conditions
(Gunde-Cimerman et al., 2003). We believe that the genetic resources of these fungi need to
be investigated for their potential to be used to improve the stress tolerance of economically
important fungi and plants.

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 4
Patents concerning fungal drought tolerance are few, and focused on S. cerevisiae
(Somvanshi, 2009) although some other fungal species are much more tolerant to a variety
of extreme stress conditions (Gunde-Cimerman et al., 2003). Therefore, in the current paper,
a method for the identification of genes from stress-tolerant fungi that can increase the stress
tolerance of recipient organisms is described. This convenient high-throughput functional
screening methodology enables the investigation of whole transcriptomes in S. cerevisiae in
short periods of time. The method facilitates identification of biotechnologically promising
genes, in our case, genes for improving salt tolerance. The effectiveness of this approach
was demonstrated by identifying 12 genes from the halotolerant basidiomycetous yeast, R.
mucilaginosa, that improved salt tolerance when expressed in S. cerevisiae. These include
the genes encoding a phosphoglucomutase and phosphomannomutase. The activities of the
corresponding proteins were investigated.
2. Materials and methods
2.1 Media, strains and growth conditions
The halotolerant basidiomycetous yeast, R. mucilaginosa (strain EXF-1630), was isolated
from an Arctic glacier in Kongsfjorden, Spitsbergen, Svalbard (Norway) and maintained in the
Ex-Culture Collection of the Department of Biology, Biotechnical Faculty, University of
Ljubljana (Slovenia). It was cultivated using the chemically defined medium Yeast Nitrogen
Base (YNB, Qbiogene), with 0.5% ammonium sulphate (w/v) and 2% glucose (w/v). Liquid
cultures were grown at 28 °C on a rotary shaker at 180 rpm.
S. cerevisiae W303a was cultivated using YNB medium (Qbiogene), with 0.5% ammonium
sulphate (w/v), 0.8% Complete Amino Acid Supplement Mixture (CSM) with/without uracil
(w/v; Qbiogene), and with 2% glucose (w/v). In the induction medium, glucose was
substituted with filter-sterilised 2% galactose (w/v) and 1% raffinose (w/v). The screening
media were prepared by adding the minimum inhibitory concentrations (MICs) of various
osmolytes to the induction medium: 1.37 M or 1.71 M NaCl, 2.7 M sorbitol, 3.4 M glycerol, or
0.4 M LiCl. The selection medium for curing of the plasmids with the URA3 selection marker
was prepared by autoclaving 20 g agar (Difco) in 750 ml distilled water, and mixing with 250
ml of a filter-sterilised solution of 5 g ammonium sulphate, 1.7 g YNB (Qbiogene), 20 g
glucose, 0.8 g CSM (Qbiogene), 40 mg/l final concentration of Uracil, and 1 g 5-fluoro-orotic
acid. The pH of the medium was not adjusted. Yeast Peptone Dextrose (YPD) medium was
prepared with 1% yeast extract, 2% peptone, 2% glucose, and 2% agar (all w/v) (Difco; in the
case of solid medium). The pH was adjusted to 6.5 with 1 M NaOH, and the medium was
sterilised by autoclaving.
E. coli (ElectroMAXTM DH10BTM T1 Phage Resistant cells; Invitrogen) was grown at 37 °C
on Luria-Bertani medium (LB, Merck) with the addition of the appropriate antibiotics. Liquid
cultures were grown at 37 °C on a rotary shaker at 200 rpm.
2.2 RNA isolation
For RNA isolation, R. mucilaginosa was grown in YNB liquid medium with 10% NaCl (w/v),
and harvested by centrifugation at mid-exponential growth phase (OD600 = 0.8-1.0). The
pellet was frozen in liquid nitrogen and homogenised using a pestle and mortar. Total RNA
was isolated from 500 mg of homogenised biomass using the TRI REAGENTTM (Sigma),
according to the manufacturer instructions. Poly(A)RNA was isolated using the FastTrack 2.0

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 5
mRNA Isolation kits (Invitrogen). The integrity and purity of the total RNA and the isolated
mRNA were evaluated with an RNA/DNA Bioanalyser (Agilent).
2.3 Library construction in E. coli
The cDNA library was constructed using the CloneMinerTM cDNA Library Construction kit
(Invitrogen), using the non-radiolabelling method and according to the manufacturer
instructions, with minor modifications. Briefly, cDNA was synthesised from 5 g poly(A)RNA
using the Biotin-attB2-Oligo(dT) Primer and SuperScript II RT. The products of the reaction
were used for second-strand cDNA synthesis using E. coli DNA polymerase I, E. coli DNA
ligase, and E. coli RNase H, followed by ligation of the attB1 adapter at the 5'-end. The cDNA
was size fractionated by column chromatography, to remove excess primers, adapters and
small cDNAs. The concentration and yield of the cDNA fractions were estimated using a
NanoDrop 2000 instrument (Thermo Scientific). Then, 150 ng of the cDNA was pooled,
precipitated, and used for the BP recombination reaction between the attB-flanked cDNA and
the entry vector pDONRTM222. The products of the reaction were electroporated into
ElectroMAXTM DH10BTM T1 Phage Resistant cells (Invitrogen), at 1,700 V with an
Electroporator 2510 (Eppendorf). The library was mixed with the freezing medium (40%
glycerol, 60% Super Optimal broth with catabolite repression (SOC) medium; Invitrogen),
and frozen. The titre of the library was determined by serial dilutions in SOC medium (10-2 to
10-4), plating on prewarmed LB plates containing kanamycin (50 mg/l), and counting of the
colonies after overnight incubation at 37 °C. The integrity of the inserts was estimated by
sequencing of 20 randomly picked clones.
2.4 Library construction in S. cerevisiae
An aliquot of the library in the entry vector pDONRTM222 that contained approximately 6.5 ×
106 transformants was inoculated into 50 ml liquid LB medium with 50 mg/l kanamycin, and
grown on a rotary shaker at 200 rpm and 37 °C. When the culture reached an OD600 of 1.0,
total plasmid DNA was isolated with the GenElute High Performance Plasmid MidiPrep Kit
(Sigma). The DNA was precipitated with the addition of 1 μl glycogen (20 mg/ml, Fermentas),
0.1 vol. 7.5 M ammonium acetate and 2.5 vol. ice-cold absolute ethanol to 1 ml of the
sample, and then centrifuged at 20,000 × g for 25 min at 4 °C. The pellet was washed twice
with 150 ml ice-cold 70% ethanol and centrifuged at 20,000 × g for 2 min at 4 °C. The
pelleted DNA was air-dried (10 min at room temperature) and dissolved in 30 μl TE buffer (10
mM Tris-HCl, pH 8, 1 mM EDTA). Then 50 ng plasmid DNA diluted to 7 μl with TE buffer was
used for the LR recombination reaction, with 450 ng pYES-DEST52 GatewayTM vector
(Invitrogen), 4 μl 5 × LR Clonase Reaction Buffer, and 6 μl LR ClonaseTM II (Invitrogen). The
reaction was performed at 25 °C for 1 h, and terminated with 1 μl Proteinase K, and an
incubation at 30 °C for 15 min, followed by an incubation at 75 °C for 10 min. The DNA was
precipitated and washed as described above, and resuspended in 9 μl TE buffer. The
products of the reaction were electroporated into ElectroMAXTM DH10BTM T1 Phage
Resistant cells (Invitrogen) in six aliquots (1.5 μl DNA suspension and 50 μl cell suspension),
as described above; the library was mixed with freezing medium, and frozen. The titre of the
library was determined by serial dilution of the sample aliquots with SOC medium (10-2 to 10-
4), plating on prewarmed LB plates containing 100 mg/l ampicillin. The colonies were counted
after an overnight incubation at 37 °C.
An aliquot of the library in pYES-DEST52 GatewayTM vector containing approximately 1 × 106
transformants was inoculated into 50 ml liquid LB medium with 100 mg/l ampicillin and grown

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 6
on a rotary shaker at 200 rpm and 37 °C. When the culture reached an OD600 of 1.0, total
plasmid DNA was isolated with the GeneJETTM Plasmid Miniprep Kit (Fermentas). The
plasmid DNA was transformed into S. cerevisiae W303a by the Library Screen
Transformation Protocol, as described by Gietz and Woods (2006), with minor modifications.
Briefly, 5 × 107 yeast cells from an overnight culture were added to 50 ml prewarmed 2 ×
YPD and incubated on a rotary shaker at 180 rpm and 28 °C until the cell density reached 2
× 107 cells/ml (determined by counting on a haemocytometer). The cells were harvested by
centrifugation (3,000 × g for 5 min), washed twice with 25 ml sterile water, and pelleted
again. The cells were mixed with the transformation mixture, prepared from 2.4 ml PEG 3500
(50%), 360 μl Li-acetate (1.0 M), 500 μl carrier single stranded salmon DNA (2 mg/ml,
denatured in boiling water for 5 min), and plasmid DNA (10 μg, diluted with sterile water to
340 μl). The mixture was vortexed and incubated at 42 °C for 1 h with inversion mixing every
5 min. Following this heat shock, the cells were pelleted by centrifugation (3,000 × g, 5 min).
The pellet was resuspended in 35 ml liquid YNB+CSM-Ura medium with 2% glucose and
0.5% ammonium sulphate (w/v). The titre of the library was determined by 10 × dilution of the
mixture prior to incubation with sterile water and plating 100 μl and 10 μl aliquots on
prewarmed plates of the same medium with 2% agar (w/v). The colonies were counted after
five days of incubation at 30 °C. The rest of the mixture was incubated overnight on a rotary
shaker at 180 rpm and 28 °C. After the incubation, the culture was mixed with 10.5 ml sterile
glycerol, aliquoted, and frozen at -80 °C.
2.5 Screening for osmotolerant transformants
To screen for transformants with increased stress tolerance, 4 ml of the frozen S. cerevisiae
library was thawed and centrifuged at 3,000× g for 5 min. The pelleted cells were washed in
4 ml fresh YNB+CSM-Ura medium with 0.5% ammonium sulphate, 2% galactose, and 1%
raffinose (w/v), re-pelleted, and resuspended again in 1 ml of the same medium. Then 100 μl
aliquots were streaked onto selection media with the MICs of the various osmolytes. The
MICs were determined by spotting S. cerevisiae W303a suspensions on YNB+CSM media
with 0.5% ammonium sulphate, 2% galactose and 1% raffinose (w/v) and with a range of
concentrations of a given osmolyte. The screening plates were incubated at 30 °C and
checked at regular intervals over a period of five weeks. All of the colonies that appeared
were transferred to fresh plates of the same medium, frozen in 50% glycerol at -80 °C, and
stored until further analysis.
2.6 Identification of the inserted genes in the osmotolerant transformants
Total DNA was extracted from the transformants with increased osmotolerance by
resuspending a medium-sized colony in 50 μl PrepMan Ultra Sample Preparation Reagent
(Applied Biosystems), and incubating the suspension in boiling water for 10 min and
centrifuging at 20,000 × g for 2 min. The supernatant was used for amplification of the insert
in the pYES-DEST52 vector by the polymerase chain reaction (PCR) in a 35-μl reaction
volume, with 10 pmol of vector -specific primers (5’-AATGCAAAAACTGCATAACCAC-3' and
5’-GGGATAGGCTTACCTTCGAAG-3’), 2.5 nmol of each dNTP (Applied Biosystems), 15
nmol MgCl2, and 0.625 U Taq polymerase (Fermentas). The thermal profile of the reaction
was as follows: 5 min denaturation at 94 °C, followed by 33 cycles of 30 s at 94 °C, 30 s at
the annealing temperature, and 30 s at 72 °C, with the final elongation step of 5 min at 72 °C.
The annealing temperature was 60 °C, and was decreased by 1 °C/cycle for the first 8
cycles, and then kept at 55 °C for the remaining 25 cycles. The products were checked by
agarose electrophoresis and sent for sequencing by Macrogen Inc. (Korea). The nucleotide
Identification of genes conferring stress tolerance Page numbers not for citation purposes. 7
sequences were analysed and their putative functions identified by comparison with the
GenBank database entries, using BLASTX. The complete sequences of the selected genes
were obtained by primer walking.
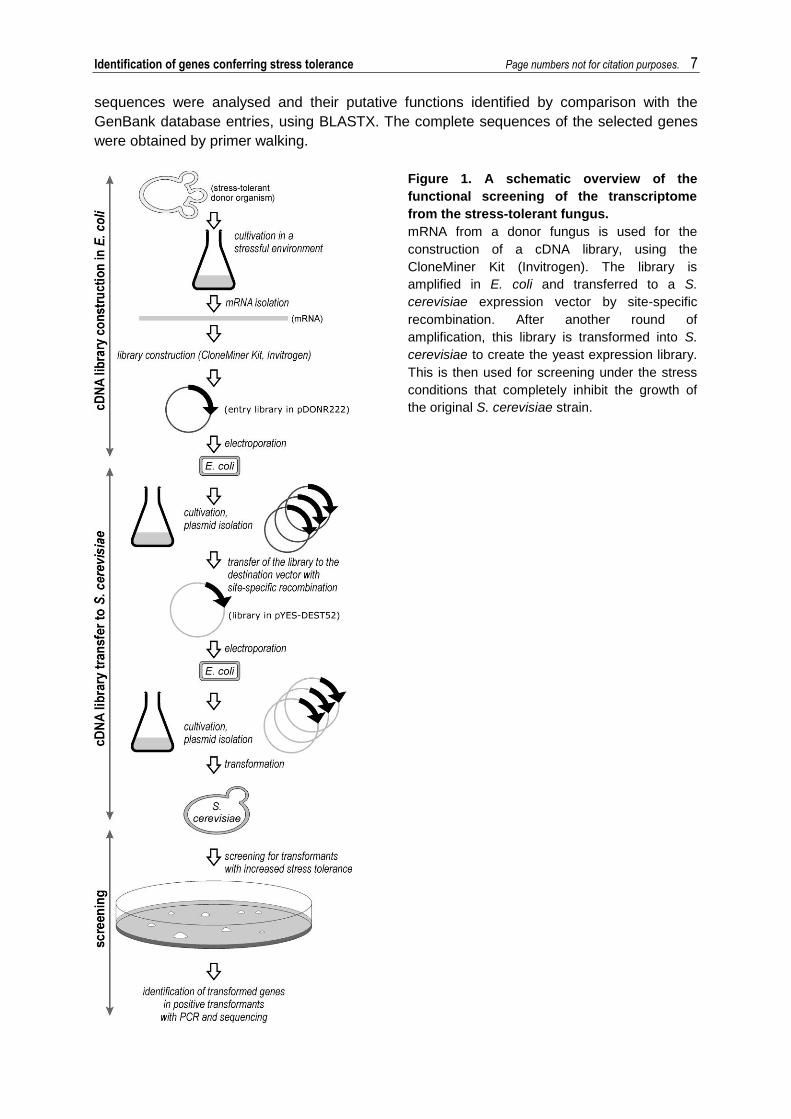
Figure 1. A schematic overview of the
functional screening of the transcriptome
from the stress-tolerant fungus.
mRNA from a donor fungus is used for the
construction of a cDNA library, using the
CloneMiner Kit (Invitrogen). The library is
amplified in E. coli and transferred to a S.
cerevisiae expression vector by site-specific
recombination. After another round of
amplification, this library is transformed into S.
cerevisiae to create the yeast expression library.
This is then used for screening under the stress
conditions that completely inhibit the growth of
the original S. cerevisiae strain.

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 8
2.7 Curing of plasmids
The selected transformants were cured of their plasmids to demonstrate the origin of the
observed stress-tolerant phenotypes. The transformants were grown overnight in liquid YPD
medium, the culture was streak-plated on solid YPD plates, and incubated at 30 °C.
Individual colonies were transferred to solid selection medium with 5-fluoro-orotic acid and
incubated at 30 °C. The loss of the plasmid in the formed colonies was confirmed through
their inability to grow on YNB+CSM-Ura medium with 0.5% ammonium sulphate, 2%
galactose, and 1% raffinose (w/v), and with PCR using vector-specific primers.
2.8 Stress-tolerance assays
S. cerevisiae cells (strain W303a) transformants with increased stress tolerance (carrying the
vector pYES-DEST52 with a gene from R. mucilaginosa) were compared to the same
tranformants cured of their plasmids. S. cerevisiae cells (strain W303a) with an empty
plasmid pYES-DEST52 was used as control. They were grown overnight in YNB+CSM-Ura
medium with 0.5% ammonium sulphate, 2% galactose, and 1% raffinose (w/v) or YNB+CSM
medium with 0.5% ammonium sulphate and 2% glucose (w/v) to mid-exponential phase,
adjusted to an OD600 of 0.5, 10-fold serially diluted (1–104 dilutions) with fresh medium, and
spotted in 3 μl aliquots onto YNB+CSM-Ura medium with 0.5% ammonium sulphate, 2%
galactose, 1% raffinose, and 2% agar (w/v), both without added osmolytes (control) and with
the various osmolytes (0.86 M or 1.37 M NaCl, 2.7 M sorbitol, 3.4 M glycerol or 0.4 M LiCl).
The plates were incubated at 30 °C (an additional plate without the added osmolytes was
incubated at 15 °C), and photographed on days 4 and 7.
2.9 Enzyme activity assays
Cell-free extracts of soluble proteins were prepared from S. cerevisiae (expressing RmPGM2
or RmSEC53, or containing the empty pYES-DEST52 plasmid) and R. mucilaginosa grown
to mid-exponential phase in liquid YNB+CSM-Ura medium with 0.5% ammonium sulphate,
2% galactose, and 1% raffinose (w/v). The cells were harvested by centrifugation (3,000 × g
for 5 min), washed twice with 50 mM Tris-HCl (pH 7.5 at 4 °C), and resuspended in an equal
volume of lysis buffer (50 mM Tris-HCl, pH 7.5, 2 mM EDTA, 0.3 M sorbitol, 1 mM ditiotreitol,
protease inhibitor cocktail 50 μl/g cells; Sigma), and kept on ice. An equal volume of acid-
washed glass beads (425-600 m; Sigma) was added to the cells in the lysis buffer. The
cells were disrupted using a Mixer Mill MM 400 (Retsch). The homogenate was centrifuged
(20,000 × g, 15 min, 4 C) and the supernatant that contained the protein was frozen at -80
°C until use.
Phosphoglucomutase activity was measured by incubating the soluble protein extracts (0.1
μg for S. cerevisiae expressing RmPGM2, 3 μg for the other samples) in 100 μl of the
following reaction mixture: 50 mM Tris-HCl (pH 7.5), 0.5 mM NAD+, 2 U/ml glucose-6-
phosphate dehydrogenase, and 1.5 mM MgCl2. The reactions were mixed in a microtitre
plate, started by the addition of 4 mM glucose-1-phosphate, and followed at 30 °C in a
Multiskan Spectrum plate reader, by measuring the production of NAD+ at 340 nm.
Phosphomannomutase activity was measured by incubation of the soluble protein extracts
(0.5 μg for S. cerevisiae expressing RmSEC53
following reaction mixture: 50 mM Tris-HCl (pH 7.5), 0.5 mM NAD+, 2 U/ml glucose-6-
phosphate dehydrogenase, 3.5 μg/ml phosphomannose isomerase, 0.2 U/ml
phosphoglucose isomerase, 0.025 mM glucose-1,6-diphosphate, and 5 mM MgCl. The

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 9
reactions were started by the addition of 0.1 mM mannose-1-phosphate. The other
conditions were the same as for the phosphoglucomutase assays.
The enzyme assays were conducted in three biological replicates, for each of which the
enzyme activities were calculated as means of two measurements. The statistical
significances of the differences between the activities of the different species/ transformants
or different LiCl concentrations were tested using the T-test in the open-source PSPP
software.
3 Results
3.1 Library construction in E. coli, and transfer to S. cerevisiae
The total size of the library in E. coli was approximately 5 × 107 CFU. Sequencing of 20
randomly picked clones and subsequent BLASTX analysis showed that all of the sequenced
vectors contained inserts, of which 12 encoded complete coding sequences (CDS) with a
homologue in the GenBank database (e-value cut-off, 1.00 e-10), four encoded a CDS with a
homologue in the GenBank database but were truncated at the 5’-end, and four were not
identified (data not shown). The library in S. cerevisiae contained approximately 2.5 × 105
CFU.
3.2 Screening for osmotolerant transformants
The screening of the cDNA library yielded several S. cerevisiae transformants that grew on
low water activity media that prevented the growth of S. cerevisiae W303a. Fifty-one colonies
appeared after one to five weeks on a medium with 1.37 M NaCl, 47 colonies on a medium
with 1.71 M NaCl, two with 2.7 M sorbitol, 35 with 3.4 M glycerol, and 20 with 0.4 M LiCl.
3.3 R. mucilaginosa genes in osmotolerant transformants
R. mucilaginosa cDNA sequences were obtained from 92 transformants that showed
increased osmotolerance. A total of 71 unique sequences were found (data not shown).
Putative functions were identified for 44 genes by comparison with GenBank database
entries. Eight sequences were similar only to hypothetical proteins, while for 19 sequences,
no similar proteins were found. Twelve transformants were selected for further analyses
(Table 1).
Redundancy-check analysis revealed that the five most-abundant expressed sequence tags
accounted for 21.7% of the sequences (Table 2). However, the 5' and 3' cDNA ends of the
redundant clones differed in length by at least a few nucleotides, which indicated their
independent integration into the library, rather than a clonal origin.
3.4 Expression of selected genes from R. mucilaginosa in S. cerevisiae increases salt
tolerance of the transformants
Spotting assays of selected transformants on different media (Fig. 2A) did not show any
differences in growth in the absence of osmolytes and at 0.86 M NaCl. In contrast, all of the
transformants grew at 1.37 M NaCl, and three of them (carrying the RmSEC53, RmFACB
and RmPGM2 genes) also grew at 0.4 M LiCl; the control strain carrying an empty plasmid
did not grow under these conditions. Growth on galactose was not a prerequisite for incrased
salt tolerance of RmSEC52 and RmPGM2 transfromants. Although less pronounced, when

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 10
they were constitutively expressed by the TPI promotor in the plasmid pYX142, these genes
also increased halotolerance of S. cerevisiae during growth on glucose (data not shown).
Growth at 15 °C showed no substantial differences between the transformants and none
grew on 2.7 M sorbitol or 3.4 M glycerol (with the exception of RmPgm2, which grew on
glycerol, data not shown).
Growth of transformants cured of their plasmids (Fig. 2B) was comparable to growth of the
original S. cerevisiae strain without the plasmid. The exceptions were the cured
transformants for RmFACB and RmPGM2, which grew better on 0.4 M LiCl and at 15 °C,
respectively, possibly due to unknown mutations in the genome.
Figure 2. Increased salt tolerance of selected S. cerevisiae transformants by genes from R.
mucilaginosa.
A: Ten-fold serially diluted cultures of transformants carrying the indicated inserts and a transformant
carrying an empty vector (pYES-DEST52) as a control. The dilutions were plated on YNB media
supplemented with all amino acids except uracil and with 2% galactose and 1% raffinose, without
additional osmolytes (control) or with added osmolytes (as indicated). The plates were incubated at 30
°C (except one plate that was incubated at 15 °C) and photographed after 4 or 7 days (as indicated).
B: Ten-fold serially diluted cultures of the same clones as above, cured of their plasmids by treatment
with 5-fluoro-orotic acid. S. cerevisiae W303a without a plasmid was used as the control. The media
were supplemented with all amino acids, and the other growth conditions were the same as above.

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 11
3.5 Phosphoglucomutase, but not phosphomannomutase, from R. mucilaginosa is sensitive
to LiCl
The phosphoglucomutase and phosphomannomutase activities were more than 10-fold
higher in the S. cerevisiae transformants containing the RmPGM2 and RmSEC53 genes,
respectively, compared to S. cerevisiae with an empty plasmid (Fig. 3). Phosphoglucomutase
activity was also significantly (T-test, p <0.05) higher in R. mucilaginosa, compared to S.
cerevisiae.
The phosphoglucomutase activity in whole cell lysates was extremely sensitive to the
presence of LiCl and was significantly (T-test, p <0.05) lowered by 1 mM LiCl, and even
further inhibited by 5 mM LiCl, in all of the samples. In contrast, phosphomannomutase
activity was not significantly lowered by any of the LiCl concentrations tested in any of the
samples, even at 50 mM LiCl. Surprisingly, the phosphoglucomutase activity of the
transformant with the overexpressed phosphomannomutase enzyme (RmSEC53) was less
sensitive to LiCl compared to that of the other samples (Fig. 3A).
Figure 3. Effect of lithium on phosphoglucomutase and phosphomannomutase activities.
Enzyme activities were measured in cell-free extracts of soluble proteins from S. cerevisiae W303a
expressing the genes encoding phosphoglucomutase (RmPGM2) (A) or phosphomannomutase
(RmSEC53) (B), or containing the corresponding empty pYES-DEST52 plasmid (Sc) and R.
mucilaginosa EXF-1630 (Rm). The reactions were followed spectrophotometrically by measuring the
production of NAD+ at 340 nm. The activities are expressed as millimoles NAD
+ produced per min per
μg total protein. The reactions for both of the enzymes were performed in the absence of LiCl and
additionally at 0.1 mM, 1 mM and 5 mM LiCl for phosphoglucomutase (A), and at 5 mM, 20 mM and
50 mM LiCl for phosphomannomutase (B).
4. Discussion
To facilitate the identification of fungal genes involved in stress tolerance that are of potential
biotechnological interest, a high-throughput screening system was established in this study
(Fig. 1). This approach has several advantages: (a) It uses naturally stress-tolerant
organisms as gene donors; (b) Fungi are more promising gene donors for improving stress
tolerance of plants and industrially important fungi than structurally different and
phylogenetically distant prokaryotes; (c) Similarly, S. cerevisiae is a more appropriate

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 12
screening system than prokaryotes, and it is easier to work with in the laboratory, compared
to screening in plant systems; (d) As eukaryotic mRNAs are monocistronic, the use of cDNA
libraries eliminates the need for determining which of the cloned genes is responsible for
changes in the transformant. The absence of introns further reduces the required
bioinformatic work; (e) The use of commercially available chemicals and kits and the
Gateway cloning technology (Invitrogen) simplifies the procedures and increases the
flexibility of the method, by offering the ability to transfer the complete cDNA library to a
vector of choice.
In addition, several modifications of the method can be imagined. The use of a vector with a
dominant marker would allow for screening in prototrophic strains of S. cerevisiae (e.g. in
industrial strains). Screening on other carbon sources would be possible with the use of a
constitutive promoter. Changes in the cultivation conditions for the mRNA for library
construction, and changes in the screening conditions, can be used for the identification of
genes involved in various other stress responses (e.g. low or high pH, oxygen stress).
Finally, the use of environmental metatranscriptomes would further expand the pool of
genetic resources to unknown and non-culturable organisms.
We have shown the efficiency of this method for the identification of halotolerance-conferring
genes by screening the transcriptome of the stress-tolerant yeast R. mucilaginosa. All of the
12 selected S. cerevisiae transformants showed increased tolerance to NaCl, three
transformants also showed increased tolerance to LiCl, while none except RmPgm2 grew on
media containing sorbitol or glycerol at concentrations inhibitory to S. cerevisiae W303a. This
indicates that the proteins identified specifically counteract toxicity of the anorganic cations
Na+ or Li+, and not osmotic stress. These genes could therefore be used to provide increased
salt tolerance, rather than for increasing drought tolerance. Genes identified by screening on
non-ionic osmolytes (e.g. sorbitol) might prove more suitable for drought tolerance.
Although adaptations to high salinity and low temperatures sometimes overlap, substantial
differences in growth of the selected transformants at 15 °C were not observed. Screening at
low temperatures would be better suited for the identification of genes that are involved in
growth and survival at low temperatures.
The genes that were shown to increase halotolerance of S. cerevisiae fall into different
functional categories. The majority of these can be associated to responses to salt stress or
other stress factors.
The two most redundant genes identified in the screening encode two different
phosphomutases. Their redundancy might result from a combination of growth on galactose
and LiCl. One of these, RmPGM2, encodes a putative phosphoglucomutase, an enzyme that
catalyses the interconversion of glucose-6-phosphate and glucose-1-phosphate (the latter
being the product of galactose catabolism). Interestingly, when expressed with a constitutive
promoter, both RmPgm2 and RmSec53 increased halotolerance even when galactose in the
medium was substituted with glucose (data not shown). This enzyme is involved in
glycolysis, the pentose-phosphate shunt, and the metabolism of glycogen, trehalose, and
galactose (Boles et al., 1994). The expression of PGM2 in S. cerevisiae increases in
response to various abiotic stresses, including salt stress and lithium stress (Masuda et al.,
2001). Pgm2 is inhibited with high affinity by lithium, which acts as a competitive inhibitor of

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 13
yeast phosphoglucomutase activity by competing with magnesium, a cofactor of Pgm2
(Masuda et al., 2001). The phosphoglucomutase activity of S. cerevisiae cells expressing
RmPgm2 did not exhibit greater tolerance to lithium, and it was significantly inhibited even at
1 mM LiCl. However, the loss of activity was probably compensated for by the higher initial
activity due to the overexpression of the RmPgm2 enzyme, which results in increased lithium
tolerance of the transformants. The higher phosphoglucomutase activity seen for R.
mucilaginosa in comparison to S. cerevisiae even in the absence of salt, might contribute to
the greater lithium tolerance of R. mucilaginosa.
RmSEC53 is a homologue of genes that encode phosphomannomutases. These are
enzymes that catalyse an early step in the pathway of yeast O-linked and N-linked
mannosylation, the interconversion of mannose-6-phosphate and mannose-1-phosphate on
the cytosolic surface of the endoplasmic reticulum (Kepes and Schekman, 1988). This
process is important for protein glycosylation, protein sorting and secretion, and maintenance
of a functional endomembrane system in eukaryotic cells (Herscovics and Orlean, 1993). In
our analysis, phosphomannomutases proved to be relatively resistant to lithium even at 50
mM LiCl. As phosphomannomutases are known to also catalyse the interconversion of
glucose-1 phosphate and glucose-6 phosphate (Boles et al., 1994), the RmSec53
transformant was tested for this activity as well. Interestingly, while the expression of
RmSec53 the S. cerevisiae did not cause a detectable increase in phosphoglucomutase
activity, the activity appeared to be less sensitive to LiCl than the activity of the
phosphoglucomutase enzymes. Overexpression of RmSec53 could therefore increase the
lithium tolerance of S. cerevisiae in at least two ways: by altering its protein mannosylation;
or by rescuing its lithium-inhibited phosphoglucomutase activity.
In addition, four other genes identified by the functional screening are associated with
carbohydrate metabolism and energy production: RmGPA2, RmACK1, RmMCP1 and
RmPET9.
RmGPA2 is similar to the S. cerevisiae gene for the nucleotide binding alpha subunit of the
heterotrimeric G protein, which is required for the detection of extracellular glucose through
the activation of cAMP synthesis (Colombo et al., 1998). A GPA2 null-mutant of S. cerevisiae
has been shown to have decreased resistance to hyperosmotic stress (Yoshikawa et al.,
2009).
No homologue was found for RmACK1 in S. cerevisiae. However, the deduced protein
shares strong similarity with acetate kinases from other fungi. These enzymes are required
for the conversion of acetate to acetyl-CoA, and they are thus important in energy
metabolism. In Listeria monocytogenes, a decrease in the levels of this enzyme after salt
shock was related to a decrease in the intracellular acetyl-CoA pool (Duche et al., 2002).
RmMCP1 and RmPET9 encode putative mitochondrial transporters: the first has no known
function, while the second is an ADP/ATP carrier of the mitochondrial inner membrane
(Lawson and Douglas, 1988).
A further gene identified by the functional screening, RmARO4, codes for a putative 3-deoxy-
D-arabino-heptulosonate-7-phosphate (DAHP) synthase, an enzyme that catalyses the first
step in the biosynthesis of aromatic amino acids (Braus, 1991). A homologue of this gene

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 14
from the halotolerant yeast D. hansenii shows increased expression under high salinity
conditions (Calderon-Torres et al., 2006). It has also been proposed that amino-acid
availability is physiologically interconnected with high salinity (Pascual-Ahuir et al., 2001).
RmRIB4 is a gene for a putative lumazine synthase (6,7-dimethyl-8-ribityllumazine
synthase), an enzyme that catalyses the penultimate step in the riboflavin biosynthesis
pathway (Mortl et al., 1996).
Finally, four of the selected genes are involved in global regulation of the cell machinery.
RmFACB does not show significant similarity to any of the genes in S. cerevisiae, but its
protein sequence is similar to the sequence of C6 transcription factor from Aspergillus
fumigatus, and its homologues. The cloned gene is possibly 5’-truncated; however, it has a
start codon at the 5’-end.
RmANB1 is similar to a gene for translation elongation factor eIF-5A. It has been shown in
animal cells that it has a modest role in protein synthesis under normal conditions; however,
it becomes much more important under stress conditions, and it is possibly involved in the
reprogramming of protein synthesis in cells under stress (Li et al., 2010).
RmATG18 encodes a putative phosphoinositide binding protein that is involved in autophagy
vesicle formation. It is responsible for vacuolar fission in response to osmotic stress, and it
interacts with the transcriptional activator Rtg3. In turn, Rtg3 regulates the expression of
specific genes upon osmostress, in a Hog1-dependent manner (Georgakopoulos et al.,
2001; Noriega Esteban, 2009). In Arabidopsis thaliana it was suggested that the AtATG18
genes are required under multiple environmental stress conditions (Xiong et al., 2005).
RmCPR encodes a protein that is similar to cytoplasmic peptidyl-prolyl cis-trans isomerase,
which catalyses peptidyl-prolyl cis-trans isomerisation, the rate-limiting step in protein folding
(Wang and Heitman, 2005). Expression of the stress-inducible homologue CcCYP from the
pigeon-pea plants in A. thaliana increased tolerance to drought, salinity and extreme
temperatures (Sekhar et al., 2010). Similarly, the homologue from Thellungiella halophila
confers salt tolerance in S. pombe and Nicotiana tabacum (Chen et al., 2007).
Since S. cerevisiae transformants were constructed from a haploid auxotrophic laboratory
strain W303a, they could not be used for industrial application. For this purpose the genes
would have to be cloned and expressed in an industrial yeast strain, preferably by a stable
integration into the genome using a dominant selection marker. Since the impact of the
genes identified in this study on plant halotolerance has not been tested yet, several of the
described genes should be expressed in model plant organisms to select the most efficient
targets for the improvement of crops.
5 Conclusions
A high-throughput method for identification of stress-tolerance-conferring genes in fungal
transcriptomes was described, and the efficiency of this method by screening the
basidiomycetous yeast R. mucilaginosa was demonstrated. Twelve selected genes
conveyed increased NaCl and/or LiCl tolerance to S. cerevisiae. The present study

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 15
demonstrates that methods for high-throughput mining of underexploited genetic resources
of stress-tolerant fungi could facilitate the improvement of organisms used in biofuel and food
production.
Acknowledgments
The authors wish to thank Dr. Tjaša Danevčič for help with measurements of enzymatic
activities and Prof. Børge Diderichsen for critical reading of the manuscript. The scientific
work was partly financed via operation “Centre of excellence for integrated approaches in
chemistry and biology of proteins” number OP13.1.1.2.02.0005, financed by European
regional development fund (85% share of financing) and by Slovenian Ministry of higher
education, science and technology (15% share of financing). The authors also acknowledge
the financial support from the state budget by the Slovenian Research Agency
(Infrastructural Centre Mycosmo, and grant no. J4-2022).
References
Bhatnagar-Mathur, P., Vadez, V., Sharma, K.K., 2008.
Transgenic approaches for abiotic stress tolerance in
plants: retrospect and prospects. Plant Cell Rep. 27, 411-
424.
Boles, E., Liebetrau, W., Hofmann, M., Zimmermann, F.K.,
1994. A family of hexosephosphate mutases in
Saccharomyces cerevisiae. Eur. J. Biochem. 220, 83-96.
Braus, G.H., 1991. Aromatic Amino-Acid Biosynthesis in the
Yeast Saccharomyces cerevisiae - a Model System for
the Regulation of a Eukaryotic Biosynthetic-Pathway.
Microbiol. Rev. 55, 349-370.
Calderon-Torres, M., Pena, A., Thome, P.E., 2006.
DhARO4, an amino acid biosynthetic gene, is stimulated
by high salinity in Debaryomyces hansenii. Yeast 23, 725-
734.
Chen, A.P., Wang, G.L., Qu, Z.L., Lu, C.X., Liu, N., Wang,
F., Xia, G.X., 2007. Ectopic expression of ThCYP1, a
stress-responsive cyclophilin gene from Thellungiella
halophila, confers salt tolerance in fission yeast and
tobacco cells. Plant Cell Rep. 26, 237-245.
Colombo, S., Ma, P.S., Cauwenberg, L., Winderickx, J.,
Crauwels, M., Teunissen, A., Nauwelaers, D., de Winde,
J.H., Gorwa, M.F., Colavizza, D., Thevelein, J.M., 1998.
Involvement of distinct G-proteins, Gpa2 and Ras, in
glucose- and intracellular acidification-induced cAMP
signalling in the yeast Saccharomyces cerevisiae. EMBO
J. 17, 3326-3341.
Du, J., Huang, Y.P., Xi, J., Cao, M.J., Ni, W.S., Chen, X.,
Zhu, J.K., Oliver, D.J., Xiang, C.B., 2008. Functional
gene-mining for salt-tolerance genes with the power of
Arabidopsis. Plant J. 56, 653-664.
Duche, O., Tremoulet, F., Namane, A., Labadie, J.,
Consortiu, E.L.G., 2002. A proteomic analysis of the salt
stress response of Listeria monocytogenes. FEMS
Microbiol. Lett. 215, 183-188.
Georgakopoulos, T., Koutroubas, G., Vakonakis, I.,
Tzermia, M., Prokova, V., Voutsina, A., Alexandraki, D.,
2001. Functional analysis of the Saccharomyces
cerevisiae YFR021w/YGR223c/YPL100w ORF family
suggests relations to mitochondrial/peroxisomal functions
and amino acid signalling pathways. Yeast 18, 1155-
1171.
Gietz, R.D., Woods, R.A., 2006. Yeast transformation by
the LiAc/SS Carrier DNA/PEG method. Second Edition
ed. in: Yeast Protocol, Vol. 313, Humana Press, pp. 107-
20.
Gostinčar, C., Grube, M., Gunde-Cimerman, N., 2011.
Evolution of Fungal Pathogens in Domestic
Environments? Fungal. Biol. 115, 1008-1018.
Gunde-Cimerman, N., Sonjak, S., Zalar, P., Frisvad, J.C.,
Diderichsen, B., Plemenitaš, A., 2003. Extremophilic fungi
in Arctic ice: a relationship between adaptation to low
temperature and water activity Phys. Chem. Earth 28,
1273-1278.
Herscovics, A., Orlean, P., 1993. Glycoprotein-biosynthesis
in yeast. FASEB J. 7, 540-550.
Holmberg, N., Bulow, L., 1998. Improving stress tolerance
in plants by gene transfer. Trends Plant Sci. 3, 61-66.
Joshi, A., Dang, H.Q., Vaid, N., Tuteja, N., 2009. Isolation
of high salinity stress tolerant genes from Pisum sativum
by random overexpression in Escherichia coli and their
functional validation. Plant Signal. Behav. 4, 400-12.
Kapardar, R.K., Ranjan, R., Grover, A., Puri, M., Sharma,
R., 2010. Identification and characterization of genes
conferring salt tolerance to Escherichia coli from pond
water metagenome. Bioresour. Technol. 101, 3917-24.

Identification of genes conferring stress tolerance Page numbers not for citation purposes. 16
Kepes, F., Schekman, R., 1988. The yeast Sec53 gene
encodes phosphomannomutase. J. Biol. Chem. 263,
9155-9161.
Lawson, J.E., Douglas, M.G., 1988. Separate genes
encode functionally equivalent ADP/ATP carrier proteins
in Saccharomyces cerevisiae - Isolation and analysis of
Aac2. J. Biol. Chem. 263, 14812-14818.
Li, C.H., Ohn, T., Ivanov, P., Tisdale, S., Anderson, P.,
2010. eIF5A Promotes Translation Elongation, Polysome
Disassembly and Stress Granule Assembly. Plos One 5,
e9942.
Masuda, C.A., Xavier, M.A., Mattos, K.A., Galina, A.,
Montero-Lomeli, M., 2001. Phosphoglucomutase is an in
vivo lithium target in yeast. J. Biol. Chem. 276, 37794-
37801.
Mortl, S., Fischer, M., Richter, G., Tack, J., Weinkauf, S.,
Bacher, A., 1996. Biosynthesis of riboflavin - lumazine
synthase of Escherichia coli. J. Biol. Chem. 271, 33201-
33207.
Munns, R., 2002. Comparative physiology of salt and water
stress. Plant Cell Environ. 25, 239-250.
Noriega Esteban, N. 2009. The Rtg1 AND Rtg3 proteins are
novel transcription factors regulated by the yeast Hog1
MAPK upon stress. in: Unitat de Senyalització Cellular,
Departament de Ciencies Experimentals i de la Salut
(CEXS), Universitat Pompeu Fabra, pp. 153.
Pascual-Ahuir, M., Serrano, R., Proft, M., 2001. The Sko1p
repressor and Gcn4p activator antagonistically modulate
stress-regulated transcription in Saccharomyces
cerevisiae. Molecular and Cellular Biology 21, 16-25.
Prista, C., Soeiro, A., Vesely, P., Almagro, A., Ramos, J.,
Loureiro-Dias, M.C., 2002. Genes from Debaryomyces
hansenii increase salt tolerance in Saccharomyces
cerevisiae W303. FEMS Yeast Res. 2, 151-7.
Rai, M.K., Kalia, R.K., Singh, R., Gangola, M.P., Dhawan,
A.K., 2010. Developing stress tolerant plants through in
vitro selection - an overview of the recent progress.
Environ. Exp. Bot. 71, 89-98.
Rozema, J., Flowers, T., 2008. Crops for a salinized world.
Science 322, 1478-1480.
Sekhar, K., Priyanka, B., Reddy, V.D., Rao, K.V., 2010.
Isolation and characterization of a pigeonpea cyclophilin
(CcCYP) gene, and its over-expression in Arabidopsis
confers multiple abiotic stress tolerance. Plant Cell
Environ. 33, 1324-1338.
Shima, J., Takagi, H., 2009. Stress-tolerance of baker's-
yeast (Saccharomyces cerevisiae) cells: stress-protective
molecules and genes involved in stress tolerance.
Biotechnol. Appl. Biochem. 53, 155-64.
Somvanshi, V.S., 2009. Patenting drought tolerance in
organisms. Recent Pat. DNA Gene Seq. 3(1), 16-25.
Wang, P., Heitman, J., 2005. The cyclophilins. Genome
Biol. 6, 226.
Xiong, Y., Contento, A.L., Bassham, D.C., 2005. AtATG18a
is required for the formation of autophagosomes during
nutrient stress and senescence in Arabidopsis thaliana.
Plant J. 42, 535-546.
Yoshikawa, K., Tanaka, T., Furusawa, C., Nagahisa, K.,
Hirasawa, T., Shimizu, H., 2009. Comprehensive
phenotypic analysis for identification of genes affecting
growth under ethanol stress in Saccharomyces
cerevisiae. FEMS Yeast Res. 9, 32-44.
Zheng, D.Q., Wu, X.C., Tao, X.L., Wang, P.M., Li, P., Chi,
X.Q., Li, Y.D., Yan, Q.F., Zhao, Y.H., 2010. Screening
and construction of Saccharomyces cerevisiae strains
with improved multi-tolerance and bioethanol fermentation
performance. Bioresour. Technol. 102, 3020-3027.
Related Documents











