RESEARCH ARTICLE Genetic relationships in Cucurbita pepo (pumpkin, squash, gourd) as viewed with high frequency oligonucleotide– targeting active gene (HFO–TAG) markers Harry S. Paris • Adi Doron-Faigenboim • Umesh K. Reddy • Ryan Donahoo • Amnon Levi Received: 6 October 2014 / Accepted: 12 January 2015 Ó Springer Science+Business Media Dordrecht 2015 Abstract Cucurbita pepo is a highly diverse, eco- nomically important member of the Cucurbitaceae. C. pepo encompasses hundreds of cultivars of pump- kins, squash, and gourds. Although C. pepo has been scrutinized with various types of DNA markers, the relationships among the cultivar-groups of C. pepo subsp. pepo, the more widely grown subspecies, have not heretofore been adequately resolved. We assessed genetic relationships among 68 accessions of Cucur- bita pepo, including 48 from C. pepo subsp. pepo, using polymorphisms in 539 high frequency oligonu- cleotide–targeting active gene (HFO–TAG) frag- ments, that preferably represent coding regions of the genome. Dissimilarities among accessions were calculated, a dendrogram was constructed, and prin- cipal component analyses were conducted. Dissimilarities demarcated the four edible-fruited cultivar-groups of C. pepo subsp. pepo, Cocozelle, Pumpkin, Vegetable Marrow, and Zucchini. Further- more, the results indicate that the Old World pumpkins as well as the long-fruited cultivar-groups of C. pepo subsp. pepo (cocozelle, vegetable marrow, and zuc- chini) evolved from spontaneous crossing and gene exchange between pumpkins derived from northern North America and pumpkins derived from southern North America. Consistent with pictorial and narrative historical records, such crossing appears to have occurred in Renaissance Europe within the first decades of the European contact with North America. The Old World pumpkins are more closely related to the long-fruited cultivar-groups than are the native North American pumpkins. Keywords Crop evolution Cucurbita pepo subsp. pepo Cucurbitaceae Nuclear DNA markers Plant breeding Pumpkin Squash Zucchini Introduction Cucurbita pepo L. (2n = 2x = 40) is a highly diverse, economically important crop species encompassing summer squash as well as many pumpkins, winter squash, and ornamental gourds (Paris et al. 2012). This species is native to North America and is thought to have been domesticated first in southern Mexico H. S. Paris (&) Newe Ya‘ar Research Center, Agricultural Research Organization, P. O. Box 1021, 30-095 Ramat Yishay, Israel e-mail: [email protected] A. Doron-Faigenboim Volcani Center, Agricultural Research Organization, P. O. Box 6, 50-250 Bet Dagan, Israel U. K. Reddy Department of Biology, West Virginia State University, Institute, WV 25112-1000, USA R. Donahoo A. Levi U.S. Vegetable Laboratory, USDA, ARS, 2700 Savannah Highway, Charleston, SC 29414, USA 123 Genet Resour Crop Evol DOI 10.1007/s10722-015-0218-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
Genetic relationships in Cucurbita pepo (pumpkin, squash,gourd) as viewed with high frequency oligonucleotide–targeting active gene (HFO–TAG) markers
Harry S. Paris • Adi Doron-Faigenboim •
Umesh K. Reddy • Ryan Donahoo • Amnon Levi
Received: 6 October 2014 / Accepted: 12 January 2015
� Springer Science+Business Media Dordrecht 2015
Abstract Cucurbita pepo is a highly diverse, eco-
nomically important member of the Cucurbitaceae.
C. pepo encompasses hundreds of cultivars of pump-
kins, squash, and gourds. Although C. pepo has been
scrutinized with various types of DNA markers, the
relationships among the cultivar-groups of C. pepo
subsp. pepo, the more widely grown subspecies, have
not heretofore been adequately resolved. We assessed
genetic relationships among 68 accessions of Cucur-
bita pepo, including 48 from C. pepo subsp. pepo,
using polymorphisms in 539 high frequency oligonu-
cleotide–targeting active gene (HFO–TAG) frag-
ments, that preferably represent coding regions of
the genome. Dissimilarities among accessions were
calculated, a dendrogram was constructed, and prin-
cipal component analyses were conducted.
Dissimilarities demarcated the four edible-fruited
cultivar-groups of C. pepo subsp. pepo, Cocozelle,
Pumpkin, Vegetable Marrow, and Zucchini. Further-
more, the results indicate that the Old World pumpkins
as well as the long-fruited cultivar-groups of C. pepo
subsp. pepo (cocozelle, vegetable marrow, and zuc-
chini) evolved from spontaneous crossing and gene
exchange between pumpkins derived from northern
North America and pumpkins derived from southern
North America. Consistent with pictorial and narrative
historical records, such crossing appears to have
occurred in Renaissance Europe within the first
decades of the European contact with North America.
The Old World pumpkins are more closely related to
the long-fruited cultivar-groups than are the native
North American pumpkins.
Keywords Crop evolution � Cucurbita pepo subsp.
pepo � Cucurbitaceae � Nuclear DNA markers � Plant
breeding � Pumpkin � Squash � Zucchini
Introduction
Cucurbita pepo L. (2n = 2x = 40) is a highly diverse,
economically important crop species encompassing
summer squash as well as many pumpkins, winter
squash, and ornamental gourds (Paris et al. 2012). This
species is native to North America and is thought to
have been domesticated first in southern Mexico
H. S. Paris (&)
Newe Ya‘ar Research Center, Agricultural Research
Organization, P. O. Box 1021, 30-095 Ramat Yishay,
Israel
e-mail: [email protected]
A. Doron-Faigenboim
Volcani Center, Agricultural Research Organization,
P. O. Box 6, 50-250 Bet Dagan, Israel
U. K. Reddy
Department of Biology, West Virginia State University,
Institute, WV 25112-1000, USA
R. Donahoo � A. Levi
U.S. Vegetable Laboratory, USDA, ARS, 2700 Savannah
Highway, Charleston, SC 29414, USA
123
Genet Resour Crop Evol
DOI 10.1007/s10722-015-0218-6

approximately 10,000 years ago (Whitaker and Cutler
1971, 1986; Lira and Montes 1994; Smith 1997).
Considerable evidence places a subsequent, indepen-
dent domestication in the eastern or central United
States about 5,000 years ago (Whitaker and Carter
1946; Smith 2006). These domestication events,
geographically quite distant, must have occurred in
genetically distinct populations. Allozyme variation
and DNA sequence polymorphisms have revealed a
strong dichotomy in cultivated C. pepo (Decker-
Walters 1990). Descendants of the older, more south-
erly domestication have been given subspecific status
as C. pepo subsp. pepo. However, wild populations of
subsp. pepo have not been described (Nee 1990).
Descendants of the more recent and northerly domes-
tication have been given subspecific status as C. pepo
subsp. texana (Scheele) Filov. Wild populations of
subsp. texana have been described from Texas and
much of the southeastern and central United States
(Erwin 1931; Bailey 1943; Decker and Wilson 1987;
Cowan 1997). Most cultigens belong to the former
subspecies but many others belong to the latter (Paris
et al. 2006, 2012). Wild populations of a third lineage
occur in a geographically intermediate region, the
states of Tamaulipas and Nuevo Leon in northeastern
Mexico (Bailey 1943; Andres 1987; Nee 1990). They
have a separate taxonomic designation, C. pepo subsp.
fraterna (Bailey) Lira, Andres et Nee, but as far as is
known have no domesticated descendants.
Cucurbita pepo, under thousands of years of
cultivation, has diversified greatly into an astonishing
range of fruit sizes, shapes, topographies, and colors
(Duchesne 1786; Paris 2007). Much of the stark
diversity in fruit shape has been driven by differing
uses of the fruits as food, for consumption of whole
immature fruits or of mature fruit flesh or seeds (Paris
et al. 2012). Differences in fruit shape, a polygenic
characteristic (Emerson 1910; Sinnott 1935) form the
basis of a horticultural classification recognizing eight
groups of edible-fruited cultivars (Paris 1986).
Accordingly, these are Acorn (turbinate, furrowed),
Scallop (flat, lobed), Crookneck (long, narrow
necked), Straightneck (long, with proximal constric-
tion), Pumpkin (round or nearly round), Vegetable
Marrow (short, tapered cylindrical), Cocozelle (long,
bulbous cylindrical) and Zucchini (uniformly cylin-
drical). The first four cultivar-groups are in C. pepo
subsp. texana and the last four are in C. pepo subsp.
pepo (Paris 2000). Both subspecies also encompass
various cultivars of small-fruited, ornamental gourds,
those of the former subspecies can be referred to as
‘‘ovifera gourds’’ and those of the latter as ‘‘pepo
gourds’’.
Various nuclear DNA-sequence polymorphisms
have been employed to examine genetic relationships
among Cucurbita pepo accessions with ever-increas-
ing precision (Lebeda et al. 2007). The markers used
have included random amplified polymorphic DNAs
(RAPDs), amplified fragment length polymorphisms
(AFLPs), inter-simple sequence repeats (ISSRs), sim-
ple sequence repeats (SSRs), and sequence-related
amplified polymorphisms (SRAPs). The results of
these investigations have helped clarify a number of
genetic relationships within the species, most notably
differentiating the subspecies of C. pepo and their
constituent cultivar-groups. Using the various mark-
ers, accessions within each of the four edible-fruited
cultivar-groups (fruit shapes) of C. pepo subsp. texana
were clearly seen to be associated with one another. Of
the four edible-fruited cultivar-groups of C. pepo
subsp. pepo, only the Zucchini Group (cultivars with
uniformly cylindrical-shaped fruits) was as clearly
defined, the other three being less clearly separated
from one another (Paris et al. 2003; Gong et al. 2012).
Recently, a new type of polymerase chain reaction
(PCR) marker named ‘‘high frequency oligonucleo-
tide–targeting active genes (HFO–TAG)’’ was devel-
oped (Levi et al. 2010). HFO–TAG markers are
dominant, produced by primers representing oligonu-
cleotides that exist in high frequency in expressed
sequence-tag (EST) unigenes, and are highly repro-
ducible and polymorphic. Therefore, HFO–TAGs
could be expected to depict genetic relationships more
in accordance with phenotypic variability than DNA
markers that represent non-coding regions. Indeed,
HFO–TAGs have resulted in a much improved
assessment of genetic diversity within Citrullus
(watermelon and colocynth) (Levi et al. 2013).
The present work focused mainly on Cucurbita
pepo subsp. pepo because relationships among culti-
var-groups of this subspecies have not been as well
resolved as in C. pepo subsp. texana. The objective
here is to obtain an improved definition of the elements
within this subspecies and to increase understanding of
the evolution and interrelationships of its cultivar-
groups by employing HFO–TAG markers.
Genet Resour Crop Evol
123

Materials and methods
Plant material
Sixty-eight accessions of Cucurbita pepo were chosen
for study, most of which are of C. pepo subsp. pepo
(Table 1). Some of the subsp. pepo accessions have
been examined with other DNA markers (Paris et al.
2003; Gong et al. 2012) and are included here for
comparative purposes. Others, some of which exhibit
intriguing combinations of traits, have not previously
been subjected to such scrutiny. Four accessions of
Citrullus Schrad. ex Eckl. et Zeyh. and two of
Lagenaria Ser. were included as checks and to obtain
a picture of phylogenetic relationships among these
three cultivated genera of the Cucurbitaceae.
Each of the 68 accessions of Cucurbita pepo was
assigned a tripartite abbreviation (Table 1). The first part
is formed by one letter designating subspecies (P = pepo,
T = texana, F = fraterna, and accessions of question-
able or uncertain subspecific affiliation were designated as
Q = questionable and U = uncertain) (Paris et al. 2003;
Gong et al. 2012). The second part is formed by a two-
letter abbreviation designatingcultivar-grouporaffiliation
(for example, PU = Pumpkin, CO = Cocozelle,
SN = Straightneck, GO = Ovifera Gourd, GT = Wild
texana Gourd, ZU = Zucchini). The third part is formed
by a three-letter abbreviation of the name of the accession
(for example, CTF = ‘Connecticut Field’, SFF = ‘Saf-
fron’, TRF = ‘True French’). In such fashion, the subsp.
pepo Pumpkin ‘Connecticut Field’ was abbreviated
P-PU-CTF, the subsp. texana Straightneck squash ‘Saf-
fron’ was abbreviated T-SN-SFF, and the subsp. pepo
Zucchini ‘True French’ was abbreviated P-ZU-TRF.
One mature fruit from many of the accessions is
depicted in Fig. 1 and others are depicted elsewhere
(Paris 2001; Paris and Nerson 2003; Goldman 2004;
Paris et al. 2012). Seeds of each accession of Cucurbita
pepo were derived from the collection maintained at the
Agricultural Research Organization, Newe Ya‘ar
Research Center (Paris 2001). The original seed samples
were obtained frompublic and private sources and nearly
all of them are derived from North America or Europe
(Table 1). The identity of each accession was checked
and verified by sowing seeds and growing out at least
eight plants in the field. Plant growth habit was scored
from field observation as bush, open bush, or vine.
Most of the seeds used for producing plants for
DNA extraction were not from the original seed
samples but instead derived from seeds obtained by
self- or sib-pollinations at Newe Ya‘ar (Table 1). A
few of the original seed samples produced plants that
had noticeable variation among one another in vege-
tative and reproductive characteristics; in such cases
the plants selected for self- or sib-pollination were
those that were most representative of the accession,
based on its name, catalogue description, or the
majority of the plants observed.
The four Citrullus and two Lagenaria accessions
were also assigned tripartite abbreviations, beginning
with C for the former and L for the latter. The four
accessions of Citrullus used for this study were the
dessert watermelons, C. lanatus (Thunb.) Matsum. et
Nakai ‘Black Diamond’ (C-LA-BDI) and ‘Charleston
Gray’ (C-LA-CHG), the citron watermelon, C. lanatus
subsp. lanatus syn. C. amarus Schrad. PI 296341 (C-
LA-341), and the colocynth, C. colocynthis (L.)
Schrad. PI 386015 (C-CC-015). The two accessions
of Lagenaria were L. siceraria (Mol.) Standl. PI
271477 (L-SI-477) and PI 442369 (L-SI-369). The
seeds of these accessions of Citrullus and Lagenaria
are maintained at the U.S. Vegetable Laboratory,
A.R.S./U.S.D.A., Charleston, SC USA.
DNA extraction
Young leaves were collected from three or four two-
week-old plants of each accession and stored at
-80 �C for later DNA isolation. The DNA was
isolated from the frozen leaves using the method
described by Levi and Thomas (1999).
PCR amplification and analysis using HFO–TAG
primers
The identification and development of HFO–TAG
primers using expressed-sequence-tag (EST) data of
watermelon fruit have been described by Levi et al.
(2010). Twenty-two of these HFO–TAG primers
(Table 2) were used in this study.
DNA amplification conditions and analysis
The PCR reaction cocktail (25 lL) contained 20 lM
NaCl, 50 mM Tris–HCl pH 9, 0.1 % Triton-X-100,
0.01 % gelatin, 1.6 mM MgCl2, 200 lM each of
dNTPs (Sigma-Aldrich, St. Louis, MO), 100 lM
primer, five units GoTaq� DNA polymerase
Genet Resour Crop Evol
123

Table 1 Classification and sources of seeds of 68 Cucurbita pepo accessions
Accession name Accession
abbreviation
Accession pedigreea Origin (seed source) Plant growth
habit
Royal Acorn T-AC-RAC RACc-S USA (Twilley) Vine
Table Queen T-AC-TQE TQEd USA (Global) Vine
Early Golden Crookneck T-CN-EAC EAC USA (Henry Field) Bush, open
Early Summer Crookneck T-CN-ESC ESCc-S USA (Ledden) Bush, open
Golden Bush Scallop T-SC-GBS GBS-1-4-2 USA (Dessert) Bush
White Bush Scallop T-SC-WBS WBS-3-Sa USA (Mandeville) Bush
Saffron T-SN-SFF SFF-S,Sa USA (Burpee) Bush, open
Straightneck Early Yellow T-SN-SNE SNE USA (American) Bush, open
Bicolor Pear T-GO-BPR BPRa-S USA (Harris) Vine
Crown of Thorns T-GO-CRT CRTa USA (Henry Field) Vine
Shenot’s Crown of Thorns T-GO-SHC SHCc USA (Jung) Vine
Spoon T-GO-SPN SPNd USA (Harris) Vine
PI 285213 T-GT-213 PI 285213-Sa,Sb USA (North Central P.I.S.) Vine
Wild Texas T-GT-WTX WTX-10-Sf,Si USA (Texas A&M Univ.) Vine
Wild Mexico 2 F-GF-WM2 WMX2-Sa Tamaulipas, Mexico
(Texas A&M U.)
Vine
Flat U-GU-FLA FLAa Canada (Stokes)b Vine
Flat Striped U-GU-FLS FLS Canada (Stokes) Vine
Miniature Ball U-GU-MNB MNB-5 Canada (Stokes) Vine
Green Warted Q-GQ-
GWA
GWA USA (Agway) Vine
Small Flat Warted Q-GQ-SFW SFW-S,Sa USA (Park)c Vine
Little Gem P-GP-LGM LGM-S,Sc South Africa (MayFord) Vine
Orange P-GP-ORA ORA-22 USA (Harris) Vine
Orange Ball P-GP-ORB ORB-3 Canada (Stokes) Vine
Orange Warted P-GP-OWA OWA-1,4 Canada (Stokes) Vine
Rolet P-GP-ROL ROL-Sa,Sb South Africa (Unknown) Vine
Genovese P-CO-GNV GNVc-S,Sa France (Vilmorin) Bush, open
Lungo Bianco di Sicilia P-CO-LBS LBS-S Italy (S.A.I.S.) Bush
Long Cocozelle P-CO-LCO LCO-1,3,7 USA (Ledden) Bush, open
Romanesco P-CO-ROM ROM-3 Italy (S.A.I.S.) Bush
Striato d’Italia P-CO-STI STIa-1,25 Italy (S.A.I.S.) Bush, open
Valery P-CO-VAA VAA Italy (La Semiorto Sementi) Vine
Verte non-coureuse d’Italie P-CO-VNI VNIc France (Delbard) Bush, open
PI 165018 P-CO-018 PI165018-Sa Turkey (North Central P.I.S.) Bush
PI 379307 P-CO-307 PI379307-5-3,7 Kosovo (North Central P.I.S.) Bush
PI 261610 P-CO-610 PI261610-S,Sa Spain (North Central P.I.S.) Vine
PI 175710 P-CO-710 PI175710-Sa,Sb Turkey (North Central P.I.S.) Bush
Citrouille de Touraine P-PU-CIR CIR-Sa France (Ducrettet) Vine
Connecticut Field P-PU-CTF CTFa USA (Ledden) Vine
Jack O’Lantern P-PU-JOL JOL-1-21 USA (Excel) Vine
Mogango sul Mineiro P-PU-MOG MOG-S,Sb Brazil (Feltrin)d Vine
Nonkadi P-PU-NOK NOK-23,S Uzbekistan (I.V.S.Q.) Vine
Porqueira P-PU-PRQ PRQ-S,Sa,Sb Portugal (Unknown) Vine
Genet Resour Crop Evol
123

Table 1 continued
Accession name Accession
abbreviation
Accession pedigreea Origin (seed source) Plant growth
habit
Small Sugar P-PU-SSU SSUd-1 USA (Ledden) Vine
Tender and True P-PU-TAT TAT-S UK (Sutton’s) Bush
Uzbekistan Local Pumpkin P-PU-ULP ULP-S Uzbekistan (N.I.P.B.) Vine
Winter Luxury P-PU-WLU WLUa USA (Jung) Vine
Yugoslavia 7 P-PU-YU7 YU7 Former Yugoslavia (Unknown) Vine
PI 212000 P-PU-000 PI212000-S,Sa Iran (North Central P.I.S.) Vine
PI 311102 P-PU-102 PI311102 Guatemala (North Central P.I.S.) Vine
PI 442298 P-PU-298 PI442298-S Mexico (North Central P.I.S.) Vine
PI 442309 P-PU-309 PI442309-S Mexico (North Central P.I.S.) Vine
PI 442313 P-PU-313 PI442313-S Mexico (North Central P.I.S.) Vine
PI 169473 P-PU-473 PI169473 Turkey (North Central P.I.S.) Vine
PI 171628 P-PU-628 PI171628-SR,Sa,Sb Turkey (North Central P.I.S.) Vine
PI 458750 Guicoy Grande Verde P-PU-750 PI458750 Guatemala (North Central P.I.S.) Vine
All Green Bush P-VM-AGB AGB-7 UK (Asmer) Bush
Bolognese P-VM-BOG BOG-1,8 Italy (C.C.S.A.) Bush, open
Sihi Lavan P-VM-SLA SLA-26-17-1-1-29 Israel (Aratan) Bush
Table Dainty P-VM-TDA TDA-5 UK (Sutton’s) Vine
Verte Petite d’Alger P-VM-VPA VPAa-1,3 France (Abondance) Bush, open
Vegetable Spaghetti P-VM-VSP VSPe-S Japan (Sakata) Vine
Yakor P-VM-YAK YAK-2 Russia (U.N.E.E.S.S.O.K.) Bush
PI 525179 Alexandria P-VM-179 PI525179-S,Sa,Sb Egypt (North Central P.I.S.) Bush
M2546 P-VM-546 M2546-Sd,e Israel (P.I.S.) Vine
PI 181764 P-VM-764 PI181764-3 Lebanon (North Central P.I.S.) Bush
Fordhook Zucchini P-ZU-FZU FZUc-2-S USA (Burpee) Bush, open
True French P-ZU-TRF TRF-4-2-1-24-18-2-20-20-3-
16
UK (Thompson and Morgan) Bush, open
Zucchini Select P-ZU-ZSL ZSL-2-6-S Canada (Stokes) Bush, open
Tripartite designation is for subspecies, cultivar-group, and abbreviated accession name, respectively. For subspecies: T = texana,
F = fraterna, P = pepo. Other designations used for some of the accessions with regard to their subspecies affiliations are
U = uncertain and Q = questionable. For cultivar-group: AC = Acorn, CN = Crookneck, SC = Scallop, SN = Straightneck,
GO = Ovifera Gourd, GT = Texana (wild) Gourd, GF = Fraterna (wild) Gourd, GU = Uncertain Gourd, GQ = Questionable
Gourd, GP = Pepo Gourd, CO = Cocozelle, PU = Pumpkin, VM = Vegetable Marrow, ZU = Zucchinia Accession pedigrees begin with three-letter abbreviations or introduction numbers. Those not followed by hyphens indicate original
seed stocks. The number of hyphens following the three-letter abbreviations or introduction numbers indicates the number of
generations of self- or sib-pollination. A number following a hyphen indicates self-pollination whilst a capital letter ‘‘S’’ following a
hyphen, with or without a lower-case letter, indicates a sib-pollination. For example, for ‘Sihi Lavan’ (P-VM-SLA), SLA-26-17-1-1-
29 indicates that the seed stock used was derived from 5 generations of self-pollination. For ‘Fordhook Zucchini’ (P-ZU-FZU), FZU-
2-S indicates that the seed stock used was derived by one generation of self-pollination followed by one generation of sib-pollination.
For ‘Wild Texas’ (T-GT-WTX), WTX-10-Sf,Si indicates that the seed stock used was derived by one generation of self-pollination
followed by one generation of sib-pollination, but the second (sib-pollinated) generation actually consists of seeds derived from two
separate sib-pollinations that were bulkedb ‘Flat’ (U-GU-FLA) is a true-breeding derivative taken from ‘Small Warted Blend’c ‘Small Flat Warted’ (Q-GQ-SFW) is a true-breeding derivative from ‘Ovifera’d ‘Mogango sul Mineiro’ (P-PU-MOG), although sold by a Brazilian seed company, has the strong ribbing characteristic of
Guatemalan and Mexican pumpkins
Genet Resour Crop Evol
123

(Promega, Madison, WI), and 7 ng template DNA.
Amplifications were carried out for 40 cycles in a
PTC-200 thermocycler (MJ Research, Watertown,
MA) for 60 s to denature the DNA at 92 �C, 70 s for
primer annealing at 35, 40, 45, 48, 50, 55, or 60 �C (as
determined for each primer in Table 2, based on
primer melting temperature-Tm), and 120 s for primer
extension at 72 �C. The fragments were analyzed
using a CEQ 8800 DNA Genetic Analysis System
(Beckman Coulter, Fullerton, CA) which has high
accuracy, distinguishing between DNA fragments
ranging in size from 75 to 450 base pairs and differing
in size by one base pair (Levi et al. 2010). For
visualization of DNA fragments on the CEQ 8800, the
forward primers were labeled with one of three
WellRED dye labels (D2, D3, or D4; Proligo, Boulder,
CO) as described for SRAP markers (Levi et al. 2006).
Marker data collection and analysis
The HFO–TAG fragments (Table 2) were scored as
‘‘1’’ (present) or ‘‘0’’ (absent) using the built-in
fragment analysis software provided with the Beck-
man Coulter (Fullerton, CA) CEQ-8800 Genetic
Analysis System. The resulting binary matrix was
uploaded to the PAST program version 2.17 (Hammer
et al. 2001). Dissimilarities among accessions were
calculated using Euclidean distances, which are pre-
ferred for comparisons involving dominant markers in
populations within a single diploid species (Kosman
and Leonard 2005). From the similarity matrix, a
dendrogram was constructed using the unweighted
pair-group average (UPGMA) clustering method. The
robustness of the phylogenetic tree was evaluated by
bootstrap analysis with 5,000 replicates using the
bootstrap function of PAST. Principal component
analyses (PCA) were carried out using the multivariate
data analysis of PAST. Distributions of the accessions
were depicted in two-dimensional scatter plots using
the first and second principal coordinates.
Results
Dissimilarity values
Dissimilarity values among the 74 accessions (68
Cucurbita pepo, 4 Citrullus, and 2 Lagenaria) ranged
from 0.00 for two accessions of Cucurbita pepo
pumpkins, PI 442309 (P-PU-309) and PI 442313 (P-
PU-313), to over 16.00 between many of the C. pepo
and two of the Citrullus lanatus accessions (Table 3).
The average dissimilarities among accessions
within taxa (P, T, F, Q, U, C, L) were always smaller
than those between taxa, except for one. This one
exception was within Cucurbita pepo, in which the
two Q accessions were slightly more distant from each
Fig. 1 Mature fruits of 33 accessions of Cucurbita pepo subsp.
pepo (P) and five other accessions of questionable (Q) or
uncertain (U) subspecific status. Left to right, with their
dimensions (cm, polar diameter 9 equatorial diameter): top
row, four pumpkins, P-PU-NOK (18 9 24), P-PU-CTF
(20 9 21), P-PU-PRQ (29 9 28), P-PU-000 (17 9 27); second
row, five pumpkins, P-PU-TAT (14 9 17), P-PU-WLU
(13 9 16), P-PU-MOG (16 9 13), P-PU-628 (15 9 14),
P-PU-473 (22 9 18); third row, eight vegetable marrows,
P-VM-VSP (22 9 13), P-VM-BOG (20 9 11), P-VM-SLA
(20 9 8), P-VM-546 (32 9 11), P-VM-AGB (28 9 12),
P-VM-TDA (23 9 11), P-VM-179 (32 9 14), P-VM-764
(25 9 9); fourth row, eight cocozelles and three zucchinis,
P-CO-610 (52 9 12), P-CO-VAA (42 9 10), P-CO-ROM
(40 9 11); P-CO-018 (47 9 10), P-CO-710 (39 9 11), LCO
(42 9 7), P-CO-STI (49 9 9), P-CO-LBS (41 9 8), P-ZU-TRF
(33 9 8), P-ZU-FZU (41 9 10), P-ZU-ZSL (40 9 10); bottom
row, five pepo gourds, two questionable gourds, and three
uncertain gourds, P-GP-ROL (6 9 6), P-GP-LGM (6 9 6),
P-GP-ORA (6 9 6), P-GP-ORB (7 9 6), P-GP-OWA (6 9 9),
Q-GQ-GSW (5 9 7); Q-GQ-SFW (5 9 7), U-GU-FLS
(5 9 7), U-GU-FLA (4 9 5), U-GU-MNB (4 9 4)
Genet Resour Crop Evol
123
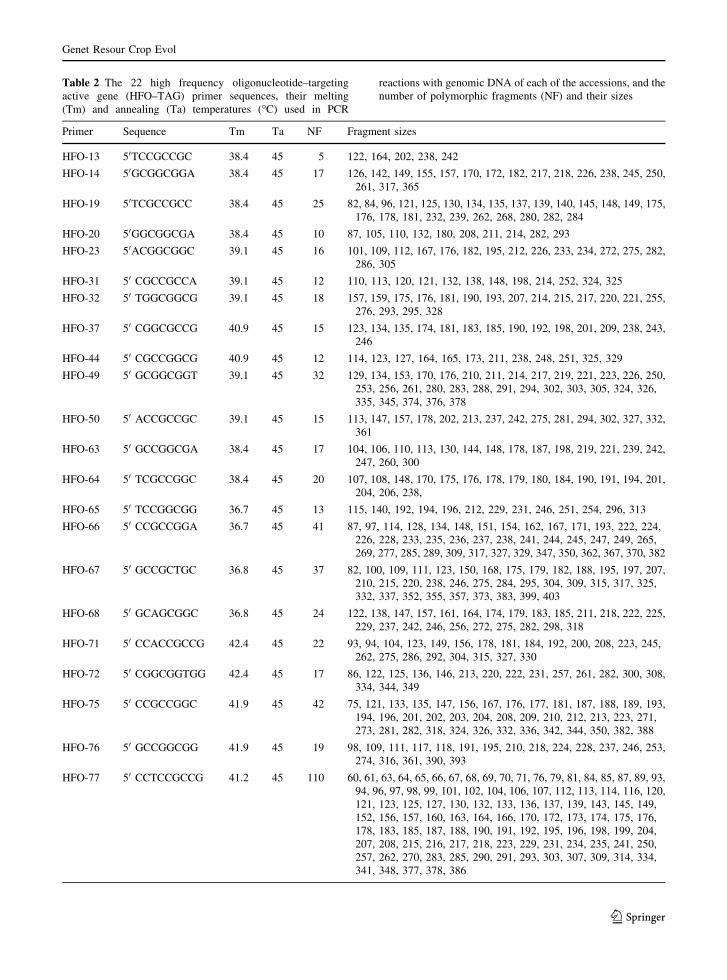
Table 2 The 22 high frequency oligonucleotide–targeting
active gene (HFO–TAG) primer sequences, their melting
(Tm) and annealing (Ta) temperatures (�C) used in PCR
reactions with genomic DNA of each of the accessions, and the
number of polymorphic fragments (NF) and their sizes
Primer Sequence Tm Ta NF Fragment sizes
HFO-13 50TCCGCCGC 38.4 45 5 122, 164, 202, 238, 242
HFO-14 50GCGGCGGA 38.4 45 17 126, 142, 149, 155, 157, 170, 172, 182, 217, 218, 226, 238, 245, 250,
261, 317, 365
HFO-19 50TCGCCGCC 38.4 45 25 82, 84, 96, 121, 125, 130, 134, 135, 137, 139, 140, 145, 148, 149, 175,
176, 178, 181, 232, 239, 262, 268, 280, 282, 284
HFO-20 50GGCGGCGA 38.4 45 10 87, 105, 110, 132, 180, 208, 211, 214, 282, 293
HFO-23 50ACGGCGGC 39.1 45 16 101, 109, 112, 167, 176, 182, 195, 212, 226, 233, 234, 272, 275, 282,
286, 305
HFO-31 50 CGCCGCCA 39.1 45 12 110, 113, 120, 121, 132, 138, 148, 198, 214, 252, 324, 325
HFO-32 50 TGGCGGCG 39.1 45 18 157, 159, 175, 176, 181, 190, 193, 207, 214, 215, 217, 220, 221, 255,
276, 293, 295, 328
HFO-37 50 CGGCGCCG 40.9 45 15 123, 134, 135, 174, 181, 183, 185, 190, 192, 198, 201, 209, 238, 243,
246
HFO-44 50 CGCCGGCG 40.9 45 12 114, 123, 127, 164, 165, 173, 211, 238, 248, 251, 325, 329
HFO-49 50 GCGGCGGT 39.1 45 32 129, 134, 153, 170, 176, 210, 211, 214, 217, 219, 221, 223, 226, 250,
253, 256, 261, 280, 283, 288, 291, 294, 302, 303, 305, 324, 326,
335, 345, 374, 376, 378
HFO-50 50 ACCGCCGC 39.1 45 15 113, 147, 157, 178, 202, 213, 237, 242, 275, 281, 294, 302, 327, 332,
361
HFO-63 50 GCCGGCGA 38.4 45 17 104, 106, 110, 113, 130, 144, 148, 178, 187, 198, 219, 221, 239, 242,
247, 260, 300
HFO-64 50 TCGCCGGC 38.4 45 20 107, 108, 148, 170, 175, 176, 178, 179, 180, 184, 190, 191, 194, 201,
204, 206, 238,
HFO-65 50 TCCGGCGG 36.7 45 13 115, 140, 192, 194, 196, 212, 229, 231, 246, 251, 254, 296, 313
HFO-66 50 CCGCCGGA 36.7 45 41 87, 97, 114, 128, 134, 148, 151, 154, 162, 167, 171, 193, 222, 224,
226, 228, 233, 235, 236, 237, 238, 241, 244, 245, 247, 249, 265,
269, 277, 285, 289, 309, 317, 327, 329, 347, 350, 362, 367, 370, 382
HFO-67 50 GCCGCTGC 36.8 45 37 82, 100, 109, 111, 123, 150, 168, 175, 179, 182, 188, 195, 197, 207,
210, 215, 220, 238, 246, 275, 284, 295, 304, 309, 315, 317, 325,
332, 337, 352, 355, 357, 373, 383, 399, 403
HFO-68 50 GCAGCGGC 36.8 45 24 122, 138, 147, 157, 161, 164, 174, 179, 183, 185, 211, 218, 222, 225,
229, 237, 242, 246, 256, 272, 275, 282, 298, 318
HFO-71 50 CCACCGCCG 42.4 45 22 93, 94, 104, 123, 149, 156, 178, 181, 184, 192, 200, 208, 223, 245,
262, 275, 286, 292, 304, 315, 327, 330
HFO-72 50 CGGCGGTGG 42.4 45 17 86, 122, 125, 136, 146, 213, 220, 222, 231, 257, 261, 282, 300, 308,
334, 344, 349
HFO-75 50 CCGCCGGC 41.9 45 42 75, 121, 133, 135, 147, 156, 167, 176, 177, 181, 187, 188, 189, 193,
194, 196, 201, 202, 203, 204, 208, 209, 210, 212, 213, 223, 271,
273, 281, 282, 318, 324, 326, 332, 336, 342, 344, 350, 382, 388
HFO-76 50 GCCGGCGG 41.9 45 19 98, 109, 111, 117, 118, 191, 195, 210, 218, 224, 228, 237, 246, 253,
274, 316, 361, 390, 393
HFO-77 50 CCTCCGCCG 41.2 45 110 60, 61, 63, 64, 65, 66, 67, 68, 69, 70, 71, 76, 79, 81, 84, 85, 87, 89, 93,
94, 96, 97, 98, 99, 101, 102, 104, 106, 107, 112, 113, 114, 116, 120,
121, 123, 125, 127, 130, 132, 133, 136, 137, 139, 143, 145, 149,
152, 156, 157, 160, 163, 164, 166, 170, 172, 173, 174, 175, 176,
178, 183, 185, 187, 188, 190, 191, 192, 195, 196, 198, 199, 204,
207, 208, 215, 216, 217, 218, 223, 229, 231, 234, 235, 241, 250,
257, 262, 270, 283, 285, 290, 291, 293, 303, 307, 309, 314, 334,
341, 348, 377, 378, 386
Genet Resour Crop Evol
123
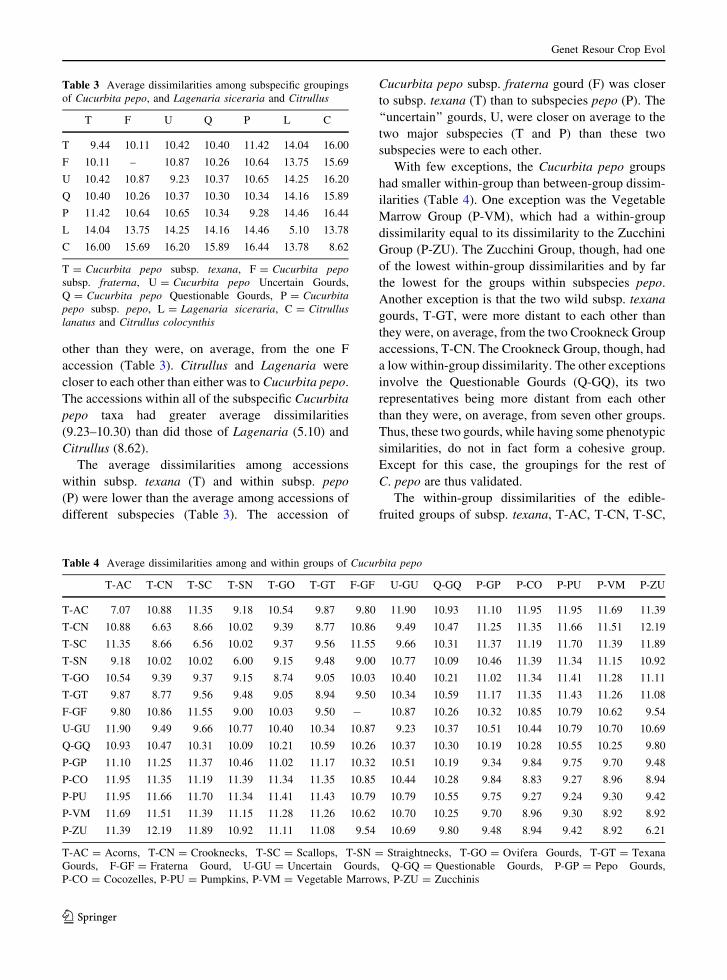
other than they were, on average, from the one F
accession (Table 3). Citrullus and Lagenaria were
closer to each other than either was to Cucurbita pepo.
The accessions within all of the subspecific Cucurbita
pepo taxa had greater average dissimilarities
(9.23–10.30) than did those of Lagenaria (5.10) and
Citrullus (8.62).
The average dissimilarities among accessions
within subsp. texana (T) and within subsp. pepo
(P) were lower than the average among accessions of
different subspecies (Table 3). The accession of
Cucurbita pepo subsp. fraterna gourd (F) was closer
to subsp. texana (T) than to subspecies pepo (P). The
‘‘uncertain’’ gourds, U, were closer on average to the
two major subspecies (T and P) than these two
subspecies were to each other.
With few exceptions, the Cucurbita pepo groups
had smaller within-group than between-group dissim-
ilarities (Table 4). One exception was the Vegetable
Marrow Group (P-VM), which had a within-group
dissimilarity equal to its dissimilarity to the Zucchini
Group (P-ZU). The Zucchini Group, though, had one
of the lowest within-group dissimilarities and by far
the lowest for the groups within subspecies pepo.
Another exception is that the two wild subsp. texana
gourds, T-GT, were more distant to each other than
they were, on average, from the two Crookneck Group
accessions, T-CN. The Crookneck Group, though, had
a low within-group dissimilarity. The other exceptions
involve the Questionable Gourds (Q-GQ), its two
representatives being more distant from each other
than they were, on average, from seven other groups.
Thus, these two gourds, while having some phenotypic
similarities, do not in fact form a cohesive group.
Except for this case, the groupings for the rest of
C. pepo are thus validated.
The within-group dissimilarities of the edible-
fruited groups of subsp. texana, T-AC, T-CN, T-SC,
Table 3 Average dissimilarities among subspecific groupings
of Cucurbita pepo, and Lagenaria siceraria and Citrullus
T F U Q P L C
T 9.44 10.11 10.42 10.40 11.42 14.04 16.00
F 10.11 – 10.87 10.26 10.64 13.75 15.69
U 10.42 10.87 9.23 10.37 10.65 14.25 16.20
Q 10.40 10.26 10.37 10.30 10.34 14.16 15.89
P 11.42 10.64 10.65 10.34 9.28 14.46 16.44
L 14.04 13.75 14.25 14.16 14.46 5.10 13.78
C 16.00 15.69 16.20 15.89 16.44 13.78 8.62
T = Cucurbita pepo subsp. texana, F = Cucurbita pepo
subsp. fraterna, U = Cucurbita pepo Uncertain Gourds,
Q = Cucurbita pepo Questionable Gourds, P = Cucurbita
pepo subsp. pepo, L = Lagenaria siceraria, C = Citrullus
lanatus and Citrullus colocynthis
Table 4 Average dissimilarities among and within groups of Cucurbita pepo
T-AC T-CN T-SC T-SN T-GO T-GT F-GF U-GU Q-GQ P-GP P-CO P-PU P-VM P-ZU
T-AC 7.07 10.88 11.35 9.18 10.54 9.87 9.80 11.90 10.93 11.10 11.95 11.95 11.69 11.39
T-CN 10.88 6.63 8.66 10.02 9.39 8.77 10.86 9.49 10.47 11.25 11.35 11.66 11.51 12.19
T-SC 11.35 8.66 6.56 10.02 9.37 9.56 11.55 9.66 10.31 11.37 11.19 11.70 11.39 11.89
T-SN 9.18 10.02 10.02 6.00 9.15 9.48 9.00 10.77 10.09 10.46 11.39 11.34 11.15 10.92
T-GO 10.54 9.39 9.37 9.15 8.74 9.05 10.03 10.40 10.21 11.02 11.34 11.41 11.28 11.11
T-GT 9.87 8.77 9.56 9.48 9.05 8.94 9.50 10.34 10.59 11.17 11.35 11.43 11.26 11.08
F-GF 9.80 10.86 11.55 9.00 10.03 9.50 - 10.87 10.26 10.32 10.85 10.79 10.62 9.54
U-GU 11.90 9.49 9.66 10.77 10.40 10.34 10.87 9.23 10.37 10.51 10.44 10.79 10.70 10.69
Q-GQ 10.93 10.47 10.31 10.09 10.21 10.59 10.26 10.37 10.30 10.19 10.28 10.55 10.25 9.80
P-GP 11.10 11.25 11.37 10.46 11.02 11.17 10.32 10.51 10.19 9.34 9.84 9.75 9.70 9.48
P-CO 11.95 11.35 11.19 11.39 11.34 11.35 10.85 10.44 10.28 9.84 8.83 9.27 8.96 8.94
P-PU 11.95 11.66 11.70 11.34 11.41 11.43 10.79 10.79 10.55 9.75 9.27 9.24 9.30 9.42
P-VM 11.69 11.51 11.39 11.15 11.28 11.26 10.62 10.70 10.25 9.70 8.96 9.30 8.92 8.92
P-ZU 11.39 12.19 11.89 10.92 11.11 11.08 9.54 10.69 9.80 9.48 8.94 9.42 8.92 6.21
T-AC = Acorns, T-CN = Crooknecks, T-SC = Scallops, T-SN = Straightnecks, T-GO = Ovifera Gourds, T-GT = Texana
Gourds, F-GF = Fraterna Gourd, U-GU = Uncertain Gourds, Q-GQ = Questionable Gourds, P-GP = Pepo Gourds,
P-CO = Cocozelles, P-PU = Pumpkins, P-VM = Vegetable Marrows, P-ZU = Zucchinis
Genet Resour Crop Evol
123
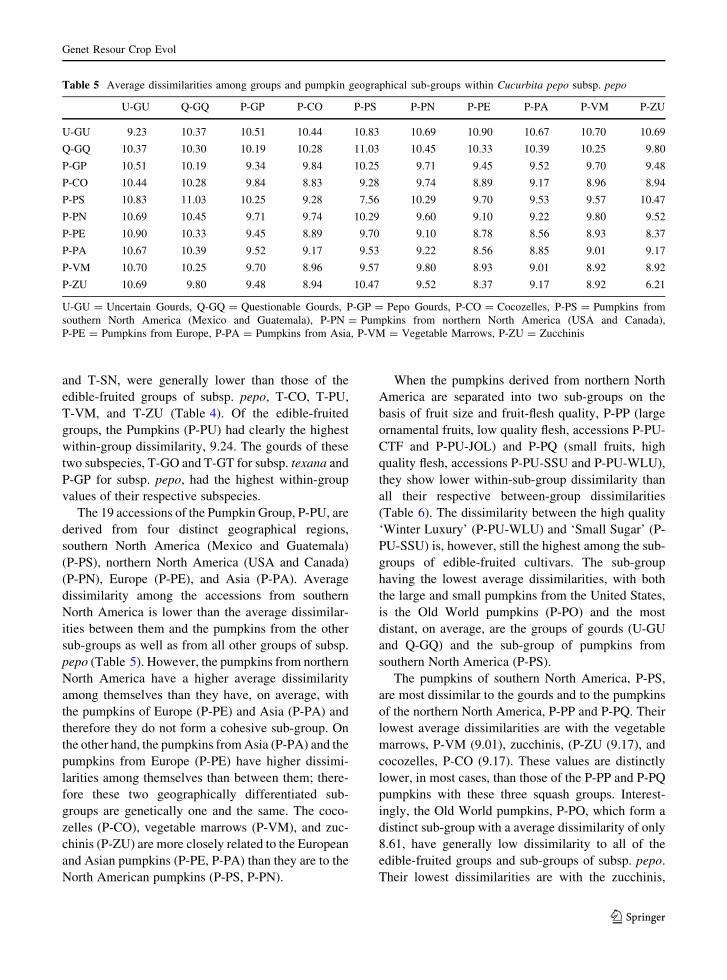
and T-SN, were generally lower than those of the
edible-fruited groups of subsp. pepo, T-CO, T-PU,
T-VM, and T-ZU (Table 4). Of the edible-fruited
groups, the Pumpkins (P-PU) had clearly the highest
within-group dissimilarity, 9.24. The gourds of these
two subspecies, T-GO and T-GT for subsp. texana and
P-GP for subsp. pepo, had the highest within-group
values of their respective subspecies.
The 19 accessions of the Pumpkin Group, P-PU, are
derived from four distinct geographical regions,
southern North America (Mexico and Guatemala)
(P-PS), northern North America (USA and Canada)
(P-PN), Europe (P-PE), and Asia (P-PA). Average
dissimilarity among the accessions from southern
North America is lower than the average dissimilar-
ities between them and the pumpkins from the other
sub-groups as well as from all other groups of subsp.
pepo (Table 5). However, the pumpkins from northern
North America have a higher average dissimilarity
among themselves than they have, on average, with
the pumpkins of Europe (P-PE) and Asia (P-PA) and
therefore they do not form a cohesive sub-group. On
the other hand, the pumpkins from Asia (P-PA) and the
pumpkins from Europe (P-PE) have higher dissimi-
larities among themselves than between them; there-
fore these two geographically differentiated sub-
groups are genetically one and the same. The coco-
zelles (P-CO), vegetable marrows (P-VM), and zuc-
chinis (P-ZU) are more closely related to the European
and Asian pumpkins (P-PE, P-PA) than they are to the
North American pumpkins (P-PS, P-PN).
When the pumpkins derived from northern North
America are separated into two sub-groups on the
basis of fruit size and fruit-flesh quality, P-PP (large
ornamental fruits, low quality flesh, accessions P-PU-
CTF and P-PU-JOL) and P-PQ (small fruits, high
quality flesh, accessions P-PU-SSU and P-PU-WLU),
they show lower within-sub-group dissimilarity than
all their respective between-group dissimilarities
(Table 6). The dissimilarity between the high quality
‘Winter Luxury’ (P-PU-WLU) and ‘Small Sugar’ (P-
PU-SSU) is, however, still the highest among the sub-
groups of edible-fruited cultivars. The sub-group
having the lowest average dissimilarities, with both
the large and small pumpkins from the United States,
is the Old World pumpkins (P-PO) and the most
distant, on average, are the groups of gourds (U-GU
and Q-GQ) and the sub-group of pumpkins from
southern North America (P-PS).
The pumpkins of southern North America, P-PS,
are most dissimilar to the gourds and to the pumpkins
of the northern North America, P-PP and P-PQ. Their
lowest average dissimilarities are with the vegetable
marrows, P-VM (9.01), zucchinis, (P-ZU (9.17), and
cocozelles, P-CO (9.17). These values are distinctly
lower, in most cases, than those of the P-PP and P-PQ
pumpkins with these three squash groups. Interest-
ingly, the Old World pumpkins, P-PO, which form a
distinct sub-group with a average dissimilarity of only
8.61, have generally low dissimilarity to all of the
edible-fruited groups and sub-groups of subsp. pepo.
Their lowest dissimilarities are with the zucchinis,
Table 5 Average dissimilarities among groups and pumpkin geographical sub-groups within Cucurbita pepo subsp. pepo
U-GU Q-GQ P-GP P-CO P-PS P-PN P-PE P-PA P-VM P-ZU
U-GU 9.23 10.37 10.51 10.44 10.83 10.69 10.90 10.67 10.70 10.69
Q-GQ 10.37 10.30 10.19 10.28 11.03 10.45 10.33 10.39 10.25 9.80
P-GP 10.51 10.19 9.34 9.84 10.25 9.71 9.45 9.52 9.70 9.48
P-CO 10.44 10.28 9.84 8.83 9.28 9.74 8.89 9.17 8.96 8.94
P-PS 10.83 11.03 10.25 9.28 7.56 10.29 9.70 9.53 9.57 10.47
P-PN 10.69 10.45 9.71 9.74 10.29 9.60 9.10 9.22 9.80 9.52
P-PE 10.90 10.33 9.45 8.89 9.70 9.10 8.78 8.56 8.93 8.37
P-PA 10.67 10.39 9.52 9.17 9.53 9.22 8.56 8.85 9.01 9.17
P-VM 10.70 10.25 9.70 8.96 9.57 9.80 8.93 9.01 8.92 8.92
P-ZU 10.69 9.80 9.48 8.94 10.47 9.52 8.37 9.17 8.92 6.21
U-GU = Uncertain Gourds, Q-GQ = Questionable Gourds, P-GP = Pepo Gourds, P-CO = Cocozelles, P-PS = Pumpkins from
southern North America (Mexico and Guatemala), P-PN = Pumpkins from northern North America (USA and Canada),
P-PE = Pumpkins from Europe, P-PA = Pumpkins from Asia, P-VM = Vegetable Marrows, P-ZU = Zucchinis
Genet Resour Crop Evol
123
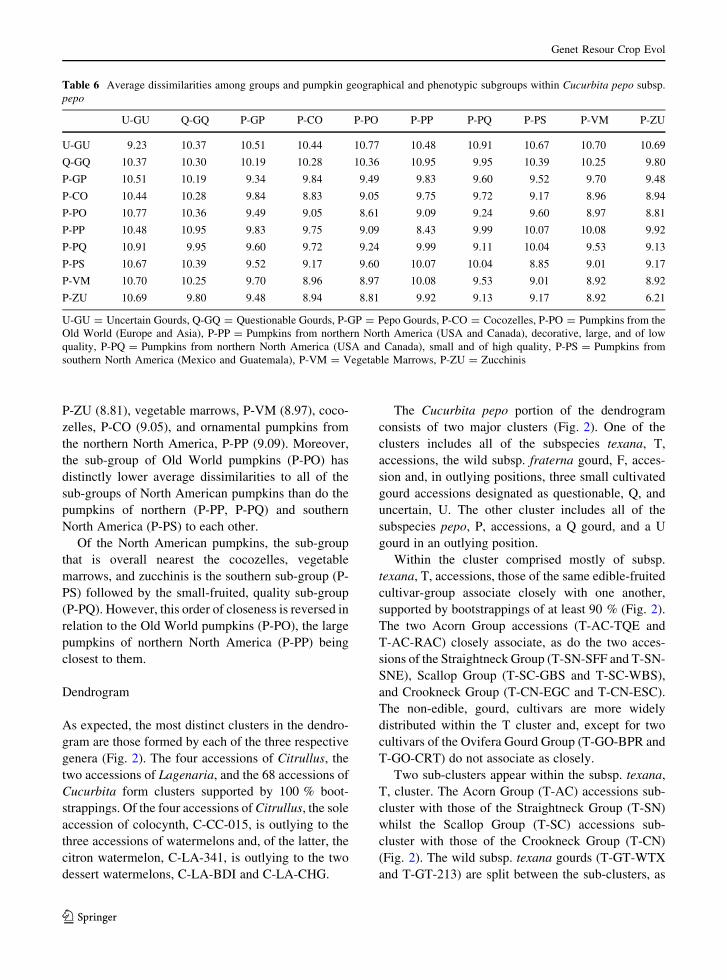
P-ZU (8.81), vegetable marrows, P-VM (8.97), coco-
zelles, P-CO (9.05), and ornamental pumpkins from
the northern North America, P-PP (9.09). Moreover,
the sub-group of Old World pumpkins (P-PO) has
distinctly lower average dissimilarities to all of the
sub-groups of North American pumpkins than do the
pumpkins of northern (P-PP, P-PQ) and southern
North America (P-PS) to each other.
Of the North American pumpkins, the sub-group
that is overall nearest the cocozelles, vegetable
marrows, and zucchinis is the southern sub-group (P-
PS) followed by the small-fruited, quality sub-group
(P-PQ). However, this order of closeness is reversed in
relation to the Old World pumpkins (P-PO), the large
pumpkins of northern North America (P-PP) being
closest to them.
Dendrogram
As expected, the most distinct clusters in the dendro-
gram are those formed by each of the three respective
genera (Fig. 2). The four accessions of Citrullus, the
two accessions of Lagenaria, and the 68 accessions of
Cucurbita form clusters supported by 100 % boot-
strappings. Of the four accessions of Citrullus, the sole
accession of colocynth, C-CC-015, is outlying to the
three accessions of watermelons and, of the latter, the
citron watermelon, C-LA-341, is outlying to the two
dessert watermelons, C-LA-BDI and C-LA-CHG.
The Cucurbita pepo portion of the dendrogram
consists of two major clusters (Fig. 2). One of the
clusters includes all of the subspecies texana, T,
accessions, the wild subsp. fraterna gourd, F, acces-
sion and, in outlying positions, three small cultivated
gourd accessions designated as questionable, Q, and
uncertain, U. The other cluster includes all of the
subspecies pepo, P, accessions, a Q gourd, and a U
gourd in an outlying position.
Within the cluster comprised mostly of subsp.
texana, T, accessions, those of the same edible-fruited
cultivar-group associate closely with one another,
supported by bootstrappings of at least 90 % (Fig. 2).
The two Acorn Group accessions (T-AC-TQE and
T-AC-RAC) closely associate, as do the two acces-
sions of the Straightneck Group (T-SN-SFF and T-SN-
SNE), Scallop Group (T-SC-GBS and T-SC-WBS),
and Crookneck Group (T-CN-EGC and T-CN-ESC).
The non-edible, gourd, cultivars are more widely
distributed within the T cluster and, except for two
cultivars of the Ovifera Gourd Group (T-GO-BPR and
T-GO-CRT) do not associate as closely.
Two sub-clusters appear within the subsp. texana,
T, cluster. The Acorn Group (T-AC) accessions sub-
cluster with those of the Straightneck Group (T-SN)
whilst the Scallop Group (T-SC) accessions sub-
cluster with those of the Crookneck Group (T-CN)
(Fig. 2). The wild subsp. texana gourds (T-GT-WTX
and T-GT-213) are split between the sub-clusters, as
Table 6 Average dissimilarities among groups and pumpkin geographical and phenotypic subgroups within Cucurbita pepo subsp.
pepo
U-GU Q-GQ P-GP P-CO P-PO P-PP P-PQ P-PS P-VM P-ZU
U-GU 9.23 10.37 10.51 10.44 10.77 10.48 10.91 10.67 10.70 10.69
Q-GQ 10.37 10.30 10.19 10.28 10.36 10.95 9.95 10.39 10.25 9.80
P-GP 10.51 10.19 9.34 9.84 9.49 9.83 9.60 9.52 9.70 9.48
P-CO 10.44 10.28 9.84 8.83 9.05 9.75 9.72 9.17 8.96 8.94
P-PO 10.77 10.36 9.49 9.05 8.61 9.09 9.24 9.60 8.97 8.81
P-PP 10.48 10.95 9.83 9.75 9.09 8.43 9.99 10.07 10.08 9.92
P-PQ 10.91 9.95 9.60 9.72 9.24 9.99 9.11 10.04 9.53 9.13
P-PS 10.67 10.39 9.52 9.17 9.60 10.07 10.04 8.85 9.01 9.17
P-VM 10.70 10.25 9.70 8.96 8.97 10.08 9.53 9.01 8.92 8.92
P-ZU 10.69 9.80 9.48 8.94 8.81 9.92 9.13 9.17 8.92 6.21
U-GU = Uncertain Gourds, Q-GQ = Questionable Gourds, P-GP = Pepo Gourds, P-CO = Cocozelles, P-PO = Pumpkins from the
Old World (Europe and Asia), P-PP = Pumpkins from northern North America (USA and Canada), decorative, large, and of low
quality, P-PQ = Pumpkins from northern North America (USA and Canada), small and of high quality, P-PS = Pumpkins from
southern North America (Mexico and Guatemala), P-VM = Vegetable Marrows, P-ZU = Zucchinis
Genet Resour Crop Evol
123
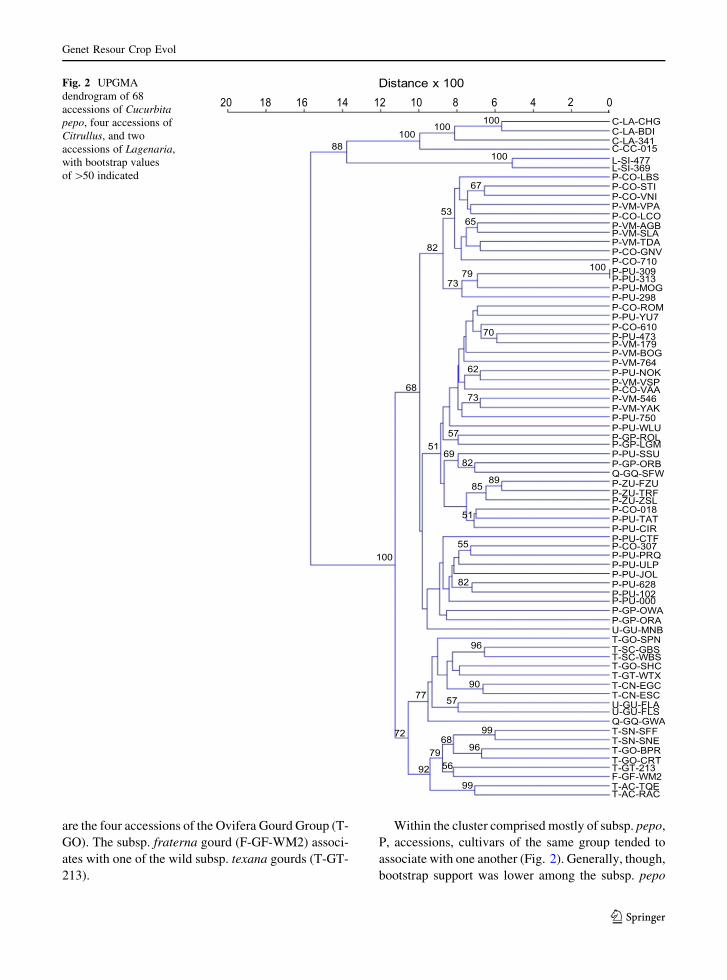
are the four accessions of the Ovifera Gourd Group (T-
GO). The subsp. fraterna gourd (F-GF-WM2) associ-
ates with one of the wild subsp. texana gourds (T-GT-
213).
Within the cluster comprised mostly of subsp. pepo,
P, accessions, cultivars of the same group tended to
associate with one another (Fig. 2). Generally, though,
bootstrap support was lower among the subsp. pepo
Fig. 2 UPGMA
dendrogram of 68
accessions of Cucurbita
pepo, four accessions of
Citrullus, and two
accessions of Lagenaria,
with bootstrap values
of [50 indicated
Genet Resour Crop Evol
123
accessions than among the subsp. texana accessions.
The three accessions of the Zucchini Group (P-ZU)
sub-clustered with one another, supported by fairly
high bootstrapping. Four strongly ribbed pumpkins
from Guatemala and Mexico, P-PU-298, -309, -313,
and -MOG, also sub-clustered with one another. One
sub-cluster containing 10 accessions consisted only of
cultivars of the Cocozelle Group (P-CO) and the
Vegetable Marrow Group (P-VM), and this sub-
cluster associated with the one consisting of four
strongly ribbed pumpkins. Another sub-cluster con-
taining 10 accessions consisted almost entirely of
pumpkins and gourds.
Principal component analyses
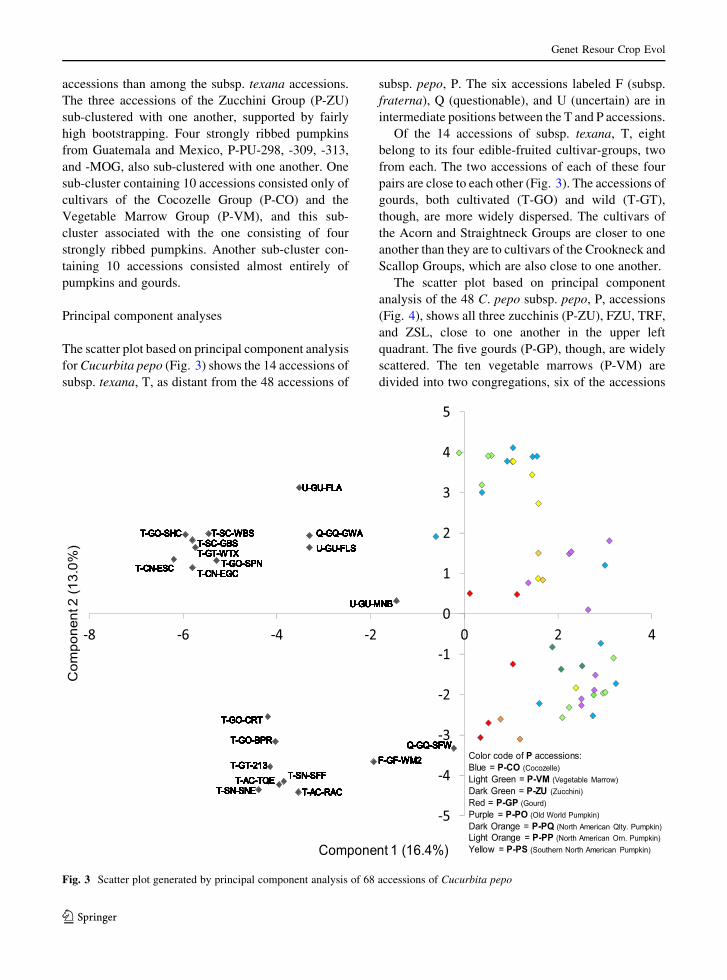
The scatter plot based on principal component analysis
for Cucurbita pepo (Fig. 3) shows the 14 accessions of
subsp. texana, T, as distant from the 48 accessions of
subsp. pepo, P. The six accessions labeled F (subsp.
fraterna), Q (questionable), and U (uncertain) are in
intermediate positions between the T and P accessions.
Of the 14 accessions of subsp. texana, T, eight
belong to its four edible-fruited cultivar-groups, two
from each. The two accessions of each of these four
pairs are close to each other (Fig. 3). The accessions of
gourds, both cultivated (T-GO) and wild (T-GT),
though, are more widely dispersed. The cultivars of
the Acorn and Straightneck Groups are closer to one
another than they are to cultivars of the Crookneck and
Scallop Groups, which are also close to one another.
The scatter plot based on principal component
analysis of the 48 C. pepo subsp. pepo, P, accessions
(Fig. 4), shows all three zucchinis (P-ZU), FZU, TRF,
and ZSL, close to one another in the upper left
quadrant. The five gourds (P-GP), though, are widely
scattered. The ten vegetable marrows (P-VM) are
divided into two congregations, six of the accessions
-5
-4
-3
-2
-1
0
1
2
3
4
5
-8 -6 -4 -2 0 2 4
Com
pone
nt 2
(13.
0%)
Component 1 (16.4%)
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Q-GQ-SFW
T-SN-SFFT-AC-TQET-SN-SNE T-AC-RAC
F-GF-WM2T-GT-213
T-GO-BPR
T-GO-CRT
U-GU-MNB
T-GT-WTXT-SC-GBS
T-GO-SPNT-CN-EGC
T-SC-WBS
T-CN-ESC
T-GO-SHC
U-GU-FLA
Q-GQ-GWAU-GU-FLS
Color code of P accessions:Blue = P-CO (Cocozelle)Light Green = P-VM (Vegetable Marrow)Dark Green = P-ZU (Zucchini)Red = P-GP (Gourd)Purple = P-PO (Old World Pumpkin)Dark Orange = P-PQ (North American Qlty. Pumpkin)Light Orange = P-PP (North American Orn. Pumpkin)Yellow = P-PS (Southern North American Pumpkin)
Fig. 3 Scatter plot generated by principal component analysis of 68 accessions of Cucurbita pepo
Genet Resour Crop Evol
123

toward the left and four toward the extreme right. Six
of the 11 cocozelles (P-CO) associate with the latter,
so the upper right quadrant contains exclusively ten
accessions derived from these two cultivar-groups.
The remaining five cocozelles are more widely
scattered. Of the pumpkins, the two small, quality
(P-PQ) pumpkins are toward the extreme left, distant
from the large, ornamental (P-PP) pumpkins and from
pumpkins of southern North America (P-PS), most of
which are at the extreme lower right. In contrast, the
Old World pumpkins (P-PO) gravitate more toward
the center, nearest the ornamental pumpkins (P-PP),
and appear to be the closest sub-group to the
cocozelles (P-CO) and vegetable marrows (P-VM).
Discussion
HFO–TAG markers have been used to obtain an
improved definition of relationships in Citrullus spp.
germplasm and have contributed to mapping the
watermelon genome (Levi et al. 2010, 2011, 2013).
In PCR experiments examining the ability of primers
to amplify fragments from a watermelon cDNA
library, the HFO–TAGs primers produced consider-
ably more fragments (an average of 6.44 fragments per
primer) compared with ISSR and RAPD primers (an
average of 3.59 and 2.49 fragments per primer,
respectively). Indeed, HFO–TAG markers had far
greater capability than RAPD and ISSR markers for
revealing polymorphisms, readily distinguishing
among various dessert watermelon cultivars and
indicating relationships that are in close agreement
with known pedigrees (Levi et al. 2010). For Cucur-
bita pepo, the HFO–TAG markers provide results that
are consistent with those obtained using other nuclear
DNA markers (Lebeda et al. 2007; Formisano et al.
2012; Gong et al. 2012), but in addition reveal
relationships concerning the evolution of its econom-
ically more important lineage, C. pepo subsp. pepo.
-5
-4
-3
-2
-1
0
1
2
3
4
5
-4 -3 -2 -1 0 1 2 3 4 5
TRF
FZU
ZSL
TAT018
CIRVAA
ORB
WLU
SSU
BOGNOK
LGM
546 179764
YAK 473ROM
750VSP YU7
307
610
ROL
LBS
309, 313
CTF ULP
OWA102
JOL 628
ORA 000
PRQMOG
298
710
STI
LCO VPAAGB
TDA
GNV
VNI
SLA
Component 1 (15.6%)
Com
pone
nt 2
(8.9
%)
Color code:Blue = CO (Cocozelle)Light green = VM (Vegetable Marrow)Dark green = ZU (Zucchini)Red = GP (Gourd)Purple = PO (Old World Pumpkin)Dark Orange = PQ (North America Quality Pumpkin)Light Orange = PP (North America Ornamental Pumpkin)Yellow = PS (Southern North America Pumpkin)
Fig. 4 Scatter plot generated by principal component analysis of 48 accessions of Cucurbita pepo subsp. pepo
Genet Resour Crop Evol
123

The HFO–TAG markers present Cucurbita pepo as
more distant from Citrullus and Lagenaria (average
dissimilarities 16.04 and 14.13, respectively) than
either of those two genera are from each other (average
dissimilarity 13.78; Table 3). Both Citrullus and
Lagenaria are likely natives of Africa (Robinson and
Decker-Walters 1997) and of the tribe Benincaseae
whilst Cucurbita is one of 11 genera in the tribe
Cucurbitae, the distribution of which is almost entirely
in the Americas (Schaefer and Renner 2011).
As expected, the HFO–TAGs clearly show the
dichotomy within Cucurbita pepo, between subsp.
texana and subsp. pepo (Table 3; Fig. 2). The HFO–
TAGs also show that the smallest-fruited gourds,
labelled Q (Questionable) and U (uncertain), are
generally intermediate between the two major culti-
vated subspecies (Tables 3 and 4; Fig. 3), as has been
observed using AFLPs and SSRs (Paris et al. 2003;
Gong et al. 2012).
The HFO–TAGs are, in general, consistent with the
horticultural classification into cultivar-groups based
mainly on fruit shape (Paris 1986). Nonetheless, the
results differ in some ways from those obtained with
other nuclear markers. Presently, a dichotomy is
observed within C. pepo subsp. texana above the level
of cultivar-group, with the acorn squash, straightneck
squash, and some gourds separated from the crook-
neck squash, scallop squash, and other gourds
(Table 4; Figs. 2 and 3). The four edible-fruited
cultivar-groups of subsp. pepo, Cocozelle, Pumpkin,
Vegetable Marrow, and Zucchini, are distinguishable
from one another by having lower within-group
dissimilarity values than their respective between-
group dissimilarity values (Table 4). They are thus
more clearly separated than observed with various
other nuclear DNA markers (Katzir et al. 2000; Ferriol
et al. 2003; Paris et al. 2003; Formisano et al. 2012;
Gong et al. 2012), but nonetheless the first three are not
completely separated in the dendrogram (Fig. 2) and
scatter plots (Figs. 3 and 4). However, analysis of the
present results with HFO–TAG markers reveals for the
first time why the cocozelles, pumpkins, and vegetable
marrows are not completely separated from one
another and can explain how the long-fruited groups,
the cocozelles and vegetable marrows, evolved.
Of the cultivar-groups of Cucurbita pepo subsp.
pepo, the Pumpkin Group contains the highest within-
group dissimilarity (Table 4). This is to be expected as
the Pumpkin Group is the oldest of the edible-fruited
cultivar-groups (Paris 2000), having originated
approximately 10,000 years ago (Smith 1997; Tepp-
ner 2004). Pumpkins having the same geographical
origin and set of phenotypic characteristics would be
expected to be more closely related to one another than
to pumpkins of different geographical origins and
appearance. Indeed, the pumpkin accessions of south-
ern North America, which bear fruits of modest size
(usually \5 kg) and are often striped and strongly
ribbed, usually with thick lignified rinds, differ greatly
in appearance from the pumpkins of northern North
America, which are grooved with non-lignified rinds,
and orange without stripes (Paris 2000; Teppner
2000). The pumpkins derived from different regions
of the North American continent are genetically quite
distant from one another (Table 5). The pumpkin
cultigens of northern North America, though, are split
among themselves into two distinct sub-groups
(Table 6) which can be distinguished phenotypically
by differing fruit size and quality. One of these sub-
groups, consisting of ‘Small Sugar’ and ‘Winter
Luxury’, has smaller fruits with high-quality flesh
and are usually consumed. The other, consisting of
‘Connecticut Field’ and ‘Jack-O-Lantern’, has larger
fruits (usually [5 kg) with low-quality flesh that are
usually used as ornaments for Hallowe’en. The three
pumpkin sub-groups from North America became
differentiated from one another on that continent
before the arrival of Europeans, probably long before
1492. All three are abundantly depicted in botanical
herbals of the Renaissance (Paris 2000; Teppner 2000)
and two of them appear in festoons produced in Rome
between 1515 and 1518 (Janick and Paris 2006). All of
the pumpkins are ultimately derived from southern
North America (Decker-Walters 1990) but it is not
known where and when the two more northerly sub-
groups of pumpkins were founded. Perhaps each was
derived through gradual diffusion northward, with
selection occurring in the United States for orange
external color and larger size or improved culinary
value. All three of the North American sub-groups are
closer to the sub-group of Old World (Euro-
pean ? Asian) pumpkins than they are to one another
(Table 6).
The Old World pumpkins encompass wide pheno-
typic variation (Paris 2000; Ferriol and Pico 2008),
ranging in size from medium, approximately 2 kg, to
quite large, 20 kg or more (Fig. 1). Most have thin
lignified rinds and are not grooved. Many are ribbed,
Genet Resour Crop Evol
123

but usually not as prominently as the pumpkins of
southern North America. Many have dark green
stripes on an orange, yellow, or light green back-
ground. They flower early enough so as to be able to
set and ripen their fruits in short-season areas of
Europe and Asia. Phenotypically, therefore, the Old
World pumpkins possess a combination of traits which
is intermediate to that possessed by pumpkins from the
southern part versus the northern part of North
America. The genetic intermediacy of the Old World
pumpkins (Table 6) is consistent with their having
originated from spontaneous crossing among pump-
kins of the different North American subgroups. The
intercrossing among pumpkin subgroups had to have
occurred after they were brought to Europe and grown
in close proximity. For example, Leonhart Fuchs, the
German botanist, had a garden in which he grew exotic
plants. He produced a large collection of plant
illustrations in approximately 1562, referred to as the
Vienna Codex (Meyer et al. 1999), among which are
images of Cucurbita pepo. Some of the C. pepo
images are clearly of inter-group hybrids (Paris 2000).
Indeed, southern and northern North American sub-
groups of pumpkins were grown in near proximity to
one another in the environs of Rome already within
25 years of the first voyage of Columbus to the
Americas (Janick and Paris 2006).
The accessions of the long-fruited cultivar-groups
of subsp. pepo, Cocozelle, Vegetable Marrow, and
Zucchini, are each more closely related to the Old
World subgroup of pumpkins than they are to any of
the North American pumpkin subgroups (Table 6).
Some cocozelle and vegetable marrow accessions also
show affinity with pumpkins from southern North
America (Fig. 2). Variant plants of southern North
American landrace pumpkins can produce elongate
fruits, but there is no evidence for the existence of
uniformly long-fruited cultivars of C. pepo subsp.
pepo in North America in the first centuries after the
European contact with that continent (Zhiteneva 1930;
Hernandez Bermejo and Leon 1994; Paris 2000;
Teppner 2000). Therefore, the development of the
long-fruited cultivars of subsp. pepo seems to have
occurred in the Old World over the past 500 years.
Iconographic records indicate that incipient cocozelles
and vegetable marrows were present in Europe within
a century of the arrival of C. pepo pumpkins from
North America (Paris 2000; Paris and Janick 2005).
Gene exchange between pumpkins from northern
North America and pumpkins from southern North
America not only led to the development of the
European and Asian pumpkins, but also to the
development of three entirely new cultivar-groups
having elongate fruits, the cocozelles, vegetable
marrows and, eventually, the zucchinis.
Selection for deviation from ancestral round fruit-
edness was not a one-time occurrence in the develop-
ment of Cucurbita pepo cultivars. In C. pepo subsp.
texana, several cultivars of the flat scallop squash are
illustrated in mid-sixteenth century herbals and the
elongate crookneck squash is also an ancient group
(Paris 2000; Teppner 2000). The de novo evolution of
long-fruited cultigens of C. pepo subsp. pepo, within a
century of the arrival of round-fruited cultigens of this
subspecies in Europe, suggests that this germplasm
underwent intensive selection there during the six-
teenth century. The deviation from ancestral fruit
roundness in the cultivated cucurbits is associated with
a greater proportion of the firm, fleshy mesocarp tissue
and colored exocarp at the expense of the relative
volume of the seedy endocarp (Sinnott and Durham
1929). Such developments are therefore associated
with abandoning the original uses of cucurbit fruits by
people, consumption of the mature fruit flesh or seeds,
in favor of consuming the whole young fruits (Paris
1989). Selection for elongated cucurbit fruits has a
long tradition in the Old World. Snake melons,
Cucumis melo L., and long bottle gourds, Lagenaria
siceraria, were already in widespread use as food by
Latin-, Greek-, and Hebrew-speaking cultures of the
Mediterranean Basin during the time of the Roman
Empire (Janick et al. 2007). Indeed, the snake melon
was the most important cucurbit crop in Mediterra-
nean lands through medieval times (Paris 2012). So it
is easily understood how, when a cucurbit newcomer
arrived in this region, Cucurbita pepo subsp. pepo, it
was directionally selected for elongate fruits. This
selection occurred most intensively in Italy but also in
Asia Minor (Gong et al. 2012) and Spain (Formisano
et al. 2012). In each of these places, the founder
populations of pumpkins from which elongate fruits
were selected may well have been different, the result
being the fuzzy demarcation between the cocozelles,
vegetable marrows, and pumpkins seen today (Figs. 2
and 4).
Of the subsp. pepo gourds, all of which are small in
size and round in shape, two cultivars from South
Africa, ‘Rolet’ and ‘Little Gem’, show affinity to each
Genet Resour Crop Evol
123

other (Fig. 2). ‘Orange Ball’ is closest to the Ques-
tionable Gourd ‘Small Flat Warted’, one of the
smallest gourds and near the genetic center of the
species (Gong et al. 2012). Although ‘Orange’ and
‘Orange Warted’ are closest to several pumpkins
(Figs. 2 and 4), neither they nor any of the P-GP,
Q-GQ, or U-GU gourds seems to have made a
significant contribution to the radiation of the edible-
fruited groups and sub-groups of subsp. pepo, as they
are distant (dissimilarity [ 9.40) to all of them
(Tables 4, 5, and 6).
Conclusions
Polymorphisms of HFO–TAG markers, which are
derived mostly from coding regions of the genome,
have resulted in an assessment of genetic diversity
within Cucurbita pepo that closely corresponds with
the morphological variation exhibited by this species.
The four edible-fruited cultivar-groups of C. pepo
subsp. texana are clearly distinguished, Acorn, Crook-
neck, Scallop, and Straightneck. The four edible-
fruited cultivar-groups of C. pepo subsp. pepo, Coco-
zelle, Pumpkin, Vegetable Marrow, and Zucchini, are
more clearly differentiated by the HFO–TAGs than
heretofore seen with other DNA markers, but are
nonetheless not as distinct as those of C. pepo subsp.
texana. A dichotomy was observed within C. pepo
subsp. texana above the level of cultivar-group, with
the acorn squash, straightneck squash, and some
gourds separated from the crookneck squash, scallop
squash, and other gourds. In C. pepo subsp. pepo, three
populations of North American pumpkins are differ-
entiated, one southerly and two northerly, one of the
latter having small, high-quality fruits and the other
having large, ornamental fruits. Apparently, these
three pumpkin sub-groups had been isolated from one
another on their native continent, but records indicate
that they were grown in close proximity in Europe
soon after the initial contacts of European explorers
with various parts of North America over 500 years
ago. The present results show that the cocozelles,
vegetable marrows, and zucchinis are more closely
allied with the Old World cultivars of pumpkins than
with any of the North American source populations.
Acknowledgments We thank Laura Massey of the USDA–
ARS, Charleston, for expert technical assistance.
References
Andres TC (1987) Cucurbita fraterna, the closest wild relative
and progenitor of C. pepo. Cucurbit Genet Coop Rep
10:69–71
Bailey LH (1943) Species of Cucurbita. Gentes Herb 6:266–322
Cowan CW (1997) Evolutionary changes associated with the
domestication of Cucurbita pepo. In: Gremillion KJ (ed)
People, plants and landscapes: studies in paleoethnobota-
ny. University of Alabama Press, Tuscaloosa, pp 63–85
Decker DS, Wilson HD (1987) Allozyme variation in the Cu-
curbita pepo complex: C. pepo var. ovifera vs C. texana.
Syst Bot 12:263–273
Decker-Walters DS (1990) Evidence for multiple domestica-
tions of Cucurbita pepo. In: Bates DM, Robinson RW,
Jeffrey C (eds) Biology and utilization of the Cucurbita-
ceae. Cornell University Press, Ithaca, pp 96–101
Duchesne AN (1786) Essai sur l’histoire naturelle des courges.
Panckoucke, Paris
Emerson RA (1910) The inheritance of sizes and shapes in
plants. Am Nat 44:739–746
Erwin AT (1931) Nativity of the cucurbits. Bot Gaz 91:105–108
Ferriol M, Pico B (2008) Pumpkin and winter squash. In: Pro-
hens J, Nuez F (eds) Handbook of plant breeding, vegeta-
bles I. Springer, New York, pp 317–349
Ferriol M, Pico B, Nuez F (2003) Genetic diversity of a germ-
plasm collection of Cucurbita pepo using SRAP and AFLP
markers. Theor Appl Genet 107:271–282
Formisano G, Roig C, Esteras C, Ercolano MR, Nuez F, Mon-
forte AJ, Pico MB (2012) Genetic diversity of Spanish
Cucurbita pepo landraces: an unexploited resource for
summer squash breeding. Genet Resour Crop Evol
59:1169–1184
Goldman A (2004) The compleat squash. Artisan, New York
Gong L, Paris HS, Nee MH, Stift G, Pachner M, Vollmann J,
Lelley T (2012) Genetic relationships and evolution in
Cucurbita pepo (pumpkin, squash, gourd) as revealed by
simple sequence repeat polymorphisms. Theor Appl Genet
124:875–891
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleonto-
logical statistics software package for education and data
analysis. Palaeontol Electron 4(1 art. 4):9
Hernandez Bermejo JE, Leon J (1994) Neglected crops, 1492
from a different perspective. FAO plant production and
protection ser no 26. Rome: FAO, pp 63–77
Janick J, Paris HS (2006) The cucurbit images (1515–1518) of
the Villa Farnesina, Rome. Ann Bot 97:165–176
Janick J, Paris HS, Parrish DC (2007) The cucurbits of Medi-
terranean antiquity: identification of taxa from ancient
images and descriptions. Ann Bot 100:1441–1457
Katzir N, Tadmor Y, Tzuri G, Leshzeshen E, Mozes-Daube N,
Danin-Poleg Y, Paris HS (2000) Further ISSR and preliminary
SSR analysis of relationships among accessions of Cucurbita
pepo. In: Katzir N, Paris HS (eds) Proceedings of Cucurbita-
ceae 2000, the 7th Eucarpia meeting on cucurbit genetics and
breeding, vol 510. Acta Horticulturae, pp 433–439
Kosman E, Leonard KJ (2005) Similarity coefficients for
molecular markers in studies of genetic relationships
between individuals for haploid, diploid, and polyploid
species. Mol Ecol 14:415–424
Genet Resour Crop Evol
123

Lebeda A, Widrlechner MP, Staub J, Ezura H, Zalapa J,
Kristkova E (2007) Cucurbits (Cucurbitaceae; Cucumis
spp., Cucurbita spp., Citrullus spp.). In: Singh RJ (ed)
Genetic resources, chromosome engineering, and crop
improvement, vol 3. CRC Press, Boca Raton, pp 271–376
Levi A, Thomas C (1999) An improved procedure for isolation
of high quality DNA from watermelon and melon leaves.
Cucurbit Genet Coop Rep 22:41–42
Levi A, Thomas CE, Trebitsh T, Salman A, King J, Karalius J,
Newman M, Reddy OUK, Xu Y, Zhang X (2006) An
extended linkage map for watermelon based on SRAP,
AFLP, SSR, ISSR, and RAPD markers. J Am Soc Hortic
Sci 131:393–402
Levi A, Wechter WP, Harris-Shultz KR, Davis AR, Fie Z (2010)
High-frequency oligonucleotides in watermelon expressed
sequenced tag-unigenes are useful in producing polymor-
phic polymerase chain reaction markers among water-
melon genotypes. J Am Hortic Sci 135:369–378
Levi A, Wechter WP, Massey LM, Carter L, Hopkins D (2011)
Genetic linkage map of Citrullus lanatus var. citroides
chromosomal segments into the watermelon cultivar
Crimson Sweet (Citrullus lanatus var. lanatus) genome.
Am J Plant Sci 2:93–110
Levi A, Thies JA, Wechter WP, Harrison HF, Simmons AM,
Reddy UK, Nimmakayala P, Fei Z (2013) High frequency
oligonucleotides: targeting active gene (HFO-TAG)
markers revealed wide genetic diversity among Citrullus
spp. accessions useful for enhancing disease or pest resis-
tance in watermelon cultivars. Genet Resour Crop Evol
60:427–440
Lira R, Montes S (1994) Cucurbits (Cucurbita spp.). In: Her-
nandez JE, Leon J (eds) Neglected crops: 1492 from a
different perspective. FAO, Rome, pp 63–77
Meyer FG, Trueblood EE, Heller JL (1999) The great herbal of
Leonhart Fuchs, vol 1, commentary. Stanford University
Press, Stanford
Nee M (1990) The domestication of Cucurbita. Econ Bot 44(3
suppl):56–68
Paris HS (1986) A proposed subspecific classification for Cu-
curbita pepo. Phytologia 61:133–138
Paris HS (1989) Historical records, origins, and development of
the edible cultivar groups of Cucurbita pepo (Cucurbita-
ceae). Econ Bot 43:423–443
Paris HS (2000) History of the cultivar-groups of Cucurbita
pepo. Hortic Rev 25(2001):71–170
Paris HS (2001) Characterization of the Cucurbita pepo col-
lection at the Newe Ya‘ar Research Center, Israel. Plant
Genet Resour Newsl 126:41–45
Paris HS (2007) The drawings of Antoine Nicolas Duchesne for
his natural history of the gourds. In: Erard C (ed). Des
planches et des mots. Publications Scientifiques, Museum
National d’Histoire Naturelle, Paris
Paris HS (2012) Semitic-language records of snake melons
(Cucumis melo, Cucurbitaceae) in the medieval period and
the ‘‘piqqus’’ of the ‘‘faqqous’’. Genet Resour Crop Evol
59:31–38
Paris HS, Janick J (2005) Early evidence for the culinary use of
squash flowers in Italy. Chron Hortic 45(2):20–21
Paris HS, Nerson H (2003) Seed dimensions in the subspecies
and cultivar-groups of Cucurbita pepo. Genet Resour Crop
Evol 50:615–625
Paris HS, Yonash N, Portnoy V, Mozes-Daube N, Tzuri G,
Katzir N (2003) Assessment of genetic relationships in
Cucurbita pepo (Cucurbitaceae) using DNA markers.
Theor Appl Genet 106:971–978
Paris HS, Burger Y, Schaffer AA (2006) Genetic variability and
introgression of horticulturally valuable traits in squash and
pumpkins of Cucurbita pepo. Isr J Plant Sci 54:223–231
Paris HS, Lebeda A, Kristkova E, Andres TC, Nee MH (2012)
Parallel evolution under domestication and phenotypic
differentiation of the cultivated subspecies of Cucurbita
pepo. Econ Bot 66:71–90
Robinson RW, Decker-Walters DS (1997) Cucurbits. CAB
International, Wallingford, pp 85, 89–90
Schaefer H, Renner SS (2011) Cucurbitaceae. In: Kubitzki K
(ed) The families and genera of vascular plants, vol 10,
Eudicots. Springer, New York, pp 112–174
Sinnott EW (1935) Evidence for the existence of genes con-
trolling shape. Genetics 20:12–21
Sinnott EW, Durham GB (1929) Developmental history of the
fruit in lines of Cucurbita pepo differing in fruit shape. Bot
Gaz 87:411–421
Smith BD (1997) The initial domestication of Cucurbita pepo in
the Americas 10,000 years ago. Science 276:932–934
Smith BD (2006) Eastern North America as an independent
center of plant domestication. Proc Natl Acad Sci USA
103:12223–12228
Teppner H (2000) Cucurbita pepo (Cucurbitaceae)—history,
seed coat types, thin coated seeds and their genetics. Phy-
ton 40:1–42
Teppner H (2004) Notes on Lagenaria and Cucurbita (Cucur-
bitaceae)—review and new contributions. Phyton
44:245–308
Whitaker TW, Carter GF (1946) Critical notes on the origin and
domestication of the cultivated species of Cucurbita. Am J
Bot 33:10–15
Whitaker TW, Cutler HC (1971) Prehistoric cucurbits from the
Valley of Oaxaca. Econ Bot 25:123–127
Whitaker TW, Cutler HC (1986) Cucurbits from preceramic
levels at Guila Naquitz. In: Flannery KV (ed) Guila Na-
quitz: archaic foraging and early agriculture in Oaxaca,
Mexico. Academic Press, Orlando, pp 275–279
Zhiteneva NE (1930) The world’s assortment of pumpkins.
Trudy Prikl Bot Genet Selek 23:157–207
Genet Resour Crop Evol
123
Related Documents






![Genetic relationships and evolution in Cucurbita pepo (pumpkin, squash, gourd) as revealed by simple sequence repeat polymorphisms [2012]](https://static.cupdf.com/doc/110x72/6331e2a483bb92fe9804247c/genetic-relationships-and-evolution-in-cucurbita-pepo-pumpkin-squash-gourd-as.jpg)




