Genetic evidence that mycobacterial FtsZ and FtsW proteins interact, and colocalize to the division site in Mycobacterium smegmatis Malini Rajagopalan a, * , Erin Maloney a , Jaroslaw Dziadek b , Marta Poplawska b , Hava Lofton a , Ashwini Chauhan a , Murty V.V.S. Madiraju a a The University of Texas Health Center at Tyler, Biomedical Research, 11937 US Hwy 271, Tyler, TX 75708, United States b Medical Biology Centre, Polish Academy of Sciences, Lodowa 106, 93-232 Lodz, Poland Received 24 April 2005; received in revised form 13 June 2005; accepted 19 June 2005 First published online 11 July 2005 Edited by R.S. Buxton Abstract We provide genetic evidence to show that the Mycobacterium tuberculosis FtsZ and FtsW proteins interact, and that these inter- actions are biologically relevant. Furthermore, we show by fluorescence microscopy that Mycobacterium smegmatis FtsW is part of its septasomal complex and colocalizes with FtsZ to the midcell sites. Colocalization experiments reveal that approximately 27% of the cells with septal Z-rings contain FtsW whereas 93% of the cells with FtsW bands are associated with FtsZ indicating that FtsW is late recruit to the septum, as in Escherichia coli. Our results suggest that mycobacterial FtsZ can localize to the septum independent of FtsW, and that interactions of FtsW with FtsZ are critical for the formation of productive FtsZ-rings and the cell division process in mycobacteria. Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Mycobacteria; Cell division; Green-fluorescent protein fusions; Recombination; Z-ring localization; Fluorescence microscopy 1. Introduction Genetic studies on the cell division process in the uni- cellular organism Escherichia coli indicate that it is a well-orchestrated event involving the concerted action of several proteins [1]. FtsZ, a key player involved in the cell division process, exhibits GTP-dependent poly- merization and GTPase activities and initiates the cell division process by assembling at the midcell site distinct structure called the Z-ring [2]. Interactions of FtsZ with itself and other cell division proteins and their orderly localization to the Z-ring are believed to result in the formation of stable cytokinetic ring and subsequent cell division. Among the several cell division proteins identi- fied, FtsA and ZipA bind to the C-terminus of FtsZ and localize independent of each other to the Z-ring [3,4]. Although FtsZ rings assemble in the absence of either FtsA or ZipA, they cannot assemble in the absence of both [5]. Other studies indicate that ZipA and FtsA have overlapping functions and that ZipA role in the cell division process in E. coli can be bypassed in the pres- ence of a gain of function mutation in ftsA [6]. FtsA, which is more conserved than ZipA, contains a con- served C-terminal amphipathic helix needed for target- ing FtsA, and consequently FtsZ to the membrane in E. coli [7]. Several studies have identified interactions of FtsZ with other cell division proteins (reviewed in [1]) and recently, using the bacterial two-hybrid system, 0378-1097/$22.00 Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsle.2005.06.043 * Corresponding author. Tel.: +1 903 877 7731; fax: +1 903 877 5969. E-mail address: [email protected] (M. Rajagopalan). www.fems-microbiology.org FEMS Microbiology Letters 250 (2005) 9–17

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.fems-microbiology.org
FEMS Microbiology Letters 250 (2005) 9–17
Genetic evidence that mycobacterial FtsZ and FtsW proteins interact,and colocalize to the division site in Mycobacterium smegmatis
Malini Rajagopalan a,*, Erin Maloney a, Jaroslaw Dziadek b, Marta Poplawska b,Hava Lofton a, Ashwini Chauhan a, Murty V.V.S. Madiraju a
a The University of Texas Health Center at Tyler, Biomedical Research, 11937 US Hwy 271, Tyler, TX 75708, United Statesb Medical Biology Centre, Polish Academy of Sciences, Lodowa 106, 93-232 Lodz, Poland
Received 24 April 2005; received in revised form 13 June 2005; accepted 19 June 2005
First published online 11 July 2005
Edited by R.S. Buxton
Abstract
We provide genetic evidence to show that the Mycobacterium tuberculosis FtsZ and FtsW proteins interact, and that these inter-actions are biologically relevant. Furthermore, we show by fluorescence microscopy that Mycobacterium smegmatis FtsW is part ofits septasomal complex and colocalizes with FtsZ to the midcell sites. Colocalization experiments reveal that approximately 27% ofthe cells with septal Z-rings contain FtsW whereas 93% of the cells with FtsW bands are associated with FtsZ indicating that FtsW islate recruit to the septum, as in Escherichia coli. Our results suggest that mycobacterial FtsZ can localize to the septum independentof FtsW, and that interactions of FtsW with FtsZ are critical for the formation of productive FtsZ-rings and the cell division processin mycobacteria.� 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Mycobacteria; Cell division; Green-fluorescent protein fusions; Recombination; Z-ring localization; Fluorescence microscopy
1. Introduction
Genetic studies on the cell division process in the uni-cellular organism Escherichia coli indicate that it is awell-orchestrated event involving the concerted actionof several proteins [1]. FtsZ, a key player involved inthe cell division process, exhibits GTP-dependent poly-merization and GTPase activities and initiates the celldivision process by assembling at the midcell site distinctstructure called the Z-ring [2]. Interactions of FtsZ withitself and other cell division proteins and their orderlylocalization to the Z-ring are believed to result in the
0378-1097/$22.00 � 2005 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2005.06.043
* Corresponding author. Tel.: +1 903 877 7731; fax: +1 903 8775969.
E-mail address: [email protected] (M. Rajagopalan).
formation of stable cytokinetic ring and subsequent celldivision. Among the several cell division proteins identi-fied, FtsA and ZipA bind to the C-terminus of FtsZ andlocalize independent of each other to the Z-ring [3,4].Although FtsZ rings assemble in the absence of eitherFtsA or ZipA, they cannot assemble in the absence ofboth [5]. Other studies indicate that ZipA and FtsA haveoverlapping functions and that ZipA role in the celldivision process in E. coli can be bypassed in the pres-ence of a gain of function mutation in ftsA [6]. FtsA,which is more conserved than ZipA, contains a con-served C-terminal amphipathic helix needed for target-ing FtsA, and consequently FtsZ to the membrane inE. coli [7]. Several studies have identified interactionsof FtsZ with other cell division proteins (reviewed in[1]) and recently, using the bacterial two-hybrid system,
. Published by Elsevier B.V. All rights reserved.

10 M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17
one study confirmed direct interactions between FtsZand ZipA, FtsZ–FtsA, FtsW–FtsQ, FtsW–FtsI proteinsof E. coli; although interactions between FtsZ and FtsWproteins have not been identified in this study [8].
The cell division process in members of mycobacteriais just beginning to be understood. These studies suggestthat FtsZ and WhmD proteins of mycobacteria areessential cell division proteins and that Mycobacterium
tuberculosis FtsZ (FtsZTB) is a target of FtsHTB protease[9–13]. Optimal levels of FtsZ are required for celldivision [11] and that M. tuberculosis ftsZ (ftsZTB) canreplace the function of Mycobacterium smegmatis coun-terpart indicating the FtsZ catalyzed steps in the celldivision processes of slow- and fast-growing membersof mycobacteria are similar [12]. Fluorescence micros-copy studies with GFP fusion proteins of FtsZTB andM. smegmatis FtsZ (FtsZSmeg) led to visualization ofFtsZ structures in the filamentous cells of M. smegmatis
overproducing FtsZ [11]. The genome sequences of M.
tuberculosis [14], Mycobacterium avium and M. smegma-
tis (www.tigr.org) lack identifiable analogs of ZipAand FtsA proteins. Consequently, the mechanism(s)involved in targeting mycobacterial FtsZ to putative celldivision site are unknown. Recently, using biochemicalapproaches in vitro interactions between FtsZTB andFtsWTB proteins have been demonstrated [15]. Thisstudy also showed that a stretch of four aspartic acidresidues located at the carboxy (C-) terminus of FtsZTB
are important for these interactions as mutant FtsZTB
protein containing alanine in place of aspartic acid(FtsZD367–370A) does not interact with FtsWTB [15]. Itis, however, unknown whether the in vitro observedinteractions between FtsZTB and FtsWTB are observedin vivo and if so, whether these interactions are biolog-ically relevant. It is also unknown whether mycobacte-rial FtsW is part of the septasomal complex, i.e.localizes to the septal ring and if so, whether it colocal-izes with FtsZ? These questions form the basis of thispaper.
2. Materials and methods
2.1. Bacterial strains and culture conditions
All plasmids were propagated in E. coli Top10 strain(Invitrogen Inc.). M. smegmatis mC2155 was originallyobtained from Dr. Bill Jacobs Jr. (Albert Einstein Col-lege of Medicine, Bronx, New York) and was culturedin Middlebrook 7H9 broth supplemented with albu-min–dextrose–sodium chloride. Mycobacterial transfor-mants were selected in the same media supplementedwith agar containing either kanamycin (25 lg/ml),hygromycin (10 lg/ml) or both [11]. Conditions for cul-turing yeast Y19 strain are as described in Clontechmanual (PT3062-1).
2.2. Molecular biology techniques: Yeast two-hybrid
plasmids and measurement of beta-galactosidase assays
Yeast matchmaker two-hybrid system (Clontech)was used to determine interactions between FtsZ andFtsW proteins. The full length and truncated formsof ftsZTB and ftsWTB were cloned downstream of yeastGAL4 activating (pGAD10) and GAL4 DNA binding(pAS2-1) domains following manufacturer�s instruc-tions (see Table 1). Details of truncations in ftsZ andftsW are described in Table 1. These plasmids invarious combinations were introduced into yeast Y19strain and the resultant transformants were analyzedqualitatively by Colony-lift Filter Assay and quantita-tively by Liquid Culture Assay essentially following theYeast Protocols Handbook PT3024-1 (Clontech).Briefly, for liquid culture assay, overnight cultureswere diluted, and grown fresh at 30 �C to OD600 of0.5 to 0.6. Cells were washed, resuspended in Z-buffer(60 mM Na2HPO4, 40 mM NaH2PO4,10 mM MgCl2,50 mM b-mercaptoethanol), permeabilized by freeze-thawing, incubated with 5-bromo-4-chloro-3-indolyl-b-D-galactoside (1 mg/ml) for 1–2 h at 30 �C. Aftersufficient yellow color developed, reactions werestopped, absorbance recorded at 435 nM and Millerunits were calculated as described (PT3024-1). Positive(pVA3-1, pTD1) and negative (pLAM5-1) controlplasmids (see Table 1) were always included in theseassays. The background b-galactosidase activity ofyeast reporter strains transformed with individualplasmids expressing respective fts genes were alsomeasured and were found to be comparable to thatproduced by the untransformed reporter strain.
2.3. Construction of plasmids expressing ftsZ
Self-replicating plasmids expressing M. smegmatis
ftsZ (ftsZsmeg, pJFR78) and ftsZsmeg–gfp (pJFR79) arepreviously described [12]. With pJFR79 as template,overlay PCR mutagenesis protocol was used to generatemutant ftsZsmeg–gfp containing alanine in place ofaspartic acid at positions 374–376 (designated asftsZ*–gfp). Primers 5 0-GACGGGGCATCGCGGCGG-
CCGCGGTCGACGTGCCGCCC-3 0 (MVM362) and50-GGGCGGCACGTCGACCGCGGCCGCCGCGAT-GC-CCCCGTC-3 0 (MVM363) were used in combina-tion with 5 0-GCTCTAGATTTGTATAGTTCATCC-3 0
(MVM189) that binds to the 3 0-end of gfp and 5 0-CA-GCCATATGACCCCCCCGCATAAC-3 0 (MVM144)that binds to the 5 0-end of ftsZ coding region, respec-tively, to generate 1.1 and 0.7 kb PCR products. Muta-tion generating sequences are shown in bold andrestriction endonuclease recognition sequences incorpo-rated for cloning are underlined. Overlay PCR reactionwas carried out using the two PCR products from theprevious step as templates along with primers

Table 1Plasmids used
Name Description Reference
pJAM2 E. coli–Mycobacterium shuttle vector, Kmr [19]pMV306H Mycobacterial integrating vector, Hygr Med-Immune Inc.pET15b E. coli expression vector, Apr Novagen Inc.pEYFP E. coli vector with EYFP coding region, Apr ClontechpECFP E. coli vector with ECFP coding region, Apr ClontechpGAD10 Yeast 2 hybrid vector with GAL4 activating domain ClontechpAS2-1 Yeast 2 hybrid vector with GAL4 DNA binding domain ClontechpVA3 Murine p53–GAL4 DNA-BD fusion in pAS2-1 ClontechpTD1 SV40-large T-antigen–GAL4 AD fusion in pGAD10 ClontechpLAM50-1 Human Lamin C-DNA-BD fusion in pAS2-1 ClontechpJfr78 amip–ftsZsmegin pMV306H, Hygr [12]pJfr79 ftsZsmeg–gfp in pJAM2, Kmr [12]pEM18 ftsZ*–gfp (ftsZD374–376Asmeg–gfp) in pJAM2, Kmr This studypEM21 amip–ftsZ* (ftsZD374–376Asmeg) in pMV306H, Hygr This studypLR21 cfp–ftsWsmeg in pJAM2, Kmr This studypRR13 amip–ftsZsmeg-yfp in pMV306H, Hygr This studypLR12 ftsZ* (ftsZD374–376Asmeg) in pET15b, Apr This studypAS2-1-ftsZ ftsZTB cloned in NcoI–BamHI sites of pAS2-1 This studypGAD10-ftsZ ftsZTB cloned in BamHI site of pGAD10 This studypAS2-1-ftsZtr Truncated ftsZTB, lacking last 54 nucleotides, cloned in
NcoI–BamHI sites of pAS2-1This study
pAS2-1–30ftsW 3 0 end of ftsWTB (nucleotides 1233 to 1572, encoding thesoluble cytoplasmic C-terminal end of FtsW) cloned inNcoI–BamHI sites of pAS2-1
This study
pGAD10-30ftsW 3 0 end of ftsWTB (nucleotides 1233 to 1572) cloned in BamHI–EcoRI sites of pGAD10 This study
M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17 11
MVM189 and MVM144 to generate a full length ftsZ*–gfp. Similarly, MVM144 and MV240 (5 0-GCTCT-AGAGTGCCGCATGAAGGGCGGC; binds to 3 0 theend of ftsZ) were used as primers to generate ftsZ*smeg
and (referred to as ftsZ* from here on). All PCR prod-ucts were confirmed by sequencing. The ftsZ* andftsZ*–gfp were cloned downstream of amidasepromoter (amip), respectively, in pMV306 and inpJAM2 to create pEM21 and pEM18, and introducedinto M. smegmatis (Table 1). For some experiments,gene for enhanced yellow fluorescent protein (eyfp)was PCR amplified from pEYFP and cloned down-stream of ftsZ to create pRR13 (Table 1). The ftsZ* cod-ing region was also cloned downstream of T7 promoterin pET15b resulting in pLR12 (Table 1).
2.4. Construction of M. smegmatis ecfp–ftsW
Blast search of the TIGR unfinished genome website(www.tigr.org) with the M. tuberculosis ftsW sequenceidentified the M. smegmatis ftsW ortholog. Oligonucleo-tide primer pair MVM439 (5 0-TCGCTAGATTATCA-CCCGTAACGCTGACCTTCCAAT-30) and MVM456(5 0-TCGCTCTAGAGTGGGCAGCATCCTCACCC-GGCTGC-3 0) was used to amplify the ftsWcoding regionfrom M. smegmatis genomic DNA, confirmed bysequencing and fused with the 3 0 end of the gene forenhanced cyan fluorescent protein (pECFP) expressingfrom amip to create pLR21.
2.5. Recombination experiments
A two-step homologous recombination strategy asdescribed by Parish and Stoker [16] was used to disruptM. smegmatis ftsZ at its native locus in the presence ofan integrated copy of either wild-type [12] or mutantftsZ. Briefly, in the first step, suicidal (non-replicative)plasmid vectors containing positive (lacZ gene for bluecolor, aph gene for kanamycin resistance) and negativecounterselectable marker (sacB for sucrose sensitivity)were used to construct ftsZ single crossover strain FZ1[12]. This strain is resistant to kanamycin, sensitive tosucrose and produces blue color on indicator plates.Plasmid construct expressing ftsZ* (pEM21) was inte-grated at mycobacteriophage attachment site in FZ1.In the next step, the resulting merodiploid SCO strainwas plated for double crossovers (DCO) that are sensi-tive to kanamycin, white in color and resistant to su-crose. Analysis of DCOs was done by PCR andSouthern hybridization as described [12].
2.6. Immunoblotting
Preparation of mycobacterial cell lysates, detection ofFtsZ by Western blotting using affinity-purified anti-M. tuberculosis FtsZ antibodies were essentially asdescribed previously [12]. Western blots were processedusing the Amersham Pharmacia ECF chemifluorescencekit and protocol, and FtsZ bands were visualized by

12 M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17
scanning the nitrocellulose blots in a Bio-Rad MolecularImager and quantified using the Quantity One software.
2.7. Polymerization assays
The M. tuberculosis ftsZ and M. smegmatis ftsZ*coding regions were expressed from T7 promoter inpET15b (pSAR1 and pLR12) and the his–tag fusionproteins were purified on two successive nickel affinitycolumns. Based on SYPRO-RUBY staining the purifiedproteins were judged to be approximately 92% pure (notshown). The purified proteins were stored in 25 mMHEPES buffer pH 7.2 containing 0.1 mM EDTA, 10%glycerol and 1 mM DTT. FtsZ polymerization wasexamined by sedimentation assay [17]. Polymerizationreactions were carried out in a 50 ll volume containing50 mMMES, pH 6.5; 5 mM MgCl2; 100 mM KCl; with-out or with 1mM GTP and 10 lM FtsZTB or FtsZ*(FtsZD374–376A). The reactions were initiated by theaddition of GTP and incubated for 10 min at 30 �C.Polymerized FtsZ was separated by centrifugation at80,000 rpm in a TLA100.1 rotor in a Beckman TL100ultracentrifuge. Pellet and supernatant fractions wereseparated on SDS–PAGE and visualized followingstaining with Coomassie brilliant blue.
2.8. Microscopy and visualization of FtsZ and FtsW
septal structures
Overnight cultures of M. smegmatis merodiploidsexpressing fluorescent fusions of wild type and mutantforms of FtsZ protein (ftsZsmeg–gfp, ftsZ*–gfp, ftsZsmeg–eyfp), and wild FtsW protein (ecfp–ftsWsmeg) werediluted in 7H9 broth and grown without or with 0.2%acetamide for 5 h. Bacteria were then examined by fluo-rescence microscopy using either a Nikon Eclipse 600microscope (with a 100· Nikon Plan fluor oil immersionobjective with a numerical aperture of 1.4; standardFITC filter set from Chroma) or a Perkin Elmer Ultra-view LCI Confocal Imaging system attached to a NikonTE2000 inverted microscope (100· DIC objective withna of 1.4; Helium–Cadmium and Argon laser for excit-ing CFP and YFP, respectively). Images were acquiredusing either a Photometrics Coolsnap ES camera andMetapmorph 6.2 imaging software (Universal ImagingCorporation) or a Hamamatsu digital camera andImaging Suite 5.2 software. All images were optimizedusing Adobe Photoshop 7.0.
3. Results and discussion
3.1. FtsZ–FtsW interactions in a yeast two-hybrid system
Yeast matchmaker two-hybrid system (Clontech) wasused to detect interactions between FtsZ and FtsW
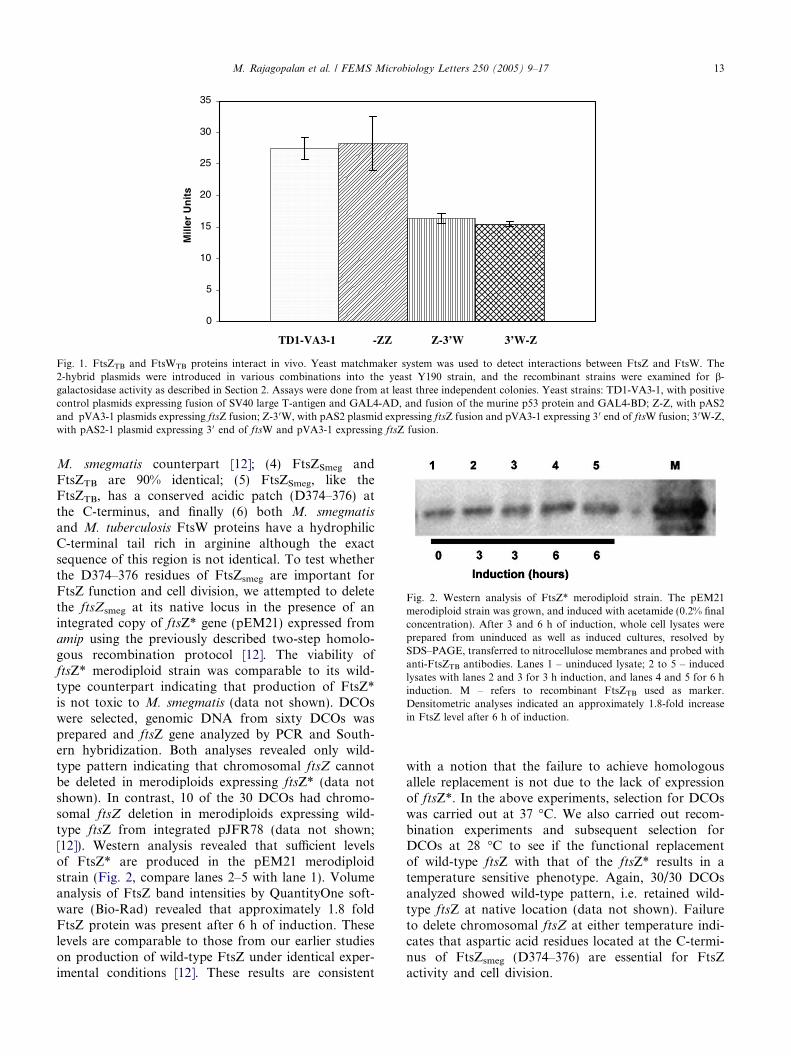
proteins. The protocol relies on the principle that posi-tive interactions between the two fusion proteins wouldresult in the production of intact b-galactosidase proteinwhich can then cleave the substrate 5-bromo-4-chloro-3-indolyl-b-D-galactoside to produce blue color. As can beseen, yeast transformants producing FtsZ from GAL4activating and DNA binding domains showed b-galac-tosidase activity comparable to that of the positive con-trols producing the well-established protein-interactionpartners, namely murine P53 protein (pVA3) andSV40 T-antigen (pTD1) (Fig. 1). These results areconsistent with the findings that FtsZTB protein exhibitsoligomerization activity in vitro [18] and polymerizationactivity in vivo [11]. Yeast transformants producingintact FtsZTB and the soluble cytoplasmic C-terminalFtsW fragment (3 0W) from either combination ofexpression vectors showed b-galactosidase activityindicating that the C-terminal end of FtsW is sufficientfor interaction with FtsZ (Fig. 1). In contrast, yeasttransformants producing heterologous human LamininC protein (pLAM5 0-1) along with either FtsZ(pGAD10-FtsZ) or C-terminal FtsW (pGAD10-3 0W)did not show any measurable b-galactosidase activity(data not shown). Similarly, yeast transformants pro-ducing a truncated FtsZ lacking the C-terminal domainand 3 0W also did not show b-galactosidase activity (datanot shown). Together, these results indicate that FtsZ–FtsW proteins interact in vivo, and that the C-terminaldomains of FtsZ and FtsW proteins are important forthese interactions.
3.2. Aspartic acid residues located at the C-terminal end
of FtsZ are essential for cell division and viability
Biochemical assays showed that a stretch of fouraspartic acid residues located at the C-terminal endof FtsZ (D364–367) are important for interactionsbetween FtsZ and FtsW [15]. Interactions betweenthe two proteins are completely abolished when 3 ofthe 4 aspartic acid residues were replaced with alanine(D364–366) [15]. If the presumed interactions betweenthe FtsZ and FtsW proteins are biologically relevantand are essential, then one would expect that FtsZprotein containing alanine in place of aspartic acid atthese locations, FtsZ*, would be defective for interac-tions with FtsW, hence would not be proficient forsubsequent cell division. To test this hypothesis weattempted to create a strain expressing ftsZ* as thesole source for FtsZ. These experiments were carriedout in M. smegmatis for the following reasons. (1)We showed earlier that M. tuberculosis merodiploidsexpressing ftsZ from heterologous promoters arenonviable [11]; (2) ftsZsmeg is essential for viabilityand can only be deleted in the presence of anothercopy of ftsZ integrated elsewhere on the chromosome[12]; (3) ftsZtb can replace the function of the
0
5
10
15
20
25
30
35
Mill
er U
nit
s
TD1-VA3-1 -ZZ Z-3’W 3’W-Z
Fig. 1. FtsZTB and FtsWTB proteins interact in vivo. Yeast matchmaker system was used to detect interactions between FtsZ and FtsW. The2-hybrid plasmids were introduced in various combinations into the yeast Y190 strain, and the recombinant strains were examined for b-galactosidase activity as described in Section 2. Assays were done from at least three independent colonies. Yeast strains: TD1-VA3-1, with positivecontrol plasmids expressing fusion of SV40 large T-antigen and GAL4-AD, and fusion of the murine p53 protein and GAL4-BD; Z-Z, with pAS2and pVA3-1 plasmids expressing ftsZ fusion; Z-30W, with pAS2 plasmid expressing ftsZ fusion and pVA3-1 expressing 3 0 end of ftsW fusion; 3 0W-Z,with pAS2-1 plasmid expressing 3 0 end of ftsW and pVA3-1 expressing ftsZ fusion.
Fig. 2. Western analysis of FtsZ* merodiploid strain. The pEM21merodiploid strain was grown, and induced with acetamide (0.2% finalconcentration). After 3 and 6 h of induction, whole cell lysates wereprepared from uninduced as well as induced cultures, resolved bySDS–PAGE, transferred to nitrocellulose membranes and probed withanti-FtsZTB antibodies. Lanes 1 – uninduced lysate; 2 to 5 – inducedlysates with lanes 2 and 3 for 3 h induction, and lanes 4 and 5 for 6 hinduction. M – refers to recombinant FtsZTB used as marker.Densitometric analyses indicated an approximately 1.8-fold increasein FtsZ level after 6 h of induction.
M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17 13
M. smegmatis counterpart [12]; (4) FtsZSmeg andFtsZTB are 90% identical; (5) FtsZSmeg, like theFtsZTB, has a conserved acidic patch (D374–376) atthe C-terminus, and finally (6) both M. smegmatis
and M. tuberculosis FtsW proteins have a hydrophilicC-terminal tail rich in arginine although the exactsequence of this region is not identical. To test whetherthe D374–376 residues of FtsZsmeg are important forFtsZ function and cell division, we attempted to deletethe ftsZsmeg at its native locus in the presence of anintegrated copy of ftsZ* gene (pEM21) expressed fromamip using the previously described two-step homolo-gous recombination protocol [12]. The viability offtsZ* merodiploid strain was comparable to its wild-type counterpart indicating that production of FtsZ*is not toxic to M. smegmatis (data not shown). DCOswere selected, genomic DNA from sixty DCOs wasprepared and ftsZ gene analyzed by PCR and South-ern hybridization. Both analyses revealed only wild-type pattern indicating that chromosomal ftsZ cannotbe deleted in merodiploids expressing ftsZ* (data notshown). In contrast, 10 of the 30 DCOs had chromo-somal ftsZ deletion in merodiploids expressing wild-type ftsZ from integrated pJFR78 (data not shown;[12]). Western analysis revealed that sufficient levelsof FtsZ* are produced in the pEM21 merodiploidstrain (Fig. 2, compare lanes 2–5 with lane 1). Volumeanalysis of FtsZ band intensities by QuantityOne soft-ware (Bio-Rad) revealed that approximately 1.8 foldFtsZ protein was present after 6 h of induction. Theselevels are comparable to those from our earlier studieson production of wild-type FtsZ under identical exper-imental conditions [12]. These results are consistent
with a notion that the failure to achieve homologousallele replacement is not due to the lack of expressionof ftsZ*. In the above experiments, selection for DCOswas carried out at 37 �C. We also carried out recom-bination experiments and subsequent selection forDCOs at 28 �C to see if the functional replacementof wild-type ftsZ with that of the ftsZ* results in atemperature sensitive phenotype. Again, 30/30 DCOsanalyzed showed wild-type pattern, i.e. retained wild-type ftsZ at native location (data not shown). Failureto delete chromosomal ftsZ at either temperature indi-cates that aspartic acid residues located at the C-termi-nus of FtsZsmeg (D374–376) are essential for FtsZactivity and cell division.

14 M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17
3.3. M. smegmatis FtsZ* can colocalize with the
wild-type FtsZsmeg to the Z-rings
Cell division process is believed to be initiated whenFtsZ assembles to form the Z-ring at the putativemidcell location and catalyzes a cascade of events. Thus,the defect in cell division resulting in inviability could bedue to a defect either at the assembly step of the Z-ringand or at a post assembly event. To gain insights intothese issues, we constructed M. smegmatis merodiploidstrain expressing amip–ftsZ*–gfp (pEM18, Table 1)from a self-replicating vector [19] and visualized FtsZstructures by fluorescent microscopy.
Overexpression of ftsZ from amip in self-replicatingplasmids results in filamentation and lethality [11].However, we found that the amip is leaky and confirmedby immunoblotting that FtsZ–GFP fusion protein wasproduced under uninduced conditions in merodiploidsexpressing ftsZsmeg–gfp or ftsZ*–gfp (Fig. 3A). Densito-metric quantitation indicated that comparable levels ofwild-type and mutant FtsZ–GFP fusion protein wereproduced (data not shown). Next, we examined whetherthe FtsZ–GFP structures in M. smegmatis merodiploids(pJFR79) can be visualized by fluorescence microscopy
Fig. 3. FtsZ* colocalizes with wild-type FtsZsmeg to the midcell. (A) ImmunoftsZ*–gfp (lanes 1 and 2) or ftsZsmeg–gfp (lane 3) were resolved by SDS–PAGEantibodies as described under Section 2. Lane 4 is purified FtsZTB protein lomerodiploids expressing either wild-type ftsZsmeg–gfp (a, c) or ftsZ*–gfp (b,fluorescence microscopy as described under Section 2. Images were acquirimaging software (Universal Imaging Corporation). All images were optimizebrightfield images, of the same respective fields.
under uninduced conditions. Under these conditions,30–40% cells contained one distinct FtsZ-band perdividing cell (Fig. 3B: panels – a, c) and average lengthof these cells was comparable to that of the vector con-trol strain (data not shown). These results indicate that alow level production of FtsZ–GFP fusion protein doesnot interfere with the cell division process. In theseexperiments, ftsZ–gfp was expressed from a replicatingplasmid. Under the same conditions, the M. smegmatis
merodiploids producing FtsZ*–GFP (pEM18) alsoshowed similar distinct FtsZ structures in approximatelysimilar number of cells, indicating that the mutantFtsZ*–GFP fusion protein associates with the wild-typeprotein at the presumptive division site (Fig. 3B: panels –b, d). When grown in the presence of 0.2% acetamide,the ftsZ*–gfp merodiploid cells produced ribbon-likestructures, similar to those seen with the wild-type FtsZoverproduction (data not shown, see [11]). These resultsindicate that replacement of aspartic acid with alanine atpositions 374–376 of M. smegmatis FtsZ does not com-promise its ability to associate with the Z-ring. Sincecells expressing mutant ftsZ* as the only copy of ftsZare not viable, we could not address whether or notthe FtsZ* is capable of forming Z-ring like structures
blotting. Whole cell lysates fromM. smegmatis merodiploids expressing, transferred to nitrocellulose membranes and probed with anti-FtsZTB
aded for reference. (B) Localization of FtsZ structures. M. smegmatis
d) were grown to exponential phase and examined by brightfield anded using a Photometrics Coolsnap ES camera and Metapmorph 6.2d using Adobe Photoshop 7.0. Panels a, b and c, d are fluorescence and

M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17 15
in the absence of the wild-type protein. Presumably,inviability is due to either a defective localization ofFtsZ* (FtsZD374–376A) or formation of nonproductiveFtsZ-rings.
3.4. The FtsZ* has polymerization activity
FtsZTB protein catalyzes GTP-dependent polymeri-zation and hydrolysis activities [18]. To test whetherFtsZ* is competent for polymerization, sedimentationassay was used [17]. Polymerization was initiated withthe addition of GTP and the FtsZ polymers pelleted fol-lowing high-speed centrifugation. While no polymeriza-tion was seen in the absence of GTP, FtsZ* polymerizedin the presence of GTP, like the wild-type FtsZTB
protein (Fig. 4). Presumably, the GTP dependent poly-merization activity detected in vitro does not translateto the formation of productive Z-rings in vivo.
3.5. FtsZsmeg and FtsWsmeg colocalize to the Z-ring
The above data show that the aspartic acid residuesat the C-terminus of FtsZ are important for FtsZ func-tion and cell division. The observed interactions betweenFtsZ and FtsW imply that FtsW could be a part of theseptasomal complex of M. smegmatis, and possiblycolocalize with FtsZ to the midcell site. Therefore, weexamined M. smegmatis merodiploid cells producingECFP–FtsWsmeg fusion protein by fluorescence micros-copy. The ECFP–FtsWsmeg could not be visualizedunder uninduced conditions (data not shown) whereasgrowth in the presence of acetamide for 5 h led to amoderate increase in cell length (Fig. 5, compare a andb), and enabled visualization of distinct ECFP–Fts-Wsmeg structures as bands (see Fig. 5c), similar to thestructures formed with FtsZ-enhanced yellow fluores-cent fusion protein (FtsZ–EYFP) (Fig. 5d). The filamen-tous cells were somewhat curvy and only 5% of them
Fig. 4. FtsZ* is polymerization proficient. The ftsZTB and ftsZ*(ftsZ*smeg) coding regions were expressed from pET15b, the his–tagfusion proteins were purified on two successive nickel affinity columnsand their polymerization activity was examined by sedimentation assay[17]. The polymerized FtsZ was separated by centrifugation, the pelletand supernatant fractions were run on SDS–PAGE and visualizedfollowing staining with Coomassie brilliant blue. The FtsZsmeg proteinruns slightly slower than the FtsZTB protein on SDS–PAGE [11]. S –supernatant; P – pellet. Purified FtsZTB and FtsZ* proteins were alsoloaded on the same gel for reference.
had distinct ECFP–FtsW structures, majority of whichwere at the midcell sites (Table 2). Merodiploids produc-ing GFP–FtsW fusion also had similar number of septalrings (data not shown) indicating that the lowabundance is not due to the type of fluorescent fusionprotein used. In contrast, under these conditions,20–22% of cells overproducing FtsZ had distinct FtsZ-bands and majority of which were at the midcelllocation (Table 2). In the above merodiploid strains,the FtsZ–EYFP (pRR13) was produced from an inte-grated plasmid whereas the ECFP–FtsW (pLR21) wasfrom a replicating plasmid. It is pertinent to note thatlate recruits to the division site in E. coli show reducedlocalization frequency as compared to FtsZ [20,21].The curvy nature of the filamentous cells made precisedetermination of the location of septal FtsZ–EYFPand ECFP–FtsW bands difficult and therefore only rel-atively straight filaments were scored for these determi-nations. The increase in cell length associated with FtsWoverproduction was moderate as compared to FtsZoverproduction under the same conditions (data notshown). Some FtsW filamentous cells also showed in-creased background fluorescence and few weak intensefluorescence spots. Presumably, these are nonproductivestructures associated with fusion protein overproductionand or accumulation of proteolytic breakdown products[21,22]. We lack antibodies to FtsW and consequentlycould not measure FtsW levels relative to FtsZ protein.Together, these results indicate that more than normallevels of FtsW interfere with the cell division process[23], and that FtsW is also a member of M. smegmatis
septasomal complex.Since both FtsZ and FtsW proteins of M. smegmatis
are associated with distinct structures, we next examinedwhether FtsW localizes to the same site as the FtsZ. Toaddress this question, we transformed merodiploidstrain expressing ftsZ–eyfp with pLR21 plasmid express-ing ecfp–ftsW. ECFP and EYFP are mutant GFP pro-teins with different absorption and emission spectraand do not show any fluorescence crossover whenappropriate fluorescence channels were used (data notshown; [21]). The M. smegmatis merodiploid strainexpressing both ftsZsmeg–eyfp and ecfp–ftsWsmeg wasgrown in the presence of 0.2% acetamide for 5 h andvisualized by fluorescence microscopy. We first observedthe localization of FtsZsmeg–EYFP to the presumptivemidcell site in the EYFP channel (Fig. 5f). The samefield of focus when examined in the ECFP channel alsoshowed distinct ECFP–FtsWsmeg bands that colocalizedto the EYFP sites (Fig. 5g). It is noteworthy that not allcells that showed distinct FtsZ–EYFPsmeg bands showedthe corresponding ECFP–FtsWsmegfluorescence (Fig. 5,compare f and g; Table 2). Approximately 20% cellshad septal Z-rings and 27% of these are associated withFtsW (see Table 2). On the other hand, 93% of FtsWrings showed FtsZ localization. Our results showing

Fig. 5. FtsZsmeg and FtsWsmeg colocalize to the putative septal sites. M. smegmatis merodiploids expressing either ftsZsmeg–eyfp (d) and ecfp–ftsWsmeg (a–c) or both (e–g) were grown to exponential phase in the 7H9-AD broth without (a) or with 0.2% acetamide (b–g). Bacteria were removed5 h after acetamide addition and examined by fluorescence microscopy using the Perkin Elmer Ultraview LCI Confocal Imaging system attached to aNikon TE2000 inverted microscope equipped with a 100· DIC objective with na of 1.4. Helium–Cadmium and Argon laser were used for excitingCFP and YFP, respectively. Images were captured with a Hamamatsu digital camera using the Imaging Suite 5.2 software. Panels a, b and e are DICimages and panels c, d, f and g are fluorescence images. All images were optimized using Adobe Photoshop 7.0. Arrowheads and arrows indicateECFP–FtsWsmeg and FtsZsmeg–EYFP bands, respectively.
Table 2Number of FtsZ and FtsW rings
Strain No. of cells No. of cells with FtsZ-rings No. of cells with FtsW-rings Septal ring location
Midcell Othersa
mC2-RR13 207 46 (22) – 34 (74) 12 (26)mC2-LR21 340 – 17 (5) 15 (88) 2 (12)mC2-RR13-LR21 262 52 (20) 15b 47 (90) 5 (10)
mC2-RR13:M. smegmatis merodiploids with integrated pRR13.mC2-LR21: M. smegmatis merodiploids with replicating pLR21 plasmid.mC2-RR13-LR21: M. smegmatis merodiploids with pRR13 and pLR21.Parentheses: Percent cells.a Cells with either 1 or 2 septal rings in quarter position or 1 ring in quarter and 1 ring in midcell location.b 14/15 (93%) show FtsZ–FtsW colocalization.
16 M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17
fewer septal FtsW rings as compared to FtsZ rings inM. smegmatis merodiploid strain are consistent with anotion that FtsWsmeg could be a late recruit to the divi-sion site, and that mycobacterial FtsZ could localize tothe division site independent of FtsW. The FtsZ-fluores-cent fusions are more easily detected as compared to thefluorescent protein fusions to other Fts proteins [21].The differences in protein abundance, and to someextent the different planes of focus for ECFP and EYFPchannels in the particular images may affect the numberof FtsZ and FtsW rings observed [21]. Alternatively, theYFP fusion at the C-terminus of FtsZ may sterically
interfere with the interactions with CFP–FtsW fusionresulting in fewer ECFP–FtsW rings.
4. Conclusions
The key findings of this study are that the FtsW pro-tein interacts with FtsZ in vivo; the FtsW protein is amember of mycobacterial septasome that is organizedinto ring like structures [11], and colocalizes with theFtsZ-ring in the filamentous cells. In addition, our resultssuggest that the observed FtsZ and FtsW interactions are

M. Rajagopalan et al. / FEMS Microbiology Letters 250 (2005) 9–17 17
biologically relevant because FtsZ mutant protein that isshown to be defective for interactions with FtsW in vitro,is not proficient for cell division and viability. The myco-bacterial genome lacks any identifiable analogs of ZipAand FtsA like proteins, and since FtsW contains mem-brane spanning domains [14] and associates with theFtsZ-ring structures at the midcell (Fig. 5 compare fand g), one hypothesis is that FtsW could help FtsZlocalize to the midcell site [15]. However, our FtsZ andFtsW colocalization experiments showing that only27% of FtsZ rings are associated with FtsW argueagainst the possibility that FtsW helps target FtsZ tothe putative midcell site in mycobacteria. Our results sug-gest that FtsW could be a late recruit to the division siteinM. smegmatis, and that it possibly localizes after FtsZ.These results are consistent with the situation seen in E.
coli [20]. This raises the question as to how FtsZ is local-ized to the putative midcell locations in mycobacteria.Clearly detailed studies are required to evaluate whetherFtsZ can assemble at midcell site in the absence of FtsWand vice versa. The FtsW protein of E. coli is implicatedat the initiation stage of Z-ring assembly and also at laterstages of cytokinesis [24,25]. It conceivable that the func-tion of mycobacterial FtsW is pleiotropic in that it couldpromote the targeting of FtsZ protein to the predivi-sional site and subsequently be involved in the stabiliza-tion of FtsZ rings and or for other aspects of cytokinesisfollowing the assembly of Z-rings. Finally, our colocal-ization experiments showing only a limited number ofFtsZ rings associatedwithFtsW, suggest a possibility thathitherto unidentified interactions of FtsZ with other pro-teins involved in cytokinesis are lost in ftsZ* background.
Acknowledgements
This work was supported from grants AI48417 (MR),AI41406 (MM) and KBN contract 3P04C06825 (J.D.and M.P.). We thank Dr. Zafer Hatahet for fruitful dis-cussions and Ms. A. Rosillo for technical assistance.
References
[1] Romberg, L. and Levin, P.A. (2003) Assembly dynamics of thebacterial cell division protein FTSZ: poised at the edge ofstability. Annu. Rev. Microbiol. 57, 125–154.
[2] Bi, E., Dai, K., Subbarao, S., Beall, B. and Lutkenhaus, J. (1991)FtsZ and cell division. Res. Microbiol. 142, 249–252.
[3] Hale, C.A. and de Boer, P.A. (1999) Recruitment of ZipA to theseptal ring of Escherichia coli is dependent on FtsZ andindependent of FtsA. J. Bacteriol. 181, 167–176.
[4] Mukherjee, A. and Lutkenhaus, J. (1999) Analysis of FtsZassembly by light scattering and determination of the role ofdivalent metal cations. J. Bacteriol. 181, 823–832.
[5] Pichoff, S. and Lutkenhaus, J. (2002) Unique and overlappingroles for ZipA and FtsA in septal ring assembly in Escherichia
coli. EMBO J. 21, 685–693.
[6] Geissler, B., Elraheb, D. and Margolin, W. (2003) A gain-of-function mutation in ftsA bypasses the requirement for theessential cell division gene zipA in Escherichia coli. Proc. Natl.Acad. Sci. USA 100, 4197–4202.
[7] Pichoff, S. and Lutkenhaus, J. (2005) Tethering the Z ring to themembrane through a conserved membrane targeting sequence inFtsA. Mol. Microbiol. 55, 1722–1734.
[8] Di Lallo, G., Fagioli, M., Barionovi, D., Ghelardini, P. andPaolozzi, L. (2003) Use of a two-hybrid assay to study theassembly of a complex multicomponent protein machinery:bacterial septosome differentiation. Microbiology 149, 3353–3359.
[9] Anilkumar, G., Srinivasan, R. and Ajitkumar, P. (2004) A C-terminal deletion mutant of Mycobacterium tuberculosis FtsZ
shows fast polymerization in vitro. Microbiology 150, 2629–2639.[10] Anilkumar, G., Srinivasan, R., Anand, S.P. and Ajitkumar, P.
(2001) Bacterial cell division protein FtsZ is a specific substratefor the AAA family protease FtsH. Microbiology 147, 516–517.
[11] Dziadek, J., Madiraju, M.V., Rutherford, S.A., Atkinson, M.A.and Rajagopalan, M. (2002) Physiological consequences associ-ated with overproduction of Mycobacterium tuberculosis FtsZ inmycobacterial hosts. Microbiology 148, 961–971.
[12] Dziadek, J., Rutherford, S.A., Madiraju, M.V., Atkinson, M.A.and Rajagopalan, M. (2003) Conditional expression of Mycobac-
terium smegmatis ftsZ, an essential cell division gene. Microbiol-ogy 149, 1593–1603.
[13] Gomez, J.E. and Bishai, W.R. (2000) whmD is an essentialmycobacterial gene required for proper septation and celldivision. Proc. Natl. Acad. Sci. USA 97, 8554–8559.
[14] Cole, S.T. et al. (1998) Deciphering the biology of Mycobacte-
rium tuberculosis from the complete genome sequence. Nature393, 537–544.
[15] Datta, P., Dasgupta, A., Bhakta, S. and Basu, J. (2002)Interaction between FtsZ and FtsW of Mycobacterium tubercu-
losis. J. Biol. Chem. 277, 24983–24987.[16] Parish, T. and Stoker, N.G. (2000) Use of a flexible cassette
method to generate a double unmarked Mycobacterium tubercu-
losis tlyA plcABC mutant by gene replacement. Microbiology146, 1969–1975.
[17] Mukherjee, A. and Lutkenhaus, J. (1998) Purification, assembly,and localization of FtsZ. Methods Enzymol. 298, 296–305.
[18] White, E.L., Ross, L.J., Reynolds, R.C., Seitz, L.E., Moore, G.D.and Borhani, D.W. (2000) Slow polymerization ofMycobacterium
tuberculosis FtsZ. J. Bacteriol. 182, 4028–4034.[19] Triccas, J.A., Parish, T., Britton, W.J. and Gicquel, B. (1998) An
inducible expression system permitting the efficient purification ofa recombinant antigen from Mycobacterium smegmatis. FEMSMicrobiol. Lett. 167, 151–156.
[20] Mercer, K.L. and Weiss, D.S. (2002) The Escherichia coli celldivision protein FtsW is required to recruit its cognate transpep-tidase, FtsI (PBP3), to the division site. J. Bacteriol. 184, 904–912.
[21] Chen, J.C. and Beckwith, J. (2001) FtsQ, FtsL and FtsI requireFtsK, but not FtsN, for co-localization with FtsZ duringEscherichia coli cell division. Mol. Microbiol. 42, 395–413.
[22] Ma, X., Ehrhardt, D.W. and Margolin, W. (1996) Colocalizationof cell division proteins FtsZ and FtsA to cytoskeletal structuresin living Escherichia coli cells by using green fluorescent protein.Proc. Natl. Acad. Sci. USA 93, 12998–13003.
[23] Khattar, M.M., Begg, K.J. and Donachie, W.D. (1994) Identifi-cation of FtsW and characterization of a new ftsW divisionmutant of Escherichia coli. J. Bacteriol. 176, 7140–7147.
[24] Wang, L., Khattar, M.K., Donachie, W.D. and Lutkenhaus, J.(1998) FtsI and FtsW are localized to the septum in Escherichia
coli. J. Bacteriol. 180, 2810–2816.[25] Khattar, M.M., Addinall, S.G., Stedul, K.H., Boyle, D.S.,
Lutkenhaus, J. and Donachie, W.D. (1997) Two polypeptideproducts of the Escherichia coli cell division gene ftsW and apossible role for FtsW in FtsZ function. J. Bacteriol. 179, 784–793.
Related Documents











