RESEARCH ARTICLE Genetic diversity of Mimosa pudica rhizobial symbionts in soils of French Guiana: investigating the origin and diversity of Burkholderia phymatum and other beta-rhizobia Ravi P.N. Mishra 1,5 , Pierre Tisseyre 1 , Re ´ my Melkonian 1 , Cle ´ mence Chaintreuil 1 , Lucie Miche ´ 1 , Agnieszka Klonowska 1 , Sophie Gonzalez 2 , Gilles Bena 1,4 , Gise ` le Laguerre 3 & Lionel Moulin 1 1 IRD, UMR LSTM, Montpellier, France; 2 IRD, Herbier de Guyane, Cayenne, French Guiana; 3 INRA, UMR LSTM, Montpellier, France; and 4 Laboratoire de Microbiologie et de Biologie Mole ´ culaire, University Mohammed V Agdal, Agdal, Morocco Correspondence: Lionel Moulin, Laboratoire des Symbioses Tropicales et Me ´ diterrane ´ ennes, TA-A82/J Campus de Baillarguet 34398 Montpellier Cedex 5, France. Tel.: +33 4 67 59 37 63; fax: +33 4 67 59 38 02; e-mail: [email protected] Present address: Ravi P.N. Mishra, Novartis Vaccines Research Centre, Via Fiorentina 1, Siena, 53100, Italy. Received 25 July 2011; revised 14 October 2011; accepted 19 October 2011. DOI: 10.1111/j.1574-6941.2011.01235.x Editor: Philippe Lemanceau Keywords rhizobia; symbiosis; Mimosa; Burkholderia; Cupriavidus; biodiversity. Abstract The genetic diversity of 221 Mimosa pudica bacterial symbionts trapped from eight soils from diverse environments in French Guiana was assessed by 16S rRNA PCR-RFLP, REP-PCR fingerprints, as well as by phylogenies of their 16S rRNA and recA housekeeping genes, and by their nifH, nodA and nodC symbi- otic genes. Interestingly, we found a large diversity of beta-rhizobia, with Burk- holderia phymatum and Burkholderia tuberum being the most frequent and diverse symbiotic species. Other species were also found, such as Burkholderia mimosarum, an unnamed Burkholderia species and, for the first time in South America, Cupriavidus taiwanensis. The sampling site had a strong influence on the diversity of the symbionts sampled, and the specific distributions of symbi- otic populations between the soils were related to soil composition in some cases. Some alpha-rhizobial strains taxonomically close to Rhizobium endophyti- cum were also trapped in one soil, and these carried two copies of the nodA gene, a feature not previously reported. Phylogenies of nodA, nodC and nifH genes showed a monophyly of symbiotic genes for beta-rhizobia isolated from Mimosa spp., indicative of a long history of interaction between beta-rhizobia and Mimosa species. Based on their symbiotic gene phylogenies and legume hosts, B. tuberum was shown to contain two large biovars: one specific to the mimosoid genus Mimosa and one to South African papilionoid legumes. Introduction Rhizobia are a functional class of soil bacteria able to develop a nitrogen-fixing symbiosis with legumes, and are also termed legume nodulating bacteria (or LNB). The legume nodulation ability is spread among the Alpha- and Beta- subclasses of Proteobacteria, and the names alpha- and beta-rhizobia have been proposed for conve- nience (Gyaneshwar et al., 2011). Beta-rhizobia were originally described in 2001 in two parallel studies: one describing two Burkholderia strains (STM678 and STM815) isolated from Aspalathus carnosa (Papilionoideae) in South Africa and Machaerium luna- tum (Papilionoideae) in French Guiana, respectively (Moulin et al., 2001), which were subsequently named Burkholderia tuberum and Burkholderia phymatum (Vandamme et al., 2002); and one describing Ralstonia taiwanensis from two Mimosa (Mimosoideae) species in Taiwan (Chen et al., 2001), which was later renamed Cu- priavidus taiwanensis (Vandamme & Coenye, 2004). Since then, several diversity studies have demonstrated the widespread occurrence of beta rhizobia as symbionts of Mimosa species in Costa Rica and Texas (Barrett & Parker, 2006; Andam et al., 2007), Panama (Barrett & Parker, 2005; Parker, 2008), Brazil and Venezuela (Chen et al., 2005b; Bontemps et al., 2010), Taiwan (Chen et al., 2005a), India (Verma et al., 2004), Papua New Guinea (Elliott et al., 2009), Australia (Parker et al., 2007), and China (Liu et al., 2011). To date, several nodulating beta-rhizobia species have been described, most of them belonging to the Burkholde- ria genus. The B. tuberum type strain (STM678 T ) is able FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY ECOLOGY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R E S EA RCH AR T I C L E
Genetic diversity of Mimosa pudica rhizobial symbionts in soils
of French Guiana: investigating the origin and diversity of
Burkholderia phymatum and other beta-rhizobia
Ravi P.N. Mishra1,5, Pierre Tisseyre1, Remy Melkonian1, Clemence Chaintreuil1, Lucie Miche1,Agnieszka Klonowska1, Sophie Gonzalez2, Gilles Bena1,4, Gisele Laguerre3 & Lionel Moulin1
1IRD, UMR LSTM, Montpellier, France; 2IRD, Herbier de Guyane, Cayenne, French Guiana; 3INRA, UMR LSTM, Montpellier, France; and4Laboratoire de Microbiologie et de Biologie Moleculaire, University Mohammed V Agdal, Agdal, Morocco
Correspondence: Lionel Moulin, Laboratoire
des Symbioses Tropicales et
Mediterraneennes, TA-A82/J Campus
de Baillarguet 34398 Montpellier Cedex 5,
France. Tel.: +33 4 67 59 37 63;
fax: +33 4 67 59 38 02; e-mail:
Present address: Ravi P.N. Mishra, Novartis
Vaccines Research Centre, Via Fiorentina 1,
Siena, 53100, Italy.
Received 25 July 2011; revised 14 October
2011; accepted 19 October 2011.
DOI: 10.1111/j.1574-6941.2011.01235.x
Editor: Philippe Lemanceau
Keywords
rhizobia; symbiosis; Mimosa; Burkholderia;
Cupriavidus; biodiversity.
Abstract
The genetic diversity of 221 Mimosa pudica bacterial symbionts trapped from
eight soils from diverse environments in French Guiana was assessed by 16S
rRNA PCR-RFLP, REP-PCR fingerprints, as well as by phylogenies of their 16S
rRNA and recA housekeeping genes, and by their nifH, nodA and nodC symbi-
otic genes. Interestingly, we found a large diversity of beta-rhizobia, with Burk-
holderia phymatum and Burkholderia tuberum being the most frequent and
diverse symbiotic species. Other species were also found, such as Burkholderia
mimosarum, an unnamed Burkholderia species and, for the first time in South
America, Cupriavidus taiwanensis. The sampling site had a strong influence on
the diversity of the symbionts sampled, and the specific distributions of symbi-
otic populations between the soils were related to soil composition in some
cases. Some alpha-rhizobial strains taxonomically close to Rhizobium endophyti-
cum were also trapped in one soil, and these carried two copies of the nodA
gene, a feature not previously reported. Phylogenies of nodA, nodC and nifH
genes showed a monophyly of symbiotic genes for beta-rhizobia isolated from
Mimosa spp., indicative of a long history of interaction between beta-rhizobia
and Mimosa species. Based on their symbiotic gene phylogenies and legume
hosts, B. tuberum was shown to contain two large biovars: one specific to the
mimosoid genus Mimosa and one to South African papilionoid legumes.
Introduction
Rhizobia are a functional class of soil bacteria able to
develop a nitrogen-fixing symbiosis with legumes, and are
also termed legume nodulating bacteria (or LNB). The
legume nodulation ability is spread among the Alpha-
and Beta- subclasses of Proteobacteria, and the names
alpha- and beta-rhizobia have been proposed for conve-
nience (Gyaneshwar et al., 2011).
Beta-rhizobia were originally described in 2001 in two
parallel studies: one describing two Burkholderia strains
(STM678 and STM815) isolated from Aspalathus carnosa
(Papilionoideae) in South Africa and Machaerium luna-
tum (Papilionoideae) in French Guiana, respectively
(Moulin et al., 2001), which were subsequently named
Burkholderia tuberum and Burkholderia phymatum
(Vandamme et al., 2002); and one describing Ralstonia
taiwanensis from two Mimosa (Mimosoideae) species in
Taiwan (Chen et al., 2001), which was later renamed Cu-
priavidus taiwanensis (Vandamme & Coenye, 2004). Since
then, several diversity studies have demonstrated the
widespread occurrence of beta rhizobia as symbionts of
Mimosa species in Costa Rica and Texas (Barrett &
Parker, 2006; Andam et al., 2007), Panama (Barrett &
Parker, 2005; Parker, 2008), Brazil and Venezuela (Chen
et al., 2005b; Bontemps et al., 2010), Taiwan (Chen et al.,
2005a), India (Verma et al., 2004), Papua New Guinea
(Elliott et al., 2009), Australia (Parker et al., 2007), and
China (Liu et al., 2011).
To date, several nodulating beta-rhizobia species have
been described, most of them belonging to the Burkholde-
ria genus. The B. tuberum type strain (STM678T) is able
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
MIC
RO
BIO
LO
GY
EC
OLO
GY

to nodulate a range of Cyclopia species (Elliott et al.,
2007b). Strains affiliated to B. tuberum were also isolated
from Mimosa pigra in Panama (Barrett & Parker, 2005),
Mimosa spp. in Costa Rica (Barrett & Parker, 2006) and
from several diverse Mimosa species in Central Brazil
(Bontemps et al., 2010). Burkholderia mimosarum has
been isolated from M. pigra nodules in Taiwan, Venezu-
ela, Brazil, China and Australia (Chen et al., 2006; Parker
et al., 2007; Liu et al., 2011), while Burkholderia nodosa
was isolated from nodules on Mimosa scabrella and
Mimosa bimucronata in Brazil (Chen et al., 2007), and
Burkholderia sabiae from Mimosa caesalpiniifolia nodules,
also in Brazil (Chen et al., 2008). So far, only one nodu-
lating species has been described in the Cupriavidus
genus, namely C. taiwanensis (Chen et al., 2001), though
other nodulating strains have been isolated but not yet
described to species level. Cupriavidus strains were iso-
lated from nodules of Mimosa pudica, Mimosa diplotricha
and M. pigra in Taiwan (Chen et al., 2003, 2005a), Costa
Rica (Barrett & Parker, 2006), Texas (Andam et al., 2007)
and India (Verma et al., 2004).
The species B. phymatum was originally described
based on a single strain, STM815T, which was isolated
from M. lunatum nodules collected in French Guiana
(Moulin et al., 2001; Vandamme et al., 2002). This spe-
cies has since been reported as being isolated from M. pu-
dica nodules in Papua New Guinea (two strains, Elliott
et al., 2007a) and China (four strains, Liu et al., 2011),
but has not yet been found as a symbiont of Mimosa in
Australia, or in Central and South America (Chen et al.,
2003, 2005b; Barrett & Parker, 2005, 2006; Bontemps
et al., 2010). It is relevant to note that the Bontemps
et al. (2010) diversity study was performed on 47 native
(mainly endemic) Mimosa species in Central Brazil, and
not a single strain among the 148 isolates belonged to
B. phymatum. On the other hand, this is not so surpris-
ing, as the preferred symbiont of B. phymatum, M. pudic-
a, was only infrequently encountered in Central Brazil by
Bontemps et al. (2010), as its niche was occupied by the
other 200+ Mimosa spp. that are native/endemic to these
centres of Mimosa diversity (Simon & Proenca, 2000;
Simon et al., 2011). In addition, Central Brazilian endem-
ics appear not to nodulate effectively with B. phymatum
(dos Reis Junior et al., 2010). Nevertheless, although the
Cerrado/Caatinga biomes appear not to contain it, until
further sampling of M. pudica is performed in these
regions, the occurrence of B. phymatum in Central Brazil
cannot be excluded. The type strain of B. phymatum,
STM815, was originally isolated from a M. lunatum nod-
ule in a plant nursery in Paracou (French Guiana). The
demonstration of nodulation ability of M. lunatum by
STM815 has proven inconclusive because of the difficul-
ties in germinating its seeds, and growing this tree under
greenhouse conditions (L. Moulin, unpublished data),
although nodulation tests on a different species of
Machaerium (Machaerium brasilense) were also unsuccess-
ful (Elliott et al., 2007b). On the other hand, Elliott et al.
(2007b, 2009) and dos Reis Junior et al. (2010) showed
that STM815 has a very large host range on Mimosa spe-
cies, with higher symbiotic capacities (competition for
nodulation, nitrogen fixation within nodules) than C. tai-
wanensis LMG19424T. The question thus remains open
about the occurrence and diversity of B. phymatum in
South America.
Previous diversity studies have thus shown an affinity
between beta-rhizobia and Mimosa species. Some Mimosa
species also appear to have a broad host range for beta-
rhizobial strains. For example, in Bontemps et al. (2010),
among 118 strains recovered from 47 Mimosa species and
belonging to at least seven Burkholderia species, 98 were
able to nodulate M. pudica. Mimosa pudica thus appears
to be highly promiscuous, capable of nodulating with a
very diverse range of beta-rhizobial species. Based on this
fact, the aim of this study was to uncover the diversity of
beta-rhizobia using a soil-trapping strategy with M. pu-
dica as a trap host in French Guiana, a country located in
South America, which contains the principal centre of
diversification of the Mimosa genus, that is the Cerrado
region of Brazil (Simon & Proenca, 2000; Simon et al.,
2011). The specific questions we adressed were the follow-
ing: (i) What is the intra and interspecific diversity of
beta-rhizobia trapped by M. pudica in diverse soils of
French Guiana? (ii) Is B. phymatum predominant in
French Guiana? (iii) Is there a link between soil parame-
ters and beta-rhizobial diversity? (iv) What are the origins
of the symbiotic genes in the beta-rhizobial populations?
To isolate M. pudica compatible symbionts, we used a
trapping method on dilutions of soils recovered from
eight sites mainly around Cayenne with M. pudica as a
trap host. Strains were isolated from the nodules, tested
for their nodulation ability, and further characterized at
the genetical and genomic level. To determine the speci-
ficity of the symbiosis between beta-rhizobia and M. pu-
dica, we conducted the same trapping strategy on four
soils using Siratro (Macroptilium atropurpureum), a
broad-host-range legume that is able to trap both alpha
and beta-rhizobia (Lima et al., 2009). The diversity of
symbionts trapped by Siratro was then compared with
that of the symbionts trapped by M. pudica. Rhizobial
genetic diversity was assessed using 16S rRNA PCR-RFLP,
REP-PCR genomic fingerprints, as well as by phylogenies
based upon the 16S rRNA and recA housekeeping genes.
Phylogenies of the symbiosis-related genes nodA, nodC
and nifH were also performed in order to determine the
origin of symbiosis in the beta-rhizobial strains that we
isolated in the coastal lowlands of French Guiana.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
2 R.P.N. Mishra et al.

Materials and methods
Soil sampling and characteristics
Two hundred grams of soil was sampled under M. pudica
plants on eight sites in French Guiana (see Table 1),
mainly on Cayenne island (on the coastal side east of
Cayenne, near the towns of Remire and Montjoly) and
on the roadside between Cayenne and Kourou cities. GPS
coordinates and Flora composition of sampled soils are
given in Table 1 and Supporting Information Fig. S1.
Altitude of sampled sites ranged from 7 to 21 m.
Soil parameters were analysed by ‘Laboratoire d’analyse
des sols d’Arras’, INRA, France: pH (water and KCL
method), granulometry (five fractions), humidity (at
105 °C), C/N, CEC (Metson method), total nitrogen and
organic carbon, P2O5 (Joret–Hebert method), total
CaCO3, Na and H+ (BaCl2 extraction), Cu, Fe, Zn and
Mn contents (by ICP-AES).
Plant trapping of rhizobia
Mimosa pudica (B&T world seeds; ecotype M. pudica var.
hispida) was used as a legume host for trapping and for
nodulation tests of bacterial strains. Siratro (M. atropur-
pureum; UCAD, Dakar, Senegal) was also used as a trap
host in parallel experiments. Mimosa pudica seeds were
scarified and surface sterilized with 96% H2SO4 and 3%
Calcium hypochlorite (15 min each treatment, followed
by five or six washes with sterile dH2O); while Siratro
seeds were immersed for 3 min in 96% H2SO4 followed
by thorough washing. The seeds were then soaked over-
night in sterile dH2O, transferred to water agar plates
(0.8% Agar) and incubated overnight at 37 °C for germi-
nation. Seedlings were transferred to Gibson tubes or to
deep-well microplates, and incubated in a tropical plant
growth chamber (30 °C, 80% humidity, 16 h day/8 h
night). Five grams of soil was suspended in 50 mL of
sterile water and vortexed thoroughly. Serial dilutions up
to 10�2 were performed, and 0.7 mL of diluted soil sus-
pension was inoculated in to either Gibson tubes (80 mL)
or to deep-well microplates (8 mL per well) filled with
Jensen’s nutrient medium (Vincent, 1970). Eight repli-
cates were used for each dilution level.
Nodules were harvested at 28 days post-inoculation.
Nodules of varying sizes were selected for isolation (60
nodules for soils S1–S4, and 20 nodules for soils S5–S8).
Nodules were thoroughly washed in running tap water,
sterilized by immersing in 3% calcium hypochlorite for
5 min and washed five or six times with sterile dH2O.
Surface-sterilized nodules were individually crushed in
20 lL dH2O. Nodule contents were streaked on yeast
mannitol agar plates (Vincent, 1970) and incubated at
28 °C for 48 h. All colony types were picked and purified
twice by repeated streaking on YMA plates. All isolates
were retested for nodulation with M. pudica (or M. atro-
purpureum) using Gibson tubes under the growth condi-
tions described earlier, except that 1 mL of exponential
bacterial culture was inoculated into the tubes instead of
soil suspension.
All strains isolated from M. pudica and M. atropurpureum
nodules are listed in Table S1. For long-term
Table 1. Characteristics of soils used for rhizobial trapping
Soil Site characteristics and flora at proximity
GPS
coordinates
Altitude
(m)
Soil parameters
Gr pH Corg Ntot OM Cal Pho
S1 East Cayenne, Coastal gardens, Mimosa
pudica and Machaerium lunatum
4°56′46″N
52°18′03″W
8 Loamy sand 7.04 26.2 1.57 45.3 < 1 0.021
S2 East Cayenne, Montjoly town, M. pudica 4°55′19″N
52°16′38″W
8 Sand 5.95 10.6 0.806 18.3 < 1 < 0.01
S3 East Cayenne, Coastal path, M. pudica 4°56′25″N
52°17′41″W
7 Sand 6.14 10.5 0.816 18.2 < 1 0.011
S4 North Cayenne, Elysee savannah,
M. pudica
4°55′32″N;
52°24′50″W
21 Clay loam 5.39 18.2 0.977 31.5 < 1 < 0.01
S5 Cayenne South, Roadside, M. pudica 4°52′51″N;
52°18′46″W
9 Sandy loam 8.58 36.6 2.55 63.3 65.3 0.18
S6 Cayenne North, Roadside, M. pudica 5°02′50″N;
52°36′20″W
17 Clay loam 6.23 25.9 1.88 44.8 < 1 < 0.01
S7 West of Remire town, M. pudica,
Aeschynomene sp.
4°53′03″N;
52°17′34″W
8 Clay 5.44 59 4.65 102 < 1 0.013
S8 Rochambeau savannah, M. pudica 4°49′26″N;
52°21′20″W
6 Sandy loam 6.20 29.7 2.09 51.4 < 1 < 0.01
Gr, granulometry-soil texture according to particle size; pH, soil pH (in water); Corg, organic carbon content (g kg�1); Ntot, total nitrogen content
(g kg�1); OM, organic matter (g kg�1); Cal, CaCO3 content (g kg�1); Pho, phosphate content (g kg�1).
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 3

maintenance, strains were grown at 28 °C in YM broth
(Vincent, 1970) and preserved in 20% glycerol at �80 °C.
Molecular methods
Isolation of bacterial DNA was performed using the SDS-
proteinase K lysis procedure described by Moulin et al.
(2004). All PCR templates were performed with GO-Taq
Polymerase (Promega) following manufacturer instruc-
tions.
A nearly full-length16S rRNA gene was amplified by
PCR using the primer pair FGPS6 and FGPS1509 (Nor-
mand et al., 1992), as described in Moulin et al. (2001).
Ten microlitres of the 16S rRNA PCR product were
digested separately with the restriction endonucleases
MspI, HinfI and CfoI (Promega) in 20 lL reaction vol-
ume following manufacturer instructions. The digests
were analysed separately by agarose gel electrophoresis
(3% Metaphor; FMC Bioproducts).
The 800-bp recA fragments were amplified by PCR (ini-
tiation step 94 °C, 5 min, followed by 35 cycles of 94 °C,
30 s, 55 °C, 30 s, 72 °C, 30 s; and final step of 7 min at
72 °C) and sequenced using the primers recABur1F and
recABurk1R (Table 2) for beta-rhizobia, while TSrecAF
and TSrecAr were used for alpha-rhizobia under condi-
tions described in N’Zoue et al. (2009). PCR products were
purified using the Qiaquick Gel Extraction kit (Qiagen),
following manufacturers instructions, and were sequenced
using the same primers that were used for amplification.
A 440-bp fragment of nifH was amplified using primers
nifHfor and nifHrev (Table 2), as described in Chen et al.
(2003). A 500-bp fragment of nodA was amplified and
sequenced for all representative strains using nodA1F and
nodArbrad (Table 2), except strains of B. tuberum for
which specific primers pairs were designed (nodAF-190t,
nodAR1). For strains STM3625 and STM3629, two frag-
ments of the nodA gene were detected. For these two
strains, cloning of each nodA copy was performed using
the pGEMT-Easy kit (Promega), transformed into XL2
electrocompetent cells. Plasmids were extracted with the
Wizard Plus SV Miniprep kit (Promega), and inserts were
sequenced using the universal primers M13F and M13R.
An 800-bp fragment of nodC was also amplified and
sequenced for representative strains as described in Bon-
temps et al. (2010), except for C. taiwanensis strains for
which a specific primer set was designed (nodC-Ctai468F/
nodC-Ctai1231R) (Table 2).
DNA sequencing was subcontracted to Macrogen Inc.
and Genoscreen Inc., using ABI chemistry and ABI3730
sequencers, and the same primers as used for PCR.
REP-PCR fingerprints of each strain were obtained by
PCR amplification using primer pairs REP1R and REP2
(Versalovic et al., 1991). PCR mix and conditions were as
described by Kaschuk et al. (2006), except that the primer
annealing temperature was reduced to 40 °C. PCR prod-
ucts were visualized on 1% agarose gel in TAE 19 buffer
after elecrophoresis. Rep fingerprint groups were defined,
one rep-group including identical or nearly identical (up
to one band difference) REP-PCR profiles.
Phylogenetic analysis
Sequences were corrected using CHROMAS PRO v1.33 (Techn-
elysium Pty Ltd), aligned with CLUSTALX and alignments
Table 2. Primers used for amplification and sequencing of 16S rRNA, recA, nodA, nodC and nifH genes
Primer name Primer sequence (5′–3′) Target gene References
FGPS6 GGAGAG TTAGATCTTGGCTCA 16S rRNA Normand et al. (1992)
FGPS1509 AAGGAGGGGATCCAGCCGCA 16S rRNA Normand et al. (1992)
nifHfor AARGGNGGNATYGGHAARTC nifH Chen et al. (2003)
nifHrev GCRTA8ABNGCCATCATYTC nifH Chen et al. (2003)
NodA1f TGCRGTGGAARNTRBVYTGGGAAA nodA Sy et al. (2001)
NodAr.Brad TCACARCTCKGGCCCGTTCCG nodA Sy et al. (2001)
NodAF-190t AGTTGGGCCGGMGCNAGGCCTGA nodA This study
NodAR1 CAACGAACTGTTAATTGGCA nodA This study
nodCBurk2F ACTSATACTYAACGTMGAYTC nodC Bontemps et al. (2010)
nodCBurk2R GMRAAYCCRAGAAATCGAAG nodC Bontemps et al. (2010)
NodCCtai468F TACTCAAGATGCGGGACAGA nodC This study
NodCCtai1231R CCGGAGATTCCGATGATGGA nodC This study
nodCrhizo1F AYGTHGTYGAYGACGGTTC nodC Bontemps et al. (2010)
nodCrhizo1r CGYGACAGCCANTCKCTATTG nodC Bontemps et al. (2010)
TSrecAf CAACTGCMYTGCGTATCGTCGAAGG recA Stepkowski et al. (2005)
TSrecAr CGGATCTGGTTGATGAAGATCACCATG recA Stepkowski et al. (2005)
recABurk1F GATCGARAAGCAGTTCGGCAA recA This study
recABurk1R TTGTCCTTGCCCTGRCCGAT recA This study
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
4 R.P.N. Mishra et al.

were manually curated with GENEDOC software (Nicholas
& Nicholas, 1997). Phylogenies were built by distance on
MEGA4 (Tamura et al., 2007); or by Maximum likelihood
using PHYML (Guindon et al., 2005) or PAUP4. Bootstrapping
or aLRT (approximative Likelihood Ratio Tests, SH-like)
tests were conducted using the www.phylogeny.fr website
(Dereeper et al., 2008).
Statistical analyses
Shannon diversity indexes were calculated using PAST soft-
ware (Hammer et al., 2001). Genetic differentiation tests
were calculated with GENEPOP (Rousset, 2008). A principal
component analysis (PCA) of soils was performed from
the data matrix including soil physico-chemical parame-
ters and relative frequency of each rhizobial species using
XLSTAT (Addinsoft™).
Nucleotide sequence accession numbers
Sequences were deposited in GenBank under the acces-
sion numbers listed in Table S1.
Results
Trapping of M. pudica symbionts from French
Guiana soils
We trapped 221 isolates from eight soils in French
Guiana using M. pudica as a trap host. All isolates
nodulated M. pudica when tested as pure cultures under
axenic conditions, except for four Burkholderia cepacia
isolates that did not form any nodules by 4 weeks post-
inoculation.
The 221 purified isolates were distributed into 10 ribo-
types by 16S rRNA PCR-RFLP using three restriction
enzymes. Assignment of each profile to a rhizobial species
was then performed by the sequencing of the full 16S
rRNA sequence from a representative strain (see Table 3
and Fig. 1a). The isolates were distributed into three gen-
era and all of them could be assigned to a described spe-
cies based on per cent of 16S rRNA nucleotide identity
(1100 bp alignment): B. phymatum (99–100% 16S rRNA
gene identity with STM815T, two ribotypes, 63 strains),
B. tuberum (98–99% 16S rRNA gene identity with
STM678T, three ribotypes, 132 strains), B. mimosarum
(99–100% 16S rRNA gene identity with PAS44T, one rib-
otype, nine strains), Burkholderia sp. (97% with B. phy-
matum as the closest species, one ribotype, one strain),
C. taiwanensis (98–99% 16S rRNA gene identity with
LMG19424T, one ribotype, six strains), Rhizobium sp.
(98% with closest described species Rhizobium endophyti-
cum CCGE1052, one ribotype, four strains), and B. cepa-
cia (one ribotype, six strains).
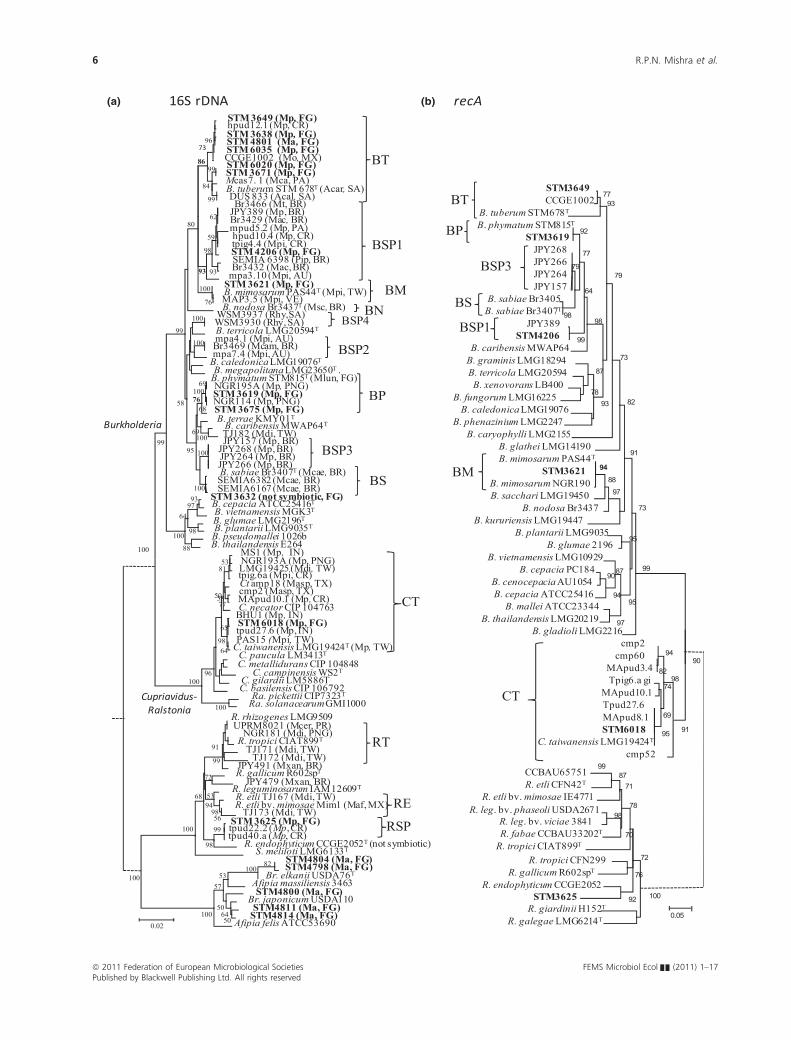
A 16S rRNA gene phylogeny (Fig. 1a) was built to infer
the evolutionary relationships among representative
strains of our collection, including selected reference
strains (mostly beta-rhizobial species). Mimosa symbionts
previously described in the literature, as well as M. pudica
strains from Central Brazil (Bontemps et al., 2010), were
also added to the analysis. The Burkholderia sequences
either fell into defined species of symbiotic Burkholderia
(B. mimosarum, B. phymatum, B. tuberum) or into clades
of undescribed species, for example STM4206 grouped in
a clade with strains isolated from diverse Mimosa species
(Mimosa acutistipula, M. pudica, Mimosa tenuiflora,
M. pigra) from Central and South America (Brazil, Pan-
ama and Costa Rica). Strain STM3632, however, fell into
Table 3. Mimosa pudica trapping: strain species assignment and distribution in soils
Species*
Representative
strain
16S-RFLP
ribotype Rep groups S1 S2 S3 S4 S5 S6 S7 S8 All
Burkholderia phymatum STM3619 AAA 13 16 – – 36 – 3 – 2 57
B. phymatum STM3675 DEG 1 – – – 2 – 4 – – 6
Burkholderia sp. STM4206 AAE 1 1 – – – – – – – 1
Burkholderia tuberum STM3649 ABF 12 – 45 55 18 1 2 2 5 128
B. tuberum STM3638 ABD 2 – 2 – – – – – – 2
B. tuberum STM6020 FBF 2 – – – – – – – 2 2
Burkholderia mimosarum STM3621 ABB 3 9 – – – – – – – 9
Cupriavidus taiwanensis STM6018 EGI 3 – – – – 6 – – – 6
Burkholderia cepacia STM3632 CDD ND 5 – – 1 – – – – 6
Rhizobium sp. STM3625 BCC 2 4 – – – – – – – 4
Total 35 47 55 57 7 9 2 9 221
All strains samples are listed in Table S1. S1–S8 refer to the soils listed in Table 1, and numbers indicates distribution of 16S-RFLP types across
soils. Burkholderia phymatum type strain STM815 belonged to RFLP profile AAA.
ND, not determined.
*Species assignment was based on 16S rRNA gene phylogeny.
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 5
B. tuberum STM 678T (Acar, SA)DUS 833 (Acal, SA)
Mcas7. 1 (Mca, PA)
STM 6020 (Mp, FG)STM 3671 (Mp, FG)
STM 6035 (Mp, FG)
STM 3638 (Mp, FG)hpud12.1 (Mp, CR)
STM 3621 (Mp, FG)B. mimosarum PAS44T (Mpi, TW) MAP3.5 (Mpi, VE)
Br3429 (Mac, BR)JPY389 (Mp, BR) Br3466 (Mt, BR)
mpud5.2 (Mp,PA)hpud10.4 (Mp,CR) tpig4.4 (Mpi, CR)STM 4206 (Mp, FG)SEMIA 6398 (Pip, BR)Br3432 (Mac, BR)
mpa3.10 (Mpi, AU)
B. nodosa Br3437T (Msc,BR)WSM3937 (Rhy,SA)WSM3930 (Rhy, SA)B. terricola LMG20594T
mpa4.1 (Mpi, AU)Br3469 (Mcam, BR) mpa7.4 (Mpi, AU)
B. caledonica LMG19076T
B. megapolitana LMG23650T .
JPY157 (Mp, BR) JPY268 (Mp, BR) JPY264 (Mp, BR) JPY266 (Mp, BR) B. sabiae Br3407T (Mcae, BR)SEMIA6382 (Mcae, BR)SEMIA6167 (Mcae, BR)
B. phymatum STM815T (Mlun, FG)NGR195A (Mp, PNG)STM 3619 (Mp, FG)NGR114 (Mp, PNG) STM 3675 (Mp, FG)B. terrae KMY01T
TJ182 (Mdi, TW)
STM 3632 (not symbiotic, FG)B. cepacia ATCC25416T
B. vietnamensis MGK3T
B. glumae LMG2196T
B. plantarii LMG9035T
B. pseudomallei 1026bB. thailandensis E264
MS1 (Mp, IN)
LMG19425 (Mdi, TW)tpig.6a (Mpi, CR)Ct amp18 (Masp, TX) cmp2 (Masp, TX) MApud10.1 (Mp,CR) C. necator CIP 104763
C. taiwanensis LMG19424T (Mp, TW)PAS15 (Mpi, TW)
STM 6018 (Mp, FG)BHU1 (Mp, IN)
tpud27.6 (Mp, IN)
C. paucula LM3413T
C. metallidurans CIP 104848 C. campinensis WS2T
C. gilardii LM5886T C. basilensis CIP 106792
Ra. pickettii CIP7323T
Ra. solanacearumGMI1000
NGR181 (Mdi, PNG)
R. rhizogenes LMG9509
TJ171 (Mdi,TW)
UPRM8021 (Mcer, PR)
R. tropici CIAT899T
TJ172 (Mdi, TW) JPY491 (Mxan, BR) R. gallicum R602spT
JPY479 (Mxan, BR)
TJ173 (Mdi,TW) R. etli bv. mimosae Mim1 (Maf, MX) R. etli TJ167 (Mdi,TW)
R. leguminosarum IAM 12609T
STM 3625 (Mp, FG)
S. meliloti LMG6133T
Br. japonicum USDA110
Afipia massiliensis 3463
Afipia felis ATCC53690
Br. elkanii USDA76T
BT
BN
BP
B. caribensis MWAP64T
NGR193A (Mp, PNG)
R. endophyticum CCGE2052T (not symbiotic)
STM4804 (Ma, FG)STM4798 (Ma, FG)
STM4800 (Ma, FG)
STM4811 (Ma, FG)STM4814 (Ma, FG)
98
9397
88
64
100
100
65
64
98
5381
775850
96
100
100
76
100
100
100
100
100
69
68
100
99
99
93
62
59
98
96
76
69
95
93
84
86
80
58
99
99
100
91
99
5698
94
72
68
100
82100
100
5064100
50
57
53
0.02
STM 3649 (Mp, FG)
STM 4801 (Ma, FG)
BM
BSP1
BS
BSP2
CT
BSP3
53
Burkholderia
Cupriavidus-
Ralstonia
16S rDNA
tpud22.2 (Mp, CR)tpud40.a (Mp, CR)
99
98
73
CCGE1002 (Mo, MX)
RT
RE
RSP
BSP4
(a)
STM3649
CCGE1002
B. tuberum STM678T
B. phymatum STM815T
STM3619
JPY268
JPY266
JPY264
JPY157
B. sabiae Br3405
B. sabiae Br3407T
JPY389
STM4206
B. caribensis MWAP64
B. graminis LMG18294
B. terricola LMG20594
B. xenovorans LB400
B. fungorum LMG16225
B. caledonica LMG19076
B. phenazinium LMG2247
B. caryophylli LMG2155
B. glathei LMG14190
B. mimosarum PAS44T
STM3621
B. mimosarum NGR190
B. sacchari LMG19450
B. nodosa Br3437
B. kururiensis LMG19447
B. plantarii LMG9035
B. glumae 2196
B. vietnamensis LMG10929
B. cepacia PC184
B. cenocepacia AU1054
B. cepacia ATCC25416
B. mallei ATCC23344
B. thailandensis LMG20219
B. gladioli LMG2216
cmp2
cmp60
MApud3.4
Tpig6.a gi
MApud10.1
Tpud27.6
MApud8.1
STM6018
C. taiwanensis LMG19424T
cmp52
CCBAU65751
R. etli CFN42T
R. etli bv. mimosae IE4771
R. leg. bv. phaseoli USDA2671
R. leg. bv. viciae 3841
R. fabae CCBAU33202T
R. tropici CIAT899T
R. tropici CFN299
R. gallicum R602spT
R. endophyticum CCGE2052
STM3625
R. giardinii H152T
R. galegae LMG6214T
97
90
94
87
95
95
94
88
97
78
87
93
99
92
98
79
64
77
98
77
93
79
73
82
91
73
99
95
74
69
82
94
98
91
90
10092
72
76
98
70
78
71
87
99
0.05
BP
BT
BSP1
recA
BS
BSP3
BM
CT
(b)
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
6 R.P.N. Mishra et al.

the same clade as ATCC25416, the type strain of the non-
symbiotic B. cepacia species. The 16S rRNA sequence
from Cupriavidus STM6018 clustered in the large C. tai-
wanensis branch of the tree, together with strains isolated
from Mimosa species Worldwide, represents the first
C. taiwanensis isolate from South America (the closest
isolate being one from Costa Rica). The alpha-rhizobial
strain STM3625 fell into the Rhizobium genus, and was
very close to two Rhizobium strains isolated from M. pu-
dica in Costa Rica (tpud22.2 and tpud40.a; sharing 99%
identity). These three strains together formed a clade
close to R. endophyticum (98% identity), which is a
nonsymbiotic endophytic strain isolated from Phaseolus
vulgaris in Mexico (Lopez-Lopez et al., 2010).
The recA gene, a house-keeping gene commonly used
in rhizobial phylogeny (N’Zoue et al., 2009; Miche et al.,
2010), was also sequenced for representative strains to
confirm their species assignment based on their 16S
rRNA gene phylogeny. The derived recA phylogeny
showed identical groupings of strains compared with their
16S rRNA gene phylogenies (Fig. 1b), except for the
BSP1 clade, whose phylogenetic postion was better
resolved using recA (and placed it closer to the B. sabiae
and B. phymatum clades).
Distribution of species and ribotypes across
soil samples
The distribution of 16S rRNA PCR-RFLP types in soils is
presented in Table 3. Rhizobial species exhibited a spe-
cific distribution for each soil, and a variable number of
species was sampled depending on the soil of origin. It is
notable that trapping in soils S5–S8 gave significantly less
nodules (c. 10 nodules per soil sample) and hence, fewer
isolates, compared with soils S1–S4.
Soil S1 hosted four species of Mimosa rhizobia (B. phy-
matum, B. mimosarum, Burkholderia sp., Rhizobium sp.),
while soils S2, S3 and S7 hosted only one (B. tuberum).
Soils S4, S6 and S8 hosted the two genotypes of B. phy-
matum and B. tuberum, and the S5 genotypes of C. tai-
wanensis and B. tuberum (Table 3). Amongst all
genotypes isolated, the most abundant were B. tuberum
(132 strains), followed B. phymatum (63), B. mimosarum
(8), C. taiwanensis (6), B. cepacia (5) and Rhizobium sp.
(4). The most frequent ribotypes in the soils were B.
tuberum (ABF) and B. phymatum (AAA). Other geno-
types were found only in one soil, such as Rhizobium sp.
(BCC), Burkholderia sp. (AAE) and B. mimosarum in soil
S1, and C. taiwanensis in soil S5. A B. cepacia genotype
was also found for several isolates (CDD) in two different
soils (S1 and S4).
Shannon diversity indexes were calculated for each site
and compared (excluding sites S5–S8 for which too few
strains could be sampled). At the species level, two sites
(S2 and S3) were monospecific with regard to isolated
M. pudica symbionts, with 100% B. tuberum (H = 0),
while S1 was the most diverse (H = 1.33), and S4 was
intermediate (H = 0.69). The three index values were sta-
tistically different from each other (v2 exact test,
P < 0.001).
As population structures of Mimosa rhizobia differed
significantly across soil samples, we compared their dis-
tribution with soil parameters. The most striking fea-
tures were that soils S2 and S3 exhibited nearly
identical soil parameters, and they hosted only a single
species, B. tuberum. Cupriavidus strains were only found
in soil S5, which, as the only basic soil amongst all the
soils sampled, was characterized by a high pH value
(8.9). Soil S1 exhibited a neutral pH value (7.04), and
hosted the highest diversity at the species level, including
Fig. 1. 16S rRNA (a) and recA (b) gene phylogenetic trees of Mimosa symbionts and related species (among Alpha- and Betaproteobacteria). The
16S rRNA gene tree was built by Neighbour Joining from a matrix corrected by the Kimura-2 parameter method. Numbers at nodes are
bootstrap% from 1000 replicates (shown only when > 50%). The recA tree was built by Maximum Likelihood following a GTR model; numbers
at nodes are aLRT tests values (Approximate Likelihood-ratio Test, SH-like; shown only when > 50%) for branch support. Scale bars indicate
numbers of substitutions per site. Broken tree lines indicate that branch length is not informative (upper branch trees were reduced to fit to
page). Both trees were rooted with Pseudomonas fluorescens Pf5 (Gammaproteobacteria). Strains isolated during this study (French Guiana) are
in bold (from Mimosa pudica and Macroptilium atropurpureum trapping). Defined bacterial species of Mimosa symbionts are indicated on each
side of the trees: BP, Burkholderia phymatum; BT, Burkholderia tuberum; BSP1–BSP3, Burkholderia spp. clade 1–clade 3; BM, Burkholderia
mimosarum; BS, Burkholderia sabiae; CT, Cupriavidus taiwanensis; RT, Rhizobium tropici; RE, Rhizobium etli; R., Rhizobium; B., Burkholderia; Br.,
Bradyrhizobium. Informations about plant host and geographical origin are only given for beta rhizobia. Abbreviations used: for Mimosa species:
Mp: Mimosa pudica, Mo: Mimosa occidentalis, Mca: Mimosa casta, Mt: Mimosa tenuiflora, Mac: Mimosa acutistipula, Mpi: Mimosa pigra, Msc:
Mimosa scabrella, Mcam: M. camporum, Mdi: Mimosa diplotricha, Mcae: Mimosa caesalpiniifolia, Masp: M. asperata, Mxan: M. xanthocentra,
Mcer: Mimosa ceratonia, Maf: Mimosa affinis; others plants: Ma: Macroptilium atropurpureum, Acar: Aspalathus carnosa, Acal: A. callosa, Pip:
Piptadenia, Rhy: Rhynchosia, Mlun: Machaerium lunatum; strain origin: FG: French Guiana, CR: Costa Rica, SA: South Africa, BR: Brazil, PA:
Panama, AU: Australia, VE: Venezuela, TW: Taiwan, PNG: Papouasie New Guinea, IN: India, TX: Texas (USA), PR: Puerto Rico. Scale bar indicates
number of substitutions per site. Accession numbers are available in Table S3.
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 7

both alpha and beta-rhizobial genotypes. All other soils
were acidic and only yielded Burkholderia strains.
Results of PCA with regard to soil physico-chemical
characteristics and frequency of each rhizobial species
are shown in Fig. S2a and b. The values for the three
first principal components of soil factors F1, F2 and F3
were 55%, 21% and 18%, respectively. In both PCA
plots, soils S2 and S3 (in which B. tuberum predomi-
nated) were not distinguished, while soil S7 was sepa-
rated from all the other soils. The alkaline pH and the
high phosphate and CaCO3 content discriminated soil
S5 (which contained C. taiwanensis) along F2. More
generally, the main soil physico-chemical characteristics
related to the distribution of rhizobial species were pH,
phosphate and CaCO3 content, and soil texture (espe-
cially broad sand).
Intraspecific variability of isolates by REP-PCR
fingerprinting
To assess the intraspecific variability of each species sam-
pled, REP-PCR fingerprints were produced for all isolates
(except the B. cepacia strains). The distribution across
soils of each REP-PCR group is given in Table S2. For
B. phymatum and B. tuberum species, 14 and 16 REP-PCR
groups could be distinguished (BP1–BP14; BT1–BT16),
respectively, with two genotypes dominating the isolates
(BP1 and BP11 for B. phymatum, BT1, BT2 for B. tube-
rum) with an even distribution across soils. Some
REP-PCR groups were found to be soil specific, such as
BT7–BT11 in soil S2. The type strain of B. phymatum,
STM815, was included in this study, and its REP-PCR
profile was close but not identical to one of the dominant
B. phymatum groups (BP1), which was found in soils S1
and S4. For B. mimosarum, Rhizobium sp. and C. taiwan-
ensis species, only two to four REP-PCR groups were found
(probably due to a low number of isolates of these species).
At the intraspecific level, and including all genotypes
(based on REP-PCR fingerprints), Shannon indexes were
H = 2.175 (S1), 1.921 (S2), 0.9689 (S3), 1.964 (S4). Soil
S3 was statistically different from the other three sites.
Despite only one species being detected in soil S2 (B. tube-
rum), the number of haplotypes was as high as in S1
(hosting four species), and surprisingly had a much
higher number of haplotypes than S3, which contained
the same rhizobial species, B. tuberum (S2: 10 haplotypes;
S3: 4 haplotypes). Burkholderia phymatum was the most
diverse species (H = 2.29). The three other rhizobial spe-
cies (B. mimosarum, B. tuberum and C. taiwanensis) exi-
hibited similar diversity patterns (H = 1.14; 1.94, 1.01,
respectively). Rhizobial populations at each site (S1–S4)
were compared on the basis of their genetic composition
using Chi-square exact tests. All four populations were
highly differentiated (P > 0.001 for each pair), meaning
that the site of sampling had a high influence on the
diversity of rhizobia sampled.
Phylogenies of nodA, nodC and nifH symbiotic
genes
Phylogenies of nodA and nifH gene fragments of repre-
sentative strains of each 16S rRNA PCR-RFLP type were
built to infer the origin and evolutionary relationships of
symbiotic genes among the bacterial collection.
The nodA data set included representative strains from
each genotype previously described plus nodA sequences
from known beta-rhizobia (essentially strains isolated
from Mimosa and Rhynchosia), as well as sequences from
alpha-rhizobia reference strains and those isolated from
Mimosa. Figure 2a shows a nodA phylogenetic tree
inferred by Maximum likelihood and rooted with Azo-
rhizobium caulinodans ORS571. All beta-rhizobial nodA
sequences grouped in a large clade including isolates from
Mimosa species. This clade was monophyletic and subdi-
vided into four subclades corresponding to: (i) B. phyma-
tum, B. sabiae and Burkholderia sp. strains (BP/BS/BSP1-2
in Fig. 2a), (ii) C. taiwanensis strains (CT), (iii) B. mimo-
sarum strains (BM), (iv) B. tuberum Mimosa strains
(BT1). The BP/BS/BSP1-2 subclade included the B. phy-
matum type strain STM815, NGR195A (isolated from
M. pudica in Papua New Guinea) and DUS751 (isolated
from M. pigra in Vietnam), as well as B. sabiae Br3407
Fig. 2. nodA (a) and nifH (B) phylogenetic trees of Mimosa symbionts and related species (among Alpha- and Betaproteobacteria). Both trees
were built by ML following a GTR model (all ML parameters estimated). Scale bar indicates numbers of substitutions per site. Numbers at nodes
are boostrap% from 100 replicates (shown only when > 50%) performed under ML criterion. Arrows indicate the monophyly of nodA and nifH
in Mimosa beta-rhizobia. Broken tree lines indicate that branch length is not informative (upper branch trees were reduced to fit to page). nodA
and nifH trees were rooted with Azorhizobium caulinodans and Azotobacter vinelandii, respectively. Strains isolated during this study (French
Guiana) are in bold (both Mimosa pudica and Siratro trapping). Species affiliations of each strain are given before each strain name following the
16S rRNA gene phylogeny (see Fig. 1 legend for abbreviations). Information about plant host and geographical origin are given for Mimosa
symbionts. Scale bar indicates number of substitutions per site. The arrow indicates the monophyletic origin of nodA and nifH genes for beta-
rhizobia isolated from Mimosa spp. Abbreviations are the same as those given in Fig. 1 legend, except: Mbi: Mimosa bimucronata, Min: M.
invisa, Acang: Acaciella angustissima, B. sp.: undefined Burkholderia species, leg. leguminosarum. Scale bar indicates number of substitutions per
site. Accession numbers are available in Table S3.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
8 R.P.N. Mishra et al.

Bm NGR190 (Min, PNG) Bm STM3621 (Mp, FG)B. sp. PTK47 (Mpi, TW) Bm PAS44T (Mpi, TW)
B. sp. DUS751 (Mpi) Bm MAP3-5 (Mpi, VE)
Bm Br3454 (Msc, BR) Bn BR3461 (Mbi, BR)
Bt STM6020 (Mp, FG)Bt STM3638 (Mp, FG)Bt STM3649 (Mp, FG)Bt STM4801 (Mp, FG)
B. vietnamensis G4 B. xenovorans CAC124
B. xenovorans LB400
Bt STM678 (Acar, SA)
Br. elkani STM4798 (Ma, FG)Bradyrhizobium sp. ANU289
Br. japonicum STM4811 (Ma., FG) Br. japonicum USDA110
Br. elkanii STM4804 (Ma. atro., FG)
photosynthetic BradyrhizobiumORS278 Azorhizobium caulinodans ORS571
Xanthobacter autotrophicus Py2
Re CFN42T
Re TJ173 (Mdi, TW) Re TJ167 (Mdi, TW)
Re Mim1 (Mafi, MX)
Rsp STM3625 (Mp, FG)R. gallicum R602sp
Rt TJ171 (Mdi, TW)Rt TJ172 (Mdi, TW)
R. leg. bv. viciae 3841R. leg. bv. trifolii K0049
S. meliloti 1021
M. loti MAFF303099
S. fredii NGR234Sinorhizobium sp. ANU240
Br. sp. STM4800 (Ma, FG)
Rhodopseudomonas palustris CGA009
BT1 bv mimosae
BSP4/BT2 bv papilionoidae
Ct MApud8.1 (Mp, CR)
Ct MApud3.4 (Mp, CR)
Ct MApud10.1 (Mp, CR)
Ct Tpud27.2 (Mp, CR) Ct tpig.6a (Mpi, CR)
Ct STM6018 (Mp, FG)Ct MS1 (Mp, IN)
Ct LMG19424T (Mp, TW) Ct LMG19425 (Mdi, TW)
Ct NGR193A (Mp, PNG)
Ct cmp2 (Masp, TX)
Ct BHU1 (Mp, IN)
Ct PAS15 (Mpi, TW) Ct amp18 (Masp, TX)
CT
Bp STM3619 (Mp, FG)Bp STM3675 (Mp, FG)
Bsp1 STM4206 (Mp, FG)
Bcar TJ182 (Mdi, TW)
Bp NGR195A (Mp, PNG) Bsp2 BR3469 (Mcam, BR) Bp STM815T (Mlun& FG)
Bs Br3407 (Mcae, BR)
BP/BS/BSP1-2
BM
BN
nifH
Bsp1 JPY389 (Mp, BR)
Bsp3 JPY266 (Mp, BR)Bsp3 JPY264 (Mp, BR)Bsp3 JPY268 (Mp, BR)Bsp3 JPY157 (Mp, BR)
Bt CCGE1002 (Mo, MX)
70
62
100
100
47
94
85
59
78
100
94
89
100
83
99
86
100
100
100
94
75
100
76 86
80
97
100
95
100
100
90
54
98
100
94
100
0.1
BSP3
Bsp4 WSM3930 (Rhy, SA) Bsp4 WSM3937 (Rhy, SA)
100
RE
RT
Azorhizobium caulinodans ORS571
Bs Br3407 T(Mcae, BR)Bsp1 STM4206 (Mp, FG)Bp STM3619 (Mp, FG)Bp STM815T (Mlun, FG)Bp NGR195 Bp STM3675 (Mp, FG)Bp DUS751 (Mpi, VT)
Bsp2 Br3469 (Mcam, BR) Ct Tpig6a (Mpi, CR) Ct MApud10.1 (Mp, CR) Ct NGR193A (Mp, PNG)Ct LMG19425 (Mdi, TW)Ct LMG19424T (Mp, TW) Ct Tpud27.6 (Mp, CR)Ct STM6018 (Mp, FG)Ct cmp2 (Masp, TX)Bcar TJ182 (Mdi, TW) Ct PAS15 (Mpi, TW)
Ct amp18 (Masp, USA)Bm PAS44T (Mpi, TW) Bm MAP3-5 (Mpi, VE) )Bm PTK47 (Mpi, TW) B. sp. mpa6.4 (Mpi, AU) B.sp. mpa2-1a (Mpi, AU) Bm STM3621 (Mp, FG)Bm NGR190 (Mdi, PNG)Bm Br3454 (Msc, BR)
Bn Br3461 (Mbi, BR)Bt STM6020 (Mp, FG)Bt STM3638 (Mp, FG)Bt STM3649 (Mp, FG)Bt CCGE1002 (Mo, MX)
Rt TJ171 (Mdi, TW)Rt TJ172 (Mdi, TW)Rt NGR181 (Mdi, PNG)Rt CFN299T
Rt ESH10 (Acang, MX)Rt CFNESH9 (Acang, MX)Rt ESH23 (Acang, MX)Rsp. STM3625 (I) (Mp, FG)Rsp. STM3629 (I) (Mp, FG)
Rsp STM3625 (II) (Mp, FG)Rsp STM3629 (II) (Mp, FG)
R. sp CFNESH34 (Acang, MX) Re Mim1 (Maf, MX)Re TJ167 (Mdi, TW) (AJ505305)
S. mexicanus ITTGS3 S. chiapanecum ITTGS1 S. chiapanecum ITTGR11 S. chiapanecum ITTHS70 Sinorhizobium sp. ITTGS8
M. plurifarium CFNESH18 M. plurifarium CFNESH19
M. plurifarium CFNESH26 S. kostiense HAMBII1489T
B. tuberum STM678T (Acar, SA) Bsp4 WSM3930 (Rhy, SA)Bsp4 WSM3937 (Rhy, SA)
Methylobacterium nodulans ORS2060T Methylobacterium sp. 4-46
S. fredii NGR234 R. etli CFN42 Br. japonicum USDA110
Br. elkanii LMG6134T
M. loti MAFF303099 R. leg. bv. trifoli WSM1325 R. leg. bv. trifoli ANU843S. meliloti 1021
R. leg. bv. viciae 248
100
56
100
100
100
82
66
97
51
85
90
56
97
99
97
83
100
78
90
100
100
96
99
76
63100
98
65
74
72
100
94
100
70
99
100
100
97
78
0.1
BP/BS/BSP1-2
CT
BM
BT1 bv mimosae
BN
nodA
RE
RT
BSP4/BT2 bv papilionoidae
(a) (b)
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 9

and Burkholderia sp. Br3469, two strains isolated from
Mimosa species in Brazil. The CT subclade contained
strains from diverse Mimosa species isolated from North
(Texas) to Central and South America, Asia and Indone-
sia. The BT subclade encompassed all B. tuberum strains
from Mimosa including CCGE1002 (isolated from
Mimosa occidentalis in Mexico). Interestingly, B. tuberum
Mimosa isolates did not group with the type strain of
B. tuberum, STM678, which was isolated from a papilio-
noid legume (A. carnosa) in South Africa. Instead,
STM678 forms a clade within a large branch that includes
all the alpha-rhizobia, and it clusters with Rhynchosia
strains (belonging to the BSP4 clade) that were also iso-
lated from South Africa (Garau et al., 2009).
Two nodA fragments from Rhizobium sp. STM3625
could be amplified by PCR. Cloning and sequencing led
to two separate nodA copies sharing 73% nucleotide iden-
tity. These two nodA copies could also be detected in the
second REP-PCR group of Rhizobium sp. (strain
STM3629). Phylogenetic analyses (Fig. 2a) revealed that
the first copy (named nodA-I) grouped together with Rhi-
zobium tropici strains isolated in Mexico from Acaciella
angustissima (Rincon-Rosales et al., 2009), while the sec-
ond copy (nodA-II) formed a new branch in the nodA
tree without close homologues.
The nifH gene tree topology of Mimosa isolates
(Fig. 2b) was very close to that of the nodA one. The only
difference was that the phylogenetic placement found for
B. tuberum isolates from Mimosa (BT1 clade) differed
from those from South African legumes (BT2 clade), that
is the B. tuberum isolates from Mimosa species were
included in the same ‘Mimosa’ panclade in terms of their
nifH and nodA phylogenies, while the B. tuberum isolates
from South Africa grouped in a separate betaproteobacte-
rial subclade with Burkholderia xenovorans and Burkhol-
deria fungorum.
A large survey of Mimosa symbionts from Central Brazil
was published during the course of this study, with the
publication of a large data set of nodC sequences (Bon-
temps et al., 2010). In order to compare the symbiotic
genes of French Guiana isolates with Brazilian ones, we
sequenced a nodC fragment from each representative
strain in our collection. As for nodA, the nodC phylogeny
(Fig. 3) grouped together all the beta-rhizobial Mimosa
strains, with the main difference being in the sub-branch-
ing of the CT subclade compared with the Burkholderia
subclades. Indeed in the nodA and nifH phylogenies, CT
formed a sister clade to (BP/BS/BSP1-2), while in the
nodC tree, the CT subclade was more deeply branched at
the base of the Mimosa beta-rhizobial panclade. Such
phylogenetic incongruence between trees was further stud-
ied using different ML models to determine whether long-
branch attraction or third base codon saturation were a
cause of bias in the nodA, nifH or nodC trees. Trees built
using a ML codon model (as defined in Moulin et al.,
2004) or by excluding the third base codon position (not
shown) produced the same topologies as the trees shown
in Fig. 2 (for nodA and nifH) and Fig. 3 (for nodC).
Trapping of rhizobia with the broad-host-range
legume M. atropurpureum
In this study, we also wondered whether the trapping
results obtained with M. pudica could be explained by
beta rhizobia being the main rhizobial populations in
French Guiana soil, or if they were because of any speci-
ficity between these symbiotic partners.
In order to assess this, we used the same soil dilutions
that were used for Mimosa trapping (only on first four
soils S1–S4), but with a broad-host-range trapping
legume species, Siratro. A summary of the diversity sam-
pled is given in Table 4, and the complete set of trapped
strains is given in Table S1. Most of the rhizobia trapped
with Siratro were Bradyrhizobium strains, and only one
isolate of Burkholderia could be trapped from soil S2
using this plant host (STM4801, a B. tuberum strain).
Discussion
Mimosa pudica has an affinity for diverse beta-
rhizobia
Specificity between beta-rhizobia and Mimosa species is a
feature reported in several studies based on their isolation
from wild nodules (Chen et al., 2003, 2005a, b, Bontemps
et al., 2010). In this study, we trapped the M. pudica
compatible symbiont diversity from soils and observed a
strong affinity of this legume species for diverse beta-
rhizobial species. To test whether this trapping strategy
(and the observed affinity) was not biased by the natural
populations of rhizobia present in the sampled soils, we
performed a parallel trapping experiment with the broad-
host-range legume Siratro, a legume species that has been
previously shown to trap both alpha and beta-rhizobia
Fig. 3. Maximum likelihood nodC phylogenetic tree of Mimosa spp. symbionts and rhizobial reference strains. ML parameters used: GTR,
estimation of all parameters. Scale bar indicates number of subsitutions per site. Numbers at nodes are aLRT values (approximate Likelihood Ratio
test). Names in bold are Mimosa pudica symbionts. Abbreviations are the same as those in Fig. 1. Plant host, geographical origin and accession
numbers used are indicated after each strain name. Accession numbers are available in Table S3. *STM815 is a broad-host-range Mimosa
symbiont, but was isolated from a Machaerium lunatum nodule.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
10 R.P.N. Mishra et al.

Azorhizobium caulinodans ORS571
C. taiwanensis LMG19424T (M. pudica, TW)Ct STM6018 (M. pudica, FG)
Sinorhizobium sp. NGR234 M. huakui R7A M. loti MAFF303099
Br. japonicum USDA110R. etli bv. mimosae Mim74 (M.affinis, MX)R. etli TJ173 (M. diplotricha, TW)
R. etli CFN42S. meliloti 1021
R. leguminosarum 3841 Methylobacterium nodulans ORS2060Rhizobium sp. JPY491 (M. xanthocentra, BR)Rhizobium sp. JPY479 (M. xanthocentra,BR)Rhizobium sp. TJ172 (M. diplotricha, TW)
Rhizobium sp. NGR181 (M. invisa, PNG)R. tropici UPRM8021 (Mimosa sp.)
Rhizobium sp. STM3625 (M. pudica, FG)
100
10038
99
95
92
79
65
93
94
94
46
100
32
81
0.1
JPY395 (M. adenocarpa, BR) JPY256 (M. setosa, BR) JPY476 (M. somnians, BRBt STM3649 (M. pudica, FG)JPY604 (M. xanthocentra, BR)JPY636 (M. xanthocentra, BR)JPY634 (M. debilis, BR) JPY637 (M. debilis, BR)JPY326 (M. xanthocentra, BR)JPY325 (M. xanthocentra, BR)JPY595 (M. gracilis, BR) JPY489 (M. thermarum, BR)JPY453 (M. melanocarpa, BR)JPY403 (M. lanuginosa, BR)JPY622 (M. gracilis, BR)JPY410 (M. humivagans, BR)JPY418 (M. gracilis, BR) JPY419 (M. discobola, BR)JPY490 (M. callithrix, BR)JPY468 (M. somnians, BR, BR)JPY605 (M. somnians, BR)
JPY407 (M. somnians, BR)JPY623 (M. radula, BR)JPY431 (M. adenocarpa, BR)JPY385 (M. crumenarioides, BR)
JPY382 (M. velloziana,BR) JPY585 (M. quadrivilis, BR)JPY346 (M. cordistipula, BR) JPY381 (M. gemmulata, BR)
JPY388 (M. lewisii, BR)JPY350 (M. gemmulata, BR)
JPY294 (M. setosissima, BR)JPY404 (M. claussenii, BR)JPY414 (M. venatorum, BR)JPY629 (M. cyclophylla, BR)JPY452 (M. gardneri, BR)JPY300 (M. setosa, BR)JPY283 (M. claussenii, BR)JPY160 (M. claussenii, BR)JPY578 (M. blanchetti, BR)JPY587 (M. velloziana, BR)JPY439 (M. ulei, BR) JPY456 (M. laniceps, BR)Bm STM3621 (M. pudica, FG)B. mimosarum PAS44T(M. pigra, TW)JPY321 (M. pigra, BR)
Bp STM3619 (M. pudica, FG)B. phymatum STM815T (Machaerium lunatum*, FG)Bsp1 STM4206 (M. pudica, FG)
Bsp3 JPY268 (M. pudica, BR)Bsp3 JPY380 (M. gemmulata, BR)
84
99
96
98
73
85
99
90
100
99
99
92
98
100
82
100
94
92
BT
BN
BM
Bsp1 JPY389 (M. pudica, BR)JPY461 (M. quadrivilis, BR)JPY359 (M. tenuiflora, BR)
BP/BSP1/
BSP3
CT
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 11

from Brazilian soils (Lima et al., 2009). In our study,
Siratro trapped mainly Bradyrhizobium (96% of isolates)
and only one Burkholderia isolate could be detected, while
M. pudica trapped mostly beta-rhizobia (98% of isolates).
Such results clearly show a specificity of M. pudica
towards beta-rhizobia and that this legume species can be
used as a trap host to recover a large number of Burk-
holderia species (and C. taiwanensis) from soil.
Populations of compatible M. pudica symbionts
differ depending on French Guiana soil under
study
Our sampling strategy revealed different assemblages of
species depending on the soil under study. Some species,
such as B. tuberum, were dominant in several soils, while
others had only minor populations (Rhizobium, B. mimo-
sarum). Compared with other published studies, we
found a large number of beta-rhizobial species in French
Guiana and more interestingly, a high differentiation of
populations between sites (at both inter and intraspecific
levels). It is clear that soil parameters as well as the geo-
graphical distribution and competitive ability to nodulate
of the various rhizobial species affected the trapping
results, and these could explain the specific distribution
of the isolates. The PCA analyses of soil parameters and
bacterial species revealed the significance of soil pH,
phosphate, CaCO3 content and granulometry on the dis-
tribution of rhizobial populations, especially on the pres-
ence of C. taiwanensis in alkaline soils. It should be
noted, however, that C. taiwanensis was found only in
one soil, so it is premature to make any definitive conclu-
sions about any affinity of C. taiwanensis strains to alka-
line soil until more soil sampling has been undertaken.
On the other hand, in the case of acidic pH, several stud-
ies have already underlined the preference of Burkholderia
species for acidic soils (Garau et al., 2009; dos Reis Junior
et al., 2010), which is a feature that might explain their
near absence from the basic soil from which Cupriavidus
strains were isolated. Rhizobium sp. strains were only
found in a soil with neutral pH, but as with C. taiwanensis,
more sampling is required to determine whether the
occurrence of Rhizobium in Mimosa nodules has any spe-
cific connection with soil pH.
Burkholderia phymatum and B. tuberum are
common M. pudica symbionts in French Guiana
This study is the first to our knowledge to use a trapping
strategy, with M. pudica as a trap host, to uncover the
genetic diversity of beta-rhizobia in soils of South America.
The diversity of M. pudica compatible symbionts sam-
pled by our trapping strategy revealed four species of
Burkholderia (B. phymatum, B. mimosarum, B. tuberum
and a yet unnamed Burkholderia species), C. taiwanensis
and a Rhizobium sp., with the diversity in species assem-
blages (or dominance of a single one) depending on the
soil under study. We also report strains of B. cepacia
inhabiting some nodules that were unable to induce any
nodulation when inoculated as pure cultures on M. pudic-
a. The most sampled species was B. tuberum which com-
prised 60% of all isolates (131 strains), with a dominant
16S rRNA RFLP type found in all soils (except one), and
highly diversified (in terms of REP-PCR profiles) in soil
S2 (10 haplotypes) but strangely not in S3 (4 haplotypes).
We chose to name all these strains B. tuberum as they
were phylogenetically grouped in the same clade as B.
tuberum STM678T (in both the 16S rRNA gene and recA
trees), although several subgroups can be observed, and so
we cannot exclude the possibility that several species are
actually present. Interestingly, this ‘large’ B. tuberum-like
species was also detected in the diversity study of Bon-
temps et al. (2010) on rhizobial isolates from 47 Mimosa
species in Brazil (constituting 54% of the 143 rhizobial
isolates), with one strain being reported as isolated from a
M. pudica nodule (JPY389). This species has also been
reported in M. pudica nodules in Costa Rica and Panama
(Barrett & Parker, 2005, 2006). Burkholderia tuberum thus
Table 4. Macroptilium atropurpureum trapping: strain species assignment and distribution in soils
Species*
Representative
strain
16S-RFLP
ribotype S1 S2 S3 S4 All
Bradyrhizobium elkanii STM4798 HMH 6 – – – 6
B. elkanii STM4804 IMI 3 6 2 5 16
Bradyrhizobium japonicum STM4811 LOK – – 1 4 5
B. japonicum STM4814 LNI – – – 1 1
Bradyrhizobium sp. STM4800 INI – 1 – – 1
Burkholderia tuberum STM4801 ABF – 1 – – 1
Total 9 8 3 10 30
S1–S4 refer to the soils listed in Table 1, and numbers indicates distribution of 16S rRNA gene-RFLP types across soils.
ND, not determined.
*Species assignment was based on 16S rRNA phylogeny.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
12 R.P.N. Mishra et al.

seems to be a well-adapted symbiont of Mimosa species in
South America, widespread in the area of origin and
diversification of the genus Mimosa (the Brazilian Cerra-
do), as well as on the French Guianan coast.
Burkholderia phymatum represented 28% of all sampled
strains with 66 isolates, and was found in four soils (out
of eight sampled), including 15 REP-PCR profiles. Burk-
holderia phymatum was named on the basis of only one
strain (STM815, Vandamme et al., 2002) and was recently
enlarged to three recently by the addition of two Mimosa
isolates from Papua New Guinea (Elliott et al., 2007a).
Our study thus enhances the number of worldwide iso-
lates of B. phymatum to a total of 66 strains, with at least
15 different REP-PCR profiles. The STM815 REP-PCR
profile was close to the most abundant one (BP1) recov-
ered from two soils in the present study. STM815 origi-
nated from a M. lunatum nodule, and so we also isolated
the nodule symbionts of this legume species where it was
present in site S1 (which hosted several B. phymatum
strains from M. pudica). The rhizobial symbionts isolated
from seven nodules from M. lunatum trees were all
restricted to the genus Bradyrhizobium (L. Moulin,
unpublished data). Taken together with the fact that
STM815 does not form nodules on another species of
Machaerium (M. brasilense, Elliott et al., 2007a, b),
M. lunatum does not appear to be a likely host for
B. phymatum, and M. pudica can be accepted as the ori-
ginal and specific host for this species. On the other
hand, Bontemps et al. (2010) did not find any B. phyma-
tum among the 147 strains in their collection from
diverse Mimosa species in the Cerrado, a result that can
be linked to the almost absence of M. pudica in the non-
urbanized areas of this region in which the (mainly ende-
mic) native Mimosa spp. grow. Burkholderia phymatum
might thus be a specific symbiont of M. pudica and be
restricted to soils where this species is present. This idea
is strengthened by the fact that dos Reis Junior et al.
(2010) clearly showed that Cerrado/Caatinga Mimosa
natives and endemics do not nodulate effectively (or at
all) with B. phymatum. However, it should be noted that
BP is capable of effectively nodulating a wide range of
widespread and weedy Mimosa species (Elliott et al.,
2007a; dos Reis Junior et al., 2010), which often share the
same environments as M. pudica, and so we cannot
exclude the possibility that these species might also have
B. phymatum as a natural symbiont. Other Mimosa sym-
biont diversity studies in Central and South America
(Costa Rica, Panama, Brazil, Venezuela), which were per-
formed on a more restricted set of Mimosa species
(including M. pigra, M. pudica, M. scabrella, M. caesalpi-
niifolia and M. diplotricha) reported nodulation only
by B. tuberum and various new species of Burkholderia
(B. mimosarum, B. nodosa, B. sabiae), but not by B. phy-
matum (Barrett & Parker, 2005, 2006; Chen et al., 2005a,
b). Taken together, these data suggest that B. phyma-
tum is less frequent as a Mimosa species symbiont
than B. tuberum, which has spread across the whole con-
tinent up to at least Central America. The apparently
weak geographical distribution of B. phymatum is uncor-
related with the broad host range and high competitive
ability of this species (Elliott et al., 2007a, 2009). On the
other hand, a recent study of Liu et al. (2011)
has described B. phymatum (as well as B. mimosarum and
C. taiwanensis) isolates from M. pudica nodules in China,
and thus, B. phymatum might have a broader geographi-
cal distribution than was previously thought.
Several strains of B. mimosarum were also found in our
sampling. This species was reported in several symbiont
diversity studies of Mimosa species in Taiwan (M. pigra;
Chen et al., 2005a, 2006), China (M. pudica, M. diplotri-
cha; Liu et al., 2011), Brazil (M. pigra, M. pudica, M. sca-
brella; Chen et al., 2005b; Bontemps et al., 2010) and
Australia (M. pigra; Parker et al., 2007), which taken
together suggest that B. mimosarum is relatively specific
to M. pigra. This hypothesis is also strengthened by com-
petitive nodulation assays (Elliott et al., 2009) showing
that B. mimosarum PAS44 is more competitive for nodu-
lation of M. pigra than B. phymatum STM815 (highly
competitive) and C. taiwanensis LMG19424, at least in a
solid medium.
The occurrence of C. taiwanensis as a M. pudica symbi-
ont in French Guiana is more enigmatic, as it had not pre-
viously been reported in South America. The detection of
this species in our study was observed in only one soil,
which was characterized by a high pH, which might
explain its predominance at this site over Burkholderia
which appears to be more adapted to acidic soils (Table 1,
Table S1, Garau et al., 2009; dos Reis Junior et al., 2010).
The presence of a low number of alpha-rhizobial iso-
lates in our collection was not surprising, as several other
studies have already found alpha rhizobia in Mimosa
nodules (Chen et al., 2003; Barrett & Parker, 2006; Elliott
et al., 2009; Bontemps et al., 2010), although they always
represented a low% of sampled isolates compared with
beta-rhizobia. The French Guiana Rhizobium isolates were
very close to the Costa Rican strains tpud22.2 and
tpud44A (99% identity in 16S rRNA), which were iso-
lated from nodules collected from wild M. pudica plants
(Barrett & Parker, 2006). All three strains might represent
a new species close to R. endophyticum (both in
16S rRNA and recA phylogenies), which is a nonsymbiot-
ic P. vulgaris endophyte from Mexico (Lopez-Lopez et al.,
2010). Moreover, the same rhizobial genotype was found
in a parallel study on M. pudica symbionts in New
Caledonia, indicating that this Rhizobium species is spe-
cific and distributed worldwide spread as a M. pudica-
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 13

interacting bacterium (A. Klonowska, unpublished data).
It is interesting to note that this group of strains is unre-
lated to other Mimosa-compatible Rhizobium species
described so far, such as R. tropici TJ171 and TJ172 iso-
lated from M. diplotricha in Taiwan and NGR181 from
Mimosa ceratonia in Papua New Guinea, or Rhizobium
etli bv. mimosae TJ167 from M. diplotricha in Taiwan and
Mim1 from Mimosa affinis in Mexico (Wang et al., 1999;
Chen et al., 2003; Elliott et al., 2009). The French Guiana
Rhizobium isolates exhibited atypical symbiotic gene
properties, hosting two separate copies of nodA gene
(Fig. 2a). Such a feature has not been reported to date to
our knowledge, and the full genome sequencing of
STM3625 confirmed the presence of two nodA copies in
two separate operons under the control of nodboxes (L.
Moulin, unpublished data). Such features are currently
under study to unravel the putative functional role of
both copies in the symbiotic interaction.
Symbiotic genes of M. pudica beta-rhizobia are
monophyletic
The nodA, nodC and nifH gene phylogenies of M. pudica
beta rhizobia from French Guiana were monophyletic and
mirrored the 16S rRNA and recA gene phylogenies. Such a
phylogenetic pattern indicates that the ancestor of con-
temporary Burkholderia species had already interacted
with legumes (probably with the ancestor of Mimosa) and
that nod genes were vertically inherited during Burkholde-
ria speciation events. Such a pattern had already been
observed in the phylogeny of nodC genes from Burkholde-
ria symbionts of 47 Mimosa species (Bontemps et al.,
2010), and the present study can add B. phymatum to this
pattern, as this species has not been recovered from any
study in Brazil. As with almost all alpha-rhizobia, the nod-
ulation genes of beta-rhizobia are assembled in operons
carried by symbiotic plasmids (Moulin et al., 2001; Chen
et al., 2003; Amadou et al., 2008) and they are supposed
to have co-evolved jointly. Therefore, finding common
evolutionary patterns for both genes (nodA and nodC) is
not surprising and adds support to their vertical inheri-
tance from a common symbiotic ancestor. The nodA and
nifH phylogenies placed the French Guiana Cupriavidus
strain in a sister clade to B. phymatum/B. sabiae, together
with other C. taiwanensis strains sampled in South Eastern
Asia. Such a phylogenetic position suggests that Cupriavi-
dus inherited nod genes from an ancestor of the B. phyma-
tum and B. sabiae species (whch are closely related to each
other according to their 16S rRNA and recA gene phyloge-
nies). However, incongruence was found in the branching
of the CT subclade in the phylogenies of the symbiotic
genes: CT grouped as a sister clade to the BP/BS clade in
the nodA and nifH trees, but in the nodC phylogeny CT
was positioned as an out-group to the Burkholderia
Mimosa panclade. Symbiotic genes in C. taiwanensis thus
appear to have a more complex evolutionary history than
those in Burkholderia, with several nod gene transfers.
As the Mimosa genus diversified into more than 496
species in the Americas (Simon et al., 2011), it is possible
that Burkholderia co-diversified with Mimosa into several
symbiotic species, each exhibiting some specificity in their
host range. Several studies have already shown the affinity
of Burkholderia spp. to Mimosa species, such as B. sabiae
with M. caesalpiniifolia (Chen et al., 2008) and B. mimo-
sarum with M. pigra (Chen et al., 2005a, b, Chen et al.,
2006), which might explain their relatively weak occur-
rence in our M. pudica trapping experiments.
The B. tuberum species contains at least two
biovars
While a pattern of codiversification between Burkholderia
and Mimosa species is suggested, a case of adaptation of a
beta-rhizobial species to different legume subfamilies was
found with B. tuberum. Indeed, B. tuberum Mimosa
isolates grouped in a subclade of the large Mimosa-
beta-rhizobia panclade in the nodA and nodC trees, while
B. tuberum from papilionoid species (STM678 isolated
from A. carnosa and able to nodulate Cyclopia species,
Moulin et al., 2001; Elliott et al., 2007b) grouped in a
branch together with Burkholderia sp. (BSP4) strains from
Rhynchosia ferulifolia (Garau et al., 2009) and was placed
outside the nodA ‘Mimosa’ panclade (with alpha rhizobia,
such as Methylobacterium and Bradyrhizobium). In this
latter case, the nodA phylogeny of beta-rhizobia correlated
with the host range rather than with the bacterial species
phylogeny, thus indicating different origins of nod genes
in B. tuberum. The nifH phylogeny also separated the
papilionoid isolates (which were close to B. xenovorans)
from the mimosoid ones (which grouped with other
Mimosa Burkholderia isolates). It is also relevant to note
that B. tuberum is the only known species of beta-rhizo-
bia with strains able to nodulate two subfamilies of
legumes (Mimosoids and Papilionoids). Burkholderia
tuberum thus appears as a more diversified legume symbi-
ont than B. phymatum in terms of geographical distribu-
tion and legume host range, and two biovars can be
defined according to their host range and nod gene phy-
logeny: biovar mimosae (Mimosa symbionts) and biovar
papilionoideae (Cyclopia symbionts), originating from
two separate horizontal transfers of nodulation genes.
Conclusion
This study has shown that M. pudica is a good trapping
host to study the diversity of soil beta rhizobia and that
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
14 R.P.N. Mishra et al.

it exhibits a high affinity for several Burkholderia species.
Our study also revealed a site-dependant distribution of
beta-rhizobial species. A vertical inheritance of symbiotic
genes among Burkholderia species was also highlighted.
The specific distribution of rhizobia in diverse soils could
be attributed occasionally to environmental factors, and it
showed variable phylogeographical patterns. The large
number of isolates sampled in this study will be further
characterized for their nodulation competitive abilities
and symbiotic efficiencies to understand how and to
which extent diverse beta-rhizobial species have adapted
to Mimosa species in comparison with alpha-rhizobia.
Moreover, the full genome sequencing of Rhizobium sp.
STM3625 and B. tuberum STM3649 at Genoscope as well
as B. mimosarum STM3621 and C. taiwanensis STM6018
as part of the GEBA-RNB project (Genome Encyclopedia
of Bacteria & Archae, Root Nodulating Bacteria) at the
Joint Genome Institute, should give insights into the
adaptation of these various species towards establishing
symbiosis with legumes.
Acknowledgements
We thank Marcelo Simon for identification of the M. pu-
dica ecotype, Euan James for helpful discussions, and the
French National Agency of Research (ANR) for funding
(Project ‘BETASYM’ ANR-09-JCJC-0046). R.P.N.M. was
funded by a postdoctoral grant from IFCPAR (Indo-
French Centre for the Promotion of Advanced Research).
References
Amadou C, Pascal G, Mangenot S et al. (2008) Genome
sequence of the beta-rhizobium Cupriavidus taiwanensis and
comparative genomics of rhizobia. Genome Res 18: 1472–
1483.
Andam CP, Mondo SJ & Parker MA (2007) Monophyly of
nodA and nifH genes across Texan and Costa Rican
populations of Cupriavidus nodule symbionts. Appl Environ
Microbiol 73: 4686–4690.
Barrett CF & Parker MA (2005) Prevalence of Burkholderia sp.
nodule symbionts on four mimosoid legumes from Barro
Colorado Island, Panama. Syst Appl Microbiol 28: 57–65.
Barrett CF & Parker MA (2006) Coexistence of Burkholderia,
Cupriavidus, and Rhizobium sp. nodule bacteria on two
Mimosa spp. in Costa Rica. Appl Environ Microbiol 72:
1198–1206.
Bontemps C, Elliott GN, Simon MF et al. (2010) Burkholderia
species are ancient symbionts of legumes. Mol Ecol 19: 44–52.
Chen WM, Laevens S, Lee TM, Coenye T, De Vos P, Mergeay
M & Vandamme P (2001) Ralstonia taiwanensis sp. nov.,
isolated from root nodules of Mimosa species and sputum
of a cystic fibrosis patient. Int J Syst Evol Microbiol 51:
1729–1735.
Chen WM, Moulin L, Bontemps C, Vandamme P, Bena G &
Boivin-Masson C (2003) Legume symbiotic nitrogen
fixation by beta-proteobacteria is widespread in nature.
J Bacteriol 185: 7266–7272.
Chen WM, James EK, Chou JH, Sheu SY, Yang SZ & Sprent JI
(2005a) Beta-rhizobia from Mimosa pigra, a newly discovered
invasive plant in Taiwan. New Phytol 168: 661–675.
Chen WM, de Faria SM, Straliotto R et al. (2005b) Proof that
Burkholderia strains form effective symbioses with legumes:
a study of novel Mimosa-nodulating strains from South
America. Appl Environ Microbiol 71: 7461–7471.
Chen WM, James EK, Coenye T, Chou JH, Barrios E, de Faria
SM, Elliott GN, Sheu SY, Sprent JI & Vandamme P (2006)
Burkholderia mimosarum sp. nov., isolated from root
nodules of Mimosa spp. from Taiwan and South America.
Int J Syst Evol Microbiol 56: 1847–1851.
Chen WM, de Faria SM, James EK, Elliott GN, Lin KY, Chou
JH, Sheu SY, Cnockaert M, Sprent JI & Vandamme P
(2007) Burkholderia nodosa sp. nov., isolated from root
nodules of the woody Brazilian legumes Mimosa
bimucronata and Mimosa scabrella. Int J Syst Evol Microbiol
57: 1055–1059.
Chen WM, de Faria SM, Chou JH, James EK, Elliott GN,
Sprent JI, Bontemps C, Young JP & Vandamme P (2008)
Burkholderia sabiae sp. nov., isolated from root nodules of
Mimosa caesalpiniifolia. Int J Syst Evol Microbiol 58: 2174–
2179.
Dereeper A, Guignon V, Blanc G et al. (2008) Phylogeny.fr:
robust phylogenetic analysis for the non-specialist. Nucleic
Acids Res 36: 465–469.
dos Reis Junior FB, Simon MF, Gross E et al. (2010)
Nodulation and nitrogen fixation by Mimosa spp. in the
Cerrado and Caatinga biomes of Brazil. New Phytol 186:
934–946.
Elliott GN, Chen WM, Chou JH et al. (2007a) Burkholderia
phymatum is a highly effective nitrogen-fixing symbiont of
Mimosa spp. and fixes nitrogen ex planta. New Phytol 173:
168–180.
Elliott GN, Chen WM, Bontemps C, Chou JH, Young JP,
Sprent JI & James EK (2007b) Nodulation of Cyclopia spp.
(Leguminosae, Papilionoideae) by Burkholderia tuberum.
Ann Bot 100: 1403–1411.
Elliott GN, Chou JH, Chen WM, Bloemberg GV, Bontemps C,
Martinez-Romero E, Velazquez E, Young JP, Sprent JI &
James EK (2009) Burkholderia spp. are the most competitive
symbionts of Mimosa, particularly under N-limited
conditions. Environ Microbiol 11: 762–778.
Garau G, Yates RJ, Deiana P & Howieson JG (2009) Novel
strains of nodulating Burkholderia have a role in
nitrogen fixation with papilionoid herbaceous legumes
adapted to acid, infertile soils. Soil Biol Biochem 41:
125–134.
Guindon S, Lethiec F, Duroux P & Gascuel O (2005)
PHYML Online – a web server for fast maximum
likelihood-based phylogenetic inference. Nucleic Acids Res
33: 557–559.
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 15

Gyaneshwar P, Hirsch AM, Moulin L et al. (2011) Legume-
nodulating betaproteobacteria: diversity, host range and
future prospects. Mol Plant Microbe Interact 24: 1276–1288.
Hammer Ø, Harper DAT & Ryan PD (2001) PAST:
Paleontological Statistics Software Package for education
and data analysis. Palaeontologia Electronica 4: 4–9.
Kaschuk G, Hungria M, Andrade DS & Campos RJ (2006)
Genetic diversity of rhizobia associated with common bean
(Phaseolus vulgaris L.) grown under no-tillage and
conventional systems in Southern Brazil. Appl Soil Ecol 32:
210–220.
Lima AS, Nobrega RSA, Barberi A, da Silva K, Ferreira DF &
Moreira FMS (2009) Nitrogen-fixing bacteria
communities occurring in soils under different uses in the
Western Amazon Region as indicated by nodulation of
siratro (Macroptilium atropurpureum). Plant Soil 319:
127–145.
Liu XY, Wu W, Wang ET, Zhang B, Macdermott J & Chen
WX (2011) Phylogenetic relationships and diversity of beta-
rhizobia associated with Mimosa spp. grown in
Sishuangbanna, China. Int J Syst Evol Microbiol 61: 334–342.
Lopez-Lopez A, Rogel MA, Ormeno-Orrillo E, Martinez-
Romero J & Martinez-Romero E (2010) Phaseolus vulgaris
seed-borne endophytic community with novel bacterial
species such as Rhizobium endophyticum sp. nov. Syst Appl
Microbiol 33: 322–327.
Miche L, Moulin L, Chaintreuil C, Contreras-Jimenez JL,
Munive-Hernandez JA, Del Carmen Villegas-Hernandez M,
Crozier F & Bena G (2010) Diversity analyses of
Aeschynomene symbionts in Tropical Africa and Central
America reveal that nod-independent stem nodulation is
not restricted to photosynthetic bradyrhizobia. Environ
Microbiol 12: 2152–2164.
Moulin L, Munive A, Dreyfus B & Boivin-Masson C (2001)
Nodulation of legumes by members of the beta-subclass of
Proteobacteria. Nature 411: 948–950.
Moulin L, Bena G, Boivin-Masson C & Stepkowski T (2004)
Phylogenetic analyses of symbiotic nodulation genes support
vertical and lateral gene co-transfer within the
Bradyrhizobium genus. Mol Phylogenet Evol 30: 720–732.
Nicholas KB & Nicholas BJ (1997) Genedoc: a tool for editing
and annotating multiple sequence alignments. Distributed
by the author.
Normand P, Cournoyer B, Simonet P & Nazaret S (1992)
Analysis of a ribosomal RNA operon in the actinomycete
Frankia. Gene 111: 119–124.
N’Zoue A, Miche L, Klonowska A, Laguerre G, de Lajudie P &
Moulin L (2009) Multilocus sequence analysis of
bradyrhizobia isolated from Aeschynomene species in
Senegal. Syst Appl Microbiol 32: 400–412.
Parker MA (2008) Symbiotic relationships of legumes and
nodule bacteria on Barro Colorado Island, Panama: a
review. Microb Ecol 55: 662–672.
Parker MA, Wurtz A & Paynter Q (2007) Nodule symbiosis of
invasive Mimosa pigra in Australia and in ancestral habitats:
a comparative analysis. Biol Invasions 9: 127–138.
Rincon-Rosales R, Lloret L, Ponce E & Martinez-Romero E
(2009) Rhizobia with different symbiotic efficiencies
nodulate Acaciella angustissima in Mexico, including
Sinorhizobium chiapanecum sp. nov. which has common
symbiotic genes with Sinorhizobium mexicanum. FEMS
Microbiol Ecol 67: 103–117.
Rousset F (2008) genepop’007: a complete re-implementation
of the genepop software for Windows and Linux. Mol Ecol
Resour 8: 103–106.
Simon MF & Proenca C (2000) Phytogeographic patterns of
Mimosa (Mimosoideae, Leguminosae) in the Cerrado biome
of Brazil: an indicator genus of high altitude centers of
endemism? Biol Conserv 96: 279–296.
Simon MF, Grether R, Queiroz LP, Sarkinen TE, Dutra VF &
Hughes CE (2011) The evolutionary history of Mimosa
(Leguminosae): towards a phylogeny of the sensitive plants.
Am J Bot 98: 1201–1221.
Stepkowski T, Moulin L, Krzyzanska A, McInnes A, Law IJ &
Howieson J (2005) European origin of Bradyrhizobium
populations infecting lupins and serradella in soils of
Western Australia and South Africa. Appl Environ Microbiol
71: 7041–7052.
Sy A, Giraud E, Jourand P et al. (2001) Methylotrophic
Methylobacterium bacteria nodulate and fix nitrogen in
symbiosis with legumes. J Bacteriol 183: 214–220.
Tamura K, Dudley J, Nei M & Kumar S (2007) MEGA4:
Molecular Evolutionary Genetics Analysis (MEGA) software
version 4.0. Mol Biol Evol 24: 1596–1599.
Vandamme P & Coenye T (2004) Taxonomy of the genus
Cupriavidus: a tale of lost and found. Int J Syst Evol
Microbiol 54: 2285–2289.
Vandamme P, Goris J, Chen WM, de Vos P & Willems A
(2002) Burkholderia tuberum sp. nov. and Burkholderia
phymatum sp. nov., nodulate the roots of tropical legumes.
Syst Appl Microbiol 25: 507–512.
Verma SC, Chowdhury SP & Tripathi AK (2004) Phylogeny
based on 16S rDNA and nifH sequences of Ralstonia
taiwanensis strains isolated from nitrogen-fixing nodules of
Mimosa pudica, in India. Can J Microbiol 50: 313–322.
Versalovic J, Koeuth T & Lupski JR (1991) Distribution of
repetitive DNA sequences in eubacteria and application to
fingerprinting of bacterial genomes. Nucleic Acids Res 19:
6823–6831.
Vincent JM (1970) A manual for the practical study of root-
nodule bacteria. I.B.P. Handbook, vol. 15. Blackwell Scientific
Publications, Ltd, Oxford.
Wang ET, Rogel MA, Garcia-de los Santos A, Martinez-
Romero J, Cevallos MA & Martinez-Romero E (1999)
Rhizobium etli bv. mimosae, a novel biovar isolated from
Mimosa affinis. Int J Syst Bacteriol 49: 1479–1491.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Ecol && (2011) 1–17
Published by Blackwell Publishing Ltd. All rights reserved
16 R.P.N. Mishra et al.

Fig. S1. Localization of sampling sites.
Fig. S2. PCA analysis representation of soils with regard
to soil physico-chemical parameters and M. pudica rhizo-
bial species distribution.
Table S1. Listing of strains trapped on French Guiana
sois by Mimosa pudica and Macroptilium atropurpureum.
Table S2. REP-PCR grouping and geographical distribution.
Table S3. Accession numbers used for reference strains in
16S rRNA, recA, nifH, nodA and nodC gene phylogenies,
by order of appearance from top to bottom in Fig. 1a
(16S rRNA), 1B (recA), 2A (nodA), 2B (nifH) and 3
(nodC).
Please note: Wiley-Blackwell is not responsible for the
content or functionality of any supporting materials sup-
plied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
FEMS Microbiol Ecol && (2011) 1–17 ª 2011 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved
Investigating the origin and diversity of beta-rhizobia 17
Related Documents











